 Luis F. Cartuche-Macas1*
Luis F. Cartuche-Macas1* José R. Pérez-González2
José R. Pérez-González2 Miguel A. Gutiérrez-Reinoso3,4Anixon G. Leal-González5
Miguel A. Gutiérrez-Reinoso3,4Anixon G. Leal-González5 Manuel García-Herreros6,7*
Manuel García-Herreros6,7*- 1Escuela Superior Politécnica Agropecuaria de Manabí (ESPAM-MFL), Calceta, Ecuador
- 2Universidad Politécnica Territorial de Maracaibo (UPTM), Maracaibo, Venezuela
- 3Facultad de Ciencias Agropecuarias y Recursos Naturales, Carrera de Medicina Veterinaria, Universidad Técnica de Cotopaxi (UTC), Latacunga, Ecuador
- 4Laboratorio de Biotecnología Animal, Departamento de Ciencia Animal, Facultad de Ciencias Veterinarias, Universidad de Concepción (UdeC), Chillán, Chile
- 5Estación Local Carrasquero, Hacienda El Laral, Instituto Nacional de Investigaciones Agrícolas (INIA), Carrasquero, Venezuela
- 6Instituto Nacional de Investigação Agrária e Veterinária (INIAV), Santarém, Portugal
- 7CIISA-AL4AnimalS, Faculty of Veterinary Medicine, University of Lisbon, Lisbon, Portugal
Criollo Limonero cattle (Bos taurus) is an endangered bovine breed reared under the dry tropical areas of Venezuela. So far, the inbreeding and genetic diversity values of the Criollo Limonero cattle have not been assessed after more than five centuries of environmental adaptation. The main aim of the present research was to carry out a detailed analysis of the demographic status, reproductive performance, inbreeding and genetic diversity of the endangered Criollo Limonero dairy cattle using official genealogical information. Six populations were considered: historical population (individuals born between 1969–2023), and five populations born from 1974 to 2023 taken at 10-year intervals (1974–1983; 1984–1993; 1994–2003; 2004–2013; 2014–2023). Population structure variables analysed were: pedigree completeness index (PCI), number of equivalent (GEqu), complete (GCom) and maximum (GMax) generations and generation interval (GI). DG variables were: inbreeding (F), inbreeding increment (ΔF), average relatedness (AR), co-ancestry (C), Non-random mating (α), effective population size (Ne) and genetic conservation index (GCI). The gene origin probability variables were: number of founders (f), effective number of founders (fe) and ancestors (fa), number of equivalent genomes (fg), fe/fa and fg/fa ratio and DG losses. The databases were analysed by ENDOG, POPREP, and CFC software. The results indicate that the Criollo Limonero dairy breed has suffered an accelerated genetic diversity loss mainly due to the bottleneck effect and genetic drift. The average number of offspring of both males and females decreased drastically during the last period. Gmax, Gcom and GEqu showed 11.70, 2.15 and 4.58 values, respectively. Moreover, F = 4.45%, AR = 3.10%, ΔF = 0.38%, C = 1.55%, α = 0.0051, GCI = 10.35 values were obtained. Ne values showed that there is a moderate-high risk of increased inbreeding, suggesting that the population could be genetically sustainable in the medium to long term if appropriate selection and breeding schemes are carried out. In conclusion, preventing genetic diversity loss and avoiding high inbreeding values should be a priority in order to conserve the endangered Criollo Limonero dairy cattle.
1 Introduction
The introduction of cattle from Spain to the Americas began in the 15th century on the second voyage of Christopher Columbus and spread first to the Caribbean islands and then to the mainland (Martínez et al., 2012). More than 500 years have passed since cattle introduction to Central and South America and many of these animals have formed creole cattle populations characterised mainly by their adaptation, resistance to different diseases, environmental conditions, and production systems (Marcuzzi et al., 2025). In the beginning, cattle were used for various purposes, such as milk, meat/fat/hide and draught production (Mignon-Grasteau et al., 2005). Later, depending on the environment, climate, production system, and natural selection, they adapted and showed unique characteristics according to their geographical location. Over time, specialised cattle breeds were introduced, with the aim of increasing production performance by crossbreeding them with local creole populations (Marcuzzi et al., 2025). As a result, some creole cattle populations were displaced to areas where the imported breeds could not survive, and in other creole cattle breeds even disappeared (Cabezas Congo et al., 2019).
In Venezuela, Criollo Limonero cattle are kept under the supervision of conservation and genetic improvement programmes, always preserving the adaptive advantages such as the ability to reproduce (fertility) and survive long periods of time (longevity) as well as their hardiness, expressed in their ability to graze and take advantage of coarse, fibrous forage of low nutritional value, great ability to traverse steep, non-mechanisable terrain, in addition to a docile temperament, with resistance to parasites and adaptation to high temperatures and wetter seasons (Araujo-Febres, 2023). The Criollo Limonero cattle are in serious danger of extinction due to several factors, such as indiscriminate crossbreeding with specialised imported beef and dairy breeds, lack of control of genetic diversity given the relatively low population size, lack of conservation policies and funding for conservation centres, and the lack of organisation of producers to conserve and disseminate these genetic resources (Landaeta-Hernández et al., 2011). The Criollo Limonero cattle were originated in the 1950s from other creole cattle raised in the region of Rivas (Nicaragua) and Costa Rica (Corrales, 2011; Contreras et al., 2012); however, the conservation and selection in Venezuela began close to the Limon River in the region of Zulia. Later, at the end of the 1960s, the Carrasquero Experimental Station (Hacienda Laral, Venezuela) was created, managed through the National Institute for Agricultural Research (INIA) with the aim of carrying out a detailed programme for the recovery of Criollo Limonero cattle (Villasmil-Ontiveros et al., 2008; Araujo-Febres, 2023). Since the 1970s, in order to avoid inbreeding problems, a breeding and selection management strategy was implemented using five genetic lines, following circular mating strategies (Kimura and Crow, 1963; Cockerham, 1970; Nomura and Yonezawa, 1996; Yenen, 2007; Gutiérrez-Reinoso et al., 2020) similar to different creole breeds in Colombia (Martinez et al., 2023). This process has allowed the population to maintain census levels below 1% from the mid-1980s to the mid-2000s, maintaining its status as an endangered breed (Yenen, 2007; Villasmil-Ontiveros et al., 2008). This breed is characterised by its milk production in tropical areas due to its great adaptability and resistance to adverse environmental conditions while maintaining fertility and dairy performance (Rojas-Espinoza et al., 2023). Conservation programmes for native and creole cattle in several countries monitor the genetic diversity of their populations through genealogical and molecular information, detecting a reduction in their population that has led to an increase in inbreeding (Cartuche-Macas et al., 2024, 2025b). Similar effects have been observed in breeds such as Romosinuano in Mexico and Blanco Orejinegro in Colombia (Ramírez-Valverde et al., 2018; Ocampo Gallego et al., 2020).
Several studies on genetic diversity have been carried out in different creole breeds of Ibero-America using genealogical information, for example, in Colombia the Costeño con Cuernos, Blanco Orejinegro, Romosinuano, and Sanmartinero breed (Martinez et al., 2008), in Nicaragua the Reyna breed (Corrales et al., 2010), in Mexico the tropical dairy criollo and Sardo Negro breed (Rosendo Ponce et al., 2018; Viveros et al., 2021), in Venezuela the Carora breed (Tullo, 2011), and in Brazil the Lageana breed (Pezzini et al., 2018). Unfortunately, there is not much information on the current status of the Criollo Limonero dairy breed, and therefore, there is a need to develop studies at the genealogical and molecular level in order to provide information on the current status of the genetic diversity of this breed, as well as to reduce the potential increase in inbreeding rates that it may be suffering from. Therefore, the main objective of the present study was to carry out an analysis of the demographic status, reproductive performance, inbreeding and genetic diversity of the Criollo Limonero dairy cattle using official genealogical information with emphasis on the ancestors and founders of the different genetic lines. Thus, the aim of the study could contribute to the development of future selection and breeding schemes necessary to conserve this breed which is currently in a critical situation.
2 Materials and methods
2.1 Ethical statement
The present research did not require any animal handling, since the study was directly carried out using the records and databases provided by National Institute for Agricultural Research (INIA, Venezuela). A possible limitation related to the database and records could be related to the relatively limited size of the data; however, considering that the Criollo Limonero is an endangered breed, the sample size was considered sufficient to carry out the present study because it was similar to other databases related to other endangered breeds.
2.2 Genealogical database
The genealogical database was provided by National Institute for Agricultural Research (INIA) of Venezuela. A total of 3,559 registered individuals were used, including 1,178 bulls and 2,381 cows, born between 1969 and 2023. For the analysis, six populations were considered: historical population (individuals born between 1969–2023), and five populations born from 1974 to 2023 taken at 10-year intervals (1974–1983; 1984–1993; 1994–2003; 2004–2013; 2014–2023) that included 850, 699, 460, 1088, and 462 individuals, respectively. Each population (reference population) was considered to be the one encompassing individuals with known sire and dam from the populations described above. Populations were defined because calculations related to genetic diversity, gene origin probabilities, and founder analyses can only be performed by considering only individuals with both parents known or by comparing them with historical and current data sets as suggested by Casanovas Arias et al. (2020). ENDOG (v 4.8) software was used to perform demographic and genetic analyses to quantify and trace genetic diversity back to ancestors (Gutiérrez and Goyache, 2005).
2.3 Demographic structure
First, the number of births was calculated in order to determine the maximum and average number of calves per sire/dam. The total and mated dam/sire ratio was also calculated dividing the total number of dams by the total number of sires, as well as the number of breeding dams divided by breeding sires, respectively. In addition, the pedigree completeness index (PCI) was calculated following the assumptions of Navas et al. (2017) from the first to the fifth generation, and also the number of maximum generations (GMax), number of complete generations (GCom) and number of equivalent generations (GEqu) in the five defined populations. Moreover, the generation interval (GI) was calculated for the 4 gametic pathways from sire and dam to son and daughter, respectively, according to James (1972). For this purpose, the record of the birth date of each individual together with that of its parents was used. In parallel, gene flow between herds was evaluated according to the contribution of sires to the population (Vassallo et al., 1986).
2.4 Strategy for breeding and selection
In order to reduce the increase in inbreeding rate, a circular mating strategy was implemented from the 1970s onwards (Kimura and Crow, 1963; Cockerham, 1970; Nomura and Yonezawa, 1996; Gutiérrez-Reinoso et al., 2022) with the formation of five families (Bella Vista, Hachiote, Joaquín Reyna, Bonita and Founders/Purchased) (Yenen, 2007) under a specific scheme (Figure 1). The dams of each family were mated with the sires of the preceding family (counterclockwise) and if the offspring was male it remained in the dam’s family (clockwise); however, and if the offspring was female it was transferred to the sire’s family (counterclockwise). The Criollo Limonero cattle breed is characterised by its milk production in tropical areas. Individuals in the present study were bred and kept in a dry tropical climate with average temperatures of 24.7 °C and 1,000 mm average annual rainfall (Figure 2).
