 Cátia Brito1,2*
Cátia Brito1,2* Sandra Martins1,2
Sandra Martins1,2 Francisco Saraiva1
Francisco Saraiva1 João Roque1
João Roque1 Margarida Arrobas3,4
Margarida Arrobas3,4 Manuel Rodrigues3,4
Manuel Rodrigues3,4 Carlos Correia1,2
Carlos Correia1,2- 1Centre for the Research and Technology of Agro-Environmental and Biological Sciences (CITAB), University of Trás-os-Montes and Alto Douro (UTAD), Vila Real, Portugal
- 2Inov4Agro–Institute for Innovation, Capacity Building and Sustainability of Agri-Food Production, University of Trás-os-Montes and Alto Douro (UTAD), Vila Real, Portugal
- 3Centro de Investigação de Montanha (CIMO), Instituto Politécnico de Bragança, Bragança, Portugal
- 4Laboratório para a Sustentabilidade e Tecnologia em Regiões de Montanha, Instituto Politécnico de Bragança, Bragança, Portugal
Well-managed cover crops are gaining traction as an alternative to conventional tillage, offering a range of agronomic and ecological benefits. This field study, conducted in a commercial vineyard in the Douro region (Portugal) during the 2022/2023 growing season, evaluated the decomposition dynamics and nutrient release from different legume cover crop residues over a period of 183 days after sowing and 114 days after rolling. These mixtures consisted of subterranean clovers (SC), low-growing (LG), and moderately-growing (MG) legumes. MG legumes produced the highest biomass (7.8 t ha−1), followed by LG (7.2 t ha−1) and SC (5.9 kg ha−1). MG residues decomposed more rapidly, potentially meeting early grapevine nutrient demands. SC residues decomposed more slowly, retaining nutrients over longer periods and acting as a sustained source of fertility, while LG legumes provided a balance between rapid and gradual nutrient release. Biomass production peaked before rolling termination, suggesting that the lower canopy foliage might have started decomposing as the crops reached maximum vegetative expansion and early seed formation, reflecting the dry, hot environmental conditions during the study year. The observed nutrient release from cover crop residues, particularly during the first 45 days post-termination, suggested a potential to support grapevine nutrient requirements, although direct uptake by grapevines was not assessed in this study. SC residues retained higher levels of key macronutrients and micronutrients over time. These findings highlight the importance of selecting legume cover crop mixtures aligned with vineyard management goals, whether rapid nutrient supply or sustained soil fertility. The study also underscores the influence of environmental conditions, residue composition and termination timing on nutrient dynamics. Long-term research is recommended to evaluate cover crops’ chemical composition, cumulative effects on soil organic carbon (C) and nutrients, microbial activity and effects on grapevine performance across diverse Mediterranean environments.
1 Introduction
The use of cover crops for permanent or seasonal soil coverage in vineyards has gained significant traction in various wine-growing regions as an alternative to conventional tillage (Marques et al., 2020; Abad et al., 2021a, 2023). Although conventional tillage is widespread, it often leaves soils vulnerable to runoff and nutrient loss, degrades soil organic matter, and reduces water-holding capacity (Bordoni et al., 2019; Marques et al., 2020).
In contrast, well-managed cover crops offer numerous agronomic and ecological benefits, including reduced soil erosion, improved soil organic matter content, carbon sequestration, enhanced aggregate stability, increased water infiltration, and improved soil fertility (Bordoni et al., 2019; Abad et al., 2021a, 2023; Opoku et al., 2024; Çerçioğlu et al., 2025). Moreover, cover crops enhance soil microbial diversity, support auxiliary insect populations, and improve vineyard landscapes’ ecological and aesthetic value, which is particularly relevant in wine tourism regions (Marques et al., 2020; Abad et al., 2021a, 2023; Opoku et al., 2024; Çerçioğlu et al., 2025).
These benefits are particularly relevant in Mediterranean steep-sloped vineyards like those in the Douro Demarcated Region, one of the most important areas for high-quality wine production in Portugal. In 2024, Portugal had 171,264 hectares of vineyards, with the Douro region alone accounting for 43,898 hectares, making it the country’s largest winegrowing region by planted area (IVV, 2025). Those regions face unique challenges, including prolonged drought periods followed by intense rainfall events and climate change impacts such as reduced annual precipitation, heightened evapotranspiration, and rising temperatures (IPCC, 2023). Furthermore, Mediterranean soils are often shallow and fragile, making them highly prone to erosion (Ferreira et al., 2022). Steep slope cultivation exacerbates these risks, with soil losses usually exceeding natural formation rates (1 t ha−1 yr−1), threatening agroecosystem sustainability and increasing downstream risks like siltation and eutrophication of water bodies (FAO, 2015; Ferreira et al., 2022).
While cover crops’ soil and ecological benefits are well-documented (Blanco-Canqui et al., 2015; Abad et al., 2021a, 2023; Çerçioğlu et al., 2025), their adoption remains low. Challenges such as labor-intensive management, potential competition with grapevines for resources and inconsistent impacts on grapevine performance, yield, and quality have limited their broader application (Lacey et al., 2020; Abad et al., 2021b; Zumkeller et al., 2023). Managing the balance between the benefits of cover crops and their competition with grapevines, especially for water and nutrients during critical growth phases, is essential for their successful integration into vineyard systems (Abad et al., 2021a, 2021b; Zumkeller et al., 2023). Proper timing of cover crop termination can mitigate these risks while allowing residues to decompose and release nutrients into the soil.
Cover crops play a dual role in nutrient dynamics by taking up nutrients during their growth and returning them to the soil through microbial decomposition (Ordóñez-Fernández et al., 2018; Poudel et al., 2023). The rate and timing of nutrient release depend on various factors, including cover crop species (e.g., legumes vs. non-legumes), residue chemical composition [e.g., C/nitrogen (N) ratio], biomass management and environmental conditions (Ordóñez-Fernández et al., 2018; Poudel et al., 2023).
Managing cover crop residues is, therefore, critical to harness their benefits. For instance, incorporating residues into the soil accelerates decomposition but can lead to nutrient losses via leaching or volatilization, while leaving residues on the surface as mulch tends to slow decomposition, conserve soil moisture, regulate soil temperature, and protect against erosion (Medrano et al., 2015; Iqbal et al., 2020).
The alignment of nutrient release with grapevine nutrient demands is essential for maximizing the benefits of cover crops. For instance, the nitrogen uptake from the soil is low from bud break to the 5–6 leaf stage, as the vine relies primarily on nitrogen reserves to support initial growth, then uptake becomes more significant from flowering, when root activity increases (Verdenal et al., 2021). Despite the recognized importance of nutrients provided by cover crop residues for supporting cash crops, there is limited information on residue decomposition and nutrient release in vineyards, particularly under Mediterranean conditions characterized by hot, dry summers. A deeper understanding of cover crop decomposition dynamics is needed to guide the selection of suitable species, improve cropping system management for sustained high productivity, and synchronize cover crop termination with cash crop development (Adhikari et al., 2024).
Leguminous cover crops offer significant economic and environmental advantages due to their unique ability to fix atmospheric N through symbiosis with Rhizobium bacteria, enhancing soil fertility by increasing total and mineral N content (Ordóñez-Fernández et al., 2018; Abad et al., 2021a; Çerçioğlu et al., 2025). In Mediterranean environments, self-reseeding annual legumes have been considered suitable species for cover cropping (Mauromicale et al., 2010; Ordóñez-Fernández et al., 2018; Abad et al., 2021a). In addition to species selection, the growth habit of cover crops is an important factor to consider, particularly in semi-arid regions. Larger, more vigorous species generally offer better weed suppression, erosion control, and produce higher biomass, while smaller, low-growing species can help to reduce water competition with grapevines (Blanco-Canqui et al., 2015; Abad et al., 2021a; 2021b).
This study aims to evaluate the decomposition and nutrient release dynamics of three legume-based cover crop mixtures with different growth habits, terminated in spring in a steep-sloped Mediterranean vineyard. By monitoring biomass loss and macro- and micronutrient release from the residues over time (spring and summer), the research seeks to understand how different legume mixtures influence the timing and quantity of nutrient availability from decomposing plant material. The findings aim to support cover crop management strategies that enhance nutrient cycling in the vineyard system while also contributing to soil conservation.
2 Material and methods
2.1 Experimental conditions
The experiment was carried out in the 2022/2023 growing season and took place in Quinta do Bomfim, Pinhão, Portugal (41°11’29”N 7°32’11”W; 150 m above sea level), Cima Corgo subregion of the Douro Demarcated Region (RDD), on a rainfed commercial vineyard (Vitis vinifera L., cv. Touriga Nacional, grafted on 196–17 rootstock). The vineyard was planted in 2015 at a 2.2 m × 1.0 m spacing. Vines were trained on a vertical ascending monoplane pruned on a unilateral Royat Cordon system. Row’s orientation was northeast-southwest with an average slope of 28%. According to the Köppen-Geiger classification, the climate of the study site is typically Mediterranean, with a hot summer (Csa) (IPMA, 2025). The monthly meteorological data for the study period, including temperature, precipitation, and relative humidity, recorded by a meteorological station located at the study site, are presented in Figure 1.
