 Mohammad Almogdad1*
Mohammad Almogdad1* Roma Semaškienė1Akvilė Jonavičienė1
Roma Semaškienė1Akvilė Jonavičienė1 Jūratė Ramanauskienė1Eimantas Venslovas1
Jūratė Ramanauskienė1Eimantas Venslovas1 Aurimas Sabeckis1
Aurimas Sabeckis1 Karolina Lavrukaitė1Yanal Alkuddsi2Manal Almukdad3
Karolina Lavrukaitė1Yanal Alkuddsi2Manal Almukdad3- 1Department of Plant Pathology and Protection, Institute of Agriculture, Lithuanian Research Centre for Agriculture and Forestry, Akademija, Kėdainiai distr., Lithuania
- 2Department of Forest Resources, Economics and Policy, Institute of Forestry, Lithuanian Research Centre for Agriculture and Forestry, Akademija, Kėdainiai distr., Lithuania
- 3Department of Laboratory Medicine, Faculty of Medicine, Damascus University, Damascus, Syria
A field experiment was conducted in Lithuania over three growing seasons (2021–2023) to evaluate the effects of sowing time, seeding rate, and insecticide application on grain damage caused by Bruchus rufimanus, as well as on grain yield and quality. The experiment included both insecticide-treated and untreated plots, along with three seeding rates and three sowing times. Early sowing resulted in the highest grain damage (46.5%), attributed to phenological synchronization with peak weevil oviposition, while late sowing reduced damage by 18.6%. Insecticide application significantly reduced grain damage and increased yield, though the extent of yield improvement depended on sowing time. Overall, seeding rate had minimal influence on yield and pest pressure, although significant variation in grain damage among seeding rates was observed in untreated plots in 2021. Crude protein content (CP) was unaffected by seeding rate but showed inter-annual variation, likely due to environmental conditions. At the optimal sowing time (late April), faba bean yield remained consistently high, and insecticide application had limited impact on yield improvement. Relatively low grain damage under these conditions suggests that insecticide use may be reduced or eliminated. These findings underscore the importance of integrating optimal sowing time with targeted insecticide application to maximize faba bean yield and minimize grain damage in Lithuania.
1 Introduction
Insect pest infestations represent a major challenge in legume cultivation (Fan et al., 2022), often leading to substantial yield losses (Hussien et al., 2022). One of the primary insects that harm faba bean (Vicia faba L.) is the broad bean weevil (Bruchus rufimanus L.) (Hamidi et al., 2021). Adults of B. rufimanus overwinter in protected places like forests, leaf litter and even in stored seeds (Tran et al., 1993). In spring, faba bean begin to flower and adults become active. In order to reach their sexual maturity, the adults begin to consume pollen and nectar (Segers et al., 2021). As soon as the first pods start forming, when daytime temperatures are higher than 15 °C, females start laying eggs (Medjdoub-Bensaad et al., 2007). The broad bean weevil larvae burrow through pod walls and feed on growing faba bean seeds, consuming the endosperm or exterminating the embryo. This feeding strategy significantly reduces the seed quality (Dell’Aglio and Tayeh, 2023), leading farmers to suffer serious economic losses (Segers et al., 2021). Damaged seeds have a major impact on agricultural economic viability and sustainability due to decreased seedling growth (Khelfane-Goucem and Medjdoub-Bensaad, 2016) and decreased germination rates (Titouhi et al., 2015). The most evident harm is the presence of the residual seed coat obscuring the larval aperture in the seeds from which the adult beetles emerge post-pupation (Segers et al., 2021). The proportion of damaged seeds varies between 18.5% and 28.9% (Kaniuczak, 2004). It is predicted that yield losses of 70–100% will occur in the absence of control measures (Gailis et al., 2022). When compared to undamaged seeds, damaged seeds frequently have reduced germination rates. The percentage of germination may drop by 32.6% (Titouhi et al., 2015).
Chemical insecticides have been widely used for managing B. rufimanus (Segers et al., 2021). However, there is a rising desire to decrease the use of chemical control methods as sustainable agriculture practices receive increased attention (Schneider et al., 2023). Insecticides can provide immediate benefits for insect pests control and crop production; however, their use raises serious concerns for human health and the environment (Pathak et al., 2022; Ahmad et al., 2024). This includes environmental pollution (Tang et al., 2021) as well as pesticide resistance (Siddiqui et al., 2023), and harm to non-target beneficial invertebrates such as honeybees, earthworms, natural enemies (predators and parasitoids) (Elhamalawy et al., 2024), and health of the soil (Gunstone et al., 2021). As a result, it is essential to investigate different options for pest management, such as agronomic practices that can naturally reduce insect populations without using chemical pesticides.
Adjusting seeding rate and sowing time are agronomic strategies for integrated pest management (Abbas et al., 2022). The microclimate of an agricultural field is influenced by the date of sowing and the rate of seeding, so they can affect pest infestation (Kaur et al., 2024). Sowing time is crucial, as it can shift crop developmental phenology in relation to the life cycle of insect pests. Depending on sowing time, phenological delays may make the plant parts exploited by weevils more or less accessible during key stages of their life cycle (Hamidi et al., 2021). Adjusting sowing times can also reduce excessive pest population densities, as plants may be less physiologically suitable for insect feeding (Shahid et al., 2014). Such delays can be advantageous by avoiding critical infestation periods, such as the peak activity of the broad bean weevil during the growing season. For instance, Szafirowska (2012) reported that the percentage of damaged seeds decreased with delayed sowing. The findings showed that the dates of sowing had a major impact on the pest’s population densities. As an example, the early sowing of maize exhibited the highest fall armyworm infection rate and density (Mbaidiro et al., 2023).
Furthermore, seeding rates are crucial for yield optimization and pest control. Changing seeding rates leads to varying lengths of the roots because of the competition between plants for supplies like water, light, and nutrients (Münzbergová, 2012; Gao et al., 2021; Xu et al., 2021), possibly decreasing each plant’s vigor. Higher-density sowing may induce a microclimate inside the crop’s canopy that is more or less conducive to the development of specific pests. Insects typically inhabit specific environments, and they may have particular preferences for microhabitats at micro-scales (Inskeep et al., 2021). High-density crops may encourage insect colonization by establishing a favorable environment and perhaps improving the overall infestation rate because the pests may more readily move through such conditions (Török et al., 2021).
Enhancing crop management and increasing yields in faba bean requires an understanding of the relationship between sowing time, seeding rate, insecticide application, and insect infestations. According to studies, by giving the plants the optimal growth conditions possible, proper agronomic techniques (sowing time and seed rate) improve harvest results (Murphy et al., 2022). Moreover, it may minimize insect damage; for instance, the early sowing time allowed most plant blooms to avoid the aphid infestation period (Elsayed et al., 2021). Higher overall yields can result from sowing at the right times, which can also improve plant tolerance to biotic and abiotic stressors and raise seed protein content (Manning et al., 2020; Yasmin et al., 2021). Additionally, research on other legume crops has shown that increasing plant density improves protein levels, which can affect seed quality (Esan et al., 2023).
This study aims to survey the effects of sowing time, seeding rate, and insecticide application on the severity of broad bean weevil infestation and the yield of faba bean. This study offers substantial knowledge to develop integrated pest control strategies by analyzing the effects of these agronomic techniques in treated and untreated fields. By decreasing the need for chemical pesticides and increasing yield potential through sowing practices, the findings could contribute to more sustainable faba bean farming practices.
2 Materials and methods
The study was carried out over three growing seasons (2021–2023) in a field belonging to the Lithuanian Research Centre for Agriculture and Forestry, Institute of Agriculture, located in Akademija, Kėdainiai district. A randomized complete block design (RCBD) was employed, consisting of 72 experimental units (3 sowing dates × 4 seeding rates × 2 insecticide treatments × 3 replicates), with 2.5m buffer zones between blocks to minimize edge effects. Each plot had a total size of 15.0 m2, with dimensions of 10.0 × 1.5m. The sandy loam soil in the experiment area had a pH of 6.1 and an average organic matter level of 3.4%. The average rainfall throughout the growing season from April to August over the research years was 323.4mm. All plots received the same recommended agronomic practices, which included applying uniform fertilization with NPK. Fungicides and herbicides were applied to control weeds and diseases. Cypermethrin in an emulsifiable concentrate (EC) formulation was applied at a dose rate of 0.05 L ha-1, following standard agricultural practices for the control of broad bean weevil. The treatment was carried out once at the end of flowering, when young pods were less than 2cm in length (BBCH stage 69). The experiment was divided into two parallel parts, one treated with insecticides and the other without, to evaluate the effectiveness of managing broad bean weevils. Each part of the experiment included the same combinations of sowing times and seeding rates.
Meteorological data were obtained from the Dotnuva weather station, located approximately 2km from the experimental fields, and managed by the Lithuanian Hydrometeorological Service. Daily air temperature and precipitation data were recorded throughout the faba bean growing season (April–August) from 2021 to 2023. These values were compared to the 1991–2020 standard climatic norm (SCN). During that reference period, the mean annual air temperature was 7.5 °C, and annual precipitation averaged 566 mm, with July typically being the warmest and wettest month. In 2021, average temperatures during the growing season were 0.7 °C above the SCN, with total rainfall exceeding the norm by 16%. The 2022 season featured near-average temperatures but a 49% increase in total precipitation, with particularly heavy rainfall in July. In 2023, average temperatures were 0.6 °C above the norm, while precipitation was 24% below average for the season (Lukošiūtė-Stasiukonienė et al., 2024).