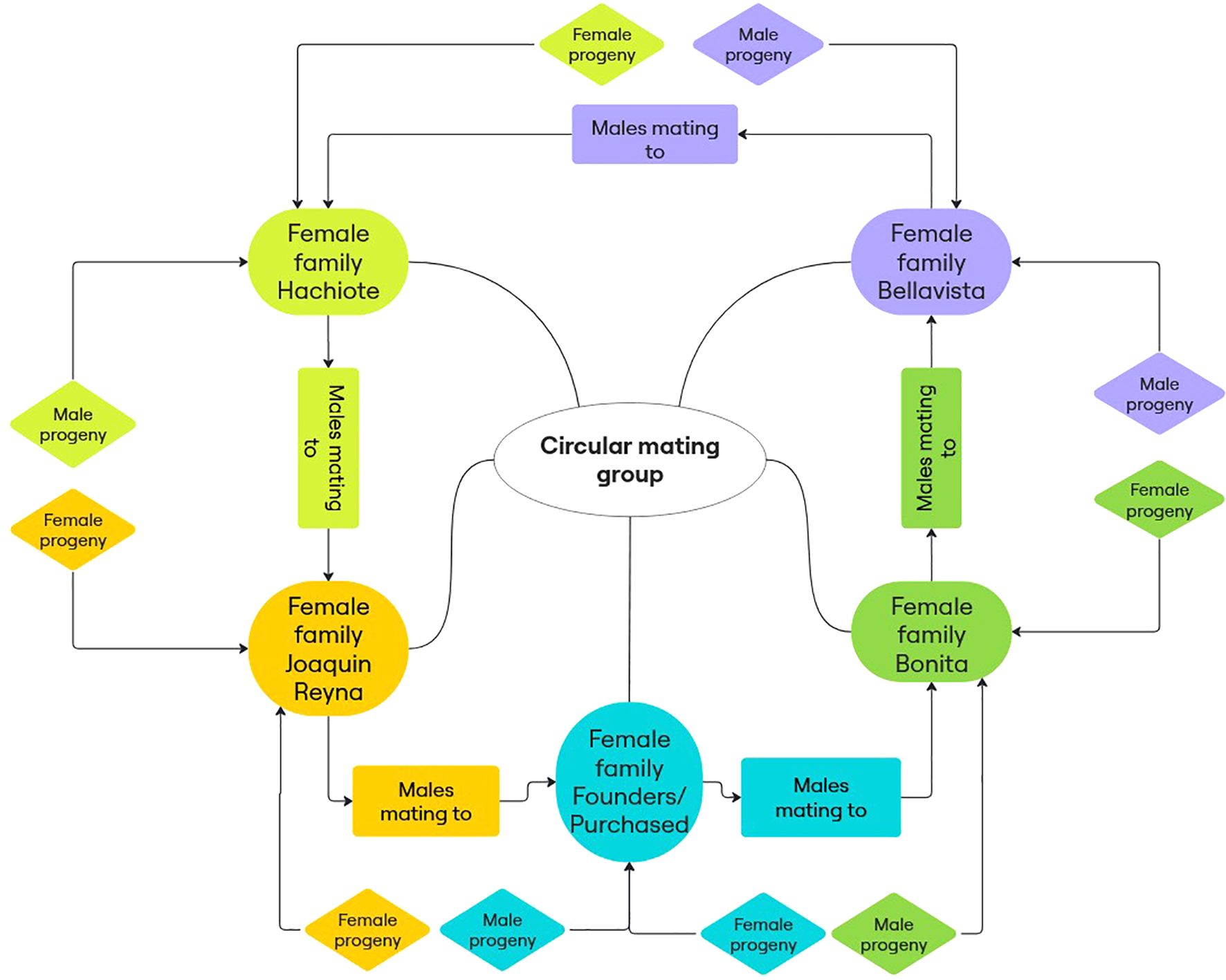
Figure 1. Breeding and selection scheme of Criollo Limonero dairy cattle during the last five decades (1970s–now).

Figure 2. Criollo Limonero cattle from Venezuela. Criollo Limonero sire (left) and dam (right).
2.5 Inbreeding, inbreeding increment, average relatedness, coancestry, and non-random mating
Inbreeding coefficient (F): The F has been defined as the probability that two alleles taken at random are identical per offspring. The F was calculated according to the algorithm proposed by Meuwissen and Luo (1992) and recursive method (Aguilar and Misztal, 2008). The inbreeding increment (ΔF) per generation was calculated using the equation proposed by Gutiérrez et al. (2008):
where Ft and Ft−1: average inbreeding of the tth generation (i = 1,…, t).
Average relatedness (AR): Each individual’s average relatedness coefficient (AR) was defined as the probability that two related individuals have inherited a particular allele of a single locus/gene from their common ancestor (this allele is known as IBD: identical by descent). AR was defined as the probability that a randomly selected allele from a population belongs to a specific individual, which was calculated using the vector c, where each element corresponds to the respective AR of an individual, defined by Gutiérrez and Goyache (2005):
where “A” is the n x n parentage matrix.
Coancestry (C): The C between two individuals is the probability that the genes, taken at random from each of the individuals, are identical by descent (Gómez-Romano et al., 2016). As a result, the C between two individuals is the F of their potential offspring. The C between two individuals is equal to the F of their offspring if the individuals are related (Nietlisbach et al., 2017). It was also used to analyse the degree of relatedness and non-random mating, α within breeds. The coancestry was calculated according to the algorithm proposed by Colleau and Sargolzaei (2008).
Individual coancestry rate (∆C): The ∆C for each generation was calculated following the methods described by Cervantes et al. (2011):
where tb and ta are the number of equivalent complete generations and Cba is the C for individuals b and a.
Non-random mating (α): The α was calculated as the correlation of genes between two individuals in relation to the correlation of genes taken at random from the population (α) according to Caballero and Toro (2000). It indicates the degree of deviation from Hardy-Weinberg proportions and is related to the F according to Sheppard and Wright (1971):
Effective size (Ne): The Ne was defined as the number of males and females contributing to genetic variability in a population. It was calculated as proposed by Hill (1981):
Three additional Ne values were also estimated using the regression coefficient (b) of the individual inbreeding coefficients on: i) the full number of generations, ii) the maximum number of generations and iii) the equivalent number of full generations, with the regression coefficient corresponding to the increment between the two inbreeding generations (Fn − Fn−1 = b) (Gutiérrez et al., 2008):
Genetic Conservation Index (GCI): The GCI was calculated from the genetic contribution of all founders, considering the proportion of genes from a founder animal in the pedigree under analysis according to Alderson et al. (1992) and Mandal et al. (2020). The following equation was used:
where “pi” is the proportion of genes of founder “i” in the individual’s pedigree.
2.6 Gene origin probabilities and ancestral contributions
Number of founders (f): The f was defined as those individuals with unknown parents assumed to be unrelated and have an inbreeding coefficient of 0.
Effective number of founders (fe): The fe was defined as the number of founders that contribute equally and are expected to produce the same genetic diversity as the study population. It was calculated from the following equation (Lacy, 1989):
where “qk” is the gene origin probability from ancestor “k”, and “f” is the real number of founders.
Effective number of ancestors (fa): The fa was defined as the minimum number of ancestors that are not necessarily founders and that account for the full genetic diversity of a population according to Boichard et al. (1997):
where “pk” is the marginal contribution of an ancestor “k” that is not explained by other chosen ancestors, and “f” is the real number of founders.
Number of founder genome equivalents (fg): The fg was defined as the number of founders that would be expected to produce the same genetic diversity as the population under study if the founders were equally represented and no allele loss occurred. This was calculated from twice the inverse of the average C according to Caballero and Toro (2000):
Fe/fa and fg/fa ratios were calculated to determine genetic bottlenecks and random genetic drift, respectively.
Genetic contributions: The genetic contributions were calculated for the top ten ancestors with the maximum genetic impact between 1999 and 2023. The marginal contribution of each major ancestor “j” was calculated as proposed by Boichard et al. (1997).
The CFC v.1.0 software was used to calculate ancestral contributions and gene origin probabilities (Sargolzaei et al., 2006).
2.7 Genetic diversity and loss for genetic contributions of founders, genetic drift and bottle necks
Genetic diversity (GD): The GD was calculated using the following equation:
Genetic diversity loss (GD-loss): The population GD-loss from the founder generation was calculated using 1 − GD. The GD-loss due to unequal contribution of founders was calculated according to the formula proposed by Caballero and Toro (2000) using 1 − GD*:
The unequal contribution of founders relates to the fact that the genetic contributions of founders of specific populations may be of different proportions due to past directional mating (human-mediated or not) during the process of population shaping. The difference between GD and GD* indicates the GD-loss due to genetic drift accumulated from the population founding (Lacy, 1989), and the effective number of non-founders (Nenf).
2.8 Statistical analysis
The software used for the database analysis was ENDOG v. 4.8 (demographic-derived parameters) (Gutiérrez and Goyache, 2005), POPREP (reproductive and generation interval-derived parameters) (Groeneveld et al., 2009), and CFC (demographic-derived parameters) (Sargolzaei et al., 2006). Finally, genetic diversity indices and gene origin probability were obtained using the equations and formulas shown in the material and methods subsections above.
3 Results
3.1 Population structure and reproductive performance evolution
3.1.1 Demographic structure
Table 1 shows the results of the Criollo Limonero population structure in the different chronological periods evaluated. It was observed that the population of individuals within each period did not exceed 1,000 individuals, except for the individuals recorded during the period 2004–2013. Similarly, there was a decrease in the number of sires and dams during the last period studied. Finally, the number of individuals with progeny also decreased drastically and was even lower in some of the periods analysed compared to the number of individuals recorded without progeny. This demographic reduction could be generated by factors such as lack of investment for maintenance, departure of animals without minimum replacements being considered, and lastly, environmental factors such as droughts.
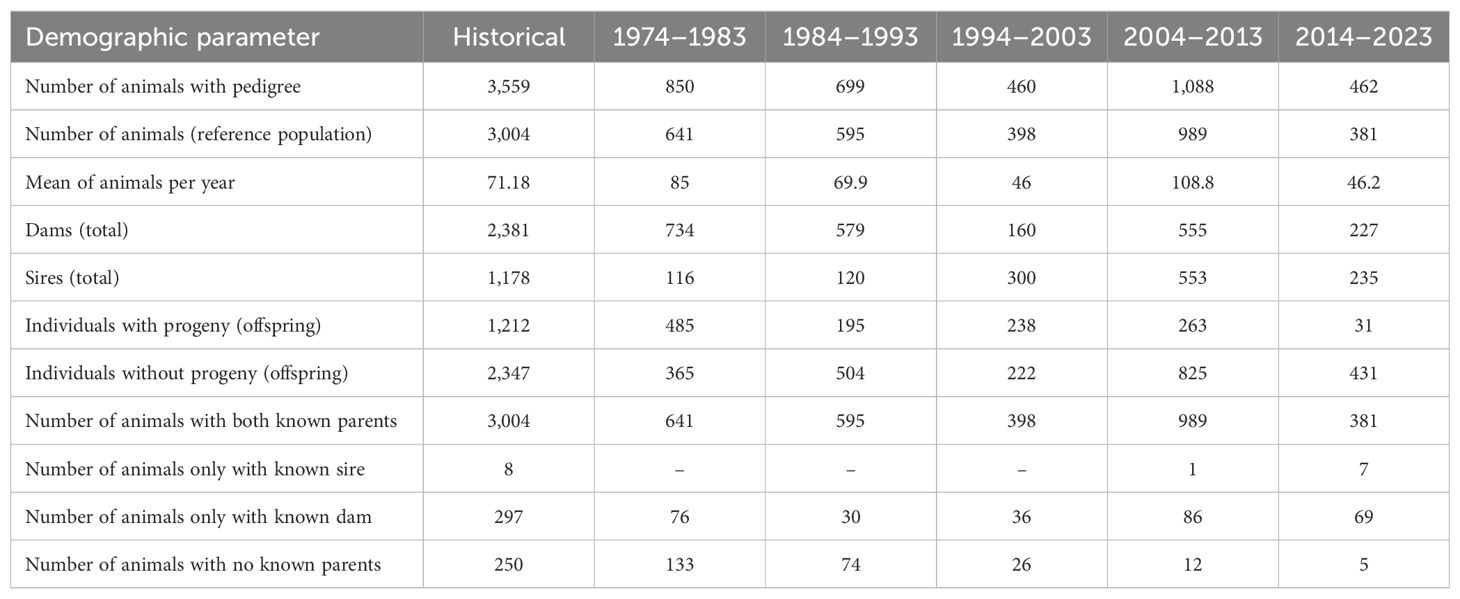
Table 1. Population structure-derived parameters in Criollo Limonero cattle.
3.1.2 Reproductive performance: number of calvings and their evolution
With regard to the reproductive parameters shown in Table 2, it was observed that the average number of offspring of both males and females decreased drastically during the last period studied, i.e. there were many females without offspring. Similarly, the same effect was observed for males (Table 1). On the other hand, Figure 3 shows that the distribution of females/year/number of births of the same females decreased in the periods 1988–2002 and 2015–2023. Moreover, since 2016 there are very few females with 1 and 2 births, although this number recovers from 2022 onwards. It should also be noted that since 2015 there has been a marked decrease in the female population.
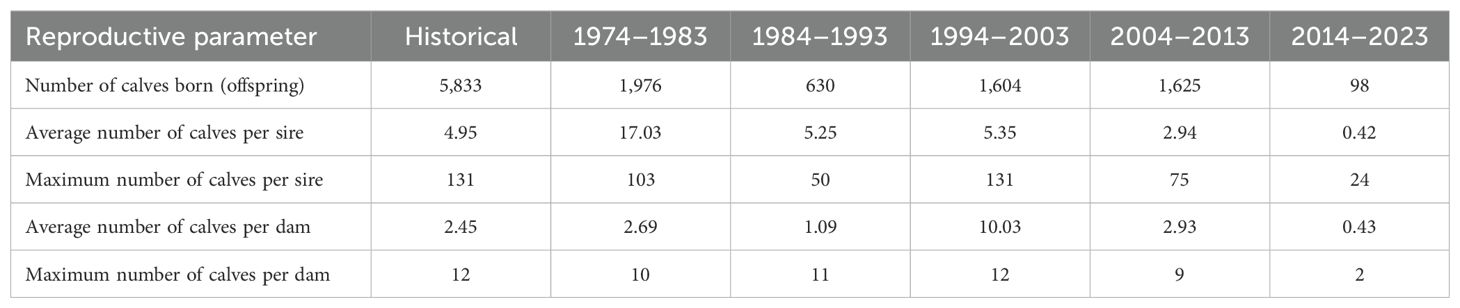
Table 2. Reproductive performance-derived parameters in Criollo Limonero cattle.
