 Fausto Freire Carrera1*
Fausto Freire Carrera1* Manuel Carrillo-Zenteno2,3Roger Pincay-Ganchozo2Jair Rendón-Holguín4
Manuel Carrillo-Zenteno2,3Roger Pincay-Ganchozo2Jair Rendón-Holguín4 Ricardo Luna-Murillo4Alberto Vecilla-Nicola2Víctor Reynel5
Ricardo Luna-Murillo4Alberto Vecilla-Nicola2Víctor Reynel5 Wilmer Tezara5,6*
Wilmer Tezara5,6*- 1Equinocial Technical University (UTE), Quito, Ecuador
- 2Department of Soil and Water Management, Soil and Water Management, National Institute of Agricultural Research, Mocache, Ecuador
- 3Quevedo State Technical University, Quevedo, Los Ríos, Ecuador
- 4Technical University of Cotopaxi, La Maná, Ecuador
- 5Luis Vargas Torres Technical University, Esmeraldas, Ecuador
- 6Tropical Botany Center, Institute of Experimental Biology, Central University of Venezuela, Caracas, Venezuela
Introduction: Cocoa plants tend to accumulate significant amounts of cadmium (Cd) in their beans and derived products due to their high affinity for Cd, which can pose challenges for commercialization. Research on the variations in Cd absorption capability among cocoa genotypes is still limited.
Aims: This study assessed physiological traits, soil pH, soil electrical conductivity (EC), macronutrient concentrations, Cd bioaccumulation, and biomass distribution in seven cocoa genotypes (CCN-51, EET-103, IMC-67, POUND-12, EET-399, EET-95 and EET-400) grown in greenhouse.
Methods: The genotypes were arranged in a randomized block design with three replicates, assessing variables such as chlorophyll content, leaf gas exchange, pH, soil EC, macronutrient concentration, Cd uptake and transfer, Cd bioaccumulation, and dry root and shoot biomass.
Results and discussion: Results showed no significant differences in chlorophyll content and gas exchange between genotypes, although EET-103 exhibited a higher transpiration rate. A significant reduction in gas exchange occurred 80 days after sowing, associated with decreases in chlorophyll content and stomatal conductance. The soil displayed a strongly acidic pH (≤5.5) and high EC (6 dS m-1) across all genotypes, with no significant inter-genotype differences observed. Cadmium accumulation was nine times higher in the shoot than in the roots, with EET-95 showing a reducedCd uptake capacity, and EET-399 demonstrating limited Cd transfer from root toshoot.
Conclusion: The genotype with the highest Cd absorption and shoot accumulation was CCN-51. Conversely, POUND-12 had the lowest Cd levels in both shoot and root, suggesting its potential as a reference genotype to limit Cd entry to aboveground tissues, especially in soils with elevated Cd concentrations.
Introduction
Cadmium (Cd) is a trace element found in the earth’s crust and is considered a natural pollutant in agricultural soils. In the aqueous phase of the soil, Cd is easily absorbed by plants and bioaccumulated in different tissues, which makes it easier for it to enter the food chain (Zhao et al., 2020). The soil-to-plant transfer rate of Cd in cocoa cultivation is greater than 1 (Oliva et al., 2020), which leads to bean Cd concentration above 10 mg kg⁻¹, which is a serious concern for chocolate production (Argüello et al., 2019). Consumed Cd is harmful is known to cause serious health issues, such as cardiovascular disease, bone disorders, renal dysfunction, and DNA damage and, consequently, cause cancer (Davidova et al., 2024).
The European Union (EU) has set a maximum permissible limit of 0.80 mg kg⁻¹ of Cd in cocoa by-products to reduce risks to its citizens (Florida Rofner, 2021). However, countries like the Russian Federation, New Zealand, and Australia have established more stringent restrictions of 0.50 mg kg⁻¹ of Cd in chocolate products (Meter et al., 2019). Currently, in Ecuador, Peru, and Colombia there is no set limit for Cd in cocoa beans, although high levels have been found in cocoa derivatives (Argüello et al., 2019; Bravo et al., 2014; Chávez et al., 2015; Florida et al., 2018; Mite et al., 2010). Ecuador’s El Oro, Manabí, and Esmeraldas provinces are the most affected by high levels of Cd contamination in cocoa-growing soils, reaching critical values ≥ 2 mg kg⁻¹ (Argüello et al., 2019; Mite et al., 2010). Ecuador’s coastal regions rely heavily on cocoa for high-value chocolate production, but increased Cd contents could hinder global trade, especially under EU rules (Morales-Rodriguez et al., 2025).
Cocoa plants are affected by heavy metals like Cd, which disrupt various biological functions such as metabolism, leaf gas exchange, chlorophyll synthesis, cell division, electron transfer rate, and photochemical efficiency in the light reactions of photosynthesis (Haider et al., 2021; Borjas-Ventura et al., 2022; Vera Pinargote et al., 2025). Plants lacking Cd tolerance experience higher toxicity, leading to decreased leaf water content, reduced stomatal conductance (gs), and increased reactive oxygen species production, necrosis, and cell damage. CCN-51 seedlings exposed to different Cd concentration showed altered mineral nutrient absorption, chlorophyll content, photosynthetic apparatus damage and reduced quantum yield, and the expression of the psbA gene was also disrupted, biomembrane rupture occurring in root and leaf cells (Pereira de Araújo et al., 2017).
It has been recently reported that the cultivation of the BN-34 genotype grafted onto the CCN-51 rootstock shows a high tolerance to Cd, which was associated with a high activity of ascorbate peroxidase, a protein that removes free radicals from plant cells, suggesting that some combinations of cocoa scions and rootstocks may be used to mitigate Cd toxicity (Almeida et al., 2023). Also, the EET-62 genotype showed strong tolerance to Cd content at 6 and 12 mg kg⁻¹; while PA-46 and IMC-67 genotypes had lower Cd accumulation, suggesting their potential use to limit Cd buildup in cocoa tissues (Galvis et al., 2023). Juvenile cocoa plants grown in soils with high Cd levels or combined Mn + Cd levels exhibit impaired photosynthesis, oxidative stress, metabolic alterations, and increased Cd absorption, transport, and accumulation in roots and leaves (Barroso et al., 2023).
Genetic differences in Cd tolerance were also noted among cocoa rootstocks, with the EET-103 genotype exhibiting lower Cd extraction and accumulation capacity compared to CCN-51 (Reyes-Perez et al., 2023a). Genetic variations in cocoa hybrids during seedling growth exhibit differences in Cd partitioning, biomass accumulation, and photosynthetic efficiency, suggesting that cocoa responses to Cd stress are primarily determined by genetic variations (Borjas-Ventura et al., 2022; Vera Pinargote et al., 2025). Previous research findings highlight the role played by genetic enhancement programs in obtaining genotypes that accumulate less Cd and achieve better physiological performance, growth, and development of cocoa plants. Agronomic strategies aim to reduce Cd transfer rates by applying organic and inorganic amendments to precipitate or adsorb Cd, decreasing its bioavailability in soil solution, and leveraging interspecific variations in Cd phytoextraction and accumulation among cocoa genotypes (Meter et al., 2019; Engbersen et al., 2019).
However, limited documentation exists on the Cd absorption capacity of commercial cocoa genotypes from Ecuador. We hypothesize that: 1. The genotypic variation among the cocoa genotypes studied will result in significant differences in physiological and agronomic characteristics, and 2. there will be significant differences among the genotypes of cocoa in Cd absorption, transfer, and bioaccumulation. Therefore, this study aimed to evaluate the physiological characteristics (SPAD values, gas exchange, macronutrient concentrations in the shoot, and Cd bioaccumulation) and biomass distribution in seedlings of seven cocoa genotypes (CCN-51, EET-103, IMC-67, POUND-12, EET-399, EET-95, and EET-400) cultivated in greenhouse conditions.
Materials and methods
Study location
This research was conducted in the greenhouse and laboratory of the Soil and Water Management Department at the Pichilingue Tropical Experimental Station (EETP), part of the National Institute of Agricultural Research (INIAP). The station is located at 79°27′W longitude and 1°06′S latitude, at an altitude of 75 meters above sea level.
Genotypes
The study included the following cocoa genotypes: IMC-67 and POUND-12 (from the Iquitos genetic group), EET-399 and EET-400 (Curaray group), EET-103 and EET-95 (National group), and CCN-51 (CCN-51 gentotype, a hybrid of IMC-67 × ICS-95) (Motamayor et al., 2008; Morillo et al., 2023; Thomas et al., 2024). The first four genotypes are resistant to machete sickness (Ceratocystis cacaofunesta) and are recommended by INIAP for use as rootstocks (Suárez Capello et al., 1993; Vera Barahona et al., 1984); whereas, EET 103 is tolerant to C. cacaofunesta and recommended for cultivation in the province of Manabí. Additionally, EET-95 is tolerant to C. cacaofunesta (Quiroz Vera, 2000). Although the CCN-51 clone is susceptible to this disease, it remains the most widely used rootstock by propagators and was therefore selected as the control for this study.
