 Jahangir Khan
Jahangir Khan André Drenth
André Drenth Olufemi A. Akinsanmi
Olufemi A. Akinsanmi- Centre for Horticultural Science, Queensland Alliance for Agriculture and Food Innovation, The University of Queensland, Brisbane, QLD, Australia
Macadamia is a tree producing high-value nuts and is indigenous to subtropical Australia. Macadamia is commercially cultivated across the globe and the macadamia industry is experiencing increased crop losses caused by fungal pathogens. Limited information is available regarding the association of fungal microbes with macadamia nursery plants. Therefore, this study aims to characterise fungal microbes associated with macadamia plants across five growth stages in leaf, stem and root during four seasons. The fungal community in macadamia nursery plants was analysed using a culture-based approach. Fungi were isolated from germinated seedlings, 3-month-old seedlings, non-grafted plants, grafted trees and 2-year-old trees in orchard. A total of 63 fungal species were identified using DNA sequencing, mainly in the phylum Ascomycota, from the different plant organs. The fungal community structure was significantly influenced by the growth stages (P = 0.001) and plant organs (P<0.001). There was no significant seasonal (P = 0.021) effect in the number of fungal genera isolated from the plants. Grafted plants and young orchards had the richest fungal composition and diversity of the 63 fungal species. Alternaria, Arcopilus, Epicoccum, Nigrospora, and Preussia were found in the leaf, Nectria in the stem, and Mortierella and Penicillium in the root organ only. Diaporthe, Neopestalotiopsis, Pestalotiopsis, Fusarium, and Phyllosticta were detected in all three organs. This study revealed that fungal community richness and diversity in macadamia plants depend on the growth stage and organ examined.
1 Introduction
Macadamias (Macadamia integrifolia and M. tetraphylla, along with their hybrids) are tropical evergreen trees in the plant family Proteaceae and originated in the subtropical rainforest of eastern Australia (Hardner et al., 2019). Macadamia is cultivated for its high-value edible nut and is the only Australian native food crop that has been cultivated on a commercial scale worldwide (Nock et al., 2019). Macadamia nut global demand is increasing, and therefore, the macadamia industry is rapidly expanding worldwide, with a total estimated production of over 78,000 metric tons (kernel basis) in 2020 (INC, 2023). Macadamia is mostly cultivated in Australia, South Africa, USA, China, Kenya, Guatemala, Vietnam, Malawi, Zimbabwe, Brazil, Mexico, Columbia, Mozambique, Paraguay, Rwanda, and New Zealand (INC, 2023). In Australia, major commercial macadamia orchards are found in the Northern Rivers and Nambucca regions of New South Wales (NSW) and Queensland (QLD) in Mackay, Emerald, Bundaberg, Maryborough, and Glasshouse Mountains. The Australian macadamia industry is rapidly expanding, particularly in the Bundaberg region. Macadamia trees occupy over 32,400 hectares under cultivation in Australia, with an annual production of 15,840 metric tons of kernels (AMS, 2024; INC, 2023).
A preliminary study on the macadamia microbial profile shows that nursery plants are inhabited by several fungal genera (Sosso et al., 2021). The structure and function of fungal communities may depend on numerous variables, including plant growth stage, season, planting density, availability of nutrients, environmental conditions, and interactions with the surrounding ecological microorganisms (Liu and Howell, 2021; Gupta et al., 2022; Wei et al., 2023; Habtewold et al., 2021). The variability of fungal communities between seasons and plants under field conditions is usually very high compared to controlled conditions (Hannula et al., 2012). The impact of fungal communities on plants may be (1) direct (expression of disease symptoms or plant death) or (2) indirect (unintended changes in the metabolism of plants, thereby affecting overall growth and production) (Hannula et al., 2012; Birch et al., 2007). The importance of the effects exerted by these factors is still largely unknown in macadamia plants.
Previous studies in other crops have focused on one growth stage and one season without investigating variability over seasons in the crops (Hannula et al., 2012; Negi et al., 2024; Habtewold et al., 2021). In macadamia, a preliminary study showed possible variation in fungal populations in macadamia seedlings and grafted plants (Sosso et al., 2021). However, there is a dearth of information on the fungal diversity at different growth stages (0–3 months old seedlings, 3–6 months old established seedlings, 6–12 months old non-grafted plants, 18–24 months old grafted plants, and 0–2 years old established orchard), seasons (Spring, Summer, Autumn, and Winter) and plant organ (Leaf, Stem, and Root). The fungal communities within plant systems change in response to plant development (Copeland et al., 2015) and have a significant impact on plant health and production (Zahid et al., 2021). Identifying the normal variation in fungal communities during plant growth stages and seasons is crucial to evaluating crop health, yield potential, and potential shifts from endophytic behavior to pathogenic behavior of fungal compositions.
It is not well understood how these non-pathogenic endophytes turn to pathogenic mode in plants (Collinge et al., 2022). The switch of endophytes into pathogenic microbes might be caused by environmental or plant stress (Eaton et al., 2011; Collinge et al., 2022). Extensive research has been conducted on endophytes in agricultural grass hosts as well as a few forest trees like maple, red alder, oak, and various conifers (Wolfe et al., 2018; Szink et al., 2016; Liao et al., 2025; Wolfe et al., 2022; Nicoletti et al., 2021; Fernandez-Conradi et al., 2019; Coelho et al., 2023). However, no research has been conducted on macadamia.
Given that the establishment and prevalence of plant–endophyte interactions are still unknown in macadamia, we sought to examine the fungal composition in batches of plants from germinated seedlings in the nursery to field establishment to answer the following research questions: (1) which fungi persist throughout the plant growth stages in different seasons and organs?, (2) which fungi are found in macadamia nursery plants?, (3) do fungal communities differ in macadamia nursery plants at different growth stages?, (4) if these species are vertically or horizontally transmitted in macadamia, and (5) what is the impact of resident endophytes on field establishment of macadamia plants. It is hypothesized that various fungal microbes infect and inhabit macadamia plants at various growth stages and seasons in nurseries in Australia. Knowledge of the species present and insight how this fungal community can be manipulated may be useful to improve productivity.
2 Materials and methods
2.1 Plant samples
To determine the fungal diversity in leaf, stem, and root tissues at different growth stages and seasons, macadamia plants were sampled from selected batches (Table 1) in Bundaberg, Queensland (24° 51′ 58″ S, 152° 20′ 52″ E; altitude 17.1 m). Sequential sampling was conducted to represent four growth stages including germinated seedlings (<3 months old), established seedlings (3–6 months old), non-grafted plants (6–12 months old) and grafted plants (18–24 months old). In addition, samples were obtained from two batches, 3 months after field establishment in an orchard (Table 1). A total of 170 plants were sampled. Each plant sample was carefully washed under running tap water, to remove debris from the surface. Leaf, stem, and root tissues were obtained from each sample of germinated seedlings (n = 10), established seedlings (n = 15), non-grafted plants (n = 20), grafted plants (n = 95), and young grafted trees in the orchard (n = 30).
