 Nadia Monjezi
Nadia Monjezi Hamid Reza Eisvand
Hamid Reza Eisvand Robert Lee2
Robert Lee2 Meir Levi
Meir Levi Donald L. Smith
Donald L. Smith- 1Plant Science Department, McGill University, Montreal, QC, Canada
- 2SeaPurAgro Inc., Montreal, QC, Canada
- 3Consultant, Montreal, QC, Canada
This study explores the potential of Cell-Free Supernatants (CFSs) derived from beneficial bacteria as a sustainable solution to enhance crop resilience in the face of environmental stress. In the context of climate change and soil salinity, CFSs emerge as a promising tool to mitigate crop losses and safeguard food security. By employing bioactive compounds extracted from microbial cultures, CFSs offer a reliable approach to support plant growth and fight abiotic stressors. The research emphasizes the effectiveness of CFSs in promoting seed germination and improving overall plant health, particularly under salinity stress. Additionally, it highlights the role of CFSs in enhancing nutrient absorption and improving plant defense mechanisms, contributing to agricultural sustainability. Despite technical limitations associated with microbial formulations, CFSs provide an alternative to conventional methods, presenting scalable and eco-friendly solutions. Among various production methods of the CFS, centrifugation only and centrifugation plus 0.22 µm filtration stand out due to their simplicity, and efficiency. However, the absence of field-level studies reveals a critical research gap, necessitating further evaluation of CFS performance under real agricultural conditions. Through collaborative research works and innovative application methods, CFSs hold the potential to transform modern agriculture, ensuring resilient crop production systems and global food security for generations to come.
1 Introduction
Numerous agricultural systems have witnessed heightened crop productivity through the use of synthetic fertilizers, which effectively lowers costs and maximizes yields. However, environmentalists’ express concerns that without action to mitigate fertilizer use, we risk creating an unsustainable situation in the long term, leading to an increased agricultural carbon footprint and elevated environmental compliance costs. These concerns have prompted a global shift towards more sustainable crop production approaches (Dhankher and Foyer, 2018; Pareek et al., 2020). In recent decades, bioactive natural products have been viewed as a positive approach, with the utilization of these eco-friendly compounds/materials in agriculture showing promise due to their low cost and potential for enhancing crop yield and quality. Biofertilizers, composed of active microbes, have emerged as cost-effective and eco-friendly alternatives to chemical-based fertilizers, being widely distributed in soil and capable of inducing specific desirable plant functions (Hug et al., 2020; Naamala and Smith, 2020).
Beneficial microbes, particularly Plant Growth-Promoting Rhizobacteria (PGPR), play essential roles in supporting plant growth, enhancing nutrient uptake, regulating phytohormone levels, and helping plants tolerate abiotic environmental stresses such as salinity, drought, and temperature fluctuations (Rai et al., 2020; Chauhan et al., 2021; Riaz et al., 2021; Khoshru et al., 2020; Parameswaran et al., 2021). However, the performance of live microbial biofertilizers in the field is often inconsistent due to variable environmental conditions (Rilling et al., 2019) and competition from other microbes already at the site of application.
Beneficial microbes, particularly Plant Growth-Promoting Rhizobacteria (PGPR) constitute sustainable crop production inputs able, among other things to help mitigate the effects of climate change related stresses and to enhance uptake of atmospheric CO2 by plants (Khoshru et al., 2020; Rai et al., 2020; Chauhan et al., 2021; Parameswaran et al., 2021; Riaz et al., 2021). However, there is a need to make their effects as consistent and as large as possible (Rilling et al., 2019). Microbial biostimulants, especially PGPR cell-free supernatants (CFSs) and isolated metabolites from the CFSs, have recently gained attention as tools to help plants withstand abiotic and biotic stresses. Specifically, these substances offer novel opportunities to promote tolerance to adverse environmental conditions during critical crop growth stages (Pellegrini et al., 2020; Li et al., 2021).
Soil salinization is a prominent abiotic stress that hampers plant functions and disrupts crop growth and yield on a global scale. Salinity reduces the plant’s water absorption capacity, creating drought-like conditions and impeding growth through osmotic stress. In addition, saline stress often leads to ion toxicity due to imbalances in cytosolic nutrients, mainly excess sodium (Na+) and chloride (Cl−) accumulation. Another consequence is the heightened synthesis and accumulation of reactive oxygen species (ROS) under salinity stress, causing oxidative stress, cellular damage, and ultimately cell death (Assaha et al., 2017; Isayenkov and Maathuis, 2019). Despite these challenges, plants have evolved an efficient network of ROS scavenging/detoxifying systems. This detoxification process involves nonenzymatic or enzymatic antioxidants (Pang and Wang, 2008). The ability of beneficial plant-associated bacteria from saline and arid environments to develop unique evolutionary adaptations with plants for coping with challenging conditions has gained significant attention (Etesami and Adl, 2020; Leontidou et al., 2020). For instance, Plant Growth-Promoting Rhizobacteria (PGPR) play crucial roles in scavenging ROS, enhancing enzymatic and nonenzymatic antioxidant activities during stress, and reducing ROS levels under abiotic stress conditions (Bharti and Barnawal, 2019; Kumar et al., 2020). Moreover, bioactive compounds produced by these beneficial microbes can stimulate crop production without the limitations faced by live microbial biofertilizers. These microbial-derived compounds are a diverse group of natural substances that contribute to the agricultural sector by enhancing resistance to biotic/abiotic stresses and acting as inter-organismal signals to improve symbioses with beneficial organisms (Naamala and Smith, 2021). Some studies highlight the positive effects of PGPR CFSs and isolated metabolites as biostimulants for enhancing plant health and growth under salinity stress (Naamala et al., 2022; Shah et al., 2022a). However, despite this evidence, considerably less attention has been directed towards utilizing PGPR-CFSs to develop eco-friendly bio-formulations for enhancing crop production under field conditions. These bioagents are designed to improve crop yield in actual fields. Despite the potential of PGPR-CFSs, using them in real-world field conditions hasn’t been explored much. This calls for more in-depth research. This effort could uncover the full potential of these biostimulants and help advance sustainable farming by connecting lab discoveries with practical use.
1.1 Definition and characteristics of CFS and microbial metabolites
Plant Growth-Promoting Rhizobacteria (PGPR) CFSs refer to the extracellular liquids obtained after removing bacterial cells from a liquid culture—typically through centrifugation and filtration. These supernatants contain an array of microbial-derived compounds such as enzymes, phytohormones (like indole-3-acetic acid), volatile organic compounds, siderophores, and antibiotics, among others (Pellegrini et al., 2020; Naamala and Smith, 2021). CFSs can be derived from various PGPR genera, including Bacillus, Pseudomonas, Devosia, and Rhizobium, known for their efficacy in promoting plant growth and resilience.
The metabolites within these CFSs can be harvested during stationary or exponential growth phases, depending on the desired activity, and are usually concentrated or freeze-dried for agricultural applications. These bioactive metabolites are typically applied as seed treatments, foliar sprays, or soil amendments. The effective concentration varies depending on the crop species and environmental conditions, but studies often use dilutions ranging from 10% to 50% (v/v) or apply dry weights in the range of 50–200 mg L-1 (Naamala et al., 2022; Shah et al., 2022a; Monjezi et al., 2023).
These microbial products have been tested on various crops, including soybean, maize, canola, cassava, and pigeon pea, under different stress conditions like salinity, drought, and temperature extremes. The positive effects include enhanced seed germination, improved antioxidant enzyme activity, greater nutrient uptake, and increased biomass and yield (Buensanteai et al., 2013; Tewari et al., 2020; Monjezi et al., 2023). An overview of the bacterial strains' CFSs, their key metabolites, application methods, and corresponding crop responses is presented in Table 1.
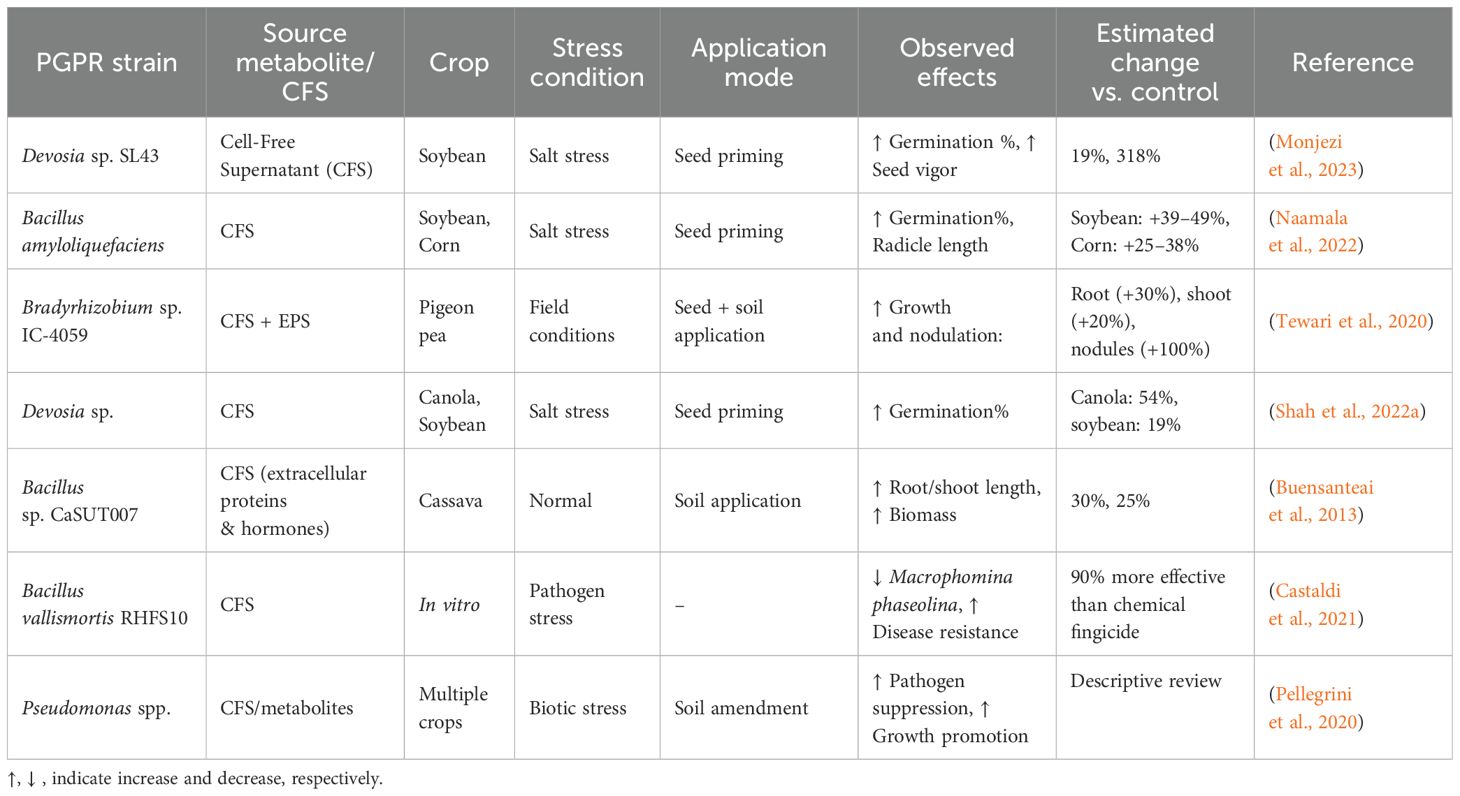
Table 1. Summarizing the bacterial strains CFSs, their derived metabolites, modes of application, and crop responses.
