 Ludivine Obry1†
Ludivine Obry1† Raissa Medina-Santos1†
Raissa Medina-Santos1† Myriam Rahmouni1
Myriam Rahmouni1 Josselin Noirel1
Josselin Noirel1 Toufik Labib1Pilar Galan2Jean-Louis Spadoni1Gaëlle Gendronneau3Randa Jdid3
Toufik Labib1Pilar Galan2Jean-Louis Spadoni1Gaëlle Gendronneau3Randa Jdid3 Sandra Courrèges3Julie Latreille3
Sandra Courrèges3Julie Latreille3 Nada André3
Nada André3 Jean-François Zagury1*
Jean-François Zagury1* Sigrid Le Clerc1*
Sigrid Le Clerc1*- 1Laboratoire Génomique, Bioinformatique, et Chimie Moléculaire, EA7528, Conservatoire National des Arts et Métiers, Paris, France
- 2Equipe de Recherche en Epidémiologie Nutritionnelle (EREN), Centre d'Epidemiologie et Biostatistiques Sorbonne Paris Cité (CRESS), Inserm U1153, Inra U1125, Cnam, Université Sorbonne Paris Nord et Sorbonne-Paris-Cité, Paris, France
- 3Biological and Clinical Department, IRD Chanel Fragrance & Beauty, Bobigny, France
Background: Skin aging is characterized by observable major changes in the composition and organization of the skin, including the appearance of wrinkles, tissue sagging, dryness, and pigmentary disorders. While both environmental and genetic factors contribute to these changes, their interaction remains underexplored. Perceived age is a biomarker of health and strongly related to facial skin aging features. Thus, we conducted a gene-environment interaction analysis on the perceived facial aging progression phenotype in 226 women, focusing on sun exposure as the environmental factor.
Methods: We assessed perceived age in 226 women at two time points, 12 years apart, allowing defining a perceived facial aging progression as phenotype. We applied a two-step gene-environment interaction approach. First, a genome-wide association study (GWAS) was performed to identify SNP effects on the perceived facial aging progression, selecting those with a p-value <5 × 10−3. In the second step, we used GEM software to test interactions between the 7,464 selected SNP and sun exposure.
Result: The GWAS identified four significant SNP associated with perceived facial aging progression, located in the CGGBP1, PGM5-AS1, and CSMD1 genes. The CGGBP1 gene is involved in DNA damage/repair, telomere and mRNA metabolism; PGM5-AS1 is an antisense RNA regulating PGM5, a component of adherens-type cell junctions; CSMD1 is involved in complement regulation and cell migration. In the second step, we tested 7,464 SNP for interactions with sun exposure in perceived facial aging progression, identifying a significant interaction signal in the VANGL1 gene. The VANGL1 gene contributes to the structural organization of the plasma membrane and has been identified as a key gene for cutaneous function and was expressed during keratinization.
Discussion: This study revealed four relevant genetic associations with skin aging and one significant G × E interaction. These findings support the known link between telomere shortening/protection and aging, and suggesting a potential role for keratinization in the context of facial aging and sun exposure, though further validation in larger cohorts is necessary. The findings could help to develop new approaches for facial aging prevention and treatment and to better understand molecular mechanisms of aging.
Introduction
Skin aging is characterized by observable major changes in the composition and organization of the skin, including the appearance of wrinkles, tissue sagging, dryness, and pigmentary disorders. These changes are also accompanied by alterations in the underlying skeletal and adipose structures of the face, which further contribute to the overall appearance of aging (Coleman and Rajiv, 2006; Ng and Chew, 2022). Both environmental and genetic factors significantly influence facial aging, and their respective contributions have been studied (Ng and Chew, 2022; Krutmann et al., 2017). While these factors have been studied individually, less is known about how they interact and contribute to the aging process collectively (Krutmann et al., 2017).
The interaction between genetic variations and environmental factors (G × E) have a significant impact on complex human traits and diseases, including skin and facial aging (Krutmann et al., 2017; Herrera-Luis et al., 2024). The skin serves as an excellent model for studying G × E interactions, as it undergoes both intrinsic aging and extrinsic aging in areas exposed to environmental factors, such as the face (Krutmann et al., 2021). Genetic predispositions may influence how skin responds to environmental stressors such as UV radiation and pollution, thus investigating these interactions could provide critical insights into the underlying biological processes of skin aging and overall progression of the aging process (Krutmann et al., 2021).
Among the high number of skin aging phenotype definitions, perceived facial age, a subjective assessment based on an individual’s appearance, stands out as a relevant phenotype for studying G × E interactions due to its inclusion of several features related to skin aging and its association with both genetic and environmental factors (Ng and Chew, 2022; Roberts et al., 2020; Avila et al., 2023; Voegeli et al., 2021). While perceived facial aging is mainly explained by skin aging features (wrinkles, sagging and hyperpigmentation), the muscles, the fat distribution, and the ligaments contribute to the complex underlying anatomy of the face that modulates perception of facial aging (Gunn et al., 2009; Nkengne et al., 2008; Merinville et al., 2015; Swift et al., 2021; Cotofana et al., 2016). Consequently, Perceived age is multifactorial process, not limited to skin changes. Moreover, perceived facial aging is a robust biomarker of ageing that predicts survival in elderly individuals and health (Christensen et al., 2009; Rippon and Steptoe, 2015). It therefore provides a holistic phenotype for G × E studies on facial aging and overall health.
To further explore the role of G × E interactions in perceived facial aging, we assessed perceived age in 226 women at two time points, 12 years apart, allowing defining a perceived facial aging progression (PFAP) as phenotype. Then, we conducted a two-step study focusing on perceived age and sun exposure, allowing reducing the multiple testing burden by focusing on SNP likely to be involved in interactions (Gauderman et al., 2017). In the first step, a genome-wide association study (GWAS) was performed to identify SNP with potential marginal genetic effect on PFAP, and in the second step, we tested these SNP for interaction with sun exposure.
Materials and methods
Cohort
A cross-sectional study was conducted to investigate skin aging in the context of the SU.VI.MAX cohort, a longitudinal cohort study, conducted in French middle-aged adults (Galan et al., 1998). The protocol was approved by the Hospital Medicals Ethics Committee of Paris-Cochin (CCPPRB no. 706) and the ‘‘Commission Nationale de l’Informatique et des Libertés’’ (CNIL no. 334641). The study was conducted according to the Declaration of Helsinki Principles. All participants gave their written, informed consent. The SU.VI.MAX cohort included 13,017 volunteers who were representative of the French adult middle-aged population for most sociodemographic features (Hercberg et al., 2004). The individuals were extracted from SU.VI.MAX in 2009 to investigate skin aging genes in women. 570 women, aged 44–70 years, took part in this study and provided their informed consent (Le Clerc et al., 2013).
Assessment of perceived age and sun exposure
Among the 570 women who took part in this study in 2002, 68 were excluded due to quality control (see details in Le Clerc et al., 2013). Among the remaining 502 women, 226 participated in the second round of phenotyping in 2014 and were available for the analysis of perceived facial aging progression.