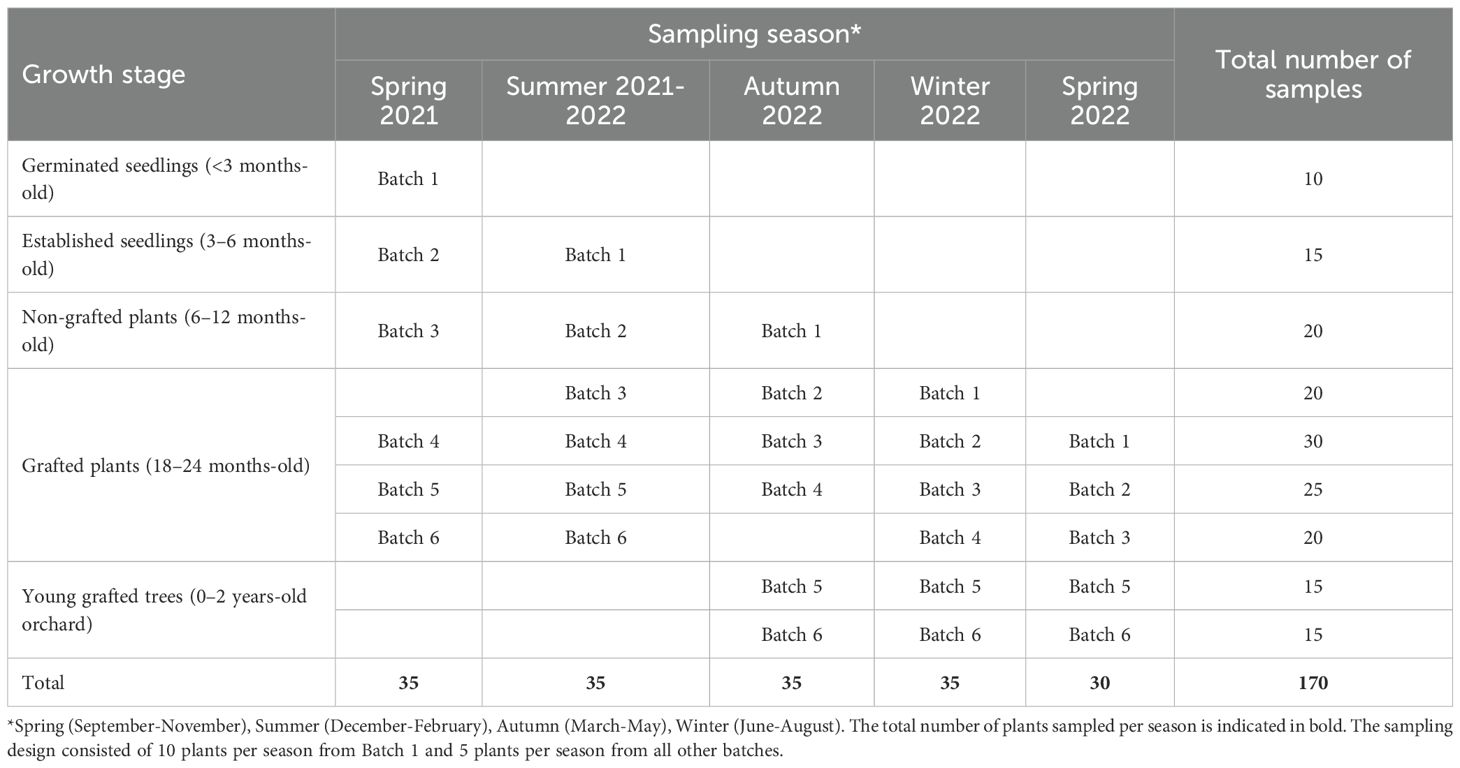
Table 1. Macadamia plants sampled at various growth stages and propagation batches.
2.2 Fungal isolation
Approximately 5–10 mm disc of each tissue segment was used for microbial isolation. A total of five tissues pieces per plate in three replicates were used for each sample. Surface sterilisation of the tissues was conducted by soaking in 70% ethanol (w/v) for 2 min, followed by 3% sodium hypochlorite solution (NaOCl) containing 0.1% Tween 20 for 2 min and in 50% ethanol (w/v) for 2 min (Araújo et al., 2001). Tissues were then rinsed twice in sterile water (Millipore Q-POD® Milli-Q, Biopak®) for 3 min and dried on sterile blotting paper before planting on 2% MEA (Malt extract agar, pH 5.5) (Merck KGaA, Darmstadt, Germany) in 90 mm plastic Petri plates. The plates were incubated at 25 °C with a 12−h photoperiod for 3−5 days. All fungal colonies were subcultured on ½-strength PDA (Potato dextrose agar, pH 7.0) (Difco™ Becton, Dickinson and Company, Sparks, USA) plates amended with streptomycin (50 µg/ml). Monoconidial and single hyphal tip cultures were derived from selected isolates as described by Akinsanmi et al. (2004) and stored at -80°C in sterile 15% glycerol solution (Akinsanmi et al., 2004).
2.3 Molecular characterisation of fungal isolates
For identification of the fungal isolates, the DNA was extracted from mycelium (≈40 mg) using robotic platform by following the procedure of BioSprint 96 DNA Plant Kit (Qiagen Pty Ltd), after the mycelium tissues was homogenised in 300 µl RLT lysis buffer with a 3 mm sterile tungsten carbide bead for 2 min at 30 Hz using TissueLyser (Qiagen Pty Ltd). Biodrop (Duo Spectrophotometer) was used for DNA quantification and the final concentration was maintained at 10 ng/µl. Polymerase Chain Reaction (PCR) was performed to amplify a portion of the 18S, 5.8S, and 28S nrRNA of the internal transcribed spacer (ITS) region using primers ITS4 and ITS5 as fungal barcoding gene (White et al., 1990). The fungal microbes identity were confirmed by multi-gene analyses of its sequences with the β-tubulin (BT) sequence of gene region using T1 and BT2b primers (Glass and Donaldson, 1995) glyceraldehyde-3-phosphate dehydrogenase (GAPDH) with primers GDF and GDR (Templeton et al., 1992); partial actin (ACT) using ACT-512F and ACT-783R (Carbone and Kohn, 1999); and translation elongation factor 1-alpha (TEF) gene with EF1-688F and EF1-1251R primers (Alves et al., 2008). PCR reaction mixes were composed of 10 μl 5X Mangotaq reaction buffer, 4 μl MgCl2, 1 μl dNTPs, 1 μl of each forward and reverse primer, 2 μl BSA (10 μg/μl), 1 μl 100% DMSO, 0.4 μl DNAtaq Polymerase, 26.6 μl of DNAse free water and 3 μl template DNA, totaling 50 μl. The PCR was performed in a Thermal Cycler (Kyratech, Fisher Biotec, Australia). Initial denaturation was performed at 95°C for 5 min, denaturation at 95°C for 30 s, annealing for ITS and BT at 49°C for 30 s, GAPDH and ACT at 61.5°C for 30 s, and for TEF at 52°C for 30 s, extension at 72°C for 45 s, and final extension at 72°C for 5 min. PCR amplicons were stained with 3X GelRed and separated on 1.0% agarose gels (Bioline, Australia) in 0.5% tris-ethylenediaminetetraacetic acid (TE) buffer and electrophoresis at 100V for 45 min. Successful amplification was determined under ultraviolet (UV) transillumination using Molecular Imager® and GelDoc™ (Bio-Rad Laboratories Inc., Segrate, and Milan, Italy). A 1 kb HyperLadder (Bioline) was used to deduce amplicon size. Both a negative control with no template DNA, and a positive control were included. Amplified products were purified using QIAquick PCR Purification Kit (Qiagen) according to manufacturer’s instructions and sequenced in both direction at Macrogen Inc. (South Korea). DNA sequences were manually trimmed, checked for quality in Geneious software version 10.2.4 (Biomatters, Ltd., Auckland, New Zealand) and compared with sequences in GenBank nucleotide database using Blast Local Alignment Search Tool (BLAST) (https://blast.ncbi.nlm.nih.gov/) in National Centre for Biotechnology Information (NCBI). Sequences with pairwise similarities > 99% were used to determine the identity of the fungal isolates.
