 Megan Woodrum Setser
Megan Woodrum Setser Daniel Brown1
Daniel Brown1 Joao HC Costa
Joao HC Costa Katherine Creutzinger
Katherine Creutzinger- 1Department of Animal and Food Science, University of Wisconsin-River Falls, River Falls, WI, United States
- 2Department of Veterinary and Animal Sciences, University of Vermont, Burlington, VT, United States
Dairy calves frequently experience hunger due to restricted milk allowance and long intervals between meals. Current methods to assess hunger in calves utilize intensive behavioral measures or training of calves for operant conditioning tests, which are generally not suitable for field research. By comparison, feeding a bitter substance to rodents has successfully been utilized to measure animal motivation to access resources, particularly in studies of addiction. Feeding a bitter substance in milk could be adapted to assess dairy calf motivation to drink milk, which could be interpreted hunger. The objectives of these experiments were to determine if 1) calf consumption of milk changed when a bitter substance was added and 2) the addition of a bitter substance to milk could be used to assess hunger in pre-weaned dairy calves. We hypothesized that calves would consume less milk as the concentration of a bitter additive increased and that calves would consume more milk with a bitter additive as the time between meals increased. Twenty-seven individually housed calves (17 Holstein, 10 dairy-beef terminal crosses; 21 ± 3 d of age) were enrolled in 2 Latin Square Design experiments. For each experiment, calves were blocked by breed and sex into one of two treatments: 1) bitter (milk altered with quinine) and 2) control (unaltered milk). In Experiment 1, calves received milk with 0.0, 0.2, 0.3, and 0.4 g/L quinine at their afternoon feeding. In Experiment 2, both bitter (0.35 g/L quinine) and control calves (0.0 g/L quinine) experienced different intervals between meals (6, 12, and 16 h). Milk intake, grain intake, and behavioral measures (time spent drinking, drinking bouts, non-drinking oral behaviors, and aversive behaviors) were recorded during the experimental feeding time for both experiments. Calves in Experiment 1 consumed less milk and performed more non-drinking oral behaviors when it was altered with quinine at 0.4 g/L compared to milk with 0.0, 0.2. and 0.3 g/L. More aversive behaviors (nostril/muzzle lick, lip smack, head bib/swing, and failed attempts to drink milk) were also performed when calves were fed milk with any quinine concentration compared to 0.0 g/L. The reduced milk consumption and performance of more aversive behaviors during the consumption of bitter milk indicates that calves find quinine aversive. Likewise, in Experiment 2, calves in the bitter treatment consumed less milk than control calves regardless of the length of time between meals. No interaction between treatment and feed withhold time was observed on milk intake, indicating calves did not change their willingness to consume bitter milk when feed was withheld for a longer time. However, calves provided unaltered milk performed more drinking bouts when milk was withheld longer, and all calves performed more non-drinking oral behaviors at 12 and 16 h and consumed more starter after longer periods between meals. These changes in behavior support increased hunger in calves despite the lack of difference in bitter milk intake with prolonged periods between meals. In conclusion, these results show that calves altered their consumption of bitter milk when the quinine concentration was greater, and that intake should not be the sole measure to consider when utilizing taste aversion to assess calf hunger.
Introduction
Hunger experienced by pre-weaned dairy calves due to restricted milk volumes has been an ongoing concern in the dairy industry. Young calves consume approximately 6 to 16 kg of milk per day and perform approximately 9 to 10 suckling bouts when housed with their dam and have unrestricted access to nurse (Kour et al., 2021; Churakov et al., 2023). Similarly, pre-weaned dairy calves fed with automatic feeders and ad libitum milk allowance consume an average of 10 L of milk per day and visit the feeder 9 times per day (Borderas et al., 2009; Curtis et al., 2018). However, under normal management on dairy farms, calves may go for long periods of time without milk and receive low milk volumes. Dairy calves are commonly fed twice per day at 12 to 16 hour intervals (USDA, 2016), which is a substantially longer than the time between meals in natural settings. The Nutrient Requirements of Dairy Cattle (NRC) recommended feeding calves 4 L of milk per day until the new suggestions were published in 2021, which increased the minimum recommended volume to 7 L of milk per day (National Academies of Sciences, E., and Medicine, 2021). The newly recommended volume is an improvement, but it is still less milk than calves would consume with unrestricted access to a milk source. Many calves are also transported and sold through livestock auctions, both events where milk is not provided and calves may not be fed for up to 28 consecutive hours or longer (Congress, U. S., 2023. 49 USC 80502).
Negative affective states, such as hunger, can be caused by restricted access to milk and may compromise calf welfare (D'Eath et al., 2009). Previous research has found that calves exhibit signs of hunger when they are fed suboptimal milk volumes (De Paula Vieira et al., 2008) and are weaned (De Passillé et al., 2011). Long periods of time between meals can also cause hunger in calves. For example, calves who are fed milk once per day play less than calves fed twice a day, which is indicative of poorer welfare (Jongman et al., 2020). Additionally, calves provided 2 meals per day from an automated feeder had more unrewarded visits than calves who were fed the same volume over 4 meals a day, which suggests that fewer meals per day (i.e., longer intervals between meals) results in increased hunger (MacPherson et al., 2019). Most of the research investigating hunger in pre-weaned calves relies on detailed behavioral analysis (De Paula Vieira et al., 2008; De Passillé et al., 2011) or training calves to perform sophisticated tasks to access milk (Lecorps et al., 2023). Post-hoc analysis of recorded video and operant training is cumbersome and requires a considerable amount of time. These methods are also impractical for testing hunger during field research (e.g., transportation or livestock auctions). Due to these reasons, research is limited in commercial settings due to limited times with access to calves as well as labor constraints for the number of animals to feasibly observe via video. New methods to assess hunger would improve our understanding of calves’ internal states regarding access to milk and create new opportunities to assess hunger in commercial settings.
Motivation tests are commonly used in animal welfare research to determine the importance of a resource to animals (Kirkden and Pajor, 2006). The theory of motivation poses that when the cost of a resource is low an animal will access a high amount of it but the desire to access the resource will vary depending on an animal’s motivation or need for it. If motivation to access a resource is low, an animal will decrease their effort to access it as the ‘cost’ increases (elastic demand), but if the motivation to access the resource is high, an animal will continue to pay the ‘cost’ (inelastic demand; Jensen and Pedersen, 2008). Motivation tests have been used to assess hunger in dairy cattle. For example, dairy cattle fed a low-energy diet had a shorter latency to feed after a feed thwarting test and attempted to feed more frequently than cows fed a normal-energy diet, indicating they were hungrier (Franchi et al., 2021). Calf motivation to consume milk, as reflected by the willingness to pay a cost for access to milk, could be a suitable measure to quantify hunger.
While limited work has been performed to directly evaluate calf motivation to consume milk, studies evaluating calf feeding behaviors have provided evidence of their high motivation to consume milk. One behavioral measure that has been linked to calf motivation to access milk is unrewarded visits to an automatic feeding system, which means that calves attempted to receive milk from an automatic feeder when they are ineligible for a meal. Calves have higher unrewarded visits to a feeder when they are provided small meal sizes (Morita et al., 1999) and are fed 6 L of milk daily compared to calves fed higher allotments of milk and during weaning period (≥8 L milk/day; Rosenberger et al., 2017); indicating that calves increase the number of times they attempt to get milk when it is restricted. When kept with their dam, calves also increased their number of feeding bouts when milk was restricted by milking the cow prior to calf access (de Passillé and Rushen, 2006). Calves increased the amount of work they were willing to exert to access milk in these artificial and semi-natural rearing systems, supporting the notion that calves are motivated to access milk, and that motivation is increased when milk is restricted.
In rodents, the willingness to consume a substance containing quinine has been utilized in models of motivation, particularly in models of alcohol addiction (Crabbe, 2012, 2014). Quinine is a bitter additive that is soluble in liquids and has an aversive flavor to rats (Contreras et al., 1995; Hopf et al., 2010; Lesscher et al., 2010). When rodents are highly motivated to access a resource, they are willing to consume quinine infused substances to gain access to the resource (Hopf et al., 2010). For example, when rats addicted to alcohol had it withheld for weeks and then were re-granted access, alcohol-dependent (inelastic demand) rats would maintain high levels of intake of an alcohol solution even when quinine was present. This is in contrast to non-addicted rodents that had previously consumed alcohol but decreased intake when quinine was included (Wolffgramm et al., 2000). Ruminants (Ginane et al., 2011), including dairy calves (Bernard, 1964) are also capable of tasting bitter flavors. For instance, calves did not show a preference for unaltered water compared to bitter water when quinine hydrochloride was included at lower concentrations, but when quinine was added at 2.5 mg quinine/100 mL water calves rejected the altered water (Goatcher and Church, 1970). Thus, the addition of a bitter substance to feed may have the potential to be adapted for use in dairy cattle to test motivation for feed, such as milk in calves.
This study aimed to explore if the addition of a bitter substance (quinine) to milk would be applicable to measure motivation to drink (i.e., hunger) in dairy calves for field research. The objectives of this study were to determine if calf consumption of milk changed when different concentrations of quinine were added (0.0, 0.2, 0.3, and 0.4 g/L) and if consumption of milk with quinine varied after different lengths of time between meals (6, 12, and 16 h). A second objective was to determine if feeding-related behaviors varied between calves fed unaltered and bitter milk. We hypothesized that calves would consume less milk as the concentration of quinine in milk increased and that calves would consume more milk with quinine when milk was withheld for longer compared to shorter periods of time. Additionally, we anticipated that feeding-related behaviors would vary between calves fed unaltered and bitter milk.
Materials and methods
This study was performed at the University of Wisconsin-River Falls Dairy Learning Center (River Falls, WI) from September 2022 to January 2023. Study protocols and calf management were approved by the Institutional Animal Care and Use Committee (20-21-47384).
