 Rahul Debnath1,2
Rahul Debnath1,2 Justin George1*
Justin George1* Ammar B. Bhandari3
Ammar B. Bhandari3 Rupesh Kariyat2
Rupesh Kariyat2 Gadi V. P. Reddy1
Gadi V. P. Reddy1 James P. Glover1
James P. Glover1 Tulsi P. Kharel3
Tulsi P. Kharel3 Krishna N. Reddy3
Krishna N. Reddy3- 1Southern Insect Management Research Unit, USDA-ARS, Stoneville, MS, United States
- 2Department of Entomology and Plant Pathology, University of Arkansas, Fayetteville, AR, United States
- 3Crop Production Systems Research Unit, USDA-ARS, Stoneville, MS, United States
Soybean production is substantially affected by insect pests, necessitating the implementation of appropriate management strategies. Effective irrigation and fertilizer regimes have been found to affect pest populations in agroecosystems. However, their interactive effects on herbivore incidence are less understood. This study reports the abundance and diversity of insect pests, especially hemipteran, lepidopteran, and stink bug species complexes, encountered on soybeans throughout their vegetative and reproductive growth stages under varying irrigation and micronutrient treatments. First, we compared total and major insect pest population densities in irrigated and rainfed conditions. Afterwards, we evaluated how the frequency of insect pests on soybean was influenced by the foliar application of various micronutrients (Fe, Zn, and Fe + Zn) compared to the control plots. This field experiment was repeated over two growing seasons, during 2023 and 2024. Results showed that total insect pest populations were significantly higher (p<0.001) in control plots than in plots treated with the combined micronutrients (Fe + Zn). However, compared to rainfed treatment, irrigated soybean attracted significantly more insect herbivores. Furthermore, micronutrients had no significant impact on reducing pest pressure when applied in conjunction with irrigation. These findings indicated that micronutrients and irrigation can have complex effects on plant-insect interactions, and the specific effects may depend on particular nutrients, crop species, and water availability. These findings also provide evidence that combining foliar applications of ferrous and zinc in soybean fields can efficiently decrease insect pest populations independent of irrigation.
1 Introduction
Soybean (Glycine max (L.) Merr.), is the sixth most commonly grown agricultural crop that generates around 53% of the world’s seed oil production (Beta and Isaak, 2016). Globally, Brazil, United States, and Argentina are the top three producers. In 2023-24, the United States produced approximately 113.27 million metric tons of soybeans, which accounted for 29% of global soybean production (USDA-FAS Production-Soybeans). However, in 2023, damage by invertebrate pests resulted in a loss of 1.6% of soybean bushels across 19 states. The total management costs were estimated to be 669.3 million USD, which also accounts for non-targeted insecticide applications (Soybean Invertebrate Loss Estimates from the United States, 2023). Due to their negative impact on crop yield through extensive defoliation, insect pests are a major concern in row crop production, and soybean production. The severity of insect pest damage depends upon the specific type of insect, their growth stage, and the crop being cultivated (Barathi et al., 2024). While numerous strategies are employed to control insect pests, chemical insecticides are the most widely adopted approach. However, the unregulated application of insecticides results in a considerable ecological footprint of pesticides on the environment, leading to the development of insect pest resistance and contamination of non-target ecosystems, including soil and water (Ren et al., 2023).
Globally, soybean production is threatened by diverse leaf-feeding and pod- damaging pests. Early monitoring of insect pest incidence is crucial for preventing severe damage; also, the use of appropriate management strategies is critical for yield. In the United States, nearly 700 insect pest species have been documented that attack soybean (Gautam et al., 2024; Greene et al., 2021). Among these, approximately 20 species or species complexes are recognized as significant economic pests (Steffy, 2015). Furthermore, various widely distributed insect pests may have considerable economic impact in certain soybean production regions while being minor in other systems. This suggests that there are additional, unidentified factors that affect the status of these pests (Steffy, 2015). The increasing impact of climate change is causing a growing concern about the adaptation of herbivore pest populations and their resistance to insecticides. Ultimately, how genetic changes shape the diversity and distribution of herbivore pest dynamics is not fully understood. Therefore, incorporating efficient agronomic practices could be a more sustainable solution for soybean pest management.
Identifying the pest’s status and population dynamics is crucial in any pest management program. Once the exact pest species and phenology are understood, management efforts can focus on species specific recommendations. Proper knowledge of the infestation and population dynamics of key insect pests facilitates the use of more targeted control methods. Lepidopteran insect species are one of the most critical pest groups in soybean (Murúa et al., 2018). Currently, Bt soybeans and foliar insecticides are used for control of the primary lepidopteran pests such as Anticarsia gemmatalis (Hübner) (Lep: Erebidae), Chrysodeixis includens (Walker) (Lep: Noctuidae), Hypena scabra (Fabricius) (Lep: Erebidae), and Spodoptera sp (McPherson and MacRae, 2009; Yu et al., 2013; Bernardi et al., 2014; Debnath et al., 2024). The soybean looper (Chrysodeixis includens Walker) has been reported as a significant pest of soybean in almost all states in the United States (Huseth et al., 2021; Ayala et al., 2024). In addition, Threecornered alfalfa hopper (TCA), Spissistilus festinus (Say) (Hemiptera: Membracidae), is also considered a pest of soybeans that causes injury and yield loss in certain scenarios. Stink bug complex (i.e., Green stink bug, Brown stink bug, Redbanded stink bug, and Southern green stink bug) (Hemiptera: Pentatomidae) are usually the late-season insect pests affecting soybean production in the United States. They utilize their piercing and sucking mouthparts to puncture plant tissues and extract fluids, which ultimately reduces both the yield and quality of the crops. Clearly, soybean hosts a wide range of insect pests that vary in their feeding guild, spatiotemporal dynamics, and taxonomic status.
Rainfall remarkably influences more than 90% of United States soybean production, resulting in substantial yield fluctuations between wet and dry years (NASS, 2013). Irrigation can significantly enhance soybean yields and profitability, particularly in soils that are prone to water deficiency (Karam et al., 2005). The reproductive stage of soybean is most vulnerable to yield reduction due to water deficiency, whereas water scarcity at the flowering stage has a comparatively lesser impact on yield (Andrade, 1995; Lich et al., 2013). Studies from other systems show that irrigation can significantly impact pest incidence. For example, in tomato, Colella et al. (2014) recorded that the population abundance of glasshouse whitefly, potato aphids, and leafhoppers was higher in the irrigated plots when compared to control. Similarly, the mustard aphid population increased significantly in the Chinese cabbage field after increased irrigation (Dhaliwal and Arora, 2002). In addition, soil moisture levels can have a direct impact on the water content of insects, subsequently influencing their physiological processes and population dynamics (Chang et al., 2008). Taken together, the dynamics of insect pest populations in an environment are determined by the physiological, morphological, and behavioral adaptations of insects in response to abiotic and biotic stressors (Djaman et al., 2019).
In addition to irrigation, fertilizer application is a major management practice that significantly affects crop nutrition and consequently, their growth, yield, and defenses. The impact of irrigation and nutrient management differs across various crop systems (Bala et al., 2018). Interestingly, insect pest preferences also can be influenced by the type and quantity of micronutrients used (Kazemi-Dinan et al., 2015; Dueli et al., 2021). Secondary nutrients such as calcium and micronutrients like zinc and ferrous enhance plant resistance against insect pests, reducing pest populations, and they can be efficiently integrated with other pest management practices (Bala et al., 2018). More importantly, a recent meta-analysis examining the nutrient enrichment effects of nitrogen (N), phosphorus (P), and (N + P) reported a significant negative impact of N enrichment and (N + P) on arthropod abundance (Nessel et al., 2023). Although the effects of fertilizer application and irrigation on pest incidence are broadly reported (Zaffaroni et al., 2020), to date, no studies are available on the impacts of the interaction between irrigation and micronutrients on insect pests in soybean. Identifying the prevalent insect pest populations is key to the selection of effective pesticides and their application rates, in addition to optimizing pest control strategies in accordance with economic thresholds.
The primary objectives of our study were: (i) to construct a comprehensive database of insect pests that attack soybean in the southern United States, (ii) to determine the pest population fluctuations during soybean growth stages, (iii) to examine the impact of irrigation on insect pest population dynamics, (iv) to explore the effect of different micronutrients (Fe, Zn, and a combination of Fe and Zn) on total and significant insect pests, and (v) to assess the possible interactive effects of micronutrients and irrigation on soybean pests under field conditions.
