 Amira M. I. Mourad
Amira M. I. Mourad Ahmed Sallam
Ahmed Sallam Andreas Börner
Andreas Börner Rania M. Hamdy4
Rania M. Hamdy4 Samar M. Esmail
Samar M. Esmail- 1Genebank Department, Leibniz Institute of Plant Genetics and Crop Plant Research (IPK), Gatersleben, Germany
- 2Agronomy Department, Faculty of Agriculture, Assuit University, Asyut, Egypt
- 3Genetics Department, Faculty of Agriculture, Assuit University, Asyut, Egypt
- 4Food Science and Technology Department, Faculty of Agriculture, Assuit University, Asyut, Egypt
- 5Wheat Disease Research Department, Plant Pathology Research Institute, Agricultural Research Center, Giza, Egypt
Wheat powdery mildew (WPM) is a devastating disease that causes significant yield losses worldwide. In Egypt, a lake in immune cultivars was reported and little is known about its genetic control. In this study, two different plant sets (the isoline set and the test set) were evaluated for WPM adult plant resistance (APR-WPM) over three cropping seasons (2021-2022, 2022-2023, and 2023-2024). The results of the isoline set revealed that the Pm1b, Pm34, Pm36, MIAG12, and NACG13 resistance genes were stable among the three cropping seasons. The results of the evaluated set revealed highly significant differences among the cropping seasons as well as among the tested genotypes, suggesting the presence of different Blumeria graminis f. sp. tritici (Bgt) races and the potential to select highly resistant genotypes. Eight resistant genotypes belonging to five different countries, excluding Egypt, were identified. A genome-wide association study (GWAS) was performed using each cropping season separately as well as the average of the three seasons. GWAS was done using two different SNP data sets, GBS and 25K-SNP array. A total of 86 significant markers associated with resistance were identified. Among these markers, only four markers were stable across at least two seasons. Three gene models harboring these markers were detected and functionally annotated to be linked with disease resistance. Gene enrichment analysis revealed that each gene model regulated a single network of different biological processes and molecular function pathways. Among the resistant genotypes, one Sudanese genotype was selected for its strong WPM resistance and high yield potential. This genotype was also found to be highly adapted to the Egyptian environment and a high-yielding genotype. The incorporation of this genotype into future breeding programs could increase the resilience of Egyptian wheat germplasm.
1 Introduction
Among cereal crops, wheat (Triticum aestivum) is considered the most important crop that provides food for 35% of the world’s population (Paux et al., 2008; Mourad et al., 2019; Mondal et al., 2021). Wheat powdery mildew (WPM, caused by Blumeria graminis f. sp tritici (Bgt)) is one of the most devasting foliar diseases affecting wheat worldwide and causes significant yield losses of 62% (Costamilan, 2005; Maxwell et al., 2009; El-Shamy et al., 2012; Huang et al., 2013; Singh et al., 2016; Jabran et al., 2023, 2024). In Egyptian fields, WPM occurs annually and has become more threatening than rust diseases (El-Shamy and Mohamed, 2022). The severity of fungal diseases, such as WPM, may increase the vulnerability of wheat to other biotic and abiotic stresses (Figueroa et al., 2018). Moreover, climate change is expected to exacerbate fungal disease outbreaks in the near future (Kissoudis et al., 2014). Therefore, greater efforts are needed to manage this disease effectively. Similarly, few studies have focused on WPM management under Egyptian conditions. These studies revealed the presence of Pm genes in Egyptian wheat germplasms and characterized different Bgt races in Egyptian fields (El-Shamy et al., 2012, 2016; Emara et al., 2016; Elsayed and Elkot, 2020; Draz et al., 2022; Mourad et al., 2024a).
As a fungal disease, WPM resistance can be classified into two main types: race-specific resistance (also known as seedling resistance) and race-non-specific resistance (also known as adult plant resistance) (Roelfs et al., 1992; Bhavani et al., 2021; Cheng et al., 2022). Pyramiding multiple seedling and adult plant resistance genes in the same genotype can provide a high level of broad-spectrum resistance to combat this serious disease (Bajgain et al., 2015; Mourad et al., 2018). Fungicide application is a potential method for WPM control. However, because of its harmful environmental effects, especially its long-term use, alternative control methods are needed (Goswami et al., 2018). One environmentally friendly approach is biological control, which involves the use of nonpathogenic organisms such as Trichoderma asperellum (T34) (Mourad et al., 2024c), Aspergillus chevalieri (BYST01) (Zhang et al., 2024), and Bacillus spp (Matzen et al., 2019; Shahin et al., 2019). However, further studies are needed to identify effective biological control agents for widespread use. Plant breeding has long been an effective method for identifying and pyramiding WPM resistant genes in wheat germplasm (Liang et al., 2019; Andersen et al., 2020; Bhavani et al., 2021). To date, 68 WPM resistant genes have been detected and mapped onto wheat chromosomes (He et al., 2021). However, some of these genes are linked to undesirable traits (Hurni et al., 2014). Therefore, identifying additional effective resistance genes or genomic regions is essential for improving WPM resistance.
Association mapping (AM) is an effective approach for detecting novel genomic regions controlling specific traits. Owing to advancements in sequencing methods in recent years, genome-wide association studies (GWASs) have been widely used to improve various traits in wheat (Alqudah et al., 2020). However, the number of GWASs conducted in the last five years to identify genomic regions controlling WPM resistance is very low (Kang et al., 2019; Mohler and Stadlmeier, 2019; Simeone et al., 2020; Miedaner et al., 2020; Alemu et al., 2021; Du et al., 2021; Hinterberger et al., 2022; Jin et al., 2022; Kaur et al., 2023; Mourad et al., 2024a; Bhadana et al., 2025). Interestingly, only one GWAS has been conducted to comprehensively understand the genetic control of WPM at the seedling stage under Egyptian conditions (Mourad et al., 2023). Additionally, numerous databases for functional annotations, gene expression, and gene enrichment have recently emerged, providing valuable insights into the functions of different wheat genomic regions (Bolser et al., 2007; Pearce et al., 2015; Ge et al., 2022). One of the valuable databases for enhancing the GWAS results is EnsemblePlants which provides detailed information on the gene models present in the same position as the significant SNP markers (Bolser et al., 2007). These gene models could be submitted to Knetminer (Hassani-Pak et al., 2021) and ShinyGo (Ge et al., 2022) databases to understand more about their role in controlling the studied trait based on their functional annotation. Therefore, integrating GWAS results with data from these databases will enhance our understanding of the genetic control of this trait. However, when GWAS and database findings are utilized to study disease resistance, it is crucial to consider the yield performance of the tested genotypes to avoid the development of highly resistant genotypes with low yield potential.
The objectives of this study are to: (1) investigate the genetic control of adult plant resistance to WPM (APR-WPM) under Egyptian field conditions; (2) identify marker-trait associations for WPM resistance across multiple cropping seasons; (3) detect stable genomic regions associated with the resistance; and (4) Select WPM-resistant genotypes with high yield potential for future breeding programmes.
2 Materials and methods
2.1 Plant materials
In this study, two distinct sets of plant materials were evaluated for WPM-APR: the “tested set” and the “isoline set”. The tested set consisted of 196 spring wheat genotypes. A detailed description of this set has been provided previously (Esmail et al., 2023b; Mourad et al., 2024c). Briefly, it represents a highly diverse collection from 22 countries worldwide (Supplementary Table S1). Among these genotypes, 35 were Egyptian, including breeding lines, old cultivars, and new cultivars. Seeds of the Egyptian genotypes were sourced from Egyptian governorate, whereas seeds of the non-Egyptian genotypes were obtained from the USDA-ARS, Aberdeen, ID, the U.S.A.
The isoline set included 20 distinct lines, each carrying a different powdery mildew resistance gene (Supplementary Table S2). This set was included to identify the possible Pm resistant genes against the Egyptian Bgt races. Seeds for this set were obtained from Dr. Abdelrazek Abdelrhim at Minia University, Minia, Egypt, and had been previously evaluated for powdery mildew resistance under Egyptian field conditions (Abdelrhim et al., 2018).
2.2 Field evaluation of powdery mildew adult plant resistance
The powdery mildew resistance of the plants in the tested set and the isoline set was evaluated across three cropping seasons: 2021-2022, 2022-2023, and 2023-2024. The evaluations were conducted at the Experimental Farm of Sakha Agricultural Research Station, Agricultural Research Center, Egypt (31°5′12″N, 30°56′49″E). The experimental layout was carried out using a randomized complete block design (RCBD) with three replications per cropping season. The genotypes were sown at the end of November (optimum sowing date) each year.
Each tested genotype was planted in a 3-meter-long row spaced 30 cm apart, with 5 g of seeds per row. During each cropping season, the experiment area was surrounded by a 1.5 m² belt of the highly powdery mildew-susceptible variety “Jagalene”. Artificial inoculation was conducted on this surrounding belt, which served as an infection spreader to ensure a uniform distribution of Bgt spores. Inoculation involved the use of Bgt conidia collected from naturally infected fields in the surrounding area. This was done by removing the propagated conidiospores from the infected plants and dispreading them over the spreader belt (Draz et al., 2023). Standard agronomic practices were applied according to recommendations for wheat cultivation in Egyptian fields.
The disease severity (DS) of powdery mildew was recorded once the susceptible check was fully covered with Bgt spores via a modified Cobb scale (0–9) as described previously in Peterson et al. (1948). The percentage of the leaf surface covered by mycelia was determined via Cobb’s scale as follows: 0% = immune (no visible sign of infection); 1 = 10% of the leaf area covered by mycelia; 2 = 20%; 3 = 30%; 4 = 40%; 5 = 50%; 6 = 60%; 7 = 70%; 8 = 80%; and 9 = 90%. Genotypes with a percentage < 20% were considered resistant.
2.3 Statistical analysis of powdery mildew adult plant resistance
Analysis of variance (ANOVA) was conducted by combining DSs from the three cropping seasons, and PLABSTAT software (Utz, 1997) was used via the following method:
where Yijk is an observation of genotype i in replication j, which was planted in experiment k; µ is the general mean; gi, ri, and Ek are the main effects of genotypes (random effects), replications, and experiments (fixed effects), respectively; and eijk is the error.
Furthermore, for each experiment, the following model was used for ANOVA:
where Yij is an observation of genotype i in replication j; μ is the general mean; gi and rj are the main effects of genotypes and replications, respectively; and eij is the error. The BLUP values were calculated following the same model using the lme4 R package (Bates et al., 2015). BLUPs were used to adjust the phenotypic data scored across different seasons as well as in each season. Broad-sense heritability was calculated via the following formula:
where and are the variances of the lines and the residuals, respectively. r is the number of replicates.
2.4 Genome-wide association study for resistance, gene models controlling WPM-APR resistance, functional annotation, and gene enrichment
To determine the genetic control of powdery mildew resistance, GWAS was conducted using DS-BLUP values of each cropping season separately as well as the average of the three cropping seasons together. The rMVP package of R software (Yin et al., 2021) was used to run the GWAS. This package allows three different models of GWAS as follows: the generalized linear model (GLM), the mixed linear model (MLM), and the fixed and random model Circulating Probability Unification (FarmCPU), which includes principle component analysis (PCA) and/or kinship as a covariate. The best model for each experiment was determined on the basis of the Quantil-Quantil-Plot (QQ-plot), which represents the deviation of the distribution of the observed -log10 (P-value) from the expected value. The significant SNP markers were identified as those whose P-value was ≤0.001 (-log10≥3.00) (Ahmed et al., 2022; Mourad et al., 2024b).
Furthermore, gene models harboring significant markers were detected on the basis of the base-pair position of the markers and the presence of gene models in the same position via the EnsemblePlants database (Bolser et al., 2007). Despite the EnsemblePlants database utilizing the International Wheat Genome Sequencing Consortium (IWGSC) RefSeq v2.0 assembly for SNP positioning, gene annotation was based on IWGSC RefSeq v1.0, which currently offers more comprehensive and functionally curated gene models. While the coordinate systems between v1.0 and v2.0 are broadly compatible in well-anchored regions, minor differences in gene structure or gene IDs may occur. Furthermore, gene enrichment of the common gene models in both evaluations was detected based on the biological process, molecular function, and cellular component pathways via the ShinyGo 0.76 database (Ge et al., 2022). To identify the most important pathways, a cutoff false discovery rate (FDR) p-value < 0.01 was applied.
2.5 Selection of superior genotypes to improve WPM-APR in Egyptian wheat germplasm
The most resistant genotypes were those with DS<20% in all three cropping seasons. Moreover, the genetic distance between each pair of selected superior genotypes was calculated via neighbor-joining via TASSEL V5.2.19 software (Bradbury et al., 2007) and visualized as a phylogenetic tree via the iTol database (Letunic and Bork, 2021). For each evaluated genotype, the thousand kernel weight (TKW, g) was scored by measuring the weight of 100 kernels from each genotype in two cropping seasons, 2022-2023 and 2023-2024.