Pre-weaned calves (n = 27; Table 1: 17 Holstein, 10 dairy-beef terminal crosses; 20 female, 7 male) were enrolled in this study at 21 ± 3 d of age to meet the required sample size per treatment (n = 12) for Experiments 1 and 2 (Table 1). The sample size was based on the number of calves per treatment in a similar study that assessed calves’ willingness to consume calf starter when a bitter flavor was added and achieved statistical significance between treatment groups (Terré et al., 2022). All calves were individually housed indoors using calf pens with wire mesh between the pens (1.9 m2, 5’ indoor calf pen, Calf-Tel, Germantown, WI; Figure 1) and bedded with sawdust (September-October) or straw (November-January). Calves were able to contact adjacent calves through the front of their pens and wire panel siding (Figure 1). Water and calf starter (Purina AMPLI-CALF Starter®: crude protein 20%, crude fat 2%, crude fiber 7.5%) were available ad libitum from the time calves were 2 d of age. Milk replacer (Land O’ Lakes Cow’s Match ColdFront PB® BOV DBZ Medicated milk replacer, Arden Hills, MN: crude protein 27%, crude fat 20%, crude fiber 0.3%; 300 g x 1 L water; 3.8 L fed at 38-40°C) was fed twice per day by pail buckets: at 0300h (‘morning feeding’) and 1500h (‘afternoon feeding’). Housing and feeding methods were set by farm procedures and adhered to for this study. To be included in the study, calves had to be healthy (i.e., no fever, signs of respiratory disease, or diarrhea) and not treated for illness within 5 d prior to the study. Calf health was monitored daily by the farm staff. Calves were removed from the study if they were identified as “sick” by the farm staff and treated with electrolytes or antibiotics during the study period.
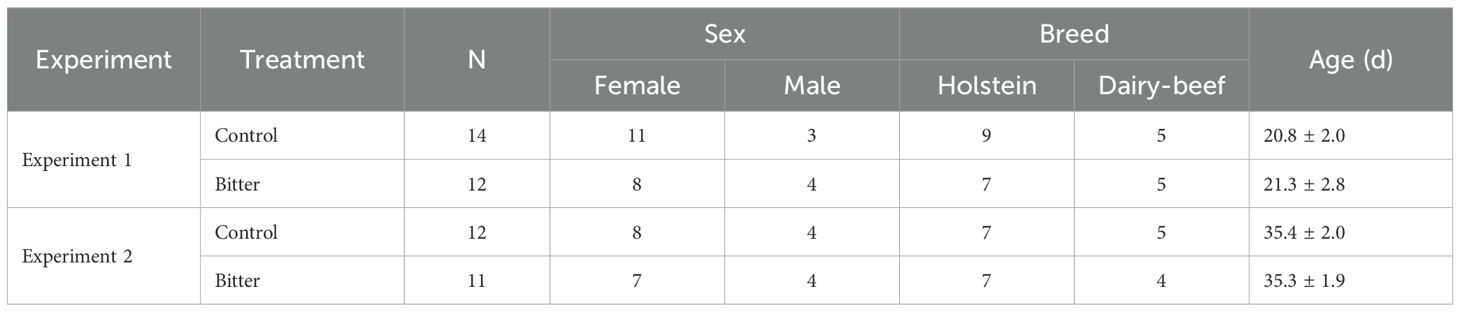
Table 1. Final calf characteristics including sex, breed, and age at enrollment in the control and bitter treatments for both Experiment 1 and Experiment 2 of the study.
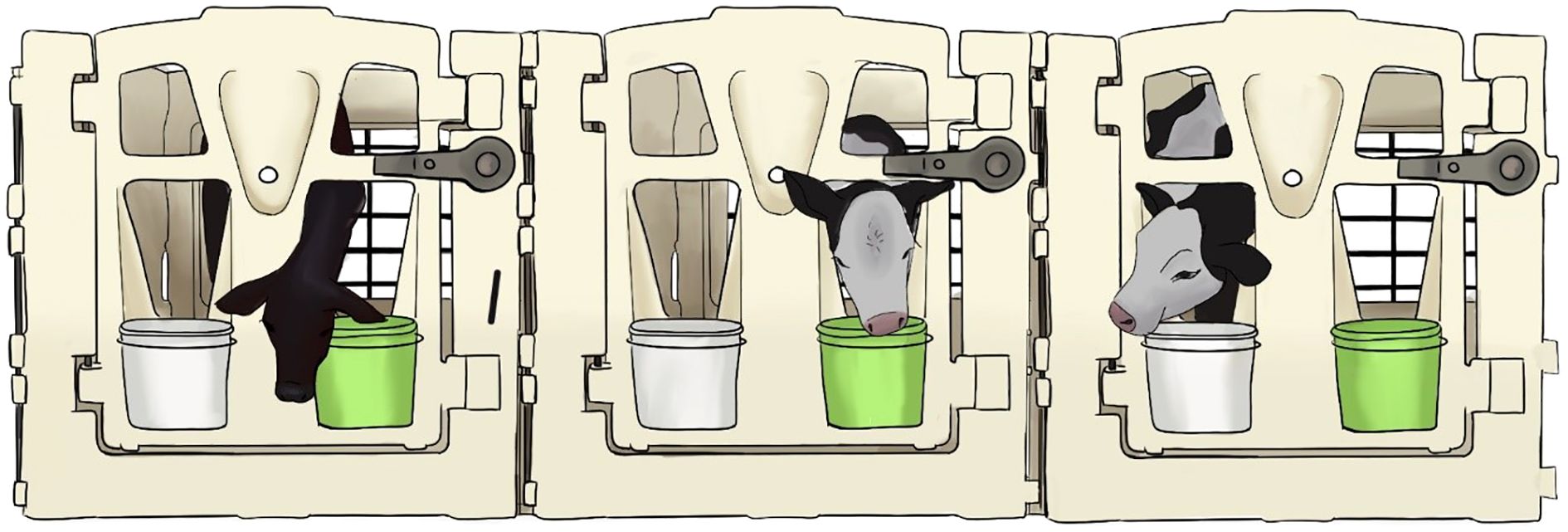
Figure 1. Diagram of calf housing and experimental set up. Calves were housed individually in pens. At the time of testing the water bucket was replaced with an experimental bucket (green). In experiment 1, calves received milk replacer with either 0.0, 0.2, 0.3, or 0.4 g/L quinine and in experiment 2, calves received either unaltered milk replacer or milk replacer with 0.3 g/L quinine after each feed withhold period. During each test period, calves still had access to grain (white bucket).
Experimentation was conducted on Monday and Thursday each week for a 2 wk period per experiment. Experiments 1 and 2 were performed successively (4 wks total). On study days, enrolled calves were fed an additional 1.9 L of milk replacer at the morning feeding (5.7 L) to ensure adequate milk was provided if milk was not consumed during test feedings. Morning milk refusals on days of experimentation were recorded (kg). At the morning feeding, calves were provided 1.0 kg of calf starter. The remaining calf starter was checked at the afternoon feeding and an additional 1.0 kg of calf starter was provided as needed. The leftover calf starter was removed and weighed during the morning feeding after experimental days (Tuesday and Friday morning feeding). Milk and calf starter consumption (kg) were determined by weighing the start weight of feed when provided and the refusals using an electronic scale (DT-580, Rujixu, China). Milk consumption of each feeding was assessed as the percentage of milk consumed per calf.
Calves were offered 3.8 L of milk replacer in a bucket during experimentation and buckets were removed 30 min after delivery, which was considered the milk feeding test period. During the milk feeding test period, calves had access to calf starter but not water. Calf behavior was continuously recorded with video cameras (EK700, Akaso, Frederick, Maryland) during the milk feeding test period. Cameras were placed approximately 0.9 m from the pen and 1.5 m from the ground, angled downward so it was possible to see the bottom of the milk bucket. Behavior data were observed from video using continuous observation in Boris software (Version 8.13, Torino, Italy) using an ethogram developed to capture feeding behaviors (Table 2). Two observers were trained to analyze videos from 10 videos that were 30 min long. A different set of 10 videos, 30 min in length were used to calculate interobserver reliability using the intraclass correlation coefficient including all duration and frequency data (R2 = 0.98). Both observers were blind to the treatment calves received during video analysis.