2 Materials and methods
2.1 Experimental site and layout
The experiments were conducted for two consecutive cropping seasons during the summers of 2023 and 2024 at the USDA-ARS farm in Stoneville, Mississippi, United States, situated at a latitude of 33° 26’57” N, and a longitude of 90° 52’08” W (Figure 1). The experiment design was a split-plot design with four replications, where irrigation was the whole plot treatment and micronutrient fertilizer treatments were subplots. The size of each plot was 60.96 m long and rows 7.73 m wide. Irrigation and micronutrient treatments were replicated four times, resulting in a total of 32 plots (Figure 2).
Figure 1. A map showing the experiment site in Mississippi, United States (A, B) and field picture of experiment location (C).
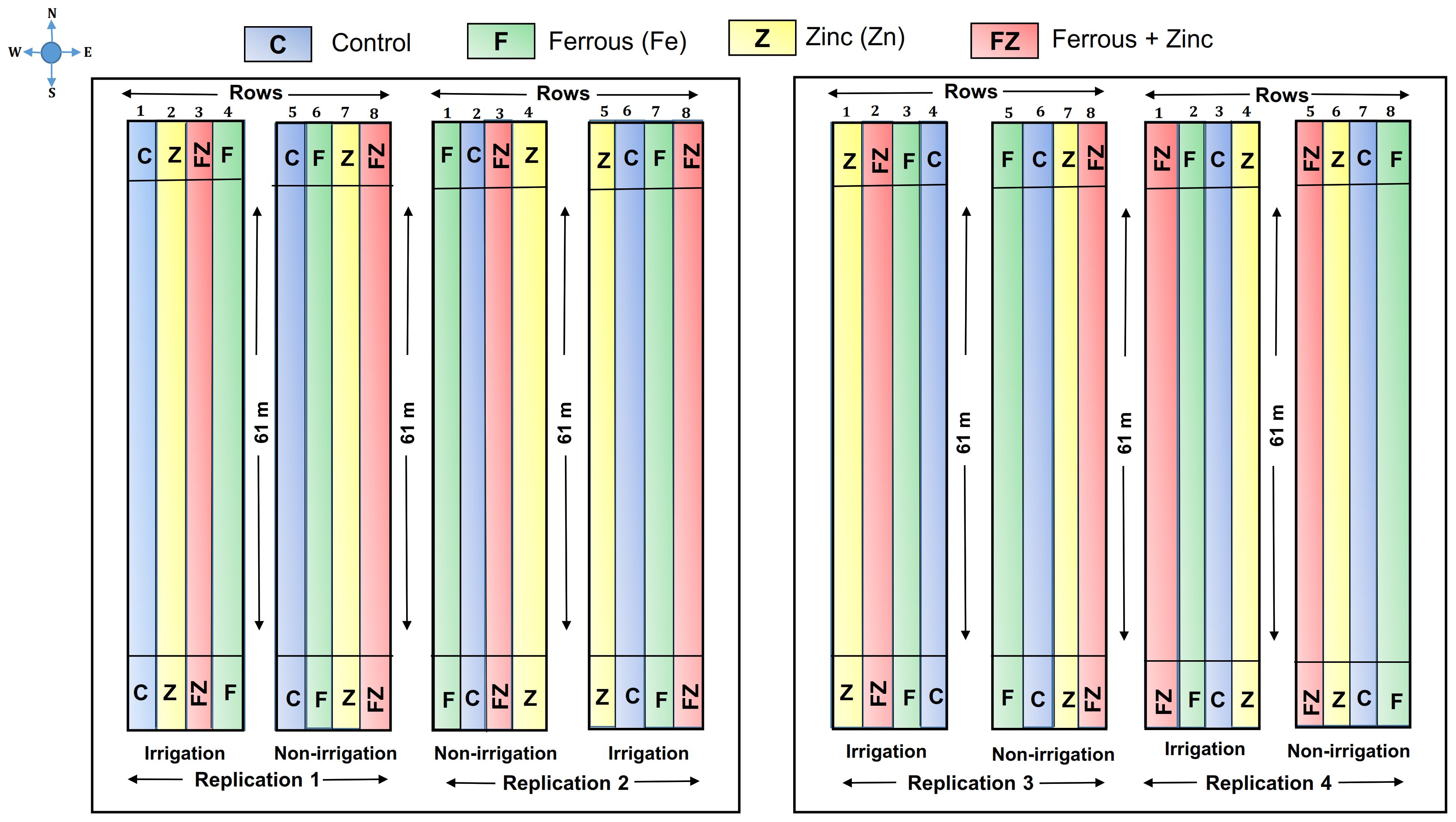
Figure 2. A schematic diagram of experimental field layout, showing four sub-plots together representing one whole plot. Irrigation treatment was applied to the whole plot, and different colors were used for sub-plots indicating particular micro-nutrient treatments applied to that plot.
2.2 Treatments
Irrigation treatments were (1) Rainfed-fully dependent on precipitation and (2) Furrow irrigation- irrigated five times in 2023 (6/8/2023, 6/27/2023, 7/27/2023, 8/4/2023, 8/23/2023) and three times in 2024 (6/24/2024, 7/9/2024, 7/31/2024) depending on amount of rainfall received. The micronutrient treatments were foliar application of (1) zinc (Zn), (2) ferrous (Fe), and (3) Zn + Fe, and (4) no micronutrient control. Soybean variety 48XFO was planted on April 18, 2023, and April 26, 2024. The Fe was applied at 2.80 kg ha-1 and Zn at 5.60 kg ha-1,respectively. The micronutrients (Zn and Fe) were foliar applied 48 days after planting (DAP) in both the 2023 and 2024 seasons.
2.3 Insect sampling
Over the two years, weekly insect collections (for 12 weeks) were made using sweep nets during the vegetative and reproductive (V3-R7) phase of soybean growth. A total of 7.62 m length (25 sweeps) was sampled in the eight-row plots. Four sweep net sampling were conducted for each treatment/week. The sweep samples were collected in a foldable mesh cage (30 x 30 x 30 cm) and then kept at -20°C before being counted. Both quantitative and qualitative analyses of the insects and their developmental stages were carried out, and insect identification and quantification were performed in the laboratory by using a stereo zoom microscope (Leica S6E, Leica, United States).
2.4 Statistical analysis
Quantitative differences in the total and significant pest population of control and micronutrient treatment were compared using one-way analysis of variance (ANOVA) (SPSS software, version 25.0). A Tukey’s HSD mean comparison test was performed when significant differences were found following ANOVA. Student’s t-test was used to analyze the population abundance of insect pests between irrigated and rainfed conditions (SPSS software, version 25.0) (Zar, 1999). A principal component analysis (PCA) was conducted to show the effect of different micronutrients on irrigation under rainfed and irrigated conditions as variables (XLSTAT version 13). Shannon-Wiener Index (H’) and Simpson’s diversity index (1-D) were used to estimate the diversity of different insect pest species/complexes. Individual effects and interaction of irrigation and micronutrients were analyzed using a two-way ANOVA (SPSS), and Sigma Plot 14.0 (Sigma Plot Software, Palo Alto, California, USA) was used to graph the plots.
The Shannon-wiener index (H’) is defined as: H=- Σ Pi In Pi
Where, Pi=S/N
S=number of individuals of one species; N=total number of all individuals in the sample
In=logarithm to base e
Simpson’s Index is expressed as:
Where, n=the total number of organisms of a particular family
N=the total number of organisms of all families
3 Results
3.1 Composition and abundance of insect pests in soybean field
A total of 13,995 insect pests belonging to five families and more than 34 different species, were collected during two growing season (as listed in Table 1). In the 2023 growing season, the total number of insects collected was 7549; among these, highest proportion was of hemipteran insects (58.54%), and the dominant hemipteran species was threecornered alfalfa hopper (TCA), Spissistilus festinus (Say) (36.1%) in 2023, and (31.55%) in 2024. The proportion of lepidopteran species complex was 6.13% in 2023 and 6.49% in 2024 of the total insect samples. Stink bug complex (i.e., Green stink bug, Brown stink bug, Redbanded stink bug, and Southern green stink bug) accounted for 1.6% in 2023 and 1.8% in 2024. Meanwhile, the remaining species in the 2023 season accounted for 56.17% in 2023 and 60.16% in 2024 out of the total insect samples collected.