Physical and chemical characteristics of the soil
The clay loam soil had the following chemical characteristics: ammonium (NH4;): 22 mg kg⁻¹; phosphorus (P): 23 mg kg⁻¹; potassium (K): 0.49 meq 100 mL⁻¹; calcium (Ca): 19 meq 100 mL⁻¹; magnesium (Mg): 2.8 meq 100 mL⁻¹; sulfur (S): 6 mg kg⁻¹; zinc (Zn): 8.2 mg kg⁻¹; Cd: 1.53 mg kg⁻¹; pH: 6.2; and electrical conductivity: 0.31 dS m⁻¹.
Experimental design
The study involved seven cocoa genotypes (treatments) arranged in a randomized block design with three replicates. Each experimental unit consisted of four plants, resulting in a total of 84 plants. The experiment lasted 120 days, comprising 90 days of plant growth, management, and evaluation under greenhouse conditions, followed by 30 days of laboratory analyses.
Experimental management
Soil for the experiment was collected from Río Negro Parish, Santa Rosa canton, El Oro province. It was dried in a greenhouse, disaggregated using a glass roller, and sieved through a 2 mm mesh. Polyethylene bags (5 × 8”) were filled with 700 g of prepared soil.
The plants were propagated from seeds obtained through assisted pollination to ensure genetic purity. Ripe fruits were harvested, and seeds were removed, cleaned of their testa, and soaked in deionized water for 24 hours. Seeds were then planted individually in pots, positioned vertically with the narrower tip facing upward. Before planting, soil moisture was adjusted to field capacity using deionized water, which was maintained through periodic irrigation.
The seedlings were grown in a greenhouse covered with a 65% polyshade screen, providing an average light intensity of 210 ± 30 μmol m⁻² s⁻¹ for 12 hours of natural light daily. Greenhouse temperatures ranged from 23.3 to 33.5 °C, with relative humidity levels between 52% and 82%.
Fertilization was based on soil analysis and the nutritional requirements of cocoa during the nursery stage, tailored for containers with 700 g of soil (Table 1). Nutrients were applied as follows: Nitrogen (N) and S were divided into three applications at 21, 42, and 63 days after sowing (DAS). K, Mg, and Ca were applied in two fractions at 42 and 63 DAS, while P was applied as a single dose at planting. The fertilizers used were Urea (46% N), diammonium phosphate (18% N, 46% P), potassium chloride (60% K), ammonium sulfate (24% S, 21% N), magnesium sulfate (27% MgO, 16% S), and calcium nitrate (15% N, 26% CaO). All fertilizers, except phosphorus, were dissolved in water and applied as a soil drench at 20 mL per plant.
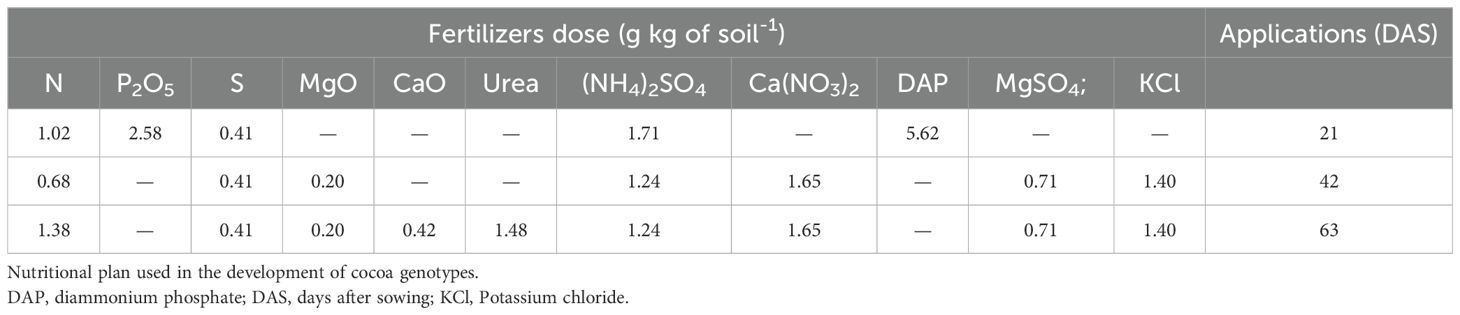
Table 1. The nutritional requirements of cocoa determined by soil analysis and fertilization at the nursery stage are shown.
Variables evaluated
pH and EC of the soil
Soil samples were collected individually for each plant after removing the casings. Each sample was weighed (20 g) on an analytical balance (A&D Weighing, model HR200, Japan) and placed into a plastic cup. Then, 50 mL of deionized water was added to create a 1:2.5 P:V ratio. The mixture was stirred for 5 minutes and left to settle for 1 hour (Henríquez et al., 1998). pH readings were recorded using a potentiometer (HACH model SensION™ MM340, Germany), and EC was measured with a conductivity meter (HACH model SensION™ EC71, Germany).
Chlorophyll index
The chlorophyll content was assessed using a SPAD-502 Plus chlorophyll meter (Konica Minolta, Inc., Tokyo, Japan). SPAD values are directly proportional to the total chlorophyll content in leaves (da Cunha et al., 2015). Measurements were taken every two weeks from 40 to 80 DAS between 9:00 a.m. and 1:00 p.m., on the same leaves used for gas exchange measurements.
Leaf gas exchange
Gas exchange parameters were measured at 40 and 80 DAS between 9:00 a.m. and 1:00 p.m. on six plants per treatment (n = 6). Measurements were performed with an infrared gas analyzer (CIRAS 2, PP Systems, Hitchin, UK) to determine gs, A, E, water use efficiency (WUE = A/E), and intercellular CO2 concentration (Ci). These measurements were taken from the mid-section of leaf No. 4, considered a fully expanded, healthy adult leaf. Experimental conditions included: ambient CO2 concentration (Ca) of 415 ± 10 µmol mol−1, 21% O2, temperature of 28 ± 1 °C, a photon flux density (PFD) of 1000 µmol m-2 s−1 to saturate photosynthesis, and a leaf-air water vapor gradient (DPV) of 1.0–1.5 kPa.
Dried shoot and root biomass
Plants harvested at 90 DAS were rinsed with tap water and sequentially immersed in hydrochloric acid (3%), distilled water, magnesium sulfate (0.001 mol L⁻¹), and deionized water. The plants were then sectioned into shoot (leaves and stems) and roots. Each section was placed into labeled kraft paper sleeves and dried in an oven (Memmert, GmbH 450, Germany) at 65 °C for 72 hours. The dry biomass was weighed on an analytical balance (A&D Weighing, model HR200, Japan).
Macronutrient concentration in the shoot
N concentration was determined using the Kjeldahl method (KjeltecTM 8400 TecatorTM Line Foss, Jinan, China) (Carrillo Zenteno et al., 2019). K, Ca, and Mg concentrations were analyzed using an atomic absorption spectrophotometer (AA-6800, Kyoto, Japan). S and P concentrations were quantified via the colorimetric method (Spercord 210 Plus, Jena, Germany) (Remache et al., 2017).
Determination of Cd in tissues
Dried plant tissues were ground using a Willey-type mill (IKA, model A11 basic, USA). To prevent contamination, the mill was cleaned with absorbent towels, diluted aqua regia (10%), and deionized water before each use. Cd was extracted through nitric-perchloric acid mineralization (8 mL HNO3 and 2 mL HClO4;; Carrillo, 2003). Cd concentrations were measured with an atomic absorption spectrophotometer (Perkin Elmer, model AAnalyst 800, Japan) equipped with a graphite furnace, at a wavelength (λ) of 228.8 nm.
Cd content in root and shoots
The Cd content in roots and shoots was calculated using the following formula:
Cd extraction capacity
The efficiencies of Cd absorption (1) and translocation (2) were determined using equations described by Wang et al. (2007):
1. Absorption efficiency (μg g-1) = .
2. Translocation efficiency = .
Statistical analysis
Data were tested for normality and homogeneity of variance using the Shapiro-Wilk and Bartlett tests, respectively. An analysis of variance (ANOVA) followed by a Tukey’s test was conducted at a 0.05 significance level. Statistical analyses were performed using Minitab version 19, and graphs were created using SigmaPlot 12 (Systat Software, San Jose, CA, USA).
Results
There were no significant differences in soil pH and EC among the genotypes. However, all genotypes exhibited strongly acidic soil pH values (≤5.5) and high EC levels (6 dS m⁻¹) (Figure 1).