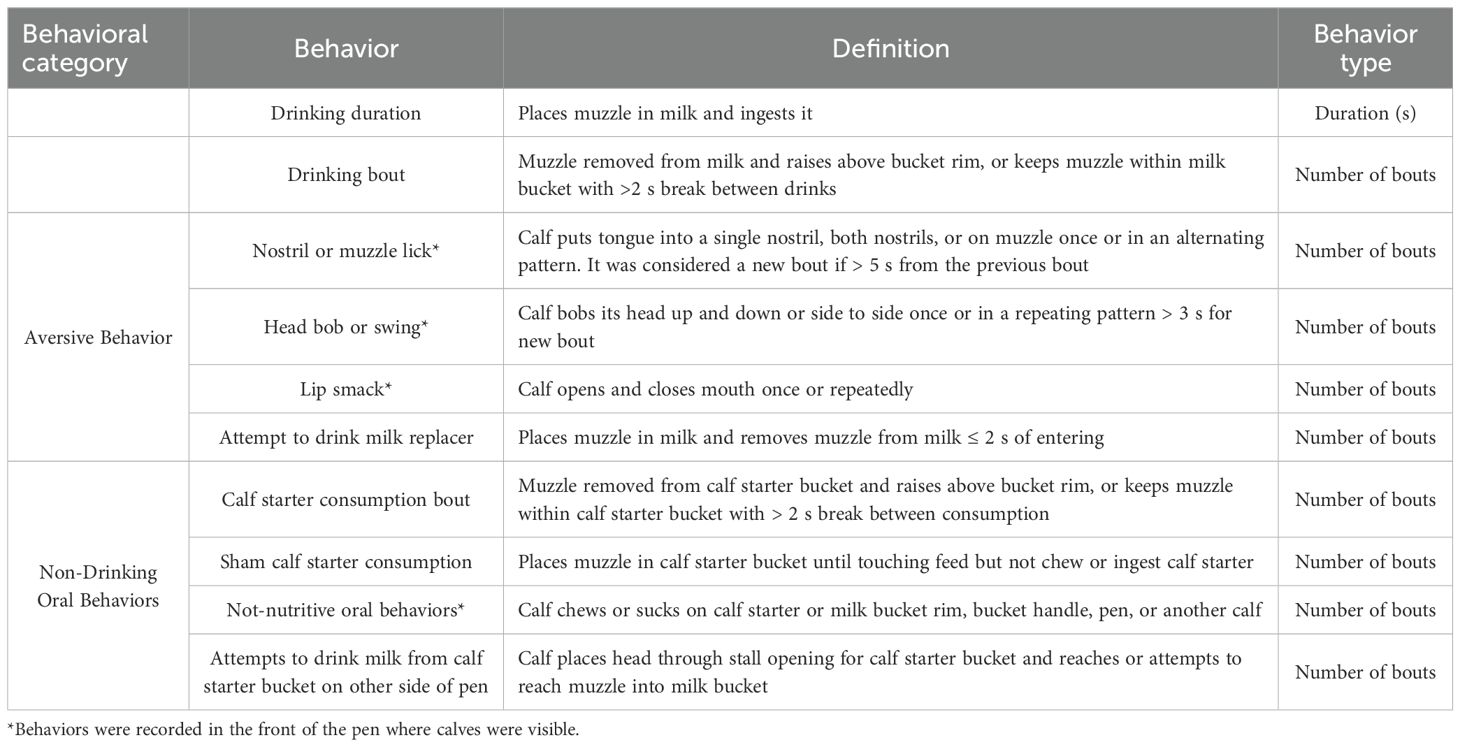
Table 2. Ethogram of behaviors measured continuously from video during the milk feeding test period (0 to 30 min after feed delivery) for Experiments 1 and 2.
Individual behaviors observed from the video were classified into categories for analysis because there were too few individual behaviors to analyze separately. In mammals other than cattle, research has assessed facial and lingual movements to evaluate aversion to bitter substances. For example, after tasting quinine, rats performed ‘gaping’ (defined as repeatedly lowering the mandible and retracting the lips around the mouth; Grill and Norgren (1978)) and humans and non-human primates performed oral and lingual movements, including tongue protrusion, gaping, and grimacing (eye squinching along with nose wrinkle (Steiner et al., 2001). In sheep, head shaking and lip smacking were observed when rams were exposed to phenylthiocarbamide, a different bitter compound (Henslee et al., 2019). Thus, the behaviors of nostril or muzzle lick, head bob or swing, lip smack, and attempt to drink milk replacer were grouped into the behavioral category ‘aversive behavior’ as they are reminiscent of the negative reaction to the presence of the quinine observed in other species. The behaviors of calf starter consumption, sham calf starter consumption, non-nutritive oral behaviors, and attempt to drink milk from the milk or calf starter bucket in front of the neighboring calf’s pen were grouped into the behavioral category ‘non-drinking oral behaviors’.
This study was comprised of two experiments. Experiment 1 tested the concentration of a bitter substance added to milk replacer (0.0, 0.2, 0.3, and 0.4 g/L quinine) on calf milk and starter consumption and feeding related behaviors. Experiment 2 tested calf consumption of milk and calf starter and feeding related behaviors after different lengths of time between feedings (6, 12, and 16 h). Calves were consecutively enrolled in Experiment 1 then 2 (Figure 2).
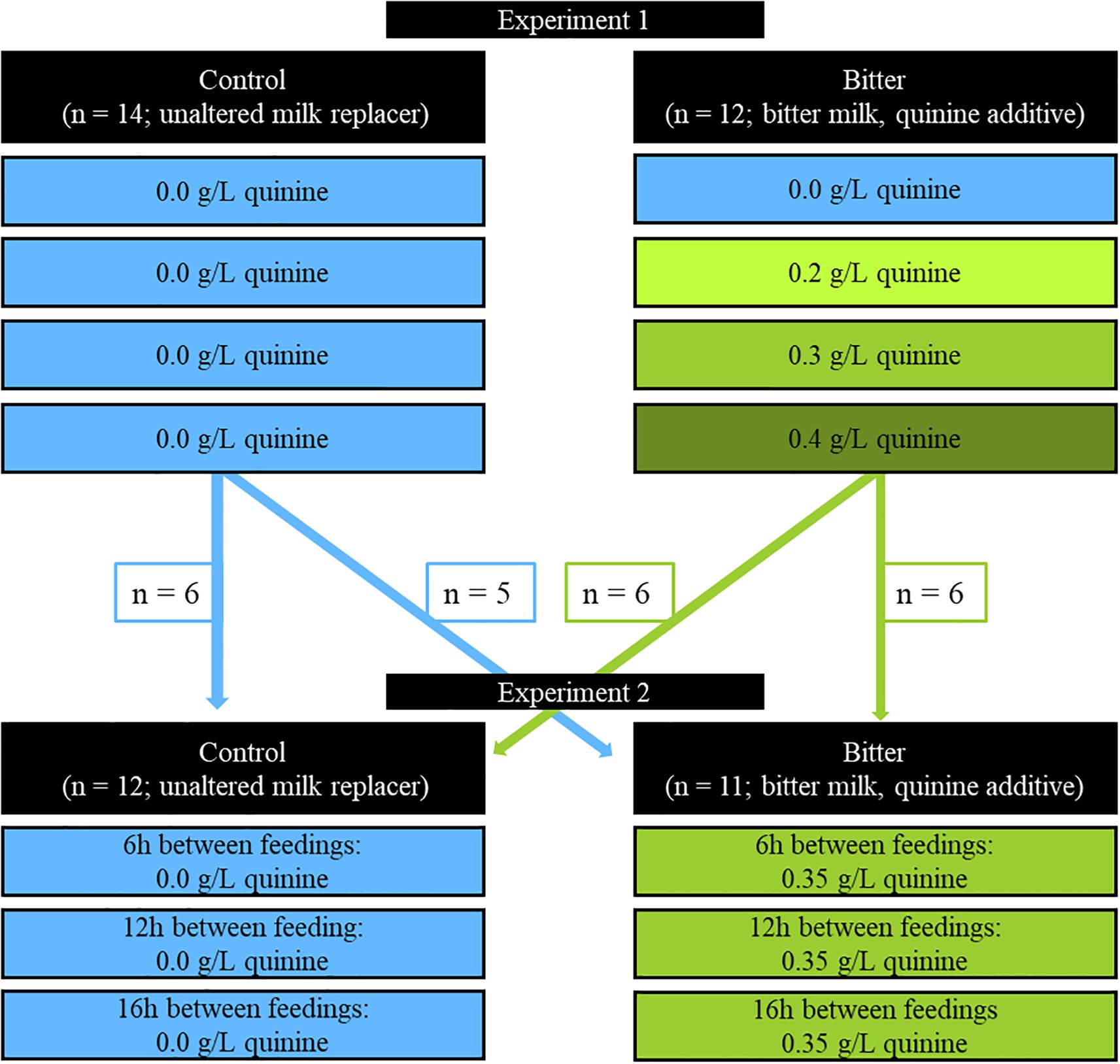
Figure 2. Overall study design showing calf enrollment in treatments in Experiment 1 and subsequent enrollment for Experiment 2. Figure shows treatments applied and possible exposures experienced by each treatment. Experiment tested the concentration of a bitter substance added to milk replacer (0.0, 0.2, 0.3, and 0.4 g/L quinine) on calf milk and starter consumption and feeding related behaviors. In Experiment 1, there were 8 possible random orders that calves received quinine concentration including 1) 0.0, 0.2, 0.3, 0.04 g/L, 2) 0.2, 0.0, 0.3, 0.4 g/L, 3) 0.0, 0.2, 0.4, 0.3 g/L, 4) 0.2, 0.0, 0.4, 0.3 g/L, 5) 0.3, 0.4, 0.0, 0.2 g/L, 6) 0.3, 0.4, 0.2, 0.0 g/L, 7) 0.4, 0.3, 0.2, 0.0 g/L, and 8) 0.4, 0.3, 0.0, 0.2 g/L. Control calves were not included in analysis for Experiment 1. Experiment 2 tested calf consumption of milk and calf starter and feeding related behaviors after different lengths of time between feedings (6, 12, and 16 h). In Experiment 2, there were 6 orders that calves received different feed withholding times, including: 1) 6, 12, 16 h, 2) 6, 16, 12 h, 3) 12, 6, 16 h, 4) 12, 16, 6 h, 5) 16, 6, 12 h, and 6) 16, 12, and 6 h. Calves were consecutively enrolled in Experiment 1 then 2.
Experiment 1
Experiment 1 included 2 treatments, a control group in which calves always received unaltered milk (“control”; n = 14) and an experimental group in which calves received various concentrations of a bitter substance in milk (“bitter”; n = 12 calves). Treatments were assigned with pseudo-randomization; the first treatment assignment was randomly assigned by coin flip, and the remaining calves were assigned to treatments in an alternating pattern. Treatments were balanced for sex and breed (Table 1).
On study days, calves were fed different concentrations of the bitter additive (Quinine hydrochloride dihydrate, MilliporeSigma, Darmstadt, Germany) in milk during the afternoon meal (1500 h). The calves were fed on a kilogram basis and offered 4.0 kg milk replacer (3.8 L). Control calves received 0.0 g/L of quinine each study day. Bitter calves were given 0.0 g/L, 0.2 g/L, 0.3 g/L, and 0.4 g/L quinine in milk on different study days. Calves in the bitter treatment received all the quinine concentrations of different days of the study using a Latin Square design. Because calf enrollment in the study was continuous, quinine concentration was blocked by day (i.e., all calves received the same quinine concentration on a single testing day). To ensure all calves received all quinine concentrations in a random order, the two weeks of study were blocked in 0.0 and 0.2 g/L quinine within a single week (alternating Monday and Thursday) and 0.3 and 0.4 g/L quinine within a single week (alternating Monday and Thursday). Thus, there were 8 possible random orders that calves received quinine concentration including 1) 0.0, 0.2, 0.3, 0.4 g/L, 2) 0.2, 0.0, 0.3, 0.4 g/L, 3) 0.0, 0.2, 0.4, 0.3 g/L, 4) 0.2, 0.0, 0.4, 0.3 g/L, 5) 0.3, 0.4, 0.0, 0.2 g/L, 6) 0.3, 0.4, 0.2, 0.0 g/L, 7) 0.4, 0.3, 0.2, 0.0 g/L, and 8) 0.4, 0.3, 0.0, 0.2 g/L.