Three sowing times were used over the research years to assess how sowing time impacted faba bean growth and broad bean weevil infestation. I) Early sowing time (April 15, 2021; April 13, 2022; April 25, 2023): when soil conditions were suitable for seedbed preparation in the early spring. II) Normal sowing time (April 29, 2021; April 29, 2022; May 4, 2023): in accordance with agronomic recommendations, sowing was carried out during the optimal time window for faba bean growth in the experimental region, when soil temperatures at sowing depth ranged between 10–14 °C and daytime air temperatures were between 15–20 °C. III) Late sowing time (May 14, 2021; May 11, 2022; May 19, 2023): the plants may avoid the insect pests’ peak activity because sowing occurred after the typical window for faba bean in the region. To evaluate the effects of plant spacing on crop yield and pest severity, the study also looked at four seeding rates. These densities varied from low to high (20, 40, 60, and 80 seeds per m2), designed to evaluate the effects of narrow and wide spacing of plants. When calculating the seed rate, the seed quantity was adjusted in each treatment based on germination rates to achieve the specified seeding rates.
Damage caused by B. rufimanus was assessed in both insecticide-treated and untreated plots. The measurements were carried out at the last phase of fruit development (BBCH stage 79). Thirty pods were randomly collected from various heights on ten plants per plot. Weevil infestation was identified by the presence of a larval entrance hole, a circular exit window, or an adult emergence hole in the seeds.
At harvest, the seeds were weighed, and the total yield was expressed in tons per hectare (t/ha) calculated to standard moisture of 14.0%. An electronic scale with a 0.001g accuracy level was utilized to measure the thousand-grain weight. Whole seed samples were weighed, and 500 seeds were randomly counted twice using the Contador seed counter (manufactured by Pfeuffer GmbH, Kitzingen, Germany). The two counts were then combined to obtain the total seed number. The test was repeated if the weight difference between the two samples exceeded 0.5g. The grain analyzer, Infratec™ 1241 (Foss, Sweden), was used to determine the crude protein content (%) which was measured on a dry weight basis.
SAS statistical software 9.4 (SAS Institute Inc., Cary, NC, USA) was used to analyze the data without any transformations. The Kolmogorov–Smirnov Test was used to check the data for homogeneity before analysis. Tukey’s test was applied at a significance level of p < 0.05 to examine the differences between each seed rate across different sowing periods as well as between different seed rates within each sowing time. Detailed ANOVA outputs (F-values, degrees of freedom, and p-values) for all measured variables are presented in Supplementary Tables S1-S5. Microsoft Excel 365 (Microsoft, Redmond, WA, USA) was used to create graphs that graphically represented the data.
3 Results and discussion
3.1 Damage caused by Bruchus rufimanus
This study showed that sowing time significantly influenced the percentage of grain damage caused by Bruchus rufimanus. Early sowing resulted in the highest damage across all study years, while late sowing exhibited the lowest (Figure 1). Additionally, insecticide-untreated plots exhibited substantially higher grain damage than treated plots. Although seeding rate had a minor influence, damage levels varied considerably between years. This variation was likely influenced by environmental factors, including the cropping density in nearby fields. Faba bean is often sown in Lithuanian agricultural systems between late April and early May (Romaneckas et al., 2018; Kimbirauskienė et al., 2023). Sowing is sometimes postponed until mid-May if unfavorable weather conditions make it difficult to prepare the seedbed. Late seeding delays blooming and pod setting, two crucial developmental phases that affect the timing of female B. rufimanus oviposition (Landry et al., 2015). Due to their mobility, the beetles move between fields to look for flowering plants (Kaniuczak, 2004). In faba bean, their oviposition activity is entirely restricted to pods (Gailis et al., 2022). The findings indicate that early seeding results in higher amounts of damage. That may be because early flowering and pod development occur at the same time as peak oviposition activity (Seidenglanz and Huňady, 2016; Huber et al., 2023). Conversely, late sowing significantly reduced grain damage, likely because pod development occurred after the pest’s primary oviposition period, thereby avoiding peak beetle activity and minimizing infestation. Bruchus rufimanus female’s sexual development correlates with an abundance of resources for flowers, which causes them to oviposit quickly on newly developed pods (Hamidi et al., 2021). Over 45 days may pass during the oviposition stage (Hamani-Aoudjit and Medjdoub-Bensaad, 2019).
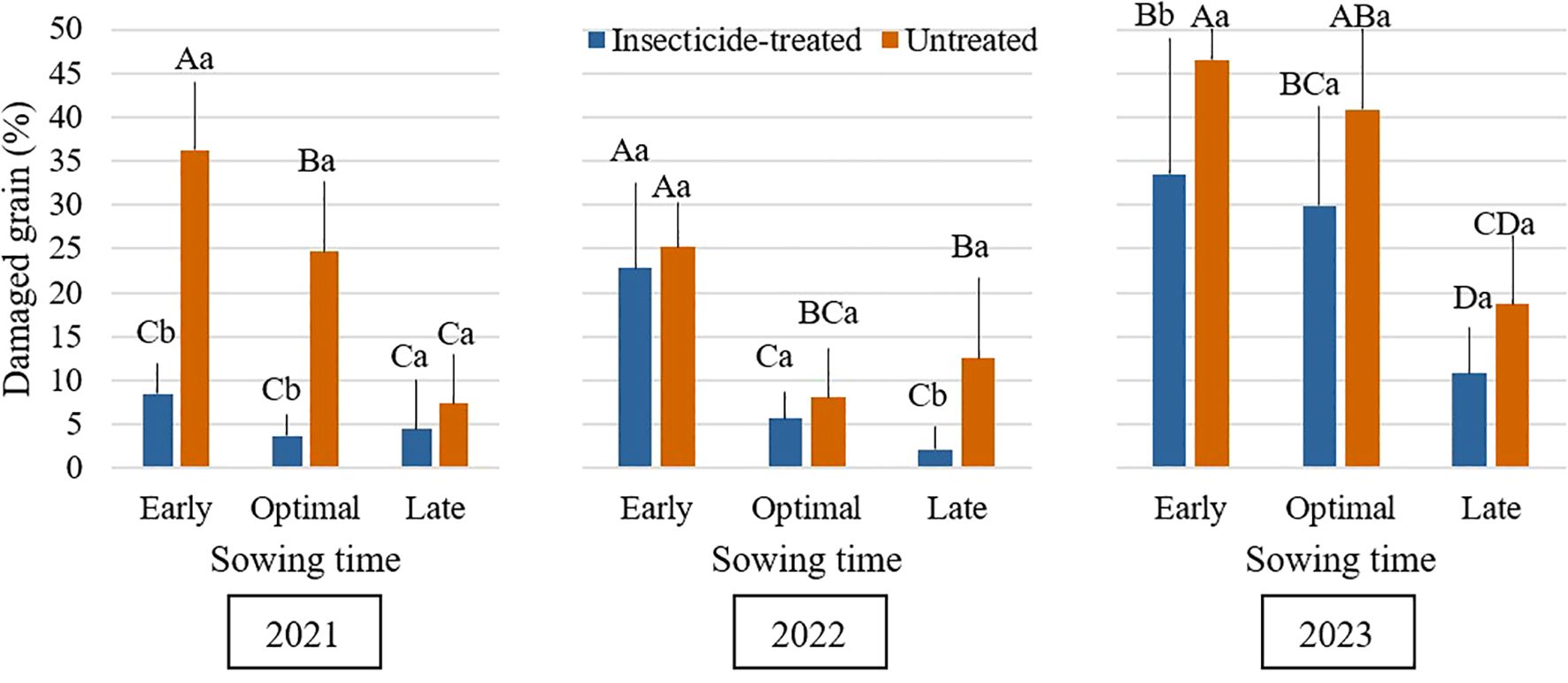
Figure 1. Mean percentage of Bruchus rufimanus-damaged grains across sowing times during the 2021–2023 growing seasons under insecticide-treated and untreated conditions. Different capital letters indicate significant differences of damaged grain at different sowing times under insecticide-treated or untreated treatment in the same year in 2021-2023 (Tukey test, P<0.05). Different lowercase letters indicate significant differences of damaged grain between the treatments of insecticide-treated and untreated under the same sowing time in the same year in 2021-2023 (Tukey test, P<0.05).
Throughout the study period, grain damage caused by B. rufimanus was consistently higher in untreated plots compared to insecticide-treated plots (Figure 2). This indicates the effectiveness of chemical control in reducing damage caused by this pest. Our results are in line with other studies that emphasize the significance of timely insecticide treatment for successful management (Segers et al., 2021). Overall, seeding rate had minimal impact on grain damage, which aligns with the absence of significant main effect in the statistical model. However, a significant difference was observed between high and low densities in untreated plots during the 2021 season. This finding is in line with research showing that by changing the microclimate conditions inside the crop canopy, higher seeding rate might affect pest dynamics (Mosedale et al., 2024). Microclimatic temperature variations, even over distances of a few meters, can significantly influence insect developmental rates and emergence timing (Greiser et al., 2022). However, in 2022 and 2023 there were no statistically significant differences in grain damage between the seeding rates in untreated plots. In the same way, there were no significant variations in grain damage between different seeding rates for treated plots throughout each of the three years. In 2021, damaged grain % was significantly higher in the untreated plots at every seeding rate. In contrast, in 2022 and 2023, despite the average grain damage % being higher in untreated plots than in treated plots, no statistically significant differences were noticed. Although these variations were not statistically significant, treated plots with the greatest seeding rate (80 plants/m2) showed the highest degrees of damage. While seeding rate had only a minor impact on damage in this study, evidence suggests that higher seeding rates could potentially increase damage due to greater flower density and associated volatile emissions. These chemicals are very attractive to B. rufimanus. This finding is consistent with that of Fezza et al. (2023), who reported that an increase in the density of baited monitoring tools led to a higher number of individuals captured. Higher plant densities in legumes can stress plants, causing them to release more volatile organic compounds (Midzi et al., 2022). These stronger scent signals can attract more herbivores (Makhlouf et al., 2024). On the other hand, denser canopies may also create less favorable microclimatic conditions for pest development, for example, higher humidity and lower temperatures, or may interfere with visual and olfactory host-finding cues. This competing hypothesis could help explain why, in our study, seeding rate exerted only a minimal overall influence on grain damage, despite the potential for contrasting mechanisms.