2.6 Yield and yield components of the selected genotypes under non-infected field conditions
The yield and yield components of the same tested plant materials (196 spring wheat genotypes) were evaluated in noninfected fields at the field of the Faculty of Agriculture, Assuit University, Asyut, Egypt (27°11′20.36′′N, 31°10′06.45′′E), for three cropping seasons: 2021–2022, 2022–2023, and 2023–2024. For each cropping season, the seeds of each tested genotype were sown in rows with a length of 1.5 m. The distance between the seeds in each row was 10 cm. The experimental design was a randomized complete block design (RCBD) with two replications in each cropping season. The irrigation and agricultural practices were performed as recommended for the wheat plants. To ensure the absence of infection, fungicides were applied as recommended.
The following traits were recorded for each genotype in each replication/cropping season: heading date (HD), plant height (PH, cm), spike length (SPL, cm), grain number of spikes (GN/SP), number of spikelets per spike (NS/SP), grain yield per spike (GY/SP, g), and thousand kernel weight (TKW, g).
3 Results
3.1 Evaluation of powdery mildew isoline set
Among the 20 evaluated isolines, none were immune in the three cropping seasons (Figure 1). However, NC06BGTAG13 line (carrying NCAG13 gene) was immune in two cropping seasons (2022-2023 and 2023-2024) and resistant (DS = 5%) in the third cropping season. Moreover, 5-BIL29, NC06BGTAG12, NC96BGTD7, NC96BGTD3, and NC99BGTAG11 isolines (carrying Pm36, MIAG12, Pm34, Pm35, and Pm37 gene, respectively) were resistant in at least two cropping seasons (DS<20%). MocZlatka (carrying Pm1b gene) was immune in only two cropping seasons (2021-2022 and 2023-2024), whereas it is not resistant in 2022-2023 (DS<20%). The remaining 13 isolines were not resistant in any cropping season.
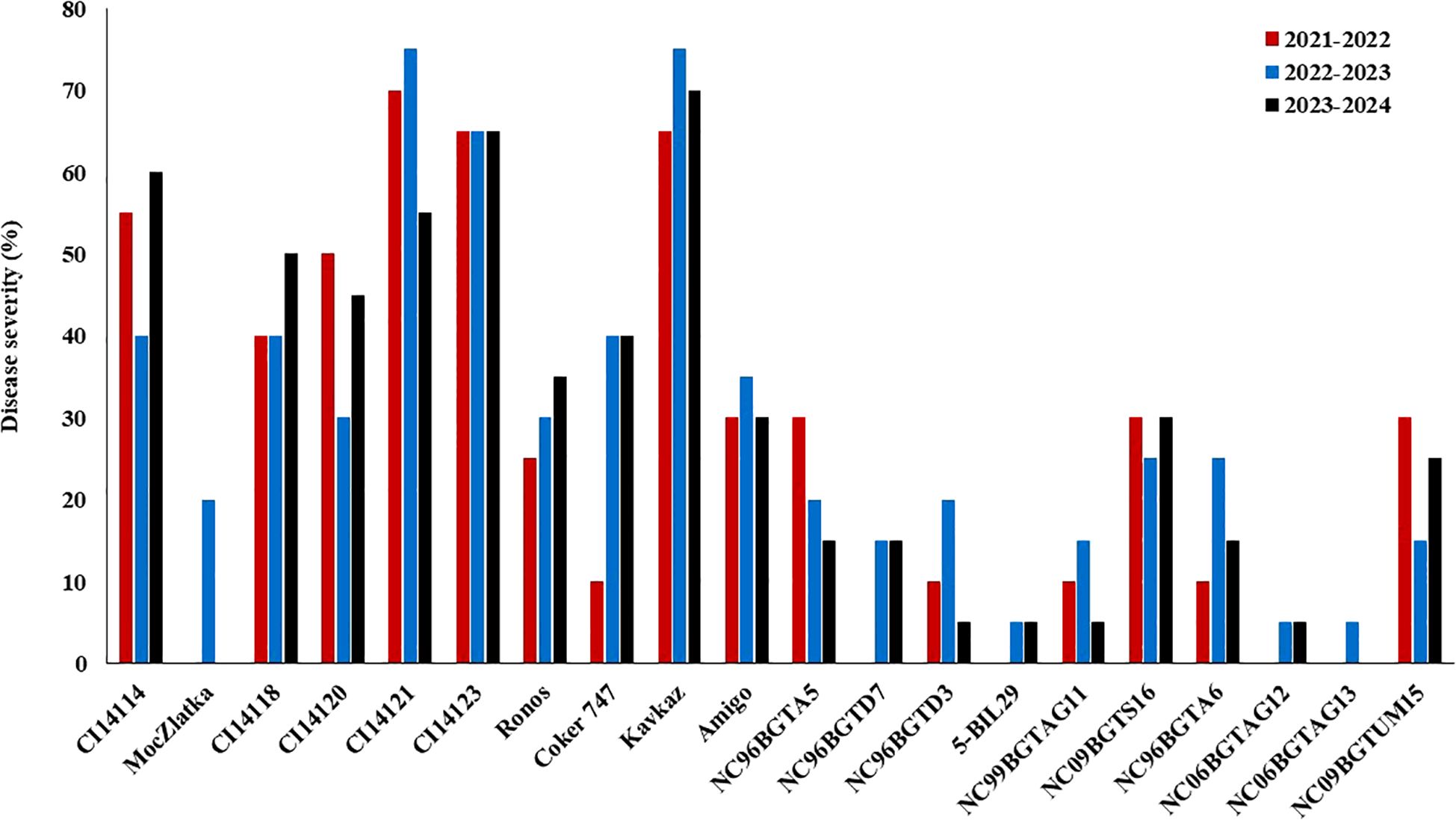
Figure 1. The response of the tested powdery mildew isolines in each cropping season under Egyptian field conditions.
3.2 Variation in WPM-APR in the tested plant set
The ANOVA revealed highly significant differences in WPM-APR among the three tested cropping seasons (Supplementary Table S3). Furthermore, a highly significant G×Y interaction was detected. The resistance of the tested genotypes significantly differed during each cropping season (Supplementary Tables S4-S6). The disease severity ranged from 13.33% to 80%, with an average of 48.02% in the first cropping season (Table 1). However, it ranged from 13.33% to 86.67%, with an average of 44.84%, and from 3.33% to 83.33%, with an average of 37.55%, in 2022-2023 and 2023-2024, respectively. High degrees of broad-sense heritability (H2) were detected in the three cropping seasons, with values of 0.89, 0.92, and 0.94 in the 2021-2022, 2022-2023, and 2023-2024 cropping seasons, respectively (Table 1). It was clearly noticed that the disease severity of WPM infection was almost the same in 2021-2022 and 2022-2023 cropping seasons. While the DS in 2023-2024 cropping season was completely different from 2021-2022 and 2022-2023 (Figure 2).
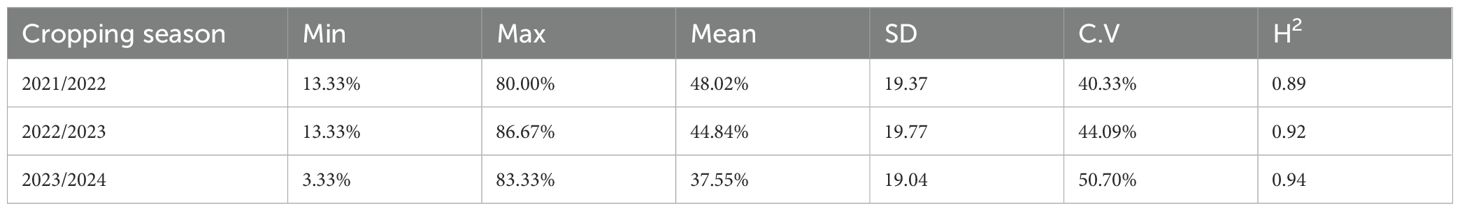
Table 1. Phenotypes of powdery mildew adult plant resistance in the three years of evaluation (2022, 2023, and 2024) representing the minimum, maximum, and mean of the resistance in each cropping season as well as standard deviation (SD), coefficient of variance (CV), and broad-sense heritability (H2).
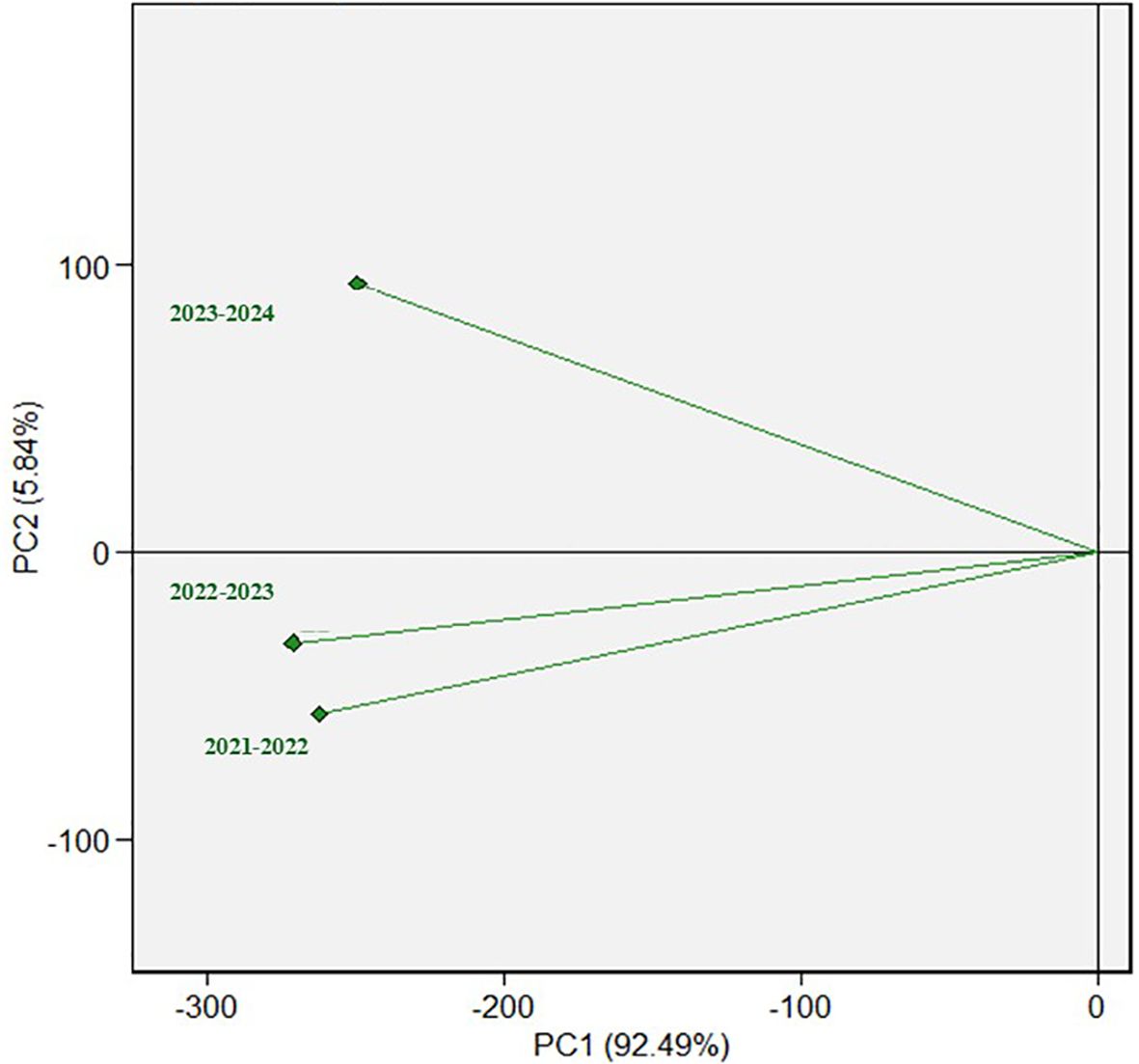
Figure 2. The relationship among disease severity in the three cropping seasons.
None of the tested genotypes were immune in any cropping season (Figure 3a). However, eight, 11, and 37 genotypes were found to be resistant to WPM (DS<20%) in 2021-2022, 2022-2023, and 2023-2024, respectively. Notably, the majority of the tested genotypes were susceptible to WPM, with 188, 185, and 159 genotypes in 2021-2022, 2022-2023, and 2023-2024, respectively. Among the resistant genotypes in each cropping season, eight genotypes were resistant in all three seasons (Figure 3b). These genotypes were from Germany (three genotypes), U.S.A. (two genotypes), Sudan (one genotype), Kenya (one genotypes), and the U.K (one gneotype) (Table 2). The DS ranged from 6.67% for the German genotype “Carstens Sommerweizen” in 2023-2024 to 16.67% for the eight genotypes in both 2021-2022 and 2022-2023. Interestingly, none of the tested 35 Egyptian genotypes were immune or resistant to WPM during the three cropping seasons (Supplementary Figure S1). However, Giza-171 and Sohag 5 were resistant in two cropping seasons (DS<20%) and nonresistant in the third season. The remaining 33 Egyptian genotypes were found to be resistant in only one cropping season or susceptible in the three cropping seasons.