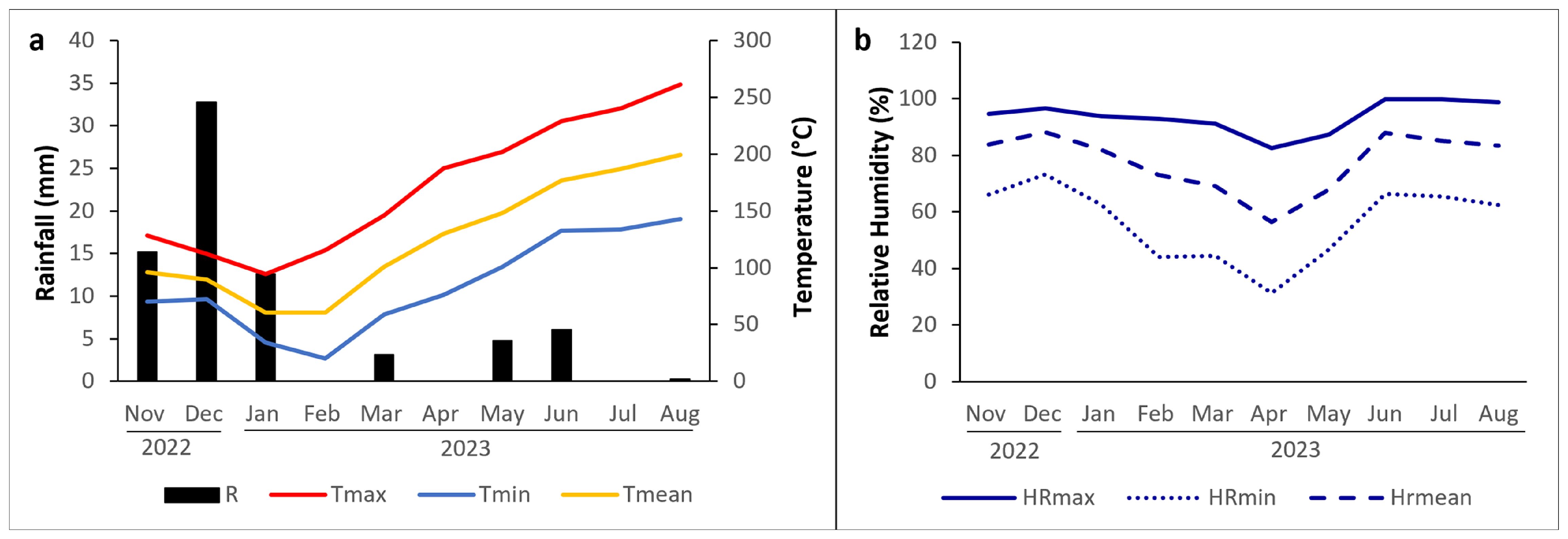
Figure 1. (a) Monthly average minimum temperature (Tmin), maximum temperature (Tmax), and medium temperature (Tmean), along with total accumulated precipitation for the study period; (b) Monthly average maximum (HRmax), minimum (HRmin), and mean relative humidity (HRmean) during the same period.
The soil physicochemical properties of the experimental plot were determined before the experiment started and are presented in Table 1.
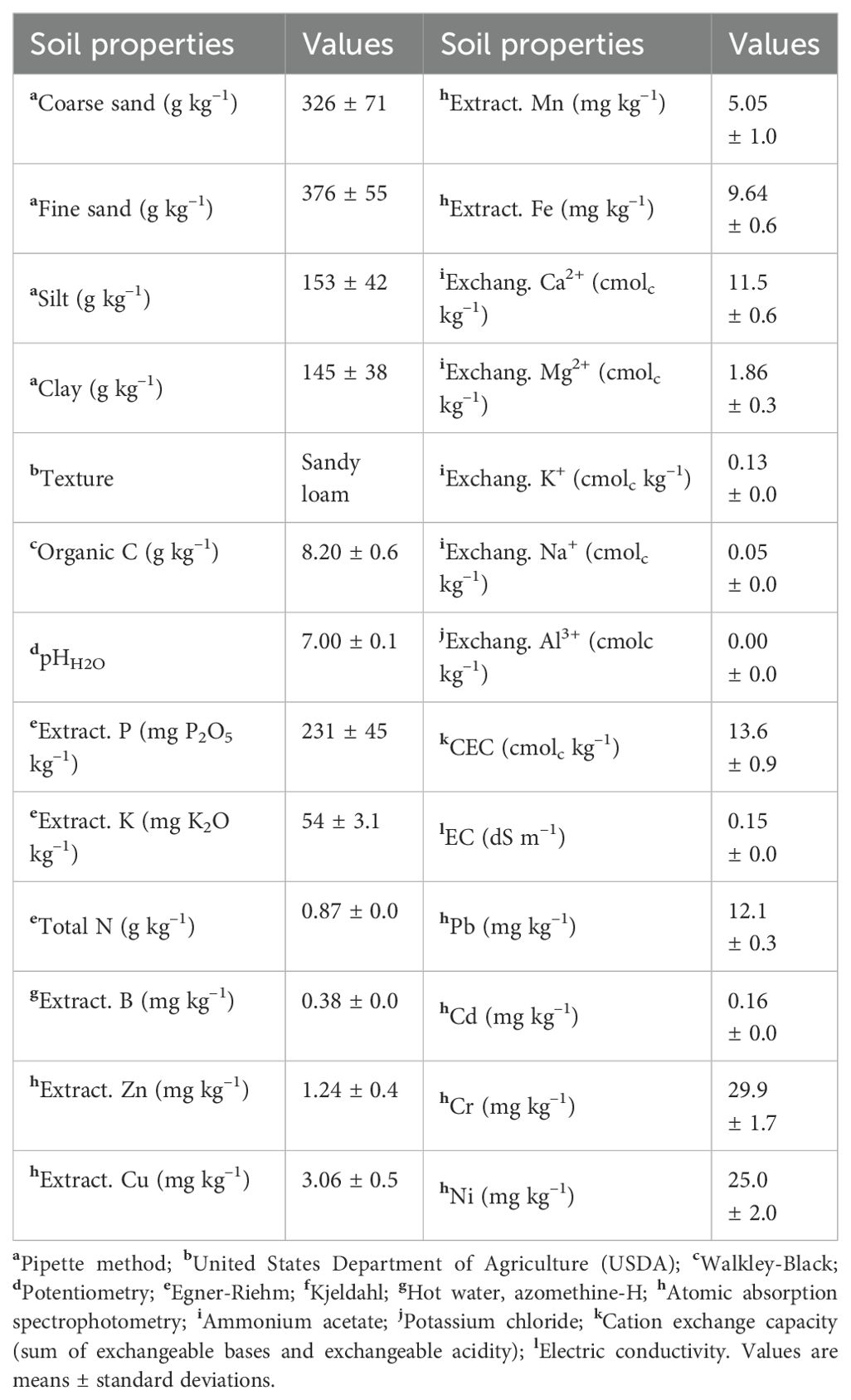
Table 1. Selected physicochemical soil properties (0-0.20 m) before the trial started.
2.2 Experimental setup and monitoring
The experiment was installed in the autumn of 2022 and consisted of three soil management systems with different mixtures of legume cover crops sown in the inter-rows: Subterranean clover (SC) (Trifolium subterraneum, ssp. subterraneum, cvs. Losa, Dalkeith, Seaton Park, Denmark, and Campeda); Low-growing (LG) legume mixture, a commercial product (Ferticover I LEG AC; Fertiprado Vaiamonte, Portugal) consisting of Ornithopus sativus, Trifolium glanduliferum, Trifolium michelianum, Trifolium resupinatum ssp. Resupinatum, Trifolium spumosum, Trifolium subterraneum ssp. subterraneum; Trifolium subterraneum ssp. yanninicum); and Moderately growing (MG) legume mixture, other commercial mixture (Ferticover II LEG AC; Fertiprado Vaiamonte, Portugal), consisting of Medicago polymorpha, Ornithopus sativus, Trifolium glanduliferum, Trifolium incarnatum, Trifolium michelianum, Trifolium resupinatum ssp. resupinatum, Trifolium subterraneum ssp. subterraneum and Trifolium subterraneum ssp. yanninicum. The subterranean clover cultivar mixture (SC) was formulated from pure seeds with the aim of creating a ground cover with a lower growth habit than any commercially available mixture. Each cultivar was included in equal proportion (20%). In the case of the commercial mixtures, the quantitative composition is not disclosed on the product label due to commercial confidentiality. The rationale behind the selection of the three treatments was to cover a gradient of canopy height, ranging from very low-growing to moderately tall mixtures.
