 Gabrielle B. Clark
Gabrielle B. Clark María Díez-León
María Díez-León Rebecca K. Meagher
Rebecca K. Meagher- 1Department of Animal Science and Aquaculture, Dalhousie University Faculty of Agriculture, Truro, NS, Canada
- 2Department of Pathobiology and Population Sciences, Royal Veterinary College University of London, London, United Kingdom
It is standard practice on mink farms in Canada to provide one manipulable environmental enrichment (EE) in each cage to benefit animal health and welfare, and once an enrichment is introduced, it must remain in the cage for the duration of that animal’s life. However, mink might habituate to permanently present enrichments, which reduces interaction with these items over time. In this study, juvenile mink were provided with multiple EEs (mobile and hanging items) that were regularly exchanged to maintain novelty in addition to a standard, permanently present EE (enriched kits; EK). EE use over time, as well as interactions based on EE type, were compared to that of two groups which were standard-housed as juveniles (control; C and enriched at whelping; EW) with access to a permanently available standard enrichment. EK housing was found to significantly increase kits’ EE use compared to standard housing (p< 0.001). Despite some evidence of rapid habituation to novel objects (e.g., decreased use of hanging rope EE in second week of access: p = 0.019), item exchange appeared to have a sustaining effect on exploration of all EEs in the cage, including familiar objects. Thus, it may be beneficial to provide farmed mink with multiple EEs, both novel and familiar, to promote their optimal use and facilitate welfare benefits. Objects with greater manipulability or malleability were also used more by kits than those with fewer possible uses (e.g., pig’s ear versus other, less malleable mobile EEs: p< 0.001; rope versus less malleable hanging EE: p< 0.001). Although this effect was persistent across all periods of observation, it is possible that kit age at time of object introduction or differences in novelty may have contributed to these differences in their use. Further research is required to determine kits’ preference for different EE types when all items are equivalently novel, and when differences in developmental stage are fully accounted for.
1 Introduction
The welfare of captive animals can be improved by providing environmental enrichment (EE) that enables the expression of natural behaviours to a greater extent than would be possible in a barren environment. In applied ethology and animal husbandry, EE refers specifically to biologically relevant stimuli that function to improve animal welfare (Newberry, 1995). Methods of EE can include incorporating fixed or structural elements to the cage environment that add complexity or stimulation for the animals, or inclusion of mobile, manipulable enrichments within the cage (Maple and Perdue, 2013). It is important that enrichments be selected based on their ability to satisfy specific behavioural needs for the target species. For instance, the provision of manipulable objects in the cage that can be chased or chewed has been shown to reduce cortisol levels, fur-chewing, and stereotypic behaviour as well as negative affective states like boredom in American mink (Neogale vison), a mustelid species farmed for their fur (Hansen et al., 2007; Meagher and Mason, 2012; Meagher et al., 2014; Meagher et al., 2017; Polanco et al., 2021). Mink are opportunistic predators who in the wild will mostly consume fish, amphibians, and crustaceans, but will also hunt rabbits and other prey on land (Gerell, 1967). In captivity, mink do not have the same opportunities for hunting; commercial fur farms typically feed their mink a meat-based paste that is placed directly on the mesh wire on top of the cage (National Farm Animal Care Council, 2013). Therefore, it is thought that the provision of mobile EEs may offer an outlet for these unfulfilled behavioural needs (e.g., chasing or chewing) and in this way confer their positive effects on abnormal behaviour, boredom, and stress in mink (Hansen et al., 2007; Meagher and Mason, 2012; Meagher et al., 2014; Meagher et al., 2017; Polanco et al., 2021).
Despite this theorized benefit of mobile EEs, existing research suggests that fixed and unmovable EEs are used by mink more often than those that are mobile and chaseable (Hansen et al., 2007; Axelsson et al., 2009; Ahloy Dallaire and Mason, 2017). In these studies, fixed enrichments seem to be used by mink as additional resting places, which may facilitate escape from the reach of newborn kits and larger males or allow them to obtain a greater vantage point from which to monitor their surroundings (Jeppesen, 2004; Hansen et al., 2007; Axelsson et al., 2009; Buob et al., 2013). It is also possible that fixed enrichments may fulfil a behavioural need for mink to be alone, as only one individual was typically observed using the structure at a given time despite having the capacity for two mink (Jeppesen, 2004). These potential benefits are thought to account for the greater use of fixed EEs (shelves or hanging tunnels and fixed lengths of rope) compared to mobile enrichment types (table tennis balls or play balls and plastic tunnels placed on the floor of the cage; Hansen et al., 2007; Axelsson et al., 2009, respectively).
The results of previous studies, however, may be impacted by several confounding factors. Since mink are inactive for a large portion of their daily time budget, even in the wild (Dunstone, 1993; Clubb and Mason, 2007), it is understandable that they may dedicate more time to the use of shelves or hanging tunnels than to items which require more active use. The mobile enrichments used in previous studies may also have been subject to a habituation effect while fixed items were not impacted to the same extent. In some instances where mobile enrichments are permanently present in the cage, other fixed object types such as hanging rope are being regularly replenished, potentially causing a novelty effect (Hansen et al., 2007). A study measuring the effect of play ball enrichments on mink welfare observed that interaction with play balls subsided within one month of introduction (Jeppesen and Falkenberg, 1990). The novelty of mobile EEs should therefore be renewed regularly by replacing familiarized objects in the cage with novel objects. Moreover, certain mobile EEs such as play balls have only one use (e.g., to be rolled) and therefore seem to attract less interaction over time, whereas other mobile EEs like loose tubes in the cage may offer multiple uses (e.g., rolling, climbing, or hiding), which may increase opportunities for object novelty (Axelsson et al., 2009). A logical next step in this area of research would be to provide mink with a variety of fixed and mobile enrichments and quantify their relative use based on how often they are exchanged and the number and/or nature of possible uses they have.
Despite the potential benefits outlined above, the continuous exchange of all items in the cage could negatively impact welfare as well as research validity, since interruptions in access to familiar items may be a source of stress for mink. Many studies in rodents support that once EE is supplied, access to the items should not be interrupted or revoked because this can induce anxiety or depression-like behaviours in the animal (e.g., Smith et al., 2017; Morano et al., 2019). To prevent this, and its potential confounding effect on further experimentation, studies investigating novelty effects on EE use must be designed with a strategy of object novelty renewal that allows one or more familiar items to remain in the cage while others are continuously exchanged. Correspondingly, it is standard practice on mink farms in Canada to provide at least one manipulable enrichment in each pen, and once an enrichment is introduced, it must remain accessible for the duration of that animal’s life (National Farm Animal Care Council, 2013). Maintenance of this practice may act to prevent the negative effects of enrichment removal on behavioural and physiological measures of stress.
It is also recommended, but not mandated, by the Mink Code of Practice in Canada that enrichments be provided in early life, because access to enrichments during early development is known to have an important role in the prevention of abnormal behaviours such as stereotypic behaviour (National Farm Animal Care Council, 2013). Supplying EE once animals have matured is shown to be less effective at reducing the performance of stereotypic behaviour (Mason, 1993; Ahola et al., 2011; Campbell et al., 2013), and animals show less motivation to gain access to enrichments when introduced to them as adults (Tilly et al., 2010). Animals raised in stimulus-poor conditions may also perceive enrichment introduction later in life as a stressor (i.e., an unpredictable environmental change; Fairhurst et al., 2011) or may have greater novelty-induced fear responses to enrichment that are resistant to habituation (e.g., Cooper et al., 1996; Jones, 2001). However, enrichment introduction typically does not begin on mink farms while false bottoms are in place (i.e., plastic net-like material placed on the floor of the cage to cover gaps between wires, for the duration of whelping, until kits become mobile) due to concerns for safety – for example, play objects may become soiled during this time due to the accumulation of faeces in the cage and pose a health issue (National Farm Animal Care Council, 2013). Interestingly, if EE is provided too early, it can also interfere with maternal care (e.g., a reduction in time spent nursing and increases in non-maternal behaviour were demonstrated in enriched rodent dams; Li et al., 2016). Thus, when provided too early in the postnatal period, EE may in fact have negative consequences for offspring development and welfare. Kits may also not develop the motor skills necessary to interact with enrichments until four weeks of age, and object play was not observed to emerge until seven weeks of age in a previous study (Jonasen, 1987), so it is unclear exactly when these items become useful in mink; there have been limited attempts thus far to provide enrichments to kits this early in development.
Few studies have been conducted to determine the preference of mink for enrichment items with various properties (e.g., fixed or mobile) as well as various degrees of novelty. In the present study, the enrichment-directed behaviours of mink kits with access to a multitude of play objects, some of which were exchanged at regular intervals to maintain novelty (enriched kits; EK), was compared to that of two standard-housed groups with access to one standard enrichment that remained permanently present in the cage (control kits [C] and kits who were enriched at whelping [EW]; these groups were kept distinct due to differential nest environments in their first week of life, which are the focus of a separate paper). It was hypothesized that the EK group would demonstrate greater EE use than standard-housed groups (C and EW) due to the novelty and variety of enrichments available. EE use was also hypothesized to be maintained at a higher level over time in the EK condition than in the standard-housed conditions due to the regular renewal of object novelty. This data may also contribute to identifying trends in EE use as kits age or as object types are exchanged. Moreover, we aimed to replicate previous research findings regarding minks’ increased use of hanging enrichments compared to mobile enrichments, although it was considered whether renewal of novelty of both enrichment types in this study would result in a change in use. There was no hypothesis regarding which of the individual hanging and mobile enrichments provided to EK kits would attract the most use, but these results will be discussed.
2 Methods
2.1 Research ethics statement
This research was approved by the Dalhousie University Faculty of Agriculture Animal Care & Use Committee (#1033575) and the Clinical Research Ethics Review Board of the Royal Veterinary College (URN 2021 2034-3). All housing conditions and husbandry practices in effect were standard for fur farms in Canada (National Farm Animal Care Council, 2013), aside from the extra enrichments provided to some treatments.
2.2 Subjects and housing
Approximately 240 female mink were bred initially to account for potential unsuccessful copulations and/or poor litter health. Litters were excluded from the study if fewer than four kits survived. Once it was confirmed that our desired sample size had survived to the post-weaning stage (a minimum of 30 litters per treatment, each with a minimum of one male-female pair, as indicated by sample size calculations prior to commencement of the study), any additional litters were excluded from further testing (resulting nC, nEW, and nEK = 45, 40, and 37 litters, respectively). Male and female mink selected at breeding for use in the study were balanced across Dark, Mahogany, Pastel, Demi, and Stardust colour types (strains). All dams were housed individually (American mink are solitary in the wild, Dunstone, 1993) indoors at the Canadian Centre for Fur Animal Research (Nova Scotia, Canada) in 75 (L) × 60 (W) × 45 (H) or 40 (H) cm wire-mesh cages with wire shelves, external wooden nest boxes, and a single physical enrichment (plastic ring). Mink were fed twice daily with a meat-based paste and had access to drinking water ad libitum.
Females that bred successfully were randomly assigned to one of three treatment groups. These included two standard-housed conditions in which kits received a standard plastic ring enrichment (control [C] and enriched at whelping [EW]; litters in the latter group were enriched through different environmental modifications than those used in this study, which were in effect from the pre-whelping period to postnatal day eight for the purpose of a separate paper), and one enriched condition (enriched kits; EK) where dams and kits received a standard plastic ring enrichment in addition to a plastic chain fixed to the cage ceiling at 28±7 days post-whelping, when kits were expected to become mobile (reported at approximately four weeks post-whelping; Jonasen, 1987). One additional mobile EE was provided to EK kits at weaning (either a plastic wiffle ball or golf ball); enrichment type was randomly assigned such that approximately half of the EK litters in each whelping cohort (dams who whelped within three days of each other) received a golf ball and half received a wiffle ball. These enrichment items were selected based on proven welfare and/or production benefits (Díez-León et al., 2013; Meagher et al., 2014; Díez-León and Mason, 2016). Groups were counterbalanced for colour type and parity, and cages for each were evenly distributed throughout the barn. All breeding females were housed in standard cages with a single enrichment (plastic ring) prior to the assignment of their respective conditions.