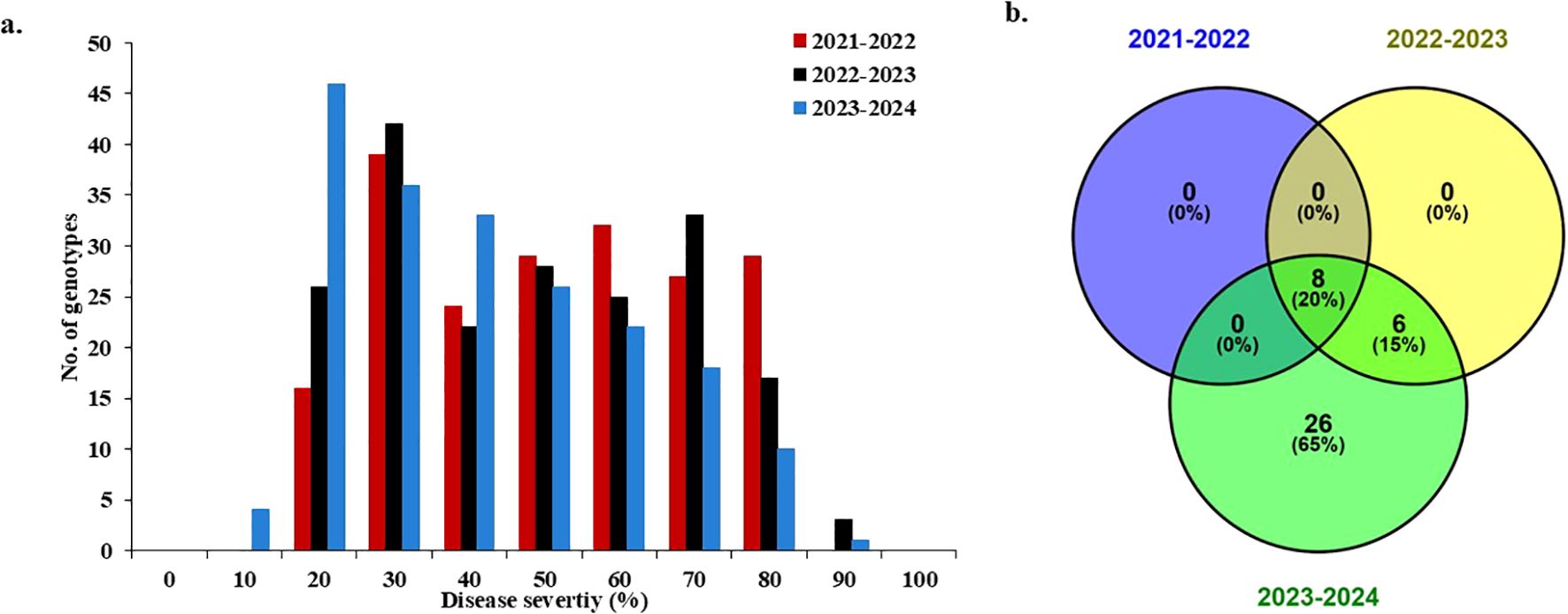
Figure 3. The response of the 196-spring wheat genotypes to powdery mildew infection at the adult growth stage in the three evaluation growing seasons: (a) histogram represents the distribution of disease severity (%) in each growing season. (b) number of resistant genotypes (with a value of disease severity<20%) in each growing season and the number of commonly resistant genotypes in the three growing seasons.
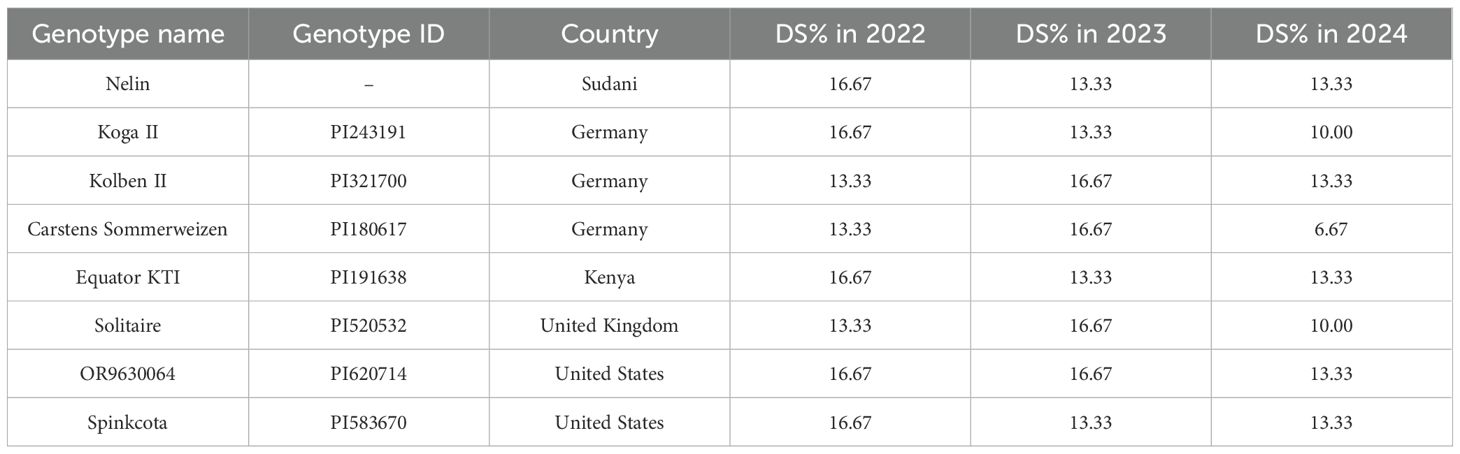
Table 2. List of common resistant genotypes (DS<20%) in the three cropping seasons 2022, 2023, and 2024, their country of origin, and plant ID.
3.3 Association mapping for WPM-APR
3.3.1 GWAS for WPM-APR in each cropping season
Owing to the highly significant differences in WPM-APR among the three cropping seasons, GWAS was carried out for each cropping season separately. Furthermore, GWAS was run on the basis of the average of the three cropping seasons to identify genomic regions commonly controlling the resistance of WPM-APR. Owing to the presence of three different subpopulations in the tested set (Mourad et al., 2020), PCA and/or kinship were included in each GWAS model. For each season, three different GWAS models were used for each SNP marker dataset. The Q-Q and Manhattan plots of the best model for each marker set in each cropping season are presented in Supplementary Figures S2, S3. The summarized results of the GWAS for each cropping season are presented in Table 3. The detailed results are presented in Supplementary Table S7.

Table 3. Summary of significant markers associated with powdery mildew adult plant resistance in each cropping season, as well as across three seasons, in the evaluated 196 spring wheat genotypes using the iSelect 25K SNP array (25K SNPs) and genotyping-by-sequencing (GBS) marker datasets.
A total of 26 markers were significantly associated with resistance in the 2021-2022 cropping season on the basis of both 25K-SNPs and GBS-SNPs (Table 3). These markers were distributed among 13 different chromosomes. The percentage of phenotypic variation explained by these markers ranged from 0.1% to 10.66%. The resistant alleles of these markers were found to reduce disease severity, ranging from -5.92 to -2.34, and were located within 11 different gene models. Among these gene models, only three were major genes, as they explained more than 10% of the phenotypic variation.
Furthermore, 37 significant markers that were distributed among 14 chromosomes were found to be significantly associated with resistance in the 2022-2023 cropping season. These markers explained 0.03–16.38% of the phenotypic variation. The allele effects of these markers ranged from -7.80 to -2.63. A total of 17 genes harbored these significant markers, with only four major genes.
The lowest number of significant markers associated with resistance was identified in the 2023-2024 cropping season, with 19 markers. These markers were distributed among eight chromosomes and explained 0.92–13.61% of the phenotypic variation. The target alleles of these markers reduced disease severity with a range of -9.49 to -3.04. A number of 14 gene models were found to harbor these significant markers with only two major genes.
On the basis of the average of the three cropping seasons, 27 markers were significantly associated with the resistance and were distributed among 9 chromosomes. These markers explained 0.50–13.16% of the phenotypic variation. Furthermore, the range of DS reduction due to the presence of the target alleles of these markers was -7.46 to -2.46. The significant markers were located within 17 gene models. Among these gene models, only two genes were found to harbor markers with a major effect.
Notably, the significant markers associated with WPM-APR in the three cropping seasons as well as the average over the three years were distributed across all 21 wheat chromosomes except 4A, 3D, and 6D (Figure 4a). The largest number of significant markers were found on the chromosomes 2B and 7B, with 11 markers each. The lowest number of markers was found on chromosomes 3A, 4B, 4D, 5D, and 7D, which carry one marker for each chromosome. The chromosomal locations of the five significant markers are not known; thus, they are located on unknown chromosomes.
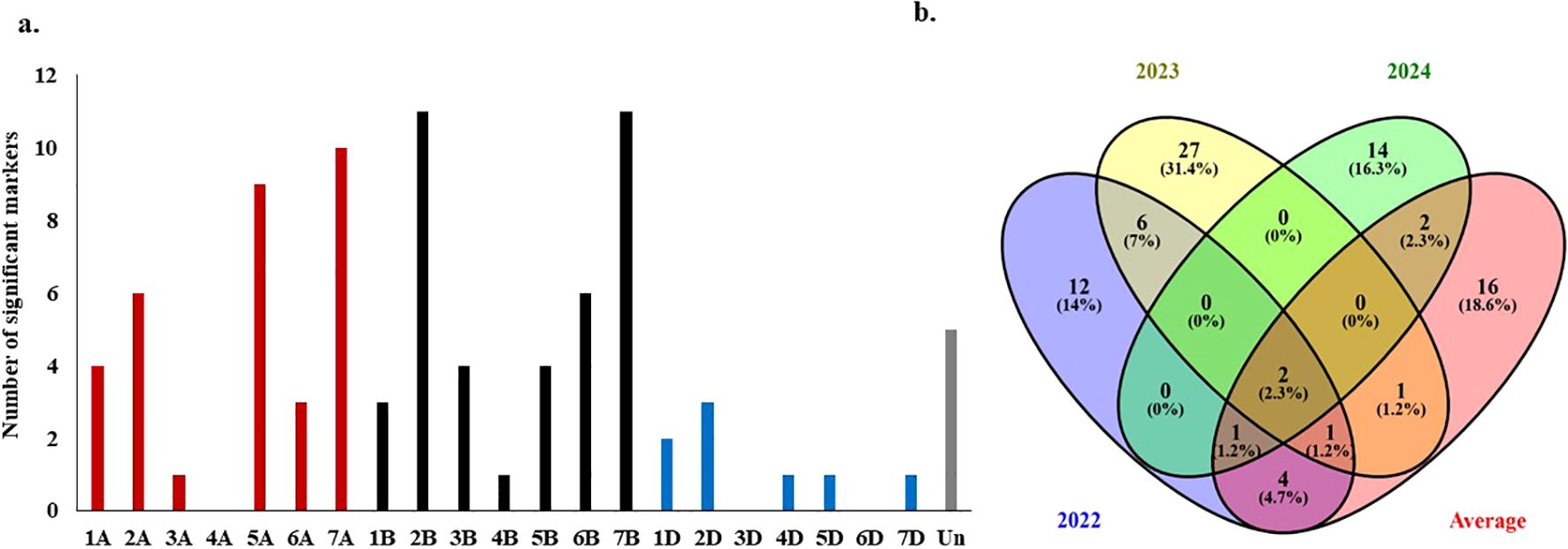
Figure 4. Significant markers associated with powdery mildew adult plant resistance in the evaluated 196-spring wheat genotypes: (a) number of signifcant markers in each growing season as well as the average of the three growing seasons, (b) number of total significant markers associated with the resistance on each one of the 21-wheat chromosomes.
It was noted that two stable markers were found to be significantly associated with WPM under all conditions (three years and the average over the three years as well). One stable marker was significant in 2021-2022, 2023-2024, and the average, while one was significant in 2021-2022, 2022-2023, and the average (Figure 4b). These four common markers are located on four different chromosomes: 1A, 2B, 3A, and 5A (Table 4). Moreover, four, one, and two markers were common between 2021-2022 and the average, 2022-2023 and the average, and 2023-2024 and the average, respectively.

Table 4. List of significant markers commonly associated with powdery mildew adult plant resistance among different cropping seasons and their average (Avr), their chromosomal location, p-value, phenotypic variation explained by marker (R2), target allele, allele effect, and gene models harboring them.
Chromosomes 1A, 2B, and 5A, which carried the four common markers across the three cropping seasons, also presented many significant markers, with 4, 11, and 9 markers, respectively (Figure 4a). Therefore, linkage disequilibrium (LD) between the significant markers and the common significant marker on each chromosome was investigated. Notably, no significant LD was detected between any of the common significant markers and the other significant markers detected on the same chromosome (Supplementary Figure S4).