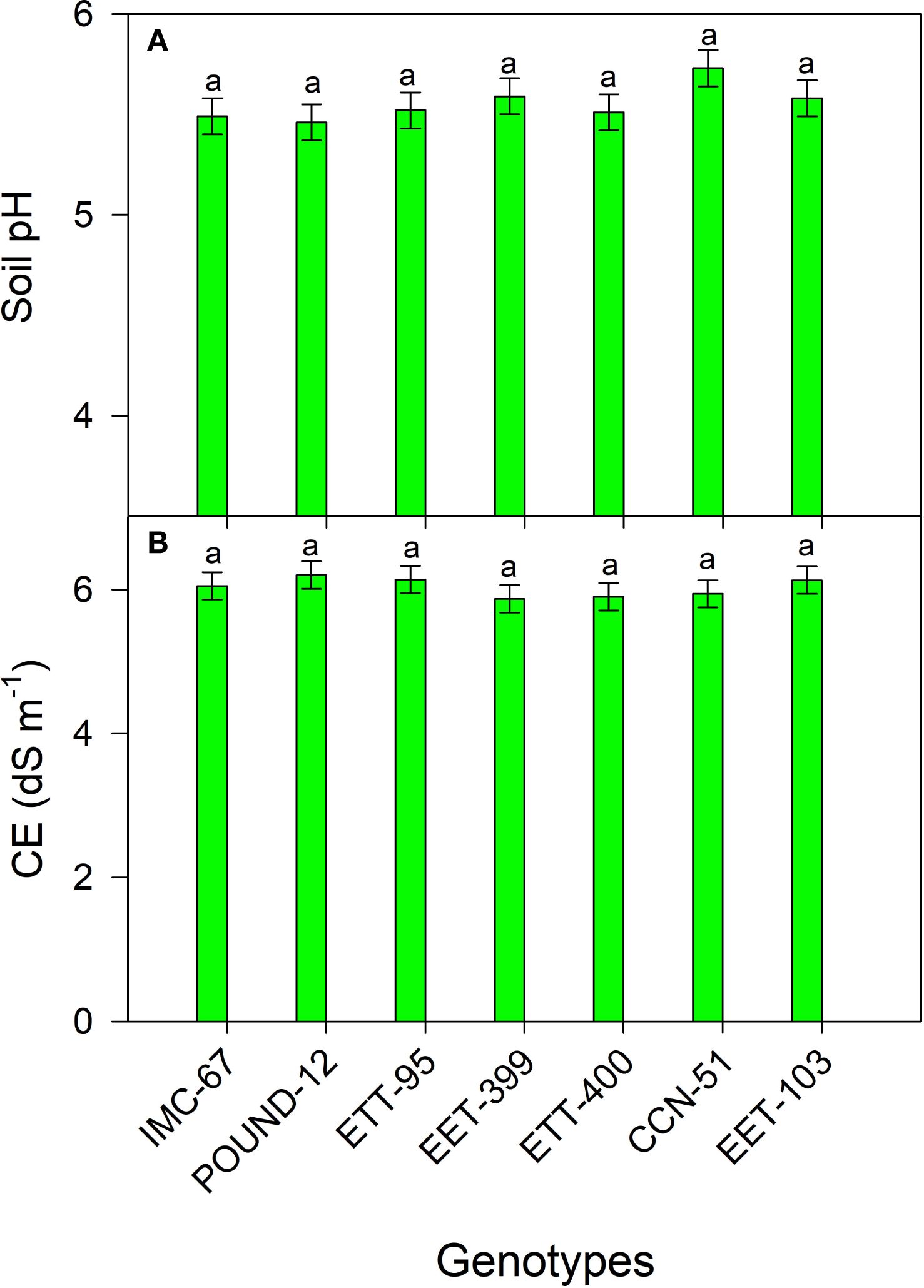
Figure 1. pH (A) and EC (B) values of the soil in which the seven cocoa genotypes were grown under greenhouse conditions. The vertical bars represent the mean ± SE (n = 6). Different letters above the bars indicate significant differences between treatments.
Chlorophyll concentrations, measured at 40, 55, and 80 DAS, showed no statistically significant differences between genotypes (Figure 2). Nevertheless, a significant reduction in SPAD units, amounting to 31.5%, was observed at 80 DAS.ç
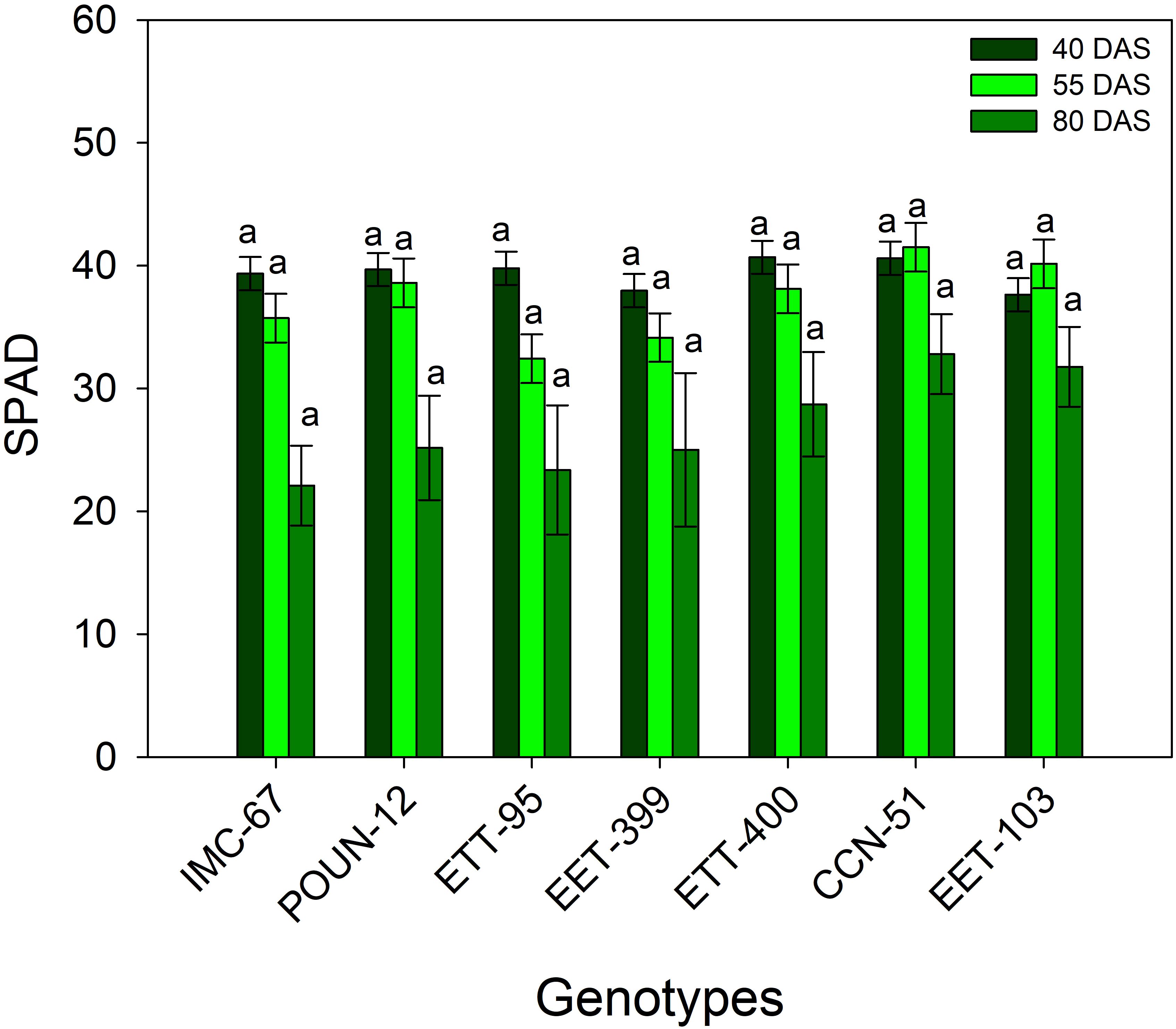
Figure 2. Chlorophyll concentration was evaluated in the seven cocoa genotypes used as standards at 40, 55, and 80 DAS. The vertical bars represent the mean ± SE (n = 6). Different letters above the bars indicate significant differences between treatments, as determined by the Tukey test (p < 0.05).
In terms of leaf gas exchange, no significant differences were found between genotypes at 40 and 80 DAS, except for transpiration, which was statistically significant at 80 DAS. The EET-103 genotype showed the highest E at 0.39 mmol m⁻² s⁻¹ (Figure 3). At 80 DAS, a reduction of 94% in A, 37% in E, 45% in gs, and 71% in WUE was observed, while Ci increased by 61%.