2.3 Post-weaning kit observations
2.3.1 Group housing
Kits were weaned at six weeks post-whelping, at which point the dam was removed from the whelping pen and placed in a separate standard cage. Kits remained in the whelping pen and were housed in groups of four to six (nC, nEW, and nEK = 218, 207, 183 kits, respectively); if there were more than eight kits in a litter, they were split across two pens (applied to ten C pens, three EW pens, and seven EK pens). Kits in EK continued to have access to prior enrichments (i.e., hanging plastic chain and standard ring enrichment) in addition to a second mobile EE introduced at this stage, while standard-housed kits (C and EW) continued to have access to a standard plastic ring. Scan sampling observations were conducted three consecutive days per week for the following four weeks (from weaning to ten weeks post-whelping). Cages were observed three times per day (two morning rounds at approximately 8:30am and 10am and one afternoon round at approximately 1pm; all rounds occurred between an 8:00am morning feeding and a 3:00pm afternoon feeding). Behaviours were recorded through scan samples every 20s over a 2-minute period before moving to the next cage. Kits’ interactions with enrichments were recorded (enrichment use defined as head in contact, licking, or sniffing the EE within 1cm, excluding sleeping with the enrichment), as well as other behaviours not relevant to this paper. The order in which cages were observed was reversed each scoring day to prevent the systematic scoring of some cages earlier than others. A schedule was implemented during this observation phase for the exchange of EK enrichments (see Figure 1). This schedule entailed exchanging golf balls for wiffle balls and vice versa at post-weaning Week 3 (eight weeks post-whelping), while the hanging chain remained present in the cage.
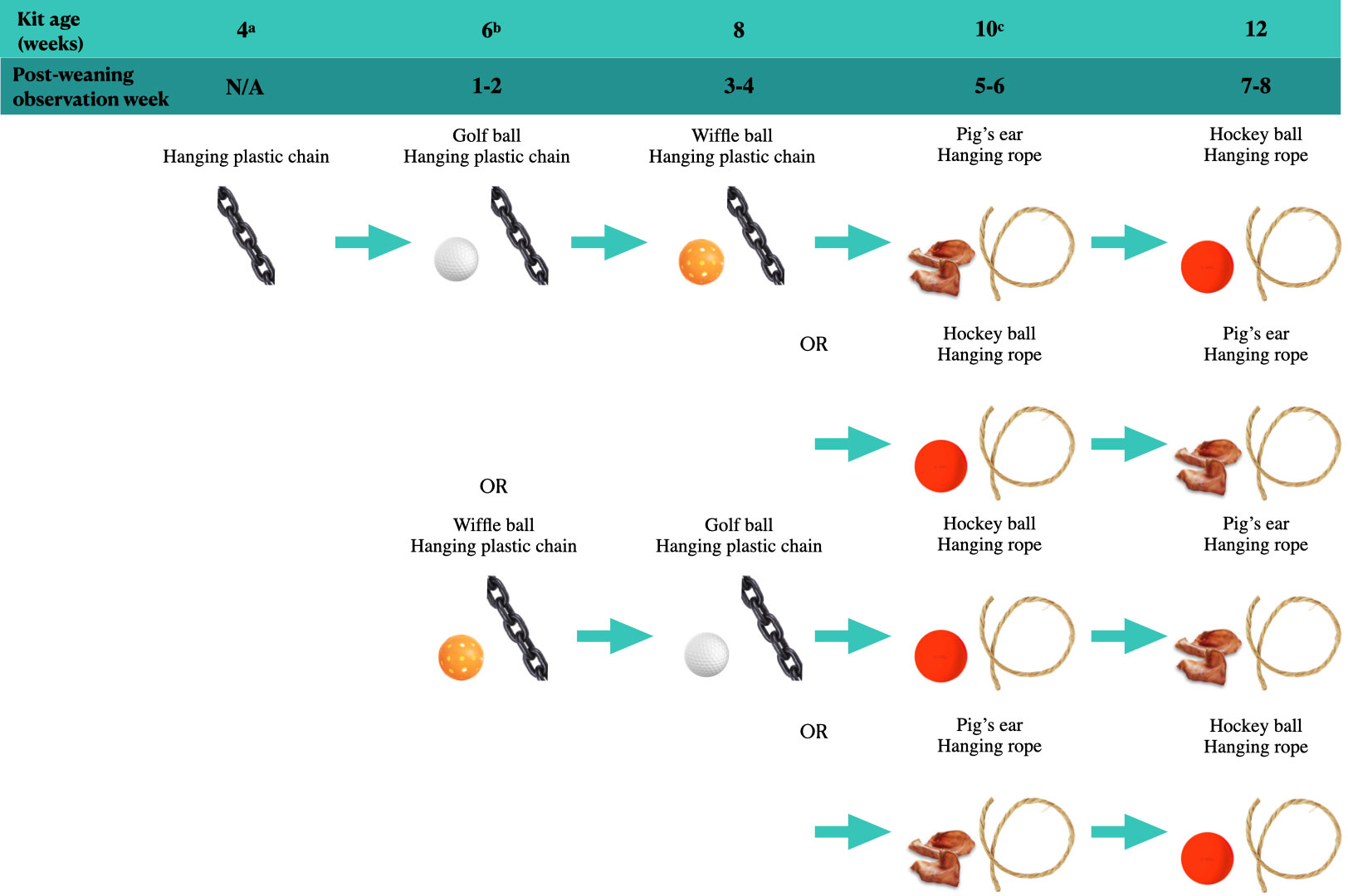
Figure 1 Schedule of enrichments for kits in EK. Enrichment type was randomly assigned such that approximately half of the litters in each whelping cohort (dams who whelped within 3 days of each other) received a golf ball and half received a wiffle ball. Exchange to pig’s ears or hockey balls at 10 weeks was counterbalanced by whelping cohort to account for the effect of age on enrichment use. aKits become mobile. EK kits begin to receive enrichments while dam is still in the pen. bWeaning. Dams moved to separate pens. Enrichments (previous and novel) remain in pen with kits. cKits separated into single- and pair-housing. One male-female pair and one single female from each litter continue to have access to the enrichments for their treatment.
2.3.2 Single- or pair-housing
At ten weeks of age (five weeks post-weaning), kits were moved to single- or pair-housing pens according to standard farm protocol. One male and one female from each litter were chosen for pair-housing and remained in the whelping cage. These male-female pairs (nC, nEW, and nEK = 55, 43, and 44 pairs, respectively) continued to be observed according to the group-housing observation protocol (see Section 2.3.1) and enrichments for EK continued to be exchanged according to the schedule in Figure 1. This included the golf or wiffle balls being exchanged either for street hockey balls or pig’s ears such that approximately half of the EK pens received each object. Hanging chains were exchanged for ropes fixed to the cage ceiling (43 cm or 38 cm long, according to cage height; see Figure 2 for an example of enrichment layout). Since the rope and pig’s ears were depletable enrichments (i.e., kits were able to chew or otherwise deplete them until they were no longer visible in the cage), these objects were replenished as needed at the beginning of each week’s observations (i.e., once weekly). This schedule of enrichment exchange continued until post-weaning Week 8 (thirteen weeks post-whelping), such that mobile enrichments were exchanged for novel enrichments every two weeks or as depletable enrichments (pig’s ears and hanging rope) needed replenishing (see Figure 1). After the conclusion of the post-weaning observation period, EK enrichments remained in the cage for two additional weeks (fifteen weeks post-whelping) until removal, at which point all kits had a standard ring for enrichment.

Figure 2 Example of enrichment for the EK housing condition in Week 5-6 or 7-8 of observation (hanging rope and hockey ball in addition to standard ring enrichment).
2.4 Statistical analyses
Data were formatted for analysis by calculating the average proportion of observations where EEs were in use per kit (henceforth referred to as average EE use) based on the total number of observations (see equation below).
Statistical analyses were conducted with jamovi statistical software (The jamovi project, 2022, v. 2.3.18.0). Significance level was set at p< 0.05. Results were defined as trending when 0.05< p< 0.10. The details of the analytical methods used for each test are as follows:
To assess levels of enrichment use across treatments, average EE use was analysed using Welch’s One-Way ANOVA in the post-weaning Week 1-4 period, Week 5-8 period, and Week 1-8 period (all observation weeks) with treatment as a factor. Data were transformed to fit the normal distribution using the square root transformation; normality was assessed visually using Q-Q plots and Levene’s test was used to assess homogeneity of variance. Post-hoc comparisons were performed using the Games-Howell correction to account for unequal variances. Group means and 95% confidence intervals were back-transformed for presentation.
To assess EE use over time within treatments, average EE use for various enrichment categories (hanging EEs, the ring EE, mobile EEs with and without the ring included, and all EEs collectively) were analysed using linear mixed model analyses with week of observation as a factor and pen number as a cluster variable. EK pens were analysed separately from C and EW pens since trends in use of EK enrichments specifically were of interest; however, treatment was an additional factor for the analysis of ring use over time in standard-housed groups (C and EW). Data were transformed using the square root transformation and visually analysed for normality using Q-Q plots. The Bonferroni correction was used for post-hoc tests to account for the number of comparisons. The Satterwaite method was applied for degrees of freedom to account for violations of homogeneity of variance. Due to emerging differences in EE use between Week 1-4 and Week 5-8 of observation, paired Student’s t-tests were performed for each enrichment category to compare EE use between these periods; these differences were presented when applicable to simplify reporting of results. Means and 95% confidence intervals were back-transformed for presentation.
EE use by item type was analysed using non-parametric Kruskal-Wallis ANOVAs and Dwass-Steel-Critchlow-Flinger pairwise comparisons as this data could not be transformed to fit the normal distribution. EE use in EK pens alone was investigated, as other treatments only had access to the ring enrichment. Average use of hanging vs. mobile EEs and individual hanging and mobile EE sub-types were compared with object type as a factor.
3 Results
3.1 Enrichment use across treatments
Treatment significantly affected the average EE use observed in both the Week 1-4 (F2,65.5 = 26.6, p< 0.001) and Week 5-8 periods (F2,63.6 = 92.0, p< 0.001; see Figure 3). Post-hoc tests revealed that, on average, EE use was observed more often in EK pens than in C and EW pens in Week 1-4 (t60.9 = -6.41, p< 0.001 and t61.3 = -6.85, p< 0.001, respectively) and Week 5-8 (t48.6 = -13.0, p< 0.001 and t48.1 = -13.0, p< 0.001, respectively; see Table 1). This effect persisted across all weeks combined (Week 1-8; F2,63.3 = 112.0, p< 0.001) towards more EE use on average in EK pens compared to C and EW pens (t46.2 = -14.4, p< 0.001 and t49.8 = -14.4, p< 0.001, respectively; see Table 1). There was no difference in EE use between C and EW pens during any of these periods (t67.0 = 0.512, p = 0.866, t66.9 = 0.063, p = 0.998, and t66.0 = 0.578, p = 0.832, respectively).
