 Maria Gallardo Paffetti
Maria Gallardo Paffetti Lucía Azócar-Aedo
Lucía Azócar-Aedo Angel Parra-Vizcaíno
Angel Parra-Vizcaíno José M. Larraín4
José M. Larraín4- 1Escuela de Medicina Veterinaria, Facultad de Medicina y Ciencias de la Salud, Universidad Mayor, Santiago, Chile
- 2Escuela de Medicina Veterinaria, Facultad de Ciencias de la Naturaleza, Universidad San Sebastián, Puerto Montt, Chile
- 3Facultad de Ciencias del Mar, Universidad Católica del Norte, Coquimbo, Chile
- 4Escuela de Agronomía, Facultad de Ciencias, Universidad Mayor, Santiago, Chile
The large intestine healthy microbiota in sheep hosts pathogenic, but mainly nonpathogenic bacteria, which are essential to intestinal metabolism, contributing energy, antigens, and metabolites that positively impact host physiology, immunity, and metabolism. However, this microbiota also poses a public health risk due to fecal contamination in animal products, such as wool. This study examined how maternal diet and pregnancy type influence the relative abundance of zoonotic bacterial DNA belonging to phyla Firmicutes and Proteobacteria in sheep feces and wool. In total, 18 Ile de France ewes, with 8 carrying twins and 10 single lambs, were divided into two groups: one fed ad libitum on naturalized pasture, the other given red clover hay plus lupine, from 45 days prepartum to 60 days postpartum. Both fecal and wool samples were collected from ewes and lambs four and three times, respectively, and analyzed via qPCR for Firmicutes (Clostridium perfringens type C, Staphylococcus aureus, Staphylococcus epidermidis, Streptococcus uberis, Enterococcus faecalis, and Streptococcus agalactiae) and Proteobacteria (Salmonella typhimurium and Escherichia coli serotype O157). Data were analyzed using repeated measures two-way ANOVA. Results showed lower bacterial abundance in fecal samples than in wool samples, with ewe’s wool exhibiting a lower bacterial abundance compared with lamb’s wool. E. faecalis (Firmicutes) and E. coli (Proteobacteria) were the most prevalent bacteria, suggesting environmental contamination related to sheep behavior. In summary, handling offspring from birth to weaning and ewes until 60 days postpartum may increase zoonotic pathogen transmission risk, raising public health concerns regarding exposure to intestinal pathogenic bacteria.
1 Introduction
The large intestine microbiota plays a crucial role in sheep intestinal metabolism, although it remains inadequately described. It likely provides energy, antigens, and metabolites, positively impacting host physiology, immunity, and metabolism. The interplay between the microbiota and mucosal physiology maintains a stable, which is essential for animal health and productivity (Khafipour et al., 2009; Tanca et al., 2017).
The microbiota in the large intestine is similar to that in the rumen (Cholewińska et al., 2020a), aiding decomposition of plant material and its conversion into energy in the form of volatile fatty acids, thereby directly affecting health, development, and productivity (Khafipour et al., 2009; Tanca et al., 2017; Wang et al., 2019; Zeng et al., 2017).
In sheep, the microbial community of large intestine consists primarily of Firmicutes, Bacteroidetes, Actinobacteria, and Proteobacteria (Cholewińska et al., 2023). Factors influencing this microbiota include environment, breed (Chang et al., 2020; Liggenstoffer et al., 2010), age (Parmar et al., 2014), feed efficiency (McLoughlin et al., 2020), diet (Wang et al., 2019), and physiological stage (Cholewińska et al., 2023), with a convergence toward mature bacterial structure as animals transition from birth to adulthood (Jami et al., 2013). Physiological stages, including conception, early pregnancy, parturition, and end of lactation in sheep, impact the digestive system’s microbiology (Szeligowska et al., 2022). Stress and dysbiosis also shape the microbial community of ruminants, which itself acts as a specific immune system preventing pathogen growth (Szeligowska et al., 2022).
Dysbiosis not only compromises animal health but also poses significant public health risks due to zoonotic spillover, particularly in small ruminants like sheep, which are reservoirs for various gastrointestinal pathogens (Hancock et al., 2001; Brown et al., 2004; Cox et al., 2005) and can transmit bacterial pathogens to humans (Battisti et al., 2006; Blanco et al., 2003; Hanlon et al., 2018). This risk is especially high for young individuals, which show higher susceptibility compared to adults (Delgado et al., 2013), posing a public health concern associated with the intake and handling of animal products contaminated with pathogenic bacteria, including wool (Mersha et al., 2010).
In a previous pilot study, it was determined that bacteria with zoonotic potential are detected in sheep feces and wool (Gallardo et al., 2019) and according Szeligowska et al. (2022), stress and dysbiosis influence the microbial community of ruminants. Considering that pregnancy, parturition and delivery are stressful factors for mothers (Nagel et al., 2019), we hypothesized that maternal diet and pregnancy type influences the bacterial phyla present in sheep feces and wool. Specifically, this study aimed to determine the effects of maternal diet and pregnancy type on the relative abundance of zoonotic bacterial DNA belonging to phyla Firmicutes and Proteobacteria in feces and wool.
2 Materials and methods
2.1 Bioethics
The methodology employed in this study was approved by the Committee for the Ethical Use of Animals in Experiments of the Universidad Austral de Chile (N°241/2015).
2.2 Location
The experiment was conducted on a farm located 12 km southeast of Villarrica city, IX Region of the Araucanía, southern Chile (39°16′0″ S, 72°13′0″ E), from July 2016 to January 2017.
2.3 Experimental design
In total, 18 animals were randomly selected from a larger flock of free-ranging Ile de France ewes in their third birth cycle, all with a similar body condition score (3.0). This group comprised eight twin-bearing and ten single lamb–bearing ewes grazing on naturalized pasture, a successional pasture that germinates spontaneously under suitable temperature and humidity conditions. Dominant species in this pasture included Agrostis tenuis, Holcus lanatus, and Trifolium repens (Gallardo et al., 2011). The selected ewes were divided into two dietary groups: one fed ad libitum on naturalized pasture (85.20% ± 1.93% dry matter, 8.80% ± 0.45% crude protein, 2.13 ± 0.07 Mcal kg−1 metabolizable energy, 64.77% ± 2.39% neutral detergent fiber, and 5.26% ± 0.14% total ashes), and the other fed red clover hay (83.12% ± 1.62% dry matter, 10.62% ± 0.12% crude protein, 2.31 ± 0.04 Mcal kg−1 metabolizable energy, 54.35% ± 0.13% neutral detergent fiber, and 5.23% ± 0.08% total ashes) supplemented with lupine (88.90% dry matter, 17.17% crude protein, 3.08 Mcal kg−1 metabolizable energy, 52.72% neutral detergent fiber, and 3.37% total ashes) to meet nutritional requirements (Gallardo et al., 2019). Ewes were divided into four groups based on diet and pregnancy type: twin-bearing ewes fed naturalized pasture (n = 4) or red clover hay (n = 4), and single lamb–bearing ewes fed naturalized pasture (n = 5) or red clover hay (n = 5). The treatment period spanned from 45 days prepartum to 60 days postpartum.
2.3.1 Sampling
Fecal and wool samples were collected from ewes 10 days prepartum (time 0) and from ewes and lambs at birth (time 1), 30 days postpartum (time 2), and 60 days postpartum (time 3) to analyze the relative abundance of bacterial DNA. Samples were placed in Eppendorf tubes, transported on ice to the Institute of Biochemistry and Microbiology, Universidad Austral de Chile, and stored at −80°C until further analysis. RNA Safer Stabilizer Reagent (E.Z.N.A.) was used to preserve samples at −80°C.
Approximately 0.5 g of feces and 8 wool fibers per animal were collected and processed using a DNA extraction protocol with 300 µL of Chelex-100 5%, adding 2 µL of lysozyme (10 mg/mL) for feces and 2 µL of proteinase K for wool. Extracted DNA was stored at −80°C until analysis. Pre-existing stock cultures for each of the eight bacteria studied were provided by the Institute of Biochemistry and Microbiology and were utilized as positive controls. DNA was extracted from the bacterial cultures using E.Z.N.A and the OMEGA-BIOTEK Tissue DNA Kit D3396-02.
2.4 Genomic DNA
Complete sequences of the 16S rRNA gene (16S rDNA) were obtained from Salmonella typhimurium (ATCC14028), Escherichia coli O157 (ATCC25922), Clostridium perfringens (ATCC13124), Staphylococcus aureus (ATCC6538), Staphylococcus epidermidis (ATCC12228), Streptococcus uberis (ATCC9927), Enterococcus faecalis (ATCC19433), and Streptococcus agalactiae (ATCC27956). Primer designs are provided in Table 1.