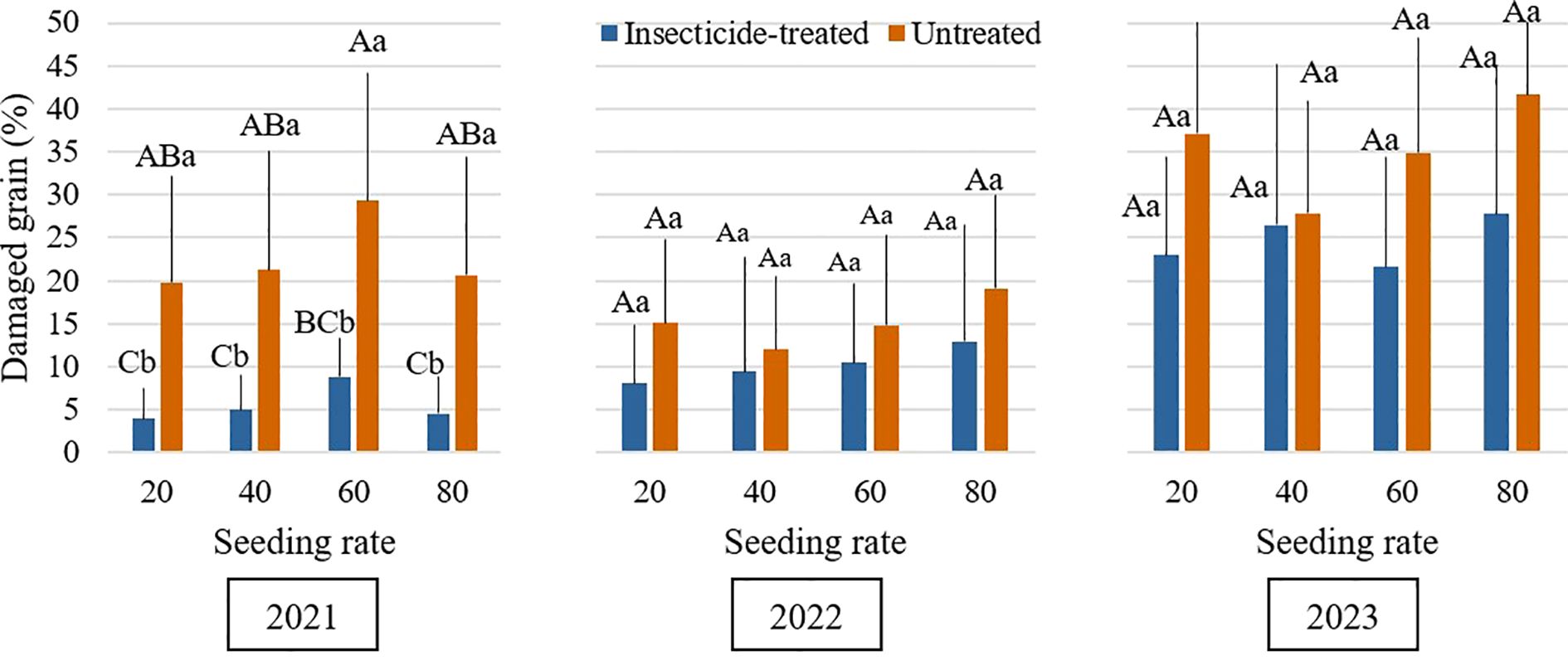
Figure 2. Mean percentage of Bruchus rufimanus-damaged grains across seeding rates during the 2021–2023 growing seasons under insecticide-treated and untreated conditions. Different capital letters indicate significant differences of damaged grain at different seeding rates under insecticide-treated or untreated treatment in the same year in 2021-2023 (Tukey test, P<0.05). Different lowercase letters indicate that the statistically significant differences of damaged grain between the treatments of insecticide-treated and untreated under the same seeding rate in the same year in 2021-2023 (Tukey test, P<0.05).
3.2 Faba bean grain yield
Figure 3 shows the effect of insecticide treatment (treated vs. untreated) at different sowing times on the yield of faba bean. According to an analysis of data covering three years, insecticide application did not significantly increase yield at the optimal sowing time. This is consistent with the results of (Koch et al., 2005), who found little variation in yield between insecticide-treated (either by foliar spray or seed treatment) and untreated plots. On the other hand, insecticide application significantly (P < 0.05) increased grain yield at both early and late sowing times. Our results showed that plots treated with insecticides consistently produced higher yields than untreated plots at both early and late sowing times every year. The yield benefit from insecticide application was most evident in early-sown plots, with increases ranging from 24.8% in 2021 to 126% in 2023, suggesting its compensatory role when phenological avoidance fails. The notably high yield increase of 126% in 2023 is likely attributable to the exceptionally low yield observed in the untreated early-sown control plots that year, which magnified the relative efficacy of the insecticide treatment. Importantly, 2023 was characterized by abiotic stress, with mean temperatures 0.6 °C above the long-term norm and precipitation 24% below average. Such drought conditions may have exacerbated pest injury by reducing plant vigor and compensatory growth capacity, while simultaneously increasing the plants’ susceptibility to herbivory. Consequently, the insecticide treatment likely provided a disproportionally higher protective effect under these stressed conditions. These findings are in line with earlier research that found that applying insecticides increased yields (Bardner et al., 1983). In the same direction, (Srinivasan et al., 2019) found that untreated control plots produced the lowest yard-long bean yields, highlighting the significance of role of insect management techniques in raising crop yield. The favorable growing season conditions in 2022 are probably the cause of the noticeably increased yield that was recorded. Air temperatures were near the long-term average, but precipitation was 49% higher than the typical climate norm, with July seeing especially heavy rainfall, which is when pod filling and seed development occur. These conditions probably helped to increase seed development, decrease water stress, and boost plant growth, which led to noticeably better yields than in 2021 and 2023 (Sehgal et al., 2018).
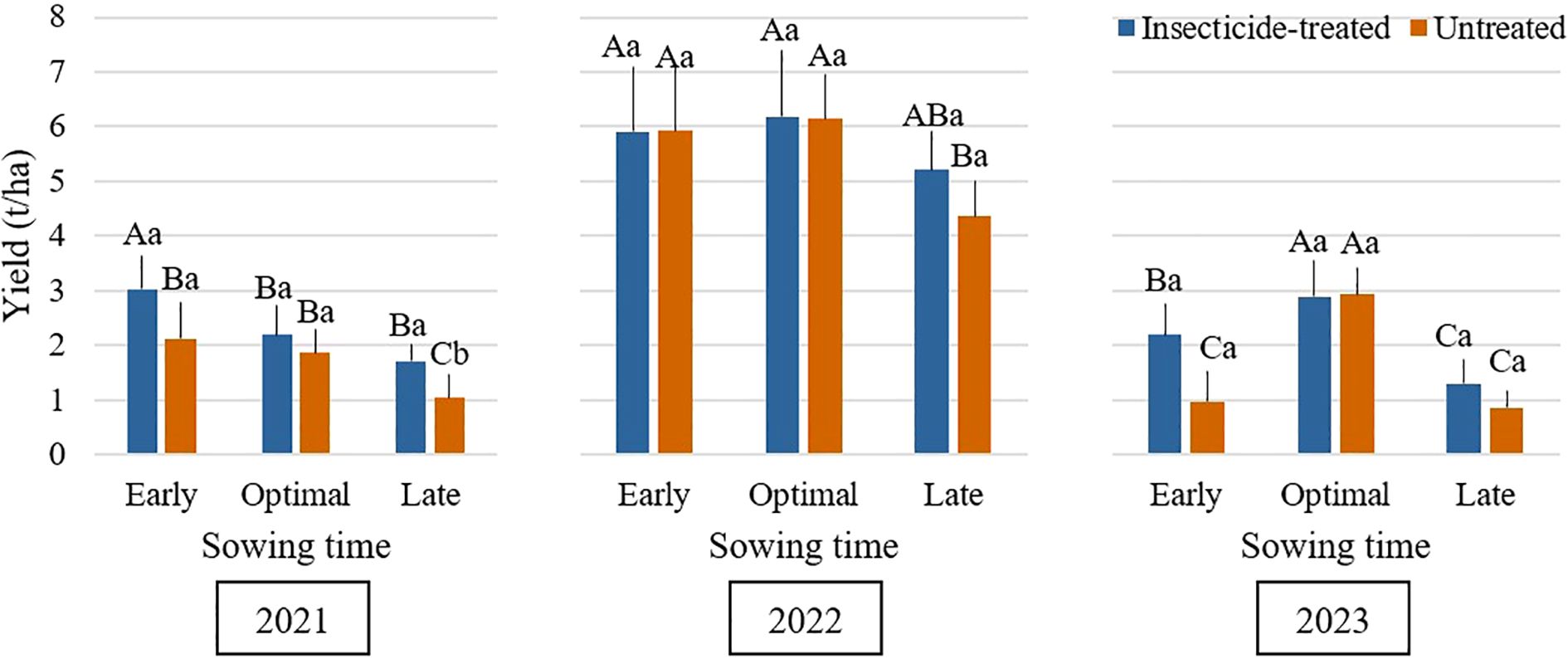
Figure 3. Influence of sowing time and insecticide application on grain yield (t/ha) of faba bean during the 2021–2023 growing seasons. Different capital letters indicate significant differences of yield at different sowing times under insecticide-treated or untreated treatment in the same year in 2021-2023 (Tukey test, P<0.05). Different lowercase letters indicate significant differences of yield between the treatments of insecticide-treated and untreated under the same sowing time in the same year in 2021-2023 (Tukey test, P<0.05).