2 Intersecting pressures: climate change, agricultural stress, and population growth
Clear evidence shows that climate change is happening, bringing various effects such as higher temperatures, drought and rising sea levels. The levels of these changes to date indicate a faster warming of the Earth than initially thought. It’s well-established that human activities are making this situation worse by releasing more greenhouse gases (GHGs), strengthening the greenhouse effect in the earth’s atmosphere. This disturbance in the natural balance causes the atmosphere to trap more heat, leading to significant changes in the global climate system (Ahmad et al., 2010; Corwin, 2021).
The effects of climate change, especially on crops, have become more significant in recent times. Abiotic stresses, such as water shortages, temperature extremes, waterlogging, and soil salinity, pose challenges for modern crop production. These issues disrupt the growth of crops, affecting their reproductive phases and ultimately reducing the harvest index (the proportion of grain yield to total above-ground biomass) and overall crop yield. The damage depends on factors like the crop’s development stage, when the stress occurs, and how severe it is (Onu et al., 2019; Raza et al., 2019). In real-world agriculture, multiple stressors come together, making the effects more complicated and harmful than isolated stresses. This complex stress situation highlights the challenges crops face under field conditions (Pandey et al., 2015).
At the same time, the growing human population is putting more pressure on arable land. This population increase adds to the challenges for agriculture, involving both living (biotic) and non-living (abiotic) stresses. For instance, farmers often try to boost productivity by using more chemical fertilizers, pesticides, irrigation, and machinery. Ironically, this intensified farming may increase vulnerability to various stress factors. The expected increase in disturbances caused by climate change will worsen both biotic and abiotic stresses, making it more difficult for agriculture and weakening industrialized farming systems (Xie et al., 2019). Considering these factors, there’s an urgent need to rethink agricultural strategies. A more comprehensive and integrated approach is crucial, taking into account the combined challenges of climate change and population growth.
The strategic use of natural agricultural biostimulants presents a promising solution for achieving multiple agricultural and environmental goals. By improving soil health, reducing greenhouse gas emissions, and enhancing nutrient-use efficiency, biostimulants can play a crucial role in the transition towards more sustainable and resilient farming systems. These agricultural biostimulants, such as plant growth-promoting rhizobacteria (PGPR), CFSs, seaweed extracts, and bioflavonoids, have become more popular as an eco-friendly method, which can help achieve the goals of reducing greenhouse gas (GHG) emissions and lowering the carbon footprint. Additionally, they improve the efficiency of nutrient use in plants. Combining and applying these natural biostimulants can enhance soil quality and further reduce GHG emissions. This approach not only supports sustainable farming but also promotes healthier crop growth. By improving soil structure and increasing microbial activity, biostimulants can enhance nutrient availability, leading to better root development and increased plant resilience against diseases and environmental stresses. Moreover, the use of these biostimulants can reduce the need for synthetic fertilizers and pesticides, which are often associated with negative environmental impacts. Natural biostimulants can also be considered a great boost for restoring degraded soils, making them more fertile and productive over time, and leading to higher crop yields and better food security, particularly in regions where soil degradation is a significant issue. In addition to their environmental benefits, natural biostimulants can also contribute to the economic sustainability of farming. By reducing the dependence on chemical inputs, farmers can lower their production costs and increase their profitability. Furthermore, the use of biostimulants can open up new markets for organic and sustainably produced crops, meeting the growing consumer demand for environmentally-friendly products (Pandey et al., 2015; Pellegrini et al., 2020; Naamala et al., 2022; Shah et al., 2022a, 2022b; Zhao et al., 2023).
2.1 Understanding how plants deal with stress: impact on global agriculture
Plants, being stationary, can’t move away from harsh conditions caused by either abiotic or biotic factors. Abiotic factors like extreme temperatures, drought, floods, heavy metals, UV radiation, and salinity stress weaken plants, making them more prone to diseases and pests. This vulnerability leads to significant drops in crop yields worldwide. Salinity, especially, is a major stressor limiting global agricultural productivity (Sahab et al., 2021). Salinity is a primary abiotic stress affecting agriculture, significantly hindering crop growth globally. Around 830 million ha of agricultural land worldwide are affected by salinity, especially in arid and semi-arid regions with limited rainfall and high evaporation rates (Sahab et al., 2021; Wang et al., 2021). In these salty soils, high salt levels cause a lack of water and essential nutrients for plants. This can happen naturally or be worsened by human activities like improper irrigation and excessive chemical fertilizer use (Raza et al., 2019; Çatav et al., 2021; Wang et al., 2021). High salt levels also negatively affect the soil ecosystem, impacting processes in plants, microorganisms, and other underground organisms (Sahab et al., 2021). Concerningly, soil salinization is increasing more than researchers predicted, indicating a need for caution as salinity-related challenges might spread to new regions (Macdonald et al., 2021). In recent years, efforts have increased to find ways to manage soil salinity. Most focus on improving soil properties and organic matter, with some looking into plant-level solutions. Combining different techniques for salt management is suggested for better results (Sahab et al., 2021). Considering the interconnected nature of how plants respond to various stresses, including salinity, has proven valuable for studying plant stress defense mechanisms. The advantage lies in consistently applying and controlling salt stress levels in various controlled environments, such as laboratories, growth chambers, greenhouses, and similar facilities (Shavrukov, 2013).
3 Plant responses to saline soil: exploring mechanisms and strategies
Soil salinization harms plants, disrupting their processes. How plants respond to salinity stress varies based on factors such as species, genotype, growth stage, and environmental conditions. Seedling emergence, a crucial phase, is vulnerable to salinity stress across crops (Naamala et al., 2022; Yaghoubian et al., 2022). While growth stage is a key determinant of stress sensitivity, the other factors—plant species, genotype, and environmental conditions—also play significant roles in shaping plant responses to salinity. Different species and genotypes exhibit variable salinity tolerance due to inherent physiological and biochemical traits. For instance, halophytes like Suaeda fruticosa exhibit superior ion compartmentalization and osmotic adjustment compared to glycophytes such as soybean (Shah et al., 2022b). Environmental factors, such as light intensity and temperature, modulate plant metabolism and antioxidant activity under salt stress, thereby influencing the degree of damage or tolerance. In this context, understanding species-specific and genotype-dependent mechanisms, alongside environment-driven modulation, is critical for targeted stress management strategies.
Moreover, although this manuscript emphasizes salinity stress as a case study, the effects of PGPR and their CFSs extend to other abiotic stressors, such as drought, extreme temperature, and heavy metal toxicity. PGPR-derived compounds help maintain water homeostasis during drought by increasing root growth and accumulation of osmoprotectants (Ayuso-Calles et al., 2021). Under cold stress, compounds like lipo-chitooligosaccharides (LCOs) enhance seed germination and early vigor, especially in canola (Schwinghamer et al., 2015). In heat stress conditions, thuricin 17 has been shown to boost biomass and root growth (Lyu et al., 2020). In the context of biotic stress, PGPR-CFSs also act as biocontrol agents, triggering systemic resistance and suppressing pathogens such as Macrophomina phaseolina (Castaldi et al., 2021). A more holistic exploration of these applications is needed, and future studies should expand beyond salinity to harness PGPR potential across diverse environmental challenges.
Challenges in seed germination due to heightened salinity are multi-dimensional. Salinity interferes with water absorption by seeds due to elevated osmotic potential, hindering germination. Harmful sodium and chloride ions in saline conditions add another layer of hindrance. These factors reduce seed germination rates under high salinity stress (de Leija et al., 2022; Huang et al., 2022; Shah et al., 2022a), as studied by Safdar et al. (2019). As plants grow beyond the vulnerable seedling stage, they develop greater resilience to salinity stress (Shah et al., 2022b). However, the speed of seed germination and seedling establishment is crucial for determining crop yield, especially with stress factors like salinity. Signs of salt stress might not be apparent later on, yet they can result in lower crop yields. This is because the adverse effects of salinity can impact the plant’s growth from an early stage, causing reduced final yields (Sabagh et al., 2021). Moreover, seeds facing challenges due to salinity may allow weeds to grow alongside or even faster than the main crop, resulting in a significant decrease in the final harvest (Arce et al., 2009). Extended exposure to salt stress significantly impacts various physiological and biochemical aspects of plants, reducing essential characteristics like leaf area, leaf water content, and photosynthetic pigments. This hampers overall photosynthetic capacity, causing a decrease in yield. These outcomes result from a complex interplay of factors, including nutritional imbalances, osmotic stress, ion effects, oxidative stress, and impaired water uptake from the soil. The cell wall becomes a primary site of salt stress effects, leading to significant changes in its physical properties (Pang and Wang, 2008; Raza et al., 2019; Çatav et al., 2021; Corwin, 2021).
The decline in photosynthesis, a crucial process in plant metabolism, in response to salt stress, is accompanied by an accumulation of reactive oxygen species (ROS). The oxidative stress-induced elevation of ROS harms plant physiology, causing the breakdown of chloroplast structure and a decrease in photosynthetic pigments (Zhao et al., 2021). In response to salinity stress, plants undergo metabolic reprogramming to maintain cellular equilibrium, involving changes in the production of primary and secondary metabolites, including proteins, responding to varying degrees of salt stress (Athar et al., 2022). The physiological shifts from salt stress exposure have far-reaching implications for plant morphological traits, affecting features like leaf area. These effects lead to a reduction in both crop yield quantity and quality. This becomes evident in the context of total leaf area reduction, a consequence of changes in cell wall regulations and integrity, leaf turgor, and photosynthetic rates. These changes negatively impact other morphological traits, including plant height, stem diameter, and root growth. Various investigations have shown the adverse negative effect of soil salinity on agronomic traits in soybean, encompassing height, stem diameter, leaf size and shape, biomass accumulation, pod count, and seed yield (Phang et al., 2008; Yu et al., 2019; Otie et al., 2021; Hasanuzzaman et al., 2022).
4 Harnessing biostimulants for enhanced plant growth and sustainable agriculture
Over the past few decades, cultivating agriculture using environmentally sustainable farming systems has become a significant challenge in the agricultural sector. Recently, there has been considerable focus on various biological substances and microorganisms known as plant biostimulants. These agents are used in agriculture to improve plant health, growth, and nutritional vigor. Plant biostimulants include beneficial fungi and bacteria, algal extracts, inorganic compounds, protein hydrolysates, humic substances, fulvic acid, and chitosan. The global biostimulant market has been steadily growing due to the favorable and environmentally friendly nature of these products, in contrast to synthetic agrochemicals, as documented by Dong et al. (2020); Sangiorgio et al. (2020), and D’Addabbo et al. (2019). In natural ecosystems, the well-being of most cultivated and wild plants is closely connected to the microbial organisms in the soil. These microorganisms have the potential to support plant growth and development by influencing hormone production, enzyme activities, signaling pathways, and various other mechanisms. Together, these factors help plants deal with both living and non-living environmental challenges. By promoting stress tolerance and resilience in plants, these mechanisms support plant growth. Additionally, the microorganisms associated with plants also play a role in making essential nutrients available through the release of complex substances, as explained by Sahab et al. (2021). Therefore, the need to create biostimulants based on microbes to lessen the negative effects of environmental stresses and ensure global food security is a significant concern. In this context, a top priority involves understanding the detailed interactions between plant roots and soil microorganisms. Elucidating and improving these interactions are crucial, not just for making agricultural systems work better, but also for keeping the soil healthy. Sangiorgio et al. (2020) and Etesami and Adl (2020) have provided valuable insights into these processes. Despite the promising use of microbial biostimulants as a smart alternative to regular chemicals in farming, there are some limitations that need careful consideration. These include issues like lower effectiveness compared to chemical alternatives and being more sensitive to environmental factors. The fact that outcomes can vary considerably among crops and locations emphasizes the need for more research. Therefore, making microbial biostimulants work better and having consistent results require collaborative scientific efforts, as highlighted by Lyu et al. (2021) and Sangiorgio et al. (2020).