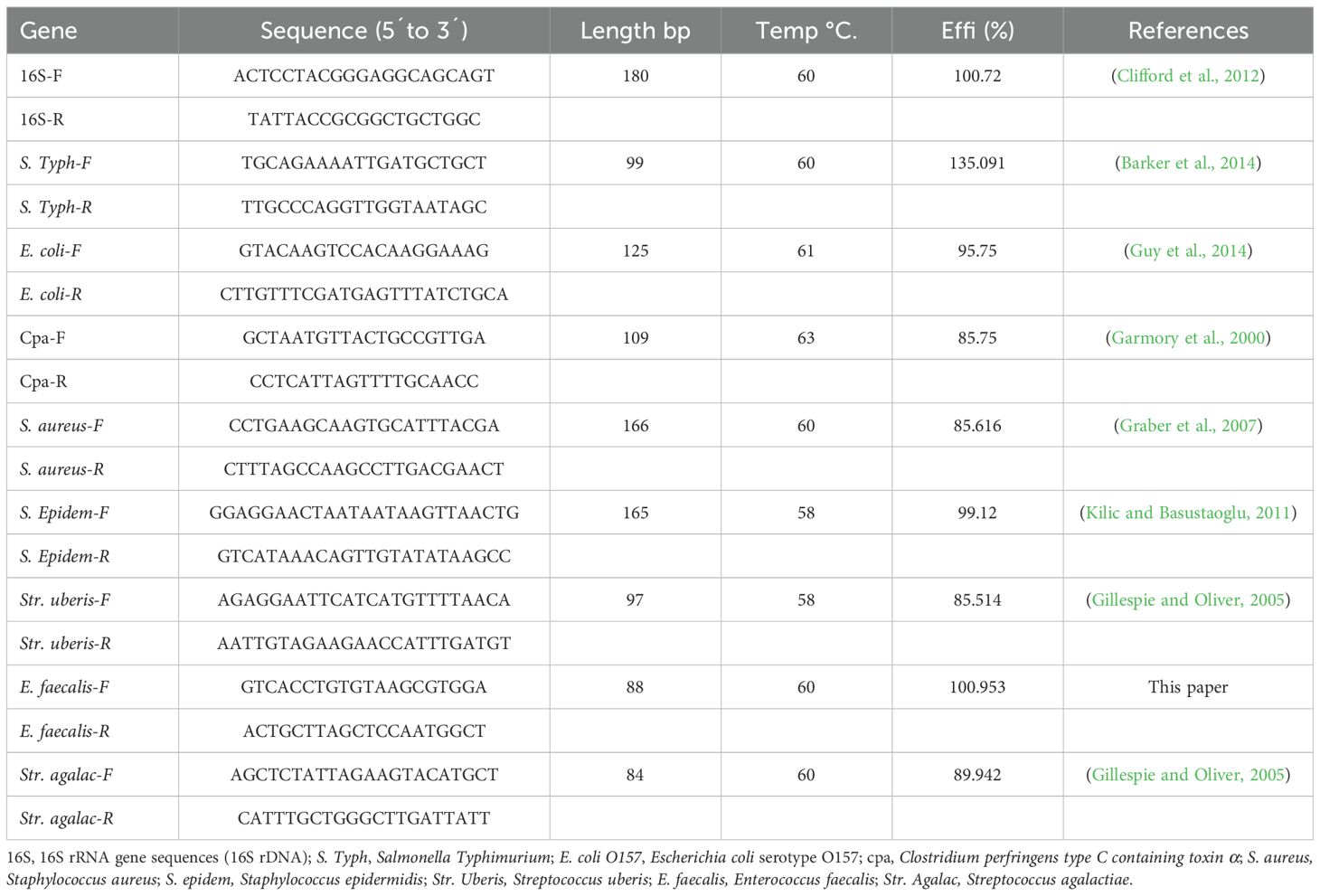
Table 1. Primer design specification for each bacteria considered in the study.
Ensured accurate thermal cycling conditions and reaction component specification, primers (Table 1) were designed using AmplifX 1.5 software, and reactions were performed on a StepOnePlus Real-Time PCR System, using 5.0 µL of Green Master Mix Promega, 1.0 µL of sample as the template, and 0.5 µL of each primer (10 µM), following a standard protocol: initial denaturation at 95°C for 10 min, followed by 40 cycles at 60°C. Cycle threshold (Ct) values were used to calculate DNA concentrations from a calibration curve prepared with positive control DNA. Initial DNA concentrations were measured spectrophotometrically at 260 nm. Bacterial load values were derived from qPCR results.
2.5 Statistical analysis
The effects of maternal diet (naturalized or red clover pasture) and pregnancy type (twins or a single lamb) on the relative abundance of eight bacterial types, in fecal and wool samples, collected at four time points (0–3) from ewes 10 days prepartum (time 0), and from ewes and lambs at birth (time 1), 30 days postpartum (time 2), and 60 days postpartum (time 3), were analyzed using a linear model. Fixed factors included maternal diet, pregnancy type, and their interaction. Treatment and measurement time effects are shown in each graph. Data were analyzed through repeated measures two-way ANOVA in R studio (version 4.0.3), with Bonferroni correction applied to identify significant mean differences. Results were considered statistically significant at p ≤ 0.05.
3 Results
Figure 1 shows the fecal pathogenic bacteria found in the studied sheep feces. Time and bacterial type exhibited significant (p < 0.0001), whereas maternal diet (p = 0.58) and pregnancy type (p = 0.51) were not significant. However, the interaction effect of time, bacterial type, maternal diet, and pregnancy type was significant (p < 0.0001). The Bonferroni test showed that time 3 (60 days postpartum) differed from times 0 (10 days prepartum), 1 (birth), and 2 (60 days postpartum) (p = 0.039). Irrespective of treatment, higher proportions of E. faecalis were observed compared to other bacteria (p < 0.001), especially 10 days prepartum (at time 0) in twin-bearing ewes fed naturalized pasture.
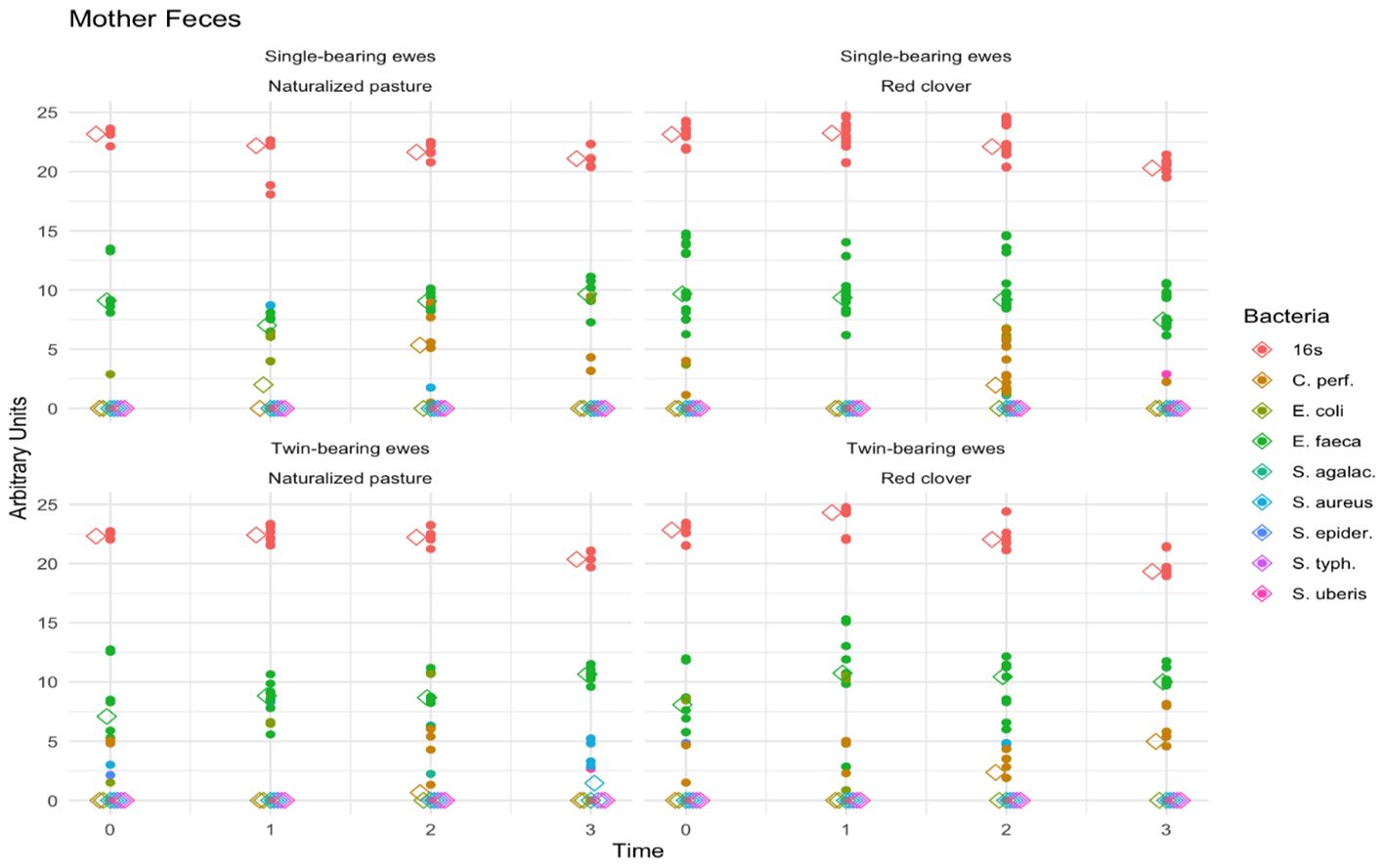
Figure 1. Fecal pathogenic bacteria in ewe feces. The scatter lines (vertical points pointing upwards) represent the abundance of the different fecal pathogenic bacteria detected in mother feces regarding to the type of pregnancy, diet and time. The colors indicate the genus and species of each microorganism identified (indicated in the legend on the right side of the figure).