The experimental layout followed a randomized complete block design with three blocks, each containing all three treatments (SC, LG, and MG), resulting in a total of nine experimental units. Each experimental unit consisted of three vine rows, each 25–30 meters in length. Prior to sowing, 250 kg ha−1 of single superphosphate (18% P2O5) was applied and incorporated into the soil to a depth of 15 cm using a cultivator. Cover crops were sown at a rate of 20 kg ha−1 on October 1, 2022, and subsequently incorporated into the soil with a light harrow to ensure a sowing depth not exceeding 5 cm. The sown cover strip occupied 60% of the vineyard area. The cover crops were maintained as live vegetation during the autumn and winter months. On May 2, 2023, 183 days after sowing (DAS), at the typical onset of water competition with grapevines, the cover crops were rolled down and left on the soil surface as mulch for the spring and summer periods. Weed control along the vine rows was carried out on February 10, 2022, using the commercial herbicide Roundup® GPS Plus (active ingredient: glyphosate 480 g L−1 in the form of potassium salts) at 0.75 L ha–1, Zagaia (active ingredient: flazasulfuron 25%) at 800 g ha–1, and Zetrola (active ingredient: 100 g L−1 or 9.6% (w/w) of propaquizafop) at 4.8 L ha–1. All other vineyard management practices, such as pruning and plant protection treatments, were carried out by the grower in accordance with standard regional practices. No fertilization was applied to the vineyard during the trial period.
Periodic aboveground biomass samples were collected to monitor biomass accumulation and nutrient concentrations over time. Sampling from 0.1 m² quadrats was randomly conducted, with three samples per row in each block and treatment. Sampling dates and their corresponding DAS or days after rolling (DAR) are as follows: March 28, 2023 (148 DAS); April 11, 2023 (162 DAS), and May 2, 2023 (183 DAS and rolling down date). Biomass assessment was performed at the time of rolling and on June 16, 2023 (45 DAR), July 19, 2023 (78 DAR), and August 24, 2023 (114 DAR).
Phenological data for the 2023 growing season were obtained from the reference plot network of the Viticultural Observatory from the Association for the Development of Viticulture in the Douro Region (ADVID) for Touriga Nacional in the Cima Corgo subregion of RDD. According to this dataset, bud burst (green tip) to bloom occurred over 49 days, approximately between the third week of March and early May. Bloom to cluster closure followed over a 36-day period, concluding by the first week of June. The transition from cluster closure to veraison occurred across 34 days, between mid-June and mid-July. Finally, the period from veraison to harvest lasted 57 days, extending from mid-July to the third week of September. In total, the phenological cycle spanned 177 days from budburst to harvest (ADVID, 2023). Meteorological data for the periods between each sampling event, including temperature, precipitation, and relative humidity, were recorded by a meteorological station located at the study site and are presented in Table 2.
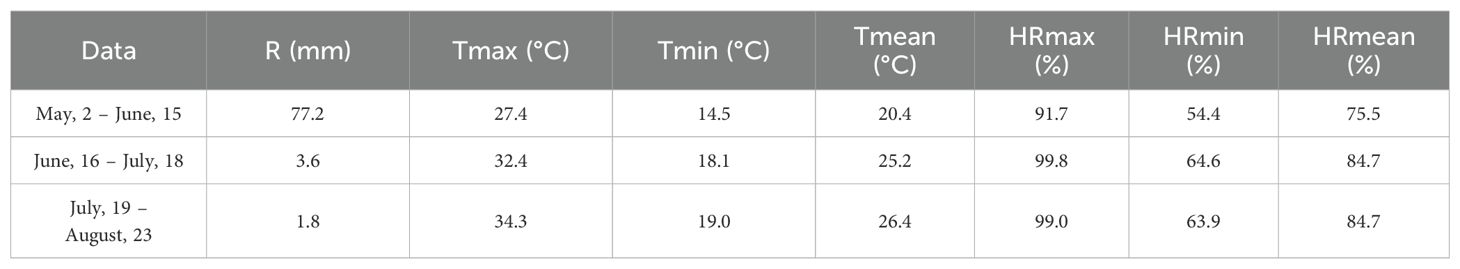
Table 2. Total accumulated precipitation (R), average maximum temperature (Tmax), minimum temperature (Tmin), mean temperature (Tmean), and average maximum (HRmax), minimum (HRmin), and mean relative humidity (HRmean) for each sampling period.
2.3 Residue processing
The collected biomass was gently cleaned to remove accumulated soil when necessary and then divided into small pieces. The samples were oven-dried at 75°C (Selecta DRY-GIG 2003741, J.P. Selecta, Barcelona) until they reached a constant weight, a difference of less than 0.1% of the sample’s total mass between two consecutive weighings at 24-hour intervals. After drying, the biomass was weighed to determine the dry matter content. Cover crop biomass production was estimated by scaling up the weight of biomass collected in the 0.1 m2 quadrat, considering that only 60% of the vineyard floor is covered by vegetation.
2.4 Nutrient in dry biomass
Dried plant samples were ground to pass through a 1 mm sieve using a Cyclotec mill (Tecator Cyclotec 1093, FOSS, Denmark) and subsequently analyzed for elemental composition. Total N was quantified using the Kjeldahl method, which involves sample mineralization with sulfuric acid and a copper-based catalyst, followed by distillation with sodium hydroxide. The ammonia (NH3) released in the steam was collected and quantified by titration using an autoanalyzer Kjeltec 8400 (FOSS, Denmark). For boron (B) analysis, samples were incinerated in the presence of calcium oxide, and B concentration was determined colorimetrically using the azomethine-H method with a UV/VIS spectrophotometer (T80+, PG Instruments, Germany). The concentrations of other nutrients were determined following microwave-assisted acid digestion with nitric acid (MARS XPress, CEM Corporation, USA). Phosphorus (P) was measured colorimetrically using the molybdenum blue method, in which ascorbic acid served as the reducing agent, with detection via the same spectrophotometer (T80+, PG Instruments, Germany). Cation concentrations, including potassium (K), calcium (Ca), magnesium (Mg), iron (Fe), copper (Cu), zinc (Zn), and manganese (Mn), were determined using atomic absorption spectrophotometry (Perkin Elmer Pinnacle 900T, Perkin Elmer, USA). For further methodological details, readers are referred to Temminghoff and Houba (2004).
2.5 Cover crop residue decomposition and nutrient release
The decomposition of cover crop residues was estimated by measuring the aboveground biomass remaining on the soil surface after being rolled down from a 0.1 m2 quadrat. Measurements were taken at different times, on the rolling down day, and at 45, 78, and 144 DAR. The percentage of biomass remaining (BR) and nutrient remaining (NR) at a given time was calculated using the formula (Poudel et al., 2023):
where Xt represents the biomass or nutrients at a given time t, and X0 is the initial cover crop biomass or nutrient mass.
2.6 Statistical analysis
Statistical analyses were performed using JMP Pro 17 (SAS Institute Inc., Cary, NC, USA). Before performing the analysis of variance (ANOVA for a randomized complete block design), assumptions were tested (homogeneity of variances with Levene’s mean test and normality with the Kolmogorov-Smirnov test). Data was analyzed considering the effect of blocks and cover crops on measured variables at each sampling time (DAS and DAR). Analyses were conducted separately for each sampling date; no formal statistical comparisons were made across dates. When significant differences were observed, means were separated using Tukey’s post-hoc test at the 5% significance level.
3 Results
In general, no significant differences were observed between blocks, indicating a high degree of plot homogeneity and limited usefulness of the blocking structure. Consequently, only the treatment effects were presented in the figures. However, a summary of the results from the analysis of variance for all measured variables was provided as Supplementary Material (Supplementary Tables S1, S2).
3.1 Cover crop biomass production and nutrients provision capacity
The biomass production of cover crops and the nutrient content (C, N, P, K, Ca, Mg, B, Fe, Mn, Zn, and Cu) in their tissues were evaluated at 148, 162, and 183 DAS, as shown in Figure 2.