Lifetime sun exposure
Lifetime sun exposure intensity was estimated in 2002 by a score based on data collected by a self-reported questionnaire. This score is a linear combination of five items weighted according to their relative contribution to the score: voluntary sun exposure, exposure of the body and the facial skin, exposure during the hottest hours of the day, intensity of self-reported lifetime sun exposure, and consideration for sunbathing. The design, validation, and description of this score have been described previously (Guinot, Malvy, and Latreille, 2001). This score was categorized into seven categories, with values ranging from 1 (lowest) to 7 (highest) exposure. The unavoidable potential bias linked to self-reported questionnaire is one limitation of our study.
Perceived age
Facial photographs were collected in 2002 and 2014 for 226 women. Three standardized, high-resolution digital images of the face were taken for each participant (one frontal view of the face and one of each profile) using a Kodak digital camera (Kodak, Paris, France) (Supplementary Figure S1). The camera was mounted on a monopod and a specifically developed chair was used to allow standardized positions of the camera with respect to the face. Lighting conditions were standardized by means of two symmetrical lamps, which provided a continuous daylight spectrum, placed at 45° to each side of the face. The participants were asked to follow specific skin care instructions; notably, application of detergents or cosmetics to the face was not authorized for at least 12 h before the study visit. They were also instructed to avoid applying makeup to their facial skin and wearing jewelry in the morning of the day of imaging. Hair was masked using a white headband. A high-resolution digital image of the frontal view of the face, with a neutral facial expression, and eyes closed was taken.
The assessment of perceived facial age involved 60 French naïve judge Caucasian women aged 30 to 70, recruited via a professional agency that provided compensation. Only women were included, as they are generally more accurate in estimating other women’s ages in everyday contexts. Inclusion criteria required participants to be French-speaking women within the target age range. Exclusion criteria included vision problems and insufficient understanding of French. Photographs were sequentially and randomly presented to the graders on a computer screen. They were requested to give subject age using E-Prime software interface that featured a slider bar spanning from 0 to 100 for age estimation (Taylor and Marsh, 2017). Photos were grouped into two blocks by year (2002 and 2014), with half the participants starting with one block or the other. To manage cognitive load, an incomplete block design was used: each participant rated 150 randomly selected photos per block. Each facial photograph was estimated by at least 33 judges (from 33 to 42 judgments). The average perceived facial age was used for the analysis.
In this study, we aimed to study a perceived facial aging progression rather than a state. For this purpose, we defined the perceived facial aging progression as:
Genotyping/imputation
The 226 women were genotyped using Illumina Infinium HumanOmni1-Quad BeadChips (Illumina, San Diego, CA) that contain 1,140,419 markers. After quality control, a total of 795,063 SNP remained for imputation. To correct for possible population stratification, genotypes were analyzed using EIGENSTRAT utility of the EIGENSOFT package version 8.0 (Price et al., 2006) to perform a principal component analysis (PCA). As a result, the 226 individuals exhibited a European ancestry and the two first component of PCA were used in the statistical analyses to avoid geographical stratification bias (Supplementary Figure S2). The genotype data were phased using SHAPEIT4 (Delaneau et al., 2019). The phased data were then imputed using IMPUTE2 (Howie et al., 2012). As reference haplotypes, we used genotype data of 2,054 individuals from the phase 3 integrated variant set of the 1000 Genomes project released in 2014 (1000 Project Consortium et al., 2010). After imputation and quality control (MAF >5%, imputation r2 > 0.3) we obtained 6,897,523 SNP for 226 individuals.
Statistical analyses
We performed a two-step procedure for analysis of G × E interaction, allowing the reduction of the multiple testing burden by focusing on SNP more likely to be involved in interactions (Gauderman et al., 2017). During the first step, we conducted a classical GWAS to assess the marginal genetic effects regarding the PFAP. We used SNPTEST software to perform linear regression with an additive model between genotype and PFAP, and we added the two first principal component of the PCA and the categorized score sun exposure as covariates (see Lifetime sun exposure), (Marchini et al., 2007). We used the genome-wide threshold of 5 × 10−8 to define significant SNP of the GWAS. For subset testing in a 2-step procedure, we considered the GWAS p-value threshold of 1 × 10−3. The second step consisted of testing the G × E interaction for perceived age phenotype and was done using GEM software with a linear regression model and with the two first principal component of the PCA as covariates and the categorized sun exposure score as interaction variable (Westerman et al., 2021). We obtained a p-value for the interaction effect and one for the genetic and interaction effects together, namely, the joint p-value. We tested the G × E interaction based on interaction effect alone between selected SNP and sun exposure regarding the PFAP phenotype. We computed linkage disequilibrium (LD) with LDlink between the selected SNPs in the GWAS, to define a set of independent SNP (LD r2 < 0.8) and the Bonferroni threshold for the interaction analysis (0.05/number of independent SNP).
Bioinformatics investigations
Following the association and interaction analyses, we carried out investigations to explore the potential functional role of the SNPs in gene expression regulation. First, we used the tool RegulomeDB (Boyle et al., 2012) and the scoring scheme of this tool refers to a higher likelihood of being functional (with 1 being higher and 7 being lower score). Then we explored several databases related to quantitative trait loci: eQTL in GTEX (Sobin et al., 2024); mQTL in mQTLdb and in EPIC DB (Gaunt et al., 2016; Villicaña et al., 2023); pQTL in SCALOP (Folkersen et al., 2020). A mQTL was considered if it existed in both mQTLdb and EPIC DB. The linkage disequilibrium was assessed using LDlink in the CEU population of 1000 genomes (Machiela and Chanock, 2015).
Results
To explore the interaction between genetic variations and sun exposure in perceived facial aging, we conducted a two-step study, using an initial screening to prioritize SNP that are more likely to be involved in an interaction and testing for interaction in a second step (Gauderman et al., 2017). We assessed perceived age in 226 women at two time points, 12 years apart, allowing us to define a perceived facial aging progression (PFAP) phenotype used as facial and skin aging phenotypes.
In the first step, we performed a classical GWAS focused on PFAP on 6,897,523 SNP (MAF >5%; imputation r2 > 0.3). We identified four significant SNP (rs114462394, rs77074982, rs567952380, rs75470026) associated with PFAP (Table 1; Figure 1). The four SNP showed imputation r2 between 0.3 and 0.4, suggesting a high level of uncertainty in the imputed genotypes. However, the MAF of these SNP in our sample were similar to those in the 1000 Genomes European population (Table 1). The genomic inflation factor did not show any deviation (λ = 1.019).