Figure 2 shows the fecal pathogenic bacteria found in ewe wool. Time (p < 0.0001), bacteria type (p < 0.0001), and pregnancy type (p < 0.0001) showed significant effects, whereas maternal diet was nonsignificant (p = 0.47). Notably, the interaction effect of time, bacteria type, maternal diet, and pregnancy type was significant (p < 0.001). A greater proportion of E. coli (p < 0.001), followed by E. faecalis (p = 0.019), was found compared to other bacteria. At time 3 (60 days postpartum), E. coli levels were higher in single lamb–bearing ewes fed naturalized pasture, whereas 30 days postpartum (at time 2), E. faecalis levels were higher in twin-bearing ewes fed naturalized pasture. At time 3 (60 days postpartum), E. coli and E. faecalis levels were higher in single lamb–bearing and twin-bearing ewes fed red clover. The Bonferroni test showed no significant time differences (p = 0.43).
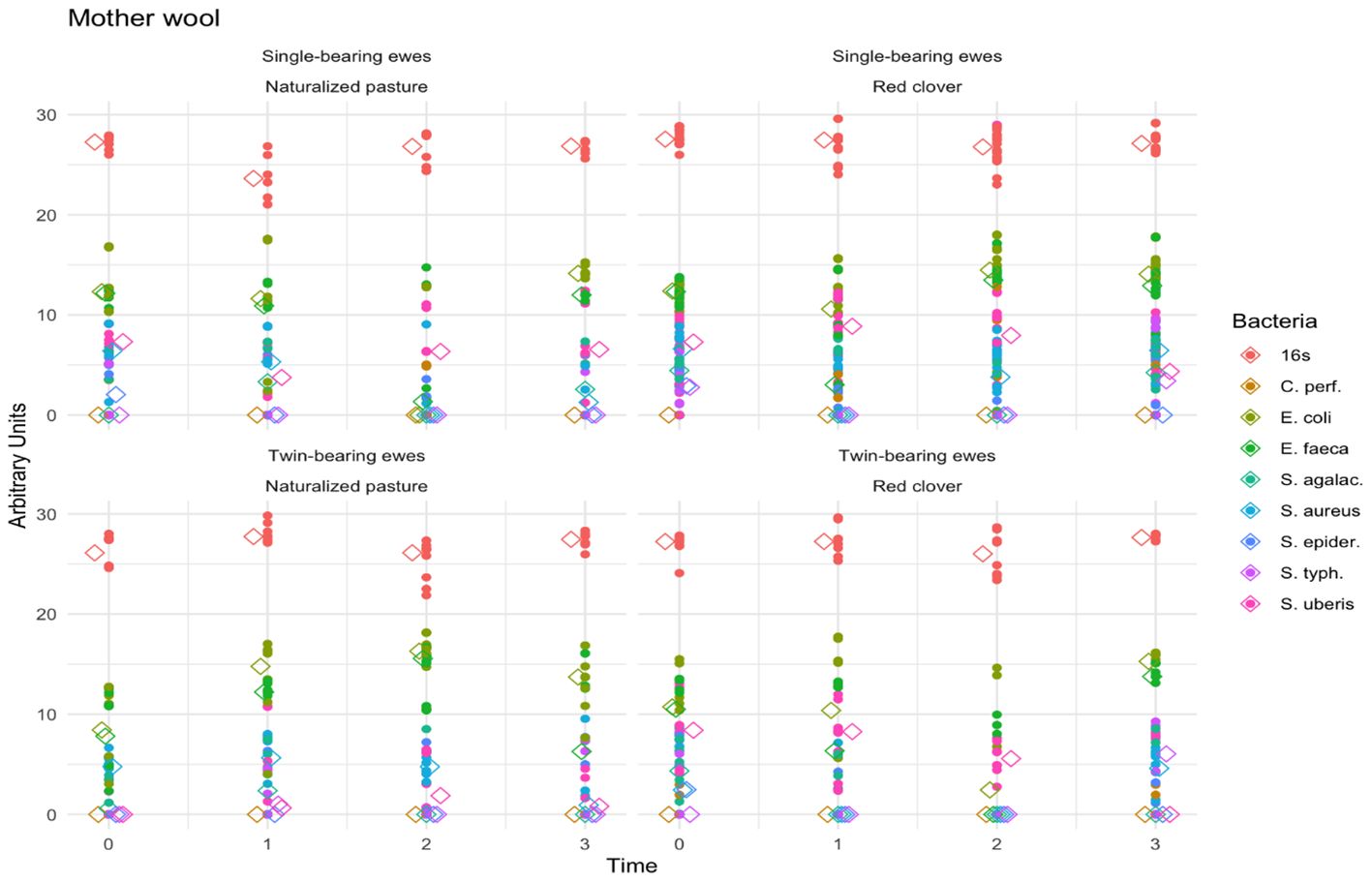
Figure 2. Fecal pathogenic bacteria in the wool of ewes. The scatter lines (vertical points pointing upwards) represent the abundance of the different fecal pathogenic bacteria detected in mother wool regarding to the type of pregnancy, diet and time. The colors indicate the genus and species of each microorganism identified (indicated in the legend on the right side of the figure).
Figure 3 shows the fecal pathogenic bacteria observed in lamb wool samples. Time (p < 0.0001), bacteria type (p < 0.0001), and maternal diet (p = 0.008) exhibited significant effects, whereas pregnancy type was nonsignificant (p = 0.58). Additionally, the interaction effect of time, bacteria type, maternal diet, and pregnancy type was nonsignificant (p = 0.59). The Bonferroni test indicated that time 3 (60 days postpartum) differed significantly from times 0, 1, and 2 (p < 0.0001). Across treatments, the offspring showed stable relative abundance of bacterial DNA from birth (time 1) to 30 days postpartum (time 2), with a decrease at 60 days postpartum (time 3) (p = 0.04). Higher proportions of E. coli and E. faecalis were observed compared to other bacteria (p < 0.001).
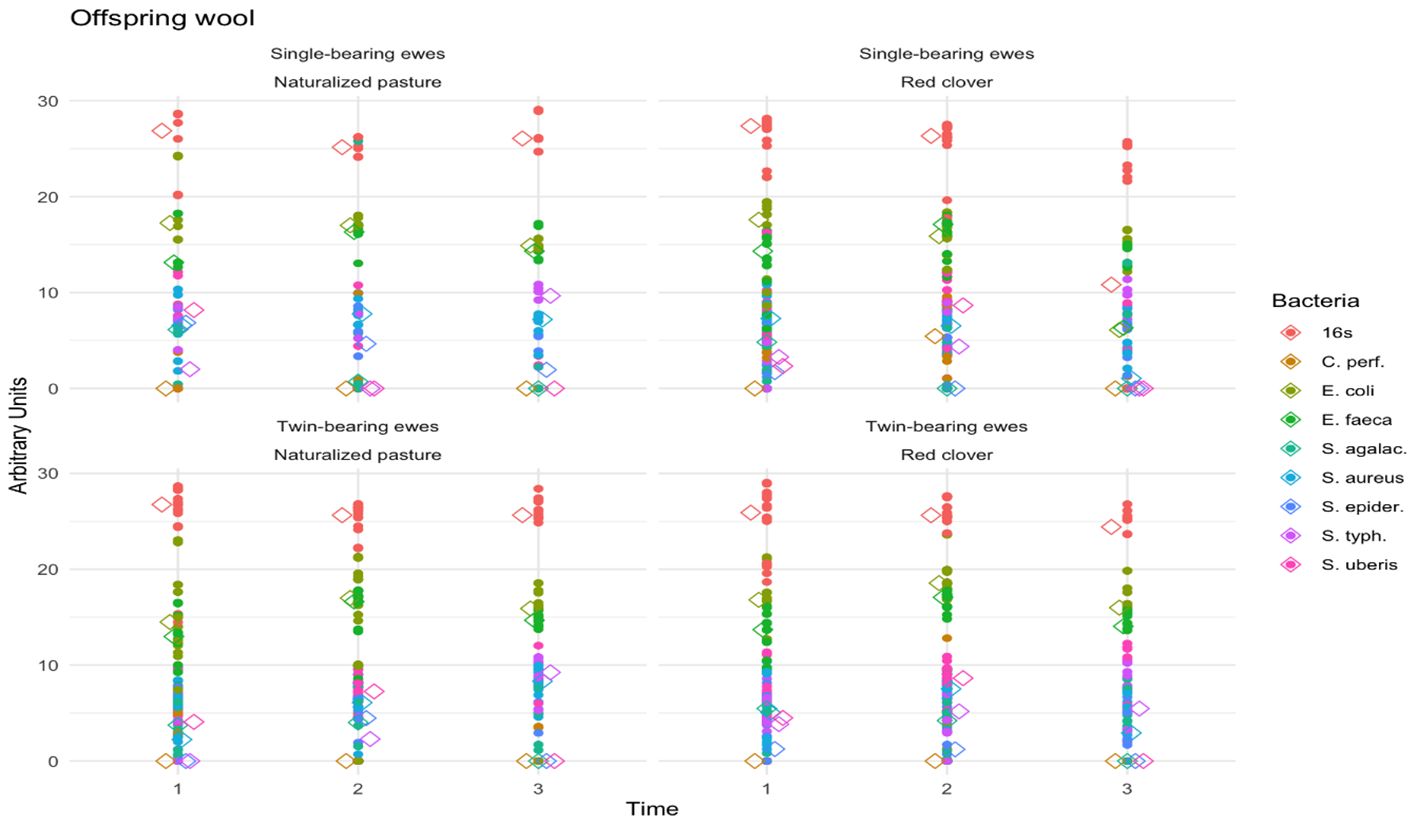
Figure 3. Fecal pathogenic bacteria in lambs’ wool. The scatter lines (vertical points pointing upwards) represent the abundance of the different fecal pathogenic bacteria detected in the offspring wool, regarding to the type of pregnancy, diet and time. The colors indicate the genus and species of each microorganism identified (indicated in the legend on the right side of the figure).