2.4 Impact and translation to field disease
To understand the impact of endophytes on plant diseases that lead to early tree mortality in young orchards, a disease survey was conducted for macadamia plant batch 5 (A203/H2) and batch 6 (HAES741/H2) transplanted into the orchard during the study. Both batches were transplanted into the same farmer’s orchard and kept under similar management practices. The macadamia plants were surveyed for visual symptoms of stem diseases, including graft dieback, stem canker, gall canker and tree mortality during 2022-2023. In each orchard during each visit, 10 rows and 20 consecutive plants from each row were observed for stem diseases. The trees with symptoms were tagged and excluded from the follow-up visit. A total of 800 trees, each of batch 5 and batch 6, were surveyed. The four border rows of each end of the orchard were excluded from the study to eliminate field border effects.
2.5 Biocontrol potential of macadamia endophytes
Macadamia endophytic isolates were evaluated in vitro to understand the potential antagonistic activity against known pathogens. Representative isolates of Aspergillus japonicus, A. hiratsukae, A. ibericus, Phyllosticta paracapitalensis, P. capitalensis, Mortierella elongate, Trichoderma atroviride, T. koningii, and T. longibrachiatum were evaluated for their antagonistic activity against known stem pathogens, Diaporthe australiana, Neofusicoccum luteum, N. parvum, Lasiodiplodia theobromae, L. pseudotheobromae, L. iraniensis and Nectria pseudotrichia. Briefly, mycelial discs of 5 mm diameter from the edge of actively growing colonies of each antagonist and pathogen were placed at opposite sites on a 90 mm diameter PDA plate (20 mm from the border). The pathogen alone on the plate was used as a control. The plates were incubated at 25°C with a 12-h photoperiod for 7 days. After 7 days, the radius of pathogen colonies facing the antagonists was measured and used to calculate the mycelial growth inhibition rate. The antagonistic levels were classified as low (≤ 50%), medium (50% - 60%), high (60% - 75%) and very high (>75%).
2.6 Statistical analysis
Fungal population data for plant organs, plant growth stages, seasons, and interactions among plant organs, growth stage, and seasons were analysed by general analysis of variance (ANOVA) procedure in GenStat 23rd edition (VSN International, Hemel Hempstead, UK). The isolation rate (%) of each fungal species was calculated using the following formula
A is the number of isolates from the same species, and S is the total number of isolates.
Fungal population diversity measures, including richness, Shannon Weaver evenness index (J’) and Shannon Weaver diversity index (H’), were calculated for each plant organs, growth stage, and seasons separately. The H’ was calculated using the following formula:
Where Pi is the proportion of individuals of a particular taxon (i), s is the total number of isolates and ln is the natural logarithm.
The Shannon Weiner evenness (J’) was calculated using following formula:
Where H′ is the Shannon index and s is the total number of isolates.
The stem Disease Prevalence (DP) was based on the number of symptomatic plants relative to the cumulative number of plants sampled within batch 5 and batch 6 plants that were transplanted into orchards using the following formula:
To validate the relationship of fungal species within batch 5 and batch 6, macadamia plants were explored using the principal component analysis procedure in GenStat 23rd edition.
The mycelial growth inhibition (%) of pathogens was calculated using the following formula,
Where Pr is the growth radius of the pathogen in control, and Ar is the growth radius of the pathogen with antagonists.
3 Results
3.1 Fungal microbes in macadamia
In this study, 5874 fungal microbes were isolated from macadamia plants in Australia (Table 2). A total of 14 fungal families consisting of 20 genera and 63 species were identified from macadamia plant samples. The majority of isolates belonged to the Diaporthe (15.8%) followed by Neopestalotiopsis (13.3%), Colletotrichum (11.1%), Alternaria (6.8%), Pestalotiopsis (6.2%), Cladosporium (5.9%), Phyllosticta (5.0%), Neofusicoccum (4.6%), and Lasiodiplodia (4.1%) while a low frequency of isolation was observed for Paraconiothyrium (0.8%) (Table 2; Figure 1). At the species level, isolation frequencies ranged from 0.2% to 4.2%. Colletotrichum siamense and C. gloeosporioides were the most common (4.2% each), followed by N. clavispora (3.8%), whereas Arcopilus navicularis was the least frequent (0.2%) (Table 2; Figure 1).
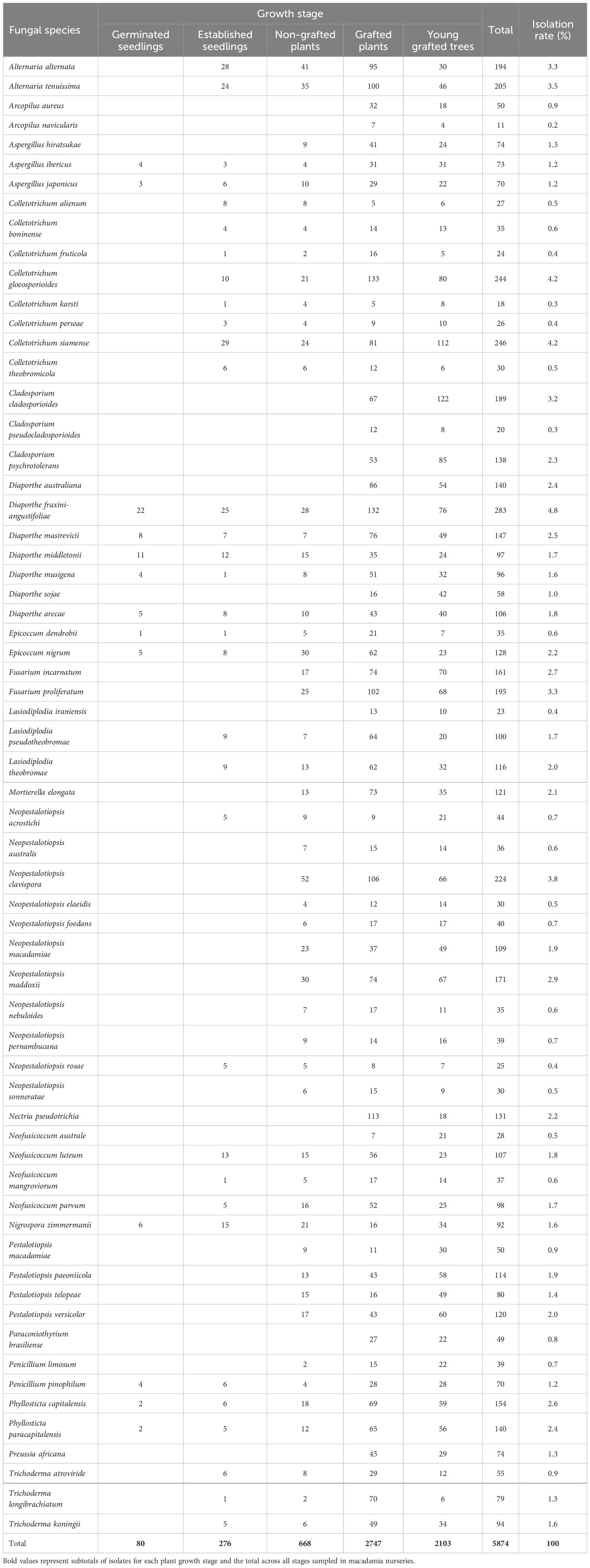
Table 2. Fungal species obtained from macadamia nursery plants at different growth stages.