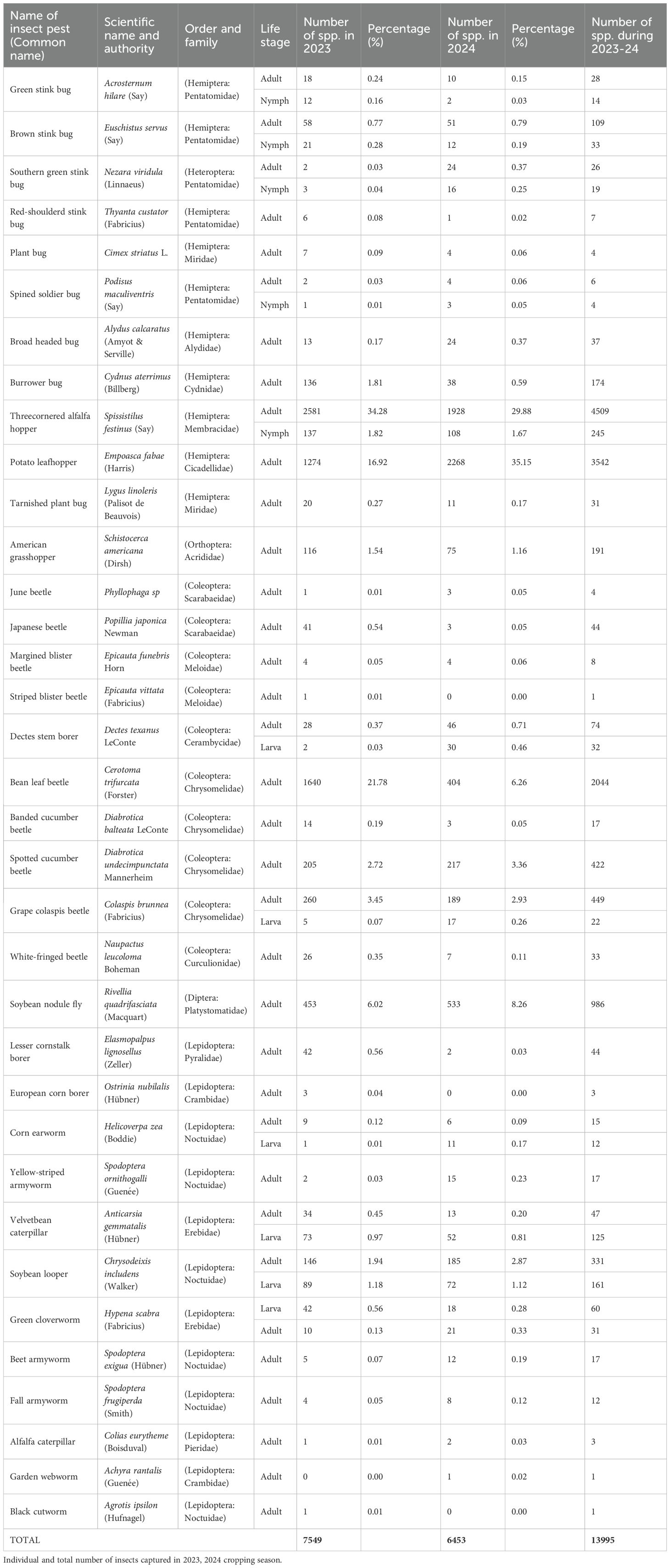
Table 1. Number and types of insect pest collected from established soybean plot during the period of 2023-2024.
3.2 Diversity of insect group/complex
In our study, we found that the lepidopteran species complex is the most diverse insect family (Shannon-Wiener Index: H’=1.40), followed by the coleopteran (H’=1.11), and stink bug complex. In terms of species evenness, species belonging to the stink bug complex were more abundant in between themselves, and the least relative abundance was found in the hemipteran group (Table 2). Simpson’s diversity index supported the Shannon index in indicating the diversity of insects. Simpson’s diversity index also indicated higher diversity in the lepidopteran group and lower diversity in the dipteran and orthopteran groups. A detailed description of diversity indices can be found in Table 2.
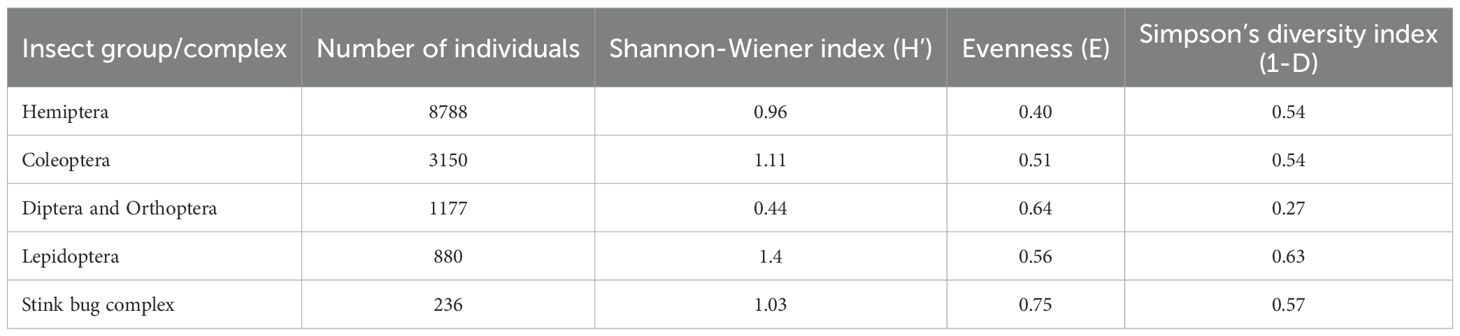
Table 2. Diversity of different insect orders caught during field collection.
3.3 Variation in the abundance of total and major insect species across soybean growth stages
The variation in abundance of the total insect species and TCA, lepidopteran, and stink bug complex during soybean growing season for the 2023–2024 period is presented in (Figure 3). The highest abundance of insect pest pressure was observed during the R5-R6 growth stages in both years. As the reproductive stage progressed, insect populations showed a gradual decline, eventually approaching the population levels observed in the R4-R5 stages of 2023 (Figure 3A). TCA population peaked during the R7-R8 growth stage in 2023. The population density in the R6-R7 stage was the highest. However, it decreased during the R7-R8 growth stage in the 2024 season (Figure 3B). The pressure from lepidopteran pest species increased steadily throughout the study, peaking in the R5-R6 stage for both seasons (Figure 3C). Stink bug infestations were minimal until soybeans reached the R5-R6 stage; after that, the population density peaked in the R7-R8 stage throughout both seasons (Figure 3D).
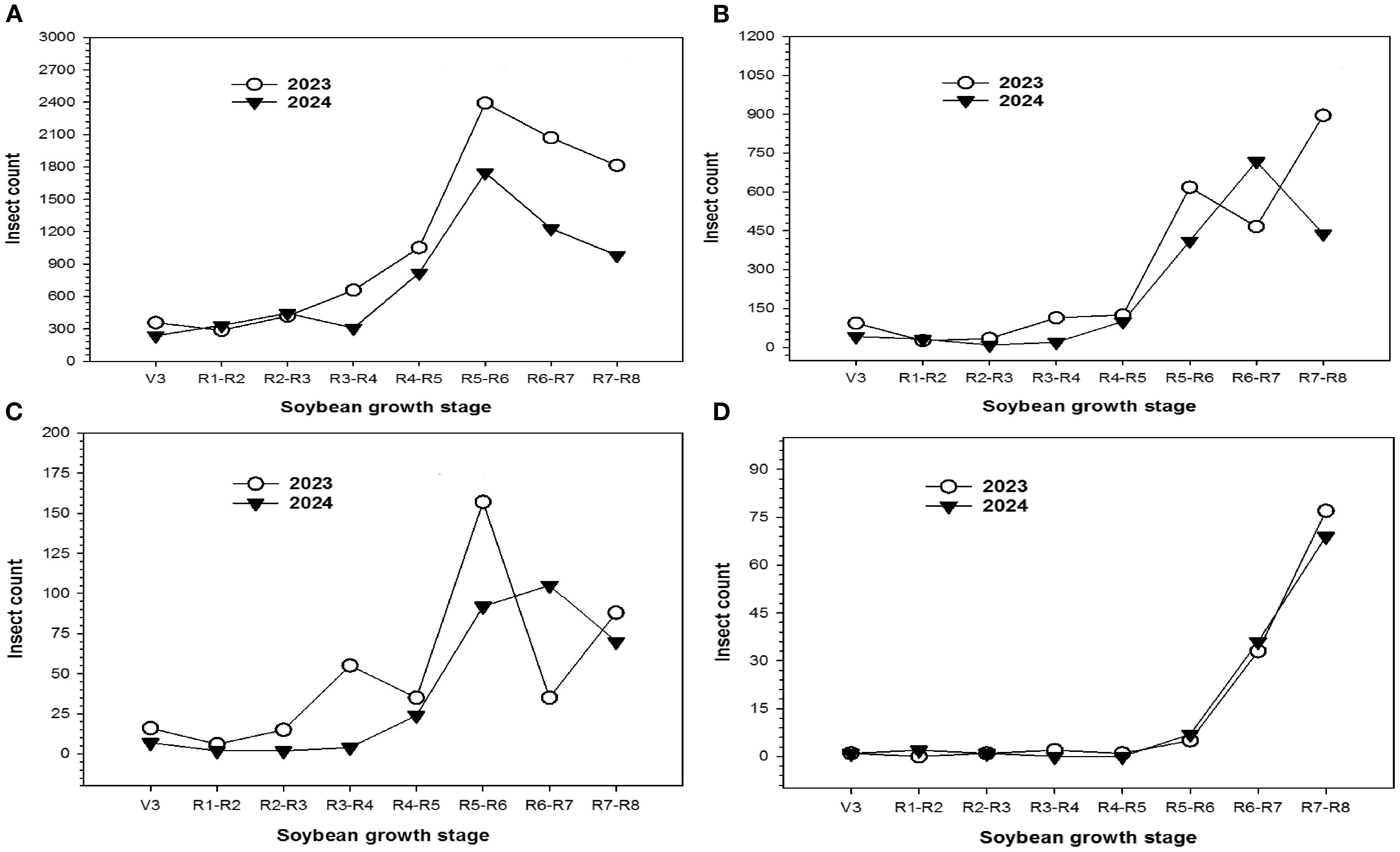
Figure 3. Variation in the population density of (A) total insect pests, (B) Threecornered alfalfa hopper, (C) Lepidopteran complex, and (D) Stink bug complex at different growth stages of soybean during the 2023–2024 crop season.