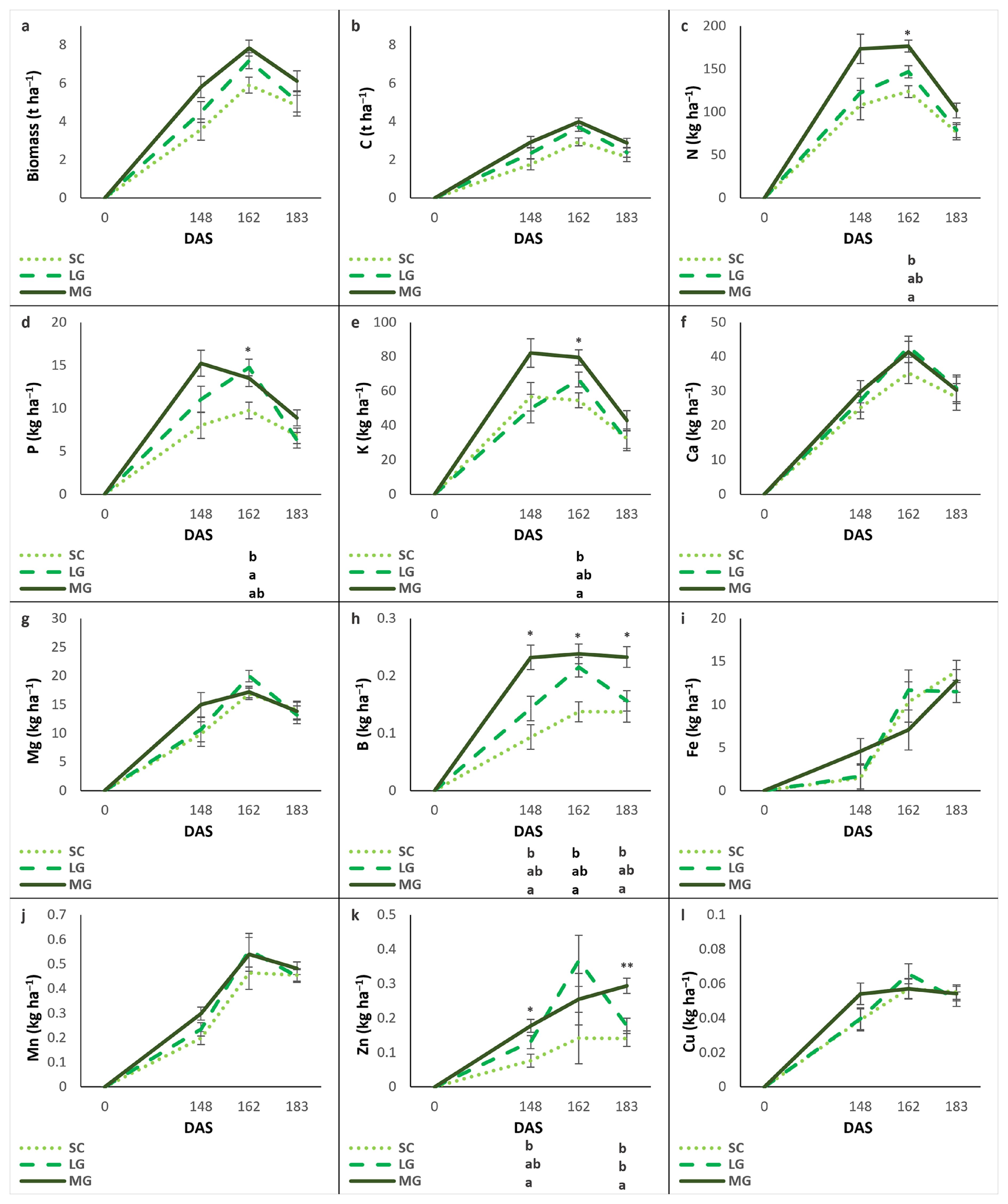
Figure 2. Evolution of biomass and nutrient accumulation over time (days after sowing, DAS) for each cover crop: subterranean clovers (SC), low-growing legumes (LG), and moderately-growing legumes (MG). Biomass (a), C (b), N (c), P (d), K (e), Ca (f), Mg (g), B (h), Fe (i), Mn (j), Zn (k), Cu (l). Values are means ± standard errors. Different letters show significant differences between cover crop mixtures in each DAR (*P<0.05; **P<0.01).
Biomass production increased from 148 to 162 DAS and declined to 183 DAS across all cover crops. The uptake of C, N, Ca, Mg, and Zn in plant tissues followed a similar pattern. For P, K, and B, an increase between 148 and 162 DAS was observed in SC and LG, while their levels remained stable in MG. After 162 DAS, a decrease was recorded in all cover crops. Fe content in plant tissues increased from 148 to 183 DAS, except in LG, where it remained relatively stable between 162 and 183 DAS. Mn increased between 148 and 162 DAS and plateaued at 183 DAS across all cover crops. Cu amount remained relatively stable throughout the study period.
Although no statistically significant differences were detected among cover crops for Biomass, MG tended to exhibit the highest biomass production, followed by LG and SC. A similar trend was observed for C, N, P, and B uptake. Statistical significance differences were found for N and K at 162 DAS, with MG having the highest values, SC the lowest, and LG intermediate values not significantly different from either; for P at 162 DAS, with LG having the highest values, SC the lowest, and MG intermediate values statistically equivalent to both; and for B at 148, 162 and 183 DAS, with MG consistently exhibiting the highest values, SC the lowest, and LG intermediate values statistically equivalent to both. Ca, Mg, Fe, Mn and Cu uptake were comparable across cover crops. For Zn, MG exhibited the highest uptake at 148 DAS, SC the lowest, and LG intermediate values statistically equivalent to the other two. At 162 DAS, Zn levels did not differ significantly between treatments, whereas at 183 DAS, MG had higher values than both SC and LG.
The highest biomass production was recorded at 162 DAS, with values of 5.9, 7.2, and 7.8 t ha−1 for SC, LG, and MG, respectively; despite observed differences, they were not statistically significant. Similarly, at the same DAS, N accumulation reached 123.7, 146.7, and 176.6 kg ha−1, P accumulation was 9.74, 14.8, and 13.5 kg ha−1, and K contents were 54.6, 66.7, and 79.5 kg ha−1 for SC, LG, and MG, respectively.
3.2 Cover crop residues decomposition
The decomposition of cover crop residues was assessed by analyzing the BR and the C/N ratio over time (DAR) for the three cover crops, as shown in Figure 3.
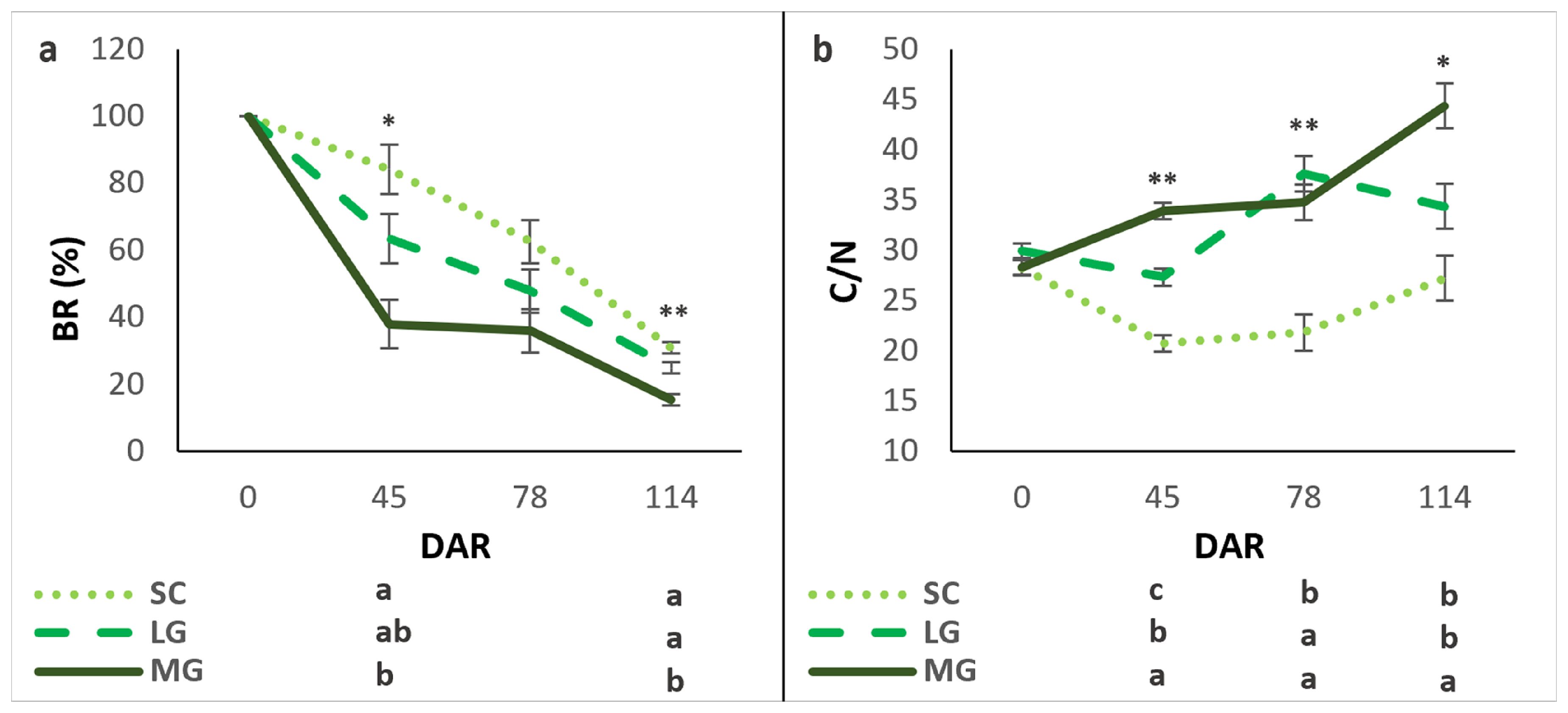
Figure 3. Biomass remaining (BR) and carbon/nitrogen (C/N) ratio over time (days after rolling, DAR) for each cover crop: subterranean clovers (SC), low-growing legumes (LG), and moderately-growing legumes (MG). BR (a), C/N (b). Values are means ± standard errors. Different letters show significant differences between cover crop mixtures in each DAR (**P<0.01, **P<0.01).
