 Sofía A. Sola1
Sofía A. Sola1 Margarita Rocca1*
Margarita Rocca1* Nadia G. Salas Gervassio1,2*
Nadia G. Salas Gervassio1,2* Julieta N. Peñalba1Rocío I. Montiel Cáceres1
Julieta N. Peñalba1Rocío I. Montiel Cáceres1 M. Gabriela Luna1,2
M. Gabriela Luna1,2- 1Centro de Estudios Parasitológicos y de Vectores (CEPAVE), CONICET-UNLP, CICPBA, La Plata, Argentina
- 2Departamento de Ciencias Exactas y Naturales, Universidad Nacional de San Antonio de Areco- UNSAdA, San Antonio de Areco, Argentina
Biological control is a key component of Integrated Pest Management (IPM) programs aimed at meeting yield and environmental goals of agriculture. In the search for effective natural enemies against the globally invasive tomato pest Phthorimaea absoluta, we evaluated the biological traits of the Neotropical mirid Campyloneuropsis cincticornis. Through laboratory bioassays, we assessed: (1) predation rates of different developmental stages of C. cincticornis on P. absoluta eggs and larvae; (2) oviposition behavior on two host plants—Solanum sisymbriifolium (sticky nightshade) and S. lycopersicum (tomato), and (3) development time on both plants. Older nymphs and adult females consumed more P. absoluta eggs and all stages (except first-instar nymphs) were able to feed on larvae, including those concealed inside leaf mines. Oviposition was markedly higher on S. sisymbriifolium, suggesting a strong preference likely shaped by offspring performance. C. cincticornis egg development time did not differ between host plants, whereas nymphal development was significantly faster on S. sisymbriifolium. These findings position C. cincticornis as a promising candidate for both augmentative and conservation biological control of P. absoluta, particularly in systems where native solanaceous vegetation is present.
Introduction
Modern agriculture requires knowledge and technologies aligned with economic and environmental objectives for sustainable food production (Robertson and Swinton, 2005). Particularly for crop protection, biological control is an important component of Integrated Pest Management (IPM) against key agricultural pests. The implementation of biological control strategies requires fundamental understanding of the biology, ecology, and behavior of a set of natural enemies of the pest. This knowledge is essential to make decisions regarding their use and/or conservation aiming to reduce pest populations to economically tolerable levels (Bueno, 2009; Jacas and Urbaneja, 2008; van Driesche et al., 2008).
The study on predatory mirid bugs as biocontrol agents has increased in the last decades since they have shown effectiveness on controlling horticultural insect pests (van Lenteren et al., 2018; Snyder, 2019). As polyphagous predators, they contribute to ecosystem stability by preying on a wide range of pest species such as aphids, whiteflies, lepidopteran eggs and larvae (Bueno et al., 2012; Calvo et al., 2012; Mollá et al., 2014; López et al., 2019; Pérez-Hedo et al., 2021; Burla et al., 2022; Montiel Cáceres et al., 2024). Globally, around 150 commercial biocontrol agents are currently registered, with mirid bugs (Hemiptera: Heteroptera) ranking among the most widely used. Approximately 20 mirid species are commercially available, with others under evaluation, primarily for use in greenhouse IPM programs (van Lenteren, 2012; van Lenteren et al., 2018, 2020).
Tomato production in Argentina is a significant agricultural activity, with an average cultivated area of approximately 17,000 hectares and an annual yield of around 1 million tons, where both open-field and greenhouse cultivation are practiced. Approximately 70% of the production is destined for fresh consumption, while the remaining 30% is processed for industrial use. This sector contributes to the national economy, creates rural jobs, and aids regional development (Secretaría de Agricultura, Ganadería y Pesca, Argentina, 2023). The South American tomato pinworm, Phthorimaea absoluta (Meyrick) (Lepidoptera: Gelechiidae) is the most relevant pest of tomato production worldwide. Females oviposit on leaves, and after hatching, larvae mine into plant tissues, causing severe damage and can reach up to 100% yield loss. Notably, larvae can exit old mines to construct new ones, temporarily exposing themselves and creating an opportunity for predation (Desneux et al., 2010). To achieve P. absoluta control, multiple IPM techniques including biological control are required (Desneux et al., 2022). Among biocontrol agents, zoophytophagous mirid bugs are commonly used against this pest. In Europe, Macrolophus pygmaeus Wagner and Nesidiocoris tenuis (Reuter) are widely used to suppress lepidopteran egg populations (Belda et al., 2010; Mollá et al., 2014; Urbaneja et al., 2009; Yiacoumi et al., 2024). Besides, predation capacity of Dicyphus bolivari Lindberg and D. errans (Wolff) on P. absoluta eggs and larvae has been evaluated (Ingegno et al., 2013, 2019). In South America, the native region of P. absoluta, Bueno et al. (2012) tested the predation capacity of the mirids Campyloneuropsis infumatus (Carvalho), Engytatus varians (Distant) and Macrolophus basicornis (Stål) on P. absoluta eggs and larvae inside mines, by drilling the plant tissues. Similar studies were conducted with the mirid Tupiocoris cucurbitaceus (Spinola) (López et al., 2012, 2019; Burla et al., 2022; Montiel Cáceres et al., 2024). Overall, evaluations of different mirid species have shown that all can prey on P. absoluta eggs and first-instar larvae, with adult females typically exhibiting higher consumption rates. At least four species are being commercialized in Europe and Latin America as biocontrol products (Table 1).
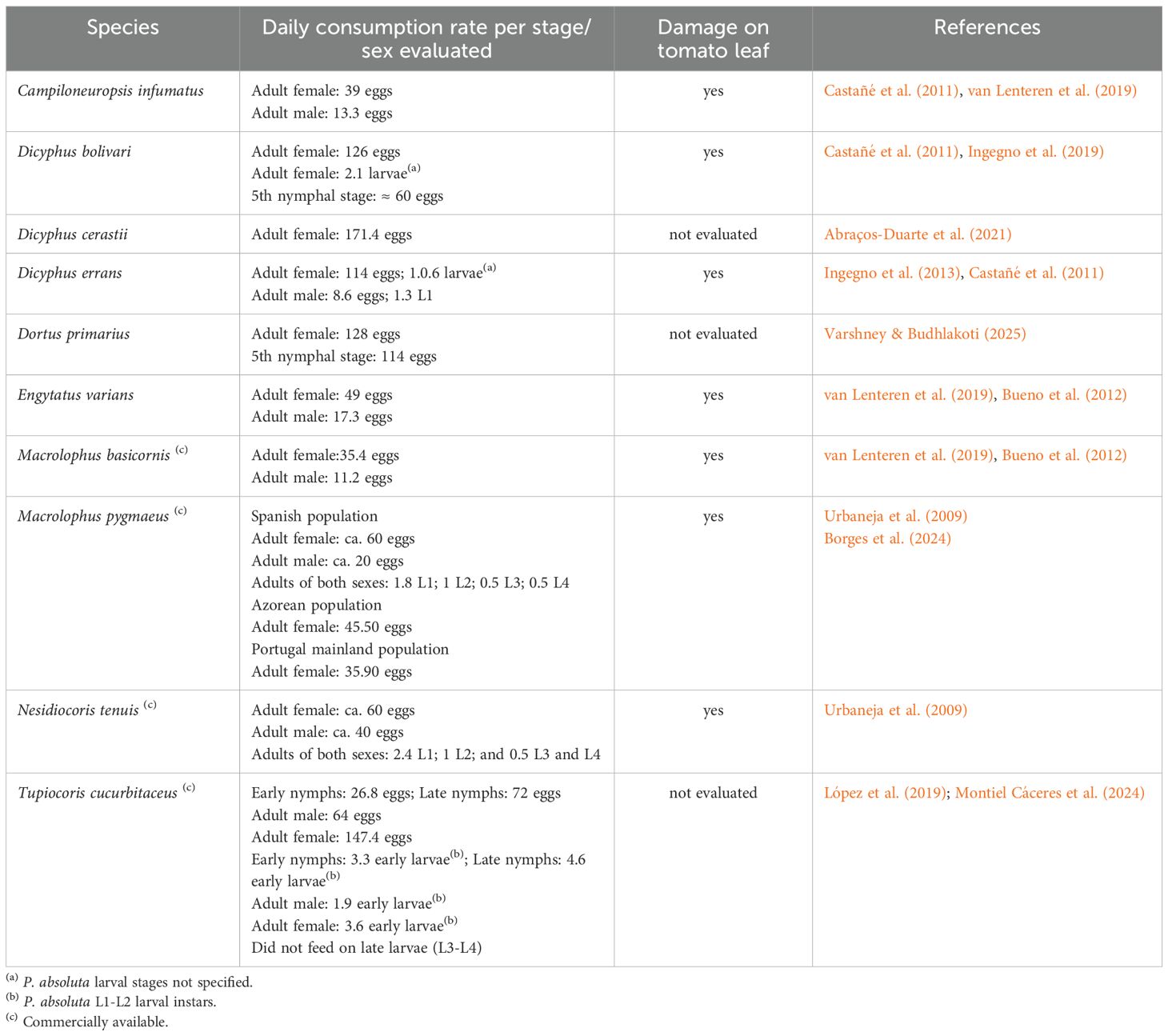
Table 1. A review of prey consumption rates of mirid species evaluated as biocontrol agents of Phthorimaea absoluta at similar conditions.