Regardless of the maternal diet, E. faecalis was found at higher levels in single lamb–bearing ewes 10 days prepartum (at time 0) compared with the other treatments. Additionally, irrespective of time, E. faecalis levels were higher in twin-bearing ewes fed red clover. However, at time 3 (60 days postpartum), E. coli abundance was higher in twin-bearing ewes fed naturalized pasture.
Figure 4 shows fecal pathogenic bacteria in sheep feces and wool samples. Across treatments, ewes exhibited higher bacterial abundance in wool than in fecal samples (p < 0.001). Bacteria type (p < 0.001) and pregnancy type (p < 0.001) showed statistically significant effects, whereas time (p = 0.36) and maternal diet (p = 0.96) were nonsignificant. Additionally, the interaction effect of time, bacteria type, and maternal diet, and pregnancy type was significant (p = 0.003). The Bonferroni test revealed no significant time differences (p = 1.00).
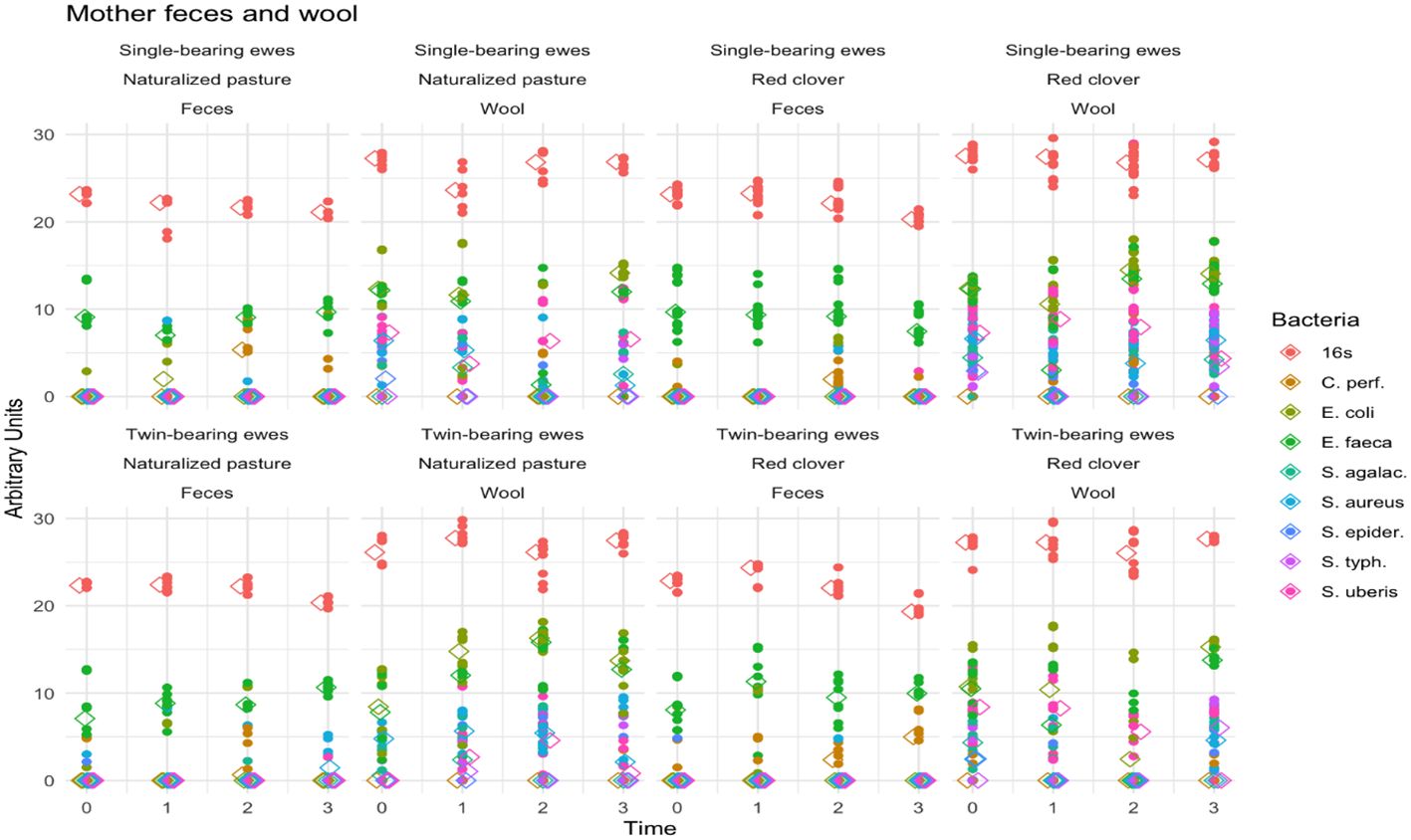
Figure 4. Pathogenic bacteria in the feces and wool of ewes. The scatter lines (vertical points pointing upwards) represent the abundance of the different fecal pathogenic bacteria detected in mother feces and wool regarding to the type of pregnancy, diet and time. The colors indicate the genus and species of each microorganism identified (indicated in the legend on the right side of the figure).
Figure 5 presents fecal pathogenic bacteria in wool from ewes and their lambs. The lambs showed higher abundance of bacterial DNA in wool compared with the ewes (p < 0.001). Time (p < 0.0001) and bacteria type (p < 0.0001) exhibited significant effects, whereas maternal diet (p = 0.13) and pregnancy type (p = 0.31) were nonsignificant. Similarly, the interaction effect of time, bacteria type, maternal diet, and pregnancy type was nonsignificant (p = 0.4). The Bonferroni test showed that time 3 (60 days postpartum) differed from times 0, 1, and 2 (p < 0.05). Higher proportions of E. coli and E. faecalis (p = 0.00036) were observed compared with all other bacteria (p < 0.001).
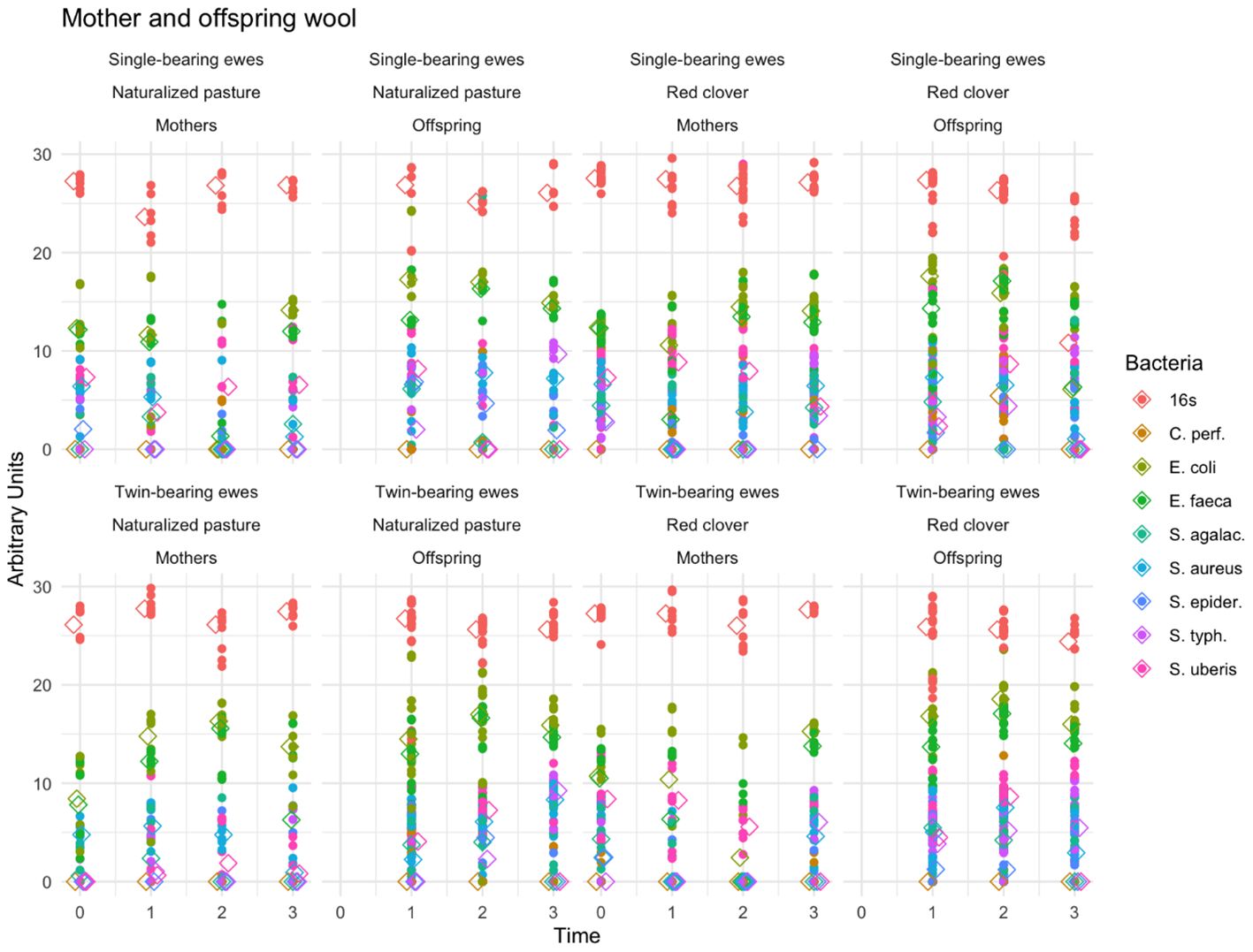
Figure 5. Fecal pathogenic bacteria in the wool of ewes and lambs. The scatter lines (vertical points pointing upwards) represent the abundance of the different fecal pathogenic bacteria detected in mother and offspring wool regarding to the type of pregnancy, diet and time. The colors indicate the genus and species of each microorganism identified (indicated in the legend on the right side of the figure).