3.4 Effects of irrigation on insect pest population dynamics
The effect of irrigation on total insect pests and TCA, lepidopteran complex, stink bug complex was identified (Figure 4). During the 2023 growing season, the number of total insect pests was significantly higher in irrigated plots compared to rainfed plots (t=2.994; df =6; P<0.05). Similar results were found during the 2024 growing season as well (t=2.681; df =6; P<0.05) (Figure 4A). TCA population was higher in irrigated plots in 2023 (t=2.973; df =6; P<0.05), and 2024 (t=2.723; df =6; P<0.05) compared to rainfed plots (Figure 4B). Significantly higher number of lepidopteran pests were observed in irrigated plots in 2023 (t=3.609; df =6; P<0.01) and 2024 season (t=2.625; df =6; P<0.05) compared with rainfed plots (Figure 4C). No significant differences were observed between irrigated and rainfed conditions for stink bug complex in 2023, whereas the population was significantly higher in irrigated conditions in 2024 (t=3.441; df=6; P<0.01) compared with rainfed plots (Figure 4D).
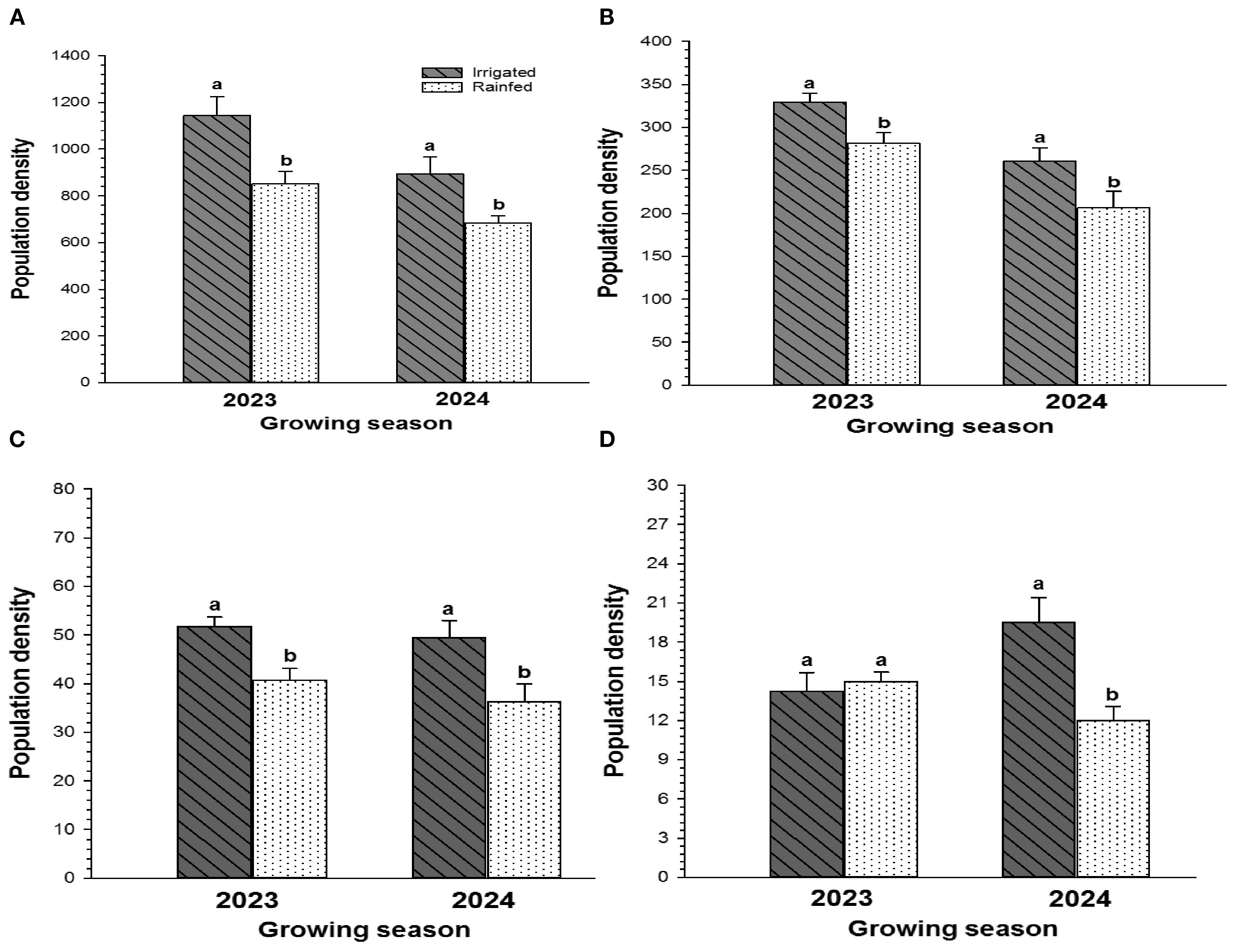
Figure 4. Impact of irrigation and rainfed treatments on (A) Total insect population, (B) Threecornered alfalfa hopper, (C) Lepidopteran complex, and (D) Stink bug complex during 2023 and 2024 cropping seasons. Different letters indicate statistically significant differences followed by (Tukey’s HSD, P<0.05).
3.5 Impact of micronutrients on insect populations
The impact of different micronutrients, ferrous (Fe), zinc (Zn), and their combination (Fe + Zn), was compared with the control plot (i.e., no micronutrients were applied) on the total insect pests and TCA, lepidopteran complex, stink bug complex (Figure 5). Significant differences were observed in the total insect population between control (316.88 ± 18.10) and individual micronutrient Fe (270.13 ± 7.04) and Zn (266.63 ± 9.14) application, and also the combination of both micronutrients (218.75 ± 9.66) in 2023 season (F3,28=11.60, P<0.0001), (Figure 5A). Similar patterns of pest infestation were observed in 2024 growing season (F3,28=12.30, P<0.0001) (Figure 5A). The combination of micronutrients (F3,28=4.802, P<0.008) significantly reduced the TCA population (70.00 ± 3.50) compared to the control plots (90.63 ± 4.38) (Figure 5B). No significant differences were found between control (90.63 ± 4.38) and individual micronutrient Fe (76.50 ± 3.96) and Zn (78.25 ± 3.85) in 2023 growing season. In 2024, significantly higher TCA population density was found in the control plot (70.63 ± 7.67), intermediate in individual treatment Fe (54.75 ± 4.59), Zn (52.88 ± 3.89), and least in Fe + Zn (48.13 ± 3.80) (F3,28=3.48, P<0.029) (Figure 5B).