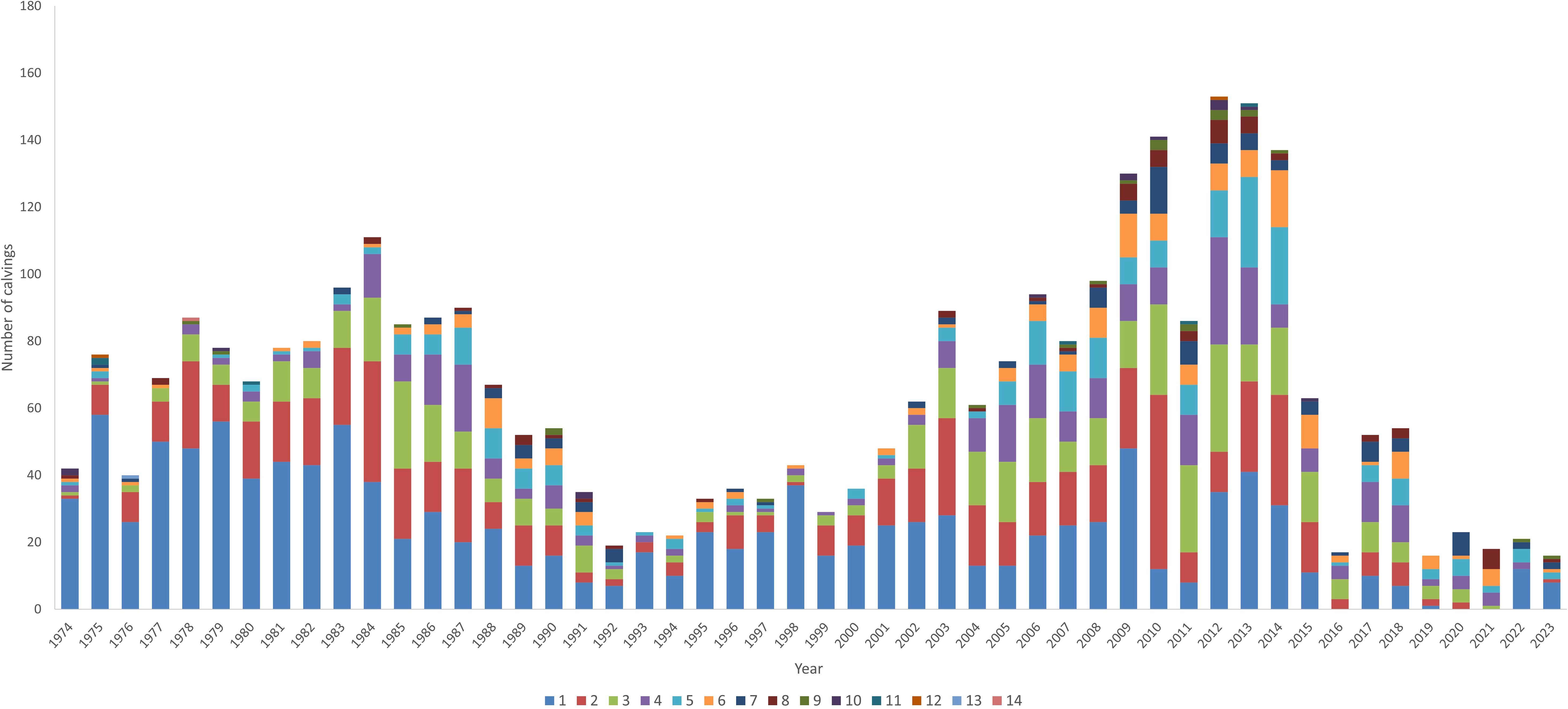
Figure 3. Distribution of dams by the number of calves/year (offspring) in a population of Criollo Limonero cattle.
3.2 Pedigree completeness and generation intervals in Criollo Limonero cattle
3.2.1 Pedigree completeness-derived parameters
The pedigree completeness for the first generation improved in each of the periods studied from 79.88% to 90.69% in the last period (Table 3). Furthermore, between the first and third generation pedigree completeness values were higher than 78.17%. These values were reflected in the maximum, complete and equivalent generations with values of 11.70, 2.15 and 4.58, respectively. Pedigree integrity values in the first two periods and between the second and fifth generation were lower than 60% which makes the diversity parameters not so reliable, but thereafter, the values were above 60% improving their reliability.
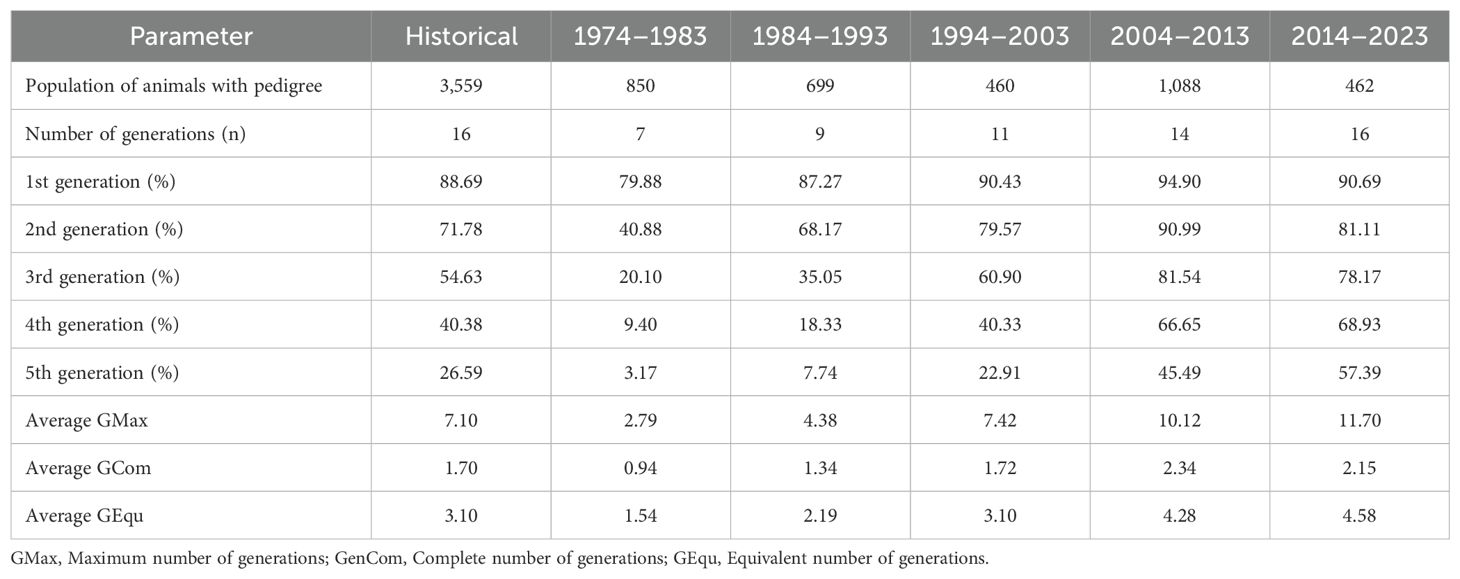
Table 3. Pedigree completeness-derived parameters in a population of the endangered Criollo Limonero cattle.
3.2.2 Generation intervals
Figure 4 shows the evolution of the generation interval in all periods. The estimated total generation interval was 6.19, and for the father–son, father–daughter, mother–son and mother–daughter paths were 6.59, 5.68, 7.42 and 6.47 years, respectively. It can be observed that the trend of the GI of the total population is relatively stable with slight increases and decreases over time. On the other hand, it was observed that between the years 2010–2014 there was an increase in the father–son pathway and a decrease in the mother-son pathway, while a decrease in the mother–daughter pathway was observed from 2006 onwards, and an increase in the mother-daughter pathway since 2009.
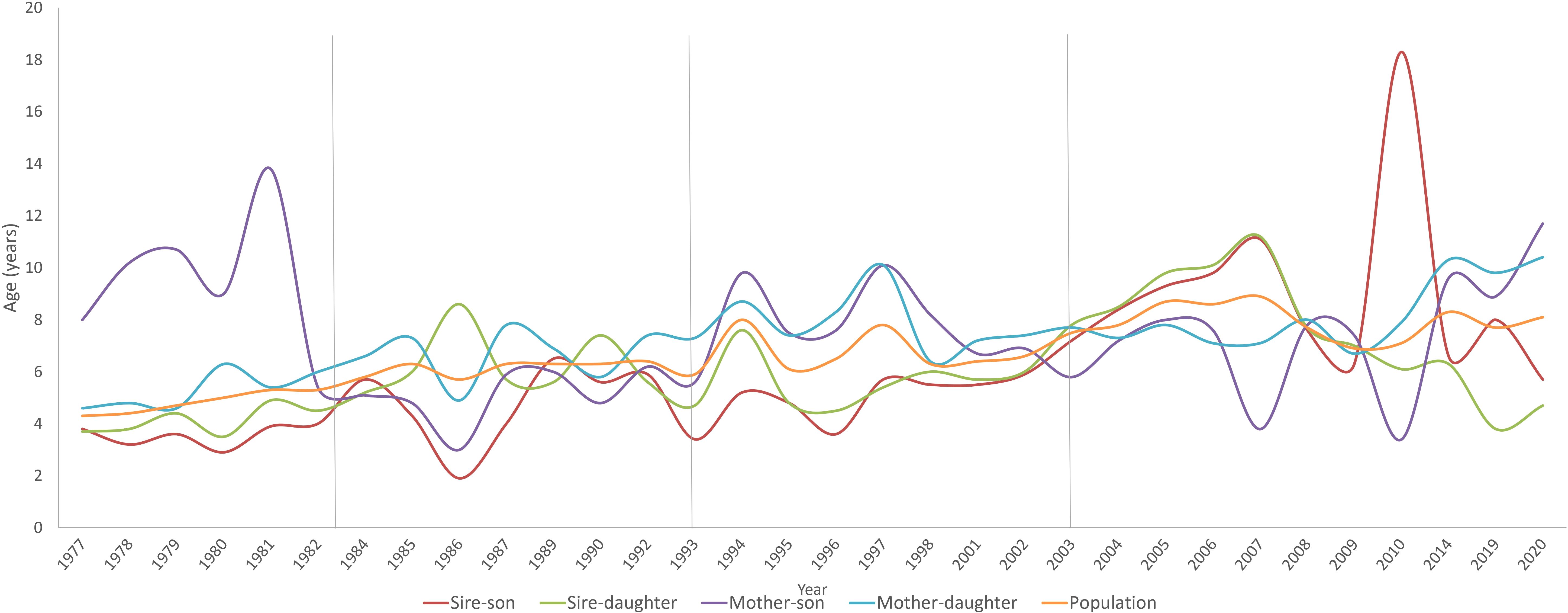
Figure 4. Generation interval (GI) evolution regarding the four gametic pathways in Criollo Limonero cattle.
3.2.3 Inbreeding, average relatedness, coancestry, and non-random mating
The inbreeding coefficient was between 0.16–2.05% and 0.61–4.45% using the Meuwisen and Recursive methods, respectively (Table 4). Furthermore, the increase in the inbreeding index remained stable in the last two periods. On the other hand, it was observed that the inbreeding population in this population was above 66.0% in the last two periods. The GCI values increased considerably, reaching 10.35 between 2014 and 2023 compared to 2.92 between 1974 and 1983. Finally, it was observed that α values were positive from the period 2004 to the present.