The evaluation of biocontrol agents involves a multi-stage process, including laboratory and field experiments to select promising candidates for application in cropping systems. Factors such as prey type, prey developmental stage, and predator sex can influence predation behavior (Coll and Guershon, 2002). For example, it has been observed that T. cucurbitaceus females consume a greater number of P. absoluta eggs and small larvae than nymphs and adult males (López et al., 2019). Moreover, the successful establishment and persistence of mirids in crop systems not only requires the presence of suitable prey but also depends critically on the availability of appropriate host plants for egg-laying (Lundgren et al., 2008). The establishment and reproduction of the natural enemies of pests in a crop strongly depends on the morphological and nutritional characteristics of the plant (Coll and Ridgway, 1995), which directly influence female oviposition behavior and the success of the offspring. In this context, the availability of oviposition sites within the crop or in the surrounding plants may be crucial to the success of both augmentative and conservation biological control strategies.
In Argentina, the augmentative release of T. cucurbitaceus has gained attention as a promising strategy for controlling P. absoluta eggs and immatures, as well as whiteflies (Hemiptera: Aleyrodidae), particularly in greenhouse crops (Polack et al., 2017). Recent field surveys have revealed that another mirid species, Campyloneuropsis cincticornis (Stål) (Hemiptera: Miridae) (Figure 1), is frequently found in horticultural farms, closely associated with tomato crops Solanum lycopersicum L. (Solanales: Solanaceae) and wild solanaceous plants, especially the sticky nightshade, Solanum sisymbriifolium Lam. (Solanales: Solanaceae) (Montiel Cáceres et al., 2023; Sola S.A., personal observation). This mirid has been reported as a zoophytophagous species inhabiting natural areas in various regions of Argentina and Brazil associated with other solanaceus plant species such as tobacco Nicotiana tabacum L., the black nightshade Solanum atriplicifolium G. Mey. and S. sisymbriifolium plants (Nogueira et al., 2019; Melo et al., 2024, 2025 https://biodar.unlp.edu.ar/cimicomorpha/es/info/21122.html). Due to its widespread presence and its association with solanaceous hosts in the main horticultural area located in northern Buenos Aires province (Carpintero et al., 2014; Montiel Cáceres et al., 2023), a better understanding of its biology is needed to assess its potential as a biological control agent, either through conservation or augmentative release strategies. To date, no studies have evaluated the predatory behavior of C. cincticornis on P. absoluta. Therefore, the present work represents the first report assessing its potential as a biological control agent against this key pest. Specifically, the objectives of this study were to evaluate: (1) the consumption rate of C. cincticornis on P. absoluta eggs and larvae, (2) the oviposition behavior of the predator on S. sisymbriifolium and tomato, and (3) its development time on both solanaceous plant species.

Figure 1. (a) Dorsal view of an adult of Campyloneuropsis cincticornis. (b) A nymph feeding on a Phthorimaea absoluta larva.
Methodology
Source of S. lycopersicum and S. sisymbriifolium plants and insects
Insects used in the experiments were obtained from colonies maintained at CEPAVE (CONICET-UNLP-CICPBA). Colonies were initially established with individuals collected from various horticultural sites in the peri-urban horticultural belt of La Plata, Buenos Aires Province, Argentina (34°54’29’’S, 58°02’25’’W). Tomato plants, S. lycopersicum var. ‘Platense’, used for insect rearing and experimental trials were obtained from a commercial nursery (Babyplant SRL) and the Gorina Experimental Station (MDA, Buenos Aires province, Argentina). Seedlings were transplanted into black plastic pots filled with a perlite-soil substrate and kept pest-free inside voile mesh cages without insecticide treatments. Plants were grown in 3-liter plastic pots with peat and perlite and fertilized every 15 days with 1.5 mg per plant of Nitrofoska foliar (PS COMPO: www.compo-expert.com, Santiago, Chile) and maintained in the greenhouse at CEPAVE (24 ± 3 °C, 65 ± 10% RH, and 14:10 (L:D) h photoperiod) during approximately 60 days until they reached the appropriate size for use (4–5 fully expanded leaves).
The P. absoluta colony was reared on tomato plants within voile cages (30 × 30 × 60 cm) under controlled environmental conditions (25 ± 2 °C, 70 ± 10% RH, 14:10 h light:dark photoperiod) in a walking room chamber. Individuals of C. cincticornis were collected from S. sisymbriifolium plants in horticultural fields and maintained in voile cages (30 × 30 × 60 cm) under the same environmental conditions. Eggs of Ephestia kuehniella Zeller (Lepidoptera: Pyralidae), obtained from a commercial supplier (Brometán SRL, Burzaco, Argentina), were used as food. Tomato cuttings placed in water-filled vials were also provided to C. cincticornis colonies for oviposition. Colonies were checked and replenished every 48 hours.
To obtain adults of standardized age, newly emerged C. cincticornis were individually placed in large plastic containers covered with voile, each containing a tomato stem in water and a paper strip with E. kuehniella eggs. Containers were labeled with the adult emergence date and monitored daily until the individuals reached 7 days of age, at which point they were used in experiments.
C. cincticornis consumption assay
We assessed the predation rate of C. cincticornis on P. absoluta eggs and early instar larvae (L1–L2), both inside and outside the leaf mines. Each experimental unit consisted of a Petri dish (9 cm diameter × 2 cm height) containing a tomato leaflet with prey corresponding to each treatment. A single predator individual was then introduced according to the assigned treatment. Trials were conducted for all nymphal instars (N1 to N5) and for 7-day-old adult females and males of the predator. When eggs were offered as prey, the number of eggs per treatment was 15, 25, 45, 80, 100, and 200 for N1 through N5 and adults, respectively. For larval predation assays, L1-L2 larval instars of P. absoluta were used because, due to their size, most mirids are unable to prey on the later instars (L3–L4) (López et al., 2019). Five larvae located inside or outside the mines, depending on the treatment, were offered to each predator stage of C. cincticornis. In the treatment with larvae outside the mines, the time the larvae were exposed, that is, the time it took them to enter a leaf and become concealed, was recorded. Control treatments (with prey but no predators) were included to account for natural mortality. Predation rates were corrected using Abbott’s formula (Abbott, 1925). Prey densities were determined through preliminary trials to estimate the maximum consumption levels, and in all cases, prey numbers exceeded the maximum consumption observed in these preliminary tests. Each treatment was replicated between 15–20 times. Experiments were conducted under controlled conditions (25 ± 2 °C, 70 ± 10% RH, 14:10 h light:dark photoperiod). After 24 h, the number of prey consumed was recorded. No prey mortality was observed in the control treatments.
Oviposition assay
The oviposition capacity of C. cincticornis was tested on tomato and S. sisymbriifolium plants in a non-choice experiment. The experimental unit consisted of one plant in a pot, enclosed in a voile cage (30 × 30 × 60 cm), each containing one male and one female adult between 7 and 9 days old. A strip of paper with E. kuehniella eggs was also provided as food. For tomato treatments, individual plants (~20 cm tall, with 4–5 fully expanded leaves) were used as oviposition substrates. After 48 hours, the adult pair was removed, and plants were monitored daily for nymph emergence over 20 days. For S. sisymbriifolium, fresh stems were collected from the field, as this plant species is difficult to maintain under greenhouse conditions. Stems (with at least two leaves, no flowers) were selected to reduce wilting. As most stems carried insects, they were immersed in water for 15 minutes to eliminate any residents. After drying, stems were placed in the same cage setup used for tomatoes. Because the egg incubation period for C. cincticornis was unknown, but ranges from 8 to 19 days in the related T. cucurbitaceus, any nymphs emerging within the first 8 days after oviposition were removed and excluded from analysis. Only nymphs emerging after day 8 were counted. Daily inspections continued for 20 days under controlled environmental conditions (25 ± 2 °C, 70 ± 10% RH, 14:10 h light:dark photoperiod).