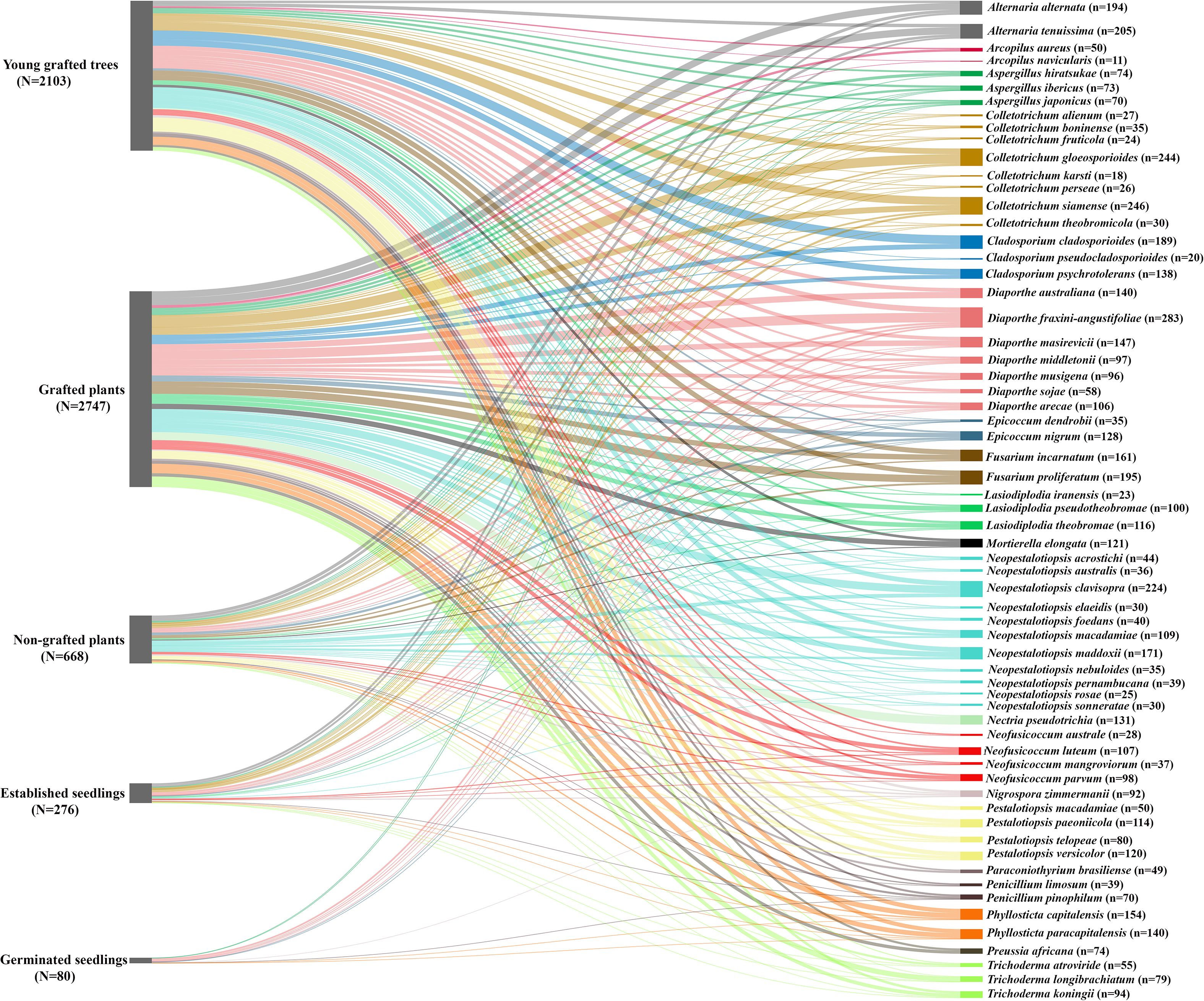
Figure 1. Relative abundance of fungal species isolated from Macadamia (Macadamia integrifolia and M. tetraphylla) at different growth stages. The width of each node represents the relative abundance of isolates. Growth stages are displayed on the left, showing the total fungal abundance at each stage, while fungal species are displayed on the right, showing their distribution across all stages. The total number of fungal isolates recorded for each growth stage is represented by N, while n denotes the total number of isolates of each fungal species across all growth stages.
Statistical analysis revealed that both growth stage and plant organ had a highly significant effect on fungal community structure (P< 0.001 for both). Although season also showed a significant effect (P = 0.021), its influence was substantially weaker than that of growth stage and organ. The interaction between growth stage and plant organs (P<0.001), growth stage and season (P = 0.009), and plant organs and season (P<0.001) was significantly different, while the interaction was not significantly different among growth stage, plant organs and season (P = 0.028). The major influencing parameters between growth stage and season, plant organs and season, growth stage, plant organs and season interactions were growth stages and plant organs. The variations in fungal diversity between growth stages and plant organs were much higher compared to seasons.
3.2 Diversity of fungal communities at various growth stages in macadamia
The fungal community does change among the different growth stages. Germinated seedlings have the lowest diversity index (H’ = 2.36), followed by established seedlings (H’ = 3.19), non-grafted plants (H’ = 3.68), grafted plants (H’ = 3.89), and young grafted trees (H’ = 3.89). The highest evenness index was in young grafted trees (J’ = 0.94), grafted plants (J’ = 0.93), and non-grated plants (J’ = 0.93), followed by established seedlings (J’ = 0.91), and germinated seedlings (J’ = 0.89). Young grafted trees and grafted plants have the richest species composition (63 species), followed by non-grafted plants (52 species), established seedlings (34 species), and germinated seedlings (14 species).
Six fungal genera, Diaporthe, Phyllosticta, Aspergillus, Epicoccum, Penicillum, and Nigrospora were detected in all five growth stages (Table 2). Six fungal genera, Neopestalotiopsis, Colletotrichum, Neofusicoccum, Alternaria, Lasiodiplodia, and Trichoderma were detected in four growth stages (Table 2). Three fungal genera, Pestalotiopsis, Fusarium, and Mortierella were detected in three growth stages, while Cladosporium, Nectria, Preussia, Arcopilus, and Paraconiothyrium were detected only in grafted plants and young grafted trees (Table 2; Figure 1). Diaporthe was the major genus identified in young grafted trees and grafted plants followed by Neopestalotiopsis and Colletotrichum (Table 2). In non-grafted plants and established seedlings, the major microbial genera were Neopestalotiopsis, Colletotrichum, Alternaria, and Diaporthe. In germinated seedlings, Diaporthe was the most prominent fungal genera (Table 2; Figure 1).
3.3 Species distribution at various growth stages in macadamia
Thirteen fungal species were detected in all five growth stages, including Diaporthe fraxini-angustifoliae, D. middletonii, D. masirevicii, D. sojae, D. musigena, Phyllosticta capitalensis, P. paracapitalensis, Aspergillus ibericus, A. japonicus, Epicoccum nigrum, E. dendrobii, Penicillium pinophilum, and Nigrospora zimmermanii (Table 2). Diaporthe australiana, D. sojae and D. litchicola were detected only in grafted plants and young grafted trees, whereas A. hiratsukae, P. limosum were detected in non-grafted, grafted and young grafted trees (Table 2; Figure 1).
Neopestalotiopsis acrostichi, N. rosae, Colletotrichum. alienum, C. boninense, C. fruticola, C. gloeosporioides, C. karsti, C. perseae, C. siamense, and C. theobromicola, Neofusicoccum luteum, N. mangroviorum, N. parvum, Alternaria alternata, A. tenuissima, Lasiodiplodia pseudotheobromae, L. theobromae, Trichoderma koningii, T. atroviride, and T. longibrachiatum were detected in four growth stages, established seedlings, non-grafted plants, grafted plants, young grafted trees. Neopestalotiopsis australis, N. clavispora, N. elaeidis, N. foedans, N. macadamiae, N. maddoxii, N. nebuloides, N. pernambucana, and N. sonneratae were detected in non-grafted, grafted plants and young grafted trees in the field. Neofusicoccum australe and L. iraniensis were detected in grafted plants and young grafted trees (Table 2; Figure 1).