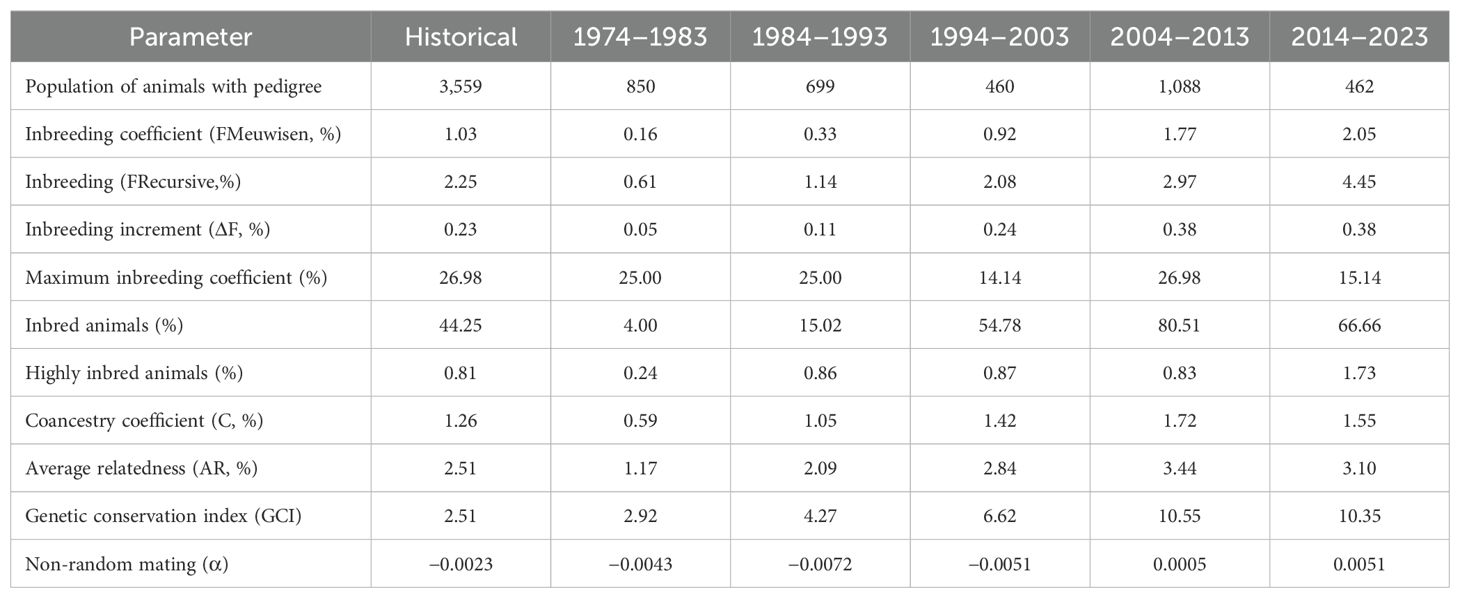
Table 4. Inbreeding (F), average relatedness (AR), coancestry (C), and non-random mating (α).
Figure 5 shows the evolution of genetic diversity parameters. In the same figure it was observed that during the last period there was a drastic drop in these parameters which could be related to the population decline as shown in Table 1. The increase in inbreeding and co-ancestry had a stable evolution over time, contrary to the rest of the parameters that had constant increases and decreases over time.
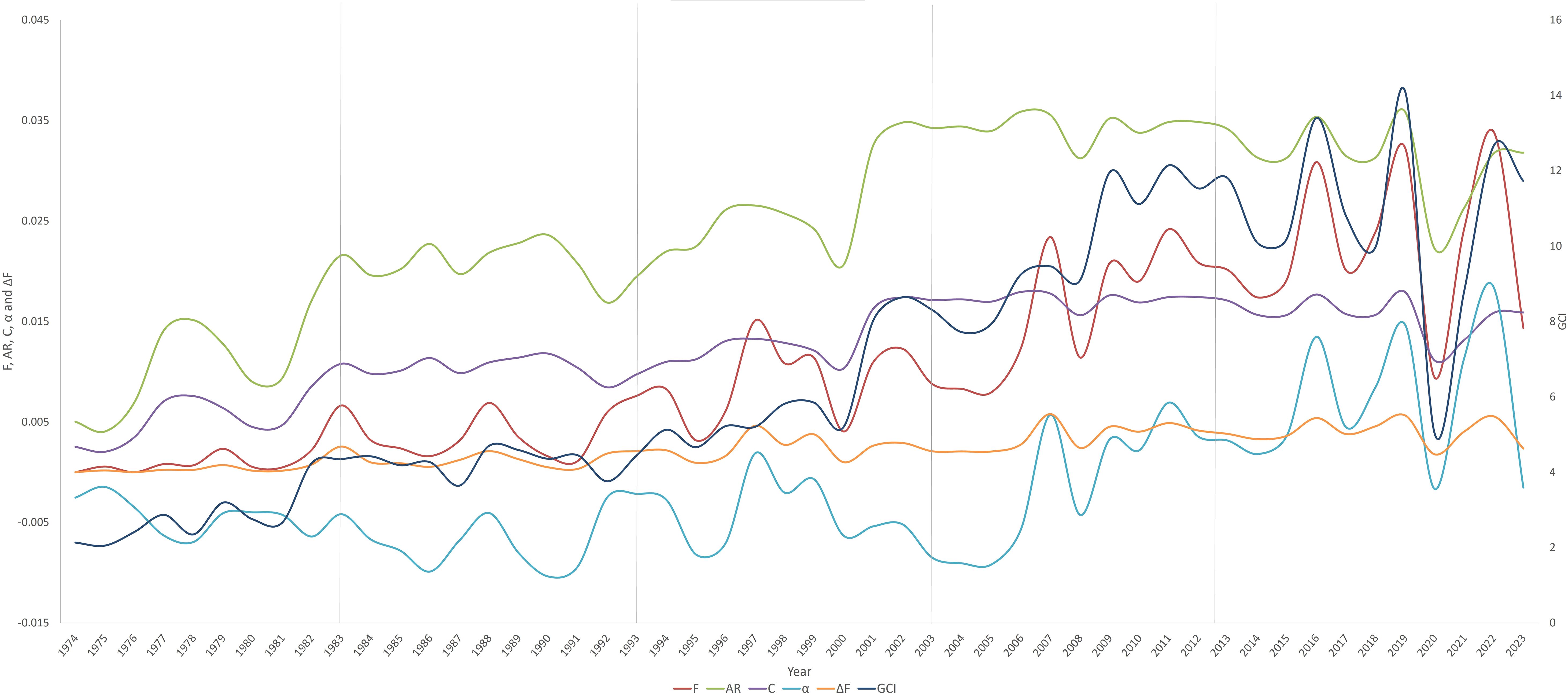
Figure 5. Inbreeding (F), average relatedness (AR), coancestry (C), and non-random mating (α) evolution (yearly) in Criollo Limonero cattle.
3.2.4 Effective size (Ne)
The Ne analysed by the population census method was estimated to be around 100, while that obtained by the generation method was between 100 and 200, except for the period 1974–1983 (Figure 6), taking into account the constant average number of offspring of the parents. The relationship between Ne and the number of offspring for both mothers and fathers was irregular in all periods analysed. The reduction of the population census (both for males and females) causes the Ne to be slightly reduced, generating a not so real effect of this reduction, increasing the appearance of genetic drift or bottlenecks that would put this breed in serious problems of extinction.
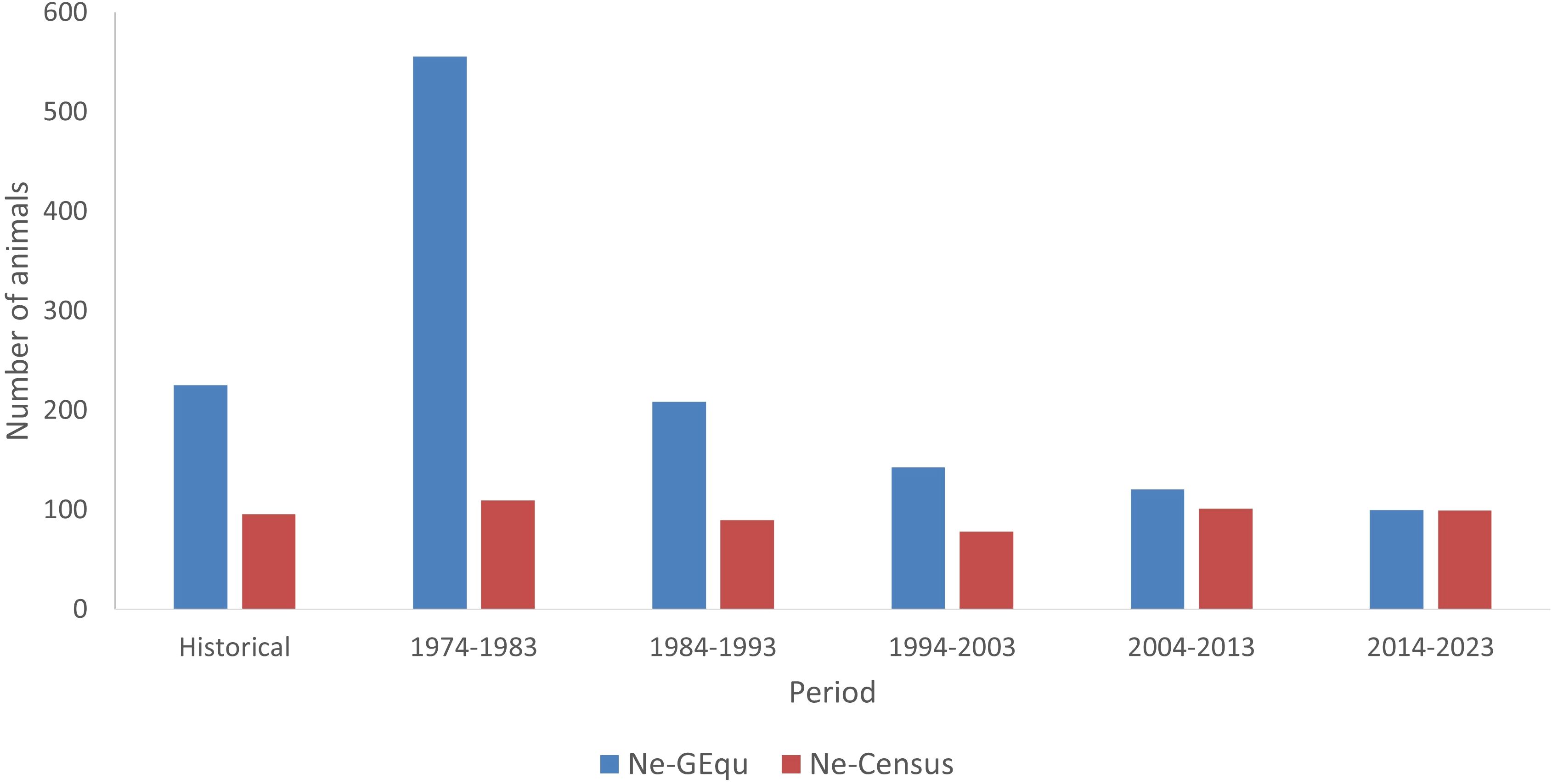
Figure 6. Effective size (Ne) evolution in Criollo Limonero cattle.
3.3 Gene origin probability, ancestral contributions and genetic diversity in Criollo Limonero cattle
3.3.1 Gene origin probability and ancestral contributions
The effective number of founders (fe) and ancestors (fa) remained between 69.35–50.84 and 49–25 between the first and last periods, marking a tendency towards reduction over time, which had a direct impact on the values of fe/fa that went from 1.42 to 2.03 in the same periods (Table 5). These values indicated a poor balance between the contributions of founders and ancestors, as well as a strong presence of the ‘bottleneck effect’. Regarding fg/fe, a reduction from 0.50 to 0.23 was observed between the first and the last period, taking into account that the lower the fg/fe ratio, the greater the effect of genetic drift. In other words, this effect possibly occurred in this population due to the various agro-climatic factors that this population has suffered over time. Similarly, the explanation of genetic diversity by ancestry was reduced to 4, 9 and 20 for 25%, 50% and 75%. On the other hand, among the 10 most important ancestors there was only one female in the last two periods. In all periods, the first ancestor contributed about 10% to the genetic diversity of the population, reaching a total of 66 offspring in the last period.