C. cincticornis developmental time assay
A no-choice experiment was conducted to evaluate the effect of host plant species (S. lycopersicum and S. sisymbriifolium) on the egg hatching time and nymphal development duration of C. cincticornis. For each plant species, individual potted plants were placed in voile cages (30 × 30 × 60 cm) and used as experimental units that were maintained under the same controlled environmental conditions described above.
One mated pair of adult C. cincticornis (1♀:1♂) was introduced onto each plant and allowed to oviposit for 48 hours. After this period, the adults were carefully removed to ensure that only a single cohort of eggs remained on each plant. Plants were then monitored daily to record the timing of egg hatching. After hatching, groups of five first-instar nymphs (NI) that emerged on the same day were placed together on the same plant and tracked to determine the duration of nymphal development until adult emergence. Observations were conducted daily, and the developmental time (in days) was recorded for each individual from the emergence of the first to the last adult. As adults emerged, they were immediately removed from the experimental unit to prevent interference with remaining nymphs and to accurately track individual development times. Both adult females during the oviposition period and nymphs throughout their development were provided with E. kuehniella eggs ad libitum as a food source. A total of 17 replicates were performed using S. sisymbriifolium and 8 replicates with tomato. Separate datasets were generated for egg development time and nymphal development time.
Statistical analysis
C. cincticornis consumption assay
Predation across developmental stages was analyzed using GLMs with binomial error distribution and a logit link function. The response variable in all models was the number of prey consumed and not consumed, specified as a two-column matrix. Egg predation was analyzed separately for nymphal instars and for adults (males and females). Nymphal instar or adult sex was used as the explanatory variable. Post hoc pairwise comparisons among instars or sexes were performed using Tukey’s Honestly Significant Difference (HSD) test. The predation of P. absoluta larvae located inside the mine was analyzed using a binomial GLM, with either nymphal instar (from NII to NV) or adult sex (male or female) as the predictor variable. First-instar nymphs (NI) were excluded from this analysis because they did not consume any larvae. The second instar (NII) was used as the reference category for nymphs. A separate binomial GLM was used to evaluate the predation of larvae outside the leaf mine. The model structure was the same as for the previous analysis, with nymphal instar or adult sex as the explanatory variable. Again, first-instar nymphs (NI) were excluded due to absence of predation. To assess the effect of larval location on predation probability, we conducted a third set of binomial GLMs in which larval position (inside vs. outside the mine) was included as the explanatory variable. Separate models were run for nymphal instars and for adults.
Oviposition assay
The number of emerging nymphs was analyzed using a GLM with a negative binomial distribution to account for overdispersion. Plant species was the explanatory variable.
C. cincticornis developmental time assay
To examine the effect of host plant species on the predatory development, generalized linear models (GLMs) assuming a Poisson distribution with a log-link function were fitted. Two models were run: one for egg hatching time and one for nymphal development (NI to adult). In both cases, host plant species were used as the predictor.
For all analyses, we calculated odds ratios (ORs) by exponentiating the estimated regression coefficients (log-odds) from the GLMs using the formula OR = e^β. These ORs represent the change in odds of a successful predation event relative to the reference level. An OR > 1 indicates a higher likelihood of predation, while an OR < 1 indicates a reduced likelihood compared to the baseline. All analyses were conducted in R (v.12.0) software using the glm function from the stats package (R Core Team, 2024). Model fit was assessed using the Akaike Information Criterion (AIC).
Results
Consumption assay
Eggs
A significant effect of nymphal instar on egg consumption was detected (LR χ² = 170.88; df = 4; p < 0.001; Figure 2A). Compared to first-instar nymphs (NI), instars NIII (β = 0.99 ± 0.19; z = 5.14; p < 0.001), NIV (β = 0.92 ± 0.19; z = 4.91; p < 0.001), and NV (β = 1.45 ± 0.18; z = 7.91; p < 0.001) showed significantly higher probabilities of egg consumption. The difference between NI and NII was not statistically significant (p = 0.236). Odds ratios indicated that NIII, NIV, and NV were 2.7, 2.5, and 4.3 times more likely to consume eggs than NI, respectively. Pairwise comparisons confirmed significant differences between most instars, except between NIII and NIV (p = 0.925).
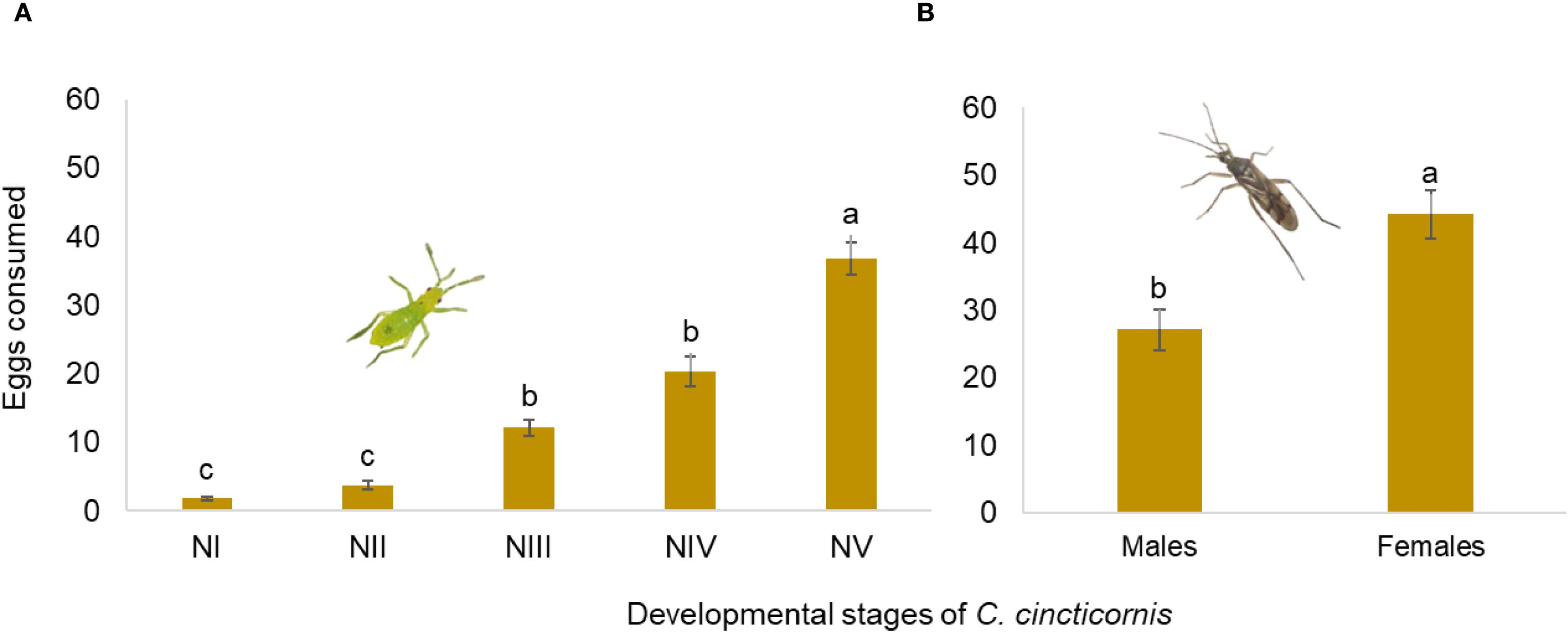
Figure 2. Mean number (± SE) of Phthorimaea absoluta eggs consumed by (A) different nymphal instars and (B) adult males and females of Campyloneuripsis cincticornis, provided in tomato leaves during 24 h. Different lowercase letters indicate significant differences according to the Tukey’s HSD test at α = 0.05.
In adults, sex significantly affected predation success (LR χ² = 128.4; df = 1; p < 0.001; Figure 2B). Females exhibited a higher predation rate than males (β = 0.76 ± 0.07; z = 11.21; p < 0.001), with an odds ratio of approximately 2.13, indicating that females were more than twice as likely to consume eggs.
Larvae inside the mine
First-instar nymphs (NI) did not consume any larvae. The model results indicated a trend toward increased predation success with advancing nymphal instar, although differences were not statistically significant at the 0.05 level (LR χ² = 6.46; df = 3; p = 0.09; Figure 3A). Predation probability was highest for fifth-instar nymphs (β = 1.86 ± 1.09; z = 1.70; p = 0.08) and fourth instars (β = 2.03 ± 1.08; z = 1.88; p = 0.06), compared to second instars. Compared to second-instar nymphs (NII), third instars (NIII) were approximately 3.08 times more likely to consume a P. absoluta larva, while fourth (NIV) and fifth instars (NV) were 7.62 and 6.44 times more likely to do so, respectively. Although these differences did not reach conventional levels of statistical significance, the magnitude of the odds ratios suggests a biologically relevant increase in predatory efficiency with nymphal instar.