Pestalotiopsis macadamiae, P. paeoniicola, P. telopeae, P. versicolor, Fusarium incarnatum, F. proliferatum and Mortierella elongate were detected in three growth stages: non-grafted plants, grafted plants and young grafted trees (Table 2). Cladosporium cladosporioides, C. psychrotolerans, C. pseudocladosporiodes, Nectria pseudotrichia, Preussia africana, Arcopilus aureus, A. navicularis, and Paraconiothyrium brasiliense were detected only in grafted plants and young grafted trees (Table 2).
3.4 Fungal diversity in macadamia leaf, stem, and root of plants
Stem tissues had the highest diversity index (H’ = 3.63) followed by leaf (H’ = 3.44) and root (H’ = 3.31). While the evenness index was highest in stem (J’ = 0.92) and root (J’ = 0.92), followed by leaf (J’ = 0.88). A total of 52 species were obtained from the stem, followed by 51 species from the leaf and 36 species from the root.
Five fungal genera Diaporthe, Neopestalotiopsis, Pestalotiopsis, Fusarium, and Phyllosticta were detected in all three organs. At the species level, D. australiana, D. sojae, and F. incarnatum were detected in the stem and root. Fusarium proliferatum was detected in the leaf and stem (Figure 1), and the remaining species were detected in all three organs (Figure 2).
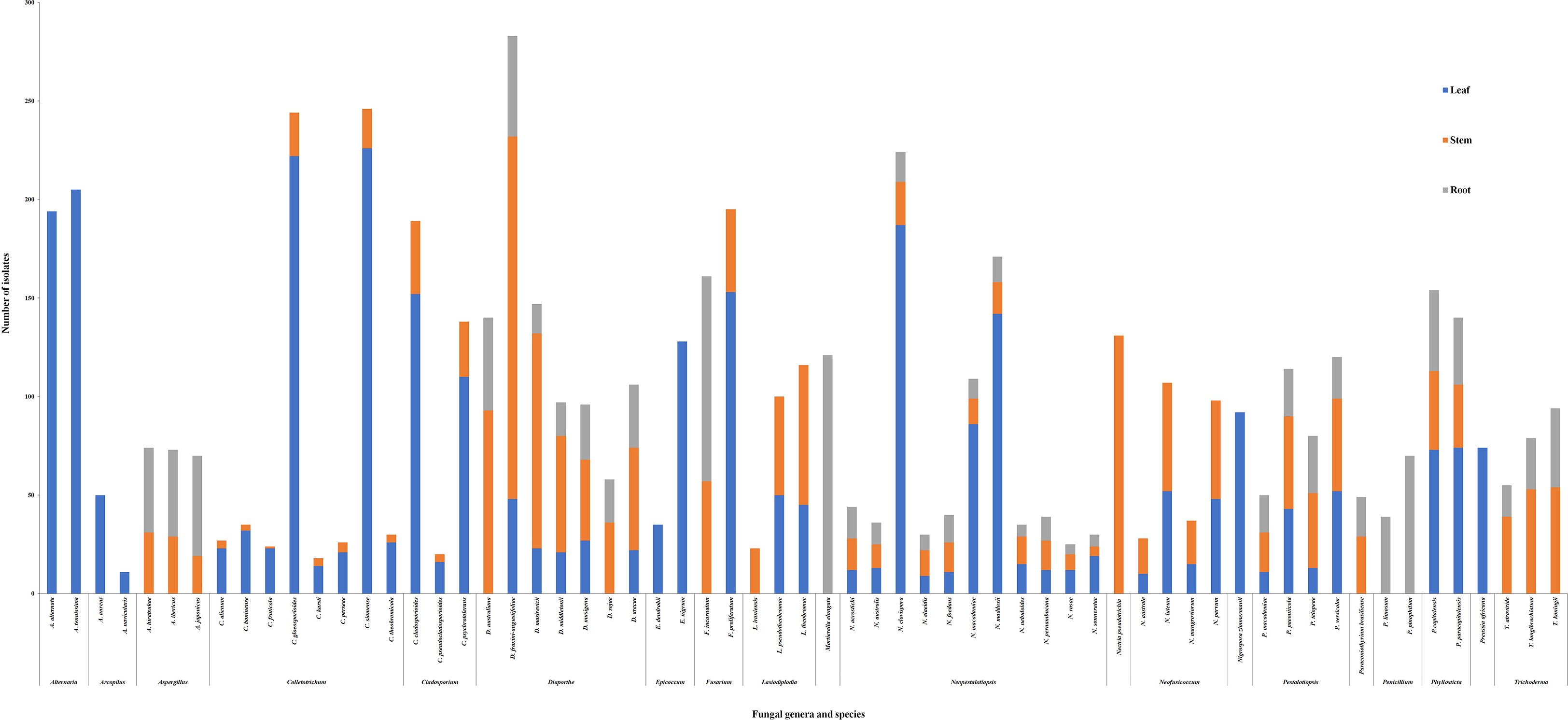
Figure 2. Fungal genera distribution in Macadamia (Macadamia integrifolia and M. tetraphylla) plants in leaf, stem and root tissues.
Four fungal genera, Colletotrichum, Cladosporium, Neofusicoccum, Lasiodiplodia, and all of their associated species were detected in leaf and stem organs. Accordingly, three fungal genera, Trichoderma, Aspergillus, and Paraconiothyrium, were detected in the stem and root.
Alternaria, Arcopilus, Epicoccum, Nigrospora, and Preussia were detected in leaves only. Nectria pseudotrichia were detected in the stem, and Mortierella and Penicillum were detected only in the roots organ only (Figure 2).
3.5 Seasonal variation among fungal communities in nursery plants
Overall, the diversity and evenness indices among the four seasons (Spring, Summer, Autumn, and Winter) are strikingly similar. All 63 species were detected in autumn, winter, and spring, while 61 species were detected in summer.
3.6 Field disease impact
There was a significant difference in stem disease prevalence and tree mortality between batch 5 (A203/H2) and batch 6 (HAES741/H2). Batch 5 shows 4% of plants with graft dieback, 1% stem canker, 2% gall canker, and 7.5% tree mortality. In contrast, 9.5% of plants with graft dieback, 2.5% stem canker, 2.5% gall canker, and 13.5% tree mortality were in batch 6.
When comparing the fungal microbial data of both batches post-grafting stages, the isolation for Trichoderma spp. is significantly lower for batch 6 (n = 10) than batch 5 (n = 93), and the number of potential pathogenic Neofusicoccum parvum (n = 33), N. luteum (n = 34), Lasiodiplodia theobromae (n = 48), L. pseudotheobromae (n = 32), and L. iraniensis (n = 18) were significantly higher in batch 5 as compared to batch 6, N. parvum (n = 9), N. luteum (n = 13), L. theobromae (n = 6), and L. pseudotheobromae (n = 2).
3.7 Relationship among fungi
To explore the interactions among plant pathogenic and beneficial fungal species, a principal component analysis (PCA) was conducted using isolates from the stem and root organs of batch 5 and 6. The first two principal components explained 71.5% of the variation. The PC1 separated the beneficial fungi from the pathogenic fungi (Figure 3). The PCA analysis revealed the beneficial fungi were separated from the pathogen fungal species, based on their aggressiveness or frequency of colonisation in the stem (Figure 3). The pathogenic species of Neofusicoccum parvum, N. luteum, Lasiodiplodia theobromae, L. pseudotheobromae, and L. iraniensis were clustered together and were segregated from Diaporthe australiana, Nectria pseudotrichia (Figure 3). The occurrence of Diaporthe australiana and Nectria pseudotrichia were unrelated to Neofusicoccum, Lasiodiplodia, and Trichoderma species (Figure 3).