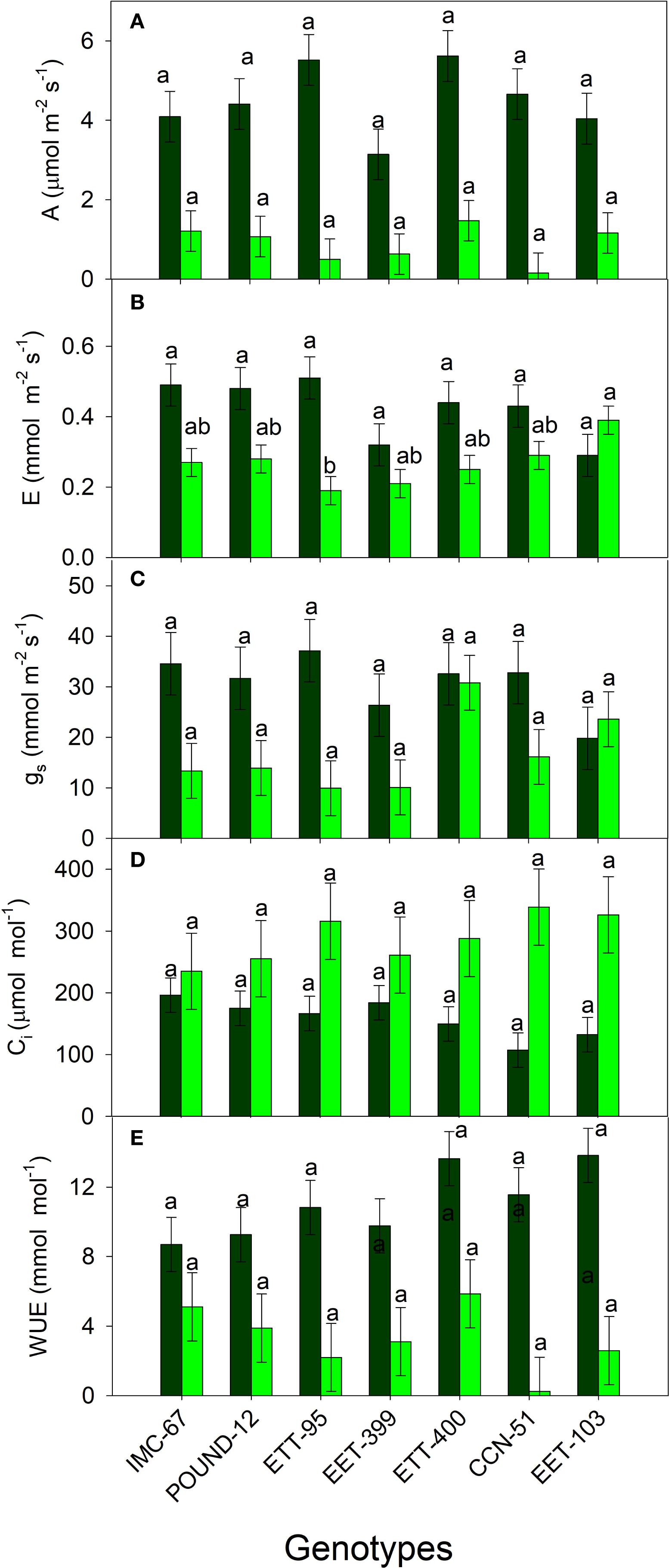
Figure 3. Photosynthesis rate (A), transpiration (B), stomatal conductance (C), intercellular CO2 concentration (D) and water use efficiency (E) measured in the leaves of seven cocoa genotypes under greenhouse conditions at 40 DAS (dark green bars) and 80 DAS (bright green bars). The vertical bars represent the mean ± SE (n = 6). Different letters above the bars indicate significant differences between treatments, as determined by the Tukey test (p < 0.05).
There were no significant differences between cocoa genotypes in the production of dry biomass in shoots and roots (Figure 4). However, for aerial dry matter, IMC-67 produced 5% more biomass than CCN-51, while the clones POUND-12, EET-103, EET-399, EET-95, and EET-400 produced 36%, 27%, 22%, 25%, and 19% less biomass than CCN-51, respectively.
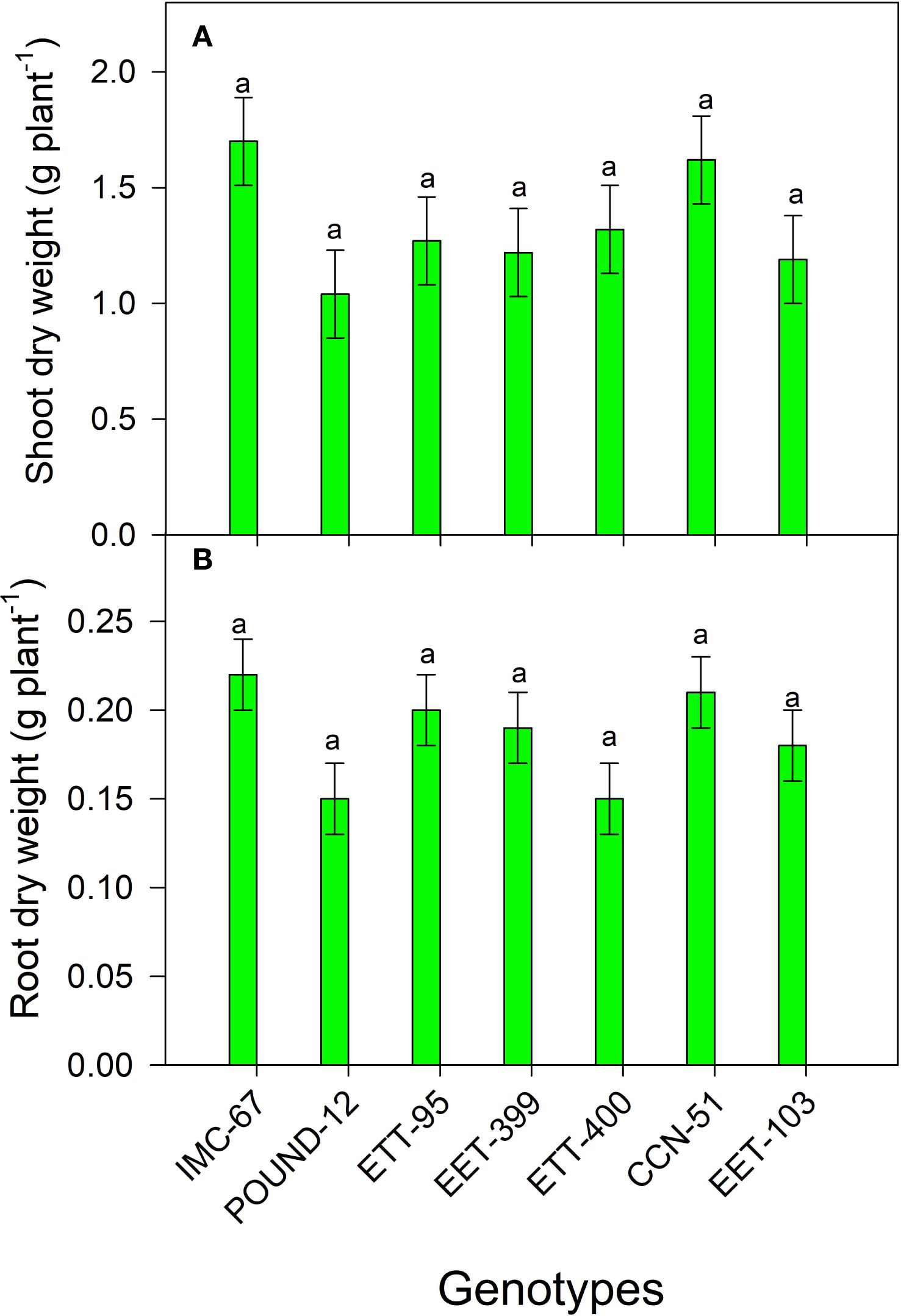
Figure 4. Production of dry biomass in shoots (A) and roots (B) of seven cocoa genotypes grown under greenhouse conditions. The vertical bars represent the mean ± SE (n = 6). Different letters above the bars indicate significant differences between treatments based on Tukey’s test (p < 0.05).
Similarly, in root dry weight, IMC-67 exhibited 5% higher biomass compared to CCN-51. In contrast, the POUND-12 and EET-400 genotypes produced 29% less biomass than CCN-51, followed by EET-103 (14% less), EET-399 (10% less), and EET-95 (5% less).
Macronutrient concentrations in the shoot (Figure 5) showed statistically significant differences only for N, P, and S. Among the genotypes, the highest concentrations were observed in CCN-51 for N (6.1 dag kg⁻¹), EET-399 for P (0.54 dag kg⁻¹), and IMC-67 for S (0.61 dag kg⁻¹).