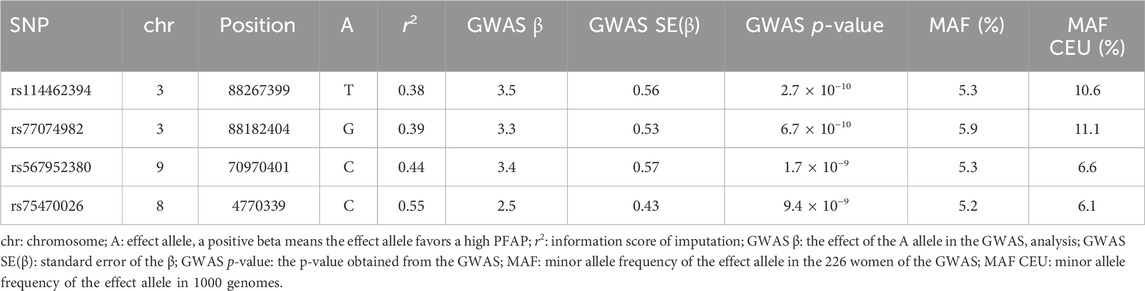
Table 1. Main associations of the GWAS.
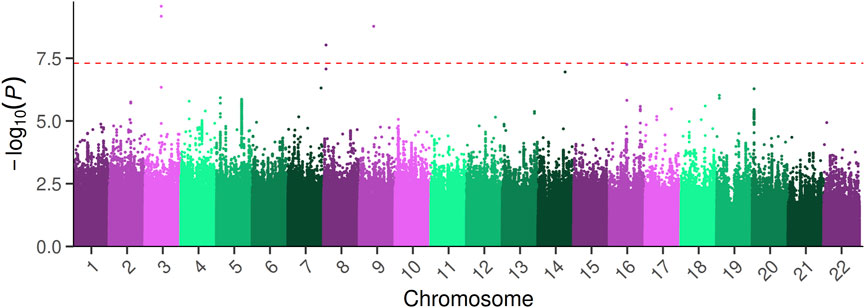
Figure 1. Manhattan plot of the GWAS analysis on PFAP. The–log10 (p-values) of the GWAS are plotted across chromosomes along the x-axis. The red dashed line indicates the genome-wide significance threshold (p-value = 5 × 10−8).
The strongest signal of the GWAS was detected at rs114462394 and rs77074982 SNP, which were in high LD (LD r2 > 0.8) with three SNP (rs114203715, rs114017876 and rs114002886) and they were all located near or in the CGGBP1 and ZNF654 genes on chromosome 3. Among them the rs114017876 and rs77074982 SNP presented several lines of evidence for their involvement in the regulation of gene expression: eQTL effects on the CGGBP1 gene in arterial aorta tissue and the CHMP2B gene in visceral adipose tissue and a high score in RegulomeDB (Table 2), (Sobin et al., 2024). The rs114017876-A allele is associated with a higher PFAP and a lower expression of CGGBP1 while the rs77074982-G allele is associated with a higher PFAP and a higher expression of CHMP2B (Tables 1,2; Figures 2A,B). The second signal was carried by the rs567952380 SNP and was located on chromosome 9 in an intron of the gene PGM5-AS1. The rs567952380-C allele was associated with a higher PFAP in our study and with an increase of PGM5 expression in cultured fibroblasts (Tables 1,2; Figure 2C). The last signal of the GWAS was observed for the rs75470026 SNP which was located on chromosome 8 in an intron of the CSMD1 gene, with the C allele associated with a higher PFAP (Tables 1,2; Figure 2D).
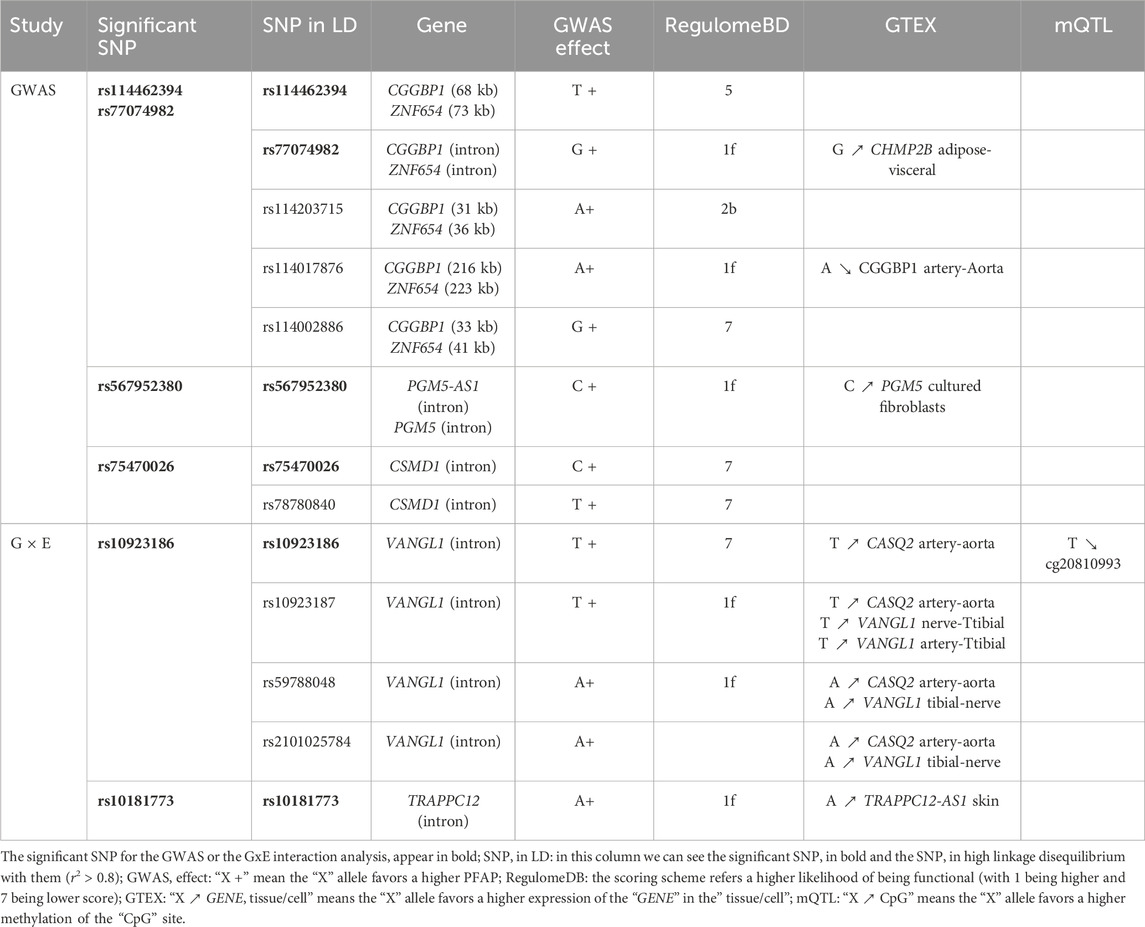
Table 2. Results of the bioinformatics investigations.