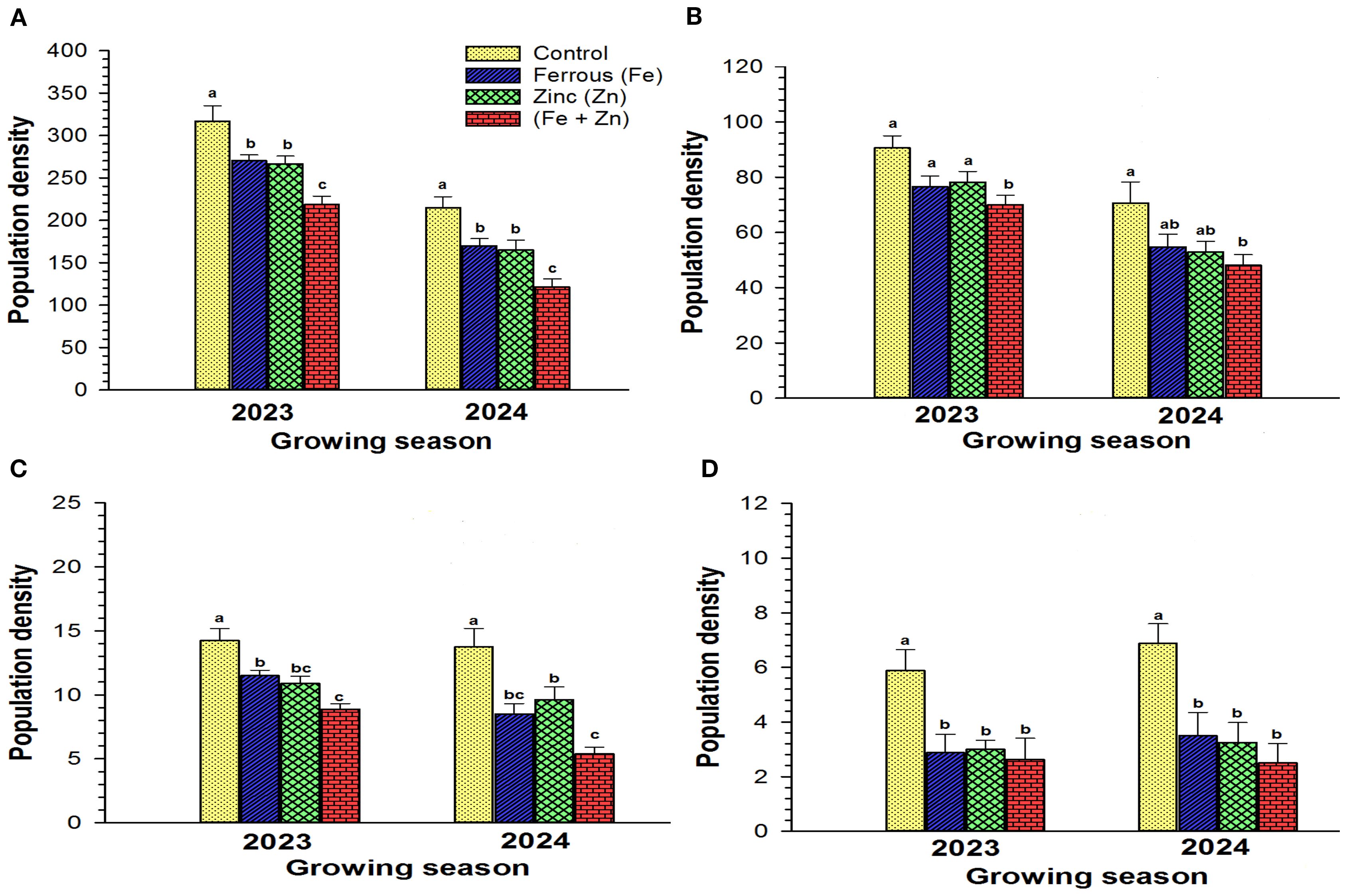
Figure 5. Effect of different micronutrients on (A) Total insect population, (B) Threecornered alfalfa hopper, (C) Lepidopteran complex, and (D) Stink bug complex on different crop season. Different letters indicate significant differences among micronutrient treatments determined by post hoc analyses using Tukey’s HSD (P<0.05).
Lepidopteran population was significantly higher in control (14.25 ± 0.94) plots compared to individual micronutrient Fe (11.50 ± 0.42), Zn (10.88 ± 0.58) and combined micronutrient (8.88 ± 0.44) treatments (F3,28=12.37, P<0.0001). No significant differences were observed between Zn and combined micronutrients (Figure 5C) in the 2023 growing season. Although the same pattern was observed as combined micronutrients significantly reduced pest population densities (F3,28 = 11.81, P<0.0001), there were no significant differences between Fe (8.50 ± 0.82) and combination of micronutrients (5.38 ± 0.53) (Figure 5C) in 2024. Stink bugs population dynamics were significantly (F3,28=5.36, P<0.005) (Figure 5D) affected by combined (2.63 ± 0.78) and individual micronutrient treatment Fe (2.88 ± 0.67) and Zn (3.00 ± 0.33) compared to control (5.88 ± 0.77). However, there were no significant differences in stink bug populations between individual and combined micronutrient applications during the 2023 growing season. Similarly, 2024 exhibited the same pattern (F3,28=6.69, P<0.002) (Figure 5D).
3.6 Interactive effects of irrigation and micronutrients
The treatment effects and interaction of irrigation and different micronutrients, ferrous (Fe), zinc (Zn), and their combination (Fe + Zn) on the total insect pest population, TCA, lepidopteran complex, and stink bug complex were explored (Table 3). A significantly higher number of total insects were found in irrigated plots (F=13.852, P<0.0001) than rainfed plots, whereas nutrient-treated plots contained a comparatively lower number of total insects (F=4.452, P<0.013) than control. Meanwhile, no significant differences were observed in plots that contained irrigation as well as micronutrients (F=2.242, P<0.109). For, TCA populations, the numbers were significantly higher in irrigated (F=12.809, P<0.002) than in rainfed treatments. Micronutrient treated plots had comparatively lower insect abundance (F=7.430, P<0.001). Meanwhile, no significant differences were observed in plots that contained irrigation as well as micronutrients (F=2.053, P<0.133) (Table 3). Significantly higher lepidopteran population was found in irrigated plots (F=27.042, P<0.0001) compared with rainfed treatments, and significantly lower lepidopteran numbers were found in micronutrient treatment (F=19.620, P<0.0001). Also, when micronutrients and irrigation were applied together, no significant difference in lepidopteran species complex was found (F=2.807, P<0.061) (Table 3). The population density of stink bugs was significantly lower in nutrient treatment (F=36.467, P<0.0001). No significance differences in species abundance were found when irrigation and micronutrients were applied together in soybean plots (F=2.365, P<0.096) (Table 3). Principal component analysis depicted variation in the total insect species abundance from irrigated and rainfed conditions and combined micronutrients completely apart from control and other individual micronutrients. The first two principal components accounted for a total cumulative variability of 100%, with PC1 explaining 69.76% and PC2 explaining 30.24% of the variation between the irrigated and rainfed conditions (Figure 6).
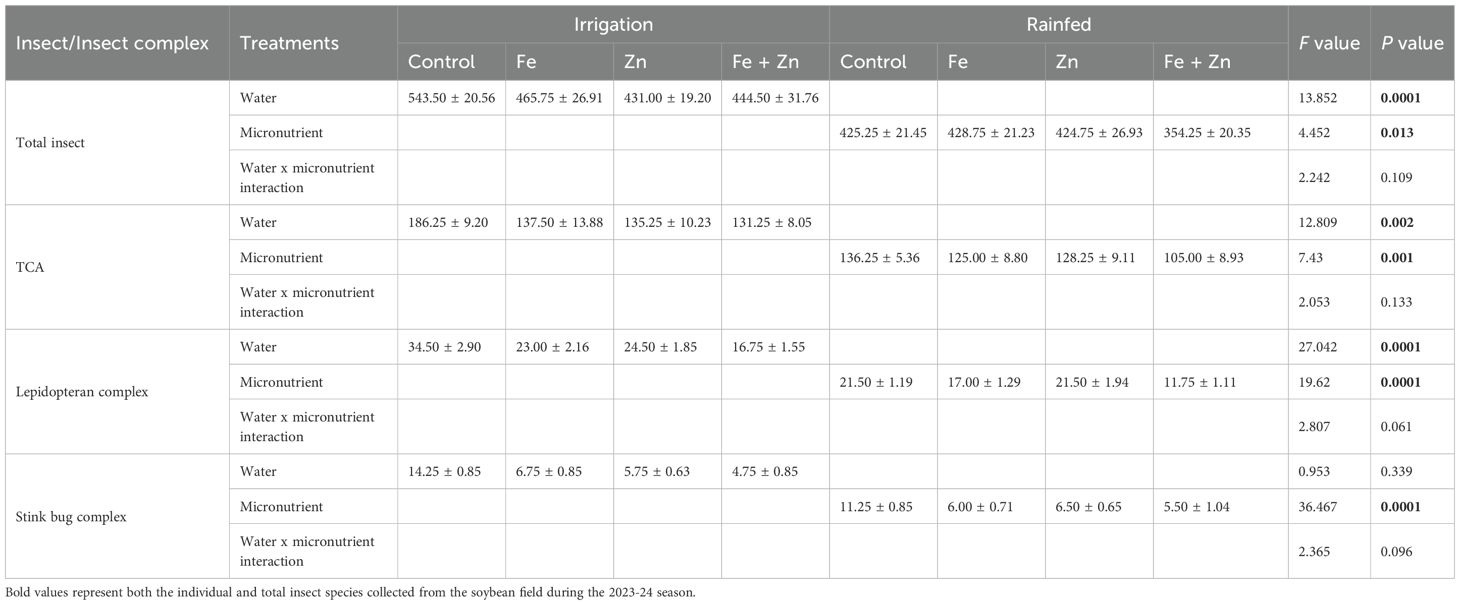
Table 3. Treatment effects and interaction of water and micronutrients on mean ( ± SEM, n =4 for each nutrient) abundance of insect pests in soybean field during the period of 2023-2024.