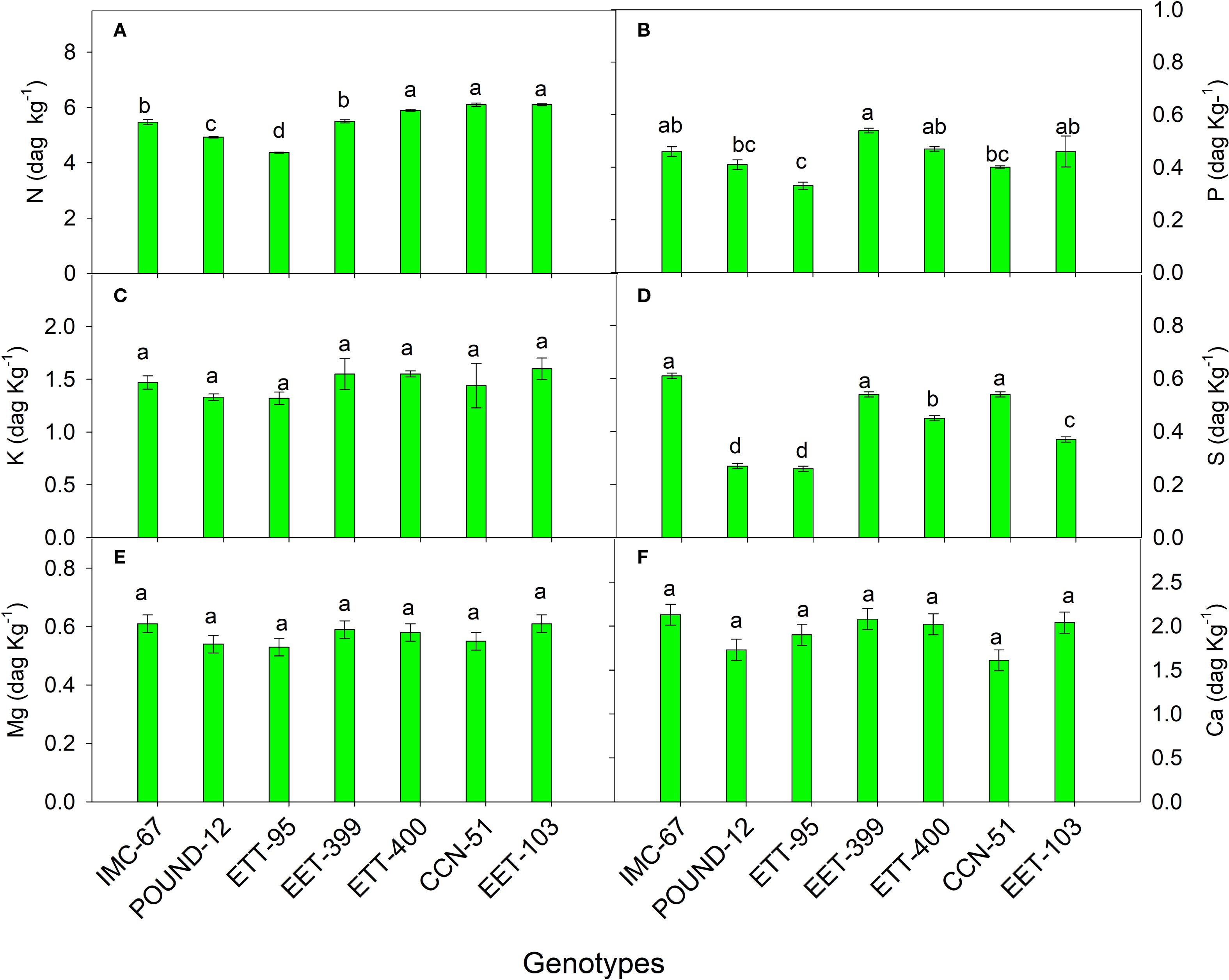
Figure 5. Concentrations of nitrogen (A), phosphorus (B), potassium (C), sulfur (D), magnesium (E), and calcium (F) in the stems of seven cocoa genotypes. The vertical bars represent the mean ± SE (n = 6). Different letters above the bars indicate significant differences between treatments after the Tukey test (p < 0.05).
Cocoa genotypes did not exhibit significant differences in Cd absorption and translocation efficiencies or Cd content in shoots, except for root Cd accumulation, which was statistically significant (Figure 6). Among the genotypes, EET-95, POUND-12, and EET-399 absorbed 26%, 24%, and 22% less Cd, respectively, compared to CCN-51, the control and the genotype with the highest Cd absorption (Figure 6A). Cadmium uptake efficiency ranged from 68.7 to 79.7 µg Cd g-1 of dry root matter, with the lowest value found in EET-95 and the highest average in CCN-51, respectively.
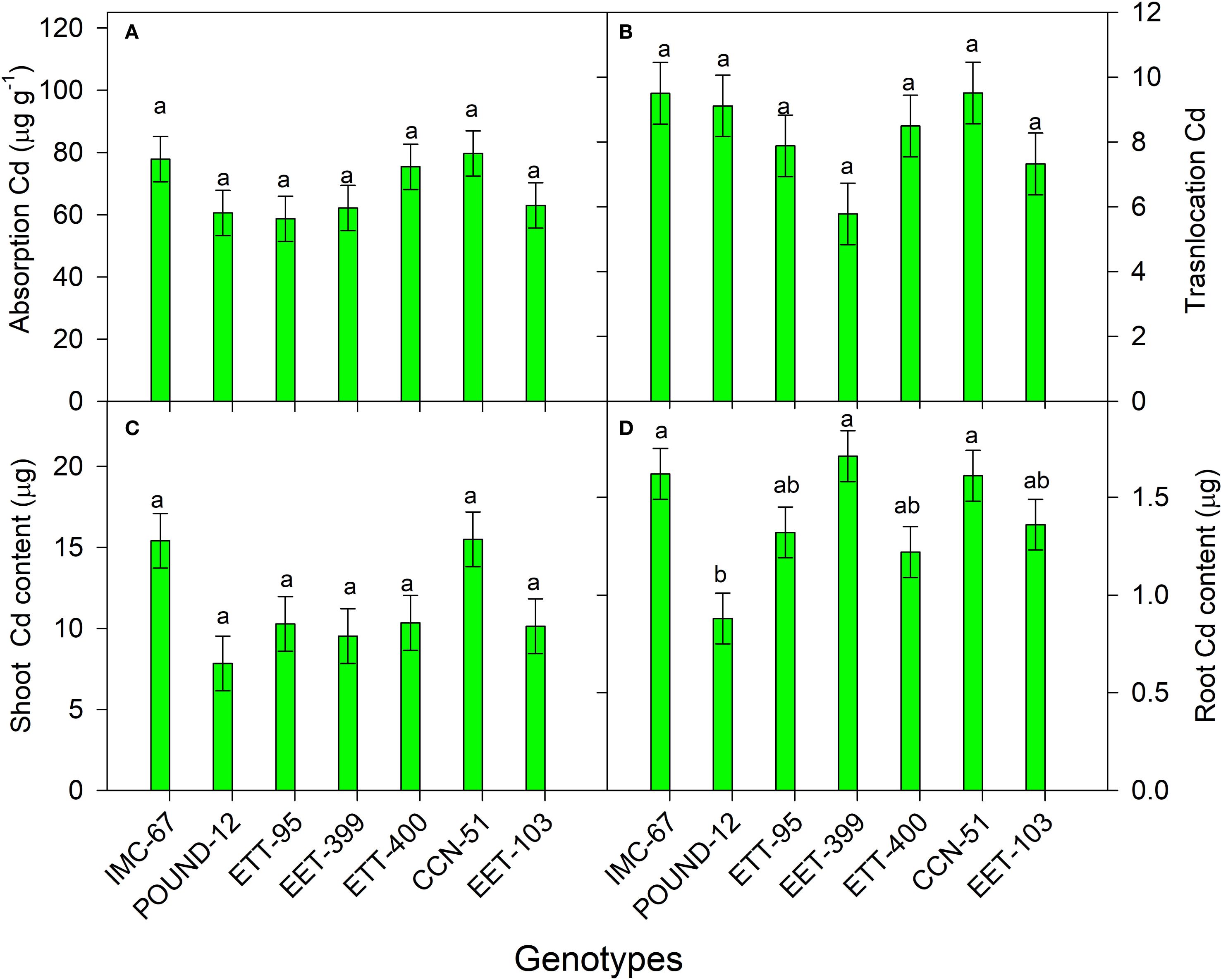
Figure 6. Absorption (A) and translocation of Cd (B), Cd contents in the shoot (C) and root (D), of seven cocoa genotypes grown under greenhouse conditions. The vertical bars represent the mean ± SE (n = 6). Different letters above the bars indicate significant differences between treatments after the Tukey test (p < 0.05).
In Cd translocation, values ranged from 5.8 to 9.5, with the lowest average obtained in EET-399 and the highest in CCN-51, respectively. In terms of Cd translocation efficiency from roots to shoots, EET-399, EET-103, and EET-95 exhibited 39%, 23%, and 17% lower translocation rates, respectively, than CCN-51 (Figure 6B). Additionally, the POUND-12 genotype bioaccumulated 49% less Cd in the shoot compared to CCN-51, followed by EET-399 (39%) and EET-400 (32%).
Discussion
The study revealed no significant differences among the evaluated cocoa genotypes in physiological variables or biomass distribution, thereby rejecting Hypothesis 1. In contrast, significant genetic differences were evident in Cd absorption efficiency, translocation, and content, supporting Hypothesis 2. Notably, POUND-12 genotype producing the least biomass and exhibited reduced Cd accumulation in shoots. This genetic effect in Cd bioaccumulation aligns with findings by Galvis et al. (2023) and Reyes-Pérez et al. (2023a), who identified similar trends in genotypes with lower biomass production, such as PA-46 and EET-103, respectively.