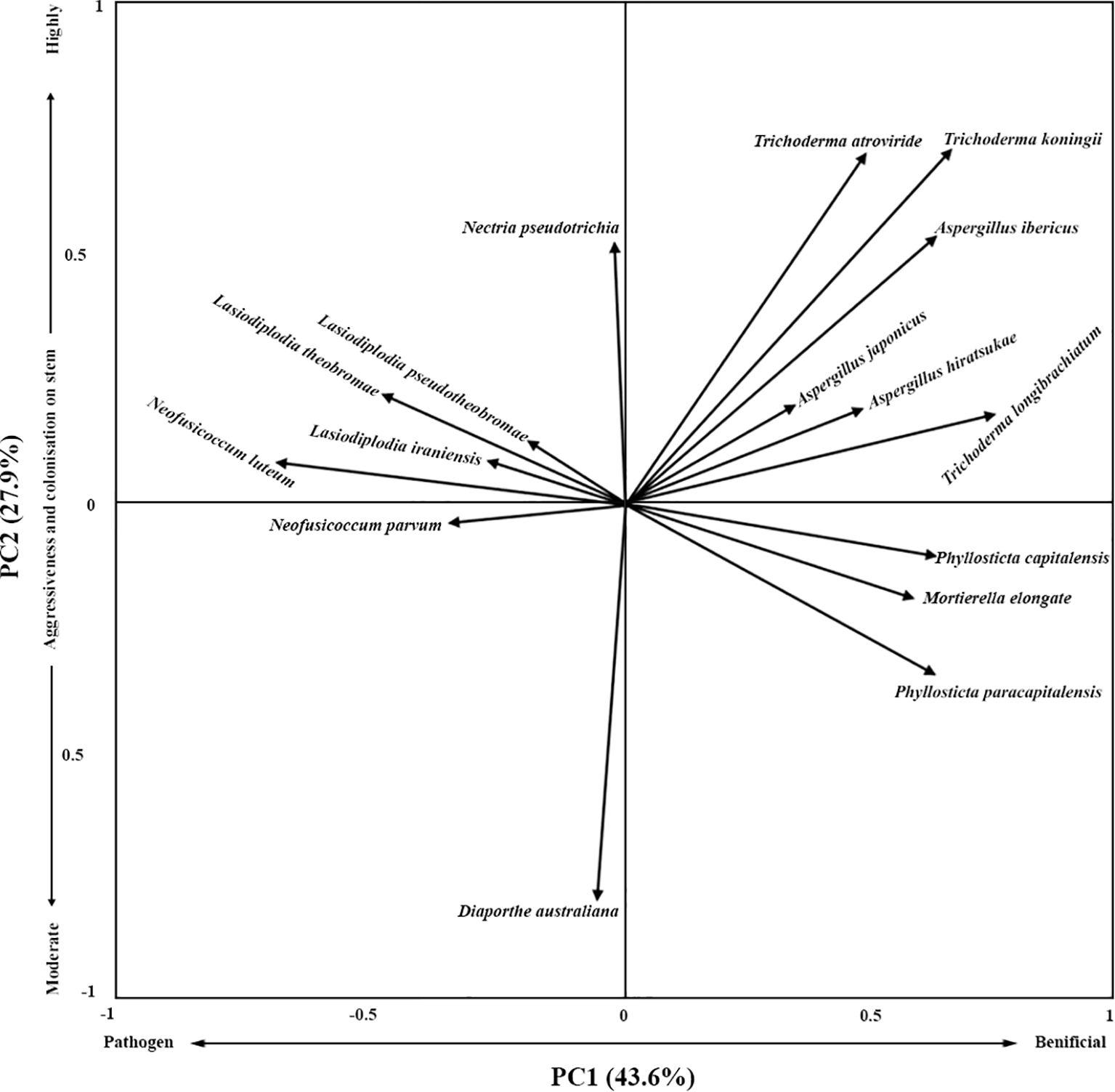
Figure 3. Ordination biplots from PCA analysis showing relationships between beneficial fungal (Trichoderma atroviride, T. koningii, T. longibrachiatum, Aspergillus ibericus, A. japonicus, A. hiratsukae, Phyllosticta paracapitalensis, P. capitalensis, and Mortierella elongate) and pathogenic species (Neofusicoccum parvum, N. luteum, Lasiodiplodia theobromae, L. pseudotheobromae, L. iraniensis, Diaporthe australiana, and Nectria pseudotrichia).
Trichoderma atroviride, T. koningii, T. longibrachiatum, Aspergillus ibericus, A. japonicus, and A. hiratsukae co-occurred. These colonisers had a negative relationship with Diaporthe australiana, Neofusicoccum. parvum, N. luteum, Lasiodiplodia theobromae, L. pseudotheobromae and L. iraniensis graft dieback and branch dieback pathogens (Figure 3). Phyllosticta paracapitalensis, P. capitalensis and Mortierella elongate seemed to co-occur and negatively related to Neofusicoccum parvum, N. luteum, Lasiodiplodia theobromae, L. pseudotheobromae, L. iraniensis, and Nectria pseudotrichia (Figure 3).
3.8 Biocontrol potential of macadamia plant endophytes
In vitro biological control of representative known macadamia pathogens Diaporthe australiana, Neofusicoccum luteum, N. parvum, Lasiodiplodia theobromae, L. pseudotheobromae, and L. iraniensis by potential biocontrol agents Aspergillus japonicus, A. hiratsukae, A. ibericus, Phyllosticta paracapitalensis, P. capitalensis, Mortierella elongate, Trichoderma atroviride, T. koningii and T. longibrachiatum by inhibition of mycelial growth of pathogens. All potential biocontrol agent isolates did not significantly reduce the mycelial growth of pathogens in dual culture tests in Petri dishes (Table 3).
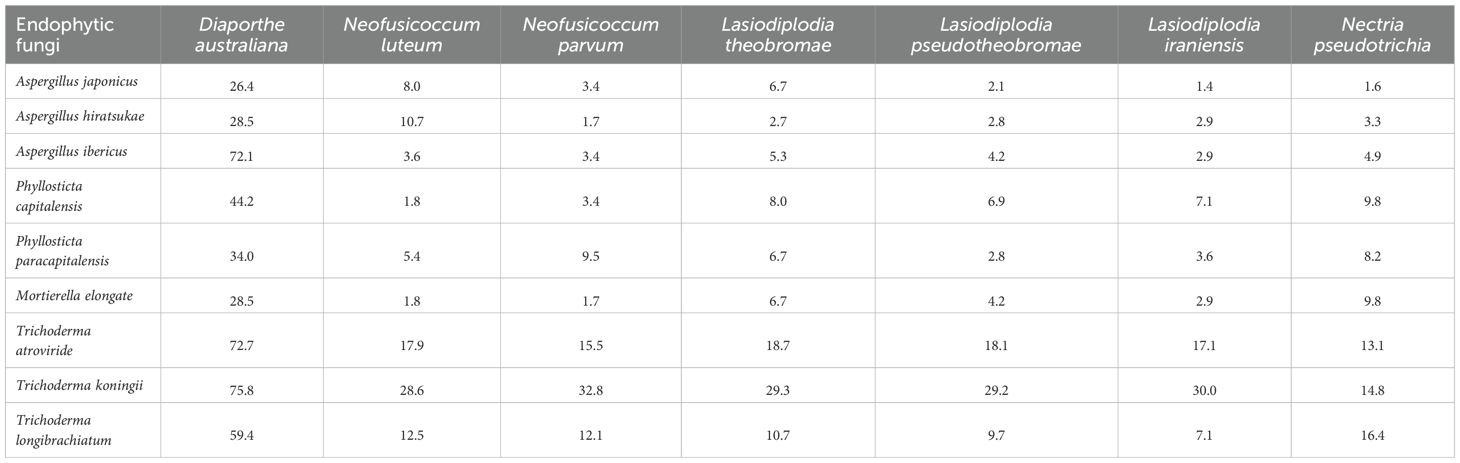
Table 3. Percent mycelial growth rate (%) of endophytic fungi against known macadamia stem pathogens.