To ensure an even distribution of quinine in milk, the quinine powder was reconstituted in 50 mL water until fully dissolved and then added to the bucket containing milk. Another 50 mL of water was added to the quinine vial and shaken, then added to the milk replacer to ensure any quinine residue was added to the milk. The reconstituted quinine-water and milk replacer were thoroughly mixed using a handheld whisk. Control milk was supplemented with an equal volume of water (100 mL) to reach an equivalent weight to bitter milk. Milk was weighed after the water and quinine-water solution had been added to the control and bitter milk, respectively.
Experiment 2
After Experiment 1, calves advanced to Experiment 2 of the study if they met the inclusion criteria. Experiment 2 included control (n = 12) and bitter (n = 11) treatments (Table 1). Calves were offered 4.0 kg of unaltered (“control”) or bitter milk (“bitter”; 0.35 g/L quinine) during the test period. This test period was defined as the second meal of a day, where the time between meals was manipulated: 6, 12, and 16 h following their morning meal. Calves experienced all three milk-withhold times regardless of treatment. The concentration of quinine was set based on Experiment 1 results, where calves reduced intake at 0.4 g/L compared to the other concentrations. For Experiment 2, this was reduced to 0.35 g/L quinine due to the pronounced decline of milk consumed observed at 0.4 g/L quinine in milk. Quinine and water were added to milk using the same protocol as Experiment 1. Treatments were balanced for sex (Table 1), and to control for treatment order from Experiment 1 to 2 (control-control = 5, control-bitter = 5, bitter-bitter = 5, bitter-control = 6); 4 and 2 calves were enrolled in only Experiment 1 or 2, respectively, due to not meeting the inclusion criteria. These times were selected based on the common feed intervals seen in the United States (USDA, 2016). This experiment was performed using a Latin-Square design; all calves received all milk withhold times in a random order. Because study enrollment was continuous, feed withholding time was blocked to individual days (i.e., all calves received the same feed withholding time on the same day). There were 6 orders that calves received different feed withholding times, including: 1) 6, 12, 16 h, 2) 6, 16, 12 h, 3) 12, 6, 16 h, 4) 12, 16, 6 h, 5) 16, 6, 12 h, and 6) 16, 12, and 6 h.
Statistical analysis
All data were recorded in Microsoft Excel (Microsoft Corp., Redmond, WA). Behavioral output was exported from Boris software (Friard and Gamba, 2016) in Microsoft Excel for analysis. Observations for each calf were summed into single values for each behavior type per calf including the total amount of time spent drinking (s) and the number of bouts of drinking, aversive behaviors, and non-drinking oral behaviors. Statistical analysis was performed in SAS (Version 9.4, Cary, NC). Data were visually inspected for distribution in SAS (PROC UNIVARIATE) before statistical analysis by each experiment to determine the appropriate model distributions to be utilized. For all models, univariable analysis (linear regression; PROC GLIMMIX) was performed between the outcomes and predictor variables. Fixed effects were included in multivariable models if P ≤ 0.20. Manual backward stepwise elimination was then used to remove fixed effects with the greatest P-value from the multivariable models; fixed effects were retained in the final model if P ≤ 0.20. Residuals were visually assessed with quantile-quantile and histogram plots for all models and models with the lowest Akaike Information Criterion (AIC) were selected. Significance and tendencies were declared at P < 0.05 and P < 0.10, respectively.
Experiment 1
For Experiment 1, control calves were excluded as to not skew the analysis from imbalanced data in the 0.0 g/L concentration group, and only calves in the bitter treatment group were evaluated. Multivariable models were utilized to determine if quinine concentration affected the percent of the milk meal consumed, calf starter intake, drinking duration, number of drinking bouts, non-drinking oral behaviors, and aversive behaviors. The univariable analysis evaluated quinine concentration (0.0, 0.2, 0.3, and 0.4 g/L), quinine concentration order, age (d), sex (male vs. female), and breed (Holstein vs. dairy-beef) for inclusion in each model. Quinine concentration was forced into the models regardless of significance. In all models, calf was included as a random effect, and Satterthwaite degrees of freedom approximation was used. Milk intake, calf starter intake, and drinking duration were analyzed using linear mixed models (PROC MIXED) used a compound symmetry covariance structure with a Gaussian distribution. The number of drinking bouts, non-drinking oral behavior bouts, and aversive behavior bouts were analyzed using generalized linear mixed methods models (PROC GLIMMIX) with a Poisson distribution). Differences in milk intake, calf starter intake, and feeding related behaviors between quinine concentrations were tested using hypothesis driven comparisons, including estimation statements between 0 vs. 0.2 g/L, 0 vs. 0.3 g/L, 0 vs. 0.4 g/L, 0.2 vs. 0.4 g/L, and 0.3 vs. 0.4 g/L.
Experiment 2
Calves in both the control and bitter treatment groups were included for analysis. Multivariable models were utilized to determine if treatment (bitter vs. unaltered milk) and time milk was withheld (6, 12, 16 h) affected the percent milk consumed, calf starter intake, drinking duration, number of drinking bouts, non-drinking oral behaviors, and aversive behaviors. The univariable analysis for Experiment 2 evaluated treatment and milk withhold time interaction, feed withholding order, Experiment 1 treatment (bitter vs. control), age (d), sex (male vs. female), and breed (Holstein vs. dairy-beef) for model inclusion. Treatment and milk withhold time were forced into all models regardless of treatment. Interactions were retained in the models if significant. In all models, calf was included as a random effect and a Satterthwaite degrees of freedom approximation was used. Models for milk intake, calf starter intake, and drinking duration used a compound symmetry covariance structure with a Gaussian distribution (linear mixed model, PROC MIXED). The models for the number of drinking bouts, non-drinking oral behavior bouts, and aversive behavior bouts had Poisson distribution using generalized linear mixed methods models (PROC GLIMMIX). For experiment 2, the model for non-drinking oral behaviors did not converge due to over dispersion and so negative binomial regression was utilized. Differences in milk intake, calf starter intake, and feeding behaviors between milk withholding times were tested using hypothesis specific comparisons, including estimation statements between 6 vs. 12 h, 6 vs. 16 h, 12 vs. 16 h, and bitter vs. control milk.
Results
Experiment 1
Fixed effect inclusion
The final fixed effects included in the multivariable models following univariable analysis and stepwise backward elimination and the final P-value for each fixed effect included are reported. The final milk consumption model included only quinine concentration, and no fixed effects were retained. The final calf starter consumption model included age (P < 0.001), as a fixed effect in addition to quinine concentration. For the models of other behaviors, non-drinking oral behaviors included age (P = 0.02), and no other fixed effects were retained in drinking duration or drinking and aversive behavior bout models.
Feed intake
The percent milk consumed by calves was associated with quinine concentration ([Test statistic df] F = 4.31 3, 33, P = 0.01; Figure 3A). Calves fed 0.4 g/L quinine consumed less milk than calved fed 0.0 g/L (LSM ± SEM; 40.5% ± 11.3% vs. 84.6% ± 11.7%, respectively; T = 3.23 33, P = 0.002), 0.2 g/L (77.9% ± 10.9%; T = 2.83 33, P = 0.006), and 0.3 g/L (74.0% ± 11.3%; T = 2.57 33, P = 0.02). However, there was no difference in milk consumption between calves fed 0.0 g/L and 0.2 g/L quinine (T = 0.51 33, P = 0.62) or 0.3 g/L (T = 0.75 33, P = 0.43). Calf starter intake was associated with quinine concentration (F = 3.93 3, 29, P = 0.02; Figure 3B). Calves consumed more calf starter when the milk had 0.4 g/L quinine (0.29 ± 0.05 Kg) than when they were fed milk with 0.2 g/L (0.16± 0.04 Kg; T = -2.79 29, P = 0.009) and 0.3 g/L quinine (0.15 ± 0.04 Kg; T = -3.05 29, P = 0.005). Age was also significantly associated with calf starter intake; calf starter intake increased as age increased (intercept = 0.02, slope = 0.003; F = 21.01 1,29, P < 0.001).