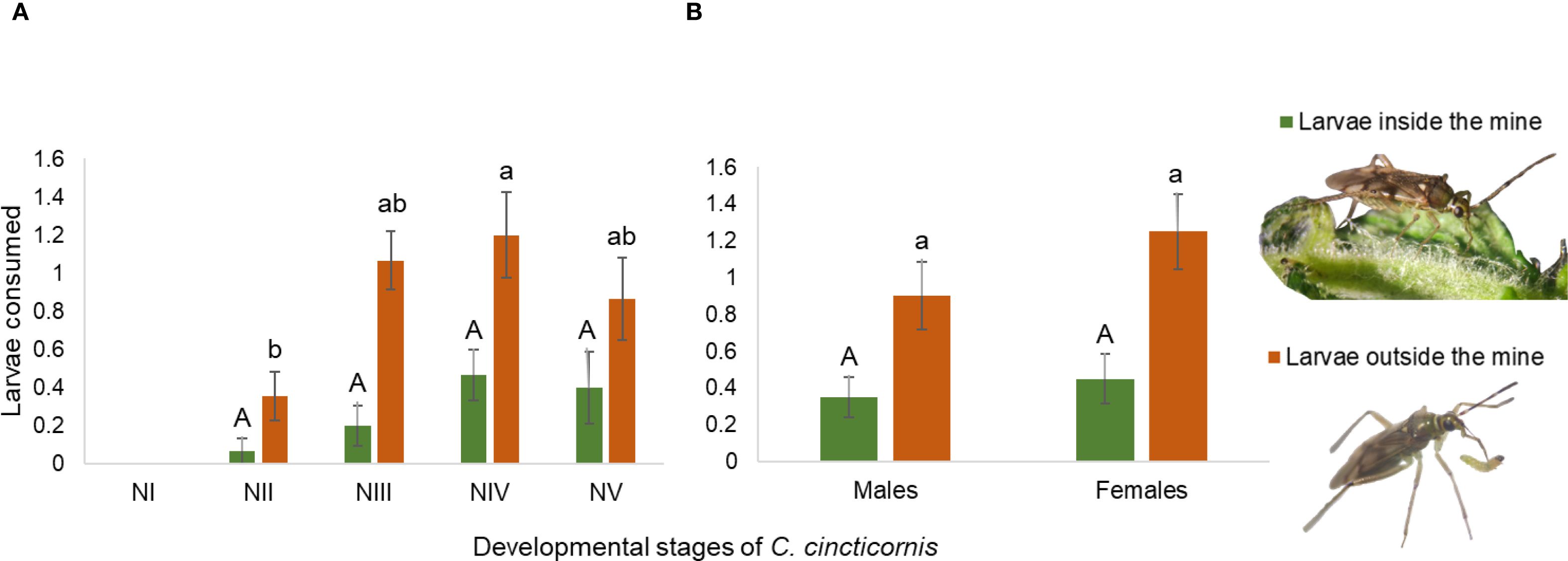
Figure 3. Mean number (± SE) of Phthorimaea absoluta larvae inside and outside tomato leaf mines consumed by (A) different nymphal instars and (B) adult males and females of Campyloneuropsis cincticornis during 24 h. Different uppercase letters indicate a statistically significant difference in the mean consumption of larvae inside the mines according to the Tukey’s HSD test at α = 0.05. Conversely, different lowercase letters denote a statistically significant difference in the mean consumption of larvae outside the mines (Tukey’s HSD test at α = 0.05).
In adults, the model indicated no significant difference in the number of larvae consumed between sexes (β = –0.25 ± 0.50 SE; z = –0.50; p = 0.62; Figure 3B). The mean number of larvae consumed by females was approximately 0.45, and males consumed on average 22% fewer larvae; however, this difference was not statistically significant.
Larvae outside the mine
The model detected a statistically significant effect of nymphal instar on predation success (LR χ² = 9.15; df = 3; p = 0.027). Compared to second-instar nymphs (NII), instars NIII and NIV showed significantly greater probabilities of predation, with log-odds estimates (± SE) of 1.26 ± 0.54 (p = 0.02) and 1.41 ± 0.54 (p = 0.009), respectively. Fifth-instar nymphs (NV) also exhibited a positive effect, but this was only marginally significant (β = 1.00 ± 0.56; p = 0.071). This corresponds to a ~3.5, 4.11 and 2.73-fold increase in the odds of predation NIII, NIV and NV, respectively, relative to NII.
Relative to adults, the model did not reveal a significant effect of sex on larval consumption (LR χ² = 1.46; df = 1; p = 0.23), and no meaningful difference in predation probability was detected between males and females. The effect of being male (β = −0.42 ± 0.35; z = −1.20; p = 0.23) suggested that males were about 34% less likely to consume larvae than females, although this difference was not statistically supported.
Consumption inside versus outside the mines
The mean exposure time of larvae, i.e., until leaf penetration and protection, was 165 ± 8.2 minutes. The model showed a statistically significant effect of larval position on the probability of predation by nymphal instars (LR χ² = 21.61; df = 1; p < 0.001). Predation probability was significantly higher when larvae were outside the leaf mine compared to when they were inside. Specifically, the odds of predation increased by 1.27 units for larvae located outside the mine (β = 1.27 ± 0.29; z = 4.34; p < 0.001). This corresponds to an approximate 3.56-fold increase in the odds of predation outside the mine, relative to inside.
In relation to adults, the analysis revealed a highly significant effect of position on predation probability (LR χ² = 37.80; df = 1; p < 0.001). Larvae positioned outside the mine were significantly more likely to be preyed upon compared to those located inside. The odds of predation increased by 1.86 units when larvae were outside the mine (β = 1.86 ± 0.33; z = 5.76; p < 0.001), corresponding to an approximate 6.42-fold increase in the odds of predation.
Oviposition assay
The number of emerged nymphs of C. cincticornis differed significantly between plant species (LR χ² = 19.82; df = 1; p < 0.001). Females laid significantly more eggs on S. sisymbriifolium than on tomato plants. Specifically, the estimated rate of nymph emergence on this noncrop solanaceous was 7.36 times higher than on tomato (β = 1.99 ± 0.46; z = 4.38; p < 0.0001), suggesting a strong oviposition preference under no-choice conditions (Figure 4).
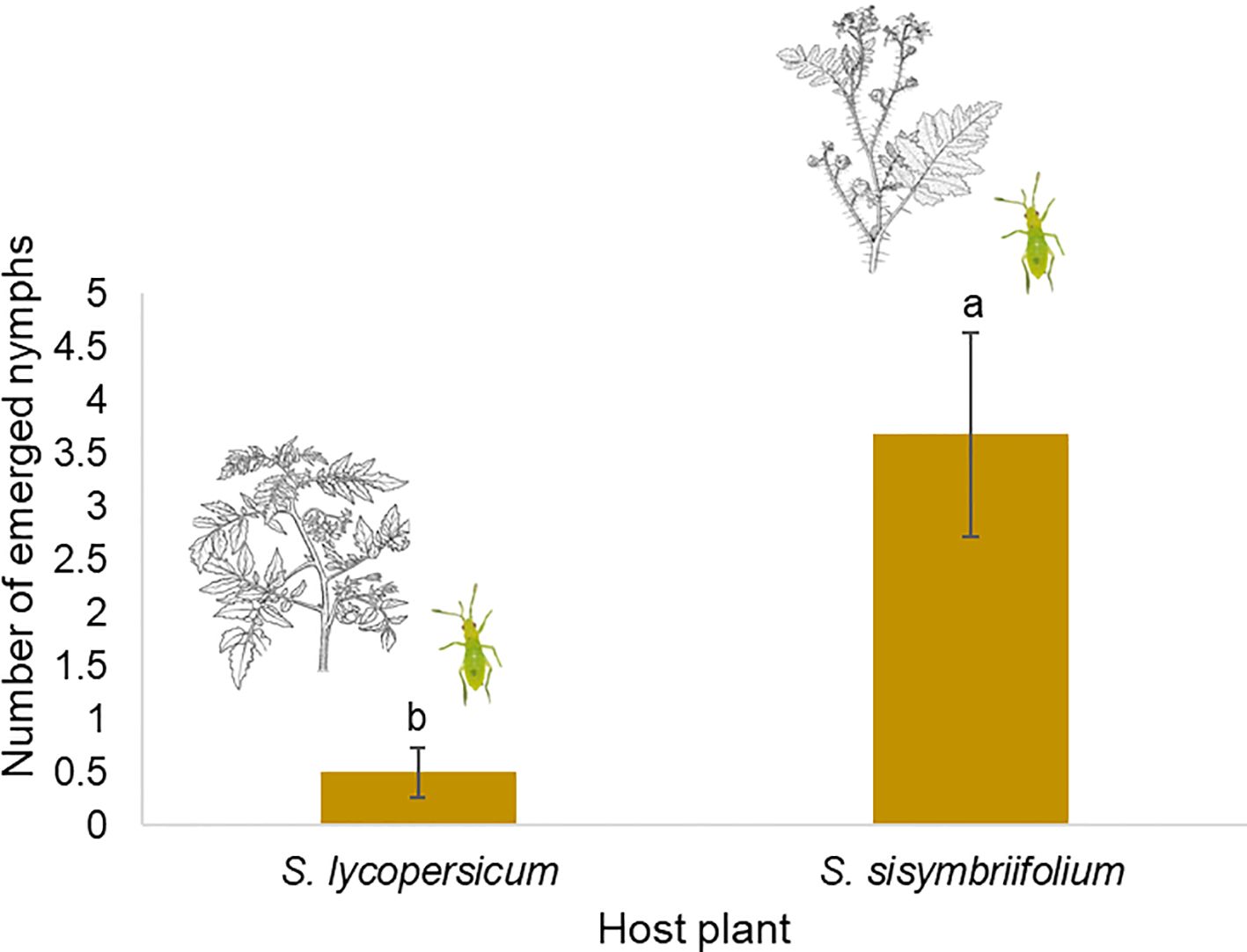
Figure 4. Mean number (± SE) of emerged nymphs from Solanum lycopersicum and S. sisymbriifolium host plants, after allowing females to oviposit for 48 h. Different lowercase letters indicate significant differences according to the Tukey’s HSD test at α = 0.05.