At the end of the experiment, chlorophyll reduction correlated with decreased leaf gas exchange. This was likely due to salt stress induced by high soil salinity (6 dS m⁻¹). A similar response was reported by Reyes-Pérez et al. (2023a) under high EC conditions (5 dS m⁻¹), which led to reduced chlorophyll (90 DAS), and gas exchange (80 DAS), along with reddish-brown leaf margins. This leaf margin color was also observed in our study. Comparable reductions in A, gs, E have also been observed in juvenile cocoa plants under water stress and Cd stress (Ortiz-Álvarez et al., 2023; Pereira de Araújo et al., 2017).
Among physiological parameters, only E at 80 DAS showed significant genetic variation, with EET-103 exhibiting the highest transpiration rate (0.39 mmol m⁻² s⁻¹). In fact, most cocoa genotypes showed low E due to small gs, probably in response to the salt stress experienced by the cocoa seedlings. Higher transpiration values were reported in cocoa genotypes by Reyes-Pérez et al. (2023a). The physiological performance of the genotypes that were evaluated in this study is not documented in the literature under the same conditions. However, the greater transpiration in IMC-67 observed by Ortiz-Álvarez et al. (2023) under water stress, linked to higher Cd translocation, was not corroborated under the present study. The physiological traits of 18 cocoa genotypes under irrigation effects, found higher gas exchange performance (A, gs and E) in genotypes such as CP-41, CP-43, and CCN-51 (Araújo et al., 2024).
Dry biomass is a key indicator of plant growth, directly linked to water content and the synthesis of primary metabolites (Huang et al., 2020). In this study, the IMC-67 and CCN-51 genotypes demonstrated the highest accumulation of dry biomass in both shoot and root compared to the other genotypes evaluated (EET-95, EET-399, EET-103, EET-400, and POUND-12). These findings are consistent with those reported by Reyes-Pérez et al. (2023a, 2023b), who observed that CCN-51 produces greater biomass than domestic cocoa genotypes like EET-103 and EET-801.
However, contrasting results were noted in the work of Borjas-Ventura et al. (2022), where POUND-7 outperformed CCN-51 in shoot biomass production. This variation highlights the influence of environmental adaptability and genetic plasticity on biomass outcomes. Studies by Héctor Ardisana et al (2018) and Palacio-López and Rodríguez-López (2007) suggest that biomass variation among genotypes is often driven by their ability to adapt to specific climatic conditions, which, in turn, affects the expression of phenotypic traits such as leaf plasticity.
Although this study was conducted with juvenile plants, the Cd content in the shoot may be related to the findings of Lewis et al. (2018), who observed a 2:1 relationship between Cd concentrations in the leaves (2.15 mg kg−1) and beans with controls (1 mg kg−1) in adult plants of 100 cocoa genotypes from eight genetic groups. This relationship was supported by a significant correlation coefficient (r = 0.61; P < 0.05). Furcal-Beriguete and Torres-Morales (2020) reported a similar pattern, with Cd concentrations of 3.44 mg kg−1 in leaves and 2.31 mg kg−1 in beans, showing a correlation coefficient of r = 0.90. Therefore, regardless of the plant’s age, genetic load, or edaphoclimatic conditions, it is considered that the beans accumulate less than 50% of the Cd found in the leaves.
Ortiz-Álvarez et al. (2023) found that increases in dry matter, biomass output, leaf area or proliferation, and Cd bioaccumulation in terms of content are all linked to Cd accumulation under greenhouse conditions. These authors also showed that when transpiration increases, there is greater Cd accumulation because sap flow carries more Cd to the leaves and shoot (Sterckeman, 2025), but it is diluted in terms of concentration (Ortiz-Álvarez et al., 2023). These explanations are reinforced by the results reported in POUND-12, IMC-67, and CCN-51. The first genotype had lower dry matter and Cd content, and the other two clones were superior in terms of shoot dry matter production and Cd bioaccumulation. In cacao seedlings, was found in CCN-51 had a greater capacity to produce dry matter and absorb nutrients and Cd (Arias-Contreras et al., 2024).
In the present study, there were differences in root Cd bioaccumulation, with the highest accumulation found in the roots of EET-399, IMC-67, and CCN-51 and the lowest allocation in POUND-12. Similarly, significant variations in Cd bioaccumulation were observed in the roots of young plants cultivated in greenhouses; the PA-121 x IMC-67 progeny exhibited the highest accumulation (Fernandez-Paz et al., 2021). Growing in a greenhouse with soil Cd concentrations of 12 mg kg−1, PA-121 was shown to have the maximum Cd allocation in root tissues at 30 and 60 DDS, in comparison to the genotypes ETT-61, EET-62, IMC-60, IMC-67, PA-150, PA-46, SCC-85, and SCC-86 (Galvis et al., 2023).
All cocoa genotypes had Cd translocation > 1, but EET-399, compared to the other clones (EET-103, IMC-67, EET-95, CCN-51, EET-400, and POUND-12) had the lowest translocation. This result is consistent with the higher Cd accumulation in the roots of this genotype, showing that it restricts Cd translocation to the aerial parts by capturing it in the root system. Reyes-Perez et al. (2023a) also detected a similar phenomenon: the EET-103 clone had higher Cd accumulation in the roots and lower Cd translocation to the shoots, compared to CCN-51, which obtained the opposite response. Also, were found a genotypic variations in Cd accumulation in the roots of five fine aroma cocoa genotypes, detecting the highest amounts of Cd (156.75 mg kg⁻¹) and lower translocation to the leaves in the INDES-38 genotype (Meléndez-Mori et al., 2023).
It was also observed that the Cd content in the shoot was nine times higher than that in the root. Similarly, Castro et al. (2015) found the highest accumulations of Cd in the offspring of two cocoa progenies (CCN-10 x SCA-6 and Catongo x Catongo) at the juvenile stage. In contrast, Oliva et al. (2020) reported that in mature trees, the leaves accumulate 123 times more Cd than the roots. However, other studies involving juvenile plants (Calva Jiménez et al., 2022) and adult trees (Llatance et al., 2018) found that the roots accumulated 4.84 and 3.36 times more Cd than the leaves, respectively. These findings demonstrate that Cd accumulation in cocoa plants varies between organs, influenced by age, genotype, and environmental conditions that affect Cd migration from one organ to another.
Of the seven genotypes studied, CCN-51 exhibited the highest absorption and accumulation of Cd in the offspring. In Ecuador, this genotype is widely used as a rootstock, covering 75% of the planted area (Latin American Center for Rural Development (RISMIP), 2023). These findings support the results of Reyes-Pérez et al. (2023a), who reported that the EET-103 genotype accumulates less Cd than CCN-51. Additionally, due to its resistance to machete sickness, EET-103 was recommended as a standard.
The POUND-12 genotype exhibited the lowest Cd content in both shoot and root. Due to its resistance to C. cacaofunesta, it is proposed as a key genetic rootstock strategy to mitigate Cd in soils with high metal concentrations. Additionally, EET-399 and EET-400 are viable alternatives, as they demonstrate low Cd accumulation, resistance to C. cacaofunesta (Suárez Capello et al., 1993), and, according to Jaimez et al. (2021), tolerance to water stress.
In contrast, Arévalo-Hernández et al. (2021) in Peru reported results that differ from the present study, finding that CCN-51, with a Cd concentration of 1.56 mg kg⁻¹, accumulated less Cd in the shoots compared to EET-400 (18.56 mg kg⁻¹). Despite its susceptibility to root diseases, CCN-51 was included in a list of 11 genotypes recommended as rootstocks for Cd mitigation, alongside AYP-22, PAS-105, UGU-126, ICT-1026, ICT-1087, ICT-1189, ICT-1292, PH-17, ICS-39, and TSH-565. Furthermore, Borjas-Ventura et al. (2022) found in the Central Jungle of Peru that CCN-51 bioaccumulates more Cd in the shoots compared to genotypes like ICS-95, ICS-60, VRAE-99, and POUND-7. These latter genotypes were recommended as standard due to their low Cd accumulation and partial resistance to Verticillium (Braga and Silva, 1989; End et al., 2021).
The highest N concentration in the shoot was detected in CCN-51, consistent with the findings of Reyes-Pérez et al. (2023a), who also reported that CCN-51 exhibited the highest N concentration in the shoot compared to the EET-103 genotype. Similarly, under field conditions, Carrillo et al. (2010) found that CCN-51 accumulated more leaf N compared to national genotypes such as EET-95 and EET-103.