4 Discussion
In the present study, the effects of maternal diet and pregnancy type on the relative abundance of the bacterial phyla Firmicutes and Proteobacteria in feces and wool were determined through a clinical trial. The presence of zoonotic bacteria, with more prevalence of E. faecalis and E. coli was established in both feces and wool across the evaluated groups, emphasizing its relevance to animal and public health.
It is known that bacterial levels can vary with diet (Choleińska et al., 2021). In the current study, although diet was not a significant factor, E. faecalis (Firmicutes) was more prevalent in fecal samples from ewes. Forage-based diets tend to increase microbial diversity in sheep, favoring Firmicutes, whereas concentrate-based diets tend to raise levels of Bacteroidetes and Actinobacteria (Marie-Etancelin et al., 2021). Cui et al. (2023) examined the adaptive flexibility of Tibetan sheep rumen microbiomes to diets of native pasture and oat hay finding higher microbial diversity in sheep fed natural pasture, although oat hay seemed to enhance beneficial bacteria abundance, potentially supporting host health and metabolic resilience in cold environments. In the present study, because sheep grazed with lupine grain supplementation (according to their requirements), bacterial growth in the large intestine remained consistent.
Across all treatments, ewe wool samples exhibited significantly higher relative abundance of bacterial DNA and greater bacterial diversity compared with fecal samples. In feces, E. faecalis was significantly more prevalent compared with other bacteria, especially in twin-bearing ewes fed naturalized pasture at 10 days prepartum. In contrast, wool samples had higher E. coli and E. faecalis levels, with greater relative abundance for these two species at 60 days postpartum in ewes fed red clover. This increase in E. coli DNA load by day 60 postpartum likely stems from environmental exposure associated with sheep behavior. A survey by Gallardo et al. (2019) in a free-grazing flock feeding on naturalized pasture in southern Chile revealed higher abundance of E. coli O157 and Salmonella typhimurium in lamb and kid feces compared with wool. Their study also showed that bacterial species influenced 16S abundance in feces and wool, and lamb and kid sex affected E. coli detection in feces, demonstrating the zoonotic potential of these bacteria in grazing animals.
Feed efficiency may also influence microbial community dynamics. Freetly and Lindholm-Perry (2023) noted that bacterial community structure at the clade level may vary with feed efficiency. McLoughlin et al. (2020) found that sheep feed efficiency was likely influenced by changes in specific archaeal populations and certain bacterial species, rather than general rumen microbiome changes. This could explain why, despite the nutritional differences between forage-based diets (naturalized pasture and red clover), treatment groups showed no significant differences in the current study.
Stress can alter digestive microbiota composition, as suggested by Szeligowska et al. (2022), who found that cortisol and glutathione-S-transferase levels in sheep may signal stress, potentially leading to shifts in digestive microbiota composition (Ezemonye and Ikpesu, 2011; Gate et al., 2004). Prolonged stress may increase gut colonization by pathogenic bacteria. Bailey et al. (2011) observed that stress reduced the number of intestinal bacteria from the Lactobacillaceae family in mice, highlighting the sensitivity of gut microbiota to physiological stress.
Physiological stage is another factor influencing the large intestine microbial community (Cholewińska et al., 2023). It is well known that different physiological stages affect the digestive microbiota. For sheep, these stages are generally divided into conception, early pregnancy, parturition, and end of lactation (Szeligowska et al., 2022). In ruminants, maintaining annual pregnancy cycles is crucial for meat and milk production, resulting in major microbial changes, likely affected by fluctuations in sex hormones (Menon et al., 2013). Cholewińska et al. (2023) reported that early pregnancy and lambing are the most microbiologically diverse periods, with notable variation in bacterial clusters and families. Koren et al. (2012) observed that pregnant female mice experienced microbial shifts, including reduced insulin sensitivity, likely in preparation for energy storage for raising offspring, and reported a decrease in Firmicutes and Actinobacteria proportions between the first and third trimesters. However, in the present study, the ewes were in late pregnancy, and wool samples from ewes and lambs showed higher relative abundance of E. coli and E. faecalis DNA load compared with other bacteria. This suggests that environmental contamination, rather than direct intestinal transmission, may explain the elevated E. coli levels, as the females’ fecal samples did not show E. coli dominance. Therefore, the maintenance and hygiene of the environment where the animals are housed are relevant to reduce the pathogenic bacterial load to which they are exposed.
As metabolic demands change in late pregnancy, nutritional imbalances can occur despite constant feed access, possibly due to increased uterine volume leading to reduced food intake and partial malnutrition. During these periods, quantitative and qualitative changes in the microbial community may develop (Szeligowska et al., 2022; Xue et al., 2020). Markle et al. (2013) found that manipulating the female microbiome by increasing testosterone levels raised Firmicutes and Bacteroidetes abundance compared with the control group, with Firmicutes increasing notably at the Clostridia genus level.
Stress during pregnancy can also impact fetal development and microbiome composition negatively (Castellazzi et al., 2018; Mackos et al., 2016). In ewes, parturition itself is a known stressor (Szeligowska et al., 2022). Given that the microbial community serves as an immune buffer, protecting against pathogenic bacterial growth (notably Clostridiaceae), stress could disrupt this balance, causing dysbiosis (Salcedo et al., 2016; Szeligowska et al., 2022). Tanca et al. (2017) noted that Clostridiaceae was among the most abundant families in Firmicutes in sheep, consistent with studies in cattle (Durso et al., 2010, 2011; Kim et al., 2014; Shanks et al., 2011). In the present study, parturition did not lead to increased levels of C. perfringens type C (Firmicutes), which may be attributed to concentrate supplementation provided during late pregnancy.
Cortisol levels in sheep are also affected by factors such as lactation, abortion, inadequate nutrition, miscarriage, and pregnancy toxemia (Davis et al., 2011; Edelmann et al., 2016; Ramin et al., 2007). Szeligowska et al. (2022) found that cortisol levels were higher in primiparous ewes than in multiparous ewes, suggesting that experienced mothers have lower stress hormone levels, which may contribute to better microbiome stability. Multiparous ewes also exhibited higher levels of certain bacterial groups, excluding Proteobacteria, compared with primiparous ewes. Given that all the ewes in the present study were multiparous, their experience with previous births may explain the stability in their microbiome composition.
Sheep breed can also influence dysbiosis levels. Selective breeding has developed local breeds and lines with greater resistance to environmental conditions (Cheng et al., 2022; Cholewińska et al., 2020a, 2021). There is evidence that microbial composition in the gastrointestinal tract varies between breeds of sheep and cattle (Douglas et al., 2016; Xin et al., 2019), likely due to differences in diet and management practices, which in turn affect the animal’s adaptation and its microbial community’s alignment with environmental conditions (Deng et al., 2017; Fonty et al., 1987; Malmuthuge and Guan, 2017; Rey et al., 2014). For example, Cholewińska et al. (2020b) reported a significant effect of breed on microbial community composition; however, this was not a factor in the present study, which included only one sheep breed.
Regarding sheep production, Yang et al. (2024a) developed a fecal scoring standard for fattening Hu sheep, finding that optimal production outcomes were associated with fecal scores of grade 3 and 4, and that these scores were closely linked to microbial composition, growth performance, and immunity. Future studies should consider measuring relative abundance of fecal bacterial DNA beyond the weaning stage into the lamb-fattening stage. In the current study, the lamb’s wool showed elevated E. coli and E. faecalis abundance from birth to 30 days postpartum, which declined by day 60 postpartum (weaning age), whereas ewe’s wool showed increased E. coli and E. faecalis DNA content at 60 days postpartum, especially among ewes fed red clover. This suggests that E. coli contamination in ewe’s wool arises mainly from environmental sources around weaning, possibly influenced by the maternal red clover diet.