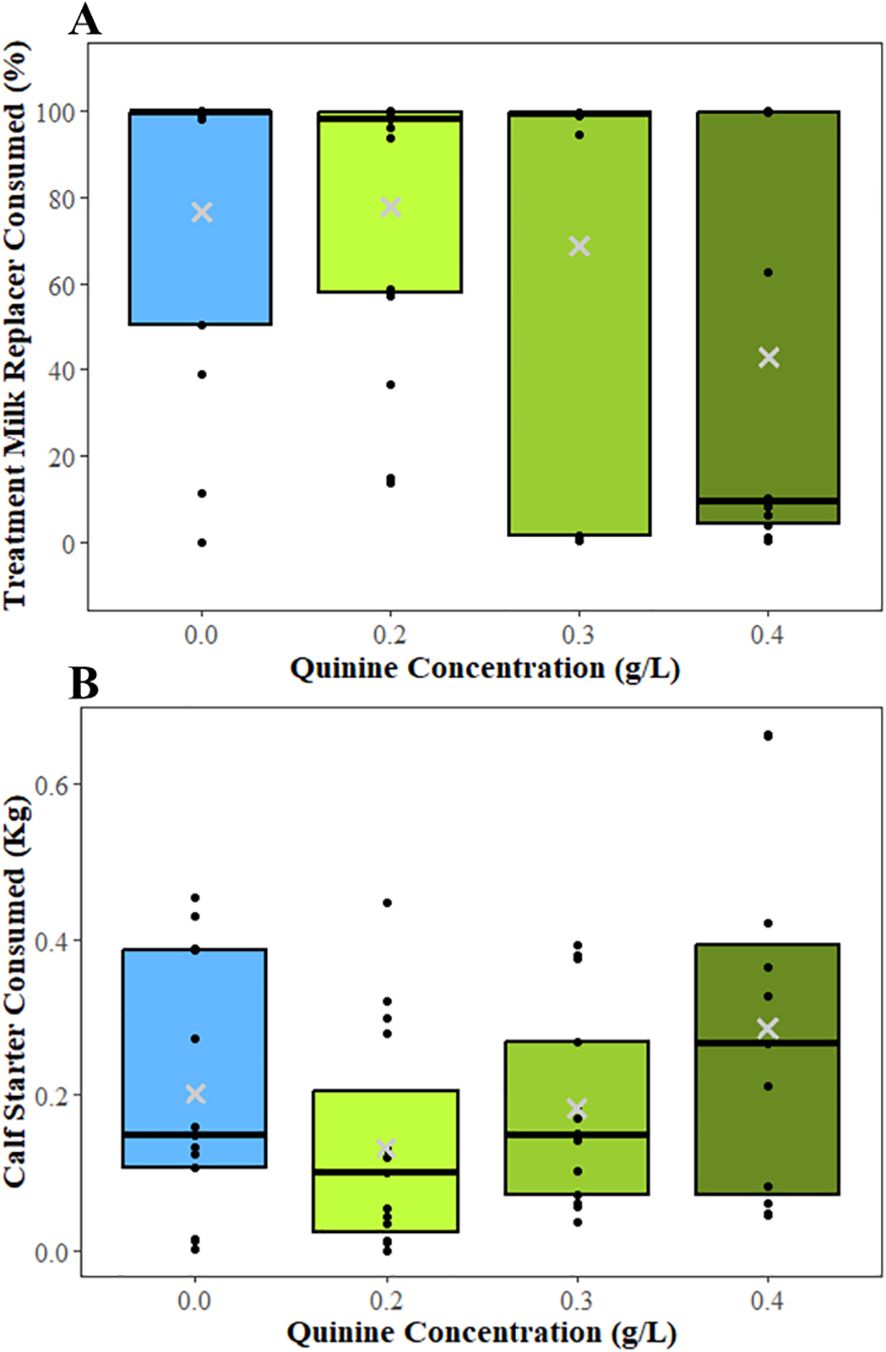
Figure 3. Boxplots showing the median (black line), interquartile range, and mean (gray ×) for (A) percent milk replacer consumed for calves that received bitter milk and (B) amount of calf starter consumed in experiment 1 when calves were provided milk with the quinine concentrations of 0.0 g/L, 0.2 g/L, 0.3 g/L, and 0.4 g/L for their evening meal. Raw values are presented in the boxplots. All calves received every concentration once in a random order. Black dots represent the values of individual calves. Differences found by quinine concentration from the mixed models are denoted with letters; boxes with differing letters were significantly different (P<0.05).
Quinine concentration and behavior
Quinine concentration was not associated with the duration of time (s) spent drinking (F = 1.74 1, 31, P = 0.18; Figure 4A). However, the number of drinking bouts varied with quinine concentration (F = 6.51 1,41, P = 0.001; Figure 4B). Calves given milk with 0.0 g/L quinine performed fewer drinking bouts (1.38 ± 0.22 bouts) than calves fed 0.2 g/L quinine (1.88 ± 0.21 bouts; T = -2.96 41, P = 0.005), 0.3 g/L quinine (1.75 ± 0.22 bouts; T = -2.09 41, P = 0.04), and 0.4 g/L quinine (2.10 ± 0.20 bouts; T = -4.34 41, P < 0.001). Additionally, calves performed fewer drinking bouts at 0.3 g/L than 0.4 g/L (T = -2.22 41, P = 0.03); there were no other differences between quinine concentrations.
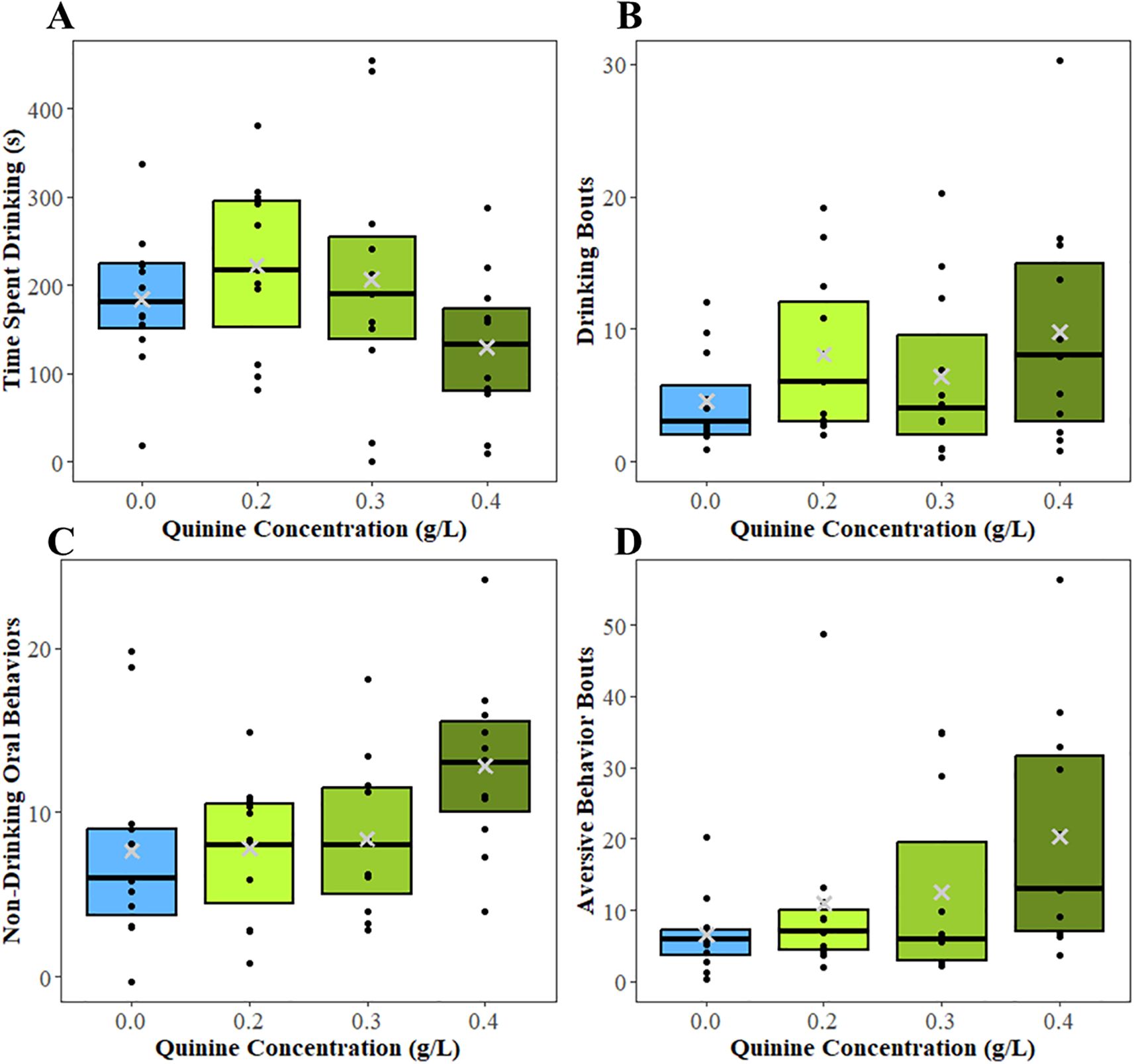
Figure 4. Boxplots showing the median (black bar), interquartile range, and mean (gray ×) for (A) time spent drinking (s), (B) number of drinking bouts, (C) number of non-drinking oral behaviors, and (D) number of aversive behavior bouts in experiment 1 when calves were provided milk with the quinine concentrations of 0.0 g/L, 0.2 g/L, 0.3 g/L, and 0.4 g/L for their evening meal. Raw values are presented in the boxplots. Behavioral analysis was performed from the time milk was delivered to 30 min after milk delivery. All calves received every quinine concentration once in a random order. Black dots represent the values of individual calves. Differences found by quinine concentration from the mixed models are denoted with letters; boxes with differing letters were significantly different (P<0.05).
There was an effect of quinine concentration on the number of non-drinking oral behavior bouts (F = 6.69 3, 40, P = 0.005; Figure 4C); calves given milk with 0.4 g/L quinine performed more non-drinking oral behaviors (2.46 ± 0.14 bouts) than calves given 0.0 g/L (2.00 ± 0.15 bouts; T = -3.96 40,P < 0.001), 0.2 g/L (2.12 ± 0.12 bouts; T = -3.29 40, P = 0.02), and 0.3 g/L (2.05 ± 0.16 bouts; T = -2.91 40, P = 0.005) quinine concentrations. Non-drinking oral behavior bouts also increased as age increased (intercept = 0.03, slope = 0.01, F = 5.77 1,40, P = 0.02). There was an effect of quinine concentration on the number of aversive behavior bouts (F = 27.00 3, 41, P < 0.001; Figure 4D). Calves fed 0.0 g/L quinine performed fewer aversive behavior bouts (1.69 ± 0.22 bouts) than calves fed 0.2 g/L (2.15 ± 0.21 bouts; T = -3.16 41, P = 0.003), 0.3 g/L (2.53 ± 0.20 bouts; T = -5.82 41, P < 0.001), and 0.4 g/L (2.79 ± 0.20 bouts; T = -8.32 41, P < 0.001); calves fed 0.4 g/L quinine also expressed more aversive bouts that calves fed 0.2 g/L (T = -5.56 41, P < 0.001) and 0.3g/L (T = -2.27 41, P = 0.03) quinine.