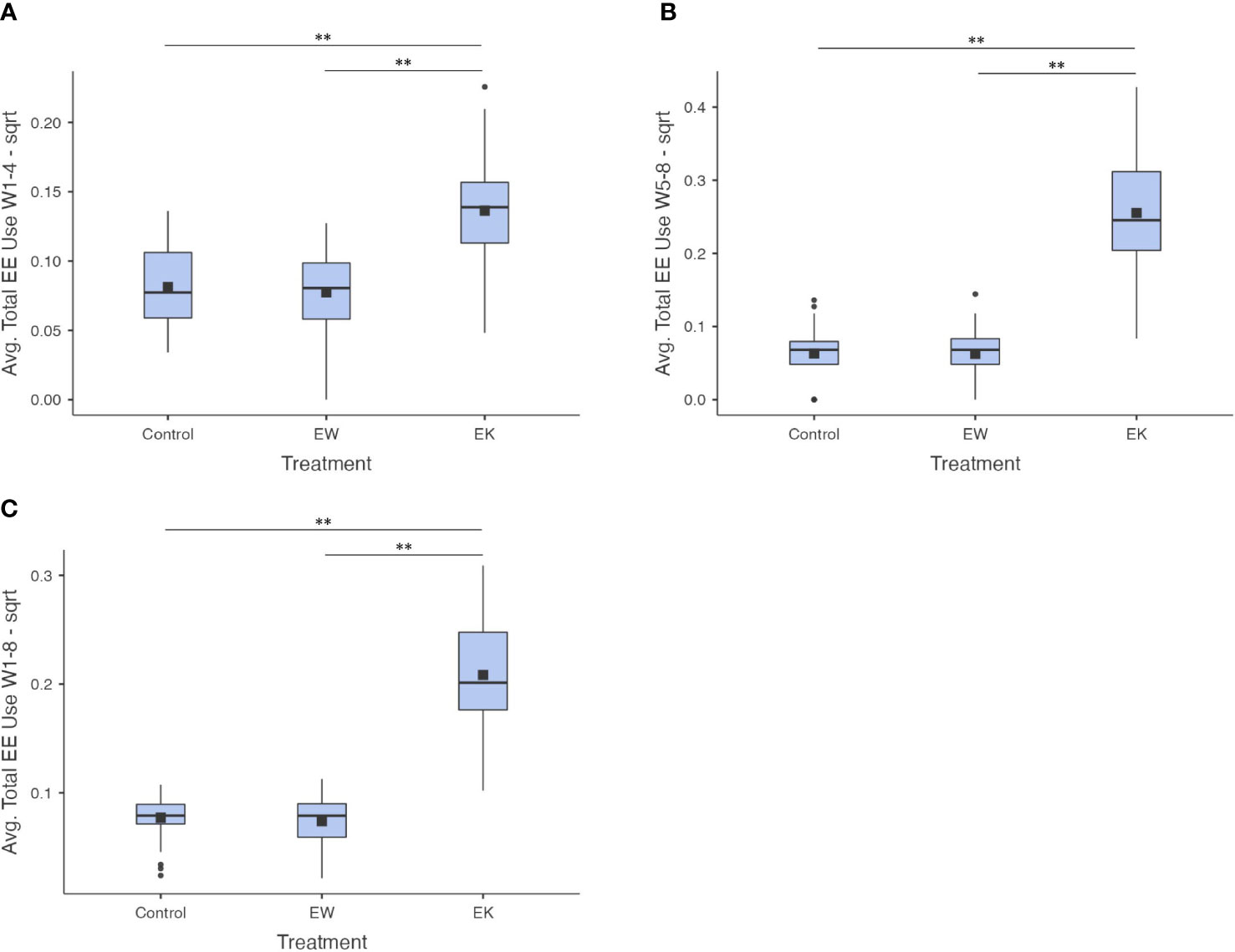
Figure 3 Square root transformed average proportions of observations where enrichments were in use across (A) Week 1-4 of observation, (B) Week 5-8 of observation, and (C) Week 1-8 of observation (all weeks). Black squares show the means; centre lines show the medians; box limits indicate the 25th and 75th percentiles as determined by jamovi software; whiskers extend 1.5 times the interquartile range from the 25th and 75th percentiles, outliers are represented by dots. N = 31, 29, and 32 sample points, respectively. Bars indicate a significant difference with ** indicating p<0.001.
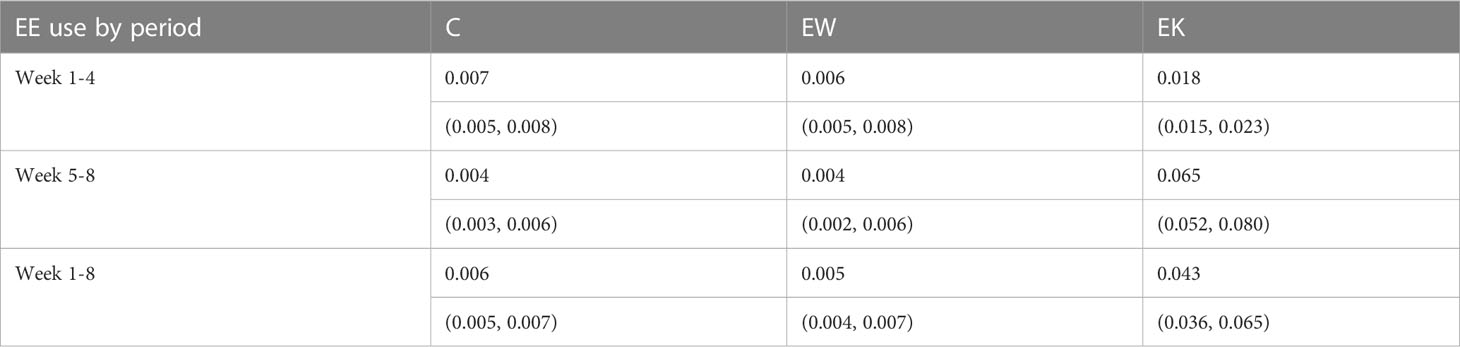
Table 1 Back-transformed average enrichment use (mean with 95% CI in parentheses) across observation periods and housing conditions.
3.2 Enrichment use over time
3.2.1 EK enrichment use over time
Average use of hanging EEs significantly differed by week of observation (F7,262 = 15.9, p< 0.001; see Figure 4A). Generally, hanging EE use was higher in the Week 5-8 period than in the Week 1-4 period (t33 = -3.553, d = -0.609, p = 0.001; see Table 2). In addition to differences between Week 1-4 and Week 5-8 collectively, hanging EEs were used more in Week 5 than in Week 6 (t256 = 3.435, p = 0.019), Week 7 (t256 = 8.238, p< 0.001), and Week 8 (t256 = 7.917, p< 0.001), respectively. Use of hanging EEs was also significantly higher in Week 6 than in Week 7 (t256 = 4.803, p< 0.001) and Week 8 (t256 = 4.482, p< 0.001; see Table 2).
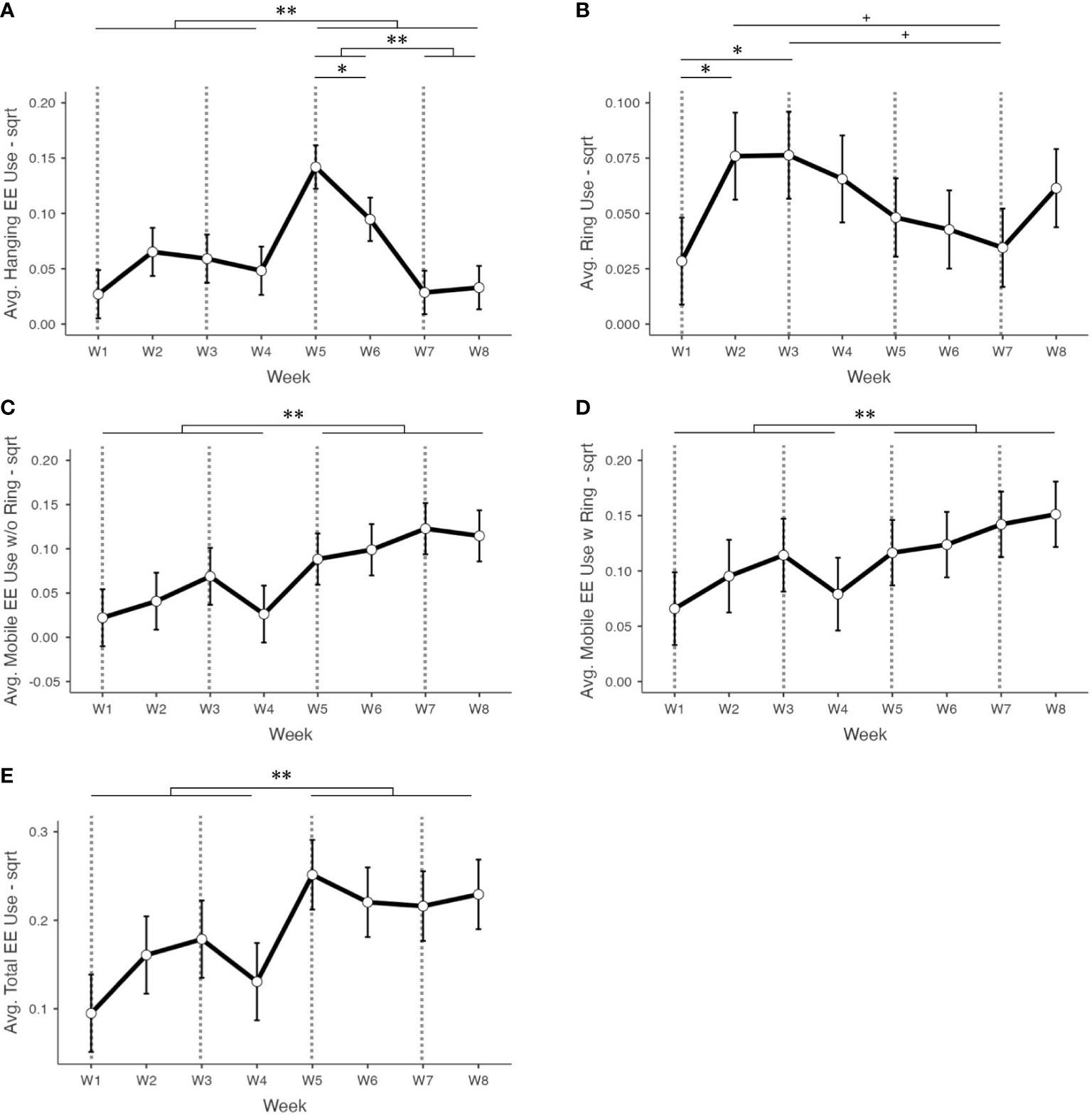
Figure 4 Square root transformed average proportions of observations where the following enrichments were in use in EK pens across weeks of post-weaning observations: (A) hanging EE, (B) ring EE, (C) mobile EEs without the ring, (D) mobile EEs with the ring, and (E) all EEs. Dashed lines indicate the exchanging of enrichments according to the schedule in Figure 1. Bars around means represent 95% CIs. By week, N = 34, 34, 34, 34, 42, 42, 42, and 42 sample points, respectively. Bars indicate a significant difference with ** indicating p<0.001, * indicating p<0.05, and + indicating 0.05<p<0.10. Differences indicated within W1-W4 and W5-W8 represent results from linear mixed model analyses, while differences indicated between W1-W4 and W5-W8 collectively represent results from paired Student’s t-tests.