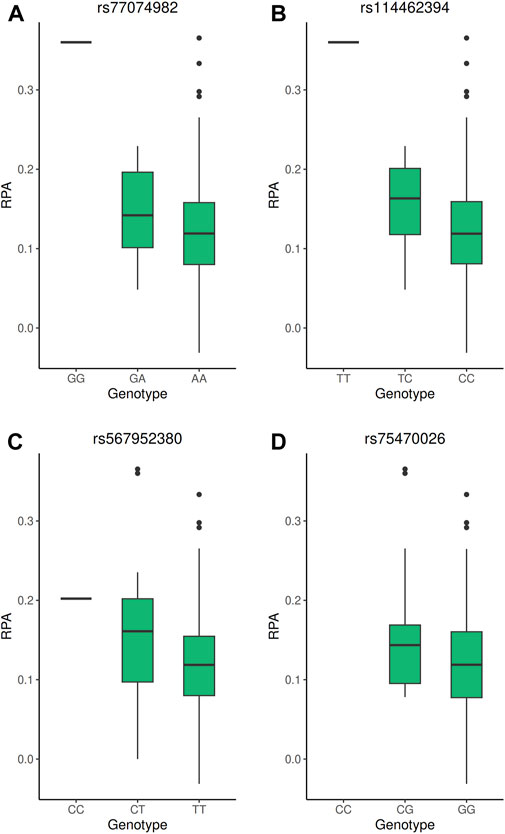
Figure 2. Boxplot of the PFAP according to the genotype of the GWAS significant SNP. (A–D) PFAP distributions according to the genotypes of the rs77074982, rs114462394, rs567952380, and rs7570026 SNP, respectively. All these SNP reached genome-wide significance threshold (5 × 10−8) in the GWAS under the additive model.
In the second step, only the SNP that passed the GWAS threshold of 1 × 10−3 (n = 7,469 SNP) were tested for an interaction with sun exposure regarding the perceived facial aging progression phenotype. In order to determine the Bonferroni threshold, we computed the LD among the 7,469 SNP to evaluate the number of independent tests (LD r2 > 0.8). We identified 1,705 independent SNP, resulting in a threshold of 2.93 × 10−5. One SNP reached the Bonferroni threshold for the G × E interaction (Table 3).
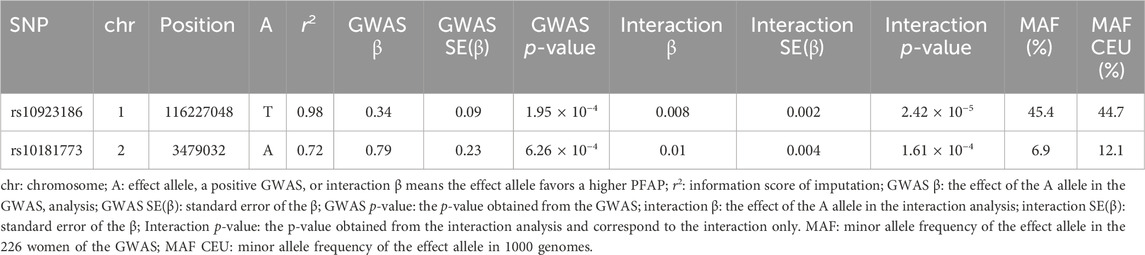
Table 3. Main associations of the G × E interaction analysis.
The only significant signal involved a group of three SNP and one indel in high LD (r2 ≃ 0.8, rs10923186, rs10923187, rs59788048 and rs111978958), with the rs10923186 SNP reaching the Bonferroni threshold (interaction p-value = 2.42 × 10−5, interaction β = 0.008, FDR = 0.04). The joint test p-value of the signal carried by rs10923186 was 9.23 × 10−8, nearly reaching the genome-wide significance threshold (5 × 10−8). To further characterize the interaction signal, we explored the genotype effects on the PFAP across sun exposure categories. These effects were estimated using an additive linear model including genotype, sun exposure, their interaction term, and adjusting for population stratification via PC1 and PC2 (Figure 3A) (R Core Team, 2024). We observed that the rs10923186-T allele favored a higher PFAP in the categories 1, 2 and 3 and, no effect or opposite effect in the categories 4, 5, 6 and 7 (Figure 3A). Overall, the rs10923186-T favored a higher PFAP in less exposed population (Figure 3B). The four variants (rs10923186, rs10923187, rs59788048 and rs111978958) were located on chromosome 1, near or within introns of the VANGL1 gene. In addition to a high score for involvement in gene expression regulation (RegulomeDB), this SNP cluster was involved in an eQTL for CASQ2 gene in artery aorta and the VANGL1 gene in tibial nerve and in a mQTL with the cg20810993 CpG site (Table 2), (Sobin et al., 2024). In summary, through the LD analysis, we observed the rs10923186-T allele was associated with higher PFAP exclusively in the low sun-exposed population, reduced methylation at cg20810993 and higher expression of CASQ2 and VANGL1 genes (Figure 3A; Tables 2,3). In addition to the first signal, we obtained a trend of G × E interaction for the rs10181773 SNP, located on chromosome 2 in an intron of the TRAPPC12 gene. The rs10181773-A allele was associated with a higher PFAP exclusively in the low sun-exposed population (FDR = 0.14, interaction p-value = 1.61 × 10−4, joint test p-value = 1.31 × 10−6; Figure 3C; Table 3). The rs10181773 SNP was involved in an eQTL with the TRAPPC12-AS1 gene in skin notably (Table 2), (Sobin et al., 2024).