3.3.2 Gene models harboring significant markers associated with WPM-APR
A total of 11, 17, 14, and 17 genes were found to harbor significant markers associated with resistance in 2021-2022, 2022-2023, 2023-2024, and the average, respectively (Table 3). The complete list of these genes and their functional annotations are presented in detail in Supplementary Table S7. Notably, the functional annotation of these genes is associated with disease resistance and plant-pathogen interactions, such as glutathione S-transferase produced by TraesCS3B02G536100, NBS–LRR–like resistance protein produced by TraesCS1D02G064200, glucan endo-1,3-beta-glucosidase produced by TraesCS7A02G069700, and serine/threonine–protein kinase produced by the TraesCS2A02G537600 gene.
The four common significant markers associated with resistance in at least two different cropping seasons were found to be located within three different gene models (Table 4). Two genes, TraesCS2B02G569400 and TraesCS5A02G314000, harbored markers associated with resistance in the three cropping seasons. The third gene, TraesCS3A02G282800, harbored a marker associated with resistance in the 2021-2022 and 2023-2024 cropping seasons. Furthermore, the TraesCS2B02G569400 gene was found to have a major effect on resistance in the three cropping seasons. The functional annotation of this gene model revealed that it controls heavy metal transport/detoxification superfamily proteins. On the other hand, the two other genes had minor effects and controlled the production of rhomboid-like proteins and nonspecific serine/threonine protein kinases by the TraesCS5A02G314000 and TraesCS3A02G282800 genes, respectively.
Gene enrichment of the three common gene models revealed 12 biological process (BP) and six molecular function (MF) pathways associated with resistance (FDR<0.05) (Figure 5a). The BP pathways were associated with cell death, negative regulation of biological processes, proteolysis, and apoptotic processes. The MF pathways were associated with serine-type peptidase, endopeptidase activity, and serine hydrolase activity. The 12 BP pathways worked in three different networks, each controlled by one gene model (Figure 5b). The six MF pathways worked in two different networks controlled by only two genes from the three identified common genes. Furthermore, TraesCS2B02G569400 was reported to be directly related to the response of wheat to fungi and bacteria and the downregulation of the defense response (Figure 5c). The TraesCS3A02G282800 gene (known as SnRK1) was reported to be associated with the defense response to bacteria, fungi, and insects in Arabidopsis thaliana. No relationship was found between TraesCS5A02G314000 and disease resistance.
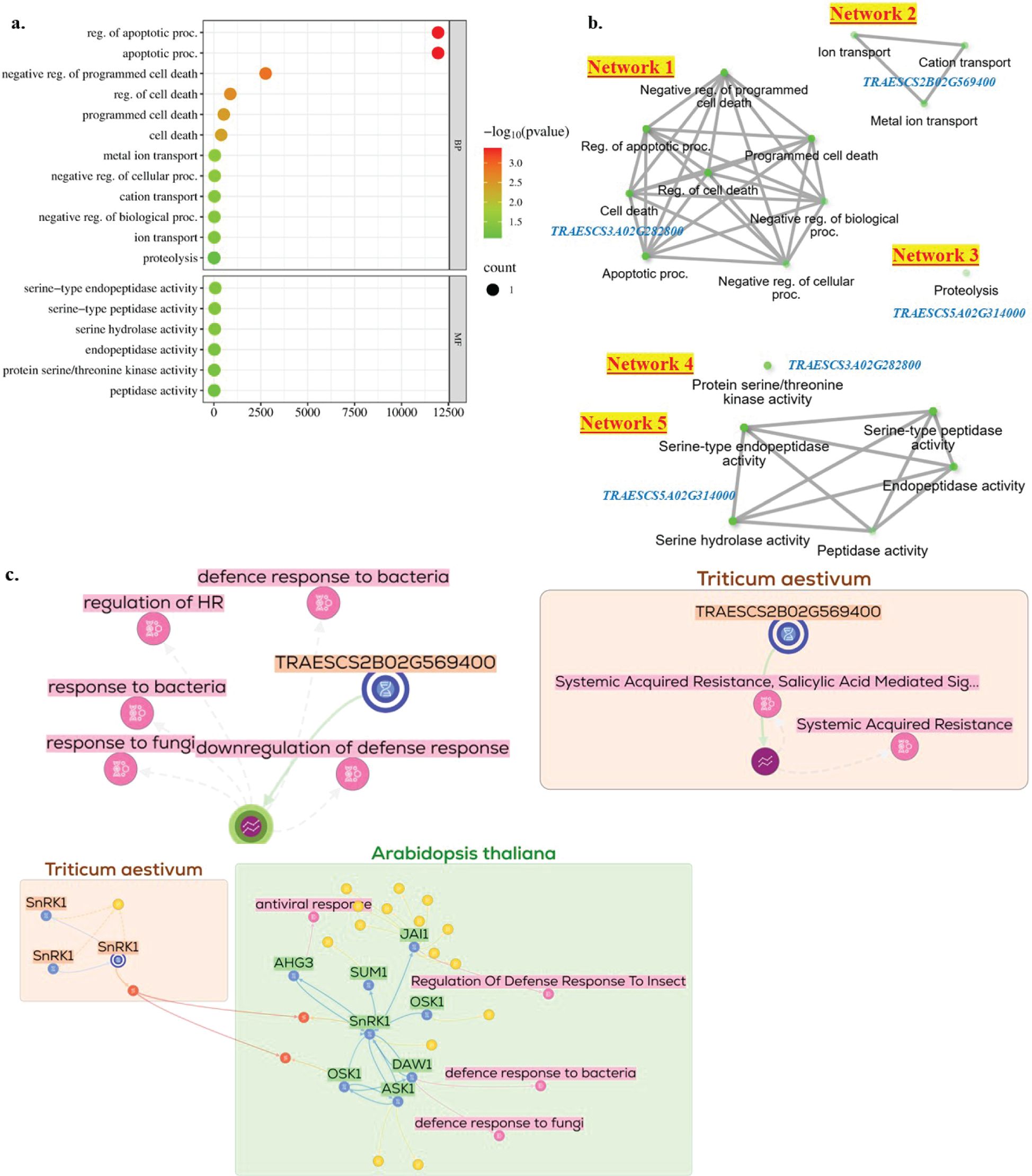
Figure 5. Gene enrichment and gene network of the three gene models harbor markers significantly associated with powdery mildew adult plant resistance in the three tested growing seasons (2022, 2023, and 2024): (a) enrichment pathways identified based on the biological process pathways (BP) and molecular function (MF), (b) gene network of the 12 BP-pathways controlled by the three gene models (Network 1, Network 2, and Network 3) and the six MF pathways (Network 4 and Network 5, (c) Network of TRAESCS2B02G569400 and TRAESCS3A02G282800 in relation to disease resistance in wheat and Arabidopsis genomes, respectively.
3.4 Selection of superior resistant and high-yield genotypes
The number of targeted alleles and different alleles of the significant markers associated with resistance were investigated in the eight selected genotypes as well as the two most resistant Egyptian genotypes. The number of target alleles ranged from 20 in the American genotype “Spinkcota” to 31 in the Egyptian genotype “Giza 171”. The Sudanese genotype “Nelin” carried 24 target alleles, which is quite similar to the other Egyptian genotype “Sohag 5” (Figure 6a). The lowest number of different alleles was 7, which was found between “Carstens Sommerweizen” and both “Koga II” and “Kolben II”. The highest number of different marker alleles was 25 between “Spinkcota” and both “Koga II” and “Kolben II” (Figure 6b). For the two Egyptian genotypes, the highest number of different alleles was 22 between “Solitaire” and both genotypes as well as between “Sohag 5” and both “Koga II” and “Kolben II”. Nelin had 13 different markers between itself and both “Sohag 5” and “Giza 171”.
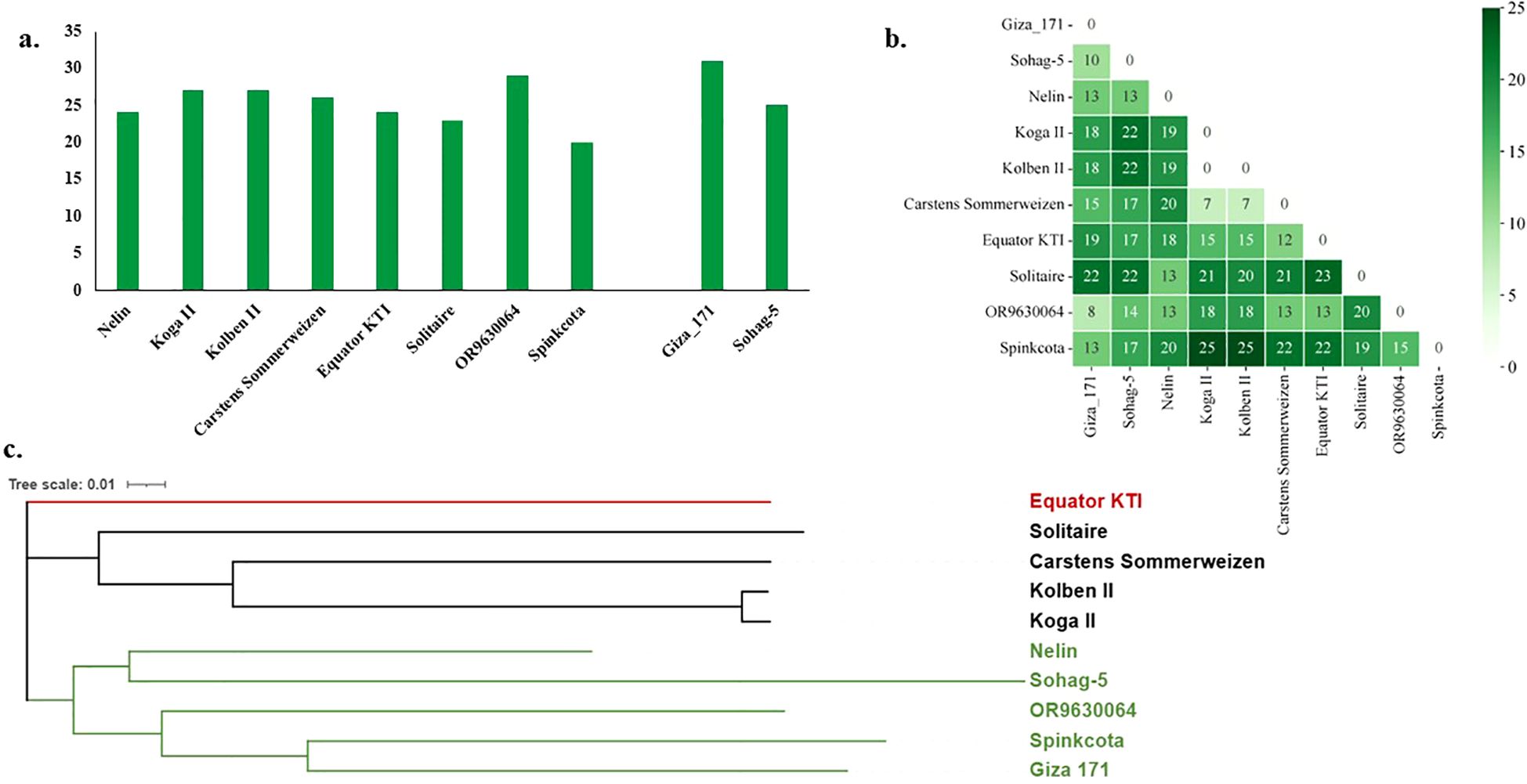
Figure 6. (a) Number of target alleles in each of the eight selected genotypes and the best two resistant Egyptian genotypes, (b) number of different alleles between each pair of the selected genotypes, and (c) phylogeny tree represents the genetic distance among each pair of the selected genotypes.
Moreover, the genetic distance between the eight selected genotypes as well as the Egyptian genotypes was calculated and presented as a matrix (Supplementary Table S8) and phylogenetic tree (Figure 6c). Three clusters were identified to contain all ten genotypes. The two Egyptian genotypes were located in the same cluster as the Sudanese genotype “Nelin” and the two American genotypes. The Kenyan genotypes were located alone in a separate cluster. The German and English genotypes were located in the same cluster together. The greatest distance was “0.451” between the Egyptian genotype “Sohag 5” and the English genotype “Solitaire”. The lowest value of genetic distance was found between the two German genotypes “Koga II” and “Kolben II”, with a value of 0.014.
To provide more understanding of the possibility of using the eight common resistant genotypes in future breeding programs, the yield of these genotypes in comparison with the best two resistant Egyptian cultivars (Sohag 5 and Giza 171) was investigated in disease-infected and noninfected fields (Figure 7). Obviously, the Sudanese genotype “Nelin” had greater TKW in both cropping seasons in infected fields (Figure 7a) and the three cropping seasons under a controlled field (Figure 7b). Furthermore, the yield of the Kenyan genotype “Equator KTI” was almost the same as that of the two Egyptian genotypes in 2022-2023 under disease conditions and in 2021-2022 under noninfected conditions. However, it has an extremely low yield in the 2023-2024 cropping season under powdery mildew-infected fields as well as in 2022-2023 and 2023-2024 under noninfected fields.