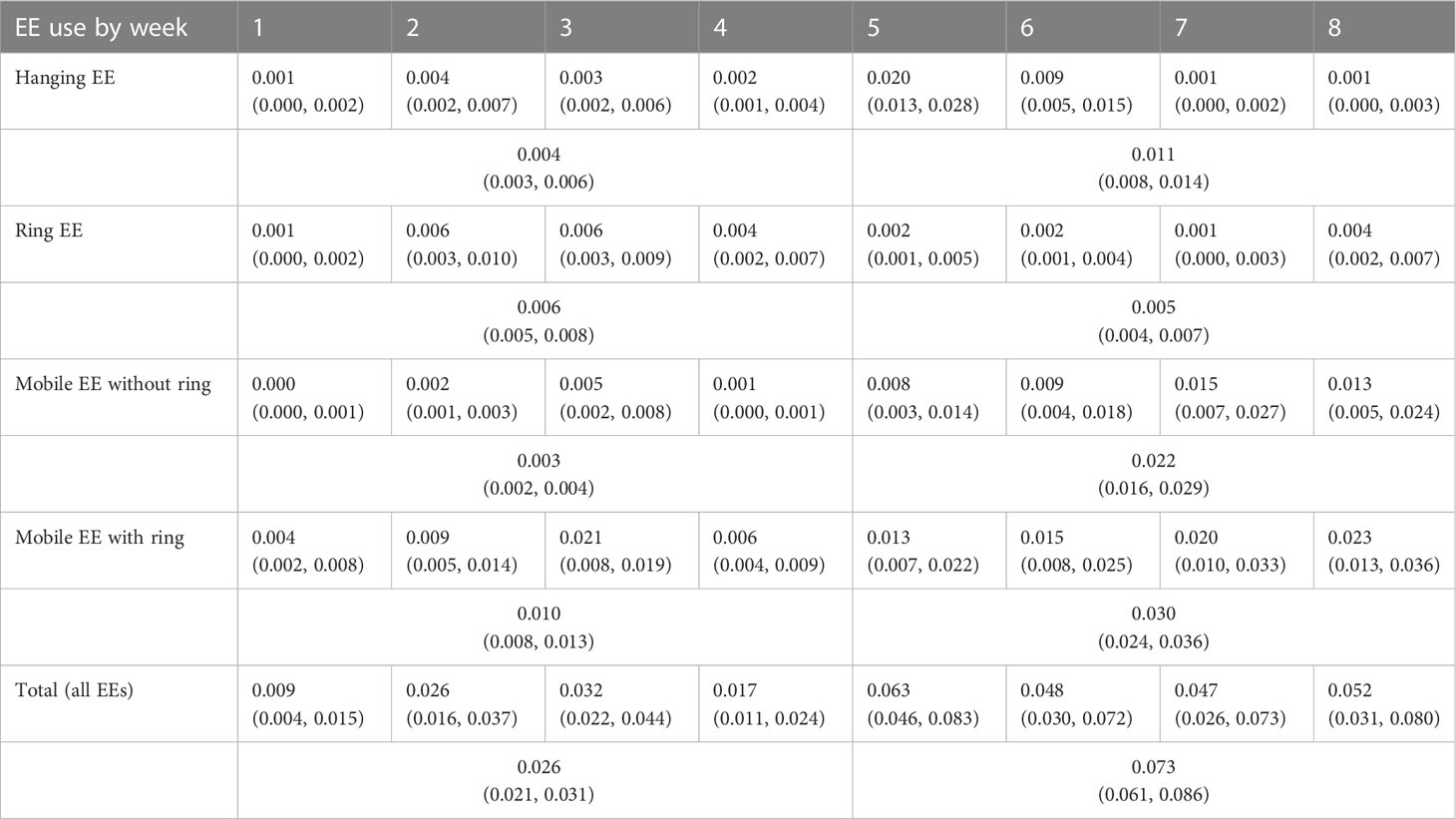
Table 2 Back-transformed EK enrichment use (mean with 95% CI in parentheses) by item type and week of observation.
Average ring use in EK pens also differed by week of observation (F7,260 = 3.61, p< 0.001; see Figure 4B). However, the only differences occurred between Week 1 and Week 2 towards more use in Week 2 (t257 = -3.375, p = 0.024), and between Week 1 and Week 3 towards more use in Week 3 (t257 = -3.403, p = 0.022; see Table 2). There were trending differences in ring use between Week 2 and Week 7 towards more use in Week 2 (t264 = 3.092, p = 0.062) and between Week 3 and Week 7 towards more use in Week 3 (t264 = 3.122, p = 0.056; see Table 2). There was no difference in the EK group’s ring use between Week 1-4 and Week 5-8 collectively (t33.0 = 0.869, d = 0.149, p = 0.391).
Use of additional mobile EEs in EK pens (i.e., excluding ring use) also varied by week of observation (F7,296 = 6.45, p< 0.001; see Figure 4C). However, aside from higher use in Week 5-8 collectively compared to Week 1-4 collectively (t33.0 = -6.690, d = -1.147, p< 0.001; see Table 2), average use of additional mobile EEs did not vary significantly within these periods (see Supplementary Table S1.3 for individual week comparisons). Similarly, with ring use included, mobile EE use in EK pens varied significantly by week of observation (F7,296 = 3.42, p = 0.002; Figure 4D), but these differences occurred primarily between Week 1-4 and Week 5-8 towards higher use in Week 5-8 (t33.0 = -6.256, d = -1.073, p< 0.001; Table 2). There were no significant differences in average mobile EE use within Week 1-4 and Week 5-8 (see Supplementary Table S1.4 for individual week comparisons).
Total average EE use in EK pens, which includes all hanging and mobile enrichments, also differed by week of observation (F7,261 = 6.45, p< 0.001) but similarly only varied between weeks in the Week 1-4 period compared to the Week 5-8 period and not within these periods (Figure 4E; see Supplementary Table S1.5 for individual week comparisons). EEs were generally in use more in Week 5-8 collectively than in Week 1-4 collectively (t33.0 = -7.102, d = -1.218, p< 0.001; see Table 2).
3.2.2 Standard-housed enrichment use over time
There was no effect of treatment (consisting of C and EW; F1,84.4 = 1.219, p = 0.273) or interaction effect between treatment and week of observation (F7,561.1 = 0.667, p = 0.701) on average EE use in standard-housed pens, i.e., use of the standard ring enrichment (Figures 5A, B). However, there was a significant effect of week of observation on EE use in these treatments collectively (F7,561.1 = 10.480, p< 0.001; Figure 5B): average ring use was higher in Week 2, Week 3, and Week 4 compared to Week 1 (t552 = -5.234, p< 0.001, t552= -6.289, p< 0.001, and t552 = -4.439, p< 0.001, respectively; see Table 3). There were no further differences in ring use within the Week 1-4 and Week 5-8 periods (see Supplementary Table 2 for remaining comparisons), though there were significant decreases in ring use in select weeks in the Week 5-8 period compared to the Week 1-4 period; this is represented as lower collective average ring use in Week 5-8 compared to Week 1-4 for both EW pens (t35.0 = 2.46, p = 0.019) and C pens (t33.0 = 2.07, p = 0.046; see Table 3).

Figure 5 Square root transformed average proportions of observations where the ring EE was in use across weeks of post-weaning observations for (A) each treatment separately and (B) Control and EW (standard-housed treatments) combined. Bars around means represent 95% CIs. By week, data points consist of an average across N = 34, 34, 34, 34, 34, 34, 34, and 34 pens for Control, N = 47, 47, 47, 47, 49, 49, 49, and 49 pens for EW, and N = 34, 34, 34, 34, 42, 42, 42, and 42 pens for EK, respectively. Bars indicate a significant difference with * indicating p<0.05 and ** indicating p<0.001. Differences indicated within W1-W4 and W5-W8 represent results from linear mixed model analyses, while differences indicated between W1-W4 and W5-W8 collectively represent results from paired Student’s t-tests.
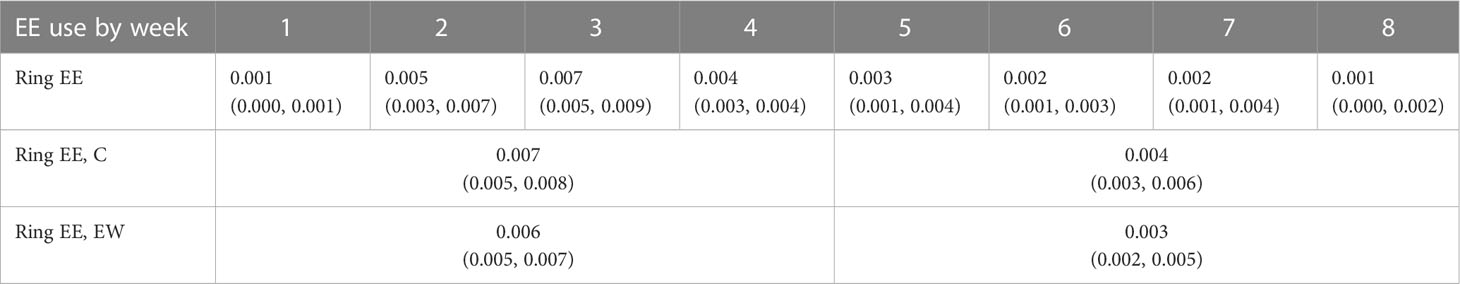
Table 3 Back-transformed standard-housed enrichment use (mean with 95% CI in parentheses) by week of observation.
3.3 EK enrichment use by object type
3.3.1 Enrichment use between mobile and hanging items
EE use varied significantly by category (hanging or mobile; x2 = 42.9, ε2 = 0.622, p< 0.001; Figure 6). Mobile enrichments (0.037 ± 0.019) were observed in use more often on average than hanging enrichments (0.009 ± 0.006).
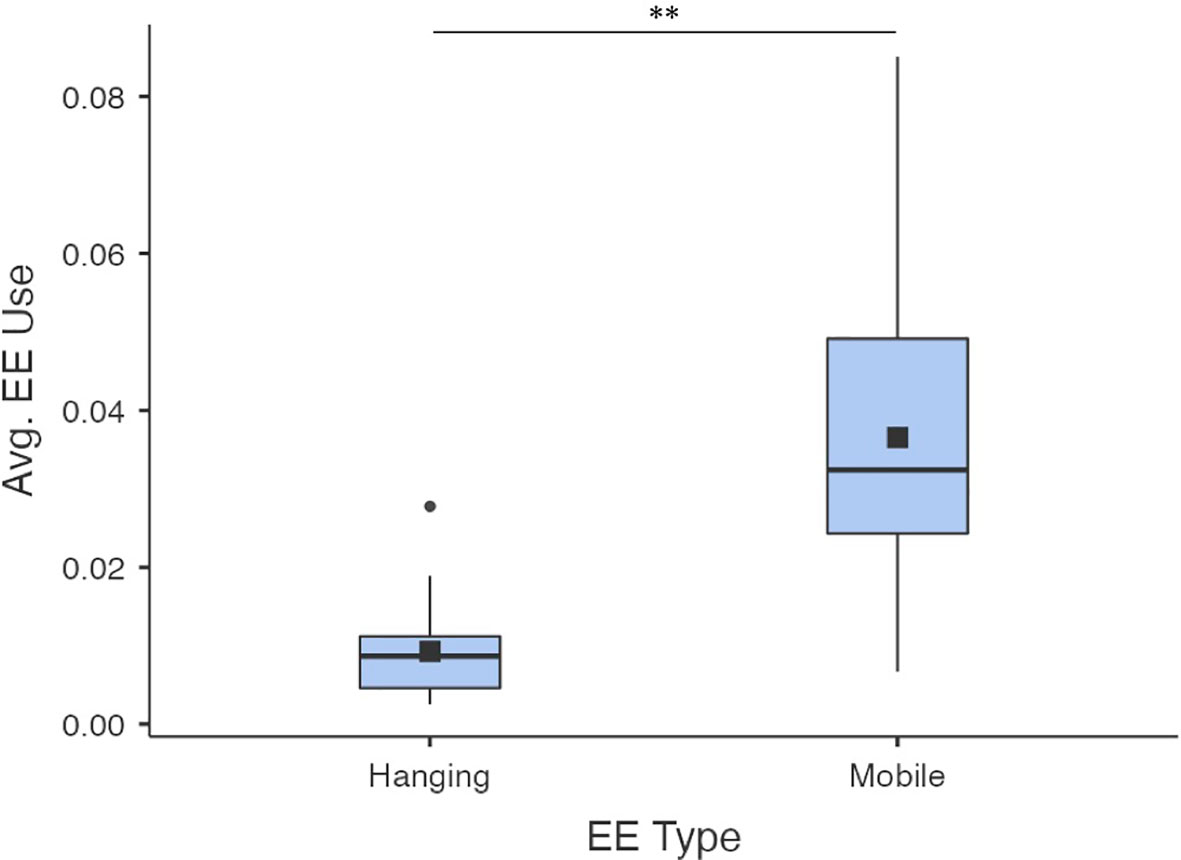
Figure 6 Average proportions of observations where hanging or mobile enrichments were in use (calculated per pen across all weeks). Data is for EK kits only (other conditions had only standard ring enrichment). Black squares show the means; centre lines show the medians; box limits indicate the 25th and 75th percentiles as determined by jamovi software; whiskers extend 1.5 times the interquartile range from the 25th and 75th percentiles, outliers are represented by dots. N = 35 and 35 sample points, respectively. Bars indicate a significant difference with ** indicating p<0.001.