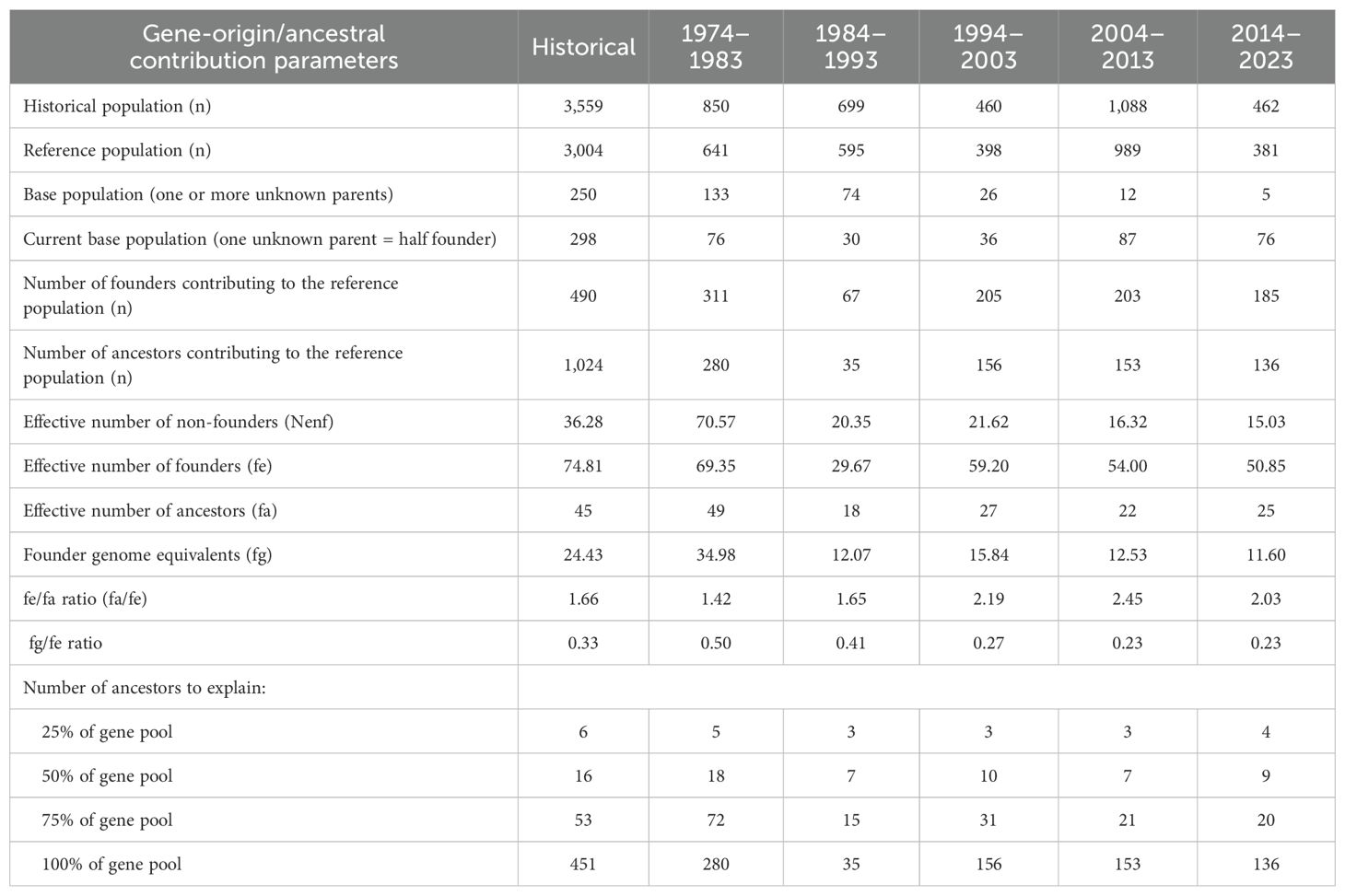
Table 5. Gene origin probability and ancestral contributions in the endangered Criollo Limonero cattle.
3.3.2 Genetic diversity loss
The parameters of genetic diversity analysed in the present study are shown in table 5. It was observed that the loss of genetic diversity (GD) in all periods was ~96%. On the other hand, genetic diversity (GD*) due to unequal contribution of founders was reduced from 99.28% to 99.02% between the first and last periods. In addition, the highest loss of GD (1-GD) in the reference population in the last period was 4.31%.
In general, an increase in total DG loss can be observed, while DG loss due to random genetic drift has increased over the periods (Table 6; Figure 7). Thus, the value of DG loss due to unequal contribution of founders has slightly increased over time (Figure 7). The values of NeFi and NeCi in the last period were 131.5 and 16.13, respectively, with the former having a tendency to decrease and NeCi to stay around a value of 16 (Table 6).
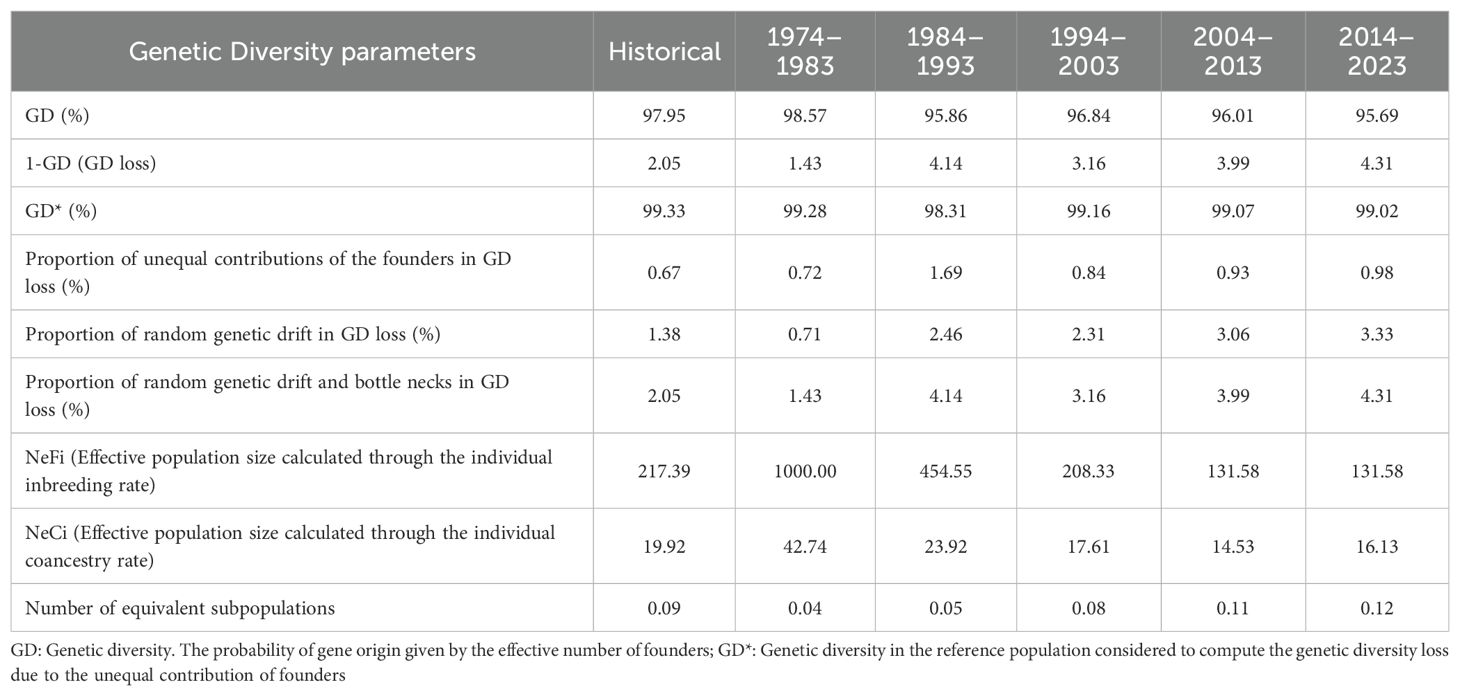
Table 6. Genetic diversity-derived parameters in the endangered Criollo Limonero cattle.
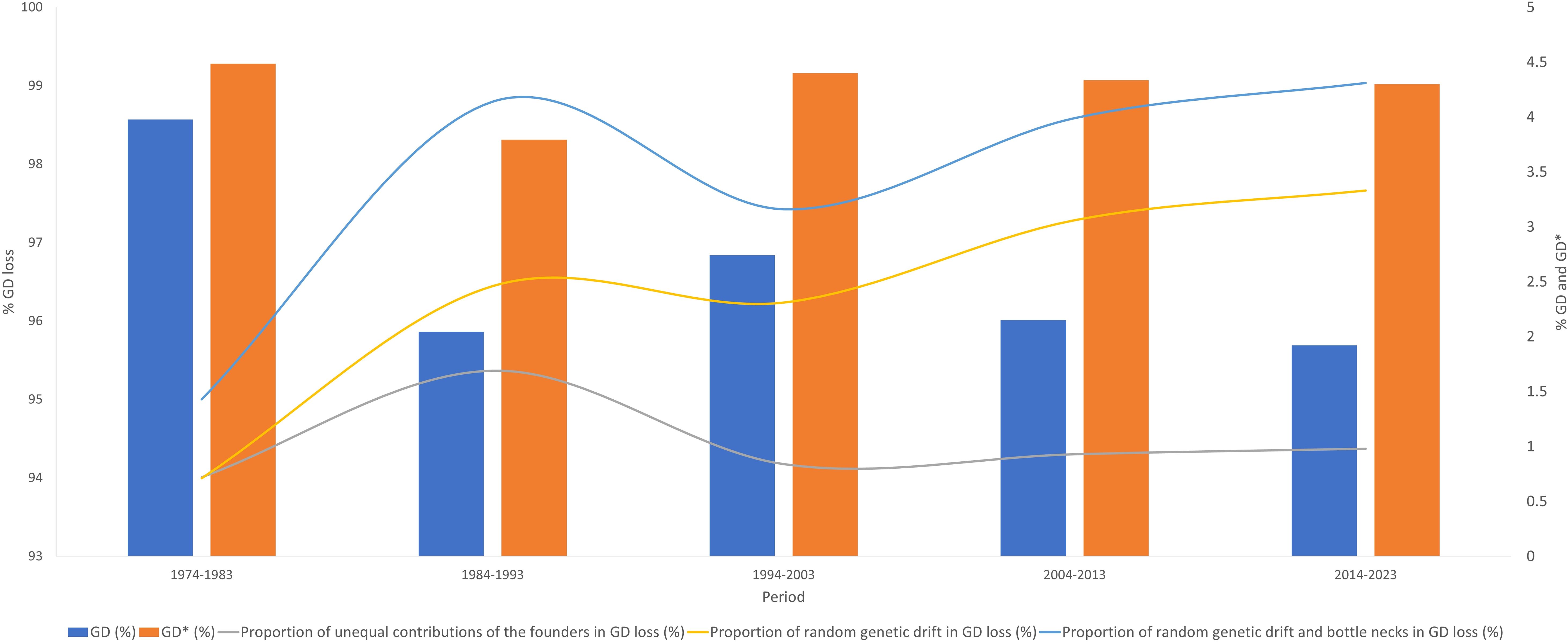
Figure 7. Proportion of the unequal contributions of the founders, random genetic drift, and bottle necks in genetic diversity (GD) loss in Criollo Limonero cattle.
4 Discussion
In the present research, the population structure, reproductive performance, inbreeding, and genetic diversity was analysed over time from 1974 to 2023 in the endangered Criollo Limonero dairy cattle. The population of Criollo Limonero breed tended to increase over time, with the exception of the period 1994–2003. The implications of the biodiversity-derived analysis in Criollo Limonero cattle lie in the development of strategies to avoid increasing inbreeding by identifying animals with a higher genetic conservation index for multiplication through the family-related breeding programme. Moreover, at the producer level, the biggest advantage is that the breeders have a list of animals with information about inbreeding values, which allows them to improve reproductive matings avoiding potential errors. Other Centre and South American creole cattle breeds, such as the San Martinero (De León et al., 2021) and Blanco Orejinegro (Ocampo Gallego et al., 2020) have also had an increase in their population during the last few years due to the fact that they have been maintained in specific research centres in their corresponding countries. On the other hand, other endangered breeds such as the Negra Andaluza (Cartuche Macas et al., 2024) and the Berrenda Negra y Colorada (González-Cano et al., 2022) have reduced their population until now. This effect could be due to the fact that in Centre and South America, the great majority of creole breeds have been maintained in public research centres that play an important role in the conservation of these genetic resources. On the other hand, in European countries, producers have been economically encouraged through projects for the conservation of native breeds, research and development of value-added products, together with breeders’ associations. Despite these efforts, the tendency to decrease the population of animals belonging to endangered breeds continues. In the present study, during the period 1994–2003 there was a drastic fall in the number of individuals, possibly due to various factors among which it can be mentioned the severe drought produced by El Niño phenomenon (1991–1992) that lasted 14 months and severely affected some Caribbean countries, including Venezuela (Orlando Olivares and Zingaretti, 2018). Since 2016, the population of Criollo Limonero dairy cattle has been affected by the factors described above, which have caused the longest-lived population to decline. Between 2019–2021, the population of first calving females was almost non-existent due to the fact that these animals were mobilised to other research centres. This fact had as a counterpart the recovery of the number of individuals during the last two years. In this way Florio Luis (2011) and Ordóñez Vela and Plasse (2020) established some critical points to take into account for the preservation of this breed, such as the reduction of purebred animals in recent years due to the lack of economic resources, unfavourable agro-climatic conditions, great inbreeding rates, introduction of different commercial genetics, reproductive and sanitary problems, reduction of breeding dams, lack of organisation of producers, lack of centralisation of genealogical registers, limited seminal supply of sires with little genetic variability and lack of a complete census of registration data. In addition, in recent years, the problems have continued, resulting in a reduction of animals due to transfers to other farms, to the point that the research centre only kept about 30 animals in 2018, which are currently being reproduced through a breeding and selection scheme (Çelik, 2024; Gutiérrez-Reinoso et al., 2023). The fact of centralising the conservation of this breed at “Hacienda Laral” (Venezuela) has allowed the breed to be maintained over time, because if the traditional management of the breed continues, it could lead to an accelerated loss of individuals.