Delayed sowing of faba bean may result in lower grain yields, as shown in Figure 3. The risk of yield reduction may be increased by potential harvesting delays, which can extend into October when the weather for crop and grain drying becomes not suitable. Greater seeding rates of 80 seeds per m² led to higher grain yields in untreated plots with optimal sowing time (Figures 4–6). Across all seeding rates, notably, in 2023, untreated plots with optimal sowing times produced more grain than those sown early or late. Grain yield from insecticide-treated and untreated plots, however, did not change much between the tested seeding rates at early and late sowing times in any of the research years. Plots that were sown at a density of 20 seeds per m², both insecticide-treated and untreated, produced the lowest grain yield. The high risk of most legumes to water scarcity, especially faba bean, is a popular challenge that is made worse by climate change (Balko et al., 2023). For faba bean to successfully develop, there must be enough soil moisture, which can be achieved by sufficient rainfall. Insufficient rainfall during crucial times like sowing time can be partly responsible for the general decline in yield for early or late sowing across the research years. (Aguilera-Diaz and Recalde-Manrique, 1995) emphasized the impact of environmental factors on crop performance by pointing out that yield is closely related to the mean temperature during flowering. One of the main variables affecting crop yield is plant density. According to studies by (Olle and Sooväli, 2020; Krob et al., 2022; Neugschwandtner et al., 2022; Ranazai et al., 2024), seeding rates have a major impact on disease severity, yield, and pest damage. Changing the seeding rate had only a minor impact on grain yield in our study. In contrast to these results, (Dahmardeh et al., 2010) found that increasing plant density from 12.5 to 20 plants per m² greatly increased yield. This variation draws attention to the complexity of yield responses to plant density, which are probably impacted by a combination of genetic features, crop management techniques, and environmental conditions.
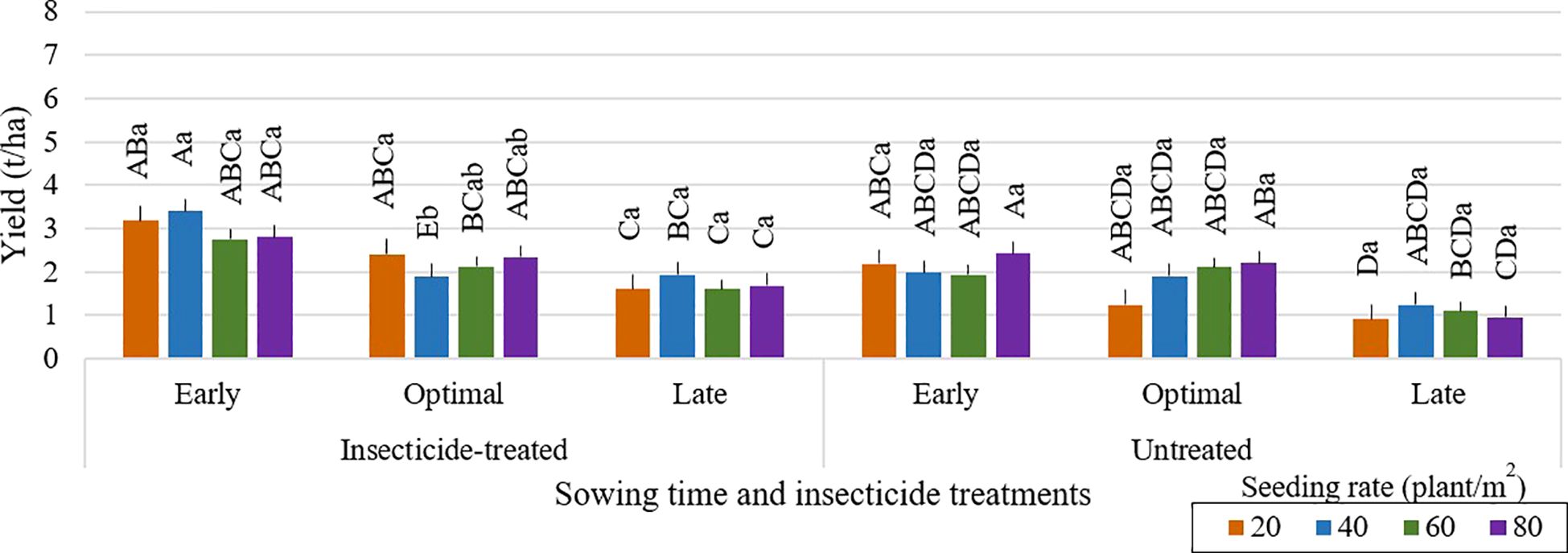
Figure 4. Faba bean grain yield (t/ha) across different sowing times and insecticide treatments at various seeding rates during the 2021 growing season. Different capital letters indicate significant differences of yield between different seeding rates under insecticide-treated or untreated treatment (Tukey test, P<0.05). Different lowercase letters indicate significant differences of yield between the different seeding rates under the same sowing time (Tukey test, P<0.05).
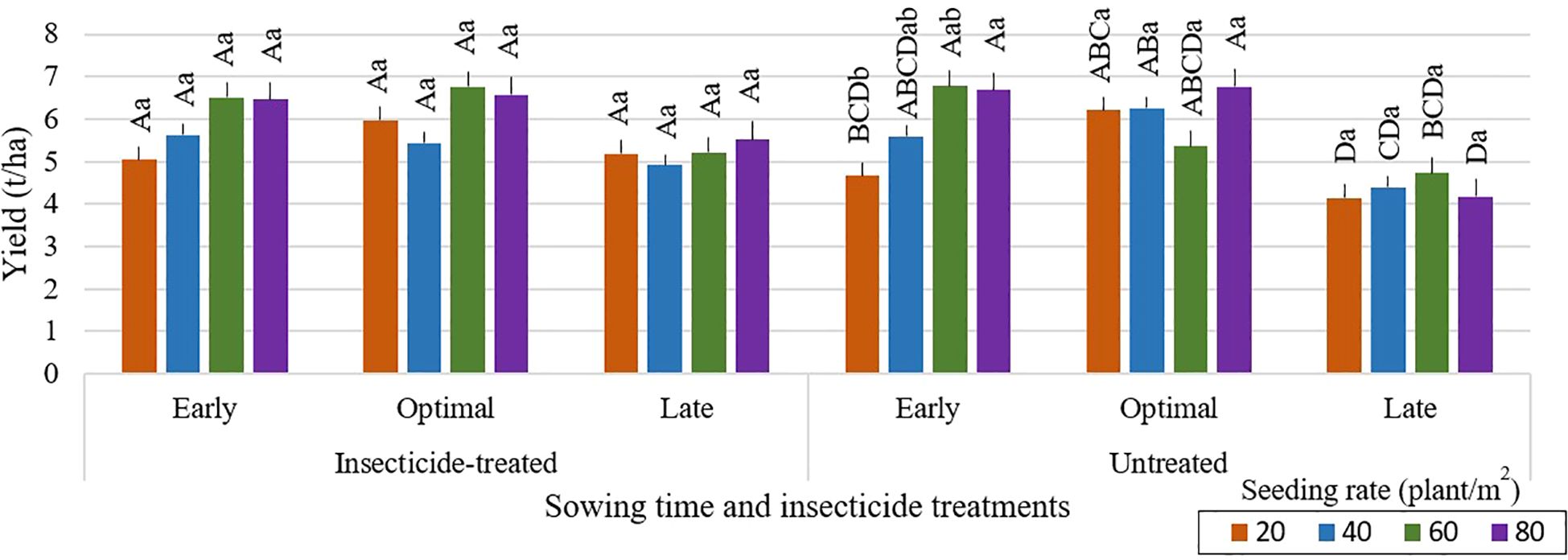
Figure 5. Faba bean grain yield (t/ha) across different sowing times and insecticide treatments at various seeding rates during the 2022 growing season. Different capital letters indicate significant differences of yield between different seeding rates under insecticide-treated or untreated treatment (Tukey test, P<0.05). Different lowercase letters indicate significant differences of yield between the different seeding rates under the same sowing time (Tukey test, P<0.05).