Some good combinations of biocontrol agents were observed. A dual culture test in Petri dishes shows that Trichoderma koningii, T. atroviride, Aspergillus ibericus and T. longibrachiatum reduce the growth of Diaporthe australiana by 75.8%, 72.7%, 72.1% and 59.4%, respectively (Table 3).
4 Discussion
This study advanced understanding of the mycobiome in macadamia production systems, highlighting key shifts from seedling germination to young orchard trees. We identified a complex community of 63 fungal species across 20 genera, revealing that this community is dynamic and shaped primarily by plant growth stage and organ type, rather than seasonal variation. Crucially, the in vitro antagonism assays, coupled with field disease data and interaction analyses, identified specific endophytic fungi with significant potential as native biocontrol agents against key macadamia pathogens.
Our findings clearly demonstrate that the macadamia mycobiome is not static but undergoes significant succession as the plant develops. The low diversity in germinated seedlings, dominated by a few genera like Diaporthe, suggests an initial colonisation by generalist fungi. The subsequent increase in diversity and evenness through established seedlings, non-grafted, and grafted plants reflects a process of ecological succession, where the developing plant recruits a more complex and likely more specialized microbial community (Hardoim et al., 2015). The convergence of high species richness in grafted plants and young trees indicates the establishment of a stable, mature mycobiome. The significant influence of plant organ, with stems harboring the greatest diversity, highlights the concept of tissue specific micro-niches, where different anatomical sites select for distinct fungal communities based on factors like nutrient availability and chemical defenses (Zimmerman and Vitousek, 2012). The relative insignificance of season contrasts with studies on foliar pathogens in annual crops, suggesting that in perennial nursery systems, the host developmental stage is a stronger driver of microbial assembly than short-term environmental fluctuations.
The dual culture assays were an effective primary screen to identify endophytes with direct antagonistic capabilities. The strong inhibition of Diaporthe australiana by isolates of Trichoderma koningii (75.8%), T. atroviride (72.7%), and notably Aspergillus ibericus (72.1%) is a key finding. The efficacy of Trichoderma spp., is consistent with their well-documented role as mycoparasites (Harman et al., 2004). However, the high antagonistic activity of Aspergillus ibericus, a less commonly reported biocontrol agent, points to the value of screening native mycobiomes for novel functional traits. The specificity of these interactions, where other endophytes showed negligible activity, underscores that biocontrol potential is a strain-specific attribute rather than a generic feature of a genus.
The in vitro findings are strongly supported by the field disease data and the Principal Component Analysis (PCA). The stark contrast in disease outcomes between Batch 5 and Batch 6 provides compelling correlative evidence. Batch 6, which suffered higher mortality and disease prevalence, had a significantly lower abundance of Trichoderma spp., and a concomitantly higher abundance of pathogenic Neofusicoccum and Lasiodiplodia species. This inverse relationship suggests that a robust community of beneficial fungi, particularly Trichoderma, may suppress pathogen populations in planta. The PCA biplot (Figure 3) statistically validates this inverse relationship, clearly segregating beneficial fungi (Trichoderma spp., Aspergillus spp., and Phyllosticta spp.), from the aggressive pathogens (Neofusicoccum spp., Lasiodiplodia spp., Diaporthe sp. and Nectria sp.). The clustering of beneficial fungi together suggests potential synergism or shared environmental preferences, while their negative correlation with pathogens indicates competitive exclusion or antagonism. The separate clustering of Diaporthe australiana and Nectria pseudotrichia from the main pathogen group is intriguing, suggesting they may occupy a different ecological niche or have a different, perhaps less aggressive, pathogenic lifestyle. The negative relationship of Phyllosticta and Mortierella with pathogens further expands the list of potential beneficial taxa beyond Trichoderma.
The higher disease pressure in Batch 6 cannot be attributed to a single pathogen but rather to an enrichment of a pathobiome, a consortium of multiple pathogens occurring simultaneously (Lamichhane and Venturi, 2015). The data suggest that the absence of a strong beneficial mycobiome may facilitate the proliferation of this pathobiome. Therefore, managing for tree health may be more effectively achieved by fostering a protective mycobiome rather than targeting individual pathogens.
The isolation of potent antagonistic endophytes native to macadamia and already adapted to its internal tissues highlights their potential as ideal candidates for probiotic inoculants, as successful colonisation not only establishes a protected niche but also confers resistance to subsequent fungal invasions by enhancing the primary coloniser’s ability to persist and resist replacement (John and White, 2017).
In conclusion, this research delineates the succession of the mycobiome in macadamia nurseries and identifies key growth stages where microbial communities are most susceptible to change. We have identified specific native endophytes, most notably Trichoderma koningii, T. atroviride, and Aspergillus ibericus, that exhibit strong in vitro antagonism against major pathogens, a finding supported by inverse correlations. These isolates represent excellent candidates for developing novel biocontrol strategies to mitigate stem canker and other diseases. Future research should priorities in planta validation, mechanistic elucidation, and the development of synergistic endophyte consortia to achieve broad-spectrum and durable disease suppression. Advancing scalable delivery strategies will be critical to enable effective colonisation and persistence, thereby establishing microbiome-informed approaches for sustainable disease management in the macadamia industry.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding authors.
Author contributions
JK: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Project administration, Resources, Software, Validation, Writing – original draft, Writing – review & editing. AD: Conceptualization, Funding acquisition, Project administration, Resources, Supervision, Writing – review & editing. OA: Conceptualization, Funding acquisition, Project administration, Resources, Supervision, Visualization, Writing – review & editing, Formal analysis, Validation.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This research was funded by Hort Innovation using the macadamia research and development levy and nursery research and development levy funds levy and funds from the Australian Government, project nos. MC21001 and NY20000. JK is a recipient of the University of Queensland Research Training Scholarship.
Acknowledgments
The authors thank the macadamia nursery industry for the collection of plant samples for this study.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Akinsanmi O. A., Mitter V., Simpfendorfer S., Backhouse D., and Chakraborty S. (2004). Identity and pathogenicity of Fusarium spp. isolated from wheat fields in Queensland and northern New South Wales. Aust. J. Agric. Res. 55, 97–107. doi: 10.1071/AR03090
Alves A., Crous P. W., Correia A., and Phillips A. (2008). Morphological and molecular data reveal cryptic speciation in Lasiodiplodia theobromae. Fungal Diversity 28, 1–13.
AMS (2024). Australian macadamias yearbook Vol. 1 (Australia: Australian Macadamia Society News Bulletin), 3–16.
Araújo W. L., Maccheroni W. Jr., Aguilar-Vildoso C. I., Barroso P. A., Saridakis H. O., and Azevedo J. L. (2001). Variability and interactions between endophytic bacteria and fungi isolated from leaf tissues of citrus rootstocks. Can. J. Microbiol. 47, 229–236. doi: 10.1139/w00-146
Birch A. N. E., Griffiths B. S., Caul S., Thompson J., Heckmann L. H., KROGH P. H., et al. (2007). The role of laboratory, glasshouse and field scale experiments in understanding the interactions between genetically modified crops and soil ecosystems: A review of the ECOGEN project. Pedobiologia 51, 251–260. doi: 10.1016/j.pedobi.2007.04.008
Carbone I. and Kohn L. M. (1999). A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 91, 553–556.