The BR values decreased consistently over time after rolling and varied significantly among the cover crops (Figure 3a). Overall, BR was highest in SC, followed by LG and MG, indicating that SC decomposed more slowly. In the first 45 days, SC retained 84% of its initial biomass, losing only 16%, while LG and MG lost 37% and 62%, respectively. By 78 DAR, no significant differences were observed in BR among the cover crops. At 114 DAR, MG exhibited the greatest decomposition, with 85% of its initial biomass lost, whereas SC and LG retained more biomass, with losses of 69% and 75%, respectively. At this stage, SC and LG were statistically similar. These results indicate that cover crops’ composition strongly affects decomposition rates, with SC showing the slowest decomposition over time.
When rolling down, the C/N ratio was statistically similar across all cover crops. However, significant differences emerged during decomposition (Figure 3b). By 45 DAR, the C/N ratio decreased in SC and LG, while it increased in MG. For LG, the ratio initially increased to 78 DAR but decreased by 114 DAR. For both SC and MG, C/N remains constant by 78 DAR, rising 25% and 27%, respectively, by 114 DAR. Over the following sampling dates, the C/N ratio increased in SC and MG. These trends highlight distinct patterns in residue quality and decomposition dynamics among the cover crops, with SC consistently exhibiting a lower C/N ratio than LG and MG after rolling down.
3.3 Cover crops’ nutrients release
The dynamics of nutrient release from cover crop residues were analyzed for C, N, P, K, Ca, Mg, B, Fe, Mn, Zn and Cu at various DAR (Figure 4). Overall, it was observed a decreasing trend in the percentage of NR in residue biomass over time, with the most pronounced release occurring within the first 45 days. SC consistently retained the highest proportion of nutrients in residues, while MG showed the lowest retention, likely reflecting its faster decomposition, and LG exhibited intermediate patterns.
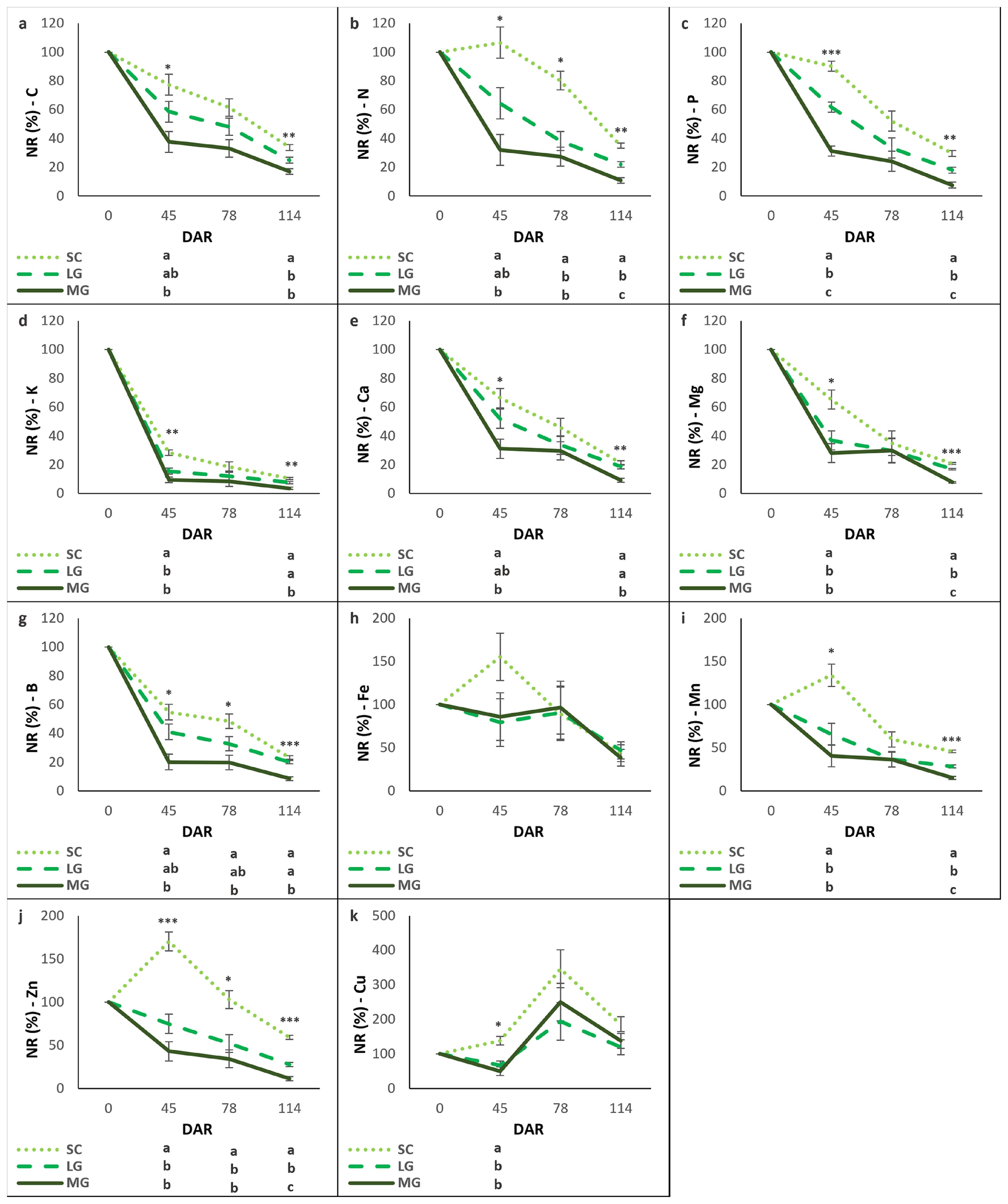
Figure 4. Nutrients remaining (NR) over time (days after rolling, DAR) for each cover crop: subterranean clovers (SC), low-growing legumes (LG), and moderately-growing legumes (MG). NR-C (a), NR-N (b), NR-P (c), NR-K (d), NR-Ca (e), NR-Mg (f), NR-B (g), NR-Fe (h), NR-Mn (i), NR-Zn (j), NR-Cu (k). Values are means ± standard errors. Different letters show significant differences between cover crop mixtures in each DAR (*P<0.05; **P<0.01; ***P<0.001).
The remaining C decreased consistently over time across all cover crops (Figure 4a). SC retained the highest levels of C, MG the lowest, and LG showed intermediate values, being LG statistically equivalent to both SC and MG at 45 DAS, and statistically equivalent to MG but significantly different from SC at 114 DAR.N generally decreased over time, except for SC, which showed an increase up to 45 DAR, before declining (Figure 4b). SC retained the highest levels of N, followed by LG, and MG had the lowest. P remaining declined steadily across all cover crops (Figure 4c). SC retained the most P, MG the least, and LG was intermediate, with no significant differences between cover crops at 78 DAR. K remaining exhibited a sharp decline by 45 DAR across all cover crops, followed by slower decreases until 114 DAR (Figure 4d). SC retained the highest levels of K throughout the study period. The remaining Ca decreased consistently over time across all cover crops (Figure 4e). SC retained higher levels of Ca, while MG retained the least. Differences between cover crops were not significant at 78 DAR. Mg remaining also declined across all cover crops (Figure 4f). SC retained the highest Mg levels, while LG and MG had similar, lower levels. Differences between cover crops were not significant at 78 DAR.