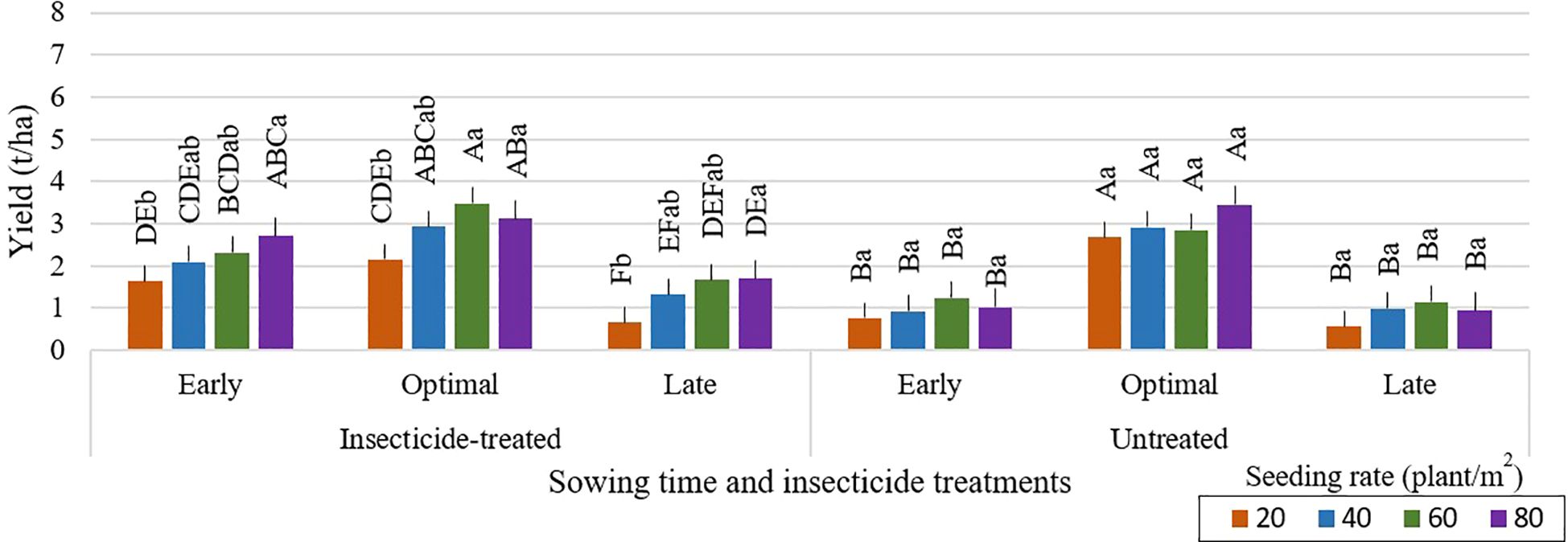
Figure 6. Faba bean grain yield (t/ha) across different sowing times and insecticide treatments at various seeding rates during the 2023 growing season. Different capital letters indicate significant differences of yield between different seeding rates under insecticide-treated or untreated treatment (Tukey test, P<0.05). Different lowercase letters indicate significant differences of yield between the different seeding rates under the same sowing time (Tukey test, P<0.05).
3.3 Crude protein content and thousand-grain weight
The impact of seeding rate and sowing time on crude protein (CP) content and thousand-grain weight (TGW) is presented in Tables 1 and 2. Results indicate that at the same sowing time, seeding rate generally did not have a significant effect on CP content or TGW. Contrary to our findings, (Anwar et al., 2015) reported that TGW significantly increased with higher seeding rates, while (Türk et al., 2011) reported a decreased in CP content under similar conditions. However, (Al-Rifaee et al., 2004) found conflicting results: in one trial year, there were no significant changes in TGW, whereas in another year, TGW was higher at lower seeding rates. Supporting our findings, (Plūduma-Pauniņa et al., 2021) reported that seeding rates had no significant effect on CP levels.
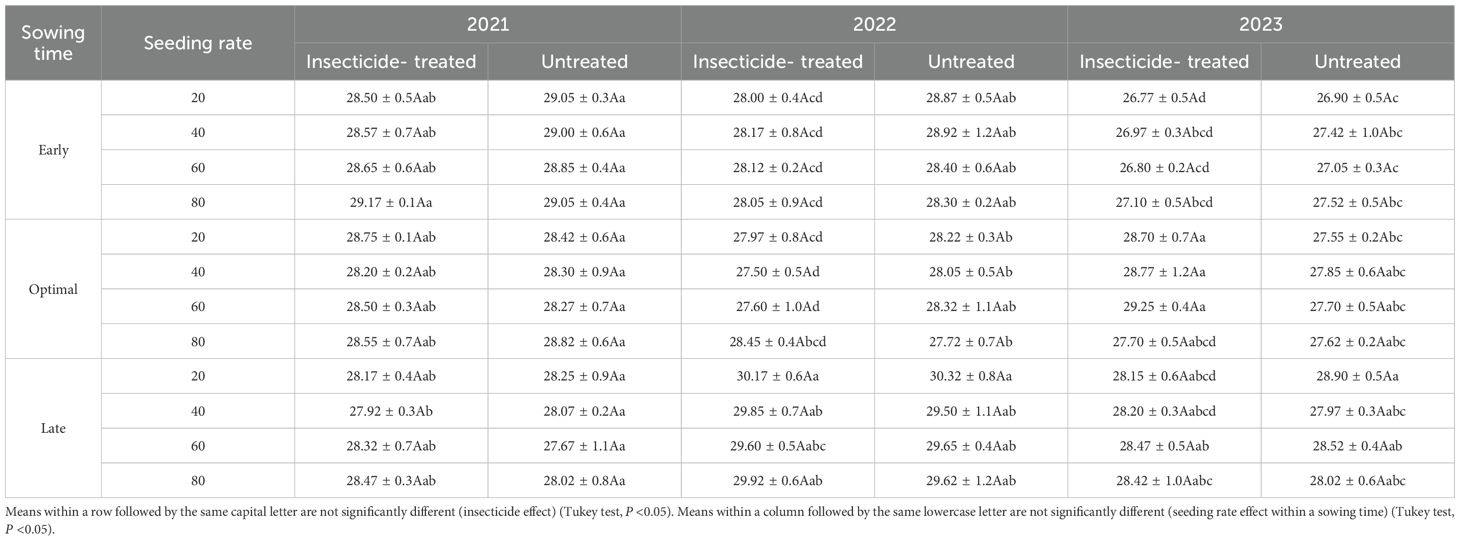
Table 1. Impact of seeding rate and sowing time on crude protein content (CP, %) of faba bean under the insecticide treated and untreated treatments in the same year in 2021-2023.
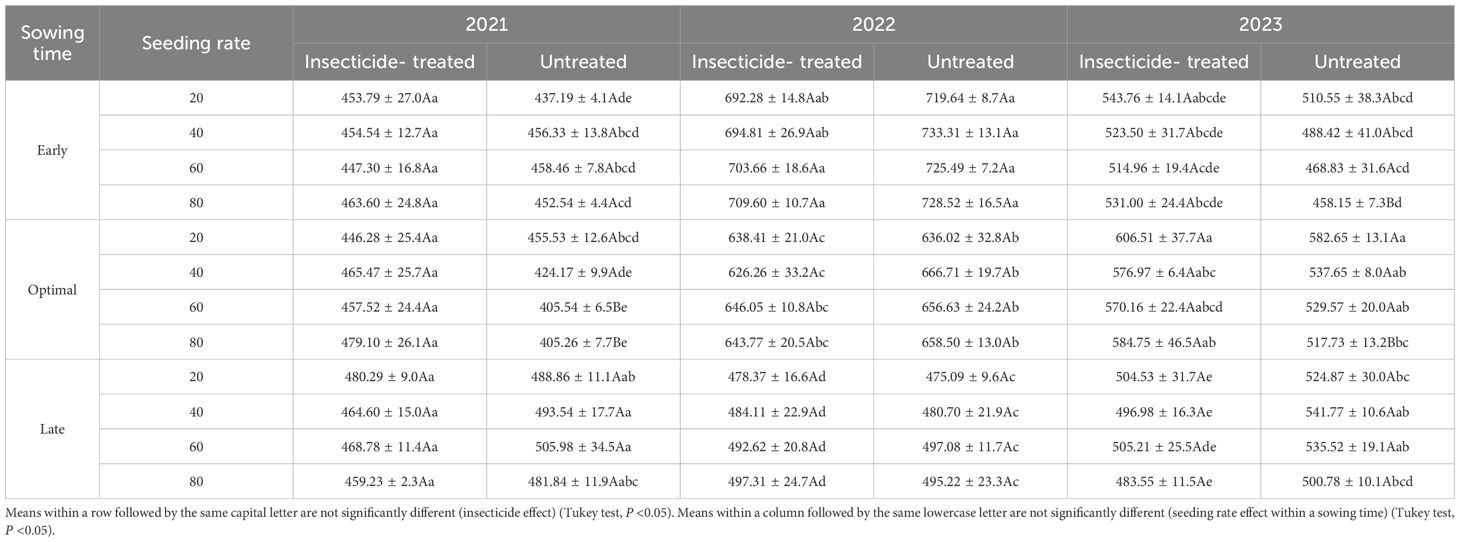
Table 2. Impact of seeding rate and sowing time on thousand-grain weight (TGW, g) of faba bean under the insecticide treated and untreated treatments in the same year in 2021-2023.
The most significant differences were noticed when comparing different sowing times. In 2021, untreated plots sown at the late sowing time had the highest TGW (492.5g on average) compared to early and optimal sowing time (Figure 7). In contrast, early sowing time treatments had the maximum TGW in 2022, with treated plots weighing 700.1g and untreated plots weighing 726.7g on average and it significantly decreased with each sowing time. In 2023, optimal sowing time resulted in the highest TGW compared to early and late sowing times, with treated plots averaging 584.6g and untreated plots averaging 541.9g. All three years demonstrated different results. There was no consistent pattern in TGW when evaluating insecticide-treated and untreated plots at different sowing times. (Anwar et al., 2015) also have noticed differences in TGW between different sowing times – TGW decreased with later sowing times as compared to earlier ones.