Small ruminants are known reservoirs for various gastrointestinal pathogens (Brown et al., 2004; Cox et al., 2005; Hancock et al., 2001) and can transmit bacteria to humans (Battisti et al., 2006; Blanco et al., 2003; Hanlon et al., 2018), with children being especially susceptible (Delgado et al., 2013). In the present study, lamb’s wool showed higher E. coli and E. faecalis levels compared to ewe’s wool, likely due to typical lamb behavior and age-related susceptibility. Previous research has shown that E. coli O157:H7 is present in Namibian sheep feces and wool (Madzingira, 2016), and multidrug-resistant strains of E. faecalis have been isolated from sheep dairy products in Poland (Gołás-Pradzynska et al., 2022). Kudva et al. (1997) demonstrated that sheep transiently shed a variety of strains, including E. coli O157:H7 with risk for transmission to humans. The acquisition of resistance genes through horizontal gene transfer, facilitated by mobile genetic elements such as plasmids and transposons is described for this pathogen (Nasrollahian et al., 2024). On the other hand, Shiga toxin-producing E. coli (STEC) represent a major issue because of the capability to cause large outbreaks and the severity of the associated illnesses; their epidemiology is related with human medicine, veterinary medicine, food safety, water, and environmental microbiology, requiring a “One Health” approach for research (Caprioli et al., 2014). Further studies are needed to determine the strains related to E. coli prevalent in small ruminants.
E. fecalis resides in the gastrointestinal tract of most animals, including humans and generally is non-pathogenic in healthy hosts, however, in the pathogenic form, it can induce life-threatening opportunistic infections whose treatments are complicated by a high degree of antimicrobial resistance (Willett and Dunny, 2024). Globally, the prevalence of drug resistant E. faecalis strains increase over time (Guan et al., 2024). The bacteria can cause complicated infections, especially in immunocompromised people by underlying conditions and their treatment or age; it is also mentioned as a nosocomial pathogen is related with the consumption of contaminated dairy products, being a public health concern (Daca and Jarzembowski, 2024).
From a public health perspective, the present findings are important and call for increased awareness of zoonotic disease risks associated with handling and consuming animal products contaminated by pathogenic bacteria, including wool (Mersha et al., 2010). Our study suggests that the risk of contamination from lamb’s wool, particularly with E. coli and E. faecalis, is highest from birth until weaning (day 60 postpartum), whereas the contamination risk from ewe’s wool increases around day 60 postpartum. Notably, E. coli O157:H7 can cause severe illness, including diarrhea, hemorrhagic colitis, and hemolytic uremic syndrome, with a low infectious dose of just 10–100 cells (Schmid-Hempel et al., 2007; Wang et al., 2024). Survivors may face chronic renal complications (Beauvais et al., 2018). Hemolytic uremic syndrome is associated with high mortality and multisystem morbidity, highlighting the need for renal monitoring and attention to extra-renal effects (Rahal et al., 2012). Additionally, E. faecalis can lead to various infections, including endocarditis, urinary tract infections, and meningitis (Yang et al., 2024b). Moreover, E. faecalis can form biofilms in endodontic infections, which have physicochemical properties that vary in response to environmental and nutritional conditions, impacting antimicrobial treatment options (Jhajharia et al., 2015).
The results of this study emphasize the relevance of incorporating biosecurity measures in animal management in case of direct contact with wool, also considering that other zoonotic agents could potentially be transmitted. For example, sheep shearing has been described to play a significant role in inducing periodic outbreaks of human brucellosis in countries such as Mongolia (Ma et al., 2022) and the detection of Coxiella burnetii in the air of an enclosed sheep barn during shearing in Germany has been reported (Schulz et al., 2005). Future research could consider the detection of other pathogens of public health concern. Some biosecurity measures that must be considering in shearing are the following: i) properly clean, and disinfect all equipment and clothing when working between different flocks, ii) using a clean, newly-sharpened comb and cutter when start shearing a new flock, iii) know the health status of the animals to be shearing to ensure that biosecurity practices are adequate, iv) properly clean clothes, gloves, hands and shoes used while shearing, v) dispose properly any excess wool before starting the next shearing, and, vii) maintain records to identify the potential origin or spread of a disease in flocks sheared (Smith, 2018).
One of the limitations of the study is that it only includes one breed of sheep and extended until the animals were weaned. It would have been very enriching if the study had included other sheep breeds and had been projected until the end of lamb fattening or until the moment of shearing. Despite this, the objectives of the study were met, but it is recommended to continue with the research line, due to its relevance to animal health, as well as to public health and the study of pathogens with zoonotic potential associated with domestic production animals.
5 Conclusion
This study showed that fecal samples from ewes had lower bacterial loads compared with wool samples, and that lamb’s wool had higher bacterial loads relative to ewe’s wool. The most prevalent bacteria were E. faecalis from Firmicutes and E. coli from Proteobacteria, indicating that wool contamination likely results from environmental exposure associated with sheep behavior. Handling lambs between birth and weaning, and handling ewes from weaning around day 60 postpartum, poses a risk of zoonotic bacterial pathogen transmission, highlighting a potential public health concern regarding exposure to large intestinal pathogens.
Data availability statement
The original contributions presented in the study are publicly available. This data can be found here: Dryad, doi: 10.5061/dryad.s4mw6m9jd.
Ethics statement
The animal study was approved by Committee for the Ethical Use of Animals in Experiments of the Universidad Austral de Chile (N&z.ousco;241/2015). The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
MG: Investigation, Project administration, Resources, Supervision, Validation, Writing – original draft, Writing – review & editing. LA-A: Methodology, Validation, Visualization, Writing – review & editing. ÁP-V: Data curation, Formal analysis, Writing – review & editing. JL: Investigation, Writing – original draft. AD: Investigation, Writing – original draft.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This study was financed by Fondo Nacional de Desarrollo Científico, Tecnológico y de Innovación Tecnológica (FONDECYT) grants 3160059, 1150934 and Fondo de Financiamiento de Centros de Investigación en Areas prioritarias (FONDAP) 15110027.
Acknowledgments
We would like to thank Dr Armin Mella and Dr. Natalia Ramirez, for their scientific guidance.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Bailey M.T., Dowd S.E., Galley J.D., Hufnagle A.R., Allen R.G., and Lyte M. (2011). Exposure to a social stressor alters the structure of the intestinal microbiota: implications for stressor-induced immunomodulation. Brain Behav Immun 25(3): 397–407. doi: 10.1016/j.bbi.2010.10.023
Barker C. S., Meshcheryakova I. V., Inoue T., and Samatey F. A. (2014). Assembling flagella in Salmonella mutant strains producing a type III export apparatus without FliO. J. Bacteriol. 196, 4001–4011. doi: 10.1128/JB.02184-14
Battisti A., Lovari S., Franco A., Di Egidio A., Tozzoli R., Caprioli A., et al. (2006). Prevalence of Escherichia coli O157 in lambs at slaughter in Rome, central Italy. Epidemiol. Infect. 134, 415–419. doi: 10.1017/S0950268805005236
Beauvais W., Gart E. V., Bean M., Blanco A., Wilsey J., McWhinney K., et al. (2018). The prevalence of Escherichia coli O157:H7 fecal shedding in feedlot pens is affected by the water-to-cattle ratio: A randomized controlled trial. PloS One 13, 192149. doi: 10.1371/journal.pone.0192149
Blanco J., Blanco M., Blanco J. E., Mora A., Gonzalez E. A., Bernardez M. I., et al. (2003). Verotoxin-producing Escherichia coli in Spain: prevalence, serotypes, and virulence genes of 0157:H7 and non-0157 VTEC in ruminants, raw beef products, and humans. Exp. Biol. Med. 228, 345–351. doi: 10.1177/153537020322800403