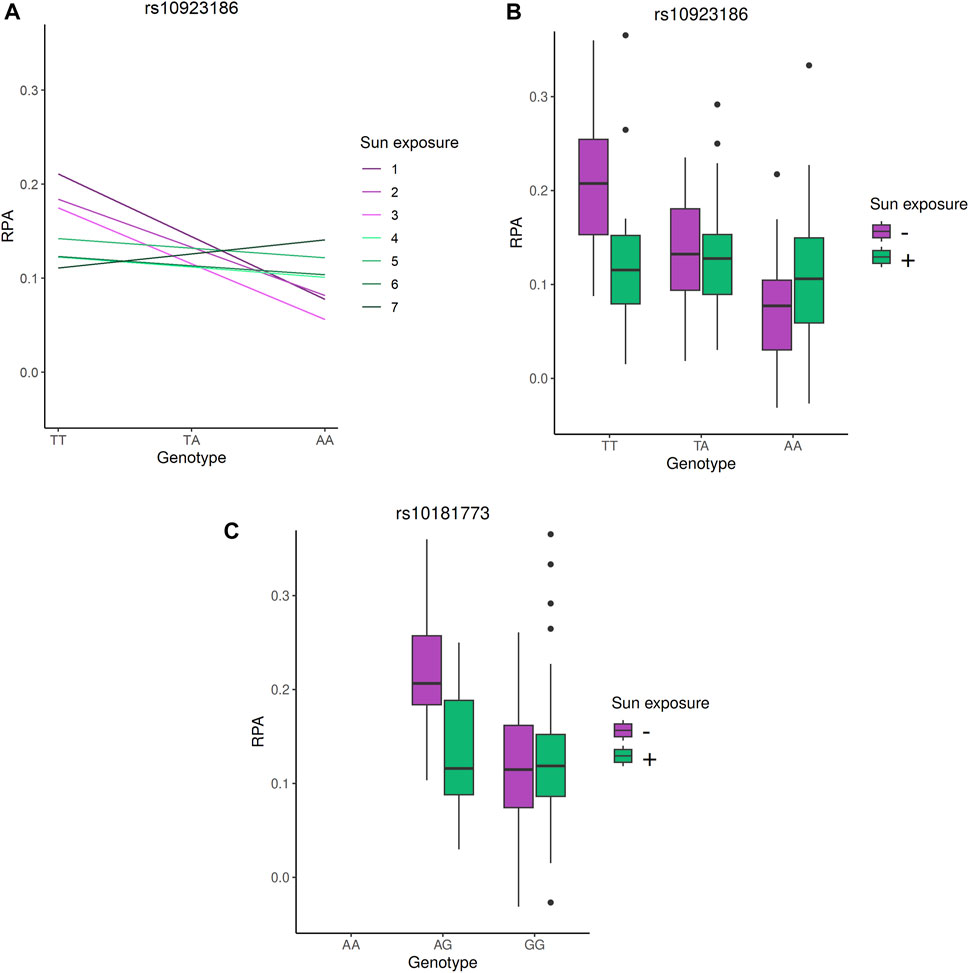
Figure 3. Genotype effects on PFAP in the G × E analysis. (A) Estimated effect of the rs10923186 T allele on PFAP within each sun exposure category. The model includes genotype, sun exposure, their interaction term, and adjustment for population structure via PC1 and PC2 and was computed with R (R Core Team, 2024). (B) and (C): PFAP distributions according to the genotypes of the rs10923186 and rs10181773 SNP, respectively. In this figure, the seven categories of sun exposure were separated into two groups: the “-” sun exposure group includes individuals with values from 1 to 3, and the “+” sun exposure group includes individuals with values from 4 to 7. This classification into two groups was used solely for visual clarity. The rs10923186 SNP reached the Bonferroni threshold (p-value <2.93 × 10−5), while the rs10181773 SNP did not (p-value = 1.61 × 10−4).
Discussion
To explore the role of G × E interactions in facial aging, we conducted a two-step study on 226 women, focusing on perceived facial aging progression with sun exposure as the environmental factor. In the first step, we performed a GWAS to identify marginal genetic effects on PFAP and prioritize SNP for G × E interaction analysis. This initial screening identified four genome-wide significant signals and prioritized 7,464 SNP. In the second step, the 7,464 SNP were tested for interactions with sun exposure in the PFAP phenotype, resulting in one significant SNP and a second one SNP showing a trend towards association.
The strongest signal from the GWAS on the PFAP phenotype was carried by six SNP in high LD (r2 > 0.8: rs114462394, rs77074982, rs114203715, rs114017876, rs1576144130 and rs114002886), all located on chromosome 3, near or within the CGGBP1 and ZNF654 genes. The CGGBP1 gene encodes a CGG repeat-binding protein involved in DNA damage/repair, telomere metabolism, and mRNA metabolism (Singh and Westermark, 2015). CGGBP1 has been associated with telomere protection, and given the known connection between telomere, cellular senescence, aging, and skin aging, it suggests that CGGBP1 could play a role in facial aging via telomere protection (Singh et al., 2014; Rossiello et al., 2022; Nakamura et al., 2002). Notably, the rs114017876-A allele was associated with a higher PFAP and a lower CGGBP1 expression, suggesting that higher CGGBP1 expression could favor telomere protection and slow down facial aging process. The ZNF654 gene is a protein coding gene predicted to be involved in transcriptional regulation and it is expressed broadly across tissues. It is has not yet been involved in skin aging, but it could influence multiple tissue systems relevant to PFAP, including muscles and connective tissues. The second GWAS signal was carried by the rs567952380 SNP, located on chromosome 9 within the PGM5-AS1 gene, and was associated with the expression of PGM5 in cultured fibroblasts. The PGM5-AS1 gene codes for an antisense RNA of PGM5 while the PGM5 itself gene codes for a component of adherens-type cell-cell and cell-matrix junctions (Belkin et al., 1994). Besides, PGM5 was among the top ten upregulated gene expression between progeroid versus wild-type mice and showed a downregulation of PGM5 protein level ex vivo following the use of marine-derived skin care ingredients (Quintana-Torres et al., 2023; Hameury et al., 2019). Our findings were consistent with these observations, as the rs567952380-C allele was linked to increased PGM5 expression and higher PFAP, suggesting that higher PGM5 expression may contribute the facial aging process. The last significant signal was carried by the rs75470026 SNP located on chromosome 8, within the CSMD1 gene. CSMD1 encodes a large membrane-bound protein and was associated with various biological processes and conditions, including schizophrenia, the complement system, cancer, metastasis and cell migration (Akyuz and Bell, 2022). Several studies have linked CSMD1 to aging and skin-related processes, including associations with telomere length, cognitive performance in the elderly, and hypertrophic scars (Son, Cui, and Xi, 2022; Stepanov et al., 2017; Gu et al., 2021).
For the gene and sun exposure interaction regarding PFAP, we obtained one significant signal, carried by the rs10923186 SNP, located on chromosome 1, near the VANGL1 gene. The VANGL1 gene encodes a protein of the tetraspanins family, which contributes to the structural organization of the plasma membrane (Singethan and Schneider-Schaulies, 2008). Interestingly, VANGL1 was identified as a key gene for cutaneous function and was expressed during keratinization according to the Human Protein Atlas (Uhlén et al., 2015; DiTommaso et al., 2014). The keratinization process seemed relevant for skin aging, since this is the process of keratinocyte cells differentiation essential to constitute the stratum corneum, which forms the skin’s barrier against the external environment (Shetty and Gokul S 2012). In summary, our analysis showed that the rs10923186-T allele was associated with higher PFAP in the less sun exposed population, a reduced methylation level of cg20810993 CpG site and higher expression of VANGL1. Regarding the known impact of UV exposure on DNA methylation in the skin and the association of rs10923186 with methylation level and VANGL1 expression level, the absence of association with PFAP in sun exposed population could be explained by UV-induced methylation changes that could lower VANGL1 expression, thereby diminishing its effect on PFAP (Vandiver et al., 2015). Both our result and the possible role of VANGL1 in skin, underlined the keratinization as a relevant candidate for further investigations to better understand the impact of UV exposure on facial aging. In addition to the first signal, a second SNP, rs10181773, showed a trend toward association exclusively in the low sun-exposed population. This SNP was located on chromosome 2 in the TRAPPC12 gene was involved in eQTL effect with TRAPPC12-AS1 gene notably in skin and skeletal muscle. TRAPPC12 encodes a protein that is a component of the TRAPP complex, which play a role in endoplasmic reticulum to Golgi apparatus trafficking (Scrivens et al., 2011; Milev et al., 2017). It also contributes to chromosome congression, kinetochore assembly and stability (Milev et al., 2015).