5 Utilization of plant growth-promoting bacteria as biotechnological tools for alleviating abiotic stress in plants
The use of plant growth-promoting bacteria (PGPB) involves various types of bacteria that can independently team up with plants in specific ways. Those found around the roots are called plant growth-promoting rhizobacteria (PGPR) (Sansinenea, 2019). These bacteria are good at living on and in plant roots and in the nearby soil, helping plants grow better. The ways they help can vary depending on the specific plant they’re working with. Importantly, the substances produced by these helpful bacteria not only boost plant growth by increasing nutrient availability or changing plant hormone levels but also have the potential to influence the entire plant’s genetic activity (Rai et al., 2020). Traditional agriculture has often relied on using lots of chemicals to make crops grow more, but this can cause considerable environmental harm, as indicated by Ramakrishna et al. (2019).
Alternatively, using naturally occurring plant growth-promoting bacteria (PGPR) is an eco-friendlier approach. It not only helps plants grow better but also improves the health of the soil. This happens because these bacteria produce different substances that make important changes in the soil around the plant, like its pH, structure, and nutrient levels. So, this method has the potential to make agriculture more sustainable in the long run (Hu et al., 2018; Ramakrishna et al., 2019; Naamala and Smith, 2020). In addition, the microbiome, acting like the ‘second genome’ of plants, has a big effect on how plant genes work. It does this by releasing signal molecules that affect the host plant, making a significant impact on the plant’s health and overall condition, including responses to stress factors. So, understanding and controlling the activities of the microbiota, especially the ones that help cultivated crops, can be very useful (Turner et al., 2013; Arif et al., 2020).
It’s well-established that microbes closely linked with plant roots play a key role in boosting plant resilience to environmental stress. This helps in enhancing growth, yield, and nutrient absorption, especially when conditions are tough. This idea is supported by the research of Leontidou et al. (2020); Etesami and Adl (2020); Singh et al. (2019), and Mhatre et al. (2019). The microbial community around plant roots is known for its various contributions in diminishing the negative effects of salinity-induced stress. By regulating and adapting plant growth and development, these microbes help plants survive and thrive. So, using halo-tolerant PGPR on the intended plant species is a practical way to lessen the negative impacts of salinity-related challenges on plant growth, as explained by Leontidou et al. (2020). In simple terms, microbes that help plants grow in salty soils have various ways of doing it. They can directly improve plant growth under saline conditions, possibly by making growth-promoting hormones like auxins, cytokinins, and gibberellins. They might also reduce the levels of ethylene, a substance that hinders plant growth. PGPR’s role in managing hormone levels, particularly reducing ethylene and its negative impact on plant growth in salty conditions, is a crucial part of how they work. At the same time, some helpful microbes boost bacterial auxin production or increase the plant’s own auxin levels. This helps the main root grow longer and creates more lateral roots. Balancing ethylene and auxin in this way helps plants efficiently absorb water, ions, and nutrients when dealing with saline conditions (Spaepen and Vanderleyden, 2011; Zerrouk et al., 2016; Iqbal et al., 2017; Kumar et al., 2020; Eichmann et al., 2021; Park et al., 2021).
In saline soil, plants encounter a primary challenge known as osmotic stress, which disrupts water balance and stomatal gas exchange, consequently impeding the rate of photosynthesis. Addressing this issue strategically involves facilitating osmotic adjustment through the accumulation of water-soluble molecules. Halo-tolerant Plant Growth-Promoting Rhizobacteria (PGPRs) have the capacity to contribute to this mechanism by synthesizing compatible solutes or osmoprotectants, such as proline and glycine betaine. The synthesis and accumulation of these stress protectants enable plants to navigate the deleterious impacts of salinity-induced stress on growth, as corroborated by Ahmed et al. (2021); Ayuso-Calles et al. (2021); Nawaz et al. (2020), and Kumar et al. (2020). Furthermore, the adverse effects of soil salinity extend to the overproduction of reactive oxygen species (ROS), resulting in oxidative damage to vital biomolecules and even to cellular demise. Nonetheless, selectively chosen microbes exhibit the capacity to stimulate various antioxidant defense enzymes, including peroxidases, catalases, superoxide dismutases, glutathione reductases, and glutathione S-transferases. This orchestrated enhancement of antioxidant defense systems effectively counters the toxic effects of ROS in stressful environments, as underscored by Leontidou et al. (2020); Otlewska et al. (2020), and Gapińska et al. (2008).
5.1 Technical preparation and application methods for PGPR and their cell-free supernatants
Plant Growth-Promoting Rhizobacteria (PGPR) are typically isolated from rhizospheric soils or plant tissues and cultured in nutrient-rich media such as Luria-Bertani (LB) broth, Nutrient Broth (NB), Tryptic Soy Broth (TSB), or Yeast Mannitol Broth (YMB), depending on the bacterial species. The incubation temperature ranges from 28°C to 32°C, and the cultures are typically grown for 24–72 hours under shaking conditions (120–180 rpm) to achieve high cell density (OD600 ≈ 1.0). Commonly used PGPR genera include Bacillus, Pseudomonas, Azospirillum, Devosia, Rhizobium, and Bradyrhizobium (Naamala and Smith, 2020; Basu et al., 2021).
PGPR are applied to crops via various methods, including seed coating, soil drenching, foliar sprays, or root dips. Application rates vary with method and strain but typically range from 107 to 109 CFU mL-1 for inoculation. These microbes have been widely studied in crops like soybean, maize, canola, wheat, tomato, cassava, and rice, among others.
To obtain CFSs, bacterial cultures are centrifuged at 8,000–12,000 rpm for 10–20 minutes at 4°C to pellet cells. The supernatant is then filtered (usually with 0.22 μm filters) to remove residual cells. In some protocols, cell lysis techniques like ultrasonication, freeze-thaw cycles, or enzymatic treatment are applied to release intracellular metabolites before filtration (Naamala et al., 2022; Shah et al., 2022a).
CFSs are rich in bioactive metabolites, including phytohormones (e.g., indole-3-acetic acid), volatiles, siderophores, enzymes, antimicrobial peptides, and lipo-chitooligosaccharides (LCOs). These components help enhance nutrient uptake, stress tolerance, and pathogen resistance in plants. The supernatants are either used fresh or preserved through freeze-drying, spray drying, or cold storage (4°C) with stabilizers to prolong shelf-life (Pellegrini et al., 2020). An integrated overview of the biochemical and physiological mechanisms triggered by CFS in plants is illustrated in Figure 1.
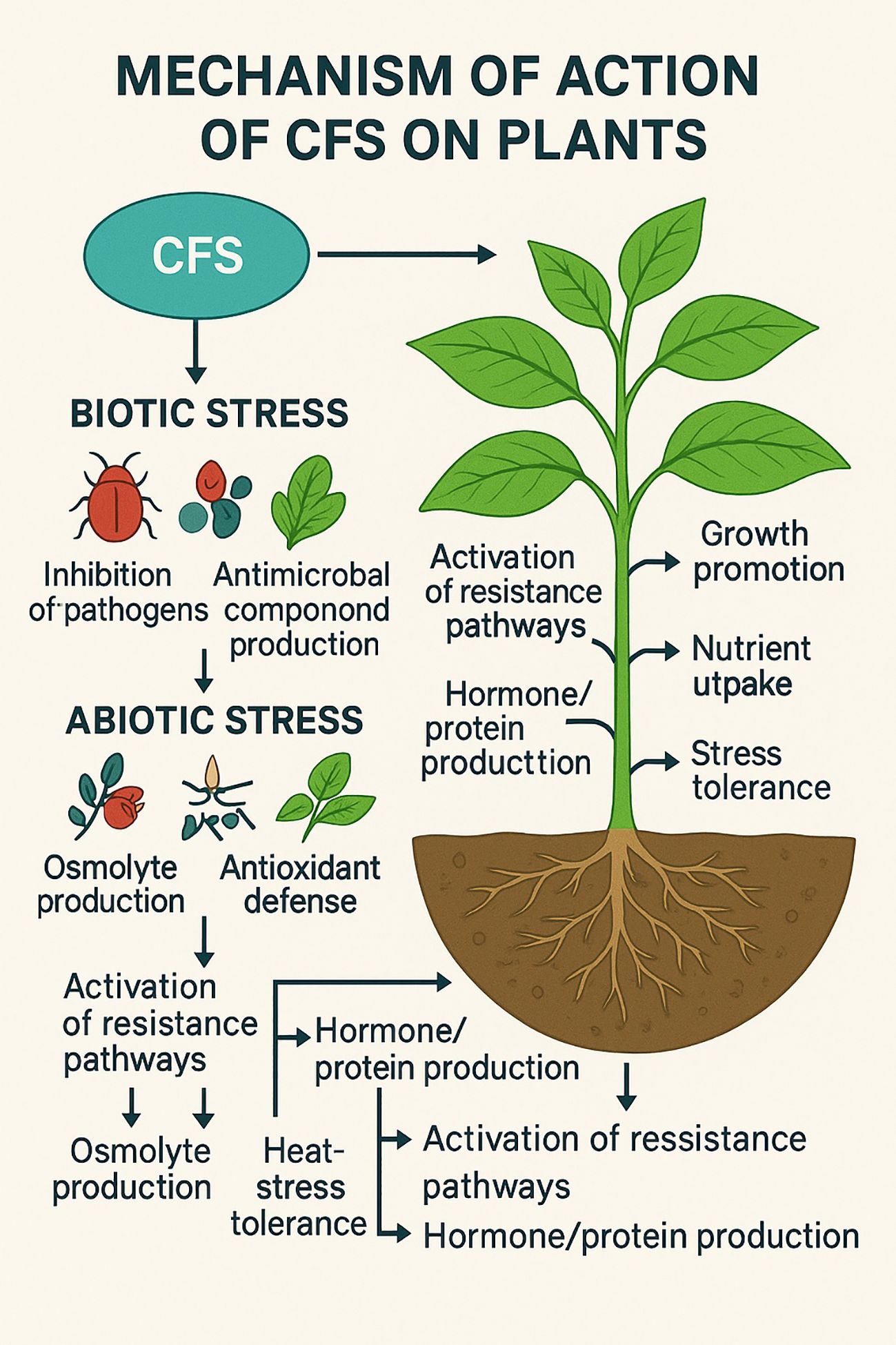
Figure 1. Mechanisms by which cell-free supernatants enhance plant performance under biotic and abiotic stress conditions.