On the other hand, it was observed that the pedigree completeness of this breed was similar to other autochthonous breeds, reaching values greater than 80% in the last two periods (1–3 generations), which guarantees reliability in the estimators of genetic diversity from the pedigree (Maccluer et al., 1983). This is important due to although in the first two periods the values were below 80%, it is worth mentioning that lower values of pedigree integrity were considered normal in populations at the beginning of their pedigree formation. Moreover, the estimated equivalent generations were 4.28–4.58, a lower value compared to other specialised breeds, such as the Brazilian Nelore (Panetto et al., 2010), although higher than the Berrenda Negra y Berrenda Colorada (González-Cano et al., 2022) and Negra Andaluza (Cartuche Macas et al., 2024), both Spanish breeds.
The GI for all periods was estimated to be between 4.58 and 8.03 years, being greater than other Centre and South American native breeds. In particular, the highest GI value was obtained in the last period studied possibly due to the fact that between 2019–2021 there were no first calvings, showing only the calvings corresponding to the second calving onwards (2019–2021) and from the third calving in the year 2021. On the other hand, in the year 2022, the second and third calvings were not shown and, finally, in the year 2023 there were no data for third and fourth calvings. These effects were possibly due to external factors such as those detailed by Florio Luis (2011) and Ordóñez Vela and Plasse (2020). The sire-calf ratio ranged from 3.51 to 7.7 years and in all periods a gradual increase over time was observed. This value was much higher than other breeds such as Blanca Ojinegra (3.72 years), Romosinuano (4.76 years), San Martinero (4.57 years) and Negra Andaluza (5.91 years). In addition, in the last period a notable increase was observed, while in the period 2004–2013 the same increase was observed due to the use of sires aged between 7–19 years and in particular in the years 2009–2010 sires aged between 16–18 years were used. This effect was similar in the creole breed Reyna de Nicaragua where sires between 10–19 years of age were used (Corrales, 2011). In the sire-daughter path, a decrease was observed since 2007, indicating that sires are being replaced by their daughters at an earlier age, i.e. daughters of breeding sires were entering the reproductive phase faster, which may generate a higher rate of genetic progress, similar to the sire–son path, which has shown a decrease since 2015. In the dam–son and dam–daughter pathways, a gradual increase has been observed as the older breeders are the ones that have provided more offspring, i.e. they are being used for longer before being replaced by their offspring, particularly in the last period.
The historical inbreeding estimated using the Meuwiwn algorithm and the Recursive method was 1.03% and 2.25%, respectively; however, in the last period it was 2.05% and 4.45%, respectively. This difference could be due to the fact that in the last period there were more sires/dams with one or two unknown parents compared to the total population, which made the F value fit larger. On the other hand, the ΔF value was 0.38%, a value slightly lower than the Blanca Ojinegra (F=2.88%; ΔF=0.40%), Romosinuano (F=2.53%; ΔF=0.42%) Colombian breeds (Ocampo Gallego et al., 2020; Cañas–Álvarez et al., 2023). Reyna breed from Nicaragua (F=2.40%) (Corrales et al., 2010) and the Creole Tropical Dairy Breed from Mexico (F=2.14%; ΔF=0.0.77%) (Rosendo Ponce et al., 2018). These low values could be due to the management of circular mating under families that has been carried out and that has avoided a greater increase. On the other hand, this inbreeding was lower than native breeds especially from Europe such as Berrenda Negra (F=7%; ΔF=0.95%) (González-Cano et al., 2022), Berrenda Colorada (F=5.70%; ΔF=0.40%) and Negra Andaluza (F=8.82%; ΔF=0.95%) (Cartuche Macas et al., 2024) from Spain and the Reggiana breed (F=7.75%; ΔF=%) from Italy (Schiavo et al., 2022).
Although in recent years strategies have been developed to encourage the consumption of products derived from these breeds, such as those derived from the Reggiana and Modonese breeds (which are currently used to produce Parmigiano-Reggiano cheese with Protected Designation of Origin), the number of individuals in these breeds has grown considerably (Schiavo et al., 2022). Another case is the Negra Andaluza breed, which is used for the production of 100% ‘native breed’ meat (both fresh meat and other meat-derived products) which has made populations improve in terms of census and genetic diversity (Cartuche Macas et al., 2024). In the case of the Criollo Limonero dairy breed, some strategies for the conservation of the breed have also been developed (Florio Luis, 2011; Ordóñez Vela and Plasse, 2020; Araujo-Febres, 2023). In this case, this breed possesses adaptive traits of commercial interest such as those related to heat tolerance (SLICK gene) as well as resistance to parasites and diseases common to tropical climates, which could justify its conservation. As a recommendation, ΔF values per generation should be below 1.0% (Boettcher et al., 2013; Tenhunen et al., 2024). In the present study, the AR value in the last period was 3.10% despite the slight reduction compared to the previous period. This indicates that this population should be carefully monitored to avoid long-term negative effects.
On the other hand, the α value in the last two periods was positive (α=0.0051) indicating a small tendency towards inbreeding (mating between genetically related individuals) given the mating between genetically similar individuals, as opposed to outbreeding (mating between unrelated individuals). Also, possible causes related to this fact could be the decrease in population size or the excessive use of certain breeders. An example of this were the 29 breeding males that had between 40 and 130 calves and the 14 breeding females that had between 10 and 14 calves. The main effect of reducing the number of males and increasing the number of offspring per male is that inbreeding potentially increases in subsequent generations with consequences on related parameters such as α. This effect can be countered by using various strategies such as the use of molecular technologies to establish more accurate parentage relationships and development of mates to reduce inbreeding potencies, as well as the use of parentage and paternity testing to reduce potential parental assignment errors. According to Groeneveld et al. (2009) the structure of the evaluated Criollo Limonero population or the geographical isolation for the conservation of the single genetic nucleus could have an influence on these results. This effect of the positive α value was similar to the endangered Negra Andaluza breed (0.061) (Cartuche Macas et al., 2024). Similar occurs in other specialised breeds which are managed through selection programmes such as Holstein and Jersey in Canada (Stachowicz et al., 2011; Gutierrez-Reinoso et al., 2021). In contrast, autochthonous and specialised breeds have differential α values (Márquez et al., 2010; Melka et al., 2013; Piccoli et al., 2014; Cavani et al., 2018; Amaya et al., 2020; de Araujo Neto et al., 2020; Wirth et al., 2021; Ablondi et al., 2022).
The estimated Ne for the last period was around 100 individuals with both methods used, although recommendations indicate that a population should have a minimum inbreeding rate of less than 1% per generation and a Ne value of 50 to avoid negative effects due to inbreeding, and above all to maintain genetic diversity at sustainable levels over time (Boettcher et al., 2013; Tenhunen et al., 2024). This value, although quite low, was similar to other creole and autochthonous breeds under similar management conditions (Ocampo Gallego et al., 2020; De León et al., 2021).
The gene origin probability analysis allows to evaluate the changes that occurred in the population in a short period of time (Sakthivel et al., 2018). This breed had an effective number of founders (fe) of 50.85, effective number of ancestors (fa) of 25, and equivalent founder genomes (fg) of 11.60. These values were lower than the Vrindavani breed which had fa=115 and fe=78 (Gangwar et al., 2024). In this breed a reduction of ancestry was observed in the last two periods, to 4 and 9 ancestors, respectively, explaining 25 and 50% of the genetic diversity. This would indicate a strong founder effect or an intense selection of certain breeders that could affect the increase of inbreeding within the population.
The value of the fe/fa ratio was >2 in the last three periods, which clearly demonstrated the existence of bottlenecks within the pedigree due to the fact that in recent years there have been massive departures of animals resulting in few founders being selected as breeders for the next generations. Another reason could be the environmental effects suffered, such as the droughts that have affected this population constantly. Similar values have been observed in northern European native breeds such as Red Dairy Cattle, Swedish Polled and Vorderwälder which showed values between 2.15–3.53 indicating bottlenecks due to inbreeding increases in the 1970s (Nyman et al., 2022). On the other hand, this ratio was higher than that obtained in the Brahman breed from Brazil (fe/fa=1–1.01) (Cavani et al., 2018) and in the high-milch Vrindavani breed from India with 1.20 which indicated possible bottleneck formations (Gangwar et al., 2024).
The value of fg/fe ratio was between 0.23–0.27 which indicates a strong impact of genetic drift, marking a reduction of genetic diversity due to random events (droughts, floods, etc.), as well as generating a higher risk of increased inbreeding rates and consequently producing inbreeding depression. This value was lower than that obtained in the Sahiwal breed in Kenya, which showed a fa/fe ratio of 0.63, indicating that there was a greater loss of genetic diversity due to genetic drift (Mwangi et al., 2016) or in the Brahman breed in Brazil (0.83) (Cavani et al., 2018) and in the Lidia breed in Spain (Cortés et al., 2014) in which the bottleneck effect was similar to that of genetic drift. Overall, given the difference between fg/fe and fa/fe it is observed that the main cause of the reduction in genetic diversity is not only the unequal contribution of the founders, but also due to genetic drift that has been accumulating even more loss of variability in successive generations. This leads to the recommendation that strategies should be implemented to mitigate this effect, such as the introduction of new sires or the control of unequal use of existing sires, as well as to better prepare to mitigate the agro-climatic effects and political decisions that have affected this population of Criollo Limonero dairy cattle and put it at serious risk of disappearing. One of the most important aspects of the conservation of creole breeds is their adaptability to changing environmental conditions (Boettcher et al., 2015), such as high temperatures, drought, and disease-transmitting parasites. This adaptability and resistance is provided by genes such as, for example, the SLICK gene for tropical zones in creole cattle breeds including the Criollo Limonero breed (Gebeyehu et al., 2024; Hansen, 2020; Pitt et al., 2019; Porto-Neto et al., 2018) or tick resistance in the Argentine creole breed (Ortega et al., 2023).
When analysing the genetic diversity loss, and given the effect of fa, fe and ge, it was ratified that the greatest losses have been generated in the last periods. The unequal contribution of the founders (GD loss = 4.31%) and the genetic drift accumulated over the non-founder generations (GD* − GD = 3.33%) were part of this effect. These values were extremely higher than the zebu breeds from Brazil (Santana et al., 2016), Ecuadorian Monbeliarde and Charolais (Cartuche-Macas et al., 2025a, 2024), Negra Andaluza (Cartuche Macas et al., 2024) and Berrenda from Spain (González-Cano et al., 2022).
The effective population size which has been calculated by the individual inbreeding rate was 131.5 and by the individual coancestry rate 16.13 in the last two generations, being different values compared to the Berrenda Negra and Colorada Spanish breeds (Berrenda Negra = 9.93 and 92.28, respectively; and Berrenda Colorada = 11.58 and 169.92, respectively) (González-Cano et al., 2022). The remarkable difference between coancestry-based Ne (16.13) and inbreeding-based Ne (131.5) suggests that, although the inbreeding rate has remained relatively low in the last generations, the number of effective ancestors contributing genetically to the population is small, which may be related to an unbalanced use of sires/dams and a closed population structure. The use of genealogical databases to track sire and dam contributions and the introduction of outcrossing strategies could be explored in the future as specific approaches to mitigate inbreeding and improve genetic diversity.
5 Conclusion
In summary, the Criollo Limonero dairy breed has suffered an accelerated genetic diversity loss mainly due to the bottleneck effect and genetic drift. Therefore, there is an urgent need to consider the implementation of breeding and selection strategies for this breed in order to mitigate the adverse effects of the genetic diversity loss. In particular, a more rigorous management of the selection and use of sire and dam breeding stock should be taken into account, taking into account the inbreeding values or genetic conservation index. Finally, the Ne values indicate that there is a moderate risk of increased inbreeding, suggesting that the population could be genetically sustainable in the medium to long term if appropriate selection and breeding schemes are carried out in order to conserve the Criollo Limonero dairy cattle breed, which is currently still in a critically endangered status.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding authors.
Ethics statement
The requirement of ethical approval was waived by ethics committee/institutional review board for the studies involving animals because the authors used a database of death animals. The studies were conducted in accordance with the local legislation and institutional requirements.