Brown P. E., Christensen O. F., Clough H. E., Diggle P. J., Hart C. A., Hazel S., et al. (2004). Frequency and spatial distribution of environmental Campylobacter spp. Appl. Environ. Microbiol. 70, 6501–6511. doi: 10.1128/AEM.70.11.6501-6511.2004
Caprioli A., Scavia G., and Morabito S. (2014). Public health microbiology of shiga toxin-producingEscherichia coli. in Microbiol Spectr. 2 (6). doi: 10.1128/9781555818791.ch13
Castellazzi A., Tagliacarne S. C., Soldi S., Perna S., Ziviani L., Milleri S., et al. (2018). Stress and immune function: there is a role for the gut microbiota? J. Clin. Gastroenterol. 52, 66–67. doi: 10.1097/MCG.0000000000001056
Chang J., Yao X., Zuo C., Qi Y., Chen D., and Ma W. (2020). The gut bacterial diversity of sheep associated with different breeds in Qinghai province. BMC Vet. Res. 16, 254. doi: 10.1186/s12917-020-02477-2
Cheng J., Zhang X., Xu D., Zhang D., Zhang Y., Song Q., et al. (2022). Relationship between rumen microbial differences and traits among Hu sheep, Tan sheep, and Dorper sheep. JAS 100, 261. doi: 10.1093/jas/skac261
Cholewińska P., Czyz K., Nowakowski P., and Wyrostek A. (2020a). The microbiome of the digestive system of ruminants-a review. Anim. Health Res. Rev. 21, 3–14. doi: 10.1017/S1466252319000069
Cholewińska P., Szeligowska N., Wojnarowski K., Nazar P., Greguła-Kania M., Junkuszew A., et al. (2023). Selected bacteria in sheep stool depending on breed and physiology state. Sci. Rep. 13, 11739. doi: 10.1038/s41598-023-38785-4
Cholewińska P., Wołoszyńska M., Michalak M., Czyż K., Rant W., and Janczak M. (2020b). Evaluation of changes in the levels of firmicutes and bacteroidetes phyla of sheep feces depending on the breed. Animals 10, 1901. doi: 10.3390/ani10101901
Cholewińska P., Wołoszyńska M., Michalak M., Czyż K., Rant W., Smoliński J., et al. (2021). Influence of selected factors on the Firmicutes, Bacteroidetes phyla and the Lactobacillaceae family in the digestive tract of sheep. Sci. Rep. 11, 23801. doi: 10.1038/s41598-021-03207-w
Clifford R. J., Milillo M., Prestwood J., Quintero R., Zurawski D. V., Kwak Y. I., et al. (2012). Detection of bacterial 16S rRNA and identification of four clinically important bacteria by real-time PCR. PloS One 7, 48558. doi: 10.1371/journal.pone.0048558
Cox P., Griffith M., Angles M., Deere D., and Ferguson C. (2005). Concentrations of pathogens and indicators in animal feces in the Sydney watershed. Appl. Environ. Microbiol. 71, 5929–5934. doi: 10.1128/AEM.71.10.5929-5934.2005
Cui X., Wang Z., Guo P., Li F., Chang S., Yan T., et al. (2023). Shift of feeding strategies from grazing to different forage feeds reshapes the rumen microbiota to improve the ability of Tibetan sheep (Ovis aries) to adapt to the cold season. Microbiol. Spectr. 11, 0281622. doi: 10.1128/spectrum.02816-22
Daca A. and Jarzembowski T. (2024). From the friend to the foe - Enterococcus faecalis diverse impact on the human immune system. Int. J. Mol. Sci. 25, 2422. doi: 10.3390/ijms25042422
Davis E. P., Glynn L. M., Waffarn F., and Sandman C. A. (2011). Prenatal maternal stress programs infant stress regulation. J. Child Psychol. Psychiatry 52, 119–129. doi: 10.1111/j.1469-7610.2010.02314.x
Delgado L., Marín J. F. G., Muñoz M., Benavides J., Juste R. A., García-Pariente C., et al. (2013). Pathological Findings in Young and Adult Sheep Following Experimental Infection With 2 Different Doses of Mycobacterium avium Subspecies paratuberculosis. Vet. Pathol. 50, 857–866. doi: 10.1177/0300985813476066
Deng L., He C., Zhou Y., Xu L., and Xiong H. (2017). Ground transport stress affects bacteria in the rumen of beef cattle: A real-time PCR analysis. Anim. Sci. J. 88, 790–797. doi: 10.1111/asj.2017.88.issue-5
Douglas J. L., Worgan H. J., Easton G. L., Poret L., Wolf B. T., Edwards A., et al. (2016). Microbial diversity in the digestive tract of two different breeds of sheep. J. Appl. Microbiol. 120, 1382–1389. doi: 10.1111/jam.13060
Durso L. M., Harhay G. P., Bono J. L., and Smith T. P. L. (2011). Virulence-associated and antibiotic resistance genes of microbial populations in cattle feces analyzed using a metagenomic approach. J. Microbiol. Methods 84, 278–282. doi: 10.1016/j.mimet.2010.12.008
Durso L. M., Harhay G. P., Smith T. P. L., Bono J. L., DeSantis T. Z., Harhay D. M., et al. (2010). Animal-to-animal variation in fecal microbial diversity among beef cattle. Appl. Environ. Microbiol. 76, 4858–4862. doi: 10.1128/AEM.00207-10
Edelmann M. N., Sandman C. A., Glynn L. M., Wing D. A., and Davis E. P. (2016). Antenatal glucocorticoid treatment is associated with diurnal cortisol regulation in term-born children. Psychoneuroendocrinol. 72, 106–112. doi: 10.1016/j.psyneuen.2016.06.012
Ezemonye L. I. N. and Ikpesu T. O. (2011). Evaluation of sub-lethal effects of endosulfan on cortisol secretion, glutathione s-transferase and acetylcholinesterase activities in clarias gariepinus. Food Chem. Toxicol. 49, 1898–1803. doi: 10.1016/j.fct.2010.10.025
Fonty G., Gouet P., Jouany J. P., and Senaud J. (1987). Establishment of the microflora and anaerobic fungi in the rumen of lambs. J. Gen. Microbiol. 133, 1835–1843. doi: 10.1099/00221287-133-7-1835
Freetly H. C. and Lindholm-Perry A. K. (2023). Rumen and cecum bacteria of beef cattle that differ in feed efficiency fed a forage diet. JAS 101, 292. doi: 10.1093/jas/skad292
Gallardo M. A., Pulido R., and Gallo C. (2011). Fatty acid composition of Longissimus dorsi muscle of Suffolk Doen lambs fed on different dryland forages. Chilean J. Agric. Res. 71 (04), 566–571. doi: 10.4067/S0718-58392011000400011
Gallardo M., Azócar-Aedo L., Arias-Darraz L., Castellaro G., Salgado M., and Cárcamo J. G. A. (2019). survey of bacterial pathogens detected in feces and wool in small ruminants (pilot study). Anim. Vet. Sci. 7, 94–98. doi: 10.11648/j.avs.20190704.13
Garmory H. S., Chanter N., French N. P., Bueschel D., Songer J. G., and Titball R. W. (2000). Occurrence of Clostridium perfringens β2-toxin amongst animals, determined using genotyping and subtyping PCR assays. Epidemiol. Infect. 124, 61–67. doi: 10.1017/S0950268899003295
Gate L., Majumdar R. S., Lunk A., and Tew K. D. (2004). Increased myeloproliferation in glutathione S-transferase π-deficient mice is associated with a deregulation of JNK and janus kinase/STAT pathways. JBC 279, 8608–8616. doi: 10.1074/jbc.M308613200
Gillespie B. E. and Oliver S. P. (2005). Simultaneous detection of mastitis pathogens, Staphylococcus aureus, Streptococcus uberis, and Streptococcus agalactiae by multiplex real-time polymerase chain reaction. JDC 88, 3510–3518. doi: 10.3168/jds.S0022-0302(05)73036-8
Gołás-Pradzynska M., Luszczynska M., and Grazyna Rola Y. (2022). Dairy Products: a potential source of multidrug-resistant Enterococcus faecalis and Enterococcus faecium strains. Foods 11, 4116. doi: 10.3390/foods11244116
Graber H. U., Casey M. G., Naskova J., Stelner A., and Schaeren W. (2007). Development of a highly sensitive and specific assay to detect Staphylococcus aureus in bovine mastitic milk. JDC 90, 4661–4669. doi: 10.3168/jds.2006-902
Guan L., Beig M., Wang L., Navidifar T., Moradi S., Tabaei F., et al. (2024). Global status of antimicrobial resistance in clinical Enterococcus faecalis isolates: systematic review and meta-analysis. Ann. Clin. Microbiol. Antimicrob. 23 (1), 80. doi: 10.1186/s12941-024-00728-w
Guy R. A., Tremblay D., Beausoleil L., Harel J., and Champagne M. J. (2014). Quantification of E. coli O157 and STEC in feces of farm animals using direct multiplex real time PCR (qPCR) and a modified most probable number assay comprised of immunomagnetic bead separation and qPCR detection. J. Microbiol. Methods 99, 44–53. doi: 10.1016/j.mimet.2014.02.002
Hancock D., Besser T., Lejeune J., Davis M., and Rice D. (2001). The control of VTEC in the animal reservoir. Int. J. Food Microbiol. 66, 71–78. doi: 10.1016/S0168-1605(00)00487-6
Hanlon K. E., Miller M. F., Guillen L. M., Echeverry A., Dormedy E., Cemo B., et al. (2018). Presence of Salmonella and Escherichia coli O157 on the hide, and presence of Salmonella, Escherichia coli O157 and Campylobacter in feces from small-ruminant (goat and lamb) samples collected in the United States, Bahamas and Mexico. Meat Sci. 135, 1–5. doi: 10.1016/j.meatsci.2017.08.003
Jami E., Israel A., Kotser A., and Mizrahi I. (2013). Exploring the bovine rumen bacterial community from birth to adulthood. ISME J. 7, 1069–1079. doi: 10.1038/ismej.2013.2
Jhajharia K., Parolia A., Shetty K. V., and Mehta L. K. (2015). Biofilm in endodontics: A review. J. Int. Soc. Prev. Community Dent. 5, 1–12. doi: 10.4103/2231-0762.151956
Khafipour E., Li S., Plaizier J. C., and Krause D. O. (2009). Rumen microbiome composition determined using two nutritional models of subacute ruminal acidosis. Appl. Environ. Microbiol. 75, 7115–7124. doi: 10.1128/AEM.00739-09