For field use, CFSs can be applied via seed priming, soil application, or foliar spray, typically at concentrations ranging from 5% to 50% (v/v) or 50 to 200 mg L-1, depending on the crop, growth stage, and environmental conditions. CFS treatments have shown strong performance under salinity, drought, heat, and pathogen pressure, especially in soybean, maize, and canola (Tewari et al., 2020; Monjezi et al., 2023).
This practical knowledge is vital for translating lab-scale benefits of PGPR and their metabolites into field-applicable bioformulations, helping ensure consistency and effectiveness in diverse agricultural systems. A comparison of various methods used for the production of CFSs is provided in Table 2, outlining their respective advantages, constraints, and potential applications in agricultural settings.
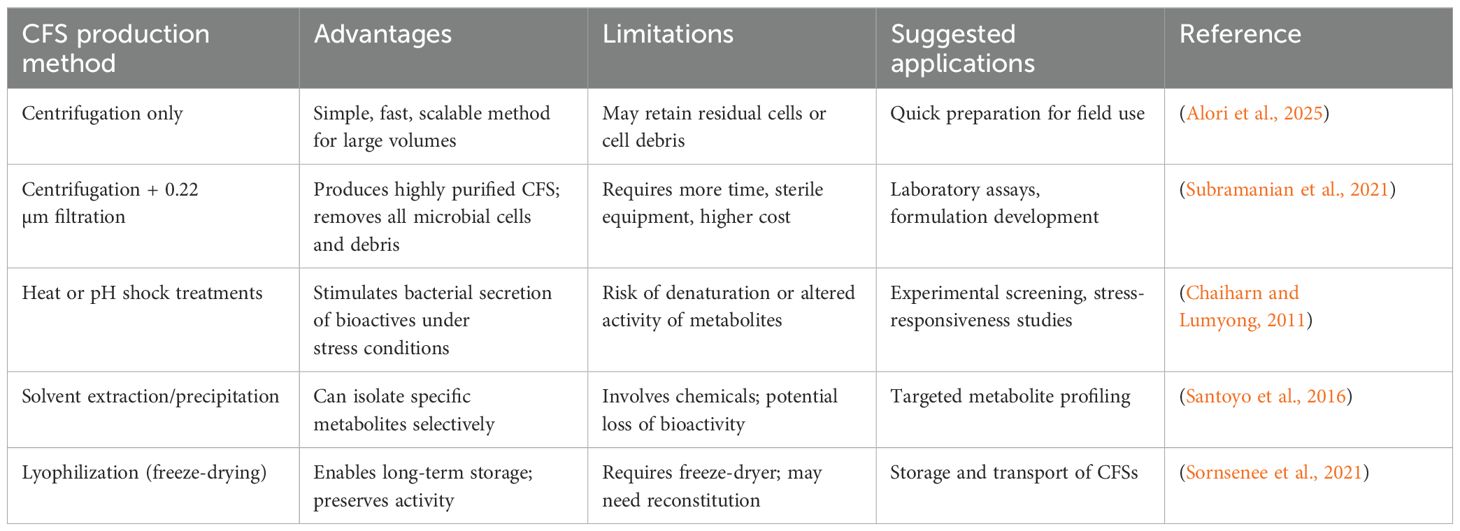
Table 2. Comparative summary of commonly used methods for the preparation of CFSs from plant growth-promoting rhizobacteria, highlighting their advantages, limitations, and suggested applications.
6 Field applications of CFSs: emerging evidence and future prospects
Although the majority of studies on CFSs have been conducted under laboratory and greenhouse conditions—where CFSs have demonstrated beneficial effects on seed germination, plant growth, and stress tolerance—recent research has begun to provide evidence of their efficacy under field conditions. For example, the application of a biosurfactant-rich CFS from Bacillus subtilis significantly reduced black scurf disease incidence in potato by 50% in a field setting (Hussain and Khan, 2020). Similarly, concentrated metabolites derived from Rhizobium tropici and Bradyrhizobium diazoefficiens enhanced grain yields of maize and soybean in multi-location field trials (Marks et al., 2013). Furthermore, a combinational bioformulation comprising Bradyrhizobium cells, its CFS, and exopolysaccharides led to significant improvements in pigeon pea growth and nodulation under field conditions (Tewari et al., 2020). These findings highlight the promising potential of CFS-based products in real-world agricultural systems. However, to fully realize this potential, further well-replicated and crop-specific field trials are needed to assess the consistency, scalability, and economic viability of CFS applications across diverse agroecological zones.
7 Comparative analysis of microbial inoculants and extracellular bioactive compounds: evaluating pros and cons
The utilization of beneficial microorganisms to enhance plant growth and alleviate environmental stresses has garnered significant attention in agricultural research. However, despite substantial evidence highlighting the potential of microbe-based fertilizers and growth enhancers, their practical application on commercial farms faces several limitations. These constraints lead to inconsistencies in plant responses to microbial interventions, especially in the presence of unpredictable and adverse environmental conditions in croplands. Therefore, addressing these challenges is crucial to achieving consistent and reliable outcomes when implementing live beneficial microbes in real-world field conditions, which encompass diverse soils, crops, and climatic nuances (Rilling et al., 2019; Naamala and Smith, 2020; Tewari et al., 2020). The effectiveness of plant growth-promoting rhizobacteria (PGPR) in soil inoculations can face challenges due to various factors, particularly soil properties influencing microbial colonization and the expression of their biocontrol mechanisms. Abiotic features, including soil pH, texture, moisture content, temperature, oxygen levels, and nutrient availability, intricately affect PGPR colonization and the display of their beneficial traits. Temperature plays a crucial role as it influences microbial growth rates and enzymatic activities, impacting the diversity of biostimulants produced by PGPR. Soil moisture content is another crucial determinant, shaping microbial growth dynamics and functional diversity. Changes in soil water content can hinder successful PGPR inoculation; insufficient water can limit microbial growth, while excess moisture can create anaerobic conditions, hampering PGPR activities (Clark et al., 2009; Borowik and Wyszkowska, 2016; Liu et al., 2025). Additionally, soil pH significantly influences the composition and diversity of beneficial soil microorganisms. Even slight deviations from the optimal pH range can cause shifts in microbial biostimulant composition, potentially undermining the benefits of microbial inoculants for plant production (Kaur et al., 2019; Wang et al., 2022).
Moreover, the performance of PGPR is influenced by biological constraints, with specific root-secreted compounds regulating interactions with various biotic and abiotic components of the environment. The nature and magnitude of these root-secreted compounds vary with soil properties, nutrient availability, plant age, and physiological state. These compounds act as mediators of plant-microbe associations, ultimately affecting the efficacy of microbial inoculants (Bais et al., 2006). Technical challenges also pose significant hurdles in the application of PGPR. As living formulations, these biofertilizers require precise storage conditions at appropriate temperatures and durations to maintain viability. Ensuring the survival of these microbes during production, distribution, and storage is crucial, as the shelf-life of microbial inoculants is often shorter than that of chemical fertilizers, leading to potential financial losses (Ngampimol and Kunathigan, 2008; Brar et al., 2012; Arriel-Elias et al., 2018; Basu et al., 2021).
In response to these limitations, the use of CFSs derived from beneficial bacteria has emerged as a promising approach. These extracellular bioactive compounds offer an innovative solution that addresses various challenges associated with PGPR application. Extracted from broth cultures through various techniques, these compounds can provide alternative methods for enhancing crop production and addressing the variability of environmental conditions. Importantly, CFS technologies avoid reliance on specific microbial strains, potentially yielding more consistent results, especially under varying conditions imposed by climate change (Naamala and Smith, 2020; 2021). In conclusion, the implementation of microbial inoculants and extracellular bioactive compounds involves a complex interplay of factors that determine their efficacy and viability within agricultural contexts. Overcoming the challenges posed by environmental variability, soil properties, and technical constraints requires a comprehensive understanding of these factors. Innovative approaches such as CFSs hold promise in mitigating the limitations associated with microbial interventions in modern agriculture.
While CFSs have emerged as a promising alternative to live microbial inoculants, their use is not without limitations. One key disadvantage is the short-lived nature of their effects, as CFSs lack the ability to replicate and sustain long-term interactions in the rhizosphere, unlike live PGPR which can colonize and adapt dynamically to their environment (Naamala and Smith, 2021). This can result in a limited duration of biostimulatory or biocontrol activity, necessitating repeated applications or complementary strategies such as slow-release formulations.
Furthermore, although CFSs eliminate certain challenges associated with microbial viability, their activity is still influenced by soil physicochemical properties, including pH, texture, organic matter content, moisture levels, and temperature. These factors can affect metabolite stability, diffusion rates, and interaction with root exudates, ultimately shaping their efficacy (Pellegrini et al., 2020; Castaldi et al., 2021). For instance, high soil pH or excessive clay content may bind or degrade bioactive compounds, while extreme moisture fluctuations can alter the bioavailability of applied metabolites, similarly to how they impact PGPR colonization.
Therefore, the claim that CFSs are categorically “better” than PGPR inoculants under stress-prone field conditions must be viewed contextually, rather than universally. In some environments, particularly where soil microbial competition, shelf-life concerns, or colonization failures hinder PGPR efficacy, CFSs may offer practical advantages. However, under conditions where sustained, root-associated activity is critical, traditional PGPR inoculation may remain superior. A combined or integrated approach—such as co-application of live PGPR with their CFSs or encapsulated metabolites—could provide synergistic benefits, offering both immediate and prolonged effects while buffering environmental variability. A comparative overview of PGPR and their CFSs regarding application requirements, risks, and modes of action is illustrated in Figure 2.