Developmental time
The host plant species had no significant effect on the egg development time of C. cincticornis (χ² = 0.24; df = 1; p = 0.62). Although the coefficient for the S. sisymbriifolium treatment was positive (β = 0.06 ± 0.13; z = 0.49; p = 0.62), it was not statistically significant, indicating similar egg development durations on both plant species (Figure 5). In contrast, the host plant had a significant effect on the nymphal development time of C. cincticornis (χ² = 14.63; df = 1; p < 0.001). Individuals reared on S. sisymbriifolium developed significantly faster than those reared on tomato (Figure 5). The estimated coefficient was negative and significant (β = –0.38 ± 0.10; z = –3.88; p < 0.001), suggesting a 31.6% reduction in development time on S. sisymbriifolium relative to tomato.
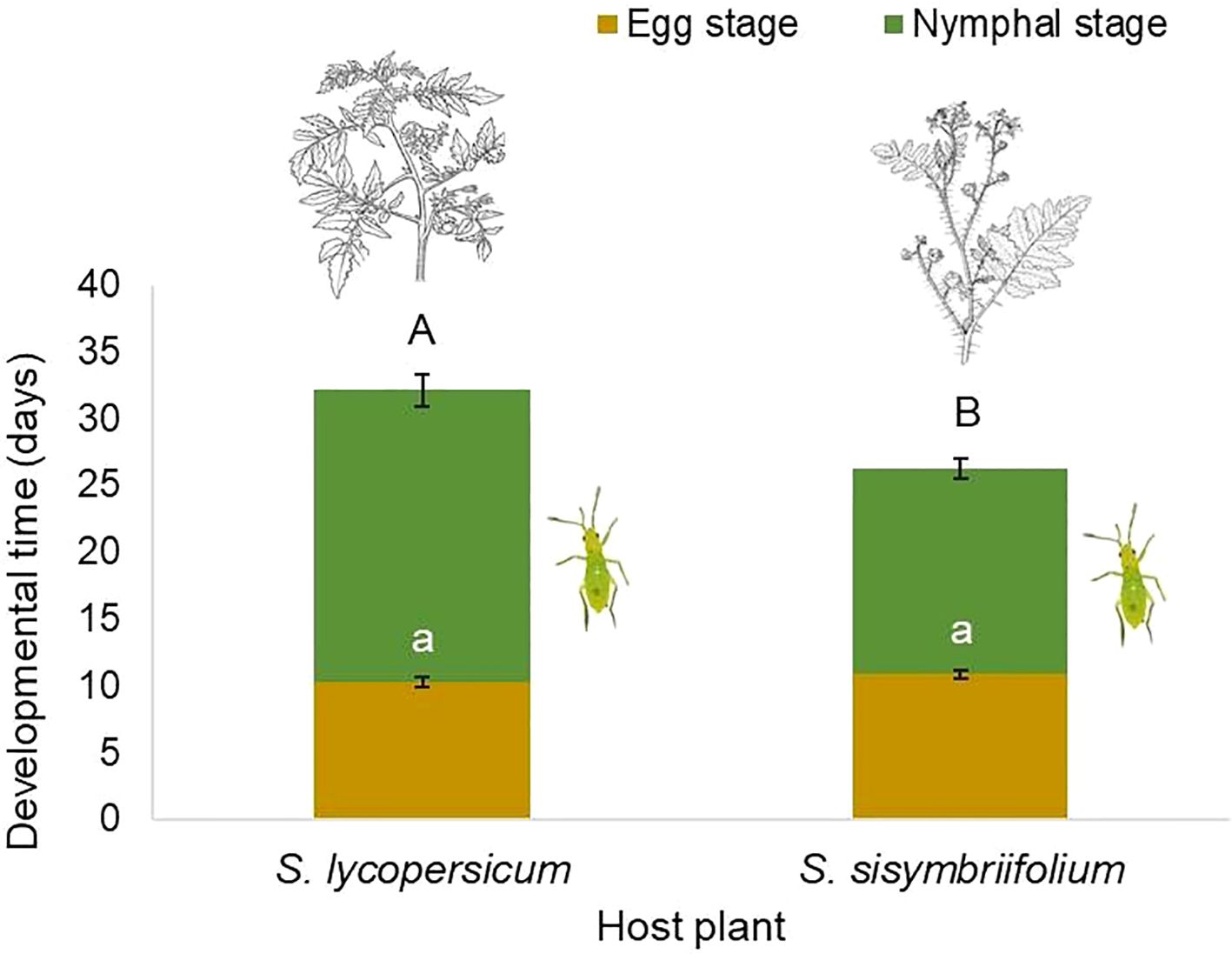
Figure 5. Egg and nymphal development times (days) of Campyloneuropsis cincticornis on Solanum lycopersicum and S. sisymbriifolium plants. Bars represent means ± SE. Similar lowercase letters indicate no statistically significant difference between egg developmental time according to the Tukey’s HSD test at α = 0.05. Conversely, different uppercase letters show a statistically significant difference between nymphal developmental time (Tukey’s HSD test at α = 0.05).
Discussion
This study is the first to evaluate the biology and predatory behavior of C. cincticornis against P. absoluta in laboratory conditions, providing novel insights into its potential as a biological control agent. Campyloneuropsis cincticornis showed the ability to feed on both eggs and larvae of P. absoluta, both inside and outside leaf mines, although its efficiency varied depending on the predator’s developmental stage, the type of prey, and its location.
The efficiency of P. absoluta egg predation by C. cincticornis increased significantly with nymphal development, with late instars (NIII–NV) consuming more eggs than early ones (NI–NII). This trend aligns with findings for other zoophytophagous mirids such as T. cucurbitaceus, where later instars also showed higher feeding activity (López et al., 2019). Adults, particularly females, were more effective predators than nymphs, likely due to higher energetic demands associated with reproduction (Calvo et al., 2012). When comparing the egg predation rates of C. cincticornis with those reported for other mirid species (Table 1), a wide variability is evident. Some species, such as D. bolivari and D. errans, show very high rates (over 100 eggs/day; Ingegno et al., 2019), while M. basicornis and E. varians also exceed 90 eggs/day (Bueno et al., 2012; van Lenteren et al., 2016). In contrast, C. cincticornis showed lower rates, comparable to C. infumatus (~51 eggs/day), and within the range reported for N. tenuis and M. pygmaeus (20–60 eggs/day) (Urbaneja et al., 2009). These interspecific differences may be attributed to distinct taxonomy, ecological niches, predator morphology, feeding behavior, or methodological differences between studies (Ingegno et al., 2013; van Lenteren et al., 2018). Therefore, the developmental stage is a key factor when considering this predatory species for biological control programs.