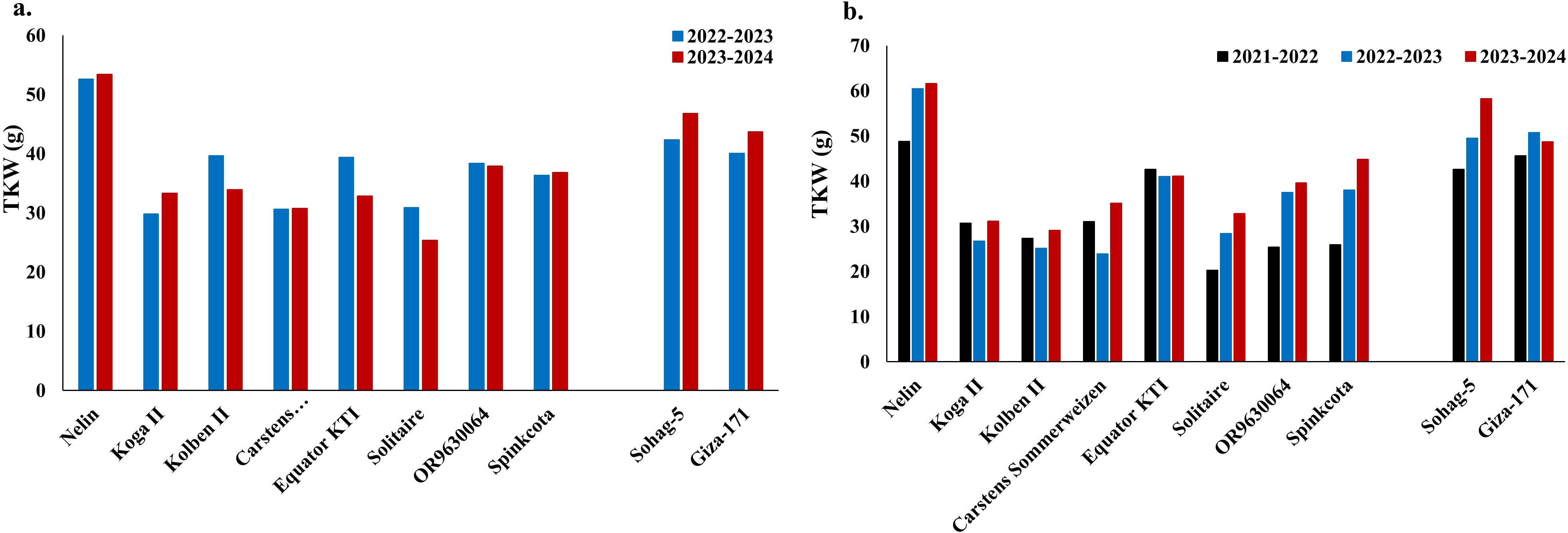
Figure 7. Thousand kernel weight (gm) of the selected eight highly resistance genotypes to powdery mildew in the three growing seasons as well as the best two Egyptian cultivars under powdery mildew infection conditions in two growing seasons (a) and disease-free field in three different growing seasons (b).
Furthermore, the yield components of the ten selected genotypes were measured under noninfected field conditions (Supplementary Figure S5). Notably, the Sudanese genotype “Nelin” had a heading date (HD) similar to that of the Egyptian genotypes, with values of 99 and 88 days in the 2022-2023 and 2023-2024 cropping seasons, respectively. The remaining resistant genotypes had higher HD values than the Egyptian genotypes did, except for “Spinkcota” (Supplementary Figure S5a). The PH values of the Egyptian genotypes were 101.5 cm and 88.5 cm for Giza 171 and 98.5 cm and 92 cm for Sohag 5 in 2022-2023 and 2023-2024, respectively. Nelin was slightly taller than the two Egyptian genotypes were, with PH values of 104 cm, 114.75 cm, and 112.67 cm in the 2021-2022, 2022-2023, and 2023-2024 cropping seasons, respectively (Supplementary Figure S5b). The remaining genotypes could be classified into two groups: shorter genotypes (Solitaire, OR9630064, and Spinkcota) and very tall genotypes in at least one cropping season (Koga II, Kolben II, Carstens Sommerweizen, and Equator KTI). All eight selected genotypes presented spike length (SPL) values within the range of the Egyptian genotypes (8.67–14.33 cm in 2021-2022, 8.5–13.5 cm in 2022-2023, and 8.17–14.34 cm in 2023-2024), except for “OR9630064”, which had a longer spike in 2022-2023 (16.5 cm) (Supplementary Figure S5c). The number of spikes per plant (NS/SP) was similar for all the selected genotypes and the Egyptian genotypes (Supplementary Figure S5d). The GY/SP ratio was lower in all eight resistant genotypes than in the Egyptian genotypes (Supplementary Figure S5e). However, Nelin had the highest GY/SP among the selected genotypes. Similarly, the grain number per spike (GN/SP) was lower in all eight selected genotypes than in the Egyptian genotypes. However, “Spinkcota” and “Nelin” had the highest GN/SP values among the selected genotypes (Supplementary Figure S5f).
4 Discussion
Wheat powdery mildew is a major disease that affects wheat-cropping areas worldwide. Limited information is available regarding resistance to this disease under Egyptian conditions (Mourad et al., 2024a). This study is the first to investigate WPM-APR under Egyptian field conditions. The evaluated plant materials were collected from 22 different countries and have been reported to be genetically diverse and well-adapted to Egyptian fields (Mourad et al., 2020, 2022c). This broad genetic diversity provides valuable resources for improving powdery mildew resistance in Egyptian wheat. WPM-APR was assessed across three cropping seasons (2021-2022, 2022-2023, and 2023-2024). Previous studies have reported the presence of different Bgt isolates in Egyptian fields across different cropping seasons (El-Shamy and Mohamed, 2022; Mourad et al., 2022). Therefore, different isolates have been identified in recent studies.
A set of 20 isolines carrying 20 different WPM resistance genes was evaluated during each cropping season. Notably, different responses of most of the evaluated isolines were recorded in each season, confirming the changes occurred in the Bgt isolates during each cropping season. Furthermore, Mourad et al. (2022) evaluated the same set of isolates at the seedling growth stage using Bgt isolates collected from Egyptian fields in the 2020-2021 and 2021-2022 cropping seasons. A comparison of the results of their study with the results obtained in the present study revealed that the NACG13 gene was highly effective against different Bgt races in both the seedling and adult growth stages. Furthermore, Pm1b, Pm34, Pm36, and MIAG12 were resistant to WPM during the two cropping seasons at the seedling and adult growth stages. Previous studies reported the resistance of these WPM resistance genes under Egyptian conditions (El-Shamy et al., 2016; Abdelrhim et al., 2018; El-Shamy and Mohamed, 2022). These five WPM resistance genes could be good sources of WPM resistance among the three cropping seasons.
4.1 Variation in WPM–adult plant resistance in the tested plant material
The highly significant differences in WPM-APR among the three cropping seasons again confirmed the presence of different Bgt isolates in each cropping season (Supplementary Table S3). Furthermore, the highly significant differences among the tested genotypes represent the high genetic variation among them and the possibility of selecting highly resistant genotypes using this plant set. The high degrees of broad-sense heritability in each cropping season indicate that the phenotypic variation in WPM resistance is mainly due to genotypic variation; thus, the selected resistant genotypes are promising for improving WPM resistance (Table 1). Previous studies reported high degrees of broad-sense heritability for WPM resistance (Hoseinzadeh et al., 2019; Hinterberger et al., 2022; Mourad et al., 2022).
Notably, none of the genotypes in the tested set were immune to WPM during any cropping season, confirming the urgent need to improve WPM in wheat crop under Egyptian field conditions. The lack of immune Egyptian wheat genotypes has been previously reported in the majority of commercial cultivars (El-Shamy et al., 2016; Draz et al., 2019, 2022; Elsayed and Elkot, 2020). Furthermore, most of the genotypes were susceptible to WPM (DS ≥20%). Few genotypes were resistant to WPM, with very few (8 genotypes) common resistant genotypes across the three cropping seasons. These genotypes belong to five different countries, and none of them are Egyptian genotypes, which again confirms the lack of WPM-resistant genotypes in the Egyptian wheat germplasm. As these 8 common genotypes belong to five different countries, crossing these genotypes with Egyptian cultivars could help improve WPM-APR in Egyptian wheat germplasms. The same set was evaluated previously for WPM at the seedling growth stage against Egyptian races, and 19 genotypes were resistant to the different Bgt isolates used in this study (Mourad et al., 2023). Among these 19 genotypes, one English genotype, “Solitaire,” was resistant at the adult growth stage in our recent study during the three different cropping seasons.
4.2 Association mapping of WPM-APR
Two types of markers, GBS-SNP and 25K-SNP, covering different parts of the wheat genome, were used in the GWAS (Liu et al., 2020; Mourad et al., 2022, 2024b, c; Esmail et al., 2023b). Furthermore, different models have been applied in GWASs, which are effective in identifying significant markers and accounting for the effects of population structure (Turuspekov et al., 2016; Kaler et al., 2020; Muhammad et al., 2020; Kiani et al., 2021). Notably, the best GWAS model varied depending on the phenotypic data and the set of markers used (Supplementary Figures S2, S3). Therefore, testing multiple GWAS models to identify markers significantly associated with the studied trait is highly recommended. The significant markers were identified based on -log10 (p-value) = 3.00. It was reported that using more stringent corrections such as Bonferroni can be overly conservative, potentially overlooking biologically meaningful associations, particularly for traits governed by multiple small-effect loci. Our choice was therefore a deliberate compromise to enhance the discovery of candidate loci, with the understanding that the identified associations should be considered suggestive. Therefore, all markers detected in this study at this threshold should be validated in independent populations or through functional characterization to confirm their roles in the traits of interest. There are many earlier studies that validated markers detected at p ≤ 0.001 in different genetic backgrounds such as drought tolerance in wheat, alkaline-saline, and disease resistance (Sallam et al., 2022, 2024; Mourad et al., 2025).
Different significant markers were found to control resistance in each cropping season, with only four common markers, which again confirmed the presence of different Bgt races in each cropping season (Table 3, Figure 3b). Combining the GWAS results from the three cropping seasons with the average, a total of 86 markers across 18 different chromosomes were identified as being significantly associated with resistance. The large number of significant markers across the different chromosomes suggests the complexity of the genetic system controlling the resistance. A similar complex genetic system that controls WPM resistance has been reported previously (Kang et al., 2019; Mohler and Stadlmeier, 2019; Miedaner et al., 2020; Simeone et al., 2020; Alemu et al., 2021; Du et al., 2021; Hinterberger et al., 2022; Jin et al., 2022; Mourad et al., 2023, 2024a). Most of the significant markers presented values of R2< 10%, suggesting that WPM-APR is a quantitative trait that is controlled by many genes with minor effects. Furthermore, most of these markers are located within gene models that control the production of proteins and enzymes that increase wheat resistance to diseases, including powdery mildew. Examples include glutathione S-transferase (Gullner et al., 2018), NBS-LRR-like resistance protein (Oladzad et al., 2023), glucan endo-1,3-beta-glucosidase (Perrot et al., 2022), and serine/threonine-protein kinase (Cao et al., 2011). Furthermore, the “S7A_35630468” marker associated with resistance in the 2022-2023 cropping season was previously reported to control WPM seedling resistance via different Bgt isolates under Egyptian conditions (Mourad et al., 2023). This marker is located in the TraesCS7A02G069700 gene, which is functionally annotated to produce glucan endo-1,3-beta-glucosidase, an enzyme that controls disease resistance in wheat, including powdery mildew (Benbow et al., 2020; Li et al., 2020; Yue et al., 2021; Esmail et al., 2023b).
Furthermore, the “wsnp_CAP8_rep_c3844_1896355” marker associated with resistance in the 2023-2024 cropping season was previously reported to control the disease severity and infection type of adult leaf rust plant resistance under Ethiopian conditions (Lhamo et al., 2023). This marker is located within the TraesCS7A02G529200 gene, which is functionally annotated to control the production of the chaperone protein dnaJ, a protein that has been reported to play an important role in several pathological processes in wheat, such as yellow mosaic virus (Liu et al., 2022).