The absence of association with major genes identified in previous skin aging GWAS, may reflect the difference in phenotype definitions (Ng and Chew, 2022). Most GWAS have focused on individual features of facial skin aging, such as lentigines, wrinkles, or sagging. While some have explored composite phenotypes, to the best of our knowledge, no one has ever investigated a dynamic facial aging phenotype as PFAP. The mechanisms underlying the perceived facial aging progression may involve different and complementary processes compared to those driving specific features like wrinkles. Indeed, these results may reflect the multifactorial process not limited to skin changes and involving components of face structure such as muscles, fat and ligaments (Swift et al., 2021; Cotofana et al., 2016). Several limitations could also account for these discrepancies with prior studies: the relatively small sample size of 226 individuals, the potential bias introduced by self-reported sun exposure questionnaire and the imputation uncertainty of the SNP identified in the GWAS. Sun-exposure questionnaires, however, have shown good concordance between self-reports and objective measures in several studies (Hillhouse et al., 2012; Jennings et al., 2013; Køster et al., 2017).
This study revealed four relevant genetic associations with perceived facial aging progression and one significant G × E interaction. These findings support the known link between telomere shortening/protection and aging, and suggest a potential role for keratinization in the context of facial aging and sun exposure. Although further validation in larger cohorts is necessary, these insights into the molecular etiology of PFAP through genetic approaches may help identify targeted interventions that could address not only cutaneous signs but also underlying facial structures through personalized approaches, notably in the context of a particular environment (Krutmann et al., 2021). Future studies could include functional assays targeting the identified variants (organotypic skin cultures) and integrate multi-omics data to get deeper into the understanding of the mechanisms at stake. Finally, by using a dynamic, holistic measure like PFAP, future genetic investigations may uncover novel biomarkers and mechanisms relevant for facial aging and aging.
Data availability statement
The data analyzed in this study is subject to the following licenses/restrictions: data privacy. Requests to access these datasets should be directed to Jean-François Zagury,emFndXJ5QGNuYW0uZnI=.
Ethics statement
The studies involving humans were approved by Hospital Medicals Ethics Committee of Paris-Cochin. The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study.
Author contributions
LO: Data curation, Formal analysis, Investigation, Writing – original draft. RM-S: Data curation, Formal analysis, Investigation, Writing – original draft. MR: Formal analysis, Investigation, Writing – review and editing. JN: Formal analysis, Investigation, Writing – review and editing. TL: Data curation, Formal analysis, Writing – review and editing. PG: Resources, Writing – review and editing. J-LS: Resources, Writing – review and editing. GG: Resources, Writing – review and editing. RJ: Resources, Writing – review and editing. SC: Conceptualization, Writing – review and editing. JL: Project administration, Conceptualization, Validation, Writing – review and editing. NA: Validation, Writing – review and editing. J-FZ: Supervision, Validation, Writing – review and editing. SL: Project administration, Conceptualization, Data curation, Formal analysis, Investigation, Writing – original draft, Supervision, Validation.
Funding
The author(s) declare that no financial support was received for the research and/or publication of this article.
Acknowledgments
The authors are grateful to all the individuals and staff who contributed to the collection of the cohorts.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fragi.2025.1519799/full#supplementary-material
Abbreviations
PFAP: perceived facial aging progression.
References
Akyuz, E., and Bell, S. M. (2022). The diverse role of CUB and sushi multiple domains 1 (CSMD1) in human diseases. Genes 13 (12), 2332. doi:10.3390/genes13122332
Avila, F. R., Carter, R. E., McLeod, C. J., Bruce, C. J., Giardi, D., Guliyeva, G., et al. (2023). Perceived age in patients exposed to distinct UV indexes: a systematic review. Indian J. Plastic Surg. 56 (2), 103–111. doi:10.1055/s-0042-1759696
Belkin, A. M., Klimanskaya, I. V., Lukashev, M. E., Lilley, K., Critchley, D. R., and Koteliansky, V. E. (1994). A novel phosphoglucomutase-related protein is concentrated in adherens junctions of muscle and nonmuscle cells. J. Cell Sci. 107 (Pt 1), 159–173. doi:10.1242/jcs.107.1.159
Boyle, A. P., Hong, E. L., Hariharan, M., Cheng, Y., Schaub, M. A., Kasowski, M., et al. (2012). Annotation of functional variation in personal genomes using RegulomeDB. Genome Res. 22 (9), 1790–1797. doi:10.1101/gr.137323.112
Christensen, K., Thinggaard, M., McGue, M., Rexbye, H., Hjelmborg, J. V. B., Aviv, A., et al. (2009). Perceived age as clinically useful biomarker of ageing: cohort study. BMJ Clin. Res. Ed. 339, b5262. doi:10.1136/bmj.b5262
Coleman, S. R., and Rajiv, G. (2006). The anatomy of the aging face: volume loss and changes in 3-dimensional topography. Aesthetic Surg. J. 26 (1S), S4–S9. doi:10.1016/j.asj.2005.09.012
Cotofana, S., Fratila, A. A. M., Schenck, T. L., Redka-Swoboda, W., Zilinsky, I., and Pavicic, T. (2016). The anatomy of the aging face: a review. Facial Plast. Surg. FPS 32 (3), 253–260. doi:10.1055/s-0036-1582234
Delaneau, O., Zagury, J.-F., Robinson, M. R., Marchini, J. L., and Dermitzakis, E. T. (2019). Accurate, scalable and integrative haplotype estimation. Nat. Commun. 10 (1), 5436. doi:10.1038/s41467-019-13225-y
DiTommaso, T., Jones, L. K., Cottle, D. L., WTSI Mouse Genetics ProgramGerdin, A. K., Vancollie, V. E., et al. (2014). Identification of genes important for cutaneous function revealed by a large scale reverse genetic screen in the mouse. PLoS Genet. 10 (10), e1004705. doi:10.1371/journal.pgen.1004705
Folkersen, L., Gustafsson, S., Wang, Q., Hansen, D. H., Hedman, Å. K., Schork, A., et al. (2020). Genomic and drug target evaluation of 90 cardiovascular proteins in 30,931 individuals. Nat. Metab. 2 (10), 1135–1148. doi:10.1038/s42255-020-00287-2
Galan, P., Yoon, H. C., Preziosi, P., Viteri, F., Valeix, P., Fieux, B., et al. (1998). Determining factors in the iron status of adult women in the SU.VI.MAX study. SUpplementation en VItamines et Minéraux AntioXydants. Eur. J. Clin. Nutr. 52 (6), 383–388. doi:10.1038/sj.ejcn.1600561
Gauderman, W. J., Mukherjee, B., Aschard, H., Hsu, L., Lewinger, J. P., Patel, C. J., et al. (2017). Update on the state of the science for analytical methods for gene-environment interactions. Am. J. Epidemiol. 186 (7), 762–770. doi:10.1093/aje/kwx228
Gaunt, T. R., Shihab, H. A., Hemani, G., Min, J. L., Woodward, G., Lyttleton, O., et al. (2016). Systematic identification of genetic influences on methylation across the human life course. Genome Biol. 17, 61. doi:10.1186/s13059-016-0926-z
Gu, S., Huang, X., Xu, X., Liu, Y., Khoong, Y., Zhang, Z., et al. (2021). Inhibition of CUB and sushi multiple domains 1 (CSMD1) expression by miRNA-190a-3p enhances hypertrophic scar-derived fibroblast migration in vitro. BMC Genomics 22 (1), 613. doi:10.1186/s12864-021-07920-8
Guinot, C., Malvy, D., and Latreille, J. (2001). “Sun exposure behaviour of a general adult population in France,” in Skin and environment - perception and protection. Editors J. Ring, S. Weidinger, and U. Darsow (Bologna: Monduzzi), 1099–1106.