Experiment 2
Fixed effects inclusion
The final fixed effects included in the multivariable models following univariable analysis and stepwise backward elimination for Experiment 2 are reported. No additional fixed effects were retained in the final model for milk consumption. The final calf starter consumption model included calf age (P = 0.002) as a fixed effect. For final behavioral analysis models, no additional fixed effects were retained for drinking duration, the interaction between treatment and milk withhold time was retained for drinking bouts (P < 0.001), breed was retained for non-drinking oral behavior bouts (P = 0.03), and sex was retained for the aversive behavior bouts (P = 0.007).
Feed intake
Calves in the bitter treatment group consumed less milk than calves in the control group (LSM ± SEM; 44.9 ± 8.0% vs. 89.7 ± 7.8% of the fed amount, respectively; ([Test statistic df] F = 16.22 1, 21, P < 0.001; Figure 5A). However, the amount of milk consumed was not associated with milk withhold time (61.2 ± 8.2% at 6 h, 64.5 ± 8.2% at 12 h, and 76.1 ± 8.1% at 16 h; F = 1.17 2, 43, P = 0.32). Calves in the control group tended to consume less calf starter than calves in the bitter group (F = 3.17 1, 40, P = 0.08; Figure 5B). Calf starter intake was associated with milk withhold time (F = 5.16 2, 40, P = 0.01; Figure 5B). Calves consumed less calf starter when feed was withheld for 6 h than when it was withheld for 12 h (0.40 ± 0.05 kg vs. 0.66 ± 0.05 kg, respectively; T = -3.21 40, P = 0.002; Figure 5B). Calves tended to consume more calf starter when feed was withheld for 16 h compared to 6 h (T = -1.88 40, P = 0.06). There was no difference in calf starter intake between 12 h and 16 h milk withhold intervals (T = 1.4940, P = 0.12). Calf starter intake also increased as age increased (intercept = 0.03, slope = 0.009; F = 10.66 1, 40, P = 0.002).
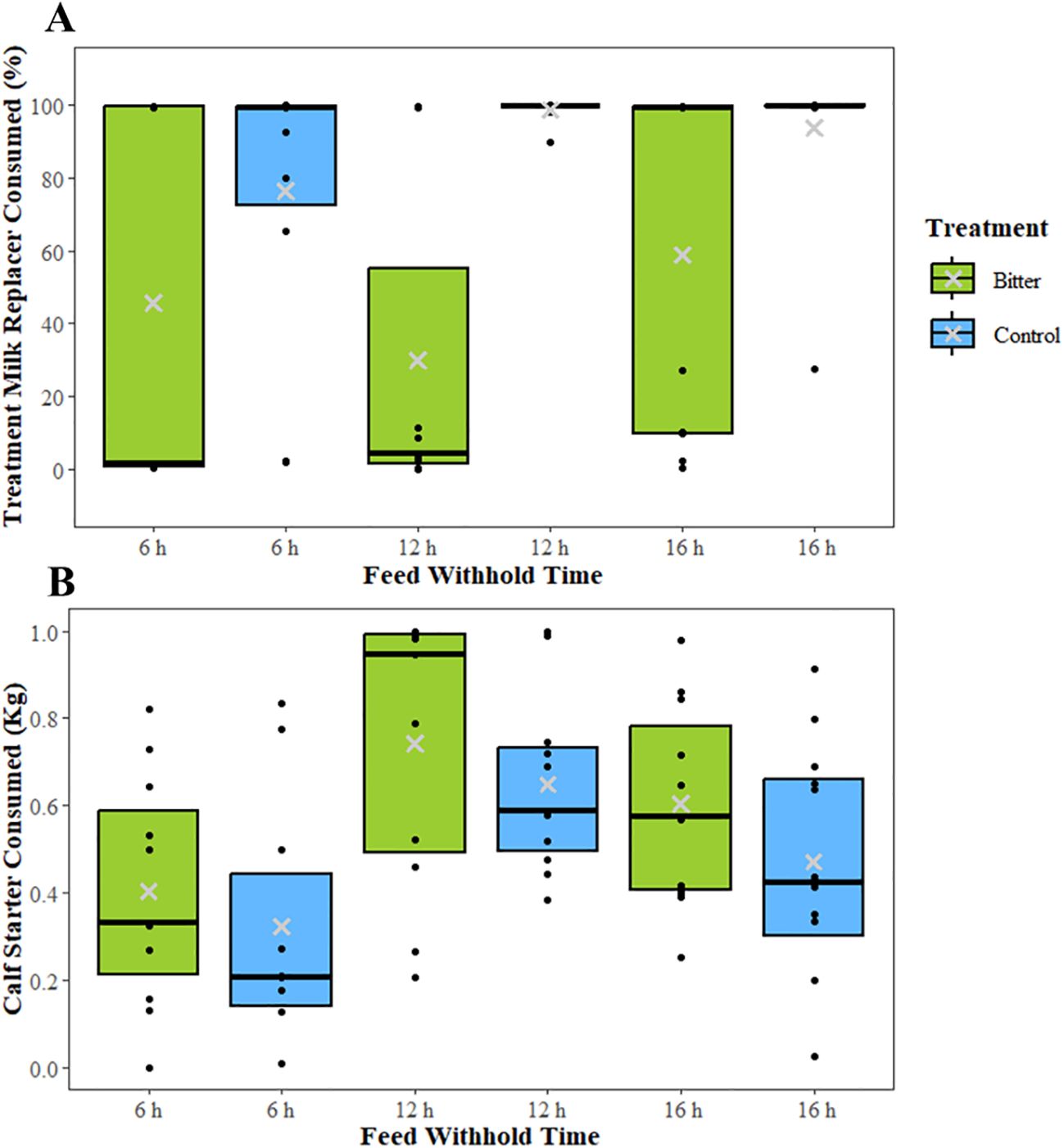
Figure 5. Boxplots showing the median (black line), interquartile range, and mean (gray ×) for (A) percent milk replacer consumed and (B) amount of calf starter consumed in experiment 2 when calves were provided either milk with 0.35 g/L quinine (bitter) or unaltered milk (control) after feed was withheld for 6 h, 12 h, or 16 h. Raw values are presented in the boxplots. Behavioral analysis was performed from the time the milk was delivered to 30 min after milk delivery. All calves received experienced each milk withhold time once in a random order. Black dots represent the values of individual calves. Differences found by quinine treatment from the mixed models are denoted with letters, and differences found by feed withhold time from the mixed models are denoted with symbols. Comparisons were made between treatments within each time point, and within treatments across withhold times. Boxes with different letters or symbols were significantly different (P < 0.05).
Milk withhold time and bitter milk on behavior
The time spent drinking milk tended to be longer for calves fed unaltered compared to bitter milk (F = 4.14 1,19, P = 0.06; Figure 6A) and there was no effect of milk withhold time (F = 1.83 2,29, P = 0.18). An interaction between milk withhold time and treatment on the number of drinking bouts was observed (F = 9.92 2,44, P < 0.001; Figure 6B). Calves fed unaltered milk (control) had fewer drinking bouts after milk was withheld for 6 h (0.85 ± 0.28 bouts) than observed at 12 h (2.68 ± 0.32 bouts; T = -6.31 44, P < 0.001) and 16 h (1.38 ± 0.27; T = -2.54 44, P = 0.01), and fewer drinking bouts after milk was withheld for 16h than at 12 h (T = 4.79 44, P < 0.001). However, this relationship between drinking bouts and time milk was withheld was not observed in calves provided bitter milk (P > 0.05).