Author contributions
LC-M: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Methodology, Writing – original draft, Writing – review & editing. JP-G: Conceptualization, Data curation, Formal Analysis, Methodology, Writing – review & editing. MG-R: Funding acquisition, Methodology, Writing – review & editing. AL-G: Conceptualization, Data curation, Methodology, Writing – review & editing. MG-H: Conceptualization, Formal Analysis, Funding acquisition, Methodology, Supervision, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that no financial support was received for the research and/or publication of this article.
Acknowledgments
The authors would like to thank the staff involved in the collection of information for the elaboration of the different databases, as well as the stall for the excellent technical assistance.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Ablondi M., Sabbioni A., Stocco G., Cipolat-Gotet C., Dadousis C., Kaam J. T., et al. (2022). Genetic diversity in the italian holstein dairy cattle based on pedigree and SNP data prior and after genomic selection. Front. Vet. Sci. 8. doi: 10.3389/FVETS.2021.773985/BIBTEX
Aguilar I. and Misztal I. (2008). Technical note: recursive algorithm for inbreeding coefficients assuming nonzero inbreeding of unknown parents. J. Dairy Sci. 91, 1669–1672. doi: 10.3168/JDS.2007-0575
Alderson G., Alderson L., and Bodó I. (1992). A system to maximize the maintenance of genetic variability in small populations. Biol. (Basel).
Amaya A., Martínez R., and Cerón-Muñoz M. (2020). Population structure and genetic diversity in Colombian Simmental cattle. Trop. Anim. Health Prod 52, 1133–1139. doi: 10.1007/S11250-019-02111-W
Araujo-Febres O. (2023). Propuesta de rescate de la raza Criollo Limonero de Venezuela. Archivos Latinoamericanos Producción Anim. 31, 363–367. doi: 10.53588/ALPA.310561
Boettcher P. J., Hoffmann I., Baumung R., Drucker A. G., McManus C., Berg P., et al. (2015). Genetic resources and genomics for adaptation of livestock to climate change. Front. Genet. doi: 10.3389/fgene.2014.00461
Boettcher P., Martin J. F., Gandini G., Joshi B. K., and Oldenbroek J. K. (2013). In vivo conservation of animal genetic resources. FAO (FAO Anim. production Health guidelines 14). 5, 461.
Boichard D., Maignel L., and Verrier É. (1997). The value of using probabilities of gene origin to measure genetic variability in a population. Genet. Selection Evol. 29, 1–19. doi: 10.1186/1297-9686-29-1-5
Caballero A. and Toro M. A. (2000). Interrelations between effective population size and other pedigree tools for the management of conserved populations. Genet. Res. 75, 331–343. doi: 10.1017/S0016672399004449
Cabezas C. R., Capote C. B., González M artínez A., Falquez O. C., Jurado J. M. L., Manuel J., et al. (2019). Biometric study of Criollo Santa Elena Peninsula cattle (Ecuador). Rev. Mexicana Ciencia Pecuaria 10, 819–836. doi: 10.22319/rmcp.v10i4.4850
Cañas–Álvarez J. J., Ossa-Saraz G. A., Garcés-Blanquiceth J. L., and Burgos–Paz W. O. (2023). Genealogical structure of the Colombian Romosinuano Creole cattle. Trop. Anim. Health Prod 55, 1–8. doi: 10.1007/S11250-023-03694-1/3
Cartuche Macas L. F., Camacho Vallejo M. E., González Ariza A., León Jurado J. M., Delgado Bermejo J. V., Marín Navas C., et al. (2024). Analysis of endangered andalusian black cattle (Negra andaluza) reveals genetic reservoir for bovine black trunk. Animals 14, 1131. doi: 10.3390/ANI14071131/S1
Cartuche-Macas L. F., Guaman Ilvay O. J., Chacón E., Gutierrez-Reinoso M. A., and Garcia-Herreros M. (2025a). Reproductive performance, inbreeding, and genetic diversity in montbeliarde dairy cattle obtained by absorption crossing. Animals 15, 322. doi: 10.3390/ANI15030322
Cartuche-Macas L. F., Gutierrez-Reinoso M. A., Chacón E., Larrea-Izurieta C. O., García-Flores J. M., and Garcia-Herreros M. (2025b). Ecuadorian Holstein-Friesian cattle paternal lineages: Demographic structure, inbreeding evolution, and genetic diversity. PloS One 20, e0318730. doi: 10.1371/JOURNAL.PONE.0318730
Cartuche-Macas L. F., Lozada E. F., Gutiérrez-Reinoso M. A., Chacón E., Navas F. J., and García-Herreros M. (2024). Evolution of population structure, reproductive performance, inbreeding, and genetic diversity in Ecuadorian charolais cattle. Veterinary Sci. 11, 566. doi: 10.3390/VETSCI11110566
Casanovas Arias D., León Jurado J. M., Bermejo Asensio L. A., Navas González F. J., Marín Navas C., and Barba Capote C. J. (2020). Genetic diversity evolution of a sheep breed reintroduced after extinction: Tracing back Christopher Columbus’ first imported sheep. Res. Vet. Sci. 132, 207–216. doi: 10.1016/J.RVSC.2020.06.007
Cavani L., Silva R. M., de O., Carreño L. O. D., Ono R. K., Bertipaglia T. S., et al. (2018). Genetic diversity of Brazilian Brahman cattle by pedigree analysis. Pesqui Agropecu Bras. 53, 74–79. doi: 10.1590/S0100-204X2018000100008
Çelik Ş. (2024). Bibliometric analysis of genomic selection in breeding of animal from 1993 to 2024: global trends and advancements. Front. Genet. 15. doi: 10.3389/FGENE.2024.1402140/BIBTEX
Cervantes I., Goyache F., Molina A., Valera M., and Gutiérrez J. P. (2011). Estimation of effective population size from the rate of coancestry in pedigreed populations. J. Anim. Breed Genet. 128, 56–63. doi: 10.1111/J.1439-0388.2010.00881.X
Cockerham C. C. (1970). Avoidance and rate of inbreeding. Math. Topics Population Genet., 104–127. doi: 10.1007/978-3-642-46244-3_4
Colleau J. J. and Sargolzaei M. (2008). A proximal decomposition of inbreeding, coancestry and contributions. Genet. Res. (Camb) 90, 191–198. doi: 10.1017/S0016672307009202
Contreras G., Chirinos Z., Molero E., and Paéz A. (2012). Medidas corporales e índices zoométricos de toros Criollo Limonero de Venezuela, Vol. 30. 175–181.
Corrales R. (2011). Population structure and phenotypic characterization as a basis for conservation and sustainable use of reyna creole cattle in Nicaragua. Agric. Food Sciences Environ. Science. 47.
Corrales R., Näsholm A., Malmfors B., and Philipsson J. (2010). Population structure of Reyna Creole cattle in Nicaragua. Trop. Anim. Health Prod 42, 1427–1434. doi: 10.1007/S11250-010-9571-9
Cortés O., Sevane N., Baro J. A., and Cañón J. (2014). Pedigree analysis of a highly fragmented population, the Lidia cattle breed. Livest Sci. 167, 1–8. doi: 10.1016/J.LIVSCI.2014.05.011
de Araujo Neto F. R., Vieira D. A., Santos D. J., de A., Pessoa M. C., Borquis R. R. A., et al. (2020). Population structure of Simmental beef cattle using pedigree analysis. Trop. Anim. Health Prod 52, 1513–1517. doi: 10.1007/S11250-019-02102-X
De León C., Martínez R., and Rocha J. F. (2021). Genealogical and genomic heritability for adaptability and reproductive traits in the Colombian creole cattle breeds blanco orejinegro and sanmartinero. Genet. Mol. Res. 20 (3), gmr18878. doi: 10.4238/GMR18878
Florio Luis J.C.G.Z. (2011). Criollo limonero’s national preservation and animal breeding program at Venezuela bolivarian republic. Actas Iberoamericanas Conservación Anim., 117–122.
Gangwar M., Ahmad S. F., Gaur G. K., Tamilarasan K., Vyas J., and XXXD.A P. (2024). Pedigree-based analysis of population structure and genetic diversity in high-milch Vrindavani crossbred cattle of India. Trop. Anim. Health Prod 56 (8), 280. doi: 10.1007/S11250-024-04179-5
Gebeyehu S., Heins B., Sonstegard T., Sölkner J., Mészáros G., Traoré A., et al. (2024). Analysis of SLICK allele in African taurine and Zebu cattle breeds. Anim. Genet. 56 (1), e13499. doi: 10.1111/age.13499
Gómez-Romano F., Villanueva B., Sölkner J., de Cara M. A. R., Mészáros G., Pérez O’Brien A. M., et al. (2016). The use of coancestry based on shared segments for maintaining genetic diversity. J. Anim. Breed. Genet. 133, 357–365. doi: 10.1111/JBG.12213
González-Cano R., González-Martínez A., Muñoz-Mejías M. E., Valera P., and Rodero E. (2022). Analyses of genetic diversity in the endangered"Berrenda" Spanish cattle breeds using pedigree data. Animals 12, 249. doi: 10.3390/ANI12030249/S1
Groeneveld E., Westhuizen B. D., Maiwashe A., Voordewind F., and Ferraz J. B. (2009). POPREP: a generic report for population management. Genet. Mol. Res. 8, 1158–1178. doi: 10.4238/VOL8-3GMR648
Gutiérrez J. P., Cervantes I., Molina A., Valera M., and Goyache F. (2008). Individual increase in inbreeding allows estimating effective sizes from pedigrees. Genet. Selection Evol. 40, 359–378. doi: 10.1051/GSE:2008008
Gutiérrez J. P. and Goyache F. (2005). A note on ENDOG: A computer program for analysing pedigree information. J. Anim. Breed. Genet. 122, 172–176. doi: 10.1111/J.1439-0388.2005.00512.X
Gutiérrez-Reinoso M. A., Aponte P. M., Cabezas J., Rodriguez-Alvarez L., and Garcia-Herreros M. (2020). Genomic evaluation of primiparous high-producing dairy cows: inbreeding effects on genotypic and phenotypic production–reproductive traits. Animals 10, 1704. doi: 10.3390/ANI10091704
Gutierrez-Reinoso M. A., Aponte P. M., and Garcia-Herreros M. (2021). Genomic analysis, progress and future perspectives in dairy cattle selection: A review. Anim. (Basel) 11, 599. doi: 10.3390/ANI11030599
Gutiérrez-Reinoso M. A., Aponte P. M., and García-Herreros M. (2022). A review of inbreeding depression in dairy cattle: current status, emerging control strategies, and future prospects. J. Dairy Res. 89, 3–12. doi: 10.1017/S0022029922000188
Gutiérrez-Reinoso M. A., Aponte P. M., and García-Herreros M. (2023). Genomic and phenotypic udder evaluation for dairy cattle selection: A review. Animals 13, 1588. doi: 10.3390/ANI13101588
Hansen P. J. (2020). Prospects for gene introgression or gene editing as a strategy for reduction of the impact of heat stress on production and reproduction in cattle. Theriogenology 154, 190–202. doi: 10.1016/j.theriogenology.2020.05.010
Hill W. G. (1981). Estimation of effective population size from data on linkage disequilibrium. Genetical Research. 38 (3), 209–216. doi: 10.1017/S0016672300020553
James J. W. (1972). Computation of genetic contributions from pedigrees. Theor. Appl. Genet. 42, 272–273. doi: 10.1007/BF00277555
Kimura M. and Crow J. F. (1963). On the maximum avoidance of inbreeding*. Genet. Res. (Camb) 4, 399–415. doi: 10.1017/S0016672300003797
Lacy R. C. (1989). Analysis of founder representation in pedigrees: Founder equivalents and founder genome equivalents. Zoo Biol. 8, 111–123. doi: 10.1002/ZOO.1430080203
Landaeta-Hernández A., Zambrano-Nava S., Hernández-Fonseca J. P., Godoy R., Calles M., Iragorri J. L., et al. (2011). Variability of hair coat and skin traits as related to adaptation in Criollo Limonero cattle. Trop. Anim. Health Prod 43, 657–663. doi: 10.1007/S11250-010-9749-1