4.3 Stable genomic regions controlling broad-spectrum resistance to different Bgt isolates
Among the many markers significantly associated with resistance, only four were associated with resistance in at least two of the three cropping seasons (Table 4, Supplementary Table S7). This low number of common significant markers represents the complexity of the wheat genome as well as the continuous changes in Bgt races. Two markers were associated with WPM resistance in the three cropping seasons, while the other two markers were significantly associated with WPM resistance in only two years, which could be due to the effect of genotype × interaction (Eltaher et al., 2021). These four markers were distributed among four different chromosomes. Interestingly, no linkage disequilibrium (LD) was detected between any of these common markers and other significant markers associated with resistance in only one cropping season. This further confirms the complexity of the wheat genome and the challenge of developing wheat genotype with broad-spectrum resistance to WPM (Supplementary Figure S4). Therefore, it is highly recommended that more GWASs be conducted on this serious disease to increase our understanding of broad-spectrum resistance to WPM (Mourad et al., 2024a). The four markers can be converted to KASP markers for further validation in different genetic backgrounds. Moreover, this marker can be used for marker-assisted selection under Egyptian conditions. It was reported that genetic validation assesses whether the same QTL or gene is consistently detected across different locations or years and whether its effect remains significant when evaluated in diverse genetic backgrounds (Sallam et al., 2023).
Only three gene models harboring the common significant markers were detected. The functional annotation of these gene models was directly and indirectly associated with disease resistance in wheat. For example, the TraesCS2B02G569400 gene is functionally annotated to produce heavy metal transport/detoxification superfamily proteins. Previous studies reported that some heavy metal transporter proteins regulate disease resistance (Chu et al., 2022; Liu et al., 2025). Furthermore, the rhomboid-like protein produced by the TraesCS5A02G314000 gene was previously reported to be an important gene for improving wheat resistance to kernel bunt (Singh et al., 2020) and both stripe and leaf rust diseases (Kumar et al., 2020). The third gene was found to control nonspecific serine/threonine protein kinases, which are key members of many powdery mildew resistance genes, such as Pm21 (Cao et al., 2011), Pm13 and Pm4 (Li et al., 2024), as well as the Sr34 stem rust resistance gene (Yu et al., 2023).
Through gene enrichment analysis, three biological process (BP) networks and two molecular function (MF) networks were identified. Each network was regulated by a single gene from the identified gene models, suggesting that each gene functions independently in enhancing WPM resistance. TraesCS3A02G282800 regulates cell death and negatively influences biological processes, playing a crucial role in initiating the plant immune response (Coll et al., 2011). Serine/threonine kinases are among the main kinases that control programmed cell death (apoptosis) (Cross et al., 2000). TraesCS5A02G314000 controls the activity of serine-type peptidase. This enzyme works in the proteolysis of cell proteins. Proteolysis was found to result in a high level of broad-spectrum resistance without any reduction in plant yield (Liu et al., 2024). TraesCS2B02G569400 works in one network to control metal ion/cation transport via the BP pathway. However, its role in MF has not been identified.
Furthermore, known Pm resistance genes located at the same chromosomal positions as the four stable markers were identified to increase our understanding. On chromosome 1A, the Pm3, Pm17, and Pm25 genes have been reported (Chen et al., 1996; Shi et al., 1998; Bougot et al., 2002). However, on the basis of the results of the differential lines, these three genes were not effective against the Egyptian races in the three cropping seasons (Figure 1). Therefore, we can conclude that the stable significant marker on this marker could be associated with a novel resistance gene. The Pm6, Pm26, Pm42, and M1AB10 genes were mapped to the 2B chromosome. Pm6 was included in the differential lines tested in this study and was not effective in resisting Egyptian Bgt races. Unfortunately, no seeds for the Pm26, Pm42, or M1AB10 genes were available for evaluation in our study. However, a previous study reported that Pm26 is located on the short arm of the 2B chromosome in the 26573946:27934056 bp region (Hinterberger et al., 2022). Compared with the location of the stable marker on this chromosome (S2B_760015865), Pm42 is located far from our stable markers (Hua et al., 2009). The Pm64, MIZec1, and M1AB10 genes were mapped to the long arm of 2B (Maxwell et al., 2010; Zhang et al., 2019). However, previous studies reported that the “wsnp_Ex_c24135_33382318” and “S2B_754153049” markers overlapped with the chromosomal position of Pm64 (Lhamo et al., 2025). Hence, our identified marker could not be Pm64. Furthermore, the MIZec1 and M1AB10 genes were mapped to 797 and 799 Mbp, respectively (Lhamo et al., 2025). Since none of the mentioned genes overlapped with the position of the “S2B_760015865” marker, we can conclude that this marker and its harboring gene are novel or unknown genes. Pm44 is located on the 3AS chromosome (Jin et al., 2020). Based on our findings, the “Ra_c88203_468” marker was located on the long arm of this chromosome. Pm2026 and Pm55 are effective resistance genes that have been mapped to the 5AL chromosome (Xu et al., 2008; Zhang et al., 2016; Liu et al., 2017; Lu et al., 2024). However, no information is available about the base pair location of this gene. Therefore, we could not confirm whether the significant marker on this chromosome was associated with Pm2026 and Pm55 due to a lack of information.
4.4 Selection of superior resistant and high-yield genotypes
The best parents for crossing are those with high genetic distance and a high number of different alleles associated with the target allele (Bertan et al., 2007; Mourad et al., 2020; Esmail et al., 2023a; Mourad et al., 2025; Sharshar et al., 2025). Based on this concept, “Solitaire” could be considered the best parent for crossing with the two Egyptian genotypes to improve WPM-APR; however, selecting superior genotypes that are both disease-resistant and high-yielding is challenging. Therefore, it is crucial to consider yield as a selection criterion in addition to a high level of WPM resistance. In this study, eight resistant genotypes were identified as stable across the three cropping seasons. Additionally, two Egyptian genotypes were considered the most resistant Egyptian genotypes, exhibiting resistance in one or two cropping seasons. “Sohag 5” has been reported as a high-yielding durum wheat genotype (Hamada et al., 2016), whereas “Giza 171” is a high-yielding bread wheat genotype resistant to stem and leaf rust (Hamada et al., 2015). Both cultivars displayed an intermediate level of resistance in at least one or two cropping seasons. Therefore, improving their resistance to WPM will result in high-yielding and resistant genotypes. Solitaire had lower yield and yield components than the two other tested genotypes did. Thus, using this genotype as a parent in future breeding programs will increase WPM-APR but may negatively impact the yield of Egyptian wheat germplasm. Among the eight selected stable resistant genotypes, only “Nelin” presented a greater TKW than did the two Egyptian cultivars in both the control and infected fields. Therefore, despite having an intermediate number of different and target alleles and having a low genetic distance from the two Egyptian genotypes, “Nelin” could be a valuable parent in future breeding programs to improve resistance in Egyptian wheat germplasm.
5 Conclusion
In conclusion, the high genetic variation in WPM-APR found in the tested panel confirms the possibility of selecting highly resistant genotypes. The absence of immune genotypes in the three cropping seasons, especially in the Egyptian wheat genotypes, confirms the urgent need to improve WPM resistance. This study is the first to elucidate the genetic control of WPM-APR under Egyptian conditions. Four stable significant markers across the three cropping seasons were identified. These markers are located on four different chromosomes and work separately in different networks, suggesting that they are four different resistance genes. Eight superior resistant genotypes that belong to five different countries other than Egypt were selected. The Sudanese genotype “Nelin” was found to be the best parent for improving WPM resistance and yield traits in Egyptian wheat germplasm.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author/s.
Author contributions
AM: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. AS: Data curation, Writing – review & editing. AB: Writing – review & editing. RH: Writing – review & editing. SE: Writing – review & editing, Data curation.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This paper is based upon work supported by the Academy of Scientific Research and Technology (ASRT) under grant number 87/2021. Molecular marker sequencing was funded by the Alexander von Humboldt Foundation. The publication of this article was funded by the Deutsche Forschungsgemeinschaft (DFG, German Research Foundation) –801 HE 9114/1-1.
Acknowledgments
The authors would like to thank Sakha’s station team of wheat pathology for helping in conducting the experiment.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fagro.2025.1595621/full#supplementary-material
References
Abdelrhim A., Abd-Alla H. M., Abdou E. S., Ismail M. E., and Cowger C.. (2018). Virulence of Egyptian Blumeria graminis f. Sp. tritici population and response of Egyptian wheat cultivars. Plant Dis. 102, 391–397. doi: 10.1094/PDIS-07-17-0975-RE
Ahmed A. A. M., Dawood M. F. A., Elfarash A., Mohamed E. A., Hussein M. Y., Börner A., et al. (2022). Genetic and morpho-physiological analyses of the tolerance and recovery mechanisms in seedling stage spring wheat under drought stress. Front. Genet. 13. doi: 10.3389/fgene.2022.1010272
Alemu A., Brazauskas G., Gaikpa D. S., Henriksson T., Islamov B., Jørgensen L. N., et al. (2021). Genome-wide association analysis and genomic prediction for adult-plant resistance to septoria tritici blotch and powdery mildew in winter wheat. Front. Genet. 12. doi: 10.3389/fgene.2021.661742
Alqudah A. M., Sallam A., Baenziger P. S., and Börner A. (2020). GWAS : Fast-forwarding gene identification and characterization in temperate Cereals : lessons from Barley – A review. J. Adv. Res. 22, 119–135. doi: 10.1016/j.jare.2019.10.013
Andersen E. J., Nepal M. P., Purintun J. M., Nelson D., Mermigka G., and Sarris P. F.. (2020). Wheat disease resistance genes and their diversification through integrated domain fusions. Front. Genet. 11. doi: 10.3389/fgene.2020.00898
Bajgain P., Rouse M. N., Bulli P., Bhavani S., Gordon T., Wanyera R., et al. (2015). Association mapping of North American spring wheat breeding germplasm reveals loci conferring resistance to Ug99 and other African stem rust races. BMC Plant Biol. 15, 249. doi: 10.1186/s12870-015-0628-9
Bates D., Mächler M., Bolker B., and Walker S. (2015). Fitting linear mixed-effects models using lme4. J. Stat. Softw 67, 1–48. doi: 10.18637/JSS.V067.I01
Benbow H. R., Brennan C. J., Zhou B., Christodoulou T., Berry S., Uauy C., et al. (2020). Insights into the resistance of a synthetically-derived wheat to Septoria tritici blotch disease: Less is more. BMC Plant Biol. 20, 1–23. doi: 10.1186/s12870-020-02612-z
Bertan I., De Carvalho F. I. F., and De Oliveira A. C. (2007). Parental selection strategies in plant breeding programs. J. Crop Sci. Biotechnol. 10, 211–222.
Bhadana D., Kaur P., Ramandeep K., Ravat V. K., Kumar R., and Vasistha N. K.. (2025). Genome-wide association study for powdery mildew resistance in CIMMYT’s spring wheat germplasm. Plant Pathol. 74, 455–464. doi: 10.1111/ppa.14031
Bhavani S., Singh P. K., Qureshi N., He X., Biswal A. K., Juliana P., et al. (2021). “Globally important wheat diseases: status, challenges, breeding and genomic tools to enhance resistance durability,” in Genomic Designing for Biotic Stress Resistant Cereal Crops. Ed. Kole C.(New Delhi, India: Springer), 59–128.