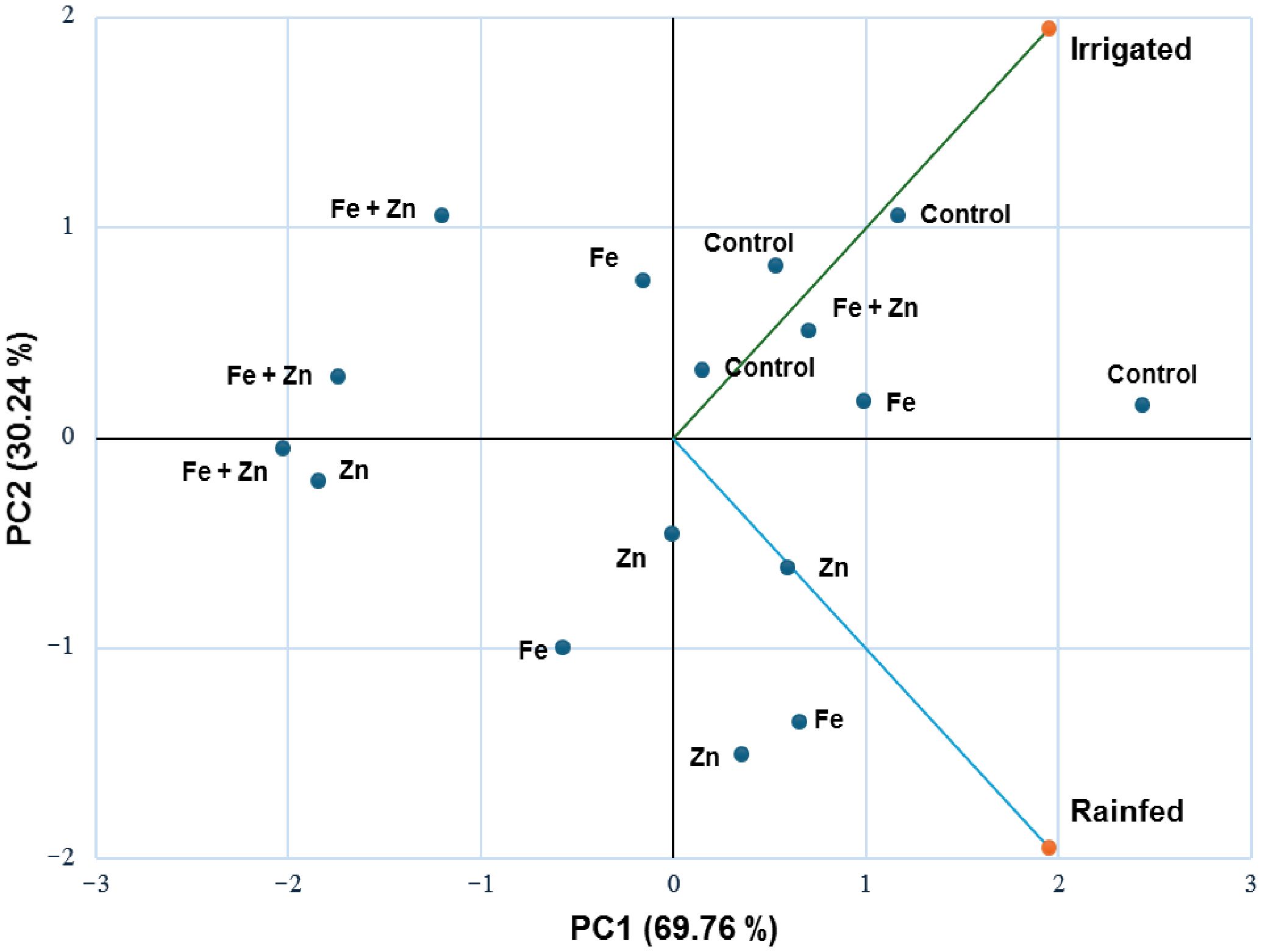
Figure 6. Principal component analysis (PCA) of total insect pest abundance on different micronutrient treatments under irrigated and rainfed conditions. The PCA shows the variation in insect abundance under irrigated and rainfed condition.
4 Discussion
In this study, we examined the role of irrigation and micronutrients in pest incidence in soybeans during the two summer seasons. Our results show that irrigation positively impacts insect pest incidence, but when combined with micronutrients, it leads to a significant reduction in pest numbers. The results from this research collectively suggest that the application of combined micronutrients, even without irrigation, can efficiently decrease insect pest populations in soybeans.
4.1 Soybeans host a wide range of insect pest species
Effective pest management of soybean production requires a thorough understanding of the population dynamics of the major insect pests. A total of nearly 14,000 insects across a variety of families and species were collected in our study, highlighting the complex ecological interactions they have with soybean. We identified the threecornered alfalfa hopper (TCA) as the most abundant hemipteran species in both seasons. TCA has led to substantial yield reductions in soybeans (roughly 25%) and increased control costs in some years in several mid-southern states (Hesler et al., 2018). Beyer et al. (2017) documented that damaging populations of TCA have historically been restricted to the southern United States. Survey results by Huseth et al. (2021) showed that Anticarsia gemmatalis (velvetbean caterpillar) and Chrysodeixis includens (soybean looper) remain widespread soybean pests in southern states. In contrast, our study showed that the lepidopteran pest pressure peaked during the R5 growth stage, and these two species were caught in more than 65% of the total Lepidoptera collections. Pest pressure from the stink bug complex peaked in the R7-R8 stage, which is mainly considered a late-season pest. Musser et al. (2022) reported that stink bugs represented the most expensive insect pest affecting soybean crops in the United States during 2022, considering both yield losses and management expenses. In our survey, based on the diversity index, we found that the lepidopteran species complex has the highest diversity among all other species, and the least diversified insect groups are the diptera and orthoptera order of insects. Although the number of species is highest in Hemiptera, it has low diversity in comparison to other groups.
4.2 Irrigation positively impacts insect pest incidence
The influence of irrigation on insect pest abundance has been found to vary across species. The abundance of phytophagous insects in tomatoes can be reduced by water stress, which also depends on cultivars and other stress levels (Inbar et al., 2001; Rivelli et al., 2013). Raderschall et al. (2021) showed that the abundance of aphids as higher in irrigated plants than in water-stressed plants. Melloul et al. (2024) reported that irrigation negatively affected the abundance of several beneficial arthropods in vineyards. However, after few weeks, direct effects from irrigation seem to disappear (Srikanth et al., 1997). The present study demonstrated that total insect pest population density was significantly higher in irrigated plots in both seasons. We found that irrigated plots resulted in a higher population density for both TCA and the lepidopteran complex in both growing seasons compared to rainfed or non-irrigated plots. The likely reason could be that the herbivore insects benefit from feeding on irrigated, vigorously growing plants. On the contrary, plants with limited water availability could be less efficient in absorbing and utilizing essential nutrients (Hale et al., 2003; Mcvean and Dixon, 2001; Huberty and Denno, 2004). Studies have reported that soybean looper, a dominant herbivore, avoids water-stressed soybean plants (Gautam et al., 2024). While stink bug population densities remained stable in both irrigated and rainfed plots during the 2023 season, they were more abundant in the irrigated plots during the 2024 season. Collectively, these findings suggest that the incidence of insect pests is positively correlated with plant nutrient absorption and soil water availability through irrigation.