Gunn, D. A., Rexbye, H., Griffiths, C. E. M., Murray, P. G., Fereday, A., Catt, S. D., et al. (2009). Why some women look young for their age. PloS One 4 (12), e8021. doi:10.1371/journal.pone.0008021
Hameury, S., Borderie, L., Monneuse, J.-M., Skorski, G., and Pradines, D. (2019). Prediction of skin anti-aging clinical benefits of an association of ingredients from marine and maritime origins: ex vivo evaluation using a label-free quantitative proteomic and customized data processing approach. J. Cosmet. Dermatology 18 (1), 355–370. doi:10.1111/jocd.12528
Hercberg, S., Galan, P., Preziosi, P., Bertrais, S., Mennen, L., Malvy, D., et al. (2004). The SU.VI.MAX study: a randomized, placebo-controlled trial of the health effects of antioxidant vitamins and minerals. Archives Intern. Med. 164 (21), 2335–2342. doi:10.1001/archinte.164.21.2335
Herrera-Luis, E., Kelly, B., Volk, H., Ladd-Acosta, C., and Wojcik, G. L. (2024). Gene-environment interactions in human health. Nat. Rev. Genet. 25 (May), 768–784. doi:10.1038/s41576-024-00731-z
Hillhouse, J., Turrisi, R., Jaccard, J., and Robinson, J. (2012). Accuracy of self-reported sun exposure and sun protection behavior. Prev. Sci. Official J. Soc. Prev. Res. 13 (5), 519–531. doi:10.1007/s11121-012-0278-1
Howie, B., Fuchsberger, C., Stephens, M., Marchini, J., and Abecasis, G. R. (2012). Fast and accurate genotype imputation in genome-wide association studies through pre-phasing. Nat. Genet. 44 (8), 955–959. doi:10.1038/ng.2354
Jennings, L., Karia, P. S., Jambusaria-Pahlajani, A., Whalen, F. M., and Schmults, C. D. (2013). The sun exposure and behaviour inventory (SEBI): validation of an instrument to assess sun exposure and sun protective practices. J. Eur. Acad. Dermatology Venereol. JEADV 27 (6), 706–715. doi:10.1111/j.1468-3083.2012.4541.x
Køster, B., Søndergaard, J., Nielsen, J. B., Allen, M., Olsen, A., and Bentzen, J. (2017). The validated sun exposure questionnaire: association of objective and subjective measures of sun exposure in a Danish population-based sample. Br. J. Dermatology 176 (2), 446–456. doi:10.1111/bjd.14861
Krutmann, J., Bouloc, A., Sore, G., Bernard, B. A., and Passeron, T. (2017). The skin aging exposome. J. Dermatological Sci. 85 (3), 152–161. doi:10.1016/j.jdermsci.2016.09.015
Krutmann, J., Schikowski, T., Morita, A., and Berneburg, M. (2021). Environmentally-induced (extrinsic) skin aging: exposomal factors and underlying mechanisms. J. Investigative Dermatology 141 (4S), 1096–1103. doi:10.1016/j.jid.2020.12.011
Le Clerc, S., Taing, L., Ezzedine, K., Latreille, J., Delaneau, O., Labib, T., et al. (2013). A genome-wide association study in caucasian women points out a putative role of the STXBP5L gene in facial photoaging. J. Investigative Dermatology 133 (4), 929–935. doi:10.1038/jid.2012.458
Machiela, M. J., and Chanock, S. J. (2015). LDlink: a web-based application for exploring population-specific haplotype structure and linking correlated alleles of possible functional variants. Bioinforma. Oxf. Engl. 31 (21), 3555–3557. doi:10.1093/bioinformatics/btv402
Marchini, J., Howie, B., Myers, S., McVean, G., and Donnelly, P. (2007). A new multipoint method for genome-wide association studies by imputation of genotypes. Nat. Genet. 39 (7), 906–913. doi:10.1038/ng2088
Merinville, E., Grennan, G. Z., Gillbro, J. M., Mathieu, J., and Mavon, A. (2015). Influence of facial skin ageing characteristics on the perceived age in a Russian female population. Int. J. Cosmet. Sci. 37 (Suppl. 1), 3–8. doi:10.1111/ics.12252
Milev, M. P., Grout, M. E., Saint-Dic, D., Cheng, Y.-H. H., Glass, I. A., Hale, C. J., et al. (2017). Mutations in TRAPPC12 manifest in progressive childhood encephalopathy and Golgi dysfunction. Am. J. Hum. Genet. 101 (2), 291–299. doi:10.1016/j.ajhg.2017.07.006
Milev, M. P., Hasaj, B., Saint-Dic, D., Snounou, S., Zhao, Q., and Sacher, M. (2015). TRAMM/TrappC12 plays a role in chromosome congression, kinetochore stability, and CENP-E recruitment. J. Cell Biol. 209 (2), 221–234. doi:10.1083/jcb.201501090
Nakamura, K.-I., Izumiyama-Shimomura, N., Sawabe, M., Arai, T., Aoyagi, Y., Fujiwara, M., et al. (2002). Comparative analysis of telomere lengths and erosion with age in human epidermis and lingual epithelium. J. Investigative Dermatology 119 (5), 1014–1019. doi:10.1046/j.1523-1747.2002.19523.x
Ng, J. Y., and Chew, F. T. (2022). A systematic review of skin ageing genes: gene pleiotropy and genes on the chromosomal band 16q24.3 may drive skin ageing. Sci. Rep. 12 (1), 13099. doi:10.1038/s41598-022-17443-1
Nkengne, A., Bertin, C., Stamatas, G. N., Giron, A., Rossi, A., Issachar, N., et al. (2008). Influence of facial skin attributes on the perceived age of caucasian women. J. Eur. Acad. Dermatology Venereol. JEADV 22 (8), 982–991. doi:10.1111/j.1468-3083.2008.02698.x
Price, A. L., Patterson, N. J., Plenge, R. M., Weinblatt, M. E., Shadick, N. A., and Reich, D. (2006). Principal components analysis corrects for stratification in genome-wide association studies. Nat. Genet. 38 (8), 904–909. doi:10.1038/ng1847
Project Consortium, G., Abecasis, G. R., Altshuler, D., Auton, A., Brooks, L. D., Durbin, R. M., et al. (2010). A map of human genome variation from population-scale sequencing. Nature 467 (7319), 1061–1073. doi:10.1038/nature09534
Quintana-Torres, D., Valle-Cao, A., Bousquets-Muñoz, P., Freitas-Rodríguez, S., Rodríguez, F., Lucia, A., et al. (2023). The secretome Atlas of two mouse models of progeria. Aging Cell 22 (10), e13952. doi:10.1111/acel.13952
R Core Team (2024). A langage and environment for statistical computing. Vienna, Austria: R foundation for Statistical Computing. Available online at: https://www.R-project.org/.