Conclusions
Genetic variation was observed in the extraction and accumulation capacity of Cd in cocoa rootstocks. These results suggest that genotypes with limited Cd absorption could serve as an agronomic strategy for producing cocoa beans in soils contaminated with this metal. In leaf gas exchange, significant differences were found only in transpiration at 80 DAS, with the EET-103 genotype showing the highest average at 0.39 mmol m⁻² s⁻¹. Overall, at 80 DAS, a reduction in A, E, gs, and WUE was observed, while Ci increased, which were associated to a decrease in chlorophyll content. In this study, the CCN-51 genotype exhibited greater Cd absorption and bioaccumulation in the shoot, while also showing the highest leaf N concentration. On the other hand, EET-95 showed lower Cd absorption. The POUND-12, EET-400, and EET-399 genotypes, with their low Cd extraction and accumulation capacity and resistance to C. cacaofunesta, are proposed as suitable candidates for use as cocoa rootstocks and for introduction into genetic improvement programs aimed at developing Cd-excluding genotypes. The primary research constraint, however, was the use of cocoa seedlings with limited root volume and a brief exposure to Cd. This could have an impact on the results when extending these findings to mature, fruitful plants in the field. Thus, we think that more research in this area in field settings on plantations is very important.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
FF: Validation, Supervision, Writing – review & editing. MC-Z: Supervision, Methodology, Writing – original draft, Investigation. RP-G: Data curation, Writing – original draft. JR-H: Conceptualization, Investigation, Writing – original draft. RL-M: Conceptualization, Formal Analysis, Writing – review & editing. AV-N: Writing – review & editing, Conceptualization, Investigation. VR: Investigation, Methodology, Writing – review & editing. WT: Software, Methodology, Writing – original draft, Investigation.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. The research was funded by German Society for International Cooperation (GIZ).
Acknowledgments
The authors thank German Society for International Cooperation (GIZ) for providing financial support.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Almeida N. M., Almeida A. F., Almeida, Santos N., Mora-Ocampo I. Y., and Pirovani C. P. (2023). Foliar proteomic profiles in cocoa stem-rootstock combinations tolerant and intolerant to cadmium toxicity. Plant Physiol. Biochem. 203, 107987. doi: 10.1016/j.plaphy.2023.107987
Araújo M. S., Chaves S. F., Pereira G. R., Guimarães M. H., Alves A. K., Dias L. A. S., et al. (2024). Multi-trait selection for nutritional and physiological quality of cacao genotypes in irrigated and non-irrigated environments. Sci. Rep. 14, 6368. doi: 10.1038/s41598-024-56556-7
Arévalo-Hernández C. O., Arévalo-Gardini E., Barraza F., Farfán A., He Z., and Baligar V. C. (2021). Growth and nutritional responses of wild and domesticated cacao genotypes to soil Cd stress. Sci. Total Environ. 763, 144021. doi: 10.1016/j.scitotenv.2020.144021
Argüello D., Chavez E., Lauryssen F., Vanderschueren R., Smolders E., and Montalvo D. (2019). Soil properties and agronomic factors affecting cadmium concentrations in cacao beans: A nationwide survey in Ecuador. Sci. total Environ. 649, 120–127. doi: 10.1016/j.scitotenv.2018.08.292
Arias-Contreras M. A., Checca-Huaman N. R., Arévalo-Gardini E., Arévalo-Hernández C. O., Passamani E. C., and Ramos-Guivar J. A. (2024). Medium scale-up synthesis of nanomaghemite as an inhibitor of cadmium uptake in seedlings of Theobroma cacao L. J. Agric. Food Res. 18, 101295. doi: 10.1016/j.jafr.2024.101295
Barroso J. P., de Almeida A. A. F., do Nascimento J. L., Oliveira B. R. M., Dos Santos I. C., Mangabeira P. A. O., et al. (2023). The damage caused by Cd toxicity to photosynthesis, cellular ultrastructure, antioxidant metabolism, and gene expression in young cacao plants are mitigated by high Mn doses in soil. Environ. Sci. pollut. Res. 30, 115646–115665. doi: 10.1007/s11356-023-30561-1
Borjas-Ventura R., Bello-Medina N., Bello-Amez S., Alvarado-Huamán L., Rabaza-Fernández D., Figueroa L. T. Y., et al. (2022). Differentiated cadmium uptake and its effect on the physiology of six cacao genotypes (Theobroma cacao L.) In San Ramón, Central Peruvian Jungle. Trop. Subtropical Agroecosystems 25, 1–10. doi: 10.56369/tsunes.4000
Braga M. and Silva S. (1989). Resistance of the Cocoa tree (Theobroma cacao L.) to Verticillium dahliae Kleb. Agrotropic(Brazil) 1, 116–121.
Bravo I., Arboleda C., and Martin F. (2014). Effect of the quality of organic matter associated with soil use and management on cadmium retention in high Andean systems of Colombia. Acta Agronómica 63, 164–174. doi: 10.17163/lgr.n34.2021.08
Calva Jiménez C. C., Pinedo Fernández L. V., and Rodríguez Reis C. E. (2022). Reduction of the toxicity of Cd to Theobroma cacao using soil amendments based on commercial charcoal and lime. Toxic 10, 15.
Carrillo Z. M. D. (2003). Characterization of heavy metal forms, their bioavailability and their adsorption and mobility dynamics in soils of Ecuador (Minas Gerais, Brazil: Postgraduate Thesis Federal University of Viçosa), 56 p.
Carrillo M., Recalde M., and Sánchez J. (2010). “Nutrition management of cocoa (Theobroma cacao L.) crops, national type and CCN-51 in the establishment stage,” in XII Ecuadorian Congress of Soil Science. Ecuadorian Society of Soil Science(Santo Domingo, Ecuador), 1–10.
Carrillo Zenteno M. D., Durango Cabanilla W. D., Morales Intriago F. L., Rivadeneira Moreira B. J., and Cargua Chávez J. E. (2019). Variation in macronutrient uptake in hard maize hybrids. ACI Adv. Sci. Eng. 11, 20–31. doi: 10.18272/aci.v11i1.1077
Castro A. V., de Almeida A. A. F., Pirovani C. P., Reis G. S., Almeida N. M., and Mangabeira P. A. (2015). Morphological, biochemical, molecular and ultrastructural changes induced by Cd toxicity in seedlings of Theobroma cacao L. Ecotoxicology Environ. Saf. 115, 174–186. doi: 10.1016/j.ecoenv.2015.02.003
Chávez E., He Z. L., Stoffella P. J., Mylavarapub R. S., Li Y. C., Moyanod B., et al. (2015). Concentration of cadmium in cacao beans and its relationship with soil cadmium in southern Ecuador. Sci. Total Environ. 533, 205–214. doi: 10.1016/j.scitotenv.2015.06.106
da Cunha A. R., Katz I., de Pádua Sousa A., Uribe M., and Andres R. (2015). SPAD index in the growth and development of Lisianthus plants as a function of different nitrogen doses in a protected environment. Idesia (Arica) 33, 97–105. doi: 10.4067/S0718-34292015000200012
Davidova S., Milushev V., and Satchanska G. (2024). The mechanisms of cadmium toxicity in living organisms. Toxics 12, 875. doi: 10.3390/toxics12120875
End M. J., Daymond A. J., and Hadley P. (2021). Technical guidelines for the safe movement of cocoa germplasm, Revised version of FAO/IPGRI Technical Guidelines No. 20 (Fourth Update, 2021). Rome (Italy): Bioversity International
Engbersen N., Gramlich A., López M., Schwarz G., et al. (2019). Cadmium accumulation and allocation in different cacao cultivars. Sci. Total Environ. 678, 660–670. doi: 10.1016/j.scitotenv.2019.05.001
Fernández-Paz J., Cortés A. J., Hernández-Varela C. A., Mejía-de-Tafur M. S., Rodriguez-Medina C., and Baligar V. C.. (2021). Rootstock-mediated genetic variance in cadmium uptake by juvenile cacao (Theobroma cacao L.) genotypes, and its effect on growth and physiology. Front. Plant Sci. 12, 777842. doi: 10.3389/fpls.2021.777842
Florida R., Claudio M., and Gómez B. (2018). The pH and absorption of cadmium in organic cacao almonds (Theobroma cacao L.) in Leoncio Prado, Huánuco, Peru. Folia amazónica 27, 1–8. Available online at: https://bit.ly/3nNNxjx.
Florida Rofner N. (2021). Review of maximum limits of cadmium in cocoa (Theobroma cacao L.). La Gran. Rev. Cien. la Vid 34, 117–130. doi: 10.17163/lgr.n34.2021.08
Furcal-Beriguete P. and Torres-Morales J. (2020). Determination of cadmium concentrations in Theobroma cacao L. plantations in Costa Rica. Rev.Tec. Mar. 33, 122–137. doi: 10.18845/tm.v33i1.5027
Galvis D. A., Jaimes-Suárez Y. Y., Rojas Molina J., Ruiz R., León-Moreno C. E., and Carvalho F. E. L. (2023). Unveiling cacao rootstock-genotypes with potential use in the mitigation of cadmium bioaccumulation. Plants 12, 2941. doi: 10.3390/plants12162941
Haider F. U., Liqun C., Coulter J. A., Cheema S. A., Wu J., Zhang R., et al. (2021). Cadmium toxicity in plants: Impacts and remediation strategies. Ecotoxicology Environ. Saf. 211, 111887. doi: 10.1016/j.ecoenv.2020.111887
Héctor Ardisana E. F., Torres García A., Fosado Tellez O., Alava Alava J., Sacan Pin G. T., and León Aguilar R. (2018). Total chlorophyll content in twelve cocoa clones (Theobroma cacao L.). La Técnica 20), 11–18. doi: 10.33936/la_tecnica.v0i20.1324
Henríquez C., Bertsch F., and Salas R. (1998). Soil fertility: Laboratory manual (San José, CR: Costa Rican Association of Soil Science), 64.