Bolser D., Staines D. M., Kersey E., and Kersey P. (2007). “Ensembl Plants: Integrating tools for visualizing, mining, and analyzing plant genomics data,” in Plant Bioinformatics: Mthods and protocols. Ed. Edwards D., 115–140. doi: 10.1007/978-1-4939-3167-5_6
Bougot Y., Lemoine J., Pavoine M. T., Barloy D., and Doussinault G.. (2002). Identification of a microsatellite marker associated with Pm3 resistance alleles to powdery mildew in wheat. Plant Breed 121, 325–329. doi: 10.1046/j.1439-0523.2002.736127.x
Bradbury P. J., Zhang Z., Kroon D. E., Barloy D., and Doussinault G.. (2007). TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 23, 2633–2635. doi: 10.1093/bioinformatics/btm308
Cao A., Xing L., Wang X., Yang X., Wang W., Sun Y., et al. (2011). Serine/threonine kinase gene Stpk-V, a key member of powdery mildew resistance gene Pm21, confers powdery mildew resistance in wheat. Proc. Natl. Acad. Sci. U.S.A. 108, 7727–7732. doi: 10.1073/pnas.1016981108
Chen X., Joens S. S., and Line R. F. (1996). Chromosomal Location of Genes for Resistance to Puccinia striiformis in Seven Wheat Cultivars with Resistance Genes at the Yr3 and Yr4 Loci. Phytopathol 86, 1228–1233. doi: 10.1094/Phyto-86-1228
Cheng P., Guo M., Hao X., Guo X., Yao Q., Guo Q., et al. (2022). Evaluation of powdery mildew resistance and molecular detection of resistance genes in an international wheat collection. Crop Prot 160, 0261–2194. doi: 10.1016/j.cropro.2022.106033
Chu C., Huang R., Liu L., Tang G., Xiao J., Yoo H., et al. (2022). The rice heavy-metal transporter OsNRAMP1 regulates disease resistance by modulating ROS homoeostasis. Plant Cell Environ. 45, 1109–1126. doi: 10.1111/pce.14263
Coll N. S., Epple P., and Dangl J. L. (2011). Programmed cell death in the plant immune system. Cell Death Differ 18, 1247–1256. doi: 10.1038/cdd.2011.37
Costamilan L. M. (2005). Variability of the wheat powdery mildew pathogen Blumeria graminis f. sp. tritici. Fitopatol Bras. 30, 420–422. doi: 10.1590/s0100-41582005000400015
Cross T. G., Scheel-Toellner D., Henriquez N. V., Deacon E., Salmon M., and Lord J. M.. (2000). Serine/threonine protein kinases and apoptosis. Exp. Cell Res. 256, 34–41. doi: 10.1006/excr.2000.4836
Draz I. S., Abdelrhim A. S., Mabrouk O. I., and Esmail S. M. (2023). Potential suppression of broad-spectrum virulence of Blumeria graminis f. sp. tritici population to race-specific resistance genes in hexaploid wheat. J. Plant Pathol. 105, 1483–1496. doi: 10.1007/s42161-023-01463-5
Draz I. S., Elkot A. F., and Abdelrhim A. S. (2022). Allelism and resistance loci of powdery mildew and leaf rust in Egyptian hexaploid bread wheat. Cereal Res. Commun. 50, 85–93. doi: 10.1007/s42976-021-00163-z
Draz I. S., Esmail S. M., Abou-Zeid M. A. E. H., and Essa T. A. E. M. (2019). Powdery mildew susceptibility of spring wheat cultivars as a major constraint on grain yield. Ann. Agric. Sci. 64, 39–45. doi: 10.1016/j.aoas.2019.05.007
Du X., Xu W., Peng C., Li C., Zhang Y., and Hu L.. (2021). Identification and validation of a novel locus, Qpm-3BL, for adult plant resistance to powdery mildew in wheat using multilocus GWAS. BMC Plant Biol. 21, 1–13. doi: 10.1186/s12870-021-03093-4
Elsayed M. and Elkot A. (2020). Molecular identification of some powdery mildew resistance genes in ten Egyptian durum wheat cultivars. J. Plant Prot Pathol. 11, 205–209. doi: 10.21608/jppp.2020.87592
El-Shamy M. M., Emara H. M., and Mohamed M. E. (2016). Virulence analysis of wheat powdery mildew (Blumeria graminis f. sp. tritici) and effective genes in middle Delta, Egypt. Plant Dis. 100, 1927–1930. doi: 10.1094/PDIS-01-16-0130-RE
El-Shamy M. M. and Mohamed M. E. (2022). Virulence analysis of wheat powdery mildew races during 2019–2020 seasons in Egypt. Cereal Res. Commun. 50, 67–73. doi: 10.1007/s42976-021-00156-y
El-Shamy M., Sallam M., and Awad H. (2012). Powdery mildew infection on some Egyptian bread wheat cultivars in relation to environmental conditions. J. Plant Prot Pathol. 3, 363–372. doi: 10.21608/jppp.2012.83777
Eltaher S., Baenziger P. S., Belamkar V., Emara H. A., Nower A. A., Salem K. F. M., et al. (2021). GWAS revealed effect of genotype × environment interactions for grain yield of Nebraska winter wheat. BMC Genomics 22, 1–14. doi: 10.1186/s12864-020-07308-0
Emara H. M., Omar A. F., El-Shamy M. M., Mohamed M. E., Emara H. M., Omar A. F., et al. (2016). Identification of Pm24, Pm35 and Pm37 in thirteen Egyptian bread wheat cultivars using SSR markers. Ciec. e Agrotecnologia 40, 279–287. doi: 10.1590/1413-70542016403036315
Esmail S. M., Omar G. E., El-Orabey W. M., Börner A., and Mourad A. M. I.. (2023a). Exploring the genetic variation of stripe rust foliar and head infection in Egyptian wheat as an effect of climate change. Agronomy 13, 1509. doi: 10.3390/agronomy13061509
Esmail S. M., Omar G. E., and Mourad A. (2023b). In-depth understanding of the genetic control of stripe rust resistance (Puccinia striiformis f. sp. tritici) induced in wheat (Triticum aestivum L.) by Trichoderma asperellum T34. Plant Dis. 107, 457–472. doi: 10.1094/PDIS-07-22-1593-RE
Figueroa M., Hammond-Kosack K. E., and Solomon P. S. (2018). A review of wheat diseases—a field perspective. Mol. Plant Pathol. 19, 1523–1536. doi: 10.1111/mpp.12618
Ge S. X., Jung D., and Yao R. (2022). ShinyGO: a graphical enrichment tool for ani-mals and plants. Bioinformatics 36, 2628–2629. doi: 10.1093/bioinformatics/btz931
Goswami S. K., Singh V., Chakdar H., and Choudhary P. (2018). Harmful effects of fungicides-current status. Int. J. Agric. Environ. Biotechnol. 11, 1011–1019.
Gullner G., Komives T., Király L., and Schröder P. (2018). Glutathione S-Transferase enzymes in plant-pathogen interactions. Fontiers Plant Sci. 9. doi: 10.3389/fpls.2018.01836
Hamada A. A., Abo-Warda A. M., El-Majeed S. A. A., El-Sayed E. A. M., Meshref M. K., Tawfelis M. B., et al. (2015). Giza 171 : A new high yielding bread wheat cultivar. Egypt J. Plant Breed 19, 2063–2079. doi: 10.12816/0031680
Hamada A. A., Abo-Warda A. M., El-Majeed S. A. A., El-Sayed E. A. M., Meshref M. K., Tawfelis M. B., et al. (2016). Sohag 5 : new durum wheat cultivar. Egypt J. Plant Breed 20, 203–211. doi: 10.12816/0031587
Hassani-Pak K., Singh A., Brandizi M., Hearnshaw J., Parsons J. D., Amberkar S., et al. (2021). KnetMiner: a comprehensive approach for supporting evidence-based gene discovery and complex trait analysis across species. Plant Biotechnol. J. 19, 1670–1678. doi: 10.1111/pbi.13583
He H., Liu R., Ma P., Du H., Zhang H., Wu Q., et al. (2021). Characterization of Pm68, a new powdery mildew resistance gene on chromosome 2BS of Greek durum wheat TRI 1796. Theor. Appl. Genet. 134, 53–62. doi: 10.1007/s00122-020-03681-2
Hinterberger V., Douchkov D., Lück S., Kale S., Mascher M., Stein N., et al. (2022). Mining for new sources of resistance to powdery mildew in genetic resources of winter wheat. Front. Plant Sci. 13. doi: 10.3389/fpls.2022.836723
Hoseinzadeh P., Zhou R., Mascher M., Himmelbach A., Niks R. E., Schweizer P., et al. (2019). High resolution genetic and physical mapping of a major powdery mildew resistance locus in barley. Front. Plant Sci. 10. doi: 10.3389/fpls.2019.00146
Hua W., Liu Z., Zhu J., Xie C., Yang T., Zhou Y., et al. (2009). Identification and genetic mapping of pm42, a new recessive wheat powdery mildew resistance gene derived from wild emmer (Triticum turgidum var. dicoccoides). Theor. Appl. Genet. 119, 223–230. doi: 10.1007/s00122-009-1031-4
Huang L. S., Zhang D. Y., Liang D., Yuan L., Zhao J. L., Hu G. S., et al. (2013). Continuous wavelet analysis for diagnosing stress characteristics of leaf powdery mildew. Int. J. Agric. Biol. 15, 34–40.
Hurni S., Brunner S., Stirnweis D., Herren G., Peditto D., McIntosh R. A., et al. (2014). The powdery mildew resistance gene Pm8 derived from rye is suppressed by its wheat ortholog Pm3. Plant J. 79, 904–913. doi: 10.1111/tpj.12593
Jabran M., Ali M. A., Muzammil S., Zahoor A., Ali F., Hussain S., et al. (2024). Exploring the potential of nanomaterials (NMs) as diagnostic tools and disease resistance for crop pathogens. Chem. Biol. Technol. Agric. 11, 1–22. doi: 10.1186/s40538-024-00592-y
Jabran M., Ali M. A., Zahoor A., Muhae-Ud-Din G., Liu T., Chen W., et al. (2023). Intelligent reprogramming of wheat for enhancement of fungal and nematode disease resistance using advanced molecular techniques. Front. Plant Sci. 14. doi: 10.3389/fpls.2023.1132699
Jin P., Guo X., Guo M., Li R., Li Q., Cheng P., et al. (2022). Genome-wide association mapping of resistance to powdery mildew in regional trials of wheat mainly from China. Plant Dis. 106, 2701–2710. doi: 10.1094/PDIS-12-21-2659-RE
Jin Y., Xue F., Zhou Y., et al. (2020). Fine-mapping of the powdery mildew resistance gene mlxbd in the common wheat landrace Xiaobaidong. Plant Dis. 104, 1231–1238. doi: 10.1094/PDIS-07-19-1347-RE
Kaler A. S., Gillman J. D., Beissinger T., and Purcell L. C. (2020). Comparing different statistical models and multiple testing corrections for association mapping in soybean and maize. Front. Plant Sci. 10. doi: 10.3389/fpls.2019.01794
Kang Y., Barry K., Cao F., and Zhou M. (2019). Genome-wide association mapping for adult resistance to powdery mildew in common wheat. Mol. Biol. Rep. 47, 1241–1256. doi: 10.1007/s11033-019-05225-4
Kaur R., Vasistha N. K., Ravat V. K., Mishra V. K., Sharma S., Joshi A. K., et al. (2023). Genome-wide association study reveals novel powdery mildew resistance loci in bread wheat. Plants 12, 1–18. doi: 10.3390/plants12223864
Kiani T., Mehboob F., Hyder M. Z., Zainy Z., Xu L., Huang L., et al. (2021). Control of stripe rust of wheat using indigenous endophytic bacteria at seedling and adult plant stage. Sci. Rep. 11, 1–14. doi: 10.1038/s41598-021-93939-6
Kissoudis C., van de Wiel C., Visser R. G. F., and van der Linden G. (2014). Enhancing crop resilience to combined abiotic and biotic stress through the dissection of physiological and molecular crosstalk. Front. Plant Sci. 5. doi: 10.3389/fpls.2014.00207
Kumar D., Kumar A., Chhokar V., Gangwar O. P., Bhardwaj S. C., Sivasamy M., et al. (2020). Genome-wide association studies in diverse spring wheat panel for stripe, stem, and leaf rust resistance. Front. Plant Sci. 11. doi: 10.3389/fpls.2020.00748
Letunic I. and Bork P. (2021). Interactive tree of life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 49, W293–W296. doi: 10.1093/nar/gkab301
Lhamo D., Li G., Song G., Li X., Sen T. Z., Gu Y. Q., et al. (2025). Genome-wide association studies on resistance to powdery mildew in cultivated emmer wheat. Plant Genome 18, 1–17. doi: 10.1002/tpg2.20493
Lhamo D., Sun Q., Zhang Q., Li X., Fiedler J., Xia G., et al. (2023). Genome-wide association analyses of leaf rust resistance in cultivated emmer wheat. Theor. Appl. Genet. 136, 20. doi: 10.1007/s00122-023-04281-6
Li H., Men W., Ma C., Liu Q., Dong Z., Tian X., et al. (2024). Wheat powdery mildew resistance gene Pm13 encodes a mixed lineage kinase domain-like protein. Nat. Commun. 15, 1–13. doi: 10.1038/s41467-024-46814-7
Li Y., Qiu L., Liu X., Zhang Q., Zhuansun X., Fahima T., et al. (2020). Glycerol-induced powdery mildew resistance in wheat by regulating plant fatty acid metabolism, plant hormones cross-talk, and pathogenesis-related genes. Int. J. Mol. Sci. 21, 673–692. doi: 10.3390/ijms21020673
Liang Y., Xia Y., Chang X., Gong G., Yang J., Hu Y., et al. (2019). Comparative proteomic analysis of wheat carrying Pm40 response to blumeria graminis f. sp. tritici using two-dimensional electrophoresis. Int. J. Mol. Sci. 20, 1–18. doi: 10.3390/ijms20040933
Liu N., Bai G., Lin M., Xu X., and Zheng W.. (2017). Genome-wide association analysis of powdery mildew resistance in U.S. Winter Wheat. Sci. Rep. 7, 1–11. doi: 10.1038/s41598-017-11230-z
Liu H., Hu T., Li F., Wang Y., Mei Y., and Zhou X.. (2025). A key heavy metal-binding protein orchestrates plant resistance against a geminivirus. Fundam Res. doi: 10.1016/j.fmre.2024.12.005
Liu Y., Jackson E., Liu X., Huang X., Hoorn R. A. L., van der Zhang Y., et al. (2024). Proteolysis in plant immunity. Plant Cell 36, 3099–3115. doi: 10.1093/plcell/koae142
Liu C., Sukumaran S., Jarquin D., Gong G., Yang J., Hu Y., et al. (2020). Comparison of array- and sequencing-based markers for genome-wide association mapping and genomic prediction in spring wheat. Crop Sci. 60, 211–225. doi: 10.1002/csc2.20098
Liu T.t., Xu M., Gao S.-q., Zhang Y., Hu Y., Jin P., et al. (2022). Genome-wide identification and analysis of the regulation wheat DnaJ family genes following wheat yellow mosaic virus infection. J. Integr. Agric. 21, 153–169. doi: 10.1016/S2095-3119(21)63619-5
Lu C., Du J., Chen H., Gong S., Jin Y., Meng X., et al. (2024). Wheat Pm55 alleles exhibit distinct interactions with an inhibitor to cause different powdery mildew resistance. Nat. Commun. 15, 1–13. doi: 10.1038/s41467-024-44796-0
Matzen N., Heick T. M., and Jørgensen L. N. (2019). Control of powdery mildew (Blumeria graminis spp.) in cereals by Serenade®ASO (Bacillus amyloliquefaciens (former subtilis) strain QST 713). Biol. Control 139, 104067. doi: 10.1016/j.biocontrol.2019.104067
Maxwell J. J., Lyerly J. H., Cowger C., Marshall D., Brown-Guedira G., and Murphy J. P.. (2009). MlAG12: A Triticum timopheevii-derived powdery mildew resistance gene in common wheat on chromosome 7AL. Theor. Appl. Genet. 119, 1489–1495. doi: 10.1007/s00122-009-1150-y
Maxwell J. J., Lyerly J. H., Srnic G., Parks R., Cowger C., Marshall D., et al. (2010). MlAB10: A Triticum turgidum Subsp. dicoccoides Derived Powdery Mildew Resistance Gene Identified in Common Wheat. Crop Sci. 50, 2261–2267. doi: 10.2135/cropsci2010.04.0195
Miedaner T., Akel W., Flath K., Jacobi A., Taylor M., Longin F., et al. (2020). Molecular tracking of multiple disease resistance in a winter wheat diversity panel. Theor. Appl. Genet. 133, 419–431. doi: 10.1007/s00122-019-03472-4
Mohler V. and Stadlmeier M. (2019). Dynamic QTL for adult plant resistance to powdery mildew in common wheat (Triticum aestivum L.). J. Appl. Genet. 60, 291–300. doi: 10.1007/s13353-019-00518-7
Mondal S., Sallam A., Sehgal D., Sukumaran Sivakumar, Farhad M., Krishnan J. N., et al. (2021). “Advances in breeding for abiotic stress tolerance in wheat,” in Genomic Designing for Abiotic Stress Resistant Cereal Crops, (Germany, Springer), 71–103.