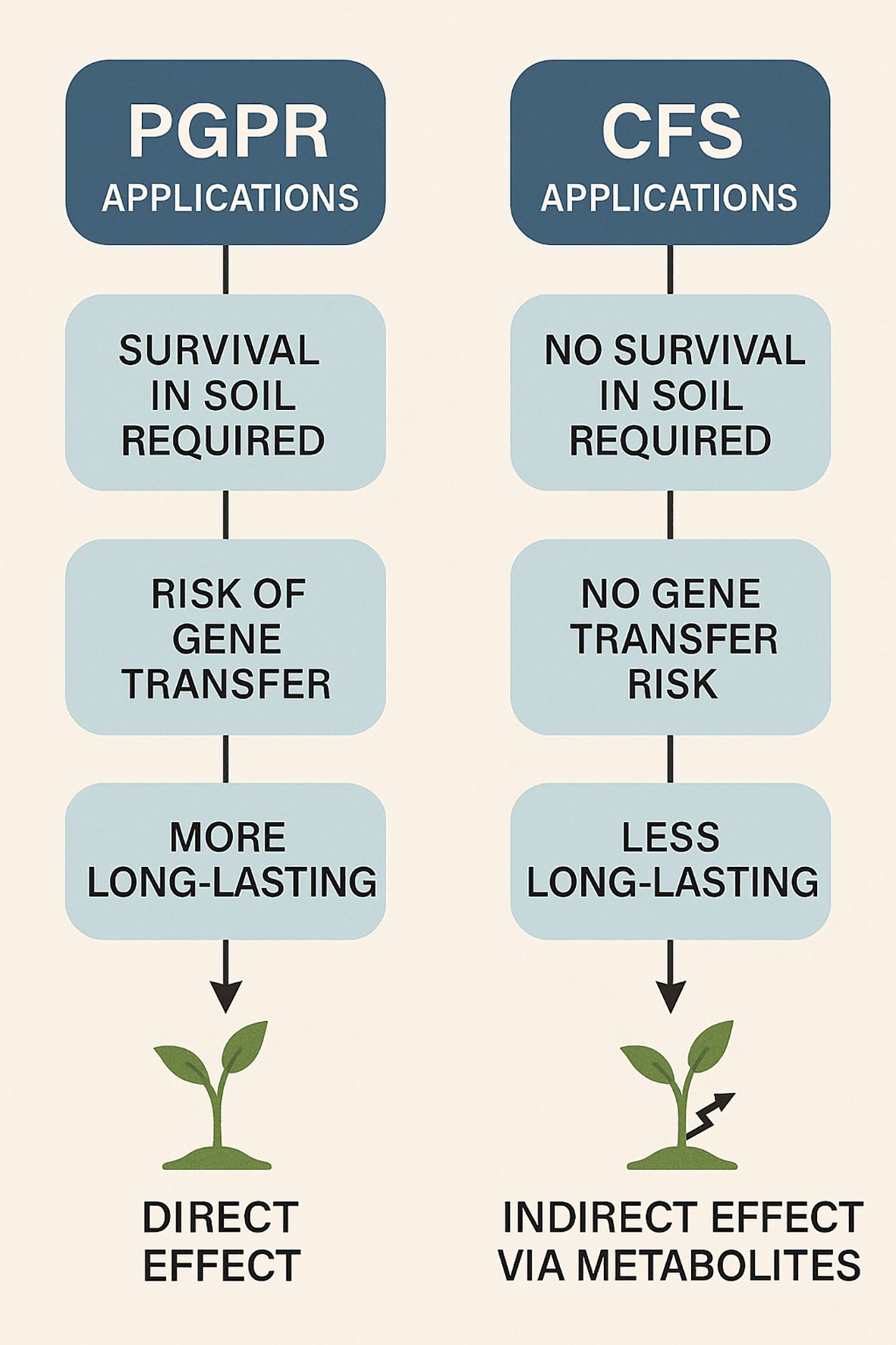
Figure 2. A visual comparison between PGPR and CFS-based applications, highlighting differences in survival, biosafety, durability, and mode of action.
However, despite the promising attributes of PGPR-derived CFSs, several challenges remain that may limit their large-scale agricultural application. For instance, bioactive metabolites within CFSs are prone to degradation in soil due to microbial activity, UV radiation, and abiotic factors, potentially reducing their efficacy (Yakhin et al., 2017; Woo and Pepe, 2018). Moreover, the composition and concentration of metabolites can vary depending on microbial strain, growth phase, and medium conditions, complicating standardization (Vurukonda et al., 2016). From a practical perspective, large-scale production and formulation of stable and cost-effective CFS-based biostimulants also remains a bottleneck (Berg et al., 2020). These limitations underscore the need for further research to optimize formulations, evaluate long-term field performance, and assess the economic viability of CFS applications.
8 Cell-free supernatants as biostimulants and biocontrol agents: a promising path for agricultural enhancement
The search for sustainable ways to boost plant growth has led to interest in organic biostimulants. These compounds provide a practical way to strengthen plant development, whether applied to seeds, young plants in the soil, or through sprays on leaves. The recent emergence of Cell-Free Supernatants from plant growth-promoting rhizobacteria (PGPR-CFSs) is a new approach with the potential to offer many benefits to plants. Plants recognize and respond to these CFSs, which act as facilitators for resource acquisition, protection against pathogens, and triggers for growth mechanisms related to the specific microorganism involved (Naamala and Smith, 2020; Monjezi et al., 2023). Microbes produce compounds that, while not always necessary for direct growth, can work in a stimulus-response system. Various stimuli, both well-known and less understood, trigger the activation of hidden gene clusters in microorganisms, resulting in the creation of useful compounds. This collection includes antibiotics, pigments, growth hormones, and signaling molecules between organisms, all positively linked to the production of valuable secondary metabolites (Singh et al., 2019; Naamala and Smith, 2021). Furthermore, CFSs not only aid in plant growth and health but also help maintain populations of PGPR, including rhizobia, by promoting the growth of the host plants on which these microorganisms rely (Tewari et al., 2020).
8.1 CFSs in mitigating abiotic stress
Research findings emphasize the potential of compounds derived from bacteria in enhancing plant growth across various stages of development, from seed germination to crop maturation. This is especially relevant in challenging conditions, such as salinity stress. Studies demonstrate the germination-promoting abilities of different microbial CFSs under salt stress. For instance, cell-free supernatants from salt-tolerant Bacillus strains have proven effective in improving corn germination and seedling growth under salinity stress (Naamala et al., 2022; Yaghoubian et al., 2022). Similarly, the combined application of flavonoids and cell-free supernatants from Devosai sp. has been observed to boost canola and soybean seed germination, particularly in high salt stress conditions (Shah et al., 2022a). Metabolites produced by rhizobial cells positively impact the growth and grain yield of maize and soybean plants (Marks et al., 2013). Tewari et al. (2020) explain the effectiveness of a treatment involving Bradyrhizobium sp. IC-4059, its cell-free culture supernatant, and exopolysaccharides (EPS) in promoting pigeon pea growth, not only in initial stages but also throughout later plant development. Buensanteai et al. (2013) demonstrated the growth-enhancing impact of phytohormones and extracellular proteins in the CFS of Bacillus sp. strain CaSUT007 broth cultures, leading to significant increases in cassava root and shoot lengths, as well as biomass production. In a recent study by Monjezi et al. (2023), the efficacy of CFS derived from a Devosia strain was further confirmed. Specifically, the CFS from Devosia sp. strain SL43 was investigated for its potential to improve soybean (Glycine max L.) seed vigor index and final germination, mitigating the detrimental impacts of salt stress. These findings highlight the significant positive effects of microbe-based CFS, emphasizing its ability to alleviate stress on plants and provide valuable insights into how microbial-derived compounds can be potent tools for enhancing plant resilience and productivity, especially in challenging environmental conditions.
Two significant signal compounds, lipo-chitooligosaccharides (LCOs) and thuricin 17, have emerged as better characterized players in influencing plant responses to various environmental stresses. These compounds have garnered attention for their potential to enhance plant resilience to stressful conditions. LCOs, acting as inter-organismal signals during the establishment of the legume-rhizobia nitrogen-fixing symbiosis, display remarkable abilities. They not only induce the formation of nitrogen-fixing nodules in leguminous host plants but also orchestrate a cascade of responses that contribute to plant adaptability to changing conditions. Similarly, thuricin 17 plays a potent role, enriching the spectrum of plant responses to a range of abiotic stressors (John McIver et al., 2007; Lee et al., 2009; Gough and Cullimore, 2011; Schwinghamer et al., 2015; Tanaka et al., 2015; Subramanian et al., 2016; Nazari and Smith, 2020). As observed in the study of Lyu et al. (2020), thuricin 17 acts as a constitutive agent, enhancing the plant stress resilience. Schwinghamer et al. (2016) validated these findings, demonstrating the tangible consequences of thuricin 17 and LCOs in saline and temperature stress conditions. Their research highlighted positive outcomes, leading to increased biomass production and the robust establishment of root systems, with clear implications for overall plant vitality. In a parallel revelation, the influential impact of LCOs became evident as they significantly influenced the rate and uniformity of canola seed germination under stressfully low temperature conditions—a crucial attribute for successful early spring sowing, initial crop establishment and agricultural productivity (Schwinghamer et al., 2015). These diverse signal compounds contribute depth to the narrative of plant growth, acting as conductors of intricate biochemical symphonies that fortify the plant’s physiological resilience against adversities. Their power to shape plant development and direct adaptive biochemical mechanisms offers an enticing avenue for scientific exploration and practical implementation in steering agricultural landscapes toward prosperity and sustainability. As we illuminate the complex details of these signal compounds, driven by their role in sculpting plant responses and adaptation, we embark on a promising path toward enhancing agricultural resilience to a wide range of stresses including those associated with climate change, nurturing productivity, and improving nutritional yields—while maintaining the integrity of our environment.
8.2 CFSs as biocontrol agents against pathogens
In addition to their effects on abiotic stress, CFSs have shown promise as biocontrol agents in combating plant pathogens. Ecological pressures stemming from excessive chemical pesticide usage to enhance plant growth under biotic stress conditions have underscored the urgency of substituting them with biopesticides. The utilization of CFSs in agriculture presents a promising avenue as a biocontrol agent against pathogens. Bacteria sourced from both domestic and undomesticated plants emerge as pivotal reservoirs of potential bioagents, owing to their varied adaptive mechanisms developed for endophytic survival (Miljaković et al., 2020; Oleńska et al., 2020). Castaldi et al. (2021) examined spore-forming bacteria from a salt-pan rhizobacterium for traits aiding plant growth and combating the plant pathogen Macophomina phaseolina. Bacillus vallismortis strain RHFS10 displayed potent antifungal effects, and its cell-free supernatants were effective at significantly lower inhibitory concentrations than a commercial fungicide, suggesting it as a promising, eco-friendly option for controlling the plant-harming fungus M. phaseolina. For example, numerous studies have highlighted the role of pathogen-antagonistic Pseudomonas types in disease reduction in soil. In this context, some studies found a connection between the quantity of Pseudomonas cells per gram of soil and the soil’s disease-fighting capacity. They also discovered that diverse Pseudomonas strains, including those in varying environments, can generate compounds with varying effects on plant pathogens. A novel approach employing CFSs in the laboratory could aid in producing and applying the most effective compounds against pathogens, considering different environmental and soil conditions (Bhattacharjee et al., 2023; Khatri et al., 2023; Peng et al., 2023; Ranjan et al., 2023). A conceptual overview of how PGPR and their metabolites influence plant stress responses and agricultural sustainability is provided in Figure 3.

Figure 3. Conceptual representation of microbial-based solutions in sustainable agriculture. The schematic illustrates how plant growth-promoting rhizobacteria (PGPR) and their CFSs contribute to plant resilience by mitigating abiotic stresses (such as salinity, drought, temperature extremes) and biotic stresses (such as pathogen attacks). Beneficial microbial metabolites enhance seed germination, root development, nutrient uptake, and systemic resistance, promoting sustainable crop growth under challenging environmental conditions.
9 Conclusions
Dealing with climate change, agricultural stress, and a growing population requires a comprehensive approach. Climate change, with its rapidly developing and human-made conditions, causes problems like water scarcity and unpredictable temperatures, affecting crops. Meanwhile, the increasing global human population puts more pressure on essential resources, making these challenges more complex. While trying to boost agricultural productivity is an important goal, intensification of standard methods may have unintended consequences and trade-offs for the environment. Considering the complex interplay of factors, it is important to have a clear strategy that addresses multiple priorities. This involves adopting coordinated approaches to tackle the adverse effects of climate change and adjusting agricultural practices to meet evolving conditions. This comprehensive strategy aims to navigate the challenges posed by climate change, ensure food security, and sustainably manage resources amid population growth. Furthermore, plants face challenges, especially from non-living factors like extreme temperatures and soil salinity, complicating modern agriculture.