Huang W., Reddy G. V., Li Y., Larsen J. B., and Shi P. (2020). Increase in absolute leaf water content tends to keep pace with that of leaf dry mass—evidence from bamboo plants. Symmetry 12, 1345. doi: 10.3390/sym12081345
Jaimez R., Vásconez G., Sotomayor I., Quijano G., Morante J., Arteaga F., et al. (2021). Physiological and root morphological responses in different combinations of rootstock-scion of cacao to water deficit. Rev. la Facultad Agronomía la Universidad del Zulia 38, 631–651. doi: 10.47280/RevFacAgron(LUZ).v38.n3.09
Latin American Center for Rural Development (RISMIP) (2023). The Cocoa Chain as an example of dynamism in the face of the pandemic. Available online at: https://www.rimisp.org/wp-content/uploads/2023/04/Policy-brief-cacao_Ecuador-.pdf (Accessed May 8, 2025).
Lewis C., Lennon A. M., Eudoxie G., and Umaharan P. (2018). Genetic variation in bioaccumulation and partitioning of cadmium in Theobroma cacao L. Sci. Total Environ. 640, 696–703. doi: 10.1016/j.scitotenv.2018.05.365
Llatance W. O., Gonza C. J., Guzmán W., and Pariente E. (2018). Bioaccumulation of cadmium in cocoa (Theobroma cacao) in the Native Community of Pakun, Peru. Rev. Forestal del Perú 33, 63–75. doi: 10.21704/rfp.v33i1.1156
Meléndez-Mori J., Guerrero-Abad J., Tejada Alvarado J., Ayala-Tocto R., and Oliva M. (2023). Genotypic variation in cadmium uptake and accumulation among fine-aroma cacao genotypes from northern Peru: a model hydroponic culture study. Environ. Pollutants Bioavailability 35, 2287710. doi: 10.1080/26395940.2023.2287710
Meter A., Atkinson R. J., and Laliberte B. (2019). Cadmium in Latin American and Caribbean cocoa: Research analysis and potential solutions for mitigation (Rome (Italy: Bioversity International), 77 p, ISBN: 978-92-9255-136-0.
Mite F., Carrillo M., and Durango W. (2010). Advances in the monitoring of the presence of cadmium in cocoa beans, soils and water in Ecuador (XII Ecuadorian Congress of Soil Sciences).
Morales-Rodriguez W. J., Morante-Carriel J., Carranza-Patino M., Ormaza-Vasquez D., Ayuso-Yuste M. C., and Bernalte-Garcia M. J. (2025). Fermentation with pectin trans-eliminase to reduce cadmium levels in nacional and CCN-51 cocoa bean genotypes. Plants 14, 2553. doi: 10.3390/plants14162553
Morillo E., Buitron J., and Quiroz-Vera J. (2023). Evaluation of Linalool gene expression in highly homozygous national cocoa genotypes from the main traditional cocoa growing areas of Ecuador. Revis Bionatura; 8, (2) 20. doi: 10.21931/RB/2023.08.02.20
Motamayor J. C., Lachenaud P., Wallace da Silva e Motta J., Loor R., Kuhn D. N., Brown J. S., et al. (2008). Geographic and genetic population differentiation of the Amazonian chocolate tree (Theobroma cacao L). PLoS One 3, e3311. doi: 10.1371/journal.pone.0003311
Oliva M., Rubio K., Epquin M., Marlo G., and Leiva S. (2020). Cadmium uptake in native cacao trees in agricultural lands of Bagua, Perú. Agronomy. 10, 1551. doi: 10.3390/agronomy1010155
Ortiz-Álvarez A., Magnitskiy S., Silva-Arero E. A., Rodríguez-Medina C., Argout X., and Castaño-Marín ÁM. (2023). Cadmium Accumulation in cacao plants (Theobroma cacao L.) under drought stress. Agronomy. 13, 2490. doi: 10.3390/agronomy13102490
Palacio-López K. and Rodríguez-López N. (2007). Phenotypic plasticity in Lippia alba (Verbenaceae) in response to water availability in two light environments. Acta biológica Colombiana 13, 187–198.
Pereira de Araújo R., Furtado de Almeida A.-A., Silva Pereira L., Mangabeira P. A. O., Olimpio Souza J., Pirovani C. P., et al. (2017). Photosynthetic, antioxidative, molecular and ultrastructural responses of young cacao plants to Cd toxicity in the soil. Ecotoxicology Environ. Saf. 144, 148–157. doi: 10.1016/j.ecoenv.2017.06.006
Quiroz Vera J. G. (2000). Agronomic characteristics of clones recommended by INIAP Quevedo, EC, INIAP, Estación Experimental Tropical Pichilingue (Programa Nacional de cacao y café). Available online at: http://repositorio.iniap.gob.ec/handle/41000/3455 (Accessed May 8, 2025).
Remache M., Carrillo M., Mora R., Durango W., and Morales F. (2017). Absorción de macronutrientes y eficiencia del N, en híbrido promisorio de maíz. Patricia Pilar, Ecuador. Agronomía Costarricense 41 (2), 103–115. doi: 10.15517/rac.v41i2.31303
Reyes-Pérez J. J., Llerena-Ramos L. T., Torres-Rodríguez J. A., and HernándezMontiel L. G. (2023b). Optimization of the dose of oligogalacturonides in the vegetative propagation of two cocoa clones (Theobroma cacao L.), under controlled nursery conditions. Terra Latinoamericana 41, e1648. doi: 10.28940/terra.v41i0.1648
Reyes-Pérez J. J., Pincay-Ganchozo R. A., Carrillo-Zenteno M. D., Reynel V., Peña-Salazar K., and Tezara W. (2023a). Macronutrient fertilization and cadmium absorption in two cocoa clones. Horticulturae 9, 1223. doi: 10.3390/horticulturae9111223
Suárez Capello C., Moreira Duque M., Vera B. J., and Vera J. (1993). Manual of cocoa cultivation (Quevedo, Ecuador: INIAP, Estación Experimental Tropical Pichilingue).
Thomas E., Zambrano F., and Loor R. (2024). Catalog of cocoas from Ecuador (Quevedo, Los Ríos, Ecuador: Bioversity International, MOCCA and the National Institute of Agricultural Research (INIAP).
Vera Barahona J., Suárez Capello C., and Mogrovejo E. (1984). Technical description of some cocoa hybrids and clones recommended by the National Institute of Agricultural Research (INIAP) (Quevedo, Ecuador: INIAP, Pichilingue Tropical Experimental Station, Cocoa Program). (Technical Communication no. 12).
Vera Pinargote L. D., Raju Maddela N., Jaimez R. E., and Corozo-Quiñonez L. (2025). Unraveling cadmium stress in cacao (Theobroma cacao L): Photosynthetic performance and differential accumulation in two new hybrid populations under controlled conditions. South Afr. J. Bot. 185, 372–382. doi: 10.1016/j.sajb.2025.08.009
Wang F. Y., Lin X. G., and Yin R. (2007). Inoculation with arbuscular mycorrhizal fungus Acaulospora mellea decreases Cu phytoextraction by maize from Cu-contaminated soil. Pedobiol 51, 99–109. doi: 10.1016/j.pedobi.2007.02.003
Keywords: Theobroma cacao, heavy metals, soils, absorption, physiology
Citation: Freire Carrera F, Carrillo-Zenteno M, Pincay-Ganchozo R, Rendón-Holguín J, Luna-Murillo R, Vecilla-Nicola A, Reynel V and Tezara W (2025) Physiological traits, cadmium bioaccumulation and biomass distribution in seven cocoa genotypes. Front. Agron. 7:1655430. doi: 10.3389/fagro.2025.1655430
Received: 27 June 2025; Accepted: 22 September 2025;
Published: 09 October 2025.
Edited by:
Naser A. Anjum, Aligarh Muslim University, IndiaReviewed by:
Hermes Pérez Hernández, The National Institute of Forestry, Agriculture and Livestock Research (INIFAP), MexicoPaurabi Das, Council of Scientific and Industrial Research (CSIR), India
Copyright © 2025 Freire Carrera, Carrillo-Zenteno, Pincay-Ganchozo, Rendón-Holguín, Luna-Murillo, Vecilla-Nicola, Reynel and Tezara. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Fausto Freire Carrera, ZmZyZWlyZUB1dGUuZWR1LmVj; Wilmer Tezara, d2lsbWVyLnRlemFyYUB1dGVsdnQuZWR1LmVj