4.3 Interactive roles of micronutrients and irrigation
The metal defense hypothesis proposes that plants can benefit from accumulating metals, potentially deterring pests and pathogens by using them as a defense mechanism (Boyd and Martens, 1992; Poschenrieder et al., 2006). Micronutrients play a crucial role in shaping insect pest populations by altering the nutritional quality of host plants and other factors such as feeding, reproduction, survival, and overall population growth. Additionally, balanced micronutrient levels in plants can lead to increased resistance against insect pests by altering the plant’s biochemical defenses and making it less suitable for feeding (Dueli et al., 2021).
Studies have also demonstrated that consuming zinc-rich diets leads to higher zinc levels in insects, resulting in lower fertility, reduced growth, and increased mortality (Kazemi-Dinan et al., 2015; Lagisz et al., 2008), and a strong deterrent effect on host choice (Kazemi-Dinan et al., 2014, 2015). The results of the current study indicated that total insect population density significantly declined by the foliar application of zinc. Likewise, due to the foliar application of ferrous, the total population density of insect pests significantly decreased. However, no significant effects were found in the case of individual insect groups. This is likely due to the fact that different insects respond differentially to the addition of micronutrients in plants. Consistent with this, ferrous accumulation in plants has been found to act as a natural defense mechanism against insect pests (Rodríguez et al., 2005). Evidence also suggests that low levels of micronutrients (ferrous and zinc) can contribute to increased plant resistance to herbivory (Ribeiro, 2017; Coleman et al., 2005). Also, the synergistic effect of using both micronutrients is more pronounced than individual application (Kazemi-Dinan et al., 2014). However, there was no significant reduction in pest abundance when irrigation and micronutrients were applied together in the same plot. The reason behind this could be that the presence of irrigation increases insect populations, while micronutrients can decrease them. Combining water treatment with micronutrients could potentially mitigate the effects of higher pest incidence under irrigation. Despite acknowledging the study’s limitations, including the lack of clarity regarding the precise mechanisms by which micronutrients affect insect pests, this research offers valuable insight into the impact of micronutrients in pest control strategies.
5 Conclusion
In the present investigation, findings on pest incidence indicates that soybeans are attacked by herbivore pests from five families and more than 34 different species across the southern United States. This study also put forward the evidence that pest populations in soybean fields fluctuate at various growth stages, with the highest incidence of pests occurring during the V5-V6 stage and stink bug peaks at the R7-R8 stage. Foliar application of micronutrients can influence plant defenses, thereby reducing the insect pest pressure on crop systems. Results from the micronutrient application on total and specific insect species or species complexes support the hypothesis that combined micronutrient applications could impact insect pest population dynamics. However, the application of irrigation in soybean plots potentially attracted more insect pests to crops. No impacts on pest population were observed when the irrigation and micronutrients were applied simultaneously. Micronutrient combinations, even without irrigation, effectively controlled insect pests. Therefore, the application of combined micronutrients (ferrous and zinc) could offer a solution for reducing the impact of insect pests on soybeans. Future studies should explore the impact of micronutrients on plant and insect physiology in relation to pest infestations.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Ethics statement
The manuscript presents research on animals that do not require ethical approval for their study.
Author contributions
RD: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Resources, Validation, Visualization, Writing – original draft, Writing – review & editing. JG: Conceptualization, Data curation, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. AB: Conceptualization, Funding acquisition, Investigation, Methodology, Resources, Validation, Writing – original draft, Writing – review & editing. RK: Conceptualization, Methodology, Project administration, Resources, Software, Validation, Visualization, Writing – original draft, Writing – review & editing. GR: Conceptualization, Funding acquisition, Methodology, Project administration, Resources, Supervision, Validation, Writing – original draft, Writing – review & editing. JG: Conceptualization, Investigation, Methodology, Resources, Supervision, Visualization, Writing – original draft, Writing – review & editing. TK: Conceptualization, Data curation, Funding acquisition, Investigation, Methodology, Resources, Validation, Writing – original draft, Writing – review & editing. KR: Conceptualization, Funding acquisition, Investigation, Project administration, Resources, Validation, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This work was supported by the U.S. Department of Agriculture, Agricultural Research Service, Research Project# 6066-22000-090-00D Insect Control and Resistance Management in Corn, Cotton, Sorghum, Soybean, and Sweet Potato, and Alternative Approaches to Tarnished Plant Bug Control in the Southern United States. The field study was also supported by the Mississippi Soybean Promotion Board project #21-2023-2024.
Acknowledgments
Special thanks to Shundalyn Moore and Russel Godbold (USDA-ARS, Stoneville, MS) for technical assistance in field studies. USDA is an equal opportunity provider and employer.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Author disclaimer
The findings and conclusions in this publication are those of the author(s) and should not be construed to represent any official USDA or U.S. Government determination or policy. Mention of trade names or commercial products in this publication is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the U.S. Department of Agriculture.
References
Andrade F. H. (1995). Analysis of growth and yield of maize sunflower and soybean grown at Balcarce, Argentina. Field Crops Res. 41, 1–12. doi: 10.1016/0378-4290(94)00107-N
Ayala J., Vasquez A., Balakrishnan D., Madrigal E., George J., and Kariyat R. (2024). Effects of fast and slow-wilting soybean genotypes on fall armyworm (Spodoptera frugiperda) growth and development. Commun. Integr. Biol. 17, 2354421. doi: 10.1080/19420889.2024.2354421
Bala K., Sood A. K., Pathania V. S., and Thakur S. (2018). Effect of plant nutrition in insect pest management: A review. J. Pharmacogn. Phytochem. 7, 2737–2742.
Barathi S., Sabapathi N., Kandasamy S., and Lee J. (2024). Present status of insecticide impacts and eco-friendly approaches for remediation-a review. Environ. Res. 240, 117432. doi: 10.1016/j.envres.2023.117432
Bernardi O., Sorgatto R., Barbosa A., Domingues F., Dourado P., and Carvalho R. (2014). Low susceptibility of Spodoptera cosmioides, Spodoptera eridania and Spodoptera frugiperda (Lepidoptera: Noctuidae) to genetically modified soybean expressing Cry1Ac protein. Crop Prot. 58, 33–40. doi: 10.1016/j.cropro.2014.01.001
Beta T. and Isaak C. (2016). Grain production and consumption: overview. In: Encyclopedia of Food Grains – The World of Food Grains, V.1 (Oxford, United Kingdom: Elsevier), 349–428.
Beyer B. A., Srinivasan R., Roberts P. M., and Abney M. R. (2017). Biology and management of the Threecornered alfalfa hopper (Hemiptera: Membracidae) in alfalfa, soybean, and peanut. J. Integr. Pest Manage. 8, 1–10. doi: 10.1093/jipm/pmx003
Boyd R. S. and Martens S. (1992). The vegetation of ultramafic (serpentine) soils (Andover, Hampshire: Intercept Limited), 279–289.
Chang X.-n., Gao H.-j., Chen F.-j., and Zhai B.-p. (2008). Effects of environmental moisture and precipitation on insects: A review. Chin. J. Ecol. 27, 619–625.
Colella T., Candido V., Campanelli G., Camele I., and Battaglia D. (2014). Effect of irrigation regimes and artificial mycorrhization on insect pest infestations and yield in tomato crop. Phytoparasitica 42, 235–246. doi: 10.1007/s12600-013-0356-3
Coleman C. M., Boyd R. S., and Eubanks M. D. (2005). Extending the elemental defense hypothesis: dietary metal concentrations below hyperaccumulator levels could harm herbivores. J. Chem. Ecol. 31, 1669–1681. doi: 10.1007/s10886-005-5919-4
Debnath R., George J., Gautam M., Shafi I., Kariyat R., and Reddy G. V. P. (2024). Ecological interactions, host plant defenses, and control strategies in managing soybean looper, Chrysodeixis includens (Lepidoptera: Noctuidae). Front. Insect Sci. e 4. doi: 10.3389/finsc.2024.1480940
Dhaliwal G. S. and Arora R. (2002). Integrated pest management: concepts and approaches (Ludhiana, Punjab, India: Kalyani Publishers), 121.