The increasing prevalence of soil salinity emphasizes the urgent need to address this challenge, which often surpasses, in severity, even the most advanced predictive models. Research efforts are therefore focused on developing versatile strategies, including innovative soil improvement techniques tailored to mitigate the extensive impacts of soil salinity. The story of sustainable and regenerative farming gets better with the increasing importance of plant biostimulants. Together, substances and microorganisms work to support plant health and encourage growth, all while aligning with the goal of preserving the environment. The organized progress and careful use of microbial-based biostimulants have the potential to contribute to global food security. However, the road ahead is filled with challenges that highlight the need for dedicated research efforts and a deep understanding of the complex relationships between plants and the various soil microorganisms.
Through collaborative scientific efforts, the improvement of biostimulant effectiveness and the cultivation of positive farming approaches global food security might be able to be maintained or increased in the face of climate change. In this a complex and detailed context, the idea of using plant growth-promoting bacteria (PGPB) emerges as a powerful approach to boost both plant growth and the inherent resilience in agricultural systems. Among these helpful bacteria, PGPR play a key role, strategically positioned in the root area to coordinate various growth-enhancing mechanisms. This includes a symphony of actions, such as adjusting nutrient accessibility and working in harmony with the subtle regulation of gene expression in plants. Unlike traditional chemical methods, using PGPR strategically has the potential to increase yields and improve soil fertility. At the same time, these microorganisms influence the nearby rhizosphere environment, signaling a significant shift in modern agricultural practices. The significant impact of the microbiome in influencing gene expression highlights the depth of the symbiotic partnership that has the potential to reshape current agricultural approaches. The vital role played by PGPR in alleviating the harmful effects of stresses showcases their diverse capabilities. This involves coordinating growth processes that result in the production of stress-reducing substances, effectively counteracting oxidative stress and providing plants with an improved survival strategy in the face of adversity. Hence, the potential of using PGPR as a sustainable agricultural strategy seems very bright, outlining a path that is set to enhance crop production while carefully maintaining the overall ecological balance.
The use of beneficial microorganisms to enhance plant growth and mitigate environmental stress presents a promising avenue for sustainable agriculture. Among these, microbial CFSs have emerged as an innovative solution, offering many of the benefits of live microbes while overcoming limitations related to survival, colonization, and environmental variability. However, the practical implementation of these microbial strategies—whether using PGPR or their derived metabolites—requires careful consideration of local soil conditions, including pH, moisture, temperature, and nutrient availability. The complex interplay between plant, microbe, and environment necessitates a deep and nuanced understanding to achieve consistent results. While laboratory and greenhouse studies have reported encouraging effects of CFSs under abiotic stresses such as salinity and drought, their real-world application remains limited. This lack of field-based validation highlights a critical gap in the literature. To bridge this divide, future efforts must prioritize optimizing CFS production, formulation, and delivery systems, alongside conducting robust, replicated field trials. Ultimately, with continued interdisciplinary research and scientific collaboration, microbial biostimulants hold immense potential to transform agriculture by enhancing crop resilience, improving soil health, and contributing meaningfully to global food security.
Author contributions
NM: Writing – original draft. HE: Writing – review & editing. RL: Writing – review & editing. ML: Writing – review & editing. DS: Conceptualization, Funding acquisition, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. The authors would like to acknowledge the support for this review paper was provided through the Biomass Canada Cluster (BMC), which is funded through Agriculture and Agri-Food Canada’s AgriScience program and industry partners.
Conflict of interest
Author RL was employed by the company SeaPurAgro Inc.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that Generative AI was used in the creation of this manuscript. Generative AI tools (ChatGPT) were used to improve the language and clarity of the manuscript, and to create schematic figures. The scientific content, interpretations, and conclusions are solely those of the authors.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Ahmad A., Diwan H., and Abrol Y. P. (2010). Global climate change, stress and plant productivity. In: Abiotic Stress Adaptation in Plants. Pareek A., Sopory S., and Bohnert H. (eds) Springer, Dordrecht. doi: 10.1007/978-90-481-3112-9_23
Ahmed S., Heo T.-Y., Roy Choudhury A., Walitang D. I., Choi J., and Sa T. (2021). Accumulation of compatible solutes in rice (Oryza sativa L.) cultivars by inoculation of endophytic plant growth promoting bacteria to alleviate salt stress. Appl. Biol. Chem. 64, 1–14. doi: 10.1186/s13765-021-00638-x
Alori E. T., Onaolapo A. O., and Ibaba A. L. (2025). Cell free supernatant for sustainable crop production. Front. Sustain. Food Syst. 9. doi: 10.3389/fsufs.2025.1549048
Arce G. D., Pedersen P., and Hartzler R. G. (2009). Soybean seeding rate effects on weed management. Weed Technol. 23, 17–22. doi: 10.1614/WT-08-060.1
Arif I., Batool M., and Schenk P. M. (2020). Plant microbiome engineering: expected benefits for improved crop growth and resilience. Trends Biotechnol. 38, 1385–1396. doi: 10.1016/j.tibtech.2020.04.015
Arriel-Elias M. T., Oliveira M. I., Silva-Lobo V. L., Filippi M. C. C., Babana A. H., Conceição E. C., et al. (2018). Shelf life enhancement of plant growth promoting rhizobacteria using a simple formulation screening method. Afr. J. Microbiol. Res. 12, 115–126. doi: 10.5897/AJMR2017.8787
Assaha D. V., Ueda A., Saneoka H., Al-Yahyai R., and Yaish M. W. (2017). The role of Na+ and K+ transporters in salt stress adaptation in glycophytes. Front. Physiol. 8, 509. doi: 10.3389/fphys.2017.00509
Athar H. U., Zulfiqar F., Moosa A., Ashraf M., Zafar Z. U., Zhang L., et al. (2022). Salt stress proteins in plants: An overview. Front. Plant Sci. 13. doi: 10.3389/fpls.2022.999058
Ayuso-Calles M., Flores-Félix J. D., and Rivas R. (2021). Overview of the role of rhizobacteria in plant salt stress tolerance. Agronomy 11 (9), 1759. doi: 10.3390/agronomy11091759
Bais H. P., Weir T. L., Perry L. G., Gilroy S., and Vivanco J. M. (2006). The role of root exudates in rhizosphere interactions with plants and other organisms. Annu. Rev. Plant Biol. 57, 233–266. doi: 10.1146/annurev.arplant.57.032905.105159
Basu A., Prasad P., Das S. N., Kalam S., Sayyed R., Reddy M., et al. (2021). Plant growth promoting rhizobacteria (PGPR) as green bioinoculants: recent developments, constraints, and prospects. Sustainability 13, 1140. doi: 10.3390/su13031140
Berg G., Rybakova D., Fischer D., Cernava T., Vergès M.-C. C., Charles T., et al. (2020). Microbiome definition re-visited: old concepts and new challenges. Microbiome 8, 103. doi: 10.1186/s40168-020-00875-0
Bharti N. and Barnawal D. (2019). “Amelioration of salinity stress by PGPR: ACC deaminase and ROS scavenging enzymes activity,” in PGPR amelioration in sustainable agriculture Singh A. K., Kumar A., Kumar P. K., and Singh, PGPR Amelioration in Sustainable Agriculture eds. (Woodhead Publishing), 85–106. doi: 10.1016/B978-0-12-815879-1.00005-7
Bhattacharjee A., Dubey S., and Sharma S. (2023). Next-generation bioformulations” for plant growth promotion and stress mitigation: A promising approach for sustainable agriculture. J. Plant Growth Regul. 42, 6741–6759. doi: 10.1007/s00344-023-10996-z
Borowik A. and Wyszkowska J. (2016). Soil moisture as a factor affecting the microbiological and biochemical activity of soil. Plant Soil Environ. 62 (6), 250–255. doi: 10.17221/158/2016-PSE
Brar S. K., Sarma S., and Chaabouni E. (2012). Shelf-life of biofertilizers: an accord between formulations and genetics. J. Biofertil Biopestici 3 (5), 1–2. doi: 10.4172/2155-6202.1000e109
Buensanteai N., Sompong M., Thamnu K., Athinuwat D., Brauman A., and Plassard C. (2013). The plant growth promoting bacterium Bacillus sp. CaSUT007 produces phytohormone and extracellular proteins for enhanced growth of cassava. Afr. J. Microbiol. Res. 7, 4949–4954. doi: 10.5897/AJMR12.1839
Castaldi S., Petrillo C., Donadio G., Piaz F. D., Cimmino A., Masi M., et al. (2021). Plant growth promotion function of Bacillus sp. strains isolated from salt-pan rhizosphere and their biocontrol potential against Macrophomina phaseolina. Int. J. Mol. Sci. 22, 3324. doi: 10.3390/ijms22073324
Çatav Ş.S., Surgun-Acar Y., and Zemheri-Navruz F. (2021). Physiological, biochemical, and molecular responses of wheat seedlings to salinity and plant-derived smoke. South Afr. J. Bot. 139, 148–157. doi: 10.1016/j.sajb.2021.02.011
Chaiharn M. and Lumyong S. (2011). Screening and optimization of indole-3-acetic acid production and phosphate solubilization from rhizobacteria aimed at improving plant growth. Curr. Microbiol. 62, 173–181. doi: 10.1007/s00284-010-9674-6
Chauhan A., Saini R., and Sharma J. C. (2021). Plant growth promoting rhizobacteria and their biological properties for soil enrichment and growth promotion. J. Plant Nutr. 45, 273–299. doi: 10.1080/01904167.2021.1952221
Clark J. S., Campbell J. H., Grizzle H., Acosta-Martìnez V., and Zak J. C. (2009). Soil microbial community response to drought and precipitation variability in the Chihuahuan Desert. Microbial Ecol. 57, 248–260. doi: 10.1007/s00248-008-9475-7
Corwin D. L. (2021). Climate change impacts on soil salinity in agricultural areas. Eur. J. Soil Sci. 72, 842–862. doi: 10.1111/ejss.13010
D’Addabbo T., Laquale S., Perniola M., and Candido V. (2019). Biostimulants for plant growth promotion and sustainable management of phytoparasitic nematodes in vegetable crops. Agronomy 9, 616. doi: 10.3390/agronomy9100616
de Leija A. C., King S. L., and Hawkins T. S. (2022). Seed germination responses to salinity for three rare wetland plants of spring-fed arid systems. J. Arid Environments 199, 104705. doi: 10.1016/j.jaridenv.2021.104705
Dhankher O. P. and Foyer C. H. (2018). Climate resilient crops for improving global food security and safety. Plant Cell Environ. 41, 877–884. doi: 10.1111/pce.13207