In contrast, larval predation showed a more complex pattern. While consumption of larvae inside mines was low—suggesting mechanical difficulty in accessing protected prey—significant predation was recorded on larvae exposed outside mines. Later nymphal instars of C. cincticornis are significantly more effective predators of P. absoluta larvae outside the mine compared to early nymphal instars. This supports the potential use of older nymphs in augmentative biological control programs, especially for targeting mobile or more exposed prey stages. Among adults, both males and females exhibited similar efficiency in preying on exposed larvae, with no significant differences under the conditions tested. In relation to larval consumption rates of C. cincticornis adults, our results were notably lower compared to those reported for other mirid predators, as females and males of D. errans preying on an average of 2.40 ± 0.54 and 1.30 ± 0.29 larvae (Ingegno et al., 2013). These findings are ecologically relevant, as P. absoluta exhibits a mining habit that complicates control efforts, especially during larval stages protected within plant tissues (Desneux et al., 2010). However, since larvae often leave mines to create new ones, they become temporarily exposed, providing a window during which C. cincticornis could act as an effective natural enemy. This reinforces its potential role as a complementary agent within IPM strategies.
Notably, our oviposition assays revealed a preference of C. cincticornis for S. sisymbriifolium under no-choice conditions, suggesting that this wild solanaceous species, that is part of the spontaneous vegetation commonly found in farms within the study area (Salas Gervassio et al., 2016), may offer favorable traits for oviposition. This result aligns with our field observations of the predominance of C. cincticornis on this plant (Montiel Cáceres et al., 2023) and highlights the importance of non-cultivated plants as a favorable reproductive substrate, which has practical implications for the development of conservation strategies aimed at improving the early establishment and persistence of mirid populations in crop systems. However, it is noteworthy that C. cincticornis also accepted the tomato plant as a suitable oviposition substrate; so it is plausible that C. cincticornis individuals may disperse from S. sisymbriifolium into tomato crops in search of prey. The ability to oviposit directly on the tomato plant may facilitate the establishment and persistence of C. cincticornis within the crop, particularly in scenarios where preferred oviposition hosts are scarce or absent. For example, the ability of D. errans to colonize tomato crops early in the season in Italy has been reported, when the European black nightshade Solanum nigrum L., one of the mirid’s hosts, is present in the environment (Ingegno et al., 2019). The same has been reported for M. pygmaeus (Ferracini et al., 2012; Lykouressis et al., 2000; Perdikis et al., 2008). Thus, predators that can move through different types of vegetation can play a strategic role in pest control throughout the crop cycle. Incorporating reservoir plants such as S. sisymbriifolium along field margins could significantly contribute to the successful implementation of IPM programs that include native mirid species. In addition, attraction of crop and noncrop plant species to predatory mirid bugs involving volatile cues is being studied to better understand the foraging behavior and prey detection (Adams et al., 2023; S.A. Sola unpublished data).
As mentioned above, the establishment and reproduction of mirid predators in a crop depends on the characteristics of the plant, which directly influences the oviposition of females and the success of the offspring. In relation to the number of eggs laid by C. cincticornis females on the evaluated host plants, we recorded a lower oviposition rate compared with the values reported for other mirid predators such as M. basicornis, E. varians, and C. infumatus on tomato, which ranged from 4 to 5 eggs per female per day (Silva et al., 2016). These differences may reflect species-specific reproductive strategies and physiological traits, as fecundity and fertility can vary considerably among zoophytophagous mirids (Castañé et al., 2004; Perdikis et al., 2011). Furthermore, methodological factors—such as the limited duration of the oviposition assay, prey availability, or the indirect count for the impossibility to detect eggs due to their endophytic nature—may also have contributed to the lower observed values. Given that C. cincticornis is a less-studied neotropical predator, further targeted studies are needed to comprehensively evaluate its reproductive potential under different conditions. In relation to offspring performance, our results showed that host plant species had no significant effect on the egg development time of C. cincticornis, indicating that both host plants provide similarly suitable conditions for embryonic development. However, nymphal development was significantly faster on S. sisymbriifolium, underscoring the role of host plant identity in shaping predator performance and may have implications for the population growth of C. cincticornis in diverse cropping systems. Compared to other neotropical mirids studied, such as C. infumatus (16.2 days), E. varians (17.4 days), and M. basicornis (18.6 days) fed with a mix of P. absoluta eggs and larva (Silva et al., 2016), C. cincticornis demonstrated comparable or even shorter development times when reared on S. sisymbriifolium. These values are also in line with those reported for the European species M. pygmaeus fed on E. kuehniella (Vandekerkhove and De Clercq, 2010), and notably shorter than those of D. errans reared on P. absoluta eggs (~24 days) and on E. kuehniella eggs (~20 days) (Ingegno et al., 2013). As previously noted for other zoophytophagous mirids, developmental times can vary substantially depending on diet and host plant quality, with unsuitable substrates often resulting in delayed development or high nymphal mortality (Gillespie and Mcgregor, 2000; Perdikis and Lykouressis, 2000). In our case, the differences in nymph developmental time observed between host plants could be attributed to the nutritional contribution of plant fluids. Due to the zoophytophagous habit of C. cincticornis, it is possible that S. sisymbriifolium provides some nutrients that enhance the developmental performance of this predator. Our results highlight the potential of S. sisymbriifolium not only as a favorable oviposition substrate but also as a host plant that enhances nymphal performance in C. cincticornis.
Mirid species such as M. pygmaeus and N. tenuis have been successfully incorporated into IPM programs in Europe for the control of P. absoluta, demonstrating the potential of mirids as key biocontrol agents in solanaceous crops (Belda et al., 2010; Mollá et al., 2011; Urbaneja et al., 2012). In Argentina, T. cucurbitaceus has shown comparable potential and is now being commercialized for IPM programs (López et al., 2019). Our findings indicate that C. cincticornis also possesses strong predatory capacity against P. absoluta, positioning it as a promising native candidate for biological control. Its ability to oviposit not only on tomato but primarily on S. sisymbriifolium, a non-cultivated solanaceous plant, reinforces its potential for their use mainly in conservation biological control strategies, where the availability of suitable non-crop hosts is crucial for sustaining predator populations. However, it should be noted that laboratory results may not fully reflect the complexity of field conditions, where factors such as climate, crop architecture, diverse prey communities, and intraguild interactions can influence predator performance. Therefore, field-based studies are necessary to validate its effectiveness before considering practical implementation (Desneux et al., 2022).
The suitability of S. sisymbriifolium as a host for both mirid species—C. cincticornis and T. cucurbitaceus— suggests its potential role as a reservoir plant, facilitating the early establishment and persistence of beneficial mirid populations in and around tomato fields. This strategy has been adopted in several countries, where alternative host plants have been shown to facilitate early colonization and the continued presence of predators throughout the crop cycles enhancing biological control outcomes (Ferracini et al., 2012; Ingegno et al., 2019; Burla et al., 2022). In this context, the incorporation of native plants such as S. sisymbriifolium into IPM strategies may serve a dual function: enhancing predator retention and preserving native biodiversity.
Within an IPM program for P. absoluta in Argentina, in addition to the use of insecticides, several bio-inputs are available, including formulations based on the bacterium Bacillus thuringiensis Berliner (Greco et al., 2020), pheromone-based monitoring tools (Triple T®, WayneAgro, CABA, Argentina), and biological control using the predatory mirid T. cucurbitaceus (López et al., 2019). Besides, semi-field releases of the endoparasitoid Pseudapanteles dignus (Muesebeck) (Hymenoptera: Braconidae) which have shown promising results (Salas Gervassio et al., manuscript submitted). In this context, the use of the mirid C. cincticornis must be compatible with other control techniques. In relation to biocontrol, understanding the interference interactions between C. cincticornis and other natural enemies is crucial for its effective incorporation into IPM programs. Some of these interactions have already been studied. For instance, positive results have been reported regarding the compatibility between T. cucurbitaceus and P. dignus (Sola et al., in press), and preliminary findings on intraguild predation between T. cucurbitaceus and C. cincticornis suggest no negative interactions between these species (J.N. Peñalba, unpublished data). In addition, ongoing experiments are evaluating intraguild predation of C. cincticornis on P. dignus. Altogether, these insights will be key to incorporating C. cincticornis into IPM programs for P. absoluta. Further studies under field conditions are essential to assess its efficacy, potential intraguild interactions, prey specificity, and possible phytophagous effects on tomato plants.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Ethics statement
Ethical approval was not required for the study involving animals in accordance with the local legislation and institutional requirements because Ethical approval for studies on Phthorimaea absoluta, is generally not required due to the low-risk nature of the research and because it is a pest species, invasive on several continents. Regarding Campyloneuropsis cincticornis, the procedures performed in this study were noninvasive and did not cause pain or stress to individuals (see Cooper 2006. Insects. In: Lewbart G.A. (ed.) Invertebrate medicine. Ed. Acribia, Spain). Once the experiments were terminated, C. cincticornis individuals were returned to the colony. It is important to mention that in Argentina, animal ethics committees for arthropods are beginning to be formed.