Maccluer J. W., Boyce A. J., Dyke B., Weitkamp L. R., Pfenning D. W., and Parsons C. J. (1983). Inbreeding and pedigree structure in standardbred horses. J. Heredity 74, 394–399. doi: 10.1093/OXFORDJOURNALS.JHERED.A109824
Mandal A., Baneh H., Subramanyam B. V., and Notter D. R. (2020). Genetic variability and population structure based on pedigree information for Muzaffarnagari sheep in India. Small Ruminant Res. 191, 106182. doi: 10.1016/J.SMALLRUMRES.2020.106182
Marcuzzi O., Cecco P.Á., Olivera L. H., Pereira Rico J. A., Calcaterra F., Vega A. L., et al. (2025). Divergent adaptation to highland and tropical environments in Bolivian Creole cattle. Gene 949, 149354. doi: 10.1016/J.GENE.2025.149354
Márquez G. C., Speidel S. E., Enns R. M., and Garrick D. J. (2010). Genetic diversity and population structure of American Red Angus cattle. J. Anim. Sci. 88, 59–68. doi: 10.2527/JAS.2008-1292
Martinez R., Bejarano D., Ramírez J., Ocampo R., Polanco N., Perez J. E., et al. (2023). Genomic variability and population structure of six Colombian cattle breeds. Trop. Anim. Health Prod 55, 1–8. doi: 10.1007/S11250-023-03574-8/FIGURES/4
Martínez A. M., Gama L. T., Cañón J., Ginja C., Delgado J. V., Dunner S., et al. (2012). Genetic footprints of Iberian cattle in America 500 years after the arrival of Columbus. PloS One 7, e49066–e49066. doi: 10.1371/JOURNAL.PONE.0049066
Martinez R. A., Garcia D., Gallego J. L., Onofre G., Perez J., and Canon J. (2008). Genetic variability in Colombian Creole cattle populations estimated by pedigree information [Article. J. Anim. Sci. 86, 545–552. doi: 10.2527/jas.2007-0175
Melka M. G., Sargolzaei M., Miglior F., and Schenkel F. (2013). Genetic diversity of Guernsey population using pedigree data and gene-dropping simulations. Animal 7, 192–201. doi: 10.1017/S1751731112001723
Meuwissen T. H. E. and Luo Z. (1992). Computing inbreeding coefficients in large populations. Genet. Sel Evol. 24, 305. doi: 10.1186/1297-9686-24-4-305
Mignon-Grasteau S., Boissy A., Bouix J., Faure J. M., Fisher A. D., Hinch G. N., et al. (2005). Genetics of adaptation and domestication in livestock. Livest Prod Sci. 93, 3–14. doi: 10.1016/J.LIVPRODSCI.2004.11.001
Mwangi S., Muasya T. K., Ilatsia E. D., and Kahi A. K. (2016). Assessment of the genetic variability using pedigree analysis of the Sahiwal breed in Kenya. Anim. Genet. Resources/Ressources génétiques animales/Recursos genéticos animales 59, 7–14. doi: 10.1017/S2078633616000199
Navas F. J., Jordana J., León J. M., Barba C., and Delgado J. V. (2017). A model to infer the demographic structure evolution of endangered donkey populations. Animal 11, 2129–2138. doi: 10.1017/S1751731117000969
Nietlisbach P., Keller L. F., Camenisch G., Guillaume F., Arcese P., Reid J. M., et al. (2017). Pedigree-based inbreeding coefficient explains more variation in fitness than heterozygosity at 160 microsatellites in a wild bird population. Proc. R. Soc. B: Biol. Sci. 284 (1850), 20162763. doi: 10.1098/RSPB.2016.2763
Nomura T. and Yonezawa K. (1996). A comparison of four systems of group mating for avoiding inbreeding. Genet. Selection Evol. 1996 28:2 28, 1–19. doi: 10.1186/1297-9686-28-2-141
Nyman S., Johansson A. M., Palucci V., Schönherz A. A., Guldbrandtsen B., Hinrichs D., et al. (2022). Inbreeding and pedigree analysis of the European red dairy cattle. Genet. Sel Evol. 54 (1), 70. doi: 10.1186/S12711-022-00761-3
Ocampo Gallego R., Ramírez Toro J., Lopera Peña J., Restrepo Castañeda G., Gallego Gil J., Ocampo Gallego R., et al. (2020). Genetic diversity assessed by pedigree analysis in the blanco orejinegro (bon) cattle breed population from the Colombian germplasm bank. Chilean J. Agric. Anim. Sci. 36, 69–77. doi: 10.29393/CHJAAS36-4D30004
Ordóñez Vela J. A. and Plasse D. (2020). Zoometría de dos poblaciones de ganado Criollo de Venezuela. Archivos Latinoamericanos Producción Anim. 28, 91–101. doi: 10.53588/ALPA283404
Orlando Olivares B. and Zingaretti M. L. (2018). Análisis de la sequía meteorológica en cuatro localidades agrícolas de Venezuela mediante la combinación de métodos multivariados. UNED Res. J. 10 (1), 192–203. doi: 10.22458/URJ.V10I1.2026
Ortega M. F., Giovambattista G., Cutullé C., Santos D. D., Nava S., Bonamy M., et al. (2023). Phenotypic evaluation of genetic resistance to the tick Rhipicephalus (Boophilus) microplus in Argentine Creole cattle. Ticks Tick-borne Dis. 14, 102223. doi: 10.1016/j.ttbdis.2023.102223
Panetto J. C. C., Gutiérrez J. P., Ferraz J. B. S., Cunha D. G., and Golden B. L. (2010). Assessment of inbreeding depression in a Guzerat dairy herd: Effects of individual increase in inbreeding coefficients on production and reproduction. J. Dairy Sci. 93, 4902–4912. doi: 10.3168/JDS.2010-3197
Pezzini T., Mariante A. S., Martins E., Paiva S., Seixas L., Costa J. B. G., et al. (2018). Population structure of Brazilian Crioula Lageana cattle (Bos taurus) breed. Rev. Colombiana Cienc. Pecuarias 31, 93–102. doi: 10.17533/udea.rccp.v31n2a02
Piccoli M. L., Braccini Neto J., Brito F. V., Campos L. T., Bértoli C. D., Campos G. S., et al. (2014). Origins and genetic diversity of British cattle breeds in Brazil assessed by pedigree analyses. J. Anim. Sci. 92, 1920–1930. doi: 10.2527/JAS.2013-7283
Pitt D., Bruford M. W., Barbato M., Orozco-terWengel P., Martínez R., and Sevane N. (2019). Demography and rapid local adaptation shape Creole cattle genome diversity in the tropics. Evolutionary Appl. 12, 105–122. doi: 10.1111/eva.12641
Porto-Neto L. R., Bickhart D. M., Landaeta-Hernandez A. J., Utsunomiya Y. T., Pagan M., Jimenez E., et al. (2018). Convergent evolution of slick coat in cattle through truncation mutations in the prolactin receptor. Front. Genet. 9. doi: 10.3389/fgene.2018.00057
Ramírez-Valverde R., Delgadillo-Zapata A. R., Domínguez-Viveros J., Hidalgo-Moreno J. A., Núñez-Domínguez R., Rodríguez-Almeida F. A., et al. (2018). Análisis de pedigrí en la determinación de la diversidad genética de poblaciones bovinas para carne mexicanas. Rev. Mex Cienc Pecu 9, 615–635. doi: 10.22319/RMCP.V9I4.4654
Rojas-Espinoza R., Macedo R., Suaña A., Delgado A., Manrique Y. P., Rodríguez H., et al. (2023). Phenotypic characterization of creole cattle in the andean highlands using bio-morphometric measures and zoometric indices. Animals 13, 1843. doi: 10.3390/ANI13111843/S1
Rosendo Ponce A., Palacios Jiménez A. L., Rosales Martínez F., Torres Hernández G., Ramírez Valverde R., and Becerril Pérez C. M. (2018). Genetic variability of Tropical Milking Criollo cattle of Mexico estimated from genealogy information. Rev. Colombiana Cienc. Pecuarias 31, 196–203. doi: 10.17533/UDEA.RCCP.V31N3A04
Sakthivel M., Balasubramanyam D., Kumarasamy P., Raja A., Anilkumar R., Gopi H., et al. (2018). Genetic structure of a small closed population of the New Zealand white rabbit through pedigree analyses. World Rabbit Sci. 26, 101–112. doi: 10.4995/WRS.2018.7426
Santana M. L., Pereira R. J., Bignardi A. B., Ayres D. R., Menezes G. R. O., Silva L. O. C., et al. (2016). Structure and genetic diversity of Brazilian Zebu cattle breeds assessed by pedigree analysis. Livest Sci. 187, 6–15. doi: 10.1016/J.LIVSCI.2016.02.002
Sargolzaei M., Iwaisaki H., and Colleau J. J. J. J. (2006). CFC: a tool for monitoring genetic diversity. INRAE - Institut Natl. Recherche pour l’agriculture l’alimentation l’environnement. 13:27–28. doi: 10.34894/VQ1DJA
Schiavo G., Bovo S., Ribani A., Moscatelli G., Bonacini M., Prandi M., et al. (2022). Comparative analysis of inbreeding parameters and runs of homozygosity islands in 2 Italian autochthonous cattle breeds mainly raised in the Parmigiano-Reggiano cheese production region. J. Dairy Sci. 105, 2408–2425. doi: 10.3168/JDS.2021-20915
Sheppard P. M. and Wright S. (1971). Evolution and the genetics of populations. Vol. 2. The theory of gene frequencies. J. Anim. Ecol. 40, 266. doi: 10.2307/3353
Stachowicz K., Sargolzaei M., Miglior F., and Schenkel F. S. (2011). Rates of inbreeding and genetic diversity in Canadian Holstein and Jersey cattle. J. Dairy Sci. 94, 5160–5175. doi: 10.3168/JDS.2010-3308
Tenhunen S., Thomasen J. R., Sørensen L. P., Berg P., and Kargo M. (2024). Genomic analysis of inbreeding and coancestry in Nordic Jersey and Holstein dairy cattle populations. J. Dairy Sci. 107, 5897–5912. doi: 10.3168/JDS.2023-24553
Tullo E. (2011). Selection methodology for a dairy breed in a tropical environment. Università degli studi di Milano (Italy). Available at: https://air.unimi.it/retrieve/handle/2434/170624/170260/phd_unimi_R08072.pdf#page=16 (Accessed January 26, 2025).
Vassallo J. M., Diaz C., and Garcia-Medina J. R. (1986). A note on the population structure of the Avileña breed of cattle in Spain. Livest Prod Sci. 15, 285–288. doi: 10.1016/0301-6226(86)90035-7
Villasmil-Ontiveros Y., Román Bravo R., Yáñez-Cuéllar L., Contreras G., Jordana J., and Aranguren-Méndez J. (2008). Genetic diversity of limonero creole breed using microsatellites molecular markers, Vol. XVIII. 415–423.
Viveros J., Reyes-Cerón A., Saiz-Pineda J., Villegas-Gutiérrez C., Aguilar-Palma G., and Rodríguez-Almeida F. (2021). Structure and genetic variability of the Mexican Sardo Negro breed Análise da estrutura e variabilidade genética da raça Sardo Negro do México. Ciec. Rural 52 (5), e20210116. doi: 10.1590/0103-8478cr20210116
Wirth A., Duda J., Distl O., Wirth A. , Duda J. , and Distl O. (2021). Genetic diversity and the impact of the breed proportions of US brown swiss in german brown cattle. Animals 11, 152. doi: 10.3390/ANI11010152
Yenen V. (2007). Caracterización Genética de Bovinos de la Raza Criollo Limonero Utilizando Marcadores Moleculares de ADN del Tipo Microsatélites. Available online at: https://www.researchgate.net/publication/262487934_caracterizacion_genetica_de_bovinos_de_la_raza_criollo_limonero_utilizando_marcadores_moleculares_de_adn_del_tipo_microsatelites (Accessed February 28, 2025).
Keywords: consanguinity, reproductive traits, diversity loss, Criollo Limonero, dairy cattle
Citation: Cartuche-Macas LF, Pérez-González JR, Gutiérrez-Reinoso MA, Leal-González AG and García-Herreros M (2025) Inbreeding, reproductive performance, and genetic diversity in a tropically adapted dairy breed: the endangered Criollo Limonero cattle. Front. Anim. Sci. 6:1590048. doi: 10.3389/fanim.2025.1590048
Received: 08 March 2025; Accepted: 15 May 2025;
Published: 02 June 2025.
Edited by:
Titus Zindove, Lincoln University, New ZealandReviewed by:
Hasitha Priyashantha, Swedish University of Agricultural Sciences, SwedenObert Tada, University of Limpopo, South Africa
Nelisiwe Mkize, Stellenbosch University, South Africa
Copyright © 2025 Cartuche-Macas, Pérez-González, Gutiérrez-Reinoso, Leal-González and García-Herreros. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Luis F. Cartuche-Macas, bC5jYXJ0dWNoZW1AZ21haWwuY29t; Manuel García-Herreros, aGVycmVyb3NnbUBnbWFpbC5jb20=