Dong C., Wang G., Du M., Niu C., Zhang P., Zhang X., et al. (2020). Biostimulants promote plant vigor of tomato and strawberry after transplanting. Scientia Hortic. 267, 109355. doi: 10.1016/j.scienta.2020.109355
Eichmann R., Richards L., and Schäfer P. (2021). Hormones as go-betweens in plant microbiome assembly. Plant J. 105, 518–541. doi: 10.1111/tpj.15135
Etesami H. and Adl S. M. (2020). Plant growth-promoting Rhizobacteria (PGPR) and their action mechanisms in availability of nutrients to plants. In: Phyto-Microbiome in Stress Regulation. Environmental and Microbial Biotechnology. Kumar M., Kumar V., and Prasad R. (eds) Singapore: Springer. doi: 10.1007/978-981-15-2576-6_9
Gapińska M., Skłodowska M., and Gabara B. (2008). Effect of short-and long-term salinity on the activities of antioxidative enzymes and lipid peroxidation in tomato roots. Acta Physiol. Plant 30, 11–18. doi: 10.1007/s11738-007-0072-z
Gough C. and Cullimore J. (2011). Lipo-chitooligosaccharide signaling in endosymbiotic plant-microbe interactions. Mol. Plant-Microbe Interact. 24, 867–878. doi: 10.1094/MPMI-01-11-0019
Hasanuzzaman M., Parvin K., Anee T. I., Masud A. A. C., and Nowroz F. (2022). Salt stress responses and tolerance in soybean. Physiology. IntechOpen. doi: 10.5772/intechopen.102835
Hu L., Robert C. A., Cadot S., Zhang X., Ye M., Li B., et al. (2018). Root exudate metabolites drive plant-soil feedbacks on growth and defense by shaping the rhizosphere microbiota. Nat. Commun. 9, 2738. doi: 10.1038/s41467-018-05122-7
Huang Z., Wang C., Feng Q., Liou R.-M., Lin Y.-F., Qiao J., et al. (2022). The mechanisms of sodium chloride stress mitigation by salt-tolerant plant growth promoting rhizobacteria in wheat. Agronomy 12, 543. doi: 10.3390/agronomy12030543
Hug J. J., Krug D., and Müller R. (2020). Bacteria as genetically programmable producers of bioactive natural products. Nat. Rev. Chem. 4, 172–193. doi: 10.1038/s41570-020-0176-1
Hussain T. and Khan A. A. (2020). Bacillus subtilis HussainT-AMU and its Antifungal activity against Potato Black scurf caused by Rhizoctonia solani on seed tubers. Biocatalysis Agric. Biotechnol. 23, 101443. doi: 10.1016/j.bcab.2019.101443
Iqbal N., Khan N. A., Ferrante A., Trivellini A., Francini A., and Khan M. (2017). Ethylene role in plant growth, development and senescence: interaction with other phytohormones. Front. Plant Sci. 8, 475. doi: 10.3389/fpls.2017.00475
Isayenkov S. V. and Maathuis F. J. (2019). Plant salinity stress: many unanswered questions remain. Front. Plant Sci. 10, 80. doi: 10.3389/fpls.2019.00080
John McIver C. C., Yuming Bai Y. Y., Schultz B., and McIver A. (2007). Foliar application of lipo-chitooligosaccharides (Nod factors) to tomato (Lycopersicon esculentum) enhances flowering and fruit production. Can. J. Plant Sci. 87, 365–372. doi: 10.4141/P06-164
Kaur C., Selvakumar G., and Ganeshamurthy A. (2019). Acid tolerant microbial inoculants: a requisite for successful crop production in acidic soils. In: Phyto and Rhizo Remediation. Microorganisms for Sustainability. Arora N. and Kumar N. (eds), vol 9. Singapore: Springer. doi: 10.1007/978-981-32-9664-0_10
Khatri S., Chaudhary P., Shivay Y. S., and Sharma S. (2023). Role of fungi in imparting general disease suppressiveness in soil from organic field. Microbial Ecol. 86, 2047–2059. doi: 10.1007/s00248-023-02211-z
Khoshru B., Moharramnejad S., Gharajeh N. H., Asgari Lajayer B., and Ghorbanpour M. (2020). Plant microbiome and its important in stressful agriculture. In: Plant Microbiome Paradigm. Varma A., Tripathi S., and Prasad R. (eds). Springer, Cham. doi: 10.1007/978-3-030-50395-6_2
Kumar A., Singh S., Gaurav A. K., Srivastava S., and Verma J. P. (2020). Plant growth-promoting bacteria: biological tools for the mitigation of salinity stress in plants. Front. Microbiol. 11. doi: 10.3389/fmicb.2020.01216
Lee K. D., Gray E. J., Mabood F., Jung W. J., Charles T., Clark S. R., et al. (2009). The class IId bacteriocin thuricin-17 increases plant growth. Planta 229, 747–755. doi: 10.1007/s00425-008-0870-6
Leontidou K., Genitsaris S., Papadopoulou A., Kamou N., Bosmali I., Matsi T., et al. (2020). Plant growth promoting rhizobacteria isolated from halophytes and drought-tolerant plants: genomic characterisation and exploration of phyto-beneficial traits. Sci. Rep. 10, 14857. doi: 10.1038/s41598-020-71652-0
Li H., Yue H., Li L., Liu Y., Zhang H., Wang J., et al. (2021). Seed biostimulant Bacillus sp. MGW9 improves the salt tolerance of maize during seed germination. AMB Express 11, 74. doi: 10.1186/s13568-021-01237-1
Liu D., Wang Z., Zhu G., Xu A., Zhang R., Bryant R., et al. (2025). Stable soil moisture promotes shoot performance and shapes the root-rhizosphere microbiome. Agric. Water Manage. 310, 109354. doi: 10.1016/j.agwat.2025.109354
Lyu D., Backer R., Subramanian S., and Smith D. L. (2020). Phytomicrobiome coordination signals hold potential for climate change-resilient agriculture. Front. Plant Sci. 11. doi: 10.3389/fpls.2020.00634
Lyu D., Zajonc J., Pagé A., Tanney C. A. S., Shah A., Monjezi N., et al. (2021). Plant holobiont theory: the phytomicrobiome plays a central role in evolution and success. Microorganisms 9, 675–612. doi: 10.3390/microorganisms9040675
Macdonald C. A., Anderson I. C., Khachane A., Singh B. P., Barton C. V. M., Duursma R. A., et al. (2021). Plant productivity is a key driver of soil respiration response to climate change in a nutrient-limited soil. Basic Appl. Ecol. 50, 155–168. doi: 10.1016/j.baae.2020.12.005
Marks B. B., Megías M., Nogueira M. A., and Hungria M. (2013). Biotechnological potential of rhizobial metabolites to enhance the performance of Bradyrhizobium spp. and Azospirillum brasilense inoculants with soybean and maize. AMB Express 3, 21. doi: 10.1186/2191-0855-3-21
Mhatre P. H., Karthik C., Kadirvelu K., Divya K. L., Venkatasalam E. P., Srinivasan S., et al. (2019). Plant growth promoting rhizobacteria (PGPR): A potential alternative tool for nematodes bio-control. Biocatalysis Agric. Biotechnol. 17, 119–128. doi: 10.1016/j.bcab.2018.11.009
Miljaković D., Marinković J., and Balešević-Tubić S. (2020). The significance of bacillus spp. in disease suppression and growth promotion of field and vegetable crops. Microorganisms 8 (7), 1037. doi: 10.3390/microorganisms8071037
Monjezi N., Yaghoubian I., and Smith D. L. (2023). Cell-free supernatant of Devosia sp. (strain SL43) mitigates the adverse effects of salt stress on soybean (Glycine max L.) seed vigor index. Front. Plant Sci. 14. doi: 10.3389/fpls.2023.1071346
Naamala J., Msimbira L. A., Antar M., Subramanian S., and Smith D. L. (2022). Cell-free supernatant obtained from a salt tolerant bacillus amyloliquefaciens strain enhances germination and radicle length under naCl stressed and optimal conditions. Front. Sustain. Food Syst. 6. doi: 10.3389/fsufs.2022.788939
Naamala J. and Smith D. L. (2020). Relevance of plant growth promoting microorganisms and their derived compounds, in the face of climate change. Agronomy 10, 1179. doi: 10.3390/agronomy10081179
Naamala J. and Smith D. L. (2021). Microbial derived compounds, a step toward enhancing microbial inoculants technology for sustainable agriculture. Front. Microbiol. 12. doi: 10.3389/fmicb.2021.634807
Nawaz A., Shahbaz M., Asadullah, Imran A., Marghoob M. U., Imtiaz M., et al. (2020). Potential of salt tolerant PGPR in growth and yield augmentation of wheat (Triticum aestivum L.) under saline conditions. Front. Microbiol. 11. doi: 10.3389/fmicb.2020.02019
Nazari M. and Smith D. L. (2020). A PGPR-produced bacteriocin for sustainable agriculture: A review of thuricin 17 characteristics and applications. Front. Plant Sci. 11. doi: 10.3389/fpls.2020.00916
Ngampimol H. and Kunathigan V. (2008). The study of shelf life for liquid biofertilizer from vegetable waste. AU JT 11, 204–208. Available online at: https://repository.au.edu/handle/6623004553/14656.
Oleńska E., Małek W., Wójcik M., Swiecicka I., Thijs S., and Vangronsveld J. (2020). Beneficial features of plant growth-promoting rhizobacteria for improving plant growth and health in challenging conditions: A methodical review. Sci. Total Environ. 743, 140682. doi: 10.1016/j.scitotenv.2020.140682
Onu O. B., Okoro G. O., Ngasoh F. G. G., and Namessan N. (2019). “The Effect of Climate Change on Abiotic Plant Atress: A review,” in Abiotic and Biotic Stress in Plants. Ed. De Oliveira A. B. (IntechOpen, Rijeka).
Otie V., Udo I., Shao Y., Itam M. O., Okamoto H., An P., et al. (2021). Salinity effects on morpho-physiological and yield traits of soybean (Glycine max L.) as mediated by foliar spray with brassinolide. Plants 10, 541. doi: 10.3390/plants10030541
Otlewska A., Migliore M., Dybka-Stępień K., Manfredini A., Struszczyk-Świta K., Napoli R., et al. (2020). When salt meddles between plant, soil, and microorganisms. Front. Plant Sci. 11. doi: 10.3389/fpls.2020.553087
Pandey P., Ramegowda V., and Senthil-Kumar M. (2015). Shared and unique responses of plants to multiple individual stresses and stress combinations: physiological and molecular mechanisms. Front. Plant Sci. 6. doi: 10.3389/fpls.2015.00723
Pang C.-H. and Wang B.-S. (2008). “Oxidative stress and salt tolerance in plants,” In: Progress in Botany. Lüttge U., Beyschlag W., and Murata J.. (eds), (Berlin, Heidelberg: Springer) 69, 231–245. doi: 10.1007/978-3-540-72954-9_9
Parameswaran K., Sitharanjithan A., Selvaraj M., Chockaiyan U., Subbaraju S., Pandi S., et al. (2021). PGPR: the redeemer of rice from abiotic stress. In: Antioxidants in Plant-Microbe Interaction. Singh H. B., Vaishnav A., and Sayyed R. (eds) (Singapore: Springer), 101–135. doi: 10.1007/978-981-16-1350-0_6
Pareek A., Dhankher O. P., and Foyer C. H. (2020). Mitigating the impact of climate change on plant productivity and ecosystem sustainability. J. Exp. Bot. 71, 451–456. doi: 10.1093/jxb/erz518
Park S., Kim A. L., Hong Y.-K., Shin J.-H., and Joo S.-H. (2021). A highly efficient auxin-producing bacterial strain and its effect on plant growth. J. Genet. Eng. Biotechnol. 19, 179. doi: 10.1186/s43141-021-00252-w
Pellegrini M., Pagnani G., Bernardi M., Mattedi A., Spera D. M., and Gallo M. D. (2020). Cell-free supernatants of plant growth-promoting bacteria: A review of their use as biostimulant and microbial biocontrol agents in sustainable agriculture. Sustainability 12, 9917. doi: 10.3390/su12239917
Peng T., Meng L., Wang Y., Jin L., Jin H., Yang T., et al. (2023). Alterations of the rhizosphere soil microbial community composition and metabolite profiles of Angelica sinensis seedlings by co-application of Nitrogen fixing bacteria and amino acids. Plant Soil 493, 535–554. doi: 10.1007/s11104-023-06248-4
Phang T.-H., Shao G., and Lam H.-M. (2008). Salt tolerance in soybean. J. Integr. Plant Biol. 50, 1196–1212. doi: 10.1111/j.1744-7909.2008.00760.x
Rai P. K., Singh M., Anand K., Saurabh S., Kaur T., Kour D., et al. (2020). “Chapter 4 - Role and potential applications of plant growth-promoting rhizobacteria for sustainable agriculture,” in New and Future Developments in Microbial Biotechnology and Bioengineering. Eds. Rastegari A. A., Yadav A. N., and Yadav N. (Elsevier), 49–60.