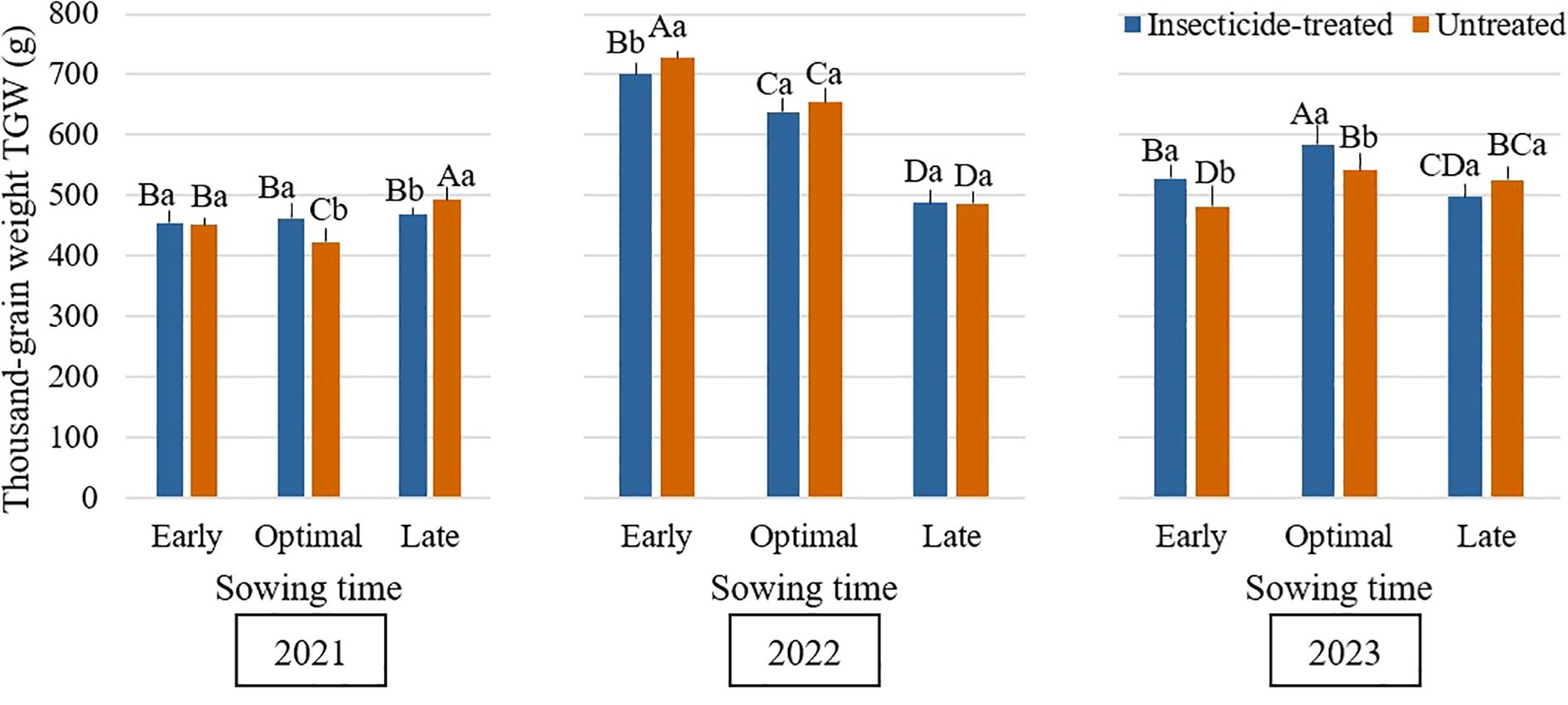
Figure 7. Influence of sowing time and insecticide application on thousand-grain weight (TGW) (g) of faba bean during the 2021–2023 growing seasons. Different capital letters indicate significant differences of thousand-grain weight (TGW) at different sowing times under insecticide-treated or untreated treatment in the same year in 2021-2023 (Tukey test, P<0.05). Different lowercase letters indicate significant differences of thousand-grain weight (TGW) between the treatments of insecticide-treated and untreated under the same sowing time in the same year in 2021-2023 (Tukey test, P<0.05).
The experimental years had an impact on the CP content of faba bean grains, although the variation was still within a small range of 2%. The sowing time impact on CP content appears to vary across years (Figure 8). In 2021, grain from early sown plots had a much higher CP percentage, but late sowing yielded the greatest CP content in 2022 and 2023. This suggests that the impact of sowing time on CP content varies each year. (Księżak and Bojarszczuk, 2022) reported that early sowing tends to result in lower CP content in soybean. Throughout all of the experiment years, the use of insecticides had no discernible effect on the amount of CP. The combination between the experimental factors and interannual weather variability likely explains the minor fluctuations in CP content observed across the study years. The results underscore that environmental conditions play a more critical role in determining faba bean grain CP content than agronomic management factors such as seeding rate and insecticide application.
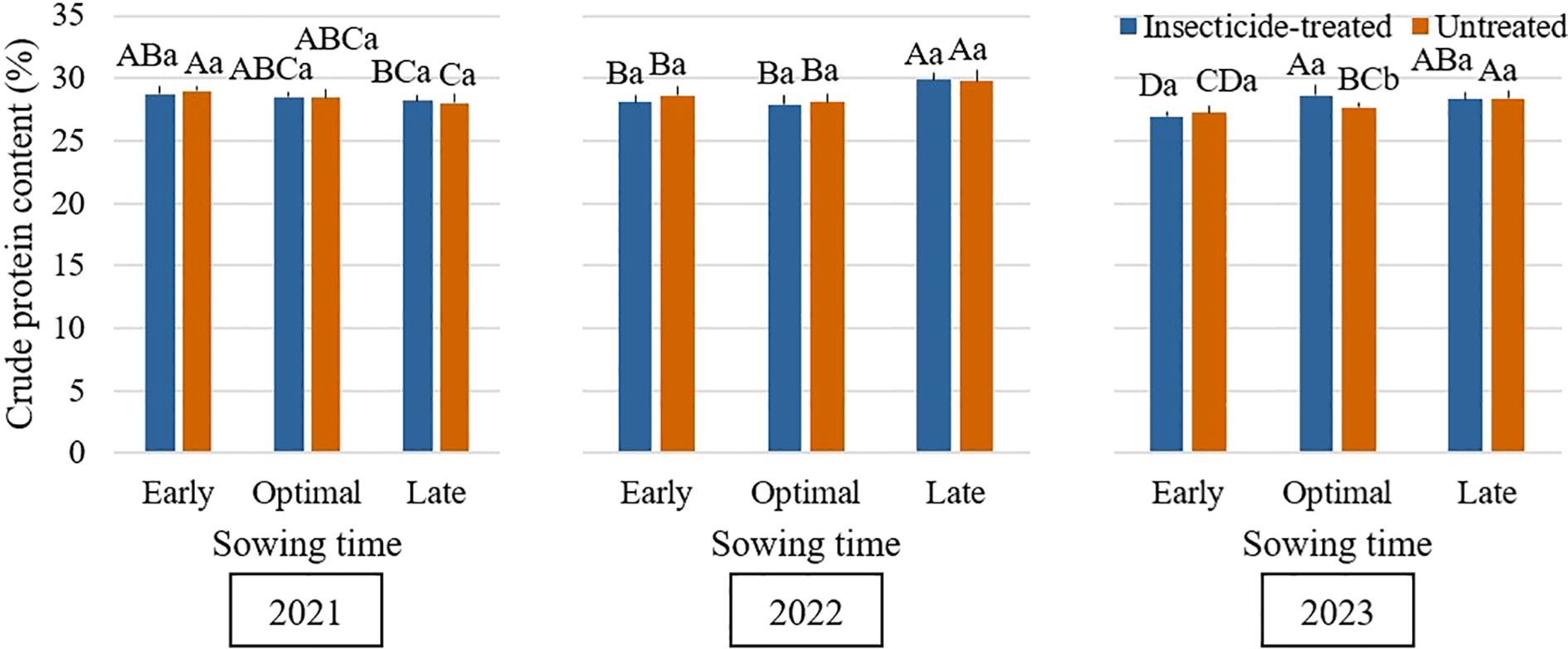
Figure 8. Influence of sowing time and insecticide application on crude protein (CP) content (%) of faba bean during the 2021–2023 growing seasons. Different capital letters indicate significant differences of crude protein (CP) content at different sowing times under insecticide-treated or untreated treatment in the same year in 2021-2023 (Tukey test, P<0.05). Different lowercase letters indicate significant differences of crude protein (CP) content between the treatments of insecticide-treated and untreated under the same sowing time in the same year in 2021-2023 (Tukey test, P<0.05).