B remaining declined steadily over time in all cover crops, with SC retaining the highest levels and MG the least (Figure 4g). While Fe generally decreased over time, SC showed an unusual increase at 45 DAR before sharply declining by 78 DAR (Figure 4h). No significant differences were observed between cover crops at any DAR. Mn followed a trend similar to Fe, with SC showing an increase at 45 DAR before declining by 78 DAR (Figure 4i). SC retained the highest Mn levels at 45 and 114 DAR, while MG retained the least. Differences between cover crops were not significant at 78 DAR. Zn levels decreased over time in all cover crops except for SC, which showed an anomalous increase at 45 DAR before declining sharply at 78 DAR (Figure 4j). SC retained the most Zn, while LG and MG were significantly lower. Cu initially decreased in LG and MG but increased in SC at 45 DAR. By 78 DAR, Cu levels rose in all cover crops before declining again by 114 DAR (Figure 4k).
4 Discussion
4.1 Cover crop biomass production and nutrients provision capacity
In Mediterranean vineyards, cover crops must be terminated in late April or early May to reduce water competition with grapevines during critical stages. Rolling down the cover crops, as performed in this study, has been considered an effective termination method, by compressing the vascular system without severing the stems, rolling halts the flow of water and sugars (Silva and Delate, 2017; Kornecki and Kichler, 2023). Unlike cutting, which accelerates decomposition by exposing more surface area to microbial activity, rolling creates a uniform mulch layer that must help to conserve soil moisture and provide better erosion control, particularly on steep slopes (Silva and Delate, 2017; Iqbal et al., 2020; Kornecki and Kichler, 2023). To better simulate these field conditions, biomass samples were collected directly from surface residues, rather than using litter bags, which artificially accelerate decomposition by fragmenting plant material. Biomass production peaked at 162 DAS but declined by 183 DAS, suggesting that senescence and partial leaf fall had already begun before cover crop termination. This was likely triggered by the atypically dry spring conditions in April 2023. These results emphasize the need to align termination timing with seed maturity to support self-reseeding potential. In semi-arid systems, such as Mediterranean vineyards, early termination is particularly critical to minimize water competition with perennial crops (Rodrigues et al., 2015).
Cover crop biomass production in these regions varies significantly by species. For instance, red clover (Trifolium pratense L.) typically produces ~ 1.6 t ha–1, crimson clover (Trifolium incarnatum L.) 1.9 t ha–1, winter pea (Pisum sativum L.) 2.5 t ha–1, hairy vetch (Vicia villosa L.) 2 t ha–1, and cowpea (Vigna unguiculata L.) up to 6 t ha–1 (Ruis et al., 2019). The legume mixtures in this study yielded between 5.9 and 7.8 t ha–1, exceeding these values and others reported for similar Mediterranean climates. For example, Ordóñez-Fernández et al. (2018) documented biomass production between 1 and 3 t ha–1 for common vetch (Vicia sativa L.), bitter vetch (Vicia ervilia L.) and chickling vetch (Lathyrus sativus L.) in olive orchards in Córdoba, Spain, across different growing seasons. The high biomass observed in this study underscores the potential of these legume mixtures to enhance soil organic C, improve water retention and promote nutrient cycling in Mediterranean vineyards, offering a valuable tool for sustainable vineyard management. Furthermore, this high biomass can be linked to N availability, as a strong correlation was reported between cover crop biomass and N fertilization effect, with positive effects starting at 1 t ha−1, the minimum threshold for providing desired ecosystem services (Notaris et al., 2025).
As expected due to their vigorous growth habit, MG mixtures consistently produced the highest biomass, followed by LG and then SC, although differences were not statistically significant. Nutrient accumulation (N, P, K) followed similar patterns, peaking at 162 DAS, with MG generally exhibiting higher nutrient levels, indicating a superior capacity for nutrient provision. This aligns with findings from Finney et al. (2016) and Ruis et al. (2019), which demonstrated that species composition influences both biomass and nutrient contributions. The observed decline in nutrients between 162 and 183 DAS further underscores the importance of termination timing to optimize nutrient return to the soil.
Grapevine nutrient requirements vary with location, variety, rootstock type, grapevine age, and production levels (Verdenal et al., 2021; Magalhães, 2015), still average macronutrient exports are 40–70 kg ha–1 year–1 of N, 4–10 kg ha–1 year–1 of P, 40–70 kg ha–1 year–1 of K, 40–80 kg ha–1 year–1 of Ca, and 6–15 kg ha–1 year–1 of Mg (Magalhães, 2015). The cover crops in this study can supply nutrient amounts that approached or exceeded these values (e.g. 124–177 kg ha–1 of N, 10–15 kg ha–1 of P, 55–80 kg ha–1 of K, 35–43 kg ha–1 of Ca and 17–20 kg ha–1 of Mg) suggesting that they can significantly contribute to grapevine nutritional needs, even accounting for the additional 5–10% required for root and trunk growth (Magalhães, 2015). However, not all nutrients released by cover crops are immediately available or taken up efficiently by grapevines. Factors such as soil characteristics, precipitation patterns, crop vigor, and local productivity must be considered. Therefore, nutrient management strategies should be site-specific and guided by regular foliar and soil analyses (Magalhães, 2015). Additionally, caution is warranted when using legume cover crops converted to mulch, due to the reduced transfer of N from legumes to the perennial crop, as observed by Rodrigues et al. (2013) in olive orchards. Thus, it is crucial to consider the timing of nutrient release concerning grapevine demand. Furthermore, initial decomposition stages may involve nutrient immobilization as microorganisms consume available nutrients, potentially reducing their immediate availability to grapevines (Adhikari et al., 2024). This reinforces the importance of managing decomposition dynamics and understanding nutrient cycling when integrating cover crops into perennial systems.
4.2 Decomposition of cover crop residues
Cover crop residues decomposition is a key process in Mediterranean vineyards, influencing nutrient cycling and organic matter dynamics, and, ultimately, grapevine productivity. This study, in agreement with prior findings (Dorissant et al., 2022; Poudel et al., 2023; Adhikari et al., 2024; Curtright et al., 2025) demonstrate that both the DAR and the type of cover crop influenced BR and the C/N ratio, highlighting the dynamic nature of residue decomposition.
Environmental conditions and residue management practices play an important role in how biomass decomposes. Rapid initial decomposition can happen under various climate conditions when microbial activity is favored (Dorissant et al., 2022; Thapa et al., 2022a; Ntonta et al., 2024). Interestingly, Ntonta et al. (2024) reported a relatively rapid mass loss of sorghum residues in South Africa’s hot and arid climate, between 20–40% within the first 14 days, and 30–75% after 42 days, depending on the sorghum cultivar. In contrast, our study showed slower initial decomposition rates, with mass loss ranging from 16% to 62% over the first 45 days. One likely factor is the method of residue management, unlike studies using litter bags, which fragment residues and expose more surface area, we preserved the structural integrity of the rolled mulch layer, reducing early microbial access and mimicking real-world vineyard conditions more closely.
In this study, decomposition patterns varied among cover crop mixtures. MG residues exhibited faster initial decomposition (within 45 DAR) than LG and SC, along with a significant increase in the C/N ratio. Residue quality and weather conditions are known to highly drive decomposition, with quality often playing a stronger role (Thapa et al., 2022b). While low C/N ratios generally promote faster breakdown due to more accessible N and simpler organic compounds (Adhikari et al., 2024; Ntonta et al., 2024), all the studied mixtures started with comparable C/N values. This suggests that decomposition was driven by factors beyond C/N alone.
One key factor appears to be biomass composition. MG mixtures likely contained a higher proportion of easily decomposable tissues such as leaves, which mineralized quickly, leaving behind more recalcitrant material (e.g., stems). This shift explains the observed increase in C/N ratio over time, a trend also reported in mixtures dominated by easily decomposable species (Dorissant et al., 2022; Thapa et al., 2022b). Although lignin and cellulose content were not directly measured, the rising C/N ratio supports this hypothesis (Thapa et al., 2022a, 2022b). The stabilization of BR between 45 and 78 DAR may mark a transition from labile to more recalcitrant biomass fractions.
Phenological differences among mixtures may also have influenced decomposition. The SC mixture began shedding leaves before rolling (observed before 0 DAR), resulting in a residue layer dominated by lignified stems, which are slower to decompose. In contrast, MG and LG mixtures retained more leaf material, potentially accelerating early-stage breakdown. Such variations in tissue type could explain the faster initial decomposition of MG despite similar starting C/N ratios.
Lastly, environmental conditions likely modulated microbial activity and decomposition rates. While favorable conditions during the early phase may have enhanced microbial colonization of residues, the subsequent drop in precipitation and rise in temperature after 45 DAR may have limited microbial activity (Ordóñez-Fernández et al., 2018; Ntonta et al., 2024), particularly affecting the breakdown of lignin-rich tissues and slowing the breakdown of recalcitrant material. This further reinforces the idea that the decomposition trajectory is governed by both intrinsic residue traits and external environmental constraints.