Author contributions
SS: Investigation, Writing – original draft, Validation, Data curation, Visualization, Methodology, Formal analysis. MR: Supervision, Methodology, Conceptualization, Writing – review & editing, Writing – original draft, Investigation, Visualization, Project administration, Formal analysis, Validation, Data curation. NSG: Writing – review & editing, Supervision, Investigation, Writing – original draft, Validation, Project administration, Formal analysis, Data curation, Methodology. JP: Data curation, Methodology, Visualization, Investigation, Writing – original draft. RMC: Methodology, Visualization, Investigation, Data curation, Writing – original draft. ML: Writing – review & editing, Funding acquisition, Writing – original draft, Project administration, Conceptualization, Supervision, Validation.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This research was funded by ANPCyT (PICT 2020-00764) and PID UNLP (N 966).
Acknowledgments
The authors would like to thank E. Nieves, J. Meaca and C.N. Roumec for providing help during the course of the study. Collection and transportation permits to conduct insect rearing were obtained from Dirección de Flora y Fauna, MDA, provincia de Buenos Aires, Argentina (EX-2021-15452539-GDEBA-DSTAMDAGP).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that Generative AI was used in the creation of this manuscript. We acknowledge the use of ChatGPT online in helping with the revision of the English language during the preparation of this manuscript. After using this tool, the authors reviewed and edited the content as needed and took full responsibility for the content of the publication.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abbott W. S. (1925). A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 18, 265–267. doi: 10.1093/jee/18.2.265a
Abraços-Duarte G., Ramos S., Valente F., Borges da Silva E., and Figueiredo E. (2021). Functional response and predation rate of Dicyphus cerastii Wagner (Hemiptera: Miridae). Insects 12, 530. doi: 10.3390/insects12060530
Adams B., Yusuf A. A., Torto B., and Khamis F. M. (2023). Non-host plant odors influence the tritrophic interaction between tomato, its foliar herbivore Tuta absoluta and mirid predator Nesidiocoris tenuis. Front. Plant Sci. 14. doi: 10.3389/fpls.2023.1014865
Belda J. E., Calvo F. J., and Giménez A. (2010). Biological control strategy for Tuta absoluta in tomato by release of Nesidiocoris tenuis in pre-transplant. Phytoma Esp. 217, 48–52.
Borges I., Oliveira L., Barbosa F., Figueiredo E., and Franco J. C. (2024). Prey consumption and conversion efficiency in females of two feral populations of Macrolophus pygmaeus, a biocontrol agent of Tuta absoluta. Phytoparasitica. 52(2). doi: 10.1007/s12600-024-01130-0
Bueno V. H. P. (2009). Biological pest control: Mass production and quality control. 2nd Edn (Lavras, MG, Brazil: UFLA Press).
Bueno V. H. P., Van Lenteren J. C., Lins J. C., Calixto A. M., Montes F. C., Silva D. B., et al. (2012). New records of Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) predation by Brazilian Hemipteran predatory bugs. J. Appl. Entomol. 137, 29–34. doi: 10.1111/jen.12017
Burla J. P., Arbulo N., Aldabe J., Fagundez C., and Castiglioni E. (2022). Companion plants for conservative management of Tupiocoris cucurbitaceus (Spinola 1852) (Heteroptera: Miridae: Dicyphini) on greenhouse tomato crops. Entomol. Commun. 4, ec04028. doi: 10.37486/2675-1305.ec04028
Calvo F. J., Bolckmans K., and Belda J. E. (2012). Release rate for a pre-plant application of Nesidiocoris tenuis for Bemisia tabaci control in tomato. BioControl. 57, 809–817. doi: 10.1007/s10526-012-9455-1
Carpintero D. L., De Biase S., and Konopko S. A. (2014). Hemiptera-Heteroptera de la Reserva Ecológica Costanera Sur (ciudad autónoma de Buenos Aires), Argentina. Rev. Mus. Argent. Cienc. Nat. 16, 67–80.
Castañé C., Alomar O., Goula M., and Gabarra R. (2004). Colonization of tomato greenhouses by the predatory mirid bugs Macrolophus caliginosus and Dicyphus tamaninii. Biol. Control. 30, 591–597. doi: 10.1016/j.biocontrol.2004.02.012
Castañé C., Arnó J., Gabarra R., and Alomar O. (2011). Plant damage to vegetable crops by zoophytophagous mirid predators. Biol. Control. 59, 22–29. doi: 10.1016/j.biocontrol.2011.03.007
Coll M. and Guershon M. (2002). Omnivory in terrestrial arthropods: mixing plant and prey diets. Annu. Rev. Entomol. 47, 267–297. doi: 10.1146/annurev.ento.47.091201.145209
Coll M. and Ridgway R. L. (1995). Functional and numerical responses of Orius insidiosus (Heteroptera: Anthocoridae) to its prey in different vegetable crops. Ann. Entomol. Soc Am. 88, 732–738. doi: 10.1093/aesa/88.6.732
Desneux N., Han P., Mansour R., Arnó J., Brévault T., Campos M. R., et al. (2022). Integrated pest management of Tuta absoluta: practical implementations across different world regions. J. Pest Sci. 95, 17–39. doi: 10.1007/s10340-021-01442-8
Desneux N., Wajnberg E., Wyckhuys K. A. G., Burgio G., Arpaia S., Narváez-Vasquez C. A., et al. (2010). Biological invasion of European tomato crops by Tuta absoluta: ecology, geographic expansion and prospects for biological control. J. Pest Sci. 83, 197–215. doi: 10.1007/s10340-010-0321-6
Ferracini C., Ingegno B. L., Mosti M., Navone P., Tavella L., and Alma A. (2012). Promising native candidates for biological control of Tuta absoluta in Italy. IOBC/WPRS Bull. 80, 51–55. doi: 10.5555/20123337594
Gillespie D. R. and Mcgregor R. R. (2000). The functions of plant feeding in the omnivorous predator Dicyphus hesperus: water places limits on predation. Ecol. Entomol. 25, 380–386. doi: 10.1046/j.1365-2311.2000.00285.x
Greco N., Walsh G. C., and Luna M. G. (2020). Biological control in Argentina, in Biological Control in Latin America and the Caribbean: Its Rich History and Bright Future, 21-42. Ed. van Lenteren et al. (CABI Publishing Wallingford). doi: 10.1079/9781789242430.0021
Ingegno B. L., Ferracini C., Gallinotti D., Alma A., and Tavella L. (2013). Evaluation of the effectiveness of Dicyphus errans (Wolff) as predator of Tuta absoluta (Meyrick). Biol. Control. 67, 246–252. doi: 10.1016/j.biocontrol.2013.08.002
Ingegno B. L., Messelink G. J., Bodino N., Iliadou A., Driss L., Woelke J. B., et al. (2019). Functional response of the mirid predators Dicyphus bolivari and Dicyphus errans and their efficacy as biological control agents of Tuta absoluta on tomato. J. Pest Sci. 92, 1457–1466. doi: 10.1007/s10340-019-01079-8
Jacas J. A. and Urbaneja A. (2008). Control biológico de plagas agrícolas (Valencia, Spain: Phytoma Ed).
López S. N., Orozco Muñoz A., Andorno A. V., Cuello E. M., and Cagnotti C. L. (2019). Predatory capacity of Tupiocoris cucurbitaceus (Hemiptera Miridae) on several pests of tomato. Bull. Insectology. 72, 201–205.
López S. N., Rojas F. A., Velásquez V. V., and Cagnotti C. (2012). Biology of Tupiocoris cucurbitaceus (Hemiptera: Miridae), a predator of the greenhouse whitefly Trialeurodes vaporariorum (Hemiptera: Aleyrodidae) in tomato crops in Argentina. Biocontrol Sci. Techn. 22, 1107–1117. doi: 10.1080/09583157.2012.705260
Lundgren J. G., Fergen J. K., and Riedell W. E. (2008). The influence of plant anatomy on oviposition and reproductive success of the omnivorous bug Orius insidiosus. Anim. Behav. 75, 1495–1502. doi: 10.1016/j.anbehav.2007.09.029
Lykouressis P., Perdikis D. C., and Chalkia C. A. (2000). The effects of νatural enemies on aphid populations on processing tomato in central Greece. Entomol. Hell. 13, 35–42. doi: 10.12681/eh.14036
Melo M. C., Castro-Huertas V., Minghetti E., Alivera L., Amaru Castelo J., and Dellapé P. M. (2024). Diversity of true bugs from the Rio Pilcomayo national park, Formosa province, Argentina. JIHS 1, 248–280. doi: 10.11646/jihs.1.4.6
Melo M. C., Montemayor S. I., Minghetti E., Varela P. S., and Dellapé P. M. (2025). Cimicomorpha (Hemiptera: Heteroptera) species from Argentina and Uruguay. Available online at: https://biodar.unlp.edu.ar/cimicomorpha/es/info/21122.html (Accessed May 20, 2025).