Coelho V., Nunes L., Moura L., and Gouveia E. (2023). Characterization of chestnut bark fungal communities in healthy trees and blight recovered through natural or introduced hypovirulence. J. Plant Pathol. 105, 1031–1043. doi: 10.1007/s42161-023-01424-y
Collinge D. B., Jensen B., and Jørgensen H. J. L. (2022). Fungal endophytes in plants and their relationship to plant disease. Curr. Opin. Microbiol. 69, 102177. doi: 10.1016/j.mib.2022.102177
Copeland J. K., Yuan L., Layeghifard M., Wang P. W., and Guttman D. S. (2015). Seasonal community succession of the phyllosphere microbiome. Mol. Plant-Microbe Interact. 28, 274–285. doi: 10.1094/MPMI-10-14-0331-FI
Eaton C. J., Cox M. P., and Scott B. (2011). What triggers grass endophytes to switch from mutualism to pathogenism? Plant Sci. 180, 190–195. doi: 10.1016/j.plantsci.2010.10.002
Fernandez-Conradi P., Fort T., Castagneyrol B., Jactel H., and Robin C. (2019). Fungal endophyte communities differ between chestnut galls and surrounding foliar tissues. Fungal Ecol. 42, 100876. doi: 10.1016/j.funeco.2019.100876
Glass N. L. and Donaldson G. C. (1995). Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 61, 1323–1330. doi: 10.1128/aem.61.4.1323-1330.1995
Gupta A., Singh U. B., Sahu P. K., Paul S., Kumar A., Malviya D., et al. (2022). Linking soil microbial diversity to modern agriculture practices: a review. Int. J. Environ. Res. Public Health 19, 3141. doi: 10.3390/ijerph19053141
Habtewold J. Z., Helgason B. L., Yanni S. F., Janzen H. H., Ellert B. H., and Gregorich E. G. (2021). Warming effects on the structure of bacterial and fungal communities in diverse soils. Appl. Soil Ecol. 163, 103973. doi: 10.1016/j.apsoil.2021.103973
Hannula S. E., De Boer W., and Van Veen J. (2012). A 3-year study reveals that plant growth stage, season and field site affect soil fungal communities while cultivar and GM-trait have minor Effects. PloS One 7, e33819. doi: 10.1371/journal.pone.0033819
Hardner C. M., Wall M., and Cho A. (2019). Global macadamia science: Overview of the special section. HortScience 54, 592–595. doi: 10.21273/HORTSCI13543-18
Hardoim P. R., Van Overbeek L. S., Berg G., Pirttilä A. M., Compant S., Campisano A., et al. (2015). The hidden world within plants: ecological and evolutionary considerations for defining functioning of microbial endophytes. Microbiol. Mol. Biol. Rev. 79, 293–320. doi: 10.1128/MMBR.00050-14
Harman G. E., Howell C. R., Viterbo A., Chet I., and Lorito M. (2004). Trichoderma species - opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2, 43–56. doi: 10.1038/nrmicro797
INC (2023). Nuts and dried fruits, Statistical yearbook (Spain: International Nut and Dried Fruit Press), 30–33.
John D. J. and White F. (2017). The fungal community: its organization and role in the ecosystem (United States: Taylor and Francis).
Lamichhane J. R. and Venturi V. (2015). Synergisms between microbial pathogens in plant disease complexes: a growing trend. Front. Plant Sci. 6, 385. doi: 10.3389/fpls.2015.00385
Liao C., Doilom M., Jeewon R., Hyde K. D., Manawasinghe I. S., Chethana K., et al. (2025). Challenges and update on fungal endophytes: classification, definition, diversity, ecology, evolution and functions. Fungal Diversity 131, 301–367. doi: 10.1007/s13225-025-00550-5
Liu D. and Howell K. (2021). Community succession of the grapevine fungal microbiome in the annual growth cycle. Environ. Microbiol. 23, 1842–1857. doi: 10.1111/1462-2920.15172
Negi R., Sharma B., Kumar S., Chaubey K. K., Kaur T., Devi R., et al. (2024). Plant endophytes: unveiling hidden applications toward agro-environment sustainability. Folia Microbiol. 69, 181–206. doi: 10.1007/s12223-023-01092-6
Nicoletti R., Beccaro G. L., Sekara A., Cirillo C., and Di Vaio C. (2021). Endophytic fungi and ecological fitness of chestnuts. Plants 10, 542. doi: 10.3390/plants10030542
Nock C. J., Hardner C. M., Montenegro J. D., Ahmad Termizi A. A., Hayashi S., Playford J., et al. (2019). Wild origins of macadamia domestication identified through intraspecific chloroplast genome sequencing. Front. Plant Sci. 10, 334. doi: 10.3389/fpls.2019.00334
Sosso J., Zakeel M., and Akinsanmi O. A. (2021). Culturable fungal endophytes in Australian macadamia nursery plants. Australas. Plant Pathol. 50, 739–746. doi: 10.1007/s13313-021-00824-x
Szink I., Davis E. L., Ricks K. D., and Koide R. T. (2016). New evidence for broad trophic status of leaf endophytic fungi of Quercus gambelii. Fungal Ecol. 22, 2–9. doi: 10.1016/j.funeco.2016.04.003
Templeton M. D., Rikkerink E. H., Solon S. L., and Crowhurst R. N.. (1992). Cloning and molecular characterization of the glyceraldehyde-3-phosphate dehydrogenase-encoding gene and cDNA from the plant pathogenic fungus Glomerella cingulata. Gene 122, 225–230.
Wei X., Fu T., He G., Cen R., Huang C., Yang M., et al. (2023). Plant types shape soil microbial composition, diversity, function, and co-occurrence patterns in cultivated land of a karst area. Land Degrad. Dev. 34, 1097–1109. doi: 10.1002/ldr.4518
White T. J., Bruns T., Lee S., and Taylor J. (1990). “Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics,” in PCR protocols A Guide to Methods and Applications, New York: Academic Press, INC. vol. 18. , 315–322.
Wolfe E. R., Dove R., Webster C., and Ballhorn D. J. (2022). Culturable fungal endophyte communities of primary successional plants on Mount St. Helens, WA, USA. BMC Ecol. Evol. 22, 18. doi: 10.1186/s12862-022-01974-2
Wolfe E. R., Kautz S., Singleton S. L., and Ballhorn D. J. (2018). Differences in foliar endophyte communities of red alder (Alnus rubra) exposed to varying air pollutant levels. Botany 96, 825–835. doi: 10.1139/cjb-2018-0085
Zahid M. S., Li D., Javed H. U., Sabir I. A., Wang L., Jiu S., et al. (2021). Comparative fungal diversity and dynamics in plant compartments at different developmental stages under root-zone restricted grapevines. BMC Microbiol. 21, 317. doi: 10.1186/s12866-021-02376-y
Keywords: Ascomycota, fungal diversity, fungal richness, nursery, tree nut
Citation: Khan J, Drenth A and Akinsanmi OA (2025) Mycobiome associated with macadamia plants at different growth stages. Front. Agron. 7:1696145. doi: 10.3389/fagro.2025.1696145
Received: 31 August 2025; Accepted: 01 October 2025;
Published: 23 October 2025.
Edited by:
Aqeel Ahmad, University of Florida, United StatesReviewed by:
Andrew D. Cartmill, Massey University, New ZealandValentim Coelho, Polytechnic Institute of Santarém, Portugal
Copyright © 2025 Khan, Drenth and Akinsanmi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jahangir Khan, amFoYW5naXIua2hhbkB1cWNvbm5lY3QuZWR1LmF1; Olufemi A. Akinsanmi, by5ha2luc2FubWlAdXEuZWR1LmF1