4.3 Nutrient release from cover crop residues
The nutrient release patterns observed in this study offer valuable insights into the decomposition dynamics of cover crop residues in Mediterranean vineyards. The decreasing trend in NR over time reflects expected residue breakdown and nutrient mineralization processes (Dorissant et al., 2022; Poudel et al., 2023; Adhikari et al., 2024). The pronounced nutrient release within the first 45 days, highlights the early stages of decomposition as critical for nutrient cycling, in line with previous findings (Lacey et al., 2020; Dorissant et al., 2022; Poudel et al., 2023; Adhikari et al., 2024).
Among the cover crops studied, SC consistently retained the highest levels of macronutrients (C, N, P, K, Ca, Mg) throughout the decomposition, also likely due to their slower decomposition rate. This slow release can support soil fertility later in the grapevine growth cycle. In contrast, MG residues exhibited the lowest nutrient retention, consistent with their rapid decomposition, potentially early-season benefits but fewer long-term contributions. The LG residues exhibited an intermediate behavior, with a moderate decomposition rate and corresponding nutrient release.
Among the nutrients, N and K are critical for grapevine growth and productivity. N is essential during early spring for bud burst and leaf growth, as well as later during fruit fill, while K plays a vital role in carbohydrate transport, which is crucial for fruit development and yield (Santos, 1996; Magalhães, 2015). While MG and LG residues released N quickly, SC showed an initial N increase up to 45 DAR, likely due to microbial immobilization, where microbes temporarily sequester N from residues to decompose C-rich materials (Grzyb et al., 2020). K was rapidly released from all cover crop residues, nearly 100% by 45 DAR, ensuring availability during early vine development. Though required in smaller quantities, P is essential for various plant processes, as energy transfer and photosynthesis (Santos, 1996; Magalhães, 2015). P release from cover crop residues followed a steady decline across all types, with SC retaining the highest levels, potentially supporting a gradual supply. Ca is particularly important for leaf and branch growth, particularly between leaf emergence and fruit set (Santos, 1996; Magalhães, 2015), a period that coincides with the peak decomposition of cover crop residues and thus aligned with grapevine Ca demand. Mg, essential for photosynthesis (Santos, 1996; Magalhães, 2015), showed significant retention in SC residues, whereas MG and LG residues released Mg more rapidly.
Micronutrients, while needed in small amounts, are essential for grapevine production, especially for high-yielding crops (Santos, 1996; Magalhães, 2015). Micronutrient release (particularly Fe, Mn, Zn, and Cu) was more variable. The anomalous increases in Fe, Mn and Zn at 45 DAR, especially in SC residues, may be attributed to methodological or environmental factors (e.g., soil moisture or temperature variations), which affect both decomposition and micronutrient solubility. SC residues retained the highest levels of micronutrients, suggesting their potential to provide a sustained source of micronutrients over time. MG residues, with the lowest retention of most micronutrients, likely released these elements more rapidly. Among micronutrients, Fe and Zn are particularly important for grapevine growth, as they are key components of biomolecules and important agents in the enzymatic system (Magalhães, 2015). Although SC residues retained the most Fe and Zn overall, MG and LG residues released these nutrients more rapidly, potentially meeting vine demand during critical growth stages. In alkaline soils, where Fe and Zn deficiencies are often exacerbated (Santos, 1996), the impact of cover crop residues on soil pH should be monitored. Similarly, B, essential for fruit set and carbohydrate transport (Magalhães, 2015), was released more gradually from SC residues, while MG and LG residues provided more rapid contributions. However, excess B can cause phytotoxicity, emphasizing the need for careful monitoring (Santos, 1996).
Synchronizing nutrient release with grapevine demand is essential. For instance, for N, P, and K, absorption increases progressively until veraison, pausing briefly during bloom and fruit set when vegetative growth slows to prioritize reproductive development. A second absorption peak occurs after veraison during maturation due to nutrient redistribution within the plant. In contrast, Ca and Mg absorption increases steadily until veraison and then declines sharply during maturation (Magalhães, 2015). In the study year, phenological stages were slightly advanced due to dry and hot spring conditions, coinciding with the natural decomposition and termination of cover crop residues. Bloom occurred on average during the first week of May for Touriga Nacional in the Cima Corgo subregion, 10 days earlier than usual. Veraison was similarly advanced by 7 days, occurring in the first week of July (ADVID, 2023).
5 Conclusions
This study evaluated the decomposition dynamics of legume-based cover crops terminated in spring in a Mediterranean vineyard. Our findings demonstrate that cover crop composition significantly impacts biomass degradation and nutrient release.
Among the mixtures studied, MG produced the highest biomass and decomposed rapidly, likely supporting immediate nutrient availability, while SC decomposed more slowly, retaining nutrients longer and potentially sustaining soil fertility. LG exhibited intermediate behavior, balancing short- and long-term nutrient release. These distinct decomposition and nutrient release patterns provide a basis for selecting cover crops and for optimizing residue management strategies to better influence nutrient availability in vineyard systems.
While these insights are valuable, the short duration and single-year focus limit long-term conclusions. Future research should consider more frequent sampling, particularly during early decomposition, assess direct links between residue decomposition, vine nutrient uptake, and soil nutrient dynamics, and evaluate effects across multiple climates and seasons. Additionally, long-term studies examining soil water retention, microbial activity, C accumulation, and detailed residue chemistry (e.g., soluble sugars, starch, hemicellulose, cellulose, and lignin) will further improve understanding of decomposition processes and inform sustainable cover crop management.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
CB: Formal Analysis, Investigation, Visualization, Writing – original draft. SM: Investigation, Visualization, Writing – review & editing. FS: Investigation, Writing – review & editing. JR: Investigation, Writing – review & editing. MA: Investigation, Writing – review & editing. MR: Investigation, Writing – review & editing. CC: Conceptualization, Project administration, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This work was funded by the Project Vine&Wine Portugal - Driving Sustainable Growth Through Smart Innovation, Ref. n.° C644866286-00000011, co-financed by the NextGenerationEU, under Reg. (EU) 2021/241 - Recovery and Resilience Facility in Portugal (PRR - C5: Capitalization and Business Innovation). This work is also supported by National Funds by FCT –Portuguese Foundation for Science and Technology, under the projects UID/04033: Centro de Investigação e de Tecnologias Agro-Ambienteis e Biológicas and LA/P/0126/2020 (https://doi.org/10.54499/LA/P/0126/2020).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fagro.2025.1604142/full#supplementary-material
References
Abad J., Hermoso de Mendoza I., Marín D., Orcaray L., and Santesteban L. G. (2021a). Cover crops in viticulture. A systematic review (1): Implications on soil characteristics and biodiversity in vineyard. OENO One 55, 1. doi: 10.20870/oeno-one.2021.55.1.3599
Abad J., Hermoso de Mendoza I., Marín D., Orcaray L., and Santesteban L. G. (2021b). Cover crops in viticulture. A systematic review (2): Implications on vineyard agronomic performance. OENO One 55, 2. doi: 10.20870/oeno-one.2021.55.2.4481
Abad J., Marín D., Imbert B., Virto I., Garbisu C., and Santesteban L. G. (2023). Under-vine cover crops: Impact on physical and biological soil proprieties in an irrigated Mediterranean vineyard. Sci. Hortic. 311, 111797. doi: 10.1016/j.scienta.2022.111797
Adhikari A., Shrestha P., Ghimire R., Liu Z., Pollock D., Acharya P., et al. (2024). Cover crop residue quality regulates litter decomposition dynamics and soil carbon mineralization kinetics in semi-arid cropping systems. Appl. Soil Ecol. 193, 105160. doi: 10.1016/j.apsoil.2023.105160
ADVID (2023). Boletim Ano Vitícola 2023. Balanço Final do Ano Vitícola (Vila Real, Portugal: ADVID - Associação para o Desenvolvimento de Viticultura Duriense). Available online at: https://www.advid.pt/uploads/DOCUMENTOS/Subcategorias/ano-vitico/Boletim_Balanco_ano_viticola_2023.pdf (Accessed March 1, 2025).
Blanco-Canqui H., Shaver T. M., Lindquist J. L., Shapiro C. A., Elmore R. W., Francis C. A., et al. (2015). Cover crops and ecosystem services: insights from studies in temperate soils. Agron. J. 107, 2449–2474. doi: 10.2134/agronj15.0086
Bordoni M., Vercesi A., Maerker M., Ganimede C., Reguzzi M. C., Capelli E., et al. (2019). Effects of vineyard soil management on the characteristics of soils and roots in the lower Oltrepo Apennines (Lombardy, Italy). Sci. Total Environ. 693, 133390. doi: 10.1016/j.scitotenv.2019.07.196
Çerçioğlu M., Udawatta R. P., and Anderson S. H. (2025). Use of cover crops for sustainable management of soil condition and health: A review. Soil Secur. 18, 100177. doi: 10.1016/j.soisec.2025.100177
Curtright A. J., Zapata-Rojas D., Horwath W. R., and Zhu-Barker X. (2025). Quantifying nitrogen provisioning and release from cover crops in walnut orchards. Agric. Ecosyst. Environ. 383, 109529. doi: 10.1016/j.agee.2025.109529
Dorissant L., Brym Z. T., and Swartz S. (2022). Residue decomposition dynamics in mixed ratios of two warm-season cover crops. Agrosyst. Geosci. Environ. 5, e20311.