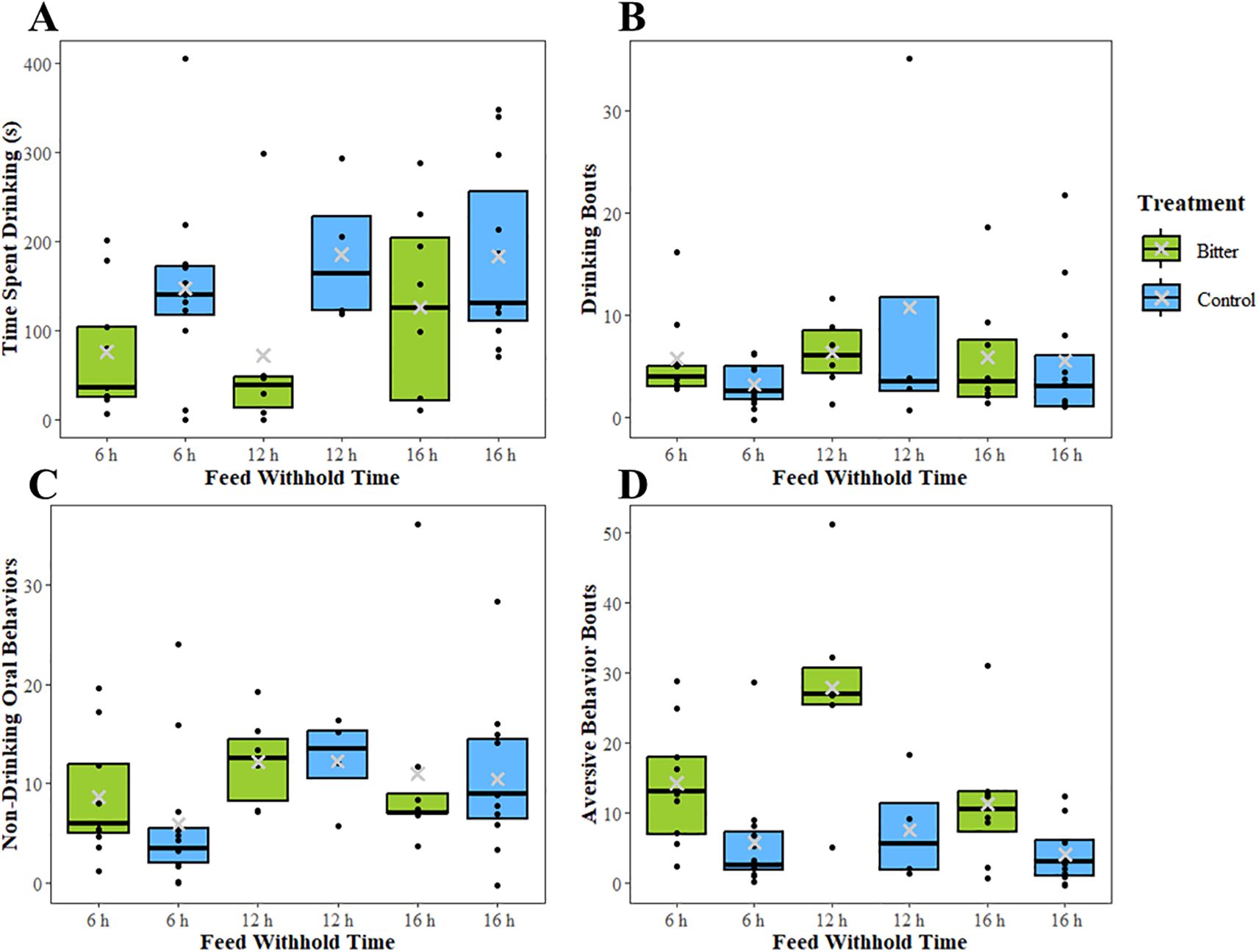
Figure 6. Boxplots showing the median (black line), interquartile range, and mean (gray ×) for (A) time spent drinking (s), (B) number of drinking bouts, (C) number of non-drinking oral behaviors, and (D) number of aversive behavior bouts in experiment 2 when calves were provided either milk with 0.35 g/L quinine (bitter) or unaltered milk (control) after feed was withheld for 6 h, 12 h, or 16 h. Raw values are presented in the boxplots. All calves received experienced each milk withhold time once in a random order. Black dots represent the values of individual calves. Differences found by quinine treatment from the mixed models are denoted with letters, and differences found by feed withhold time from the mixed models are denoted with symbols. Comparisons were made between treatments within each time point, and within treatments across withhold times. Boxes with different letters or symbols were significantly different (P < 0.05).
Milk withhold time was significantly associated with the number of non-drinking oral behaviors (T = 5.53 2, 25, P = 0.01; Figure 6C). Calves expressed fewer non-drinking oral behaviors when feed was withheld for 6 h (2.03 ± 0.18 bouts) than when it was withheld for 12 h (2.66 ± 0.22 bouts; T = -3.01 25, P = 0.006) and 16 h (2.48 ± 0.18 bouts; T = -2.60 28, P = 0.01). Additionally, Holstein calves performed fewer non-drinking oral behaviors than dairy-beef crossbreeds (2.02 ± 0.14 vs. 2.76 ± 0.28 bouts, respectively; T = 2.36 13, P = 0.03). Finally, calves in the bitter treatment group performed more aversive behaviors than control calves (2.34 ± 0.21 vs 1.28 ± 0.20 bouts respectively, F = 17.86 1, 17, P < 0.001; Figure 6D). Milk withhold time was also associated with aversive behaviors (T = 20.71 1,17, P < 0.001). Calves performed fewer aversive behaviors when feed was withheld for 16 h (1.46 ± 0.17 bouts) compared to 6 h (1.78 ± 0.17 bouts; T = 2.88 45, P = 0.006) and 12 h (2.20 ± 0.18 bouts; T = 6.38 45, P < 0.001), and fewer aversive behaviors were observed after milk was withheld 6 h compared to after 12 h (T = -3.92 45, P < 0.001). Sex was also associated with number of aversive behaviors, where heifers performed more aversive behaviors than bulls (2.28 ± 0.14 vs. 1.34 ± 0.29 bouts, respectively; T = 2.92 25, P = 0.007).
Discussion
The objectives of these experiments were to assess if calves altered their milk consumption when a bitter flavor was added and if calves consumed more milk with a bitter substance when milk was withheld for longer periods of time. This study supported the hypothesis that a bitter additive would decrease the amount of milk consumed, as calves provided milk replacer with 0.4 g/L quinine consumed less milk than calves provided milk replacer with smaller concentrations or no quinine. However, contrary to the hypothesis, intake of milk replacer with 0.35 g/L of quinine did not vary between feed withholding times.
Experiment 1
In Experiment 1, calves consumed less milk when the greatest concentration of quinine was present compared to all other concentrations (0.4 g/L vs. ≤ 0.3 g/L). Lower milk consumption was not observed when milk was altered with 0.2 g/L or 0.3 g/L quinine compared to milk without quinine. Calves also consumed more calf starter when the highest concentration of quinine was provided, compared to other lower concentrations, corresponding to the decreased milk consumption also observed at the highest concentration of quinine. An increase in calf starter intake was likely an effort to compensate for the decreased energy and nutrients typically received from milk when refused by calves. The depression of milk intake at the greatest quinine concentration is consistent with previous literature that evaluated cattle responses to bitter additives. In general, domestic ruminants are capable of tasting bitter flavors, though they may be less sensitive to bitter additives than other mammals (Ginane et al., 2011). When taste reception was evaluated in calves, quinine sulphate (bitter flavor) produced a relatively small electrophysiological response when passed over the tongue of anesthetized calves in comparison to other substances, including sucrose (sweet flavor), sodium chloride (salty flavor), acetic acid (sour flavor), and others (Bernard, 1964). Calf reactions to quinine hydrochloride in water at different concentrations showed that calves were first indifferent to quinine hydrochloride at lower concentrations (0.63 and 1.25 mg/100 ml) and then transitioned immediately to rejection at higher concentrations (2.5 mg/100 ml) (Goatcher and Church, 1970). Notably, all quinine concentrations utilized in Experiment 1 were greater than those in Goatcher and Church (1970). Therefore, this study builds upon the findings that show cattle have low sensitivity to bitter flavors but still find them unpalatable.
Calves started and stopped drinking milk more frequently (drinking bouts) and performed more aversive behaviors (nostril or muzzle lick, head bob or swing, lip smack, and attempt to drink milk replacer) at all quinine concentrations compared to unaltered milk, which may suggest that calves were able to taste quinine at all concentrations. Similarly, calves in another study also had more fragmented feeding behaviors (more pauses per suckling bout) when they were provided an acidified versus an unacidified milk replacer (Todd et al., 2018). Despite reduced intake of milk only occurring at the highest quinine concentration, the increased feeding bouts and performance of aversive behaviors at all quinine concentrations may indicate that calves can taste quinine and that it was aversive at all concentrations.
The expression of non-drinking oral behaviors only increased at a quinine concentration of 0.4 g/L compared to all other concentrations. Non-drinking oral behaviors in this study consisted of attempts to consume feed from a source other than the altered milk, including the consumption of calf starter between drinking bouts or attempting to drink from their neighbors’ milk bucket, and sucking or chewing on another calf or non-nutritive object. This increase in non-drinking oral behaviors occurred concurrently with depression in milk intake at quinine concentrations of 0.4 g/L. An increase in non-drinking oral behaviors may be an indicator of frustration because calves were unwilling to consume bitter milk but were still motivated to eat. Frustration in response to restricted access to expected feed has been documented in other livestock species. For example, when hens were trained to receive a feed reward in response to pressing a key, an increase in pecking of an alternative substrate (feathers) and repeated pecking of the trigger for the reward were observed when the feed reward was withheld (Rodenburg et al., 2002). In this study, the non-drinking oral behaviors may show frustration from palatability restriction from milk rather than physical restriction from milk. It is also possible that calves were hungry but sought an alternate feed source because they chose not to drink from the bitter milk available to them. Further research is needed to link these non-drinking oral behaviors with frustration from restriction of an expected feed reward and/or hunger in pre-weaned calves.
The results from Experiment 1 provided a basis that quinine has the potential to be used as a method to test calf motivation to consume milk through taste aversion. Application of a taste aversion test would be useful in field research where options for behavioral assessment are limited and to increase the number of experimental units enrolled in a study, which is frequently limited by the human effort required to review animal behavior for research. However, the low response of calves to quinine at smaller concentrations with an increase to high aversion at 0.4 g/L may present a challenge to the applicability of this test. Future research is encouraged test calf preference between multiple quinine concentrations in feed or the preference between different aversive flavor profiles.
Experiment 2
As in Experiment 1, calves consumed less milk when a bitter additive was present than when the milk was unaltered. However, calves did not consume significantly different amounts of bitter or unaltered milk at different lengths of time between meals. This suggests that calves were not more motivated to drink bitter milk offered after milk was withheld for up to 16 h. However, a lack of motivation to consume milk would be contrary to previous literature indicating that calves are highly motivated to consume milk when it is restricted (Morita et al., 1999; de Passillé and Rushen, 2006; Rosenberger et al., 2017). Given the novelty of using taste aversion to assess calf motivation, these results should be interpreted cautiously. For instance, individual variation in willingness to consume a novel feed may have influenced the outcomes. There was a large amount of individual variation in the willingness to consume bitter milk after milk was withheld; calf intake of bitter milk after a 12 h inter-meal interval ranged from 0.1% to 99.7% of the offered milk. Previous work in dairy calves has shown that personality is related to calf response stressors such as diarrhea, dehorning, and weaning (Woodrum Setser et al., 2024a), as well as mixing into new groups (Occhiuto et al., 2023). For example, calves with a low score for the trait ‘Active’ had more unrewarded visits to an automatic milk feeder during weaning and, thus, were considered hungrier (De Paula Vieira et al., 2008; Woodrum Setser et al., 2024c). Some individual calves in the current study may have been more sensitive to hunger (consuming all the bitter milk) or have been more sensitive to a bitter additive (refused all of it). The large amount of variation in the amount of bitter milk consumed may also suggest that a larger sample size would be required to detect a difference in motivation using this specific motivation test design, or increasing the intensity of the bitter additive may decrease the amount of variation observed.