Kilic A. and Basustaoglu A. C. (2011). Double triplex real-time PCR assay for simultaneous detection of Staphylococcus aureus, Staphylococcus epidermidis, Staphylococcus hominis, and Staphylococcus haemolyticus and determination of their methicillin resistance directly from positive blood culture bottles. Res. Microbiol. 162, 1060–1066. doi: 10.1016/j.resmic.2011.07.009
Kim M., Kim J., Kuehn L. A., Bono J. L., Berry E. D., Kalchayanand N., et al. (2014). Investigation of bacterial diversity in the feces of cattle fed different diets 1. J. Anim Sci. 92, 683–694. doi: 10.2527/jas.2013-6841
Koren O., Goodrich J. K., Cullender T. C., Spor A., Laitinen K., Kling Bäckhed H., et al. (2012). Host remodeling of the gut microbiome and metabolic changes during pregnancy. Cell. 150, 470–480. doi: 10.1016/j.cell.2012.07.008
Kudva T., Hatfield P., and Hovde P. (1997). Characterization of Escherichia coli O157:H7 and other Shiga toxin-producing E. coli serotypes isolated from sheep. J. Clin. Microbiol. 35 (4), 892-9. doi: 10.1128/jcm.35.4.892-899.1997
Liggenstoffer A. S., Youssef N. H., Couger M. B., and Elshahed M. S. (2010). Phylogenetic diversity and community structure of anaerobic gut fungi (phylum Neocallimastigomycota) in ruminant and non-ruminant herbivores. ISME J. 4, 1225–1235. doi: 10.1038/ismej.2010.49
Ma X., Li M., Zhang J., Luo X., and Sun G. (2022). Interactions of periodic birth and shearing induce outbreak of brucellosis in inner Mongolia. Int. J. Biomath. 15, 2250043. doi: 10.1142/S1793524522500437
Mackos A. R., Varaljay V. A., Maltz R., Gur T. L., and Bailey M. T. (2016). Role of the intestinal microbiota in host responses to stressor exposure. Int. Rev. Neurobiol. 131, 1–19. doi: 10.1016/bs.irn.2016.08.002
Madzingira O. (2016). Shiga toxin-producing E. coli isolated from sheep in Namibia. J. Infect. Dev. Ctries. 10, 400–403. doi: 10.3855/jidc.6903
Malmuthuge N. and Guan L. L. (2017). Understanding host-microbial interactions in rumen: Searching the best opportunity for microbiota manipulation. J. Anim. Sci. Biotechnol. 8, 8. doi: 10.1186/s40104-016-0135-3
Marie-Etancelin C., Tortereau F., Gabinaud B., Martinez Boggio G., Le Graverand Q., Marcon D., et al. (2021). Apart from the diet, the ruminal microbiota of lambs is modified in relation to their genetic potential for feed efficiency or feeding behavior. Front. Microbiol. 12. doi: 10.3389/fmicb.2021.759432
Markle J. G. M., Frank D. N., Mortin-Toth S., Robertson C. E., Feazel L. M., Rolle-Kampczyk U., et al. (2013). Sex differences in the gut microbiome drive hormone-dependent regulation of autoimmunity. Science. 339, 1084–1088. doi: 10.1126/science.1233521
McLoughlin S., Spillane C., Claffey N., Smith P. E., O’Rourke T., Diskin M. G., et al. (2020). Rumen microbiome composition is altered in sheep divergent in feed efficiency. Front. Microbiol. 11. doi: 10.3389/fmicb.2020.01981
Menon R., Watson S. E., Thomas L. N., Allred C. D., Dabney A., Azcarate-Peril M. A., et al. (2013). Diet complexity and estrogen receptor β status affect the composition of the murine intestinal microbiota. Appl. Environ. Microbiol. 79, 5763–5773. doi: 10.1128/AEM.01182-13
Mersha G., Asrat D., Zewde B. M., and Kyule M. (2010). Occurrence of Escherichia coli O157:H7 in faeces, skin and carcasses from sheep and goats in Ethiopia. Lett. Appl. Microbiol. 50, 71–76. doi: 10.1111/j.1472-765X.2009.02757.x
Nagel C., Aurich C., and Aurich J. (2019). Stress effects on the regulation of parturition in different domestic animal species. Anim Reprod. Sci. 207, 156–171. doi: 10.1016/j.anireprosci.2019.04.011
Nasrollahian S., Graham J., and Halaji M. (2024). A review of the mechanisms that confer antibiotic resistance in pathotypes of E. coli. Front. Cell Infect. Microbiol. 14. doi: 10.3389/fcimb.2024.1387497
Parmar N. R., Solanki J. V., Patel A. B., Shah T. M., Patel A. K., Parnerkar S., et al. (2014). Metagenome of mehsani buffalo rumen microbiota: An assessment of variation in feed-dependent phylogenetic and functional classification. J. Mol. Microbiol Biotechnol. 24, 249–261. doi: 10.1159/000365054
Rahal E. A., Kazzi N., Nassar F. J., and Matar G. M. (2012). Escherichia coli O157:H7-Clinical aspects and novel treatment approaches. Front. Cell Infect. Microbiol. 2. doi: 10.3389/fcimb.2012.00138
Ramin A. G., Siamak A. R., and Macali A. (2007). Evaluation on serum glucose, BHB, urea and cortisol concentrations in pregnant ewes. Med. Weter. 63, 674–677.
Rey M., Enjalbert F., Combes S., Cauquil L., Bouchez O., and Monteils V. (2014). Establishment of ruminal bacterial community in dairy calves from birth to weaning is sequential. J. Appl. Microbiol. 116, 245–257. doi: 10.1111/jam.12405
Salcedo J., Frese S. A., Mills D. A., and Barile D. (2016). Characterization of porcine milk oligosaccharides during early lactation and their relation to the fecal microbiome. JDS. 99, 7733–7743. doi: 10.3168/jds.2016-10966
Schmid-Hempel P., Frank S. A., and Manchester M. (2007). Pathogenesis, virulence, and infective dose. PloS Pathog. 3, 1372–1373. doi: 10.1371/journal.ppat.0030147
Schulz J., Runge M., Schröder C., Ganter M., and Hartung J. (2005). Detection of Coxiella burnetii in the air of a sheep barn during shearing. Dtsch Tierarztl Wochenschr. 112, 470–472.
Shanks O. C., Kelty C. A., Archibeque S., Jenkins M., Newton R. J., McLellan S. L., et al. (2011). Community structures of fecal bacteria in cattle from different animal feeding operations. Appl. Environ. Microbiol. 77, 2992–2901. doi: 10.1128/AEM.02988-10
Smith M. (2018). Biosecurity: for the sheep shearer. (Washington: Fact sheet Washington State University. Extension).
Szeligowska N., Cholewińska P., Smoliński J., Wojnarowski K., Pokorny P., Czyż K., et al. (2022). Glutathione S-transferase (GST) and cortisol levels vs. microbiology of the digestive system of sheep during lambing. BMC Vet Res. 18, 107. doi: 10.1186/s12917-022-03201-y
Tanca A., Fraumene C., Manghina V., Palomba A., Abbondio M., Deligios M., et al. (2017). Diversity and functions of the sheep faecal microbiota: a multi-omic characterization. Microb Biotechnol. 10, 541–554. doi: 10.1111/mbt2.2017.10.issue-3
Wang F., Suna H., Kanga C., Yana J., Chen A. J., Fengc X., et al. (2024). Genomic island-encoded regulatory proteins in enterohemorrhagic Escherichia coli O157:H7. Virulence. 15 (1), 2313407. doi: 10.1080/21505594.2024.2313407
Wang L., Zhang K., Zhang C., Feng Y., Zhang X., Wang X., et al. (2019). Dynamics and stabilization of the rumen microbiome in yearling Tibetan sheep. Sci. Rep. . 9, 19620. doi: 10.1038/s41598-019-56206-3
Willett J. and Dunny G. (2024). Insights into ecology, pathogenesis, and biofilm formation of Enterococcus faecalis from functional genomics. Microbiol Mol. Biol. Rev. 89 (1), e0008123. doi: 10.1128/mmbr.00081-23
Xin J., Chai Z., Zhang C., Zhang Q., Zhu Y., Cao H., et al. (2019). Comparing the microbial community in four stomach of dairy cattle, yellow cattle and three yak herds in Qinghai-Tibetan plateau. Front. Microbiol. 10. doi: 10.3389/fmicb.2019.01547
Xue Y., Lin L., Hu F., Zhu W., and Mao S. (2020). Disruption of ruminal homeostasis by malnutrition involved in systemic ruminal microbiota-host interactions in a pregnant sheep model. Microbiome. 8, 138. doi: 10.1186/s40168-020-00916-8
Yang W., Meng X., Zhen Y., Baima Q., Wang Y., Jiang X., et al. (2024b). Strategies and mechanisms targeting Enterococcus faecalis biofilms associated with endodontic infections: a comprehensive review. Front. Cell Infect. Microbiol. 14. doi: 10.3389/fcimb.2024.1433313
Yang X., Wang J., Cheng J., Zhang D., Huang K., Zhang Y., et al. (2024a). Relationship between sheep feces scores and gastrointestinal microorganisms and their effects on growth traits and blood indicators. Front. Microbiol. 15. doi: 10.3389/fmicb.2024.1348873
Keywords: relative abundance, pathogenic phyla bacteria, wool contamination, sheep, human risk, public health
Citation: Gallardo Paffetti M, Azócar-Aedo L, Parra-Vizcaíno A, Larraín JM and Díaz A (2025) Impact of maternal diet and pregnancy type on the abundance of zoonotic bacteria (Firmicutes and Proteobacteria) in sheep feces and wool. Front. Anim. Sci. 6:1583281. doi: 10.3389/fanim.2025.1583281
Received: 25 February 2025; Accepted: 07 April 2025;
Published: 17 July 2025.
Edited by:
Michael D. Flythe, United States Department of Agriculture, United StatesReviewed by:
Xiongxiong Cui, Guizhou University, ChinaMa YuHao, Naval Military Medical University, China
Copyright © 2025 Gallardo Paffetti, Azócar-Aedo, Parra-Vizcaíno, Larraín and Díaz. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Maria Gallardo Paffetti, bWFyaWEuZ2FsbGFyZG9AdW1heW9yLmNs