FAO, ITPS (2015). Status of the World’s Soil Resources (SWSR) – Main Report (Rome, Italy: Food and Agriculture Organization of the United Nations and Intergovernmental Technical Panel on Soils).
Ferreira C. S. S., Seifollahi-Aghmiuni S., Destouni G., Ghajarnia N., and Kalantari Z. (2022). Soil degradation in the European Mediterranean region: Processes, status and consequences. Sci. Total Environ. 805, 150106. doi: 10.1016/j.scitotenv.2021.150106
Finney D. M., White C. M., and Kaye J. P. (2016). Biomass production and carbon/nitrogen ratio influence ecosystem services from cover crop mixtures. Agron. J. 108, 39–52. doi: 10.2134/agronj15.0182
Grzyb A., Wolna-Maruwka A., and Niewiadomska A. (2020). Environmental factors affecting the mineralization of crop residues. Agronomy 10, 1951. doi: 10.3390/agronomy10121951
IPCC (2023). Climate change 2023: synthesis report. contribution of working groups I, II and III to the sixth assessment report of the intergovernmental panel on climate change (Geneva, Switzerland: IPCC), 1–34.
IPMA (2025). Instituto Português do Mar e Atmosfer. Available online at: https://www.ipma.pt/en/oclima/normais.clima/?print=true (Accessed March 10, 2025).
Iqbal R., Raza M. A. S., Valipour M., Saleem M. F., Zaheer M. S., Ahmad S., et al. (2020). Potential agricultural and environmental benefits of mulches—a review. Bull. Natl. Res. Cent. 44, 75. doi: 10.1186/s42269-020-00290-3
IVV (2025). Instituto da Vinha e do Vinho. Available online at: https://www.ivv.gov.pt/np4/home.html (Accessed July 10, 2025).
Kornecki T. S. and Kichler C. M. (2023). Recurring rolling/crimping effects on termination effectiveness of iron clay pea and pearl millet warm-season cover crops. Agriculture 13, 1949. doi: 10.3390/agriculture13101949
Lacey C., Nevins C., Camberato J., Kladivko E., Sadeghpour A., and Armstrong S. (2020). Carbon and nitrogen release from cover crop residues and implications for cropping systems management. J. Soil Water Conserv. 75, 505–514. doi: 10.2489/jswc.2020.00102
Magalhães (2015). Tratado de Viticultura, A Videira a Vinha e o Terroir. 2nd Edition (Lisboa: Esfera Poética), 608 p, ISBN: ISBN: 9789899820739.
Marques M., Ruiz-Colmenero M., Bienes R., García-Díaz A., and Sastre B. (2020). Effects of a permanent soil cover on water dynamics and wine characteristics in a steep vineyard in the central Spain. Air Soil Water Res. 13, 1–10. doi: 10.1177/117862212094806
Mauromicale G., Occhipinti A., and Mauro R. P. (2010). Selection of shade-adapted subterranean clover species for cover cropping in orchards. Agron. Sustain. Dev. 30, 473–480. doi: 10.1051/agro/2009035
Medrano H., Tomás M., Martorell S., Escalona J. M., Pou A., Fuentes S., et al. (2015). Improving water use efficiency of vineyards in semi-arid regions. A review. Agron. Sustain. Dev. 35, 499–517. doi: 10.1007/s13593-014-0280-z
Notaris C., Peixoto L., Mortensen E., and Rasmussen J. (2025). Cover crop biomass production as a predictor of nitrogen fertilizer replacement value - legumes secure positive effects. Agric. Ecosyst. Environ. 381, 109446. doi: 10.1016/j.agee.2024.109446
Ntonta S., Zengeni R., Muchaonyerwa P., and Chaplot V. (2024). Variability in decomposition rate of sorghum cultivar residues linked to lignin content. Rhizosphere 29, 100850. doi: 10.1016/j.rhisph.2024.100850
Opoku A., Ogunleye A., Solomon J., and Payne W. (2024). Cover crop systems impact on biomass production, carbon-to-nitrogen ratio, forage quality, and soil health in a semi-arid environment. Heliyon 10, e39600. doi: 10.1016/j.heliyon.2024.e39600
Ordóñez-Fernández R., Repullo-Ruibérriz de Torres M. A., Márquez-García J., Moreno-García M., and Carbonell-Bojollo R. M. (2018). Legumes used as cover crops to reduce fertilisation problems improving soil nitrate in an organic orchard. Eur. J. Agron. 95, 1–13. doi: 10.1016/j.eja.2018.02.001
Poudel P., Parajuli B., Park D., and Ye R. (2023). Cover crop residues decomposition and nutrient releases in a sandy ultisols of U.S. coastal plain: impacts of termination timing. Commun. in Soil Sci. Plant Anal. 54, 2394–2411. doi: 10.1080/00103624.2023.2223612
Rodrigues M. A., Correia C. M., Claro A. M., Ferreira I. C. F. R., Barbosa J. C., Moutinho-Pereira J. M., et al. (2013). Soil nitrogen availability in olive orchards after mulching legume cover crop residues. Sci. Hortic. 158, 45–51. doi: 10.1016/j.scienta.2013.04.035
Rodrigues M., Ferreira I. Q., Freitas S. L., Pires J. M., and Arrobas M. P. (2015). Self-reseeding annual legumes for cover cropping in rainfed managed olive orchards. Span. J. Agric. Res. 13, e0302. doi: 10.5424/sjar/2015132-6252
Ruis S. J., Blanco-Canqui H., Creech C. F., Koehler-Cole K., Elmore R. W., and Francis. C. A. (2019). Cover crop biomass production in temperate agroecozones. Agron. J. 111, 1535–1551. doi: 10.2134/agronj2018.08.0535
Santos Q. J. (1996). Fertilização – Fundamentos da utilização dos adubos e corretivos. 2nd Edition (Mem Martins, Portugal: Publicações Europa -América), ISBN: ISBN: 972-1-00949-0.
Silva E. M. and Delate K. (2017). A decade of progress in organic cover crop-based reduced tillage practices in the upper midwestern USA. Agriculture 7, 44. doi: 10.3390/agriculture7050044
Temminghoff E. E. and Houba V. J. (2004). Plant Analysis Procedures (London, UK: Kluwer Academic Publishers).
Thapa R., Tully K., Hamovit N., Yarwood S., Schomberg H., Cabrera M., et al. (2022b). Microbial processes and community structure as influenced by cover crop residue type and placement during repeated dry-wet cycles. Appl. Soil Ecol. 172, 104349. doi: 10.1016/j.apsoil.2021.104349
Thapa R., Tully K., Reberg-Horton C., Cabrera M., Davis B. W., Fleisher D., et al. (2022a). Cover crop residue decomposition in no-till cropping systems: Insights from multi-state on-farm litter bag studies. Agric. Ecosyst. Environ. 326, 7823. doi: 10.1016/j.agee.2021.107823
Verdenal T., Dienes-Nagy Á., Spangenberg J. E., Zufferey V., Spring J.-L., Viret O., et al. (2021). Understanding and managing nitrogen nutrition in grapevine: a review. OENO One 55, 1–43. doi: 10.20870/oeno-one.2021.55.1.3866
Keywords: agroecosystem sustainability, cover crops’ elemental composition, cover crop residues, residues decomposition, grapevine
Citation: Brito C, Martins S, Saraiva F, Roque J, Arrobas M, Rodrigues M and Correia C (2025) Biomass and nutrient release dynamics of decomposing rolling down legume cover crops in a steep-sloped Mediterranean vineyard. Front. Agron. 7:1604142. doi: 10.3389/fagro.2025.1604142
Received: 01 April 2025; Accepted: 22 July 2025;
Published: 08 August 2025.
Edited by:
Desouza Blaise, Central Institute for Cotton Research (ICAR), IndiaReviewed by:
Amit Anil Shahane, Central Agricultural University, IndiaYesica Stefania Bernaschina, National Institute for Agricultural Research (INIA), Uruguay
Copyright © 2025 Brito, Martins, Saraiva, Roque, Arrobas, Rodrigues and Correia. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Cátia Brito, Y3ZxYnJpdG9AdXRhZC5wdA==