4 Conclusions
This study demonstrates the impacts of seeding rate, insecticide use, and sowing time on faba bean yield and grain damage in Lithuanian conditions. In conclusion, late April is identified as the optimal sowing period for faba bean in Lithuania. Late sowing resulted in reduced grain yield but also less grain damage caused by Bruchus rufimanus. While early sowing carries a higher risk of damage, its yield potential can be high under favorable conditions, especially when protected with insecticides. Under unfavorable conditions, the yield penalty for late sowing may be more severe. The yield of faba bean was affected by the timing of sowing. However, during the research years, increasing the seeding rate beyond 60 seeds m-2 did not consistently increase yield. Overall, seeding rate exerted minimal influence on yield and pest damage. However, in 2021, significant variation in grain damage was observed among seeding rates in untreated plots, suggesting that under certain pest pressure conditions, seeding rate may play a role. In order to maximize yield and minimize grain damage in faba bean farming systems, our data highlight the need to combine the best possible sowing time, insect control strategies, and environmental variable adaptation. We recommend that Lithuanian farmers adopt a dual strategy: (1) prioritize sowing in late April where feasible; and (2) apply cypermethrin only in early-sown fields where damage risk exceeds 30%. Further studies are needed on the long-term impacts of integrated pest management techniques on seed quality and yield stability in different climates. Furthermore, examining how different faba bean cultivars, insecticide effectiveness, and sowing time interact could improve region-specific recommendations.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
MoA: Conceptualization, Formal Analysis, Investigation, Methodology, Writing – original draft. RS: Supervision, Writing – review & editing. AJ: Writing – review & editing. JR: Writing – review & editing. EV: Formal Analysis, Writing – review & editing. AS: Writing – review & editing. KL: Writing – review & editing. YA: Writing – review & editing. MaA: Formal Analysis, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. Part of this research was supported by the long-term research program “Harmful organisms in agro and forest ecosystems” implemented by the Lithuanian Research Centre for Agriculture and Forestry.
Acknowledgments
The authors express their gratitude to the technical team of the Department of Plant Pathology and Protection at the Institute of Agriculture, Lithuanian Research Centre for Agriculture and Forestry, for their valuable contributions to this research.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fagro.2025.1631790/full#supplementary-material
References
Abbas M., Saleem M., Hussain D., Ramzan M., Jawad Saleem M., Abbas S., et al. (2022). Review on integrated disease and pest management of field crops. Int. J. Trop. Insect Sci. 42, 3235–3243. doi: 10.1007/S42690-022-00872-W/TBLES/2
Aguilera-Diaz C. and Recalde-Manrique L. (1995). Effects of plant density and inorganic nitrogen fertilizer on field beans (Vicia faba). J. Agric. Sci. 125, 87–93. doi: 10.1017/S0021859600074530
Ahmad M. F., Ahmad F. A., Alsayegh A. A., Zeyaullah M., AlShahrani A. M., Muzammil K., et al. (2024). Pesticides impacts on human health and the environment with their mechanisms of action and possible countermeasures. Heliyon 10, e29128. doi: 10.1016/J.HELIYON.2024.E29128
Al-Rifaee M., Turk M., and Tawaha A. M. (2004). Effect of seed size and plant population density on yield and yield components of local faba bean (Vicia faba L. Major). Int. J. Agric. Biol. 6, 294–299.
Anwar S., Ali W., Inamullah K., Islam M., Bashir S., Shafi M., et al. (2015). Effect of sowing dates and seed rates on the agro-physiological traits of wheat. J. Environ. Earth Sci. 5, 135–141.
Balko C., Torres A. M., and Gutierrez N. (2023). Variability in drought stress response in a panel of 100 faba bean genotypes. Front. Plant Sci. 14. doi: 10.3389/FPLS.2023.1236147/BIBTEX
Bardner R., Fletcher K. E., and Griffiths D. C. (1983). Chemical control of the pea and bean weevil, Sitona lineatus L., and subsequent effects on the yield of field beans Vicia faba L. J. Agric. Sci. 101, 71–80. doi: 10.1017/S0021859600036388
Dahmardeh M., Ramroodi M., and Valizadeh J. (2010). Effect of plant density and cultivars on growth, yield and yield components of faba bean (Vicia faba L.). Afr J. Biotechnol. 9, 8643–8647.
Dell’Aglio D. D. and Tayeh N. (2023). Responsiveness of the broad bean weevil, Bruchus rufimanus, to Vicia faba genotypes. Entomol Exp. Appl. 171, 312–322. doi: 10.1111/EEA.13277
Elhamalawy O., Bakr A., and Eissa F. (2024). Impact of pesticides on non-target invertebrates in agricultural ecosystems. Pestic Biochem. Physiol. 202, 105974. doi: 10.1016/J.PESTBP.2024.105974
Elsayed E., Salman A., and Abdel-Rahman M. (2021). Incidence of certain insect pests infesting faba bean Vicia faba cultivars under some agriculture practices. Egypt J. Plant Prot Res. Inst 4, 440–444.
Esan V. I., Olaniyi O. C., Ogunbode T. O., Ayegboyin M. H., Sangoyomi T. E., and Akande J. (2023). Growth and nutritional potential of selected grasses and legumes for ruminants’ production. IOP Conf Ser. Earth Environ. Sci. 1219, 12011. doi: 10.1088/1755-1315/1219/1/012011
Fan K., Yip H. Y., Taddi S., Huang Y. Y., Wong F. L., Hui J. H. L., et al. (2022). Pursuing greener farming by clarifying legume-insect pest interactions and developing marker-assisted molecular breeding. Adv. Bot. Res. 102, 211–258. doi: 10.1016/BS.ABR.2022.02.018
Fezza E., Roberts J. M., Bruce T. J. A., Walsh L. E., Gaffney M. T., and Pope T. W. (2023). Decoding attraction: Improving vine weevil monitoring by exploiting key sensory cues. Pest Manag Sci. 79, 4635–4643. doi: 10.1002/PS.7665
Gailis J., Astašova N., Jākobsone E., and Ozoliņa-Pole L. (2022). Biology of broadbean seed beetle (Bruchus rufimanus; Coleoptera: Chrysomelidae) in Latvia. Acta Agric. Scand. B Soil Plant Sci. 72, 4–16. doi: 10.1080/09064710.2021.1977841
Gao Y., Zhang M., Yao C., Liu Y., Wang Z., and Zhang Y. (2021). Increasing seeding density under limited irrigation improves crop yield and water productivity of winter wheat by constructing a reasonable population architecture. Agric. Water Manag 253, 106951. doi: 10.1016/J.AGWAT.2021.106951
Greiser C., von Schmalensee L., Lindestad O., Gotthard K., and Lehmann P. (2022). Microclimatic variation affects developmental phenology, synchrony and voltinism in an insect population. Funct. Ecol. 36, 3036–3048. doi: 10.1111/1365-2435.14195
Gunstone T., Cornelisse T., Klein K., Dubey A., and Donley N. (2021). Pesticides and soil invertebrates: a hazard assessment. Front. Environ. Sci. 9. doi: 10.3389/FENVS.2021.643847/BIBTEX
Hamani-Aoudjit S. and Medjdoub-Bensaad F. (2019). Monitoring the activity and biology of pest Bruchus rufimanus (Coleoptera: Chrysomelidae) in beans crops. Indian J. Agric. Res. 53, 632–635. doi: 10.18805/A-408
Hamidi R., Taupin P., and Frérot B. (2021). Physiological synchrony of the broad bean weevil, Bruchus rufimanus Boh., to the host plant phenology, Vicia faba L. Front. Insect Sci. 1. doi: 10.3389/FINSC.2021.707323/BIBTEX
Huber J., Chaluppa N., Voit B., Steinkellner S., and Killermann B. (2023). Damage potential of the broad bean beetle (Bruchus rufimanus Boh.) on seed quality and yield of faba beans (Vicia faba L.). Crop Prot 168, 106227. doi: 10.1016/J.CROPRO.2023.106227
Hussien W. A., Elzoghby I. R., Fahmy A. M., and Ahmed M. A. (2022). Response of certain faba bean cultivars to infestation by Aphis craccivora (Hemiptera: Aphididae) in relation to the expected yield and its components. AUJES 3, 249–266. doi: 10.21608/AUJES.2022.131409.1064
Inskeep J. R., Allen A. P., Taylor P. W., Rempoulakis P., and Weldon C. W. (2021). Canopy distribution and microclimate preferences of sterile and wild Queensland fruit flies. Sci. Rep. 11, 1–14. doi: 10.1038/s41598-021-92218-8
Kaniuczak Z. (2004). Seed damage of field bean (Vicia faba L. Var. Minor Harz.) caused by bean weevils (Bruchus rufimanus Boh.) (Coleoptera: Bruchidae). J. Plant Prot Res. 44, 125–130.
Kaur H., Difonzo C., Cassida K. A., Chilvers M. I., and Singh M. P. (2024). Planting time and seeding rate impact insect feeding, ear rots, and forage nutritive value in silage corn. Agron. J. 116, 1978–1990. doi: 10.1002/AGJ2.21620
Khelfane-Goucem K. and Medjdoub-Bensaad F. (2016). Impact of Bruchus rufimanus infestation upon broad bean seeds germination. Adv. Environ. Biol. 10, 144–152.