3.3.2 Enrichment use within mobile and hanging items
EE use also varied by sub-type of mobile enrichment (x24 = 99.5, ε2 = 0.510, p< 0.001; Figures 7A, B) and hanging enrichment (x2 = 15.1, ε2 = 0.199, p< 0.001; Figure 8). The ring (0.006 ± 0.004) and the wiffle ball (0.006 ± 0.006) were used significantly more on average than the golf ball (0.0.002 ± 0.003; W= -6.95, p< 0.001 and W = 4.33, p = 0.012, respectively) and the hockey ball (0.002 ± 0.005; W = -7.35, p< 0.001 and W = -5.35, p< 0.001, respectively). Use of the ring and wiffle ball did not differ (W = -1.82, p = 0.699), nor did use of the golf ball and hockey ball (W = -1.81, p = 0.703). The pig’s ear (0.048 ± 0.033) was used more often on average than all other mobile EEs (compared to ring: W = 9.29, p< 0.001; compared to golf ball: W = 9.43, p< 0.001; compared to wiffle ball; W = 8.84, p< 0.001; and compared to hockey ball: W = 10.21, p< 0.001; Figure 7A). The rope was used more on average than the chain (0.014 ± 0.012 and 0.005 ± 0.004, respectively; W = 5.50, p< 0.001; Figure 8).
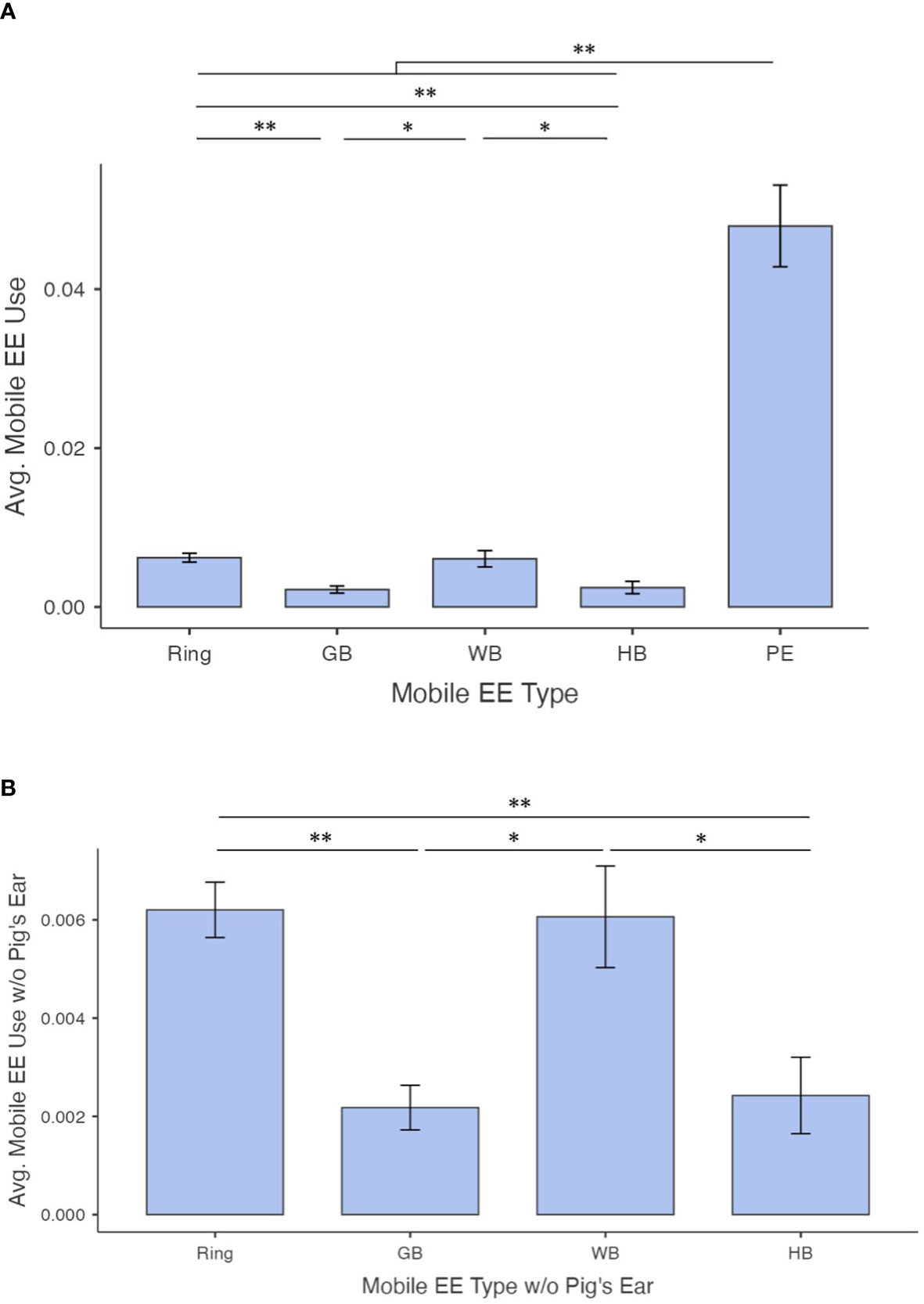
Figure 7 Average proportions of observations where enrichments were in use according to (A) enrichment type, including all mobile enrichments (Ring, standard ring enrichment; GB, golf ball; WB, wiffle ball; HB, hockey ball; and PE, pig’s ear) and (B) mobile enrichment type excluding the pig’s ear. Data is for EK kits only (other conditions had only standard ring enrichment). Error bars represent standard error. N = 42, 35, 35, 42, and 42 sample points, respectively. Bars indicate a significant difference with * indicating p<0.05 and ** indicating p<0.001.
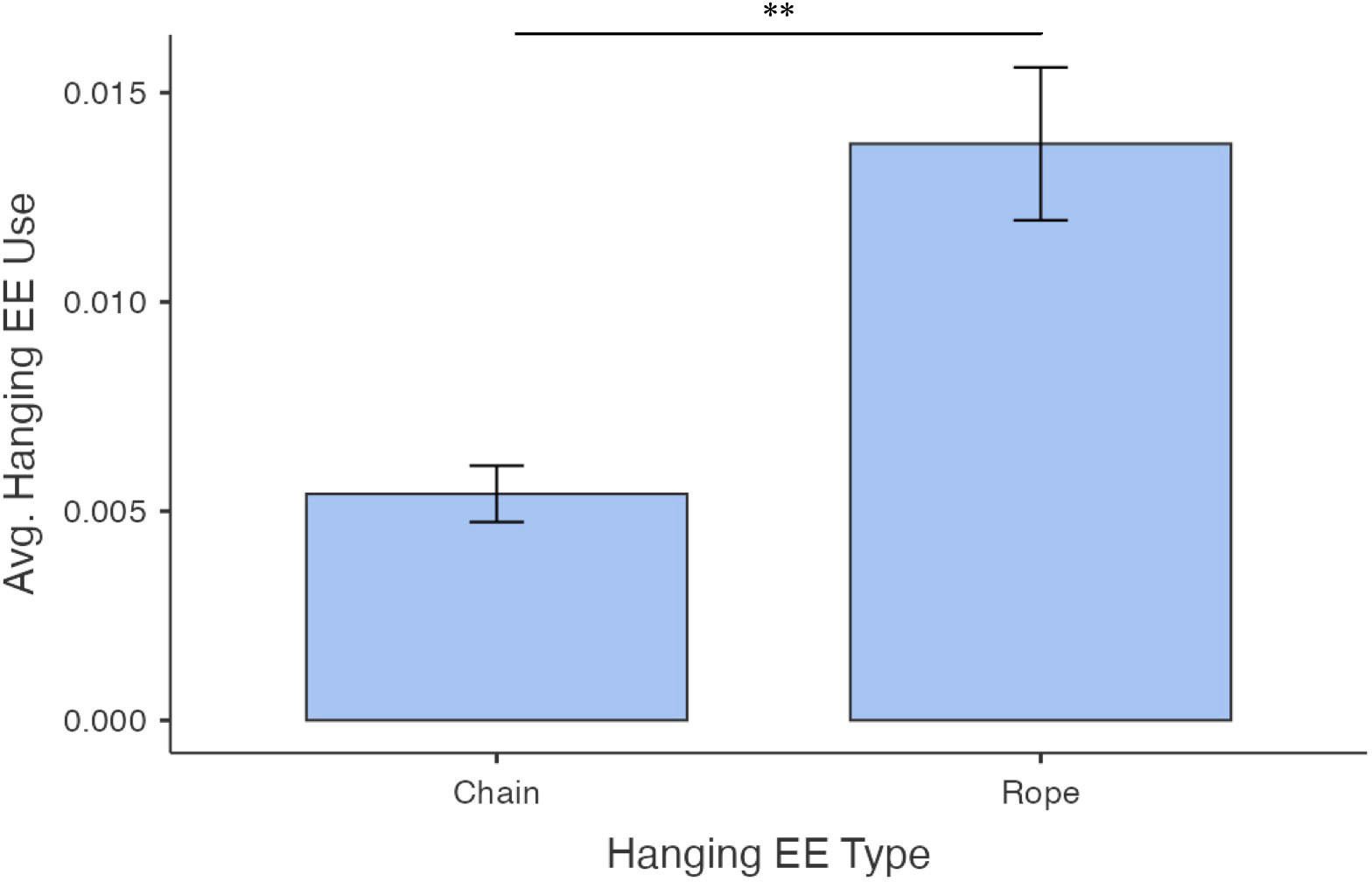
Figure 8 Average proportions of observations where enrichments were in use (calculated per pen across all weeks) according to hanging enrichment type. Data is for EK kits only (other conditions had only standard ring enrichment). Error bars represent standard error. N = 35 and 42 sample points, respectively. Bars indicate a significant difference with ** indicating p<0.001.