Mourad A. M. I., Ahmed A. A. M., Baenziger P. S., Börner A., and Sallam A.. (2024a). Broad-spectrum resistance to fungal foliar diseases in wheat: recent efforts and achievements. Front. Plant Sci. 15. doi: 10.3389/fpls.2024.1516317
Mourad A. M. I., Alomari D. Z., Alqudah A. M., Sallam A., and Salem K. F. M.. (2019). “Recent Advances in wheat (Triticum spp.) breeding,” in Advances in planr breeding strategies: cereals, (Germany, Springer), 559–593.
Mourad A. M. I., Baghdady S., Abdel-aleem F. A. M., Jazeri R. M., and Börner A.. (2024b). Novel genomic regions and gene models controlling copper and cadmium stress tolerance in wheat seedlings. Agronomy 14, 2876. doi: 10.3390/agronomy14122876
Mourad A. M. I., Belamkar V., and Baenziger P. S. (2020). Molecular genetic analysis of spring wheat core collection using genetic diversity, population structure, and linkage disequilibrium. BMC Genomics 21, 1–12. doi: 10.1186/s12864-020-06835-0
Mourad A. M. I., Draz I. S., Omar G. E., Börner A., and Esmail S. M.. (2022). Genome-Wide Screening of Broad-Spectrum Resistance to Leaf Rust (Puccinia triticina Eriks) in Spring Wheat (Triticum aestivum L.). Front. Plant Sci. 13. doi: 10.3389/fpls.2022.921230
Mourad A. M. I., Esmail S. M., and Boerner A. (2024c). Effectiveness and genetic control of Trichoderma spp. as a biological control of wheat powdery mildew disease. Phytopathol 114, 2221–34. doi: 10.1094/PHYTO-05-24-0157-R
Mourad A. M. I., Hamdy R. M., and Esmail S. M. (2023). Novel genomic regions on chromosome 5B controlling wheat powdery mildew seedling resistance under Egyptian conditions. Front. Plant Sci. 14. doi: 10.3389/fpls.2023.1160657
Mourad A. M. I., Sallam A., Belamkar V., Wegulo S., Bowden R., Jin Y., et al. (2018). Genome-wide association study for identification and validation of novel SNP markers for Sr6 stem rust resistance gene in bread wheat. Front. Plant Sci. 9. doi: 10.3389/fpls.2018.00380
Mourad A. M. I., Sallam A., Farghaly K. A., and Boerner A. (2025). Detailed genetic analyses highlight genetic variation and genomic regions for lead tolerance in spring wheat. Front. Agron. 7. doi: 10.3389/fagro.2025.1428366
Muhammad A., Hu W., Li Z., Li J., Xie G., Wang J., et al. (2020). Appraising the genetic architecture of Kernel traits in hexaploid wheat using GWAS. Int. J. Mol. Sci. 21, 1–21. doi: 10.3390/ijms21165649
Oladzad A., Roy J., Mamidi S., Miklas P. N., Lee R., Clevenger J., et al. (2023). Linked candidate genes of different functions for white mold resistance in common bean (Phaseolus vulgaris L) are identified by multiple QTL mapping approaches. Front. Plant Sci. 14. doi: 10.3389/fpls.2023.1233285
Paux E., Sourdille P., Salse J., Saintenac C., Choulet F., Leroy P., et al. (2008). A physical map of the 1-gigabase bread wheat chromosome 3B. Science 80-) 322, 101–104. doi: 10.1126/science.1161847
Pearce S., Vazquez-Gross H., Herin S. Y., Saintenac C., Choulet F., Leroy P., et al. (2015). WheatExp: An RNA-seq expression database for polyploid wheat. BMC Plant Biol. 15, 1–8. doi: 10.1186/s12870-015-0692-1
Perrot T., Pauly M., and Ramírez V. (2022). Emerging roles of β-glucanases in plant development and adaptative responses. Plants 11, 1119. doi: 10.3390/plants11091119
Peterson R., Campbell A., and Hannah A. (1948). A Diagrammitic scale for estimating rust intensity on leaves and stems of cereals. Can. J. Res. 26, 496–500. doi: 10.1139/cjr48c-033
Roelfs A. P., Singh R. P., and Saari E. E. (1992). Rust Diseases of Wheat: Concepts and methods of disease management (Mexico. D.F: CYMMIT).
Sallam A., Alqudah A. M., Baenziger P. S., and Rasheed A. (2023). Editorial: Genetic validation and its role in crop improvement. Front. Genet. 13. doi: 10.3389/fgene.2022.1078246
Sallam A., Dawood M. F. A., Jarquín D., Mohamed E. A., Hussein M. Y., Börner A., et al. (2024). Genome-wide scanning to identify and validate single nucleotide polymorphism markers associated with drought tolerance in spring wheat seedlings. Plant Genome 17, e20444, 1–26. doi: 10.1002/tpg2.20444
Sallam A., Eltaher S., Alqudah A. M., Belamkar V., and Baenziger P. S.. (2022). Combined GWAS and QTL mapping revealed candidate genes and SNP network controlling recovery and tolerance traits associated with drought tolerance in seedling winter wheat. Genomics 114, 110358. doi: 10.1016/j.ygeno.2022.110358
Shahin A., Ashmawy M., Esmail S., and El-Moghazy S. (2019). Biocontrol of wheat powdery mildew disease under field conditions in Egypt. Zagazig J. Agric. Res. 46, 2255–2270. doi: 10.21608/zjar.2019.65078
Sharshar A., Esmail S. M., Elmoghazy D. E., Elhag W. A., Genedy M. S., and Mourad A.. (2025). Effect of combined alkaline-saline and stripe rust stresses on physiological and yield traits in bread wheat. J. Crop Heal 77, 1–18. doi: 10.1007/s10343-024-01067-8
Shi A. N., Leath S., and Murphy J. P. (1998). A major gene for powdery mildew resistance transferred to common wheat from wild einkorn wheat. Phytopathology 88, 144–147. doi: 10.1094/PHYTO.1998.88.2.144
Simeone R., Piarulli L., Nigro D., Signorile M. A., Blanco E., Mangini G., et al. (2020). Mapping powdery mildew (Blumeria graminis f. sp. tritici) resistance inwild and cultivated tetraploid wheats. Int. J. Mol. Sci. 21, 1–19. doi: 10.3390/ijms21217910
Singh S., Sehgal D., Kumar S., Arif M. A. R., Vikram P., Sansaloni C. P., et al. (2020). GWAS revealed a novel resistance locus on chromosome 4D for the quarantine disease Karnal bunt in diverse wheat pre-breeding germplasm. Sci. Rep. 10, 1–11. doi: 10.1038/s41598-020-62711-7
Singh R. P., Singh P. K., Rutkoski J., Hodson D. P., He X., Jørgensen L. N., et al. (2016). Disease impact on wheat yield potential and prospects of genetic control. Annu. Rev. Phytopathol. 54, 303–322. doi: 10.1146/annurev-phyto-080615-095835
Turuspekov Y., Ormanbekova D., Rsaliev A., and Abugalieva S. (2016). Genome-wide association study on stem rust resistance in Kazakh spring barley lines. BMC Plant Biol. 16, 14–81. doi: 10.1186/s12870-015-0686-z
Xu H., Yao G., Xiong L., Yang L., Jiang Y., Fu B., et al. (2008). Identification and mapping of pm2026: A recessive powdery mildew resistance gene in an einkorn (Triticum monococcum L.) accession. Theor. Appl. Genet. 117, 471–477. doi: 10.1007/s00122-008-0791-6
Yin L., Zhang H., Tang Z., Xu J., Yin D., Zhang Z., et al. (2021). rMVP: A memory-efficient, visualization-enhanced, and parallel-accelerated tool for genome-wide association study. Genomics Proteomics Bioinf. 19, 619–628. doi: 10.1016/J.GPB.2020.10.007
Yu G., Matny O., Gourdoupis S., Rayapuram N., Aljedaani F. R., Wang Y. L., et al. (2023). The wheat stem rust resistance gene Sr43 encodes an unusual protein kinase. Nat. Genet. 55, 921–926. doi: 10.1038/s41588-023-01402-1
Yue J., Wang Y., Jiao J., and Wang H. (2021). Comparative transcriptomic and metabolic profiling provides insight into the mechanism by which the autophagy inhibitor 3-MA enhances salt stress sensitivity in wheat seedlings. BMC Plant Biol. 21, 577. doi: 10.1186/s12870-021-03351-5
Zhang S., Huang Z., Xu H., Liu Q., Jiang Z., Yin C., et al. (2024). Biological control of wheat powdery mildew disease by the termite-associated fungus Aspergillus chevalieri BYST01 and potential role of secondary metabolites. Pest Manag Sci. 80, 2011–2020. doi: 10.1002/ps.7938
Zhang R., Sun B., Chen J., Cao A., Xing L., Feng Y., et al. (2016). Pm55, a developmental-stage and tissue-specific powdery mildew resistance gene introgressed from Dasypyrum villosum into common wheat. Theor. Appl. Genet. 129, 1975–1984. doi: 10.1007/s00122-016-2753-8
Keywords: functional annotation, gene enrichment, gene model, gene network, GWAS, yield component
Citation: Mourad AMI, Sallam A, Börner A, Hamdy RM and Esmail SM (2025) Discovering stable genomic regions linked to powdery mildew adult-plant resistance in wheat. Front. Agron. 7:1595621. doi: 10.3389/fagro.2025.1595621
Received: 18 March 2025; Accepted: 05 June 2025;
Published: 04 September 2025.
Edited by:
Hakim Manghwar, Lushan Botanical Garden (CAS), ChinaReviewed by:
Muhammad Jabran, Chinese Academy of Agricultural Sciences (CAAS), ChinaDeepa Bhadana, Chaudhary Charan Singh University, India
Copyright © 2025 Mourad, Sallam, Börner, Hamdy and Esmail. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Amira M. I. Mourad, bW91cmFkQGlway1nYXRlcnNsZWJlbi5kZQ==; YW1pcmFfbW91cmFkQGF1bi5lZHUuZWc=
†Present address: Ahmed Sallam, Faculty of Biotechnology, Badr University in Assiut (BUA), Assuit, Egypt
‡ORCID: Amira M.I. Mourad, orcid.org/0000-0001-6879-2559
Andreas Börner, orcid.org/0000-0003-3301-9026
Ahmed Sallam, orcid.org/0000-0002-3917-648X