Kimbirauskienė R., Sinkevičienė A., Jonaitis R., and Romaneckas K. (2023). Impact of tillage intensity on the development of faba bean cultivation. Sustainability 15, 8956. doi: 10.3390/SU15118956
Koch R. L., Burkness E. C., Hutchison W. D., and Rabaey T. L. (2005). Efficacy of systemic insecticide seed treatments for protection of early-growth-stage snap beans from bean leaf beetle (Coleoptera: Chrysomelidae) foliar feeding. Crop Prot 24, 734–742. doi: 10.1016/J.CROPRO.2004.12.010
Krob J. L., Stewart S. D., and Brown S. A. (2022). Effects of plant density, seed spacing, and seed treatment on thrips injury to cotton. Crop Prot 161, 106059. doi: 10.1016/J.CROPRO.2022.106059
Księżak J. and Bojarszczuk J. (2022). The seed yield of soybean cultivars and their quantity depending on sowing term. Agronomy 12, 1066. doi: 10.3390/AGRONOMY12051066
Landry E. J., Coyne C. J., and Hu J. (2015). Agronomic performance of spring-sown faba bean in southeastern Washington. Agron. J. 107, 574–578. doi: 10.2134/AGRONJ14.0284
Lukošiūtė-Stasiukonienė A., Almogdad M., Semaškienė R., and Mačiulytė V. (2024). Crop density and sowing timing effect on tan spot occurrence in spring wheat. Agriculture 14, 1284. doi: 10.3390/agriculture14081284
Makhlouf L., El Fakhouri K., Kemal S. A., Maafa I., Meftah Kadmiri I., and El Bouhssini M. (2024). Potential of volatile organic compounds in the management of insect pests and diseases of food legumes: a comprehensive review. Front. Plant Sci. 15. doi: 10.3389/fpls.2024.1430863
Manning B. K., Adhikari K. N., and Trethowan R. (2020). Impact of sowing time, genotype, environment and maturity on biomass and yield components in faba bean (Vicia faba). Crop Pasture Sci. 71, 147–154. doi: 10.1071/CP19214
Mbaidiro J. T., Onzo A., Djenaissem A., and Mbaikoubou M. (2023). Influence of sowing dates on the population density of the fall armyworm Spodoptera frugiperda (JE. Smith) and its damage on maize plants in Chad. Int. J. Biol. Chem. Sci. 17, 773–786. doi: 10.4314/IJBCS.V17I3.3
Medjdoub-Bensaad F., Khelil M. A., and Huignard J. (2007). Bioecology of broad bean bruchid Bruchus rufimanus Boh. (Coleoptera: Bruchidae) in a region of Kabylia in Algeria. Afr J. Agric. Res. 2, 412–417.
Midzi J., Jeffery D. W., Baumann U., Rogiers S., Tyerman S. D., and Pagay V. (2022). Stress-induced volatile emissions and signalling in inter-plant communication. Plants 11, 2566. doi: 10.3390/plants11192566
Mosedale J. R., Eyre D., Korycinska A., Everatt M., Grant S., Trew B., et al. (2024). Mechanistic microclimate models and plant pest risk modelling. J. Pest Sci. 97, 1749–1766. doi: 10.1007/S10340-024-01777-Y
Münzbergová Z. (2012). Seed density significantly affects species richness and composition in experimental plant communities. PLoS One 7, e46704. doi: 10.1371/JOURNAL.PONE.0046704
Murphy L. C., Sparkes D. L., Spink J. H., and Alves S. (2022). Yield response of field beans (Vicia faba) to plant population and sowing date in a temperate climate. Irish J. Agr Food Res. 61, 384–396. doi: 10.15212/IJAFR-2022-0024
Neugschwandtner R. W., Bernhuber A., Kammlander S., Wagentristl H., Klimek-Kopyra A., Bernas J., et al. (2022). Effect of two seeding rates on yield and yield components of winter and spring faba bean. Acta Agric. Scand. B Soil Plant Sci. 72, 496–505. doi: 10.1080/09064710.2021.2014552
Olle M. and Sooväli P. (2020). The severity of diseases of faba bean depending on sowing rate and variety. Acta Agric. Scand. B Soil Plant Sci. 70, 572–577. doi: 10.1080/09064710.2020.1814402
Pathak V. M., Verma V. K., Rawat B. S., Kaur B., Babu N., Sharma A., et al. (2022). Current status of pesticide effects on environment, human health and it’s eco-friendly management as bioremediation: a comprehensive review. Front. Microbiol. 13. doi: 10.3389/FMICB.2022.962619
Plūduma-Pauniņa I., Gaile Z., and Bimšteine G. (2021). Sowing time effect on yield and quality of field beans in a changing meteorological situation in the Baltic region. Agron. Res. 19, 1873–1887. doi: 10.15159/AR.21.112
Ranazai S. K., Sadiq M., Baloch M. S., Qureshi H., Anwar T., Alarfaj A. A., et al. (2024). Impact of different priming and sowing techniques in combination with different seed rates on wheat growth and yield. Sci. Rep. 14, 1–10. doi: 10.1038/s41598-024-78256-y
Romaneckas K., Kimbirauskiene R., Adamaviciene A., Jasinskas A., and Sarauskis E. (2018). Impact of soil tillage intensity on faba bean cultivation. Eng. Rural Dev. 17, 34–38. doi: 10.22616/ERDev2018.17.N034
Schneider K., Barreiro-Hurle J., and Rodriguez-Cerezo E. (2023). Pesticide reduction amidst food and feed security concerns in Europe. Nat. Food 4, 746–750. doi: 10.1038/s43016-023-00834-6
Segers A., Caparros Megido R., Lognay G., and Francis F. (2021). Overview of Bruchus rufimanus Boheman 1833 (Coleoptera: Chrysomelidae): biology, chemical ecology and semiochemical opportunities in integrated pest management programs. Crop Prot 140, 105411. doi: 10.1016/J.CROPRO.2020.105411
Sehgal A., Sita K., Siddique K. H. M., Kumar R., Bhogireddy S., Varshney R. K., et al. (2018). Drought or/and heat-stress effects on seed filling in food crops: impacts on functional biochemistry, seed yields, and nutritional quality. Front. Plant Sci. 9. doi: 10.3389/fpls.2018.01705
Seidenglanz M. and Huňady I. (2016). Effects of faba bean (Vicia faba) varieties on the development of Bruchus rufimanus. Czech J. Genet. Plant Breed 52, 22–29. doi: 10.17221/122/2015-CJGPB
Shahid M. R., Mahmood A., Farooq J., Tasdeeq M., Shahid H., Asif M., et al. (2014). The impact of sowing dates and varieties on the incidence of Oxycarenus laetus and Dysdercus koingii on cotton. Com Sci. 5, 412–418.
Siddiqui J. A., Fan R., Naz H., Bamisile B. S., Hafeez M., Ghani M. I., et al. (2023). Insights into insecticide-resistance mechanisms in invasive species: Challenges and control strategies. Front. Physiol. 13. doi: 10.3389/FPHYS.2022.1112278
Srinivasan R., Paola S., Lin M.y., Chhun H. H., Sareth K., and Sor S. (2019). Development and validation of an integrated pest management strategy for the control of major insect pests on yard-long bean in Cambodia. Crop Prot 116, 82–91. doi: 10.1016/J.CROPRO.2018.10.015
Szafirowska A. (2012). The role of cultivars and sowing date in control of broad bean weevil (Bruchus rufimanus Boh.) in organic cultivation. Veg Crop Res. Bull. 77, 29–36. doi: 10.2478/V10032-012-0013-2
Tang F. H. M., Lenzen M., McBratney A., and Maggi F. (2021). Risk of pesticide pollution at the global scale. Nat. Geosci 14, 206–210. doi: 10.1038/s41561-021-00712-5
Titouhi F., Amri M., and Mediouni-Ben Jemâa J. (2015). Status of coleopteran insects infesting faba bean in Tunisia with emphasis on population dynamics and damage of Bruchus rufimanus (Chrysomylidae). Basic Res. J. Agric. Sci. Rev. 4, 225–233.
Török E., Zieger S., Rosenthal J., Földesi R., Gallé R., Tscharntke T., et al. (2021). Organic farming supports lower pest infestation, but fewer natural enemies than flower strips. J. Appl. Ecol. 58, 2277–2286. doi: 10.1111/1365-2664.13946
Tran B., Darquenne J., and Huignard J. (1993). Changes in responsiveness to factors inducing diapause termination in Bruchus rufimanus (Boh.) (Coleoptera: Bruchidae). J. Insect Physiol. 39, 769–774. doi: 10.1016/0022-1910(93)90052-S
Türk M., Albayrak S., and Yüksel O. (2011). Effect of seeding rate on the forage yields and quality in pea cultivars of differing leaf types. Turk J. Field Crops 16, 137–141.
Xu C., Li R., Song W., Wu T., Sun S., Han T., et al. (2021). High density and uniform plant distribution improve soybean yield by regulating population uniformity and canopy light interception. Agronomy 11, 1880. doi: 10.3390/AGRONOMY11091880/S1
Keywords: Bruchus rufimanus, bruchid beetle, density, grain quality, harvest, leguminous, protein, TGW
Citation: Almogdad M, Semaškienė R, Jonavičienė A, Ramanauskienė J, Venslovas E, Sabeckis A, Lavrukaitė K, Alkuddsi Y and Almukdad M (2025) Optimized sowing time combined with targeted insecticide application enhances faba bean (Vicia faba L.) yield by suppressing Bruchus rufimanus damage in Lithuania. Front. Agron. 7:1631790. doi: 10.3389/fagro.2025.1631790
Received: 20 May 2025; Accepted: 09 October 2025;
Published: 22 October 2025.
Edited by:
Wenwu Zhou, Zhejiang University, ChinaReviewed by:
Yongjian Xie, Zhejiang A&F University, ChinaLi-Lin Chen, Fujian Agriculture and Forestry University, China
Copyright © 2025 Almogdad, Semaškienė, Jonavičienė, Ramanauskienė, Venslovas, Sabeckis, Lavrukaitė, Alkuddsi and Almukdad. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mohammad Almogdad, bW9oYW1tYWQuYWxtb2dkYWRAbGFtbWMubHQ=