4 Discussion
Overall, the findings of this study support our hypothesis that kits in enriched housing (i.e., with access to multiple EEs, both novel and familiar) would demonstrate greater enrichment use than kits in standard housing (i.e., with access to only one familiar EE). There was significantly more enrichment use observed in the EK group compared to the C and EW groups across the entire post-weaning period; therefore, the enrichment provision strategy applied in this study was successful in promoting greater interaction with enrichments. Based on these results, it may be advisable that at minimum, more than one enrichment be provided in mink pens to increase their use. This concept is supported by previous studies in rodents. Although EE for rodents is typically comprised of more than one item per cage due to the antagonistic interactions that can occur when only one enrichment is provided for group-housed rodents (i.e., fighting for resources/EE devices; Van Loo et al., 2002; McQuaid et al., 2012; McQuaid et al., 2013; Weber et al., 2017; McQuaid et al., 2018), it was previously unclear whether access to multiple of the same enrichment is more beneficial than access to multiple different items. In a study where cages were either supplied several of the same EE or a variety of EEs, greater enrichment use in addition to more pronounced effects on welfare measures were observed in cages with a diverse selection of enrichment objects (Abou-Ismail, 2011). This finding may be especially relevant if the enrichments provided have different properties and can be interacted with in different ways, as was the case in the present study. It was proposed by Zhang et al. (2022) in a study with dairy calves that provision of multiple items that can be interacted with in different ways (e.g., stationary brushes, plastic chains, dry teats, and hay nets) may satisfy different intrinsic behavioural motivations of calves (e.g., grooming, suckling, and feed intake), thus increasing overall item interaction in a cumulative way rather than specifically motivated interaction with one type of object.
As such, the simultaneous provision of both hanging and mobile enrichments may have contributed to item interaction in this study since these categories of items serve different uses. We did not expect these categories to be used equally, but contrary to our predictions, mobile enrichments were used significantly more than hanging enrichments. This may be because mobile enrichments were exchanged more frequently than hanging enrichments (every two weeks as opposed to every four weeks), and kits’ habituation to each may have differed accordingly. An alternate explanation may be that there were a greater number of mobile EEs available in the cage to interact with; since there were two mobile enrichments present in enriched cages, more than one kit could easily play with one at a given time, whereas there was only one hanging enrichment provided at a time. However, multiple kits were sometimes observed interacting with the rope or chain (e.g., climbing, tugging, or chewing) at once, since the items were large enough to allow this. This explanation would also only apply to the group-housing phase, since there were more enrichments present in the pair cages than there were kits, yet the preference for mobile enrichments persisted. Alternatively, it is possible that the hanging enrichments provided did not share the same functional advantage as enrichments categorized this way in previous studies. The hanging items in such studies (e.g., shelves and tunnels) were used extensively due to their functionality as additional resting places, or opportunities to reach a higher vantage point and/or be alone in the cage, whereas mobile enrichments such as play balls and loose tubes were used to a lesser extent and were primarily chased or chewed (Hansen et al., 2007; Meagher and Mason, 2012; Meagher et al., 2017; Polanco et al., 2021). The hanging lengths of rope or chain used in this study more closely resemble the latter category of enrichments based on their use for climbing and/or chewing, but they are not mobile to the same extent as loose items on the cage floor. These results demonstrate that it may be appropriate in future studies to further subdivide the category of ‘fixed’ or ‘hanging’ enrichments into fully fixed structural enrichments, which provide an additional resting space, versus fixed but manipulable objects which can only be climbed, chewed, etc., when investigating the effects of enrichment properties on their use.
Within mobile enrichments, the pig’s ear was used most often, followed by the standard ring enrichment and the wiffle ball. The hockey ball and golf ball received the lowest amounts of use and were interacted with at similar levels despite being provided in different post-weaning periods. Although specific methods of enrichment interaction were not recorded, anecdotally, we observed that the ring was used in as many as four different ways by kits; these include being rolled, rested in (with the ring in either a vertical or horizontal position; rings were large enough to prevent mink from getting stuck), chewed, or climbed on. In contrast, the golf ball and hockey ball were only observed being rolled and/or climbed. The wiffle ball similarly had more possibilities for interaction compared to other play balls, but fewer uses than the ring; in addition to being rolled and/or climbed, the wiffle balls were often seen being dragged or carried in kits’ teeth due to their light weight and the holes perforating their surface. Both the golf ball and hockey ball were heavier than the wiffle ball and did not have surfaces that facilitated being carried in this way. A similar effect was observed within hanging enrichments: the rope was observed in use more often than the chain, potentially because the rope could be climbed, chewed, unravelled, and torn down whereas the chain could only be climbed and chewed.
The relative popularity of the standard ring enrichment is interesting because it was present in the cage the most consistently and for the longest duration of time in this study, yet its use in EK pens remained relatively consistent across all weeks of observation. This suggests that despite not being regularly exchanged, greater opportunities for object novelty may have been offered by the ring due to its more numerous uses (Axelsson et al., 2009) than by novel objects with fewer uses. This finding is similar to that of a previous study regarding enrichment novelty versus enrichment complexity in rats (Abou-Ismail and Mendl, 2016). The authors found that novelty, in the sense that multiple replicates of the same object were exchanged for multiple of another object weekly, was less influential than the characteristics of the objects available at any given time, even if one or more of these objects remained accessible throughout the entire study period (Abou-Ismail and Mendl, 2016).
These authors also proposed that increased use of EE in the condition with more diverse, permanently present objects may be due to the lack of control and predictability imposed by frequent exchange of enrichments in the cage environment, which can adversely affect welfare (Abou-Ismail and Mendl, 2016). It is important to consider that, firstly, rotating of enrichments could be considered enrichment removal in the sense that the animal loses access to an item they have come to find rewarding, even though another enrichment arrives in its place. Such removal may be perceived as a negative event or environmental challenge over which the animal has no control, similar to other routine changes to the animal’s environment such as cage cleaning that invoke physiological and behavioural responses of stress (Morgan and Tromborg, 2006). There is evidence that uncontrollable events made predictable by signalling with a cue or temporal consistency are less aversive than uncontrollable, unpredictable events, suggesting that predictability allows a perception of control over the event and thus enhances the animals ability to cope (Bassett and Buchanan-Smith, 2007). Complete enrichment removal evokes a depression-like phenotype in captive animals, including helplessness behaviour (Smith et al., 2017), which has been linked to uncontrollability and/or unpredictability of events that directly or indirectly impact the animal (Burger and Arkin, 1980; Morgan and Tromborg, 2006). Meanwhile, events or stimuli perceived as positive (such as food presentation and other food-related rewards) appear to have enhanced welfare benefits when somewhat unpredictable, potentially reflecting the probabilistic nature of food acquisition in the wild (de Oliveira and da Silva Vasconcellos, 2022). As far as we are aware, little research has been conducted regarding whether control or predictability of enrichment access or removal has benefits for item use and measures of welfare. While the schedule of EE exchange used in this study may have offered some predictability over a longer period, it is unlikely that four bi-weekly enrichment rotations were sufficient for mink to learn this schedule, especially as there were no obvious cues or signals associated with the exchange; though, some uncertainty may have arisen about the constancy of these enrichments as the study progressed. When objects are permanently present, as the ring was in this study, Abou-Ismail and Mendl (2016) suggest that opportunities for the animal to exert control over their environment may be increased, thus improving their welfare and positively impacting other factors associated with enrichment use (Abou-Ismail and Mendl, 2016). In fact, it has been proposed that controllability may be a more important aspect of EE than the complexity of the enrichment objects (Sambrook and Buchanan-Smith, 1997). Either of these hypotheses (i.e., number of item uses or degree of controllability) are plausible regarding kits’ use of the ring enrichment.
However, these hypotheses do not explain why the pig’s ear was used significantly more than all other mobile enrichments. The pig’s ear could be chewed but was not able to roll, and therefore be chased, as effectively as the ring or play ball EEs. In this sense, it had a limited number of uses compared to the ring and wiffle ball enrichments. Moreover, it was replaced/exchanged more frequently than all other mobile items. Replacements were made once per week within the two-week period of access to the pig’s ear if a pen had depleted theirs entirely, which may have contributed an additional source of novelty, since although the enrichment’s properties would not have been novel to the kits there would have been a brief period of absence before its re-introduction. The same applies to the rope, as it was replaced on a weekly basis if a pen had torn theirs down from its fixture. As such, the additional novelty component of the pig’s ear compared to other mobile enrichments, and of the rope compared to the chain, may have promoted their greater use. However, it is also possible that the pig’s ear may have satisfied an appetitive or food-related motivation for the kits, as their gradual disappearance from the cage infers that some or most of the enrichment may have been ingested. It is well-established that diets varying in textures or tastes, introduction of novel edible enrichments, or stimulation via foraging tasks can be a source of enrichment for captive carnivores (Bashaw et al., 2003; Watters et al., 2011; Ruskell et al., 2015; Riggio et al., 2019). If the pig’s ear can be considered feed enrichment, interactions with the item may have been reinforced by its incentive value as food. Studies have also found that species-specific edible enrichments were used more than nonedible enrichments by captive animals that forage for their food rather than hunt (Elmore et al., 2012; Roy et al., 2019; Zhang et al., 2022).
The previously mentioned concept of controllability may also apply to the observed differences in pig’s ear and rope use compared to other enrichments: there are suggested to be different ‘grades’ of enrichment controllability according to the sophistication of their cause-effect relationships, or ways in which they can be interacted with, even in contexts where all enrichments are novel to the cage environment (Sambrook and Buchanan-Smith, 1997). According to this concept, fixed objects (defined as the animal being able to move only in relation to the object) are claimed to have less controllability than moveable objects (defined as the animal and object moving with respect to each other and the enclosure), which in turn both have less controllability than malleable objects (defined as an action being applied to a point on an object resulting in an effect or change to that same point on the object; Sambrook and Buchanan-Smith, 1997). By this logic, the malleable enrichments provided in this study (i.e., the pig’s ear and rope, which were both destructible) may have been appealing to kits in offering a greater ability to exert control or physically manipulate the environment. Destructible or deformable enrichments have similarly been found to be preferred in other species (in chimpanzees: Videan et al., 2005; in pigs: Van de Weerd et al., 2003; Perre et al., 2011; Courboulay, 2014). Like pig’s ears, other destructible animal products (e.g., hide strips) have been shown to be very attractive to mink in pilot studies (e.g., informal pilot data from Meagher et al., 2014), thus this nature of enrichment may have potential if robust and inexpensive versions are identified. Cost and durability are important considerations for edible or easily destructible enrichments since informal observations indicated that some pens in the present study depleted their pig’s ears and/or hanging ropes in as little as two days, although other pens maintained the same pig’s ear and rope for 14+ days without markedly depleting them.
Regarding trends in enrichment use over time, it can be qualitatively observed that average use of available enrichments was higher in each week of observation in EK pens compared to standard-housed pens, and there was more evidence of habituation to enrichment in standard-housed pens compared to EK pens: ring use was lower in Week 5-8 collectively compared to Week 1-4 collectively in standard-housed pens, while ring use in EK did not differ significantly across these periods (in fact, use of other enrichment categories was increased compared to prior weeks). This aligns with our hypothesis that enrichment use would be maintained at a higher level over time in the EK condition. The twelve-week period of access to the ring in this study may therefore have been sufficient to observe habituation in housing conditions where it was the only object available in the cage, which reflects previously demonstrated habituation to permanently present mobile items within one month of introduction (e.g., Jeppesen and Falkenberg, 1990). However, despite less evidence of habituation in EK pens, average weekly use of the ring enrichment occurred in similar amounts between standard- and enriched-housed kits and followed similar trends when looking at individual weeks in the early and late observation period (e.g., significant or trending differences towards lower use in later weeks compared to earlier weeks). Notably, ring use in standard and enriched housing followed divergent trends in Week 8 of observation where an unexplained increase in ring use occurred in EK pens (though non-significant), which is likely why EK ring use did not differ between Week 1-4 and Week 5-8 collectively. It is possible that with a prolonged observation period, habituation to the ring may have become evident in the EK condition as well.