Ramakrishna W., Yadav R., and Li K. (2019). Plant growth promoting bacteria in agriculture: Two sides of a coin. Appl. Soil Ecol. 138, 10–18. doi: 10.1016/j.apsoil.2019.02.019
Ranjan A., Rajput V. D., Prazdnova E. V., Gurnani M., Bhardwaj P., Sharma S., et al. (2023). Nature’s antimicrobial arsenal: non-ribosomal peptides from PGPB for plant pathogen biocontrol. Fermentation 9, 597. doi: 10.3390/fermentation9070597
Raza A., Razzaq A., Mehmood S. S., Zou X., Zhang X., Lv Y., et al. (2019). Impact of climate change on crops adaptation and strategies to tackle its outcome: A review. Plants 8, 34. doi: 10.3390/plants8020034
Riaz U., Murtaza G., Anum W., Samreen T., Sarfraz M., and Nazir M. Z. (2021). Plant Growth-Promoting Rhizobacteria (PGPR) as Biofertilizers and Biopesticides. In: Microbiota and Biofertilizers. Hakeem K. R., Dar G. H., Mehmood M. A., and Bhat R. A. (eds) Springer, Cham. doi: 10.1007/978-3-030-48771-3_11
Rilling J. I., Acuña J. J., Nannipieri P., Cassan F., Maruyama F., and Jorquera M. A. (2019). Current opinion and perspectives on the methods for tracking and monitoring plant growth–promoting bacteria. Soil Biol. Biochem. 130, 205–219. doi: 10.1016/j.soilbio.2018.12.012
Sabagh A., Islam M., Skalicky M., Ali Raza M., Singh K., Anwar Hossain M., et al. (2021). Salinity stress in wheat (Triticum aestivum L.) in the changing climate: adaptation and management strategies. Front. Agron. 3, 661932. doi: 10.3389/fagro.2021.661932
Safdar H., Amin A., Shafiq Y., Ali A., Yasin R., Shoukat A., et al. (2019). A review: Impact of salinity on plant growth. Nat. Sci. 17, 34–40. doi: 10.7537/marsnsj170119.06
Sahab S., Suhani I., Srivastava V., Chauhan P. S., Singh R. P., and Prasad V. (2021). Potential risk assessment of soil salinity to agroecosystem sustainability: Current status and management strategies. Sci. Total Environ. 764, 144164. doi: 10.1016/j.scitotenv.2020.144164
Sangiorgio D., Cellini A., Donati I., Pastore C., Onofrietti C., and Spinelli F. (2020). Facing climate change: application of microbial biostimulants to mitigate stress in horticultural crops. Agronomy 10, 794. doi: 10.3390/agronomy10060794
Sansinenea E. (2019). “Bacillus spp.: As Plant Growth-Promoting Bacteria,” in Secondary Metabolites of Plant Growth Promoting Rhizomicroorganisms: Discovery and Applications. Eds. Singh H. B., Keswani C., Reddy M. S., Sansinenea E., and García-Estrada C. (Springer Singapore, Singapore), 225–237.
Santoyo G., Moreno-Hagelsieb G., Orozco-Mosqueda Mdel C., and Glick B. R. (2016). Plant growth-promoting bacterial endophytes. Microbiol. Res. 183, 92–99. doi: 10.1016/j.micres.2015.11.008
Schwinghamer T., Souleimanov A., Dutilleul P., and Smith D. (2015). The plant growth regulator lipo-chitooligosaccharide (LCO) enhances the germination of canola (Brassica napus [L.]). J. Plant Growth Regul. 34, 183–195. doi: 10.1007/s00344-014-9456-7
Schwinghamer T., Souleimanov A., Dutilleul P., and Smith D. (2016). Supplementation with solutions of lipo-chitooligosacharide Nod Bj V (C18:1, MeFuc) and thuricin 17 regulates leaf arrangement, biomass, and root development of canola (Brassica napus [L.]). Plant Growth Regul. 78, 31–41. doi: 10.1007/s10725-015-0072-8
Shah S. Z., Rasheed A., El-Keblawy A., Gairola S., Phartyal S. S., Gul B., et al. (2022b). Inter-provenance variation in seed germination response of a cash crop halophyte Suaeda fruticosa to different abiotic factors. Flora 292, 152079. doi: 10.1016/j.flora.2022.152079
Shah A., Subramanian S., and Smith D. L. (2022a). Seed priming with devosia sp. Cell-free supernatant (CFS) and citrus bioflavonoids enhance canola and soybean seed germination. Molecules 27, 3410. doi: 10.3390/molecules27113410
Shavrukov Y. (2013). Salt stress or salt shock: which genes are we studying? J. Exp. Bot. 64, 119–127. doi: 10.1093/jxb/ers316
Singh B. P., Rateb M. E., Rodriguez-Couto S., Polizeli M., and Li W.-J. (2019). Editorial: microbial secondary metabolites: recent developments and technological challenges. Front. Microbiol. 10. doi: 10.3389/fmicb.2019.00914
Sornsenee P., Chatatikun M., Mitsuwan W., Kongpol K., Kooltheat N., Sohbenalee S., et al. (2021). Lyophilized cell-free supernatants of Lactobacillus isolates exhibited antibiofilm, antioxidant, and reduces nitric oxide activity in lipopolysaccharide-stimulated RAW 264.7 cells. PeerJ 9, e12586. doi: 10.7717/peerj.12586
Spaepen S. and Vanderleyden J. (2011). Auxin and plant-microbe interactions. Cold Spring Harb. Perspect. Biol. 3. doi: 10.1101/cshperspect.a001438
Subramanian S., Ricci E., Souleimanov A., and Smith D. L. (2016). A proteomic approach to lipo-chitooligosaccharide and thuricin 17 effects on soybean germinationUnstressed and salt stress. PloS One 11, e0160660. doi: 10.1371/journal.pone.0160660
Subramanian S., Souleimanov A., and Smith D. L. (2021). Thuricin17 production and proteome differences in bacillus thuringiensis NEB17 cell-free supernatant under naCl stress. Front. Sustain. Food Syst. 5. doi: 10.3389/fsufs.2021.630628
Tanaka K., Cho S.-H., Lee H., Pham A. Q., Batek J. M., Cui S., et al. (2015). Effect of lipo-chitooligosaccharide on early growth of C4 grass seedlings. J. Exp. Bot. 66, 5727–5738. doi: 10.1093/jxb/erv260
Tewari S., Pooniya V., and Sharma S. (2020). Next generation bioformulation prepared by amalgamating Bradyrhizobium, cell free culture supernatant, and exopolysaccharides enhances the indigenous rhizospheric rhizobial population, nodulation, and productivity of pigeon pea. Appl. Soil Ecol. 147, 103363. doi: 10.1016/j.apsoil.2019.103363
Turner T. R., James E. K., and Poole P. S. (2013). The plant microbiome. Genome Biol. 14, 209. doi: 10.1186/gb-2013-14-6-209
Vurukonda S. S. K. P., Vardharajula S., Shrivastava M., and SkZ A. (2016). Enhancement of drought stress tolerance in crops by plant growth promoting rhizobacteria. Microbiological Res. 184, 13–24. doi: 10.1016/j.micres.2015.12.003
Wang T., Cao X., Chen M., Lou Y., Wang H., Yang Q., et al. (2022). Effects of soil acidification on bacterial and fungal communities in the jiaodong peninsula, northern China. Agronomy 12, 927. doi: 10.3390/agronomy12040927
Wang D., Zhao C., Zheng J., Zhu J., Gui Z., and Yu Z. (2021). Evolution of soil salinity and the critical ratio of drainage to irrigation (CRDI) in the Weigan Oasis in the Tarim Basin. CATENA 201, 105210. doi: 10.1016/j.catena.2021.105210
Woo S. L. and Pepe O. (2018). Microbial consortia: promising probiotics as plant biostimulants for sustainable agriculture. Front. Plant Sci. 9. doi: 10.3389/fpls.2018.01801
Xie X., He Z., Chen N., Tang Z., Wang Q., and Cai Y. (2019). The roles of environmental factors in regulation of oxidative stress in plant. BioMed. Res. Int. 2019, 9732325. doi: 10.1155/2019/9732325
Yaghoubian I., Msimbira L. A., and Smith D. L. (2022). Cell-Free Supernatant of Bacillus Strains can Improve Seed Vigor Index of Corn (Zea mays L.) Under Salinity Stress. Front. Sustain. Food Syst. 6. doi: 10.3389/fsufs.2022.857643
Yakhin O. I., Lubyanov A. A., Yakhin I. A., and Brown P. H. (2017). Biostimulants in plant science: A global perspective. Front. Plant Sci. 7. doi: 10.3389/fpls.2016.02049
Yu X., Shi P., Hui C., Miao L., Liu C., Zhang Q., et al. (2019). Effects of salt stress on the leaf shape and scaling of pyrus betulifolia bunge. Symmetry. 11, 991. doi: 10.3390/sym11080991
Zerrouk I. Z., Benchabane M., Khelifi L., Yokawa K., Ludwig-Müller J., and Baluska F. (2016). A Pseudomonas strain isolated from date-palm rhizospheres improves root growth and promotes root formation in maize exposed to salt and aluminum stress. J. Plant Physiol. 191, 111–119. doi: 10.1016/j.jplph.2015.12.009
Zhao X., Liu Y., Jin M., Zhang T., Shi W., and Li C. (2023). Rhizosphere microbial dynamics in response to Desmondesmus sp. ZM-3 and carbon footprint analysis in constructed wetland. J. Cleaner Production 408, 137128. doi: 10.1016/j.jclepro.2023.137128
Keywords: cell-free supernatants, crop resilience, environmental stress, sustainable agriculture, microbial formulations, global food security
Citation: Monjezi N, Eisvand HR, Lee R, Levi M and Smith DL (2025) Navigating complex agricultural challenges: harnessing microbial solutions for sustainable growth and resilience. Front. Agron. 7:1631654. doi: 10.3389/fagro.2025.1631654
Received: 20 May 2025; Accepted: 01 July 2025;
Published: 21 July 2025.
Edited by:
Naser A. Anjum, Aligarh Muslim University, IndiaReviewed by:
Adnan Akhter, University of the Punjab, PakistanMudassir Iqbal, Swedish University of Agricultural Sciences, Sweden
Copyright © 2025 Monjezi, Eisvand, Lee, Levi and Smith. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Donald L. Smith, RG9uYWxkLlNtaXRoQE1jR2lsbC5DYQ==