Mollá O., Biondi A., Alonso-Valiente M., and Urbaneja A. (2014). A comparative life history study of two mirid bugs preying on Tuta absoluta and Ephestia kuehniella eggs on tomato crops: implications for biological control. BioControl. 59, 175–183. doi: 10.1007/s10526-013-9553-8
Mollá O., González-Cabrera J., and Urbaneja A. (2011). The combined use of Bacillus thuringiensis and Nesidiocoris tenuis against the tomato borer Tuta absoluta. BioControl 56, 883–891. doi: 10.1007/s10526-011-9392-9
Montiel Cáceres R. I., Rocca M., and Luna M. G. (2024). Laboratory evaluation of the feeding behavior of the generalist predatory mirid bug Tupiocoris cucurbitaceus (Hemiptera: Miridae) for the biological control of Phthorimaea absoluta (Lepidoptera: Gelechiidae). Rev. Fac. Cienc. Agrar. Univ. Nac. Cuyo. 56, 97–108. doi: 10.48162/rev.39.140
Montiel Cáceres R., Salas Gervassio N. G., Minghetti E., Dellapé P., Luna M. G., and Rocca M. (2023). Heteropteran bugs assemblage associated with organic tomato farms: knowledge for pest management. Neotrop. Entomol. 52, 251–262. doi: 10.1007/s13744-022-01007-z
Nogueira B. C. F., Ferreira P. S. F., Coelho L. A., Martins D. S., and Barcellos B. D. (2019). Plant bugs predators (Hemiptera: Heteroptera: Miridae) with references to arthropods and fungi in Brazil. An. Acad. Bras. Ciênc. 91, e20181194. doi: 10.1590/0001-3765201920181194
Perdikis D., Fantinou A., and Lykouressis D. (2011). Enhancing pest control in annual crops by conservation of predatory Heteroptera. Biol. Control. 59, 13–21. doi: 10.1016/j.biocontrol.2011.03.014
Perdikis D., Kapaxidi E., and Papadoulis G. (2008). Biological control of insect and mite pests in greenhouse solanaceous crops. Europ. J. Plant Sc. Biotech. 2, 125–144.
Perdikis D. and Lykouressis D. (2000). Effects of various items, host plants, and temperatures on the development and survival of Macrolophus pygmaeus Rambur (Hemiptera: Miridae). Biol. Control. 17, 55–60. doi: 10.1006/bcon.1999.0774
Pérez-Hedo M., Riahi C., and Urbaneja A. (2021). Use of zoophytophagous mirid bugs in horticultural crops: Current challenges and future perspectives. Pest Manage. Sci. 77, 33–42. doi: 10.1002/ps.6043
Polack L. A., López S. N., Silvestre C., Viscarret M., Andorno A., del Pino M., et al. (2017). Control biológico en tomate con el mírido Tupiocoris cucurbitaceus (Buenos Aires: INTA). Available online at: https://inta.gob.ar/documentos/control-biologico-en-tomate-con-el-mirido-tupiocoris-cucurbitaceus.
R Core Team (2024). R: a language and environment for statistical computing (Vienna, Austria: R Foundation for Statistical Computing). Available online at: https://www.R-project.org/.
Robertson G. P. and Swinton S. M. (2005). Reconciling agricultural productivity and environmental integrity: a grand challenge for agriculture. Front. Ecol. Environ. 3, 38–46. doi: 10.1890/1540-9295(2005)003[0038:RAPAEI]2.0.CO;2
Salas Gervassio N. G., Luna M. G., Lee S., Salvo A., and Sánchez N. E. (2016). Trophic web associated with the South American tomato moth Tuta absoluta: implications for its conservation biological control in Argentina. Agric. For. Entomol. 18, 137–144. doi: 10.1111/afe.12146
Secretaría de Agricultura, Ganadería y Pesca, Argentina (2023). Producción de Tomate en Argentina. Available online at: https://www.Argentina.gob.ar/agricultura/subsecretaria-agricultura/informes-sectoriales-de-hortalizas (Accessed August 26, 2025).
Silva D. B., Bueno V. H. P., Montes F. C., and van Lenteren J. C. (2016). Population growth of three mirid predatory bugs feeding on eggs and larvae of Tuta absoluta on tomato. BioControl. 61, 545–553. doi: 10.1007/s10526-016-9736-1
Snyder W. E. (2019). Give predators a complement: Conserving natural enemy biodiversity to improve biocontrol. Biol. Control. 135, 73–82. doi: 10.1016/j.biocontrol.2019.04.017
Urbaneja A., González-Cabrera J., Arnó J., and Gabarra R. (2012). Prospects for the biological control of Tuta absoluta in tomatoes of the Mediterranean basin. Pest Manage. Sci. 68, 1215–1222. doi: 10.1002/ps.3344
Urbaneja A., Montón H., and Mollá O. (2009). Suitability of the tomato borer Tuta absoluta as prey for Macrolophus pygmaeus and Nesidiocoris tenuis. J. Appl. Entomol. 133, 292–296. doi: 10.1016/j.biocontrol.2019.04.017
Vandekerkhove B. and De Clercq P. (2010). Pollen as an alternative or supplementary food for the mirid predator Macrolophus pygmaeus. Biol. Control. 53, 238–242. doi: 10.1016/j.biocontrol.2010.01.005
van Driesche R. G., Hoddle M. S., and Center T. D. (2008). Control de plagas y malezas por enemigos naturales (USDA Forest Service, FHTET). Available online at: http://www.fs.fed.us/foresthealth/technology/pdfs/VANDRIESCHE_CONTROL_Y_PLAGAS_WEB.pdf.
van Lenteren J. C., Bueno V. H. P., Burgio G., Lanzoni A., Montes F. C., Silva D. B., et al. (2012). The state of commercial augmentative biological control: plenty of natural enemies, but a frustrating lack of uptake. BioControl. 57, 1–20. doi: 10.1007/s10526-011-9395-1
van Lenteren J. C., Bueno V. H. P., Burgio G., Lanzoni A., Montes F. C., Silva D. B., et al. (2019). Pest kill rate as aggregate evaluation criterion to rank biological control agents: a case study with Neotropical predators of Tuta absoluta on tomato. Bull. Entomol. Res. 109, 812–820. doi: 10.1017/S0007485319000130
van Lenteren J. C., Alomar O., Ravensberg W. J., and Urbaneja A. (2020). “Biological control agents for control of pests in greenhouses,” in Integrated Pest and Disease Management in Greenhouse Crops, 2nd edition. Ed. Gullino M. L., et al (Springer).
van Lenteren J. C., Bolckmans K., Köhl J., Ravensberg W. J., and Urbaneja A. (2018). Biological control using invertebrates and microorganisms: plenty of new opportunities. BioControl. 63, 39–59. doi: 10.1007/s10526-017-9801-4
van Lenteren J. C., Hemerik K., Lins J. C., and Bueno B. H. P. (2016). Functional responses of three neotropical mirid predators to eggs of Tuta absoluta on tomato. Insects. 7, 34. doi: 10.3390/insects7030034
Varshney R. and Budhlakoti N. (2025). Evaluation of the potential of predatory mirid bug, Dortus primarius Distant as biocontrol agent of Tuta absoluta Meyrick. J. Plant. Dis. Prot. 132, 6. doi: 10.1007/s41348-024-00997-95
Keywords: predation activity, biological control, natural enemy, tomato, Hemiptera, Miridae
Citation: Sola SA, Rocca M, Salas Gervassio NG, Peñalba JN, Montiel Cáceres RI and Luna MG (2025) Campyloneuropsis cincticornis as a potential biological control agent of the South American tomato pinworm Phthorimaea absoluta: predation activity, oviposition and relationship with solanaceous host plants. Front. Agron. 7:1635654. doi: 10.3389/fagro.2025.1635654
Received: 26 May 2025; Accepted: 25 September 2025;
Published: 17 October 2025.
Edited by:
Murray B. Isman, University of British Columbia, CanadaReviewed by:
Luis F Aristizabal, Consultant, Kailua-Kona, HI, United StatesAngeliki Paspati, Hellenic Agricultural Organization DEMETER, Greece
Srivignesh Sundaresan, Central University of Tamil Nadu, India
Copyright © 2025 Sola, Rocca, Salas Gervassio, Peñalba, Montiel Cáceres and Luna. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Margarita Rocca, bXJvY2NhQGNlcGF2ZS5lZHUuYXI=; Nadia G. Salas Gervassio, bmFkaWFzYWxhc0BjZXBhdmUuZWR1LmFy