Use of other EK enrichments was more variable over time. Most of the significant variation in enrichment use for EK pens occurred according to the properties of the enrichments provided in different periods; EE use in Week 5-8 was generally greater than use in Week 1-4, which is likely due to the provision of the rope and pig’s ear in the latter period, though it should be noted that housing changes occurred during this phase as well (i.e., split into pair-housing) and this may have had some effect. Within Week 1-4 and 5-8, there were slight, but non-significant, increases in mobile EE use in weeks where mobile EEs were exchanged compared to the following and preceding weeks. This may demonstrate a positive effect of object novelty on EE use, but this novelty effect appeared to diminish rather quickly depending on the object(s) introduced. In some cases, EE use declined within one week of novel object introduction; for instance, average use of hanging EEs declined significantly between the first and second week of access to the hanging rope enrichment. Interestingly, the same was not demonstrated for mobile EEs in the same week: when wiffle balls or golf balls were exchanged for hockey balls or pig’s ears, mobile EE use remained elevated in the week following the exchange.
These trends in EE use around times of object exchange may reflect an initial increase in object exploration due to novelty, followed by a return to interaction with familiar objects if the properties of the novel object are not sufficiently appealing to kits. A similar effect was seen in a recent study where rat cages were modified either by the addition of new objects, an increase in size to existing objects, or added complexity to existing objects (Pisula et al., 2021). In both the object size modification and object complexity modification conditions, there was more initial exploration of the changed object followed by a shift to exploration of unchanged objects. The authors theorized that if an environmental modification is not sufficiently complex, all necessary information about the object will be incorporated rather quickly without satisfying the animal’s need for sensory and informational stimulation, resulting in lingering arousal and redirection of exploratory behaviour to other aspects of the environment (Pisula et al., 2021). This may demonstrate that certain objects provided in this study (namely, the pig’s ear) offered sufficient complexity to activate more advanced, long-term exploration and prevent a redirection of exploration to familiar objects. This may also explain the slightly increased use of the ring enrichment in EK pens in Week 8 and its correspondence to somewhat decreased use of other EEs, which may reflect a return to interaction with familiar objects when novel objects have been sufficiently explored. That said, it can be concluded that maintenance of object novelty was effective at maintaining kits’ interest in the items present since EE use generally showed little fluctuation within Week 1-4 and 5-8 in EK pens, even if this attention was not always directed towards novel objects.
We also aimed to identify a time at which enrichment provision becomes useful in kits, or whether there are changes in enrichment use as kits age. The increase in use of enrichments from Week 5-8 (age 10-13 weeks) relative to Week 1-4 may reflect changes in play style preferences as mink age; social play in mink is known to emerge around five weeks of age and increase across subsequent weeks (Jonasen, 1987; Brink and Jeppesen, 2005), while object play, similar to enrichment use as defined here, emerges later. This may explain a transition to greater object play later in development. However, a previous study regarding ontogeny of play in mink found that all forms of play, including object play, peaked between eight-ten weeks of age and declined thereafter (Vinke and van Leeuwen, 2005). Although trends in other forms of play (e.g., social, locomotor) were not evaluated in this study, a decline in object play after eight to ten weeks was not observed. Rather, overall use of enrichments in EK was increased at ten weeks of age compared to prior weeks and remained consistent until thirteen weeks of age. This aligns more with the finding of Ahloy Dallaire and Mason (2016) that object play did not decrease to the same extent as social play between ten-eleven weeks and sixteen-twenty weeks.
However, any potential age effects on EE use in this study are difficult to disentangle from effects of enrichment properties on EE use. For example, it could be speculated that there was an age effect in EK kits regarding the pig’s ears’ relative popularity compared to other mobile enrichments, since it was provided in either Week 5-6 or Week 7-8 of the post-weaning period (when kits were ten-eleven or twelve-thirteen weeks old), and there was significantly more enrichment use seen during these times compared to the Week 1-2 and Week 3-4 observation period (when kits were six-seven or eight-nine weeks old). However, the hockey ball was also an enrichment provided in either Week 5-6 or Week 7-8, and use of the hockey ball was not found to be significantly different than that of the golf ball, while the wiffle ball was used significantly more than the hockey ball; these were both enrichments provided in earlier weeks. Similarly, levels of hanging enrichment use were roughly equivalent between Week 1, 2, 3, and 4 and Week 7 and 8, which may reflect a return to ‘normal’ levels of use following habituation to the hanging rope introduced in Week 5. With that said, it cannot be definitively concluded from these results whether EE use was affected by kit age; future studies should provide enrichments with similar properties (and therefore similar appeal to kits) according to a similar schedule of enrichment exchange across all post-whelping weeks.
The standard-housed groups may serve as a better representation of potential age effects in enrichment use, as there was no renewal of object novelty or provision of enrichments with varying properties across time periods. Ring use in the standard-housed groups was significantly low in post-weaning Week 1 compared to Week 2, 3, and 4, and then began to decline across subsequent weeks. This reduced EE use in post-weaning Week 1 is interesting because the same trend was demonstrated for all EEs, regardless of housing condition. It is possible that object play was not yet fully developed in kits by this age since it was not observed until seven weeks of age in a previous study (Jonasen, 1987), though enrichments were presently used by kits to some extent at six weeks of age. Alternatively, increased stress due to weaning may have influenced kits’ use of enrichment items until post-weaning Week 2, at which point there may have been some recovery. Stress is generally known to decrease exploratory behaviour, though intervention with EE is also shown to mediate these stress effects and promote explorative behaviour, particularly in early life (Mkwanazi et al., 2018; Bak and Malmkvist, 2020). It is possible that EEs were able to mediate weaning stress to some extent but were not able to influence kit behaviour in the immediate post-weaning week. However, we collected no physiological stress measures from kits around the time of weaning, so this is speculative. Such benefits of EE in the immediate post-weaning period could also be validated by determining whether kits value access to enrichments at this time (e.g., by conducting consumer-demand studies which assess ‘costs’ paid to access enrichments; Cooper and Mason, 1997; Cooper and Mason, 2000).
In conclusion, the results of this study demonstrate that enrichments with more numerous uses, greater malleability, or greater controllability were most effective at promoting EE interaction. Although the ring enrichment, which is standard to provide across all Canadian mink farms, was one of the most-used enrichments, the presence of multiple EEs in the cage was able to significantly increase overall enrichment use compared to provision of the ring alone. There also seemed to be a positive effect of object novelty on enrichment use in weeks where familiar EEs were exchanged for novel ones, however, these increases were not statistically significant, and there appeared to be rapid habituation to these items depending on their properties. More ‘complex’ novel enrichments may remain cognitively stimulating across a longer period and therefore warrant more exploration, prolonging the positive effects of novelty on EE use. It is unclear whether kit age influenced EE use in this study; further research is required in which the enrichments supplied are equivalently appealing to kits and follow a similar schedule of enrichment exchange across all post-whelping weeks. In such a study, it may also be beneficial to provide the same number of fully fixed structural enrichments, fixed but manipulable enrichments, and mobile enrichments to provide more information about kit preference for different enrichment types. Consumer-demand studies could also be used to determine whether kits value access to enrichments during certain stages of development or housing (e.g., following weaning). Measures of physiological stress should also be collected from kits around the time of weaning to investigate the relationship between EE use and weaning stress.
Data availability statement
The authors confirm that all data underlying the findings are fully available without restriction. Our data files are available from the Borealis Dataverse Repository at https://doi.org/10.5683/SP3/D8N26F.
Ethics statement
The animal study was reviewed and approved by the Dalhousie University Faculty of Agriculture Animal Care & Use Committee and the Clinical Research Ethics Review Board of the Royal Veterinary College.
Author contributions
RM and MD-L conceived and designed the experiments. RM and GC conducted the experiments. RM and GC analysed the data. MD-L offered input on statistical methods. GC wrote the paper. RM and MD-L reviewed and edited the paper. All authors contributed to the article and approved the submitted version.
Funding
This study was supported by Mitacs through the Mitacs Accelerate Program and by the Canada Mink Breeders Association (Mitacs IT23041). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Acknowledgments
Many thanks to the farm staff at the Canadian Centre for Fur Animal Research for their vital assistance in conducting this study, and to Meridyth Hardy for her assistance with enrichments and data collection. We would also like to thank Mitacs and the Canada Mink Breeders Association for funding.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fanim.2023.1228533/full#supplementary-material
References
Abou-Ismail U. A. (2011). Are the effects of enrichment due to the presence of multiple items or a particular item in the cages of laboratory rat? Appl. Anim. Behav. Sci. 134, 72–82. doi: 10.1016/j.applanim.2011.06.007
Abou-Ismail U. A., Mendl M. T. (2016). The effects of enrichment novelty versus complexity in cages of group-housed rats (Rattus norvegicus). Appl. Anim. Behav. Sci. 180, 130–139. doi: 10.1016/j.applanim.2016.04.014
Ahloy Dallaire J., Mason G. J. (2016). Play in juvenile mink: litter effects, stability over time, and motivational heterogeneity. Dev. Psychobiol. 58, 945–957. doi: 10.1002/dev.21425
Ahloy Dallaire J., Mason G. J. (2017). Juvenile rough-and-tumble play predicts adult sexual behaviour in American mink. Anim. Behav. 123, 81–89. doi: 10.1016/j.anbehav.2016.10.023
Ahola L., Mononen J., Mohaibes M. (2011). Effects of access to extra cage constructions including a swimming opportunity on the development of stereotypic behaviour in singly housed juvenile farmed mink (Neovison vison). Appl. Anim. Behav. Sci. 134, 201–208. doi: 10.1016/j.applanim.2011.06.013
Axelsson H. M. K., Aldén E., Lidfors L. (2009). Behaviour in female mink housed in enriched standard cages during winter. Appl. Anim. Behav. Sci. 121, 222–229. doi: 10.1016/j.applanim.2009.09.013
Bak A. S., Malmkvist J. (2020). Barren housing and negative handling decrease the exploratory approach in farmed mink. Appl. Anim. Behav. Sci. 222, 104901. doi: 10.1016/j.applanim.2019.104901
Bashaw M. J., Bloomsmith M. A., Marr M. J., Maple T. L. (2003). To hunt or not to hunt? A feeding enrichment experiment with captive large felids. Zoo Biol. 22, 189–198. doi: 10.1002/zoo.10065
Bassett L., Buchanan-Smith H. M. (2007). Effects of predictability on the welfare of captive animals. Appl. Anim. Behav. Sci. 102, 223–245. doi: 10.1016/j.applanim.2006.05.029
Brink A.-L., Jeppesen L. L. (2005). Behaviour of mink kits and dams (Mustela vison) in the lactation period. Can. J. Anim. Sci. 85, 7–12. doi: 10.4141/A04-028
Buob M., Meagher R., Dawson L., Palme R., Haley D., Mason G. (2013). Providing ‘get-away bunks’ and other enrichments to primiparous adult female mink improves their reproductive productivity. Appl. Anim. Behav. Sci. 147, 194–204. doi: 10.1016/j.applanim.2013.05.004
Burger J. M., Arkin R. M. (1980). Prediction, control, and learned helplessness. J. Pers. Soc. Psychol. 38, 482–491. doi: 10.1037/0022-3514.38.3.482
Campbell D. L. M., Dallaire J. A., Mason G. J. (2013). Environmentally enriched rearing environments reduce repetitive perseveration in caged mink, but increase spontaneous alternation. Behav. Brain Res. 239, 177–187. doi: 10.1016/j.bbr.2012.11.004
Clubb R., Mason G. J. (2007). Natural behavioural biology as a risk factor in carnivore welfare: How analysing species differences could help zoos improve enclosures. Appl. Anim. Behav. Sci. 102, 303–328. doi: 10.1016/j.applanim.2006.05.033
Cooper J. J., Mason G. (1997). The effect of cost of access on consumption of environmental resources in mink. BSAP Occasional Publ. 20, 129–130. doi: 10.1017/S0263967X00043639
Cooper J. J., Mason G. J. (2000). Increasing costs of access to resources cause re-scheduling of behaviour in American mink (Mustela vison): implications for the assessment of behavioural priorities. Appl. Anim. Behav. Sci. 66, 135–151. doi: 10.1016/S0168-1591(99)00069-6
Cooper J. J., Ödberg F., Nicol C. J. (1996). Limitations on the effectiveness of environmental improvement in reducing stereotypic behaviour in bank voles (Clethrionomys glareolus). Appl. Anim. Behav. Sci. 48, 237–248. doi: 10.1016/0168-1591(95)01020-3
Courboulay V. (2014). Enrichment materials for fattening pigs: summary of IFIP trials. Cahiers l’IFIP 1, 47–56.