Djaman K., Rudnick D. R., Moukoumbi Y. D., Sow A., and Irmak S. (2019). Actual evapotranspiration and crop coefficients of irrigated lowland rice (Oryza sativa L.) under semiarid climate. Ital. J. Agron. 14, 1059. doi: 10.4081/ija.2019.1059
Dueli G. F., DeSouza O., and Ribeiro S. P. (2021). Metal bioaccumulation alleviates the negative effects of herbivory on plant growth. Sci. Rep. 24, 19062. doi: 10.1038/s41598-021-98483-x
Gautam M., Shafi I., and Kariyat R. (2024). Compensation of physiological traits under simulated drought and herbivory has functional consequences for fitness in soybean (Glycine max (L.) Merrill). Environ. Exp. Bot. 226, 105944. doi: 10.1016/j.envexpbot.2024.105944
Greene A. D., Reay-Jones F. P., Kirk K. R., Peoples B. K., and Greene J. K. (2021). Associating site characteristics with distributions of pestiferous and predaceous arthropods in soybean. Environ. Entomol. 50, 477–488. doi: 10.1093/ee/nvab098
Hale B. K., Bale J. S., Pritchard J., Masters G. J., and Brown V. K. (2003). Effects of host plant drought stress on the performance of the bird cherry-oat aphid, Rhopalosiphum padi (L.): a mechanistic analysis. Ecol. Entomol. 28, 666–677. doi: 10.1111/j.1365-2311.2003.00563.x
Hesler L. S., Allen K. C., Luttrell R. G., Sappington T. W., and Papiernik S. K. (2018). Early-Season pests of soybean in the United States and factors that affect their risk of infestation. J. Integr. Pest Manage. 9, 1–15. doi: 10.1093/jipm/pmx028
Huberty A. F. and Denno R. F. (2004). Plant water stress and its consequences for herbivorous insects: a new synthesis. Ecol 85, 1383–1398. doi: 10.1890/03-0352
Huseth A. S., Koch R. L., Reisig D., Davis J. A., Paula-Moraes S. V., and Hodgson E. W. (2021). Current distribution and population persistence of five lepidopteran pests in U.S. Soybean. J. Integr. Pest Manage. 12, 1–10. doi: 10.1093/jipm/pmab004
Inbar M., Doostdar H., and Mayer R. T. (2001). Suitability of stressed and vigorous plants to various insect herbivores. Oikos 94, 228–235. doi: 10.1034/j.1600-0706.2001.940203.x
Karam F., Maasad R., Sfeir T., Mounzer O., and Rouphael Y. (2005). Evapotranspiration and seed yield of field grown soybean under deficit irrigation conditions. Agric. Water Manage. 75, 226–244. doi: 10.1016/j.agwat.2004.12.015
Kazemi-Dinan A., Barwinski A., Stein R. J., Kramer U., and Muller C. (2015). Metal hyperaccumulation in Brassicaceae mediates defense against herbivores in the field and improves growth. Entomol. Exp. Appl. 157, 3–10. doi: 10.1111/eea.12333
Kazemi-Dinan A., Thomaschky S., Stein R. J., Krämer U., and Müller C. (2014). Zinc and cadmium hyperaccumulation act as deterrents towards specialist herbivores and impede the performance of a generalist herbivore. New Phytol. 202, 628–639. doi: 10.1111/nph.12663
Lagisz M. (2008). Changes in morphology of the ground beetle Pterostichus oblongopunctatus F. (Coleoptera: Carabidae) from vicinities of a zinc-and-lead smelter. ET&C 27, 14–21. doi: 10.1897/07-661.1
Lich M. A., Wright D., and Lenssen A. W. (2013). Soybean response to drought (Ames, IA (USA: Agriculture Iowa State University Extension and Outreach).
McPherson R. and MacRae T. (2009). Evaluation of transgenic soybean exhibiting high expression of a synthetic Bacillus thuringiensis cry1A transgene for suppressing lepidopteran population densities and crop injury. J. Econ. Entomol. 102, 1640–1648. doi: 10.1603/029.102.0431
Mcvean R. I. K. and Dixon A. F. G. (2001). The effect of plant drought stress on populations of the pea aphid Acyrthosiphon pisum. Ecol. Entomol. 26, 440–443. doi: 10.1046/j.1365-2311.2001.00341.x
Melloul E., Rocher L., Gros R., Bischoff A., and Blight O. (2024). Irrigation decreases flower cover and beneficial arthropod abundances in Mediterranean vineyards. Basic Appl. Ecol. 77, 1–7. doi: 10.1016/j.baae.2024.03.003
Murúa M. G., Vera M. A., Herrero M. I., Fogliata S. V., and Michel A. (2018). Defoliation of soybean expressing Cry1Ac by lepidopteran pests. Insects 9, 93. doi: 10.3390/insects9030093
Musser F. R., Catchot A. L., Davis J. A., DiFonzo C., Graham S. H., Greene J. K., et al. (2022). Soybean insect losses in the United States. Midsouth Entomol. 15, 39–63.
National Agriculture Statistics Service (NASS). (2013). Crop Production 2012 Summary. Washington DC: US Department of Agriculture.
Nessel M. P., Konnovitch T., Romero G. Q., and González A. L. (2023). Decline of insects and arachnids driven by nutrient enrichment: a meta-analysis. Ecol 104, e3897. doi: 10.1002/ecy.3897
Poschenrieder C., Tolra R., and Barcelo J. (2006). Can metals defend plants against biotic stress? Trends Plant Sci. 11, 288–295. doi: 10.1016/j.tplants.2006.04.007
Raderschall C. A., Vico G., Lundin O., Taylor A. R., and Bommarco R. (2021). Water stress and insect herbivory interactively reduce crop yield while the insect pollination benefit is conserved. Glob. Chang. Biol. 27, 71–83. doi: 10.1111/gcb.15386
Ren Z., Cai T., Wan Y., Zeng Q., Li C., Zhang J., et al. (2023). Unintended consequences: disrupting microbial communities of Nilaparvata lugens with non-target pesticides. Pestic Biochem. Physiol. 194, 105522. doi: 10.1016/j.pestbp.2023.105522
Ribeiro S. P. (2017). Plant defense against leaf herbivory based on metal accumulation: examples from a tropical high altitude ecosystem. Plant Species Biol. 32, 147–155. doi: 10.1111/1442-1984.12136
Rivelli A. R., Toma I., Trotta V., Fanti P., De Maria S., and Battaglia D. (2013). “Combined effect of water stress and Macrosiphum euphorbiae infestation on plant growth in tomato,” in Proceedings of the 12th Congress of the European Society for Agronomy, Helsinki, Finland. eds. Stoddard F. and Makela P. (University of Helsinki), 334–335.
Rodríguez N., Menéndez N., Tornero J., Amils R., and Fuente V. D. L. (2005). Internal iron biomineralization in Imperata cylindrica, a perennial grass: chemical composition, speciation and plant localization. New Phytol. 165, 781–789. doi: 10.1111/j.1469-8137.2004.01264.x
Soybean Invertebrate Loss Estimates from the United States (2023). Crop Protection Network. doi: 10.31274/cpn-20230511-0
Srikanth J., Easwaramoorthy S., Kurup N. K., and Santhalakshmi G. (1997). Spider abundance in sugarcane: impact of cultural practices, irrigation and post-harvest trash burning. Biol. Agric. Hortic. 14, 343–356. doi: 10.1080/01448765.1997.9755169
Steffey K. L. (2015). Insects and their management, Compendium of soybean diseases and pests. 5th ed (USA: APS Press).
USDA-FAS Production-Soybeans. Available online at: https://fas.usda.gov/data/production/commodity/2222000
Yu H., Li Y., Li X., Romeis J., and Wu K. (2013). Expression of Cry1Ac in transgenic Bt soybean lines and their efficiency in controlling lepidopteran pests. Pest Manag. Sci. 69, 1326–1333. doi: 10.1002/ps.3508
Zaffaroni M., Cunniffe N. J., and Bevacqua D. (2020). An ecophysiological model of plant-pest interactions: the role of nutrient and water availability. J. R Soc. Interface 17, 20200356. doi: 10.1098/rsif.2020.0356
Keywords: metal defense, fertilizer, ferrous, field survey, zinc
Citation: Debnath R, George J, Bhandari AB, Kariyat R, Reddy GVP, Glover JP, Kharel TP and Reddy KN (2025) Impact of irrigation and micronutrient treatments on insect herbivore population dynamics in soybean. Front. Agron. 7:1571675. doi: 10.3389/fagro.2025.1571675
Received: 05 February 2025; Accepted: 17 September 2025;
Published: 03 October 2025.
Edited by:
Murray B. Isman, University of British Columbia, CanadaReviewed by:
Vijay Sheri, Texas Tech University, United StatesVlad Stoian, University of Agricultural Sciences and Veterinary Medicine of Cluj-Napoca, Romania
Copyright © 2025 Debnath, George, Bhandari, Kariyat, Reddy, Glover, Kharel and Reddy. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Justin George, anVzdGluLmdlb3JnZUB1c2RhLmdvdg==