Rippon, I., and Steptoe, A. (2015). Feeling old vs being old: associations between self-perceived age and mortality. JAMA Intern. Med. 175 (2), 307–309. doi:10.1001/jamainternmed.2014.6580
Roberts, V., Main, B., Timpson, N. J., and Haworth, S. (2020). Genome-wide association study identifies genetic associations with perceived age. J. Investigative Dermatology 140 (12), 2380–2385. doi:10.1016/j.jid.2020.03.970
Rossiello, F., Jurk, D., and Passos, J. F.Fabrizio d’Adda di Fagagna (2022). Telomere dysfunction in ageing and age-related diseases. Nat. Cell Biol. 24 (2), 135–147. doi:10.1038/s41556-022-00842-x
Scrivens, P. J., Noueihed, B., Shahrzad, N., Hul, S., Brunet, S., and Sacher, M. (2011). C4orf41 and TTC-15 are mammalian TRAPP components with a role at an early stage in ER-to-golgi trafficking. Mol. Biol. Cell 22 (12), 2083–2093. doi:10.1091/mbc.E10-11-0873
Shetty, S., and Gokul, S. (2012). Keratinization and its disorders. Oman Med. J. 27 (5), 348–357. doi:10.5001/omj.2012.90
Singethan, K., and Schneider-Schaulies, J. (2008). Tetraspanins: small transmembrane proteins with big impact on membrane microdomain structures. Commun. and Integr. Biol. 1 (1), 11–13. doi:10.4161/cib.1.1.6406
Singh, U., Maturi, V., Jones, R. E., Paulsson, Y., Baird, D. M., and Bengt, W. (2014). CGGBP1 phosphorylation constitutes a telomere-protection signal. Cell CycleGeorget. Tex. 13 (1), 96–105. doi:10.4161/cc.26813
Singh, U., and Westermark, B. (2015). CGGBP1--an indispensable protein with ubiquitous cytoprotective functions. Upsala J. Med. Sci. 120 (4), 219–232. doi:10.3109/03009734.2015.1086451
Sobin, L., Barcus, M., Branton, P. A., Engel, K. B., Keen, J., Tabor, D., et al. (2024). Histologic and quality assessment of genotype-tissue expression (GTEx) research samples: a large postmortem tissue collection. Archives Pathology and Laboratory Med. 149 (May), 217–232. doi:10.5858/arpa.2023-0467-OA
Son, N., Cui, Y., and Wang, X. (2022). Association between telomere length and skin cancer and aging: a mendelian randomization analysis. Front. Genet. 13, 931785. doi:10.3389/fgene.2022.931785
Stepanov, V., Marusin, A., Vagaitseva, K., Bocharova, A., and Makeeva, O. (2017). Genetic variants in CSMD1 gene are associated with cognitive performance in normal elderly population. Genet. Res. Int. 2017, 6293826. doi:10.1155/2017/6293826
Swift, A., Liew, S., Weinkle, S., Garcia, J. K., and Silberberg, M. B. (2021). The facial aging process from the inside out. Aesthetic Surg. J. 41 (10), 1107–1119. doi:10.1093/asj/sjaa339
Taylor, P. J., and Marsh, J. E. (2017). “E -prime (software),” in The international encyclopedia of communication research methods. Editors J. Matthes, C. S. Davis, and R. F. Potter 1st ed. (Wiley), 1–3. doi:10.1002/9781118901731.iecrm0085
Uhlén, M., Fagerberg, L., Hallström, B. M., Lindskog, C., Oksvold, P., Mardinoglu, A., et al. (2015). Proteomics. Tissue-based map of the human proteome. Sci. (New York, N.Y.) 347 (6220), 1260419. doi:10.1126/science.1260419
Vandiver, A. R., Irizarry, R. A., Hansen, K. D., Garza, L. A., Runarsson, A., Li, X., et al. (2015). Age and sun exposure-related widespread genomic blocks of hypomethylation in nonmalignant skin. Genome Biol. 16 (1), 80. doi:10.1186/s13059-015-0644-y
Villicaña, S., Castillo-Fernandez, J., Hannon, E., Christiansen, C., Tsai, P.-C., Maddock, J., et al. (2023). Genetic impacts on DNA methylation help elucidate regulatory genomic processes. Genome Biol. 24 (1), 176. doi:10.1186/s13059-023-03011-x
Voegeli, R., Schoop, R., Prestat-Marquis, E., Rawlings, A. V., Shackelford, T. K., and Fink, B. (2021). Differences between perceived age and chronological age in women: a multi-ethnic and multi-centre study. Int. J. Cosmet. Sci. 43 (5), 547–560. doi:10.1111/ics.12727
Keywords: G×E interaction, perceived age, facial aging, SNP, sun exposure, GWAS
Citation: Obry L, Medina-Santos R, Rahmouni M, Noirel J, Labib T, Galan P, Spadoni J-L, Gendronneau G, Jdid R, Courrèges S, Latreille J, André N, Zagury J-F and Le Clerc S (2025) Identification of gene–sun exposure interactions of GWAS-identified variants in perceived facial aging progression. Front. Aging 6:1519799. doi: 10.3389/fragi.2025.1519799
Received: 30 October 2024; Accepted: 26 June 2025;
Published: 23 July 2025.
Edited by:
Simon Galas, Université de Montpellier, FranceReviewed by:
Myriam Richaud, UMR5247 Institut des Biomolécules Max Mousseron (IBMM), FrancePiercarlo Minoretti, Studio Minoretti, Italy
Copyright © 2025 Obry, Medina-Santos, Rahmouni, Noirel, Labib, Galan, Spadoni, Gendronneau, Jdid, Courrèges, Latreille, André, Zagury and Le Clerc. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jean-François Zagury, emFndXJ5QGNuYW0uZnI=; Sigrid Le Clerc, c2lncmlkLmxlX2NsZXJjQGNuYW0uZnI=
†These authors have contributed equally to this work and share first authorship