de Oliveira L. B. S., da Silva Vasconcellos A. (2022). May unpredictable events affect monkey welfare under human care? Behav. Processes 200, 104665. doi: 10.1016/j.beproc.2022.104665
Díez-León M., Bowman J., Bursian S., Filion H., Galicia D., Kanefsky J., et al. (2013). Environmentally enriched male mink gain more copulations than stereotypic, barren-reared competitors. PloS One 8, e80494. doi: 10.1371/journal.pone.0080494
Díez-León M., Mason G. (2016). Effects of environmental enrichment and stereotypic behavior on maternal behavior and infant viability in a model carnivore, the American mink (Neovison vison). Zoo Biol. 35, 19–28. doi: 10.1002/zoo.21249
Elmore M. R. P., Garner J. P., Johnson A. K., Kirkden R. D., Patterson-Kane E. G., Richert B. T., et al. (2012). Differing results for motivation tests and measures of resource use: The value of environmental enrichment to gestating sows housed in stalls. Appl. Anim. Behav. Sci. 141, 9–19. doi: 10.1016/j.applanim.2012.07.010
Fairhurst G. D., Frey M. D., Reichert J. F., Szelest I., Kelly D. M., Bortolotti G. R. (2011). Does environmental enrichment reduce stress? An integrated measure of corticosterone from feathers provides a novel perspective. PloS One 6, e17663. doi: 10.1371/journal.pone.0017663
Gerell R. (1967). Food selection in relation to habitat in mink (Mustela vison Schreber) in Sweden. Oikos 18, 233–246. doi: 10.2307/3565101
Hansen S., Malmkvist J., Palme R., Damgaard B. (2007). Do double cages and access to occupational materials improve the welfare of farmed mink? Anim. Welfare 16, 63–76. doi: 10.1017/S0962728600030943
Jeppesen L. L. (2004). Mink welfare improved by combined implementation of several small initiatives. Scientifur 28, 11–18.
Jeppesen L. L., Falkenberg H. (1990). Effects of play balls on peltbiting, behavior and level of stress in ranch mink. Scientifur 14, 179–186.
Jones R. B. (2001). Does occasional movement make pecking devices more attractive to domestic chicks? Br. Poultry Sci. 42, 43–50. doi: 10.1080/00071660020035064
Li K. A., Lund E. T., Voigt J.-P. W. (2016). The impact of early postnatal environmental enrichment on maternal care and offspring behaviour following weaning. Behav. Processes 122, 51–58. doi: 10.1016/j.beproc.2015.11.008
Maple T., Perdue B. (2013). “Chapter 6: environmental enrichment,” in Zoo animal welfare (Berlin, Heidelberg: Springer-Verlag), 95–117. doi: 10.1007/978-3-642-35955-2_6
Mason G. J. (1993). Age and context affect the stereotypies of caged mink. Behaviour 127, 191–229. doi: 10.1163/156853993X00029
McQuaid R. J., Audet M.-C., Anisman H. (2012). Environmental enrichment in male CD-1 mice promotes aggressive behaviors and elevated corticosterone and brain norepinephrine activity in response to a mild stressor. Stress 15, 354–360. doi: 10.3109/10253890.2011.623249
McQuaid R. J., Audet M.-C., Jacobson-Pick S., Anisman H. (2013). Environmental enrichment influences brain cytokine variations elicited by social defeat in mice. Psychoneuroendocrinology 38, 987–996. doi: 10.1016/j.psyneuen.2012.10.003
McQuaid R. J., Dunn R., Jacobson-Pick S., Anisman H., Audet M.-C. (2018). Post-weaning environmental enrichment in male CD-1 mice: Impact on social behaviors, corticosterone levels and prefrontal cytokine expression in adulthood. Front. Behav. Neurosci. 12. doi: 10.3389/fnbeh.2018.00145
Meagher R. K., Campbell D. L. M., Mason G. J. (2017). Boredom-like states in mink and their behavioural correlates: A replicate study. Appl. Anim. Behav. Sci. 197, 112–119. doi: 10.1016/j.applanim.2017.08.001
Meagher R. K., Dallaire J. A., Campbell D. L. M., Ross M., Møller S. H., Hansen S. W., et al. (2014). Benefits of a ball and chain: simple environmental enrichments improve welfare and reproductive success in farmed American mink (Neovison vison). PloS One 9, e110589. doi: 10.1371/journal.pone.0110589
Meagher R. K., Mason G. J. (2012). Environmental enrichment reduces signs of boredom in caged mink. PloS One 7, e49180. doi: 10.1371/journal.pone.0049180
Mkwanazi M. V., Ncobela C. N., Kanengoni A. T., Chimonyo M. (2018). Effects of environmental enrichment on behaviour, physiology and performance of pigs — A review. Asian-Australas J. Anim. Sci. 32, 1–13. doi: 10.5713/ajas.17.0138
Morgan K., Tromborg C. (2006). Sources of stress in captivity. Appl. Anim. Behav. Sci. 102, 262–302. doi: 10.1016/j.applanim.2006.05.032
Morano R., Hoskins O., Smith B. L., Herman J. P. (2019). Loss of environmental enrichment elicits behavioral and physiological dysregulation in female rats. Front. Behav. Neurosci. 12. doi: 10.3389/fnbeh.2018.00287
National Farm Animal Care Council (2013). Code of practice for the care and handling of farmed mink (Lacombe, Alta: National Farm Animal Care Council).
Newberry R. C. (1995). Environmental enrichment: Increasing the biological relevance of captive environments. Appl. Anim. Behav. Sci. 44, 229–243. doi: 10.1016/0168-1591(95)00616-Z
Perre V. V., Driessen B., Thielen J. V., Verbeke G., Geers R. (2011). Comparison of pig behaviour when given a sequence of enrichment objects or a chain continuously. Anim. Welfare 20, 641–649. doi: 10.1017/S0962728600003286
Pisula W., Modlinska K., Chrzanowska A., Goncikowska K. (2021). Response to novelty induced by change in size and complexity of familiar objects in Lister-Hooded rats, a follow-up of 2019 study. Sci. Rep. 11, 10281. doi: 10.1038/s41598-021-89289-y
Polanco A., Meagher R., Mason G. (2021). Boredom-like exploratory responses in farmed mink reflect states that are rapidly reduced by environmental enrichment, but unrelated to stereotypic behaviour or ‘lying awake.’. Appl. Anim. Behav. Sci. 238, 105323. doi: 10.1016/j.applanim.2021.105323
Riggio G., Mariti C., Boncompagni C., Corosaniti S., Di Giovanni M., Ogi A., et al. (2019). Feeding enrichment in a captive pack of European wolves (Canis lupus lupus): assessing the effects on welfare and on a zoo’s recreational, educational and conservational role. Animals 9, 331. doi: 10.3390/ani9060331
Roy C., Lippens L., Kyeiwaa V., Seddon Y. M., Connor L. M., Brown J. A. (2019). Effects of enrichment type, presentation and social status on enrichment use and behaviour of sows with electronic sow feeding. Anim. (Basel) 9, 369. doi: 10.3390/ani9060369
Ruskell A. D., Meiers S. T., Jenkins S. E., Santymire R. M. (2015). Effect of Bungee-carcass enrichment on behavior and fecal glucocorticoid metabolites in two species of zoo-housed Felids. Zoo Biol. 34, 170–177. doi: 10.1002/zoo.21192
Sambrook T. D., Buchanan-Smith H. M. (1997). Control and complexity in novel object enrichment. Anim. Welfare 6, 207–216. doi: 10.1017/S0962728600019801
Smith B. L., Lyons C. E., Correa F. G., Benoit S. C., Myers B., Solomon M. B., et al. (2017). Behavioral and physiological consequences of enrichment loss in rats. Psychoneuroendocrinology 77, 37–46. doi: 10.1016/j.psyneuen.2016.11.040
Tilly S.-L. C., Dallaire J., Mason G. J. (2010). Middle-aged mice with enrichment-resistant stereotypic behaviour show reduced motivation for enrichment. Anim. Behav. 80, 363–373. doi: 10.1016/j.anbehav.2010.06.008
Van de Weerd H. A., Docking C. M., Day J. E. L., Avery P. J., Edwards S. A. (2003). A systematic approach towards developing environmental enrichment for pigs. Appl. Anim. Behav. Sci. 84, 101–118. doi: 10.1016/S0168-1591(03)00150-3
Van Loo P. L. P., Kruitwagen C. L. J. J., Koolhaas J. M., Van de Weerd H. A., Van Zutphen L. F. M., Baumans V. (2002). Influence of cage enrichment on aggressive behaviour and physiological parameters in male mice. Appl. Anim. Behav. Sci. 76, 65–81. doi: 10.1016/S0168-1591(01)00200-3
Videan E., Fritz J., Schwandt M., Smith H., Howell S. (2005). Controllability in environmental enrichment for captive chimpanzees (pan troglodytes). J. Appl. Anim. Welfare Sci. 8, 117–130. doi: 10.1207/s15327604jaws0802_4
Vinke C., van Leeuwen J. (2005). Juvenile farmed mink (Mustela vison) with additional access to swimming water play more frequently than animals housed with a cylinder and platform, but without swimming water. Anim. Welfare 14, 53–60. doi: 10.1017/S096272860002892X
Watters J. V., Miller J. T., Sullivan T. J. (2011). Note on optimizing environmental enrichment: a study of fennec fox and zoo guests. Zoo Biol. 30, 647–654. doi: 10.1002/zoo.20365
Weber E., Ahloy Dallaire J., Gaskill B., Pritchett-Corning K., Garner J. (2017). Aggression in group-housed laboratory mice: Why can’t we solve the problem? Lab. Anim. 46, 157–161. doi: 10.1038/laban.1219
Keywords: American mink, animal welfare, environmental enrichment, enrichment use, novelty
Citation: Clark GB, Díez-León M and Meagher RK (2023) Diversity and novelty in environmental enrichment increases enrichment use in juvenile American mink (Neogale vison). Front. Anim. Sci. 4:1228533. doi: 10.3389/fanim.2023.1228533
Received: 24 May 2023; Accepted: 18 July 2023;
Published: 09 August 2023.
Edited by:
Rafael Freire, Charles Sturt University, AustraliaReviewed by:
Claudia Vinke, Utrecht University, NetherlandsSarah Talbot, Charles Sturt University, Australia
Copyright © 2023 Clark, Díez-León and Meagher. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Gabrielle B. Clark, Z2FicmllbGxlYmNsYXJrQG91dGxvb2suY29t