It is also possible that calf hunger was affected by the extra 1.9 L of milk fed in the morning on test days before application of the taste aversion motivation test. Additional milk was provided to ensure adequate milk was provided if milk was not consumed during test feedings, as an ethical consideration. Access to a resource outside of the test period has been shown to influence the elasticity of demand for the resource within a motivation test. For instance, one study explored rat motivation for water after water was manipulated before the motivation test period; rats that received water 1 h, 2 h, or 4 h before the test were less willing to push a weighted lever to receive water than rats who did not receive water before the motivation test (Ladewig et al., 2002). While the additional milk provided in this study occurs with the morning meal which was well before motivation test period following feed withholding, it is unknown how long after receiving additional milk in the morning feeding calves experience increased satiety. There is some evidence that at 6 h calves were less hungry, as there is greater variation in consumption of the unaltered milk than is observed after feed was withheld for 12 and 16 h; however, this difference was numerical and not statistically significant. Further research should evaluate length of satiety based on meal size in calves.
Animals show high motivation for food across species, and food is consistently classified as a resource with inelastic demand, meaning animals will proportionately increase their effort to achieve the resource even as the cost to obtain it increases (Dawkins, 1990; Jensen and Pedersen, 2008). Kirkden et al. (2003) suggest that measuring other behaviors during a motivation test, including time and effort to obtain a resource, can give a more wholistic view of animal motivation rather than simply measuring the amount of the resource consumed. Indeed, in studies evaluating feeding behaviors in calves and cows, the assessment of other behaviors during the test, rather than just feed intake was essential to understanding individual differences in willingness to consume a novel feed (Costa et al., 2020; Woodrum Setser et al., 2024b). Therefore, it is important to consider other measures, such as calf behaviors during the test, to gain a holistic picture of calf experience when milk is withheld.
The first additional behavior to consider is calf starter intake. Calves increased their intake of calf starter when milk was withheld for longer period; calves consumed more after 12 h than at 6, and tended to consume more at 16 h than after 6h. Increased solid feed consumption in response to restricted milk allowance is a well-accepted indicator of hunger (Khan et al., 2011). For example, calves consume more concentrate when smaller milk volumes are provided before weaning (4 L vs 12 L; Borderas et al., 2009; Steele et al., 2017). Therefore, greater calf starter intake during longer inter-meal intervals may have been observed in response to calf hunger. It is also possible that calf starter intake may have influenced the degree of hunger experienced during the motivation test, as calves always had access to solid feed. However, the increase in calf starter intake was seen regardless of treatment. This suggests that calves were likely hungrier when fed 12 and 16 h after their morning meal than 6 h, especially because calves fed at the 6 h interval were also fed their normal afternoon meal. Further, calves fed bitter milk tended to consume more calf starter than calves fed unaltered milk. Calves in the bitter treatment group were likely hungrier than those in the control group because they consumed less milk, thus compensating for the limited feed intake. Withholding calf starter on test days could have prohibited calves from compensating for a lack of feed and elucidated more treatment differences. However, we felt as though removing all feed sources from calves would be an ethical concern due to negative affective states caused by hunger. Calf starter intake also increased with calf age, where older calves consumed more calf starter than younger calves, which is consistent with expectations (Neave et al., 2019). Since both feed withhold time and calf age were associated with calf starter intake, one should use caution generalizing these finding across different calf ages, where younger calves with less developed rumens may not be consuming calf starter at significant levels yet (Eckert et al., 2015). However, within this study calves were the same age at the start of Experiment 2 and the ages at which calves received different feed withholding times were balanced by the Latin Square design.
Milk withhold time and treatment also impacted the behaviors expressed during the test period. Regardless of treatment, calves increased the number of drinking bouts when milk was withheld for 12 h compared to 6 h. An interaction was also seen between treatment and feed withhold time; calves fed unaltered milk started and stopped drinking less frequently after milk was withheld for 6 h compared to 12 and 16 h and had more drinking bouts at 12 than 16 h. However, this relationship was not observed in calves fed bitter milk. Additionally, we did not observe that milk treatment affected the number of drinking bouts. These results are contrary to our findings in Experiment 1, which found calves performed more drinking bouts when a bitter additive was in their milk, and previous research, which also saw an increase in drinking bouts with acidified milk (Todd et al., 2018). This difference in results is likely due to the combined effect with withholding feed. Because this increase in drinking bouts is only observed within the control group, it can be assumed that this increase in drinking bouts is associated with the amount of time milk is withheld. A study found calves fed 5% of their body weight twice daily performed more suckling bouts than when fed 10% of their body weight (Jongman et al., 2020); further supporting the relationship between hunger and a greater number of drinking bouts at meal time. Notably, drinking bouts were greater after 12 h than after 16 h regardless of treatment, perhaps indicating there may be a ceiling effect from hunger effect on the number of drinking bouts.
The amount of time milk was withheld was associated with the number of non-drinking oral behavior bouts. Calves performed more non-drinking oral behaviors when feed was withheld for 12 and 16 h compared to 6 h. Previous research reported that non-nutritive sucking, a behavior grouped into non-drinking oral behaviors in this study, also increased with calf hunger (Herskin et al., 2010). As previously discussed, the non-drinking oral behaviors around feeding may indicate frustration, as seen in chickens (Rodenburg et al., 2002). Therefore, the increased non-drinking oral behaviors suggest that calves were hungrier and more frustrated when feed was withheld for longer periods. It is important to note that ‘longer’ periods include the 12 h interval, the time between meals on farms that feed twice daily. Most farms in the U.S. (94.6%) feed calves twice daily (USDA, 2016). Comparative hunger behaviors at 12 and 16 h without feed may suggest that calves should be fed more frequently than the current standard practice. We also observed that the dairy-beef crossbreeds expressed more non-drinking oral behaviors than Holstein calves, which may indicate that they were more susceptible to hunger than their purebred counterparts. This may be because dairy-beef crossbreeds have been shown to have greater growth rates and thus may have greater energy demands (Vestergaard et al., 2019; Berry, 2021).
As in Experiment 1, calves fed bitter milk expressed a greater number of aversive behaviors than calves fed unaltered milk. This further supports the idea that these behaviors are an aversive reaction to a bitter additive, reminiscent of gaping and lip smacking observed in rodents, primates, and sheep (Grill and Norgren, 1978; Steiner et al., 2001; Henslee et al., 2019). Data from this study is also the first to show that dairy calves perform behaviors which indicate feed aversion in response to a bitter flavor. However, milk withhold time was also associated with the number of aversive behaviors. Regardless of treatment, calves expressed fewer aversive behaviors after milk was withheld for 16 h compared to both 6 h and 12 h. This may be a result of increased hunger reducing the expression of these behaviors, but more research is needed to understand the relationship between these behaviors and hunger.
Study limitations
To the authors knowledge, this is the first test that utilized taste aversion to measure motivation to access feed in calves intended for field research. This test was intended to replace more labor-intensive behavioral observations that limit research on commercial farms. However, this study highlights the importance of additional behavioral observations to gain an accurate picture of calf hunger as the test is currently designed. The following are considerations and recommendations for future application of taste aversion to measure calf motivation for milk and areas where this motivation test may need further refinement for field application.
The concentration of the bitter additive or flavor profile utilized in this taste aversion motivation test may require further refinement. The quinine concentration used in Experiment 2 (0.35 g/L) may have been too high a stimulus to accurately measure changes in motivation to consume milk after it was withheld. This concentration was utilized due to the divergent consumption of milk at higher (0.4 g/L) vs. lower (≤ 0.3 g/L) quinine concentrations in Experiment 1. As with other behavioral assessments in animals, such as judgment bias, the sensitivity of animals to a reward or punishment needs to be carefully considered, as an incorrect stimulus may lead to false negative results (Kremer et al., 2022). Within motivation tests, animals are appraising the cost required versus their desire to obtain a resource (Jensen and Pedersen, 2008). In Experiment 2, the quinine concentration may have set the ‘cost’ too high to distinguish a change in motivation to consume milk after a 6-hour interval between meals compared to 16 hours. We suggest that this methodology should be applied to other flavors to test the efficacy of taste aversive as a model of calf motivation to drink milk. Acidic flavors may be worthwhile to explore, as cattle are able to taste these flavors and find them aversive (Ginane et al., 2011; Todd et al., 2018). Future applications of this taste aversion motivation test should consider calf sensitivity to the aversive flavor (i.e., bitter or acidic flavors) utilized and subsequently the concentration utilized.
Conclusion
This was the first experiment to our knowledge to test the impact of feeding a bitter additive in milk replacer to pre-weaned calves on their motivation to consume milk. Feeding calves bitter milk replacer resulted in less milk consumed than when calves were provided unaltered milk; though, this was only observed at relatively high concentrations of quinine (0.35 and 0.40 g/L) and not at 0.2 or 0.3 g/L. Calves performed more aversive behaviors including lip smacks, head bobs, and nostril licks at all concentrations of quinine compared to when calves consumed unaltered milk; suggesting that calves could taste a bitter flavor and found it to be aversive. In Experiment 2, bitter milk was not a successful tool for assessing calf motivation to drink because calves consumed less milk when it was bitter compared to unaltered milk at 6 h, 12 h, and 16 h after their previous meal with no effect of the milk withhold time. While this test did not observe an effect of milk withhold time on willingness to consume bitter milk, milk intake is only one measure of hunger. Withholding milk affected calf starter intake, drinking bouts, non-drinking oral behaviors, and aversive behaviors expressed by calves. Changes in feeding behaviors and calf starter consumption demonstrated that calves were likely hungrier when milk was withheld for 12 and 16 h, compared to 6 h. Future research should continue to develop taste aversion as a measure of motivation with different quinine concentrations or other flavors such as sour or acidic compounds to assess calf motivation for field research.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
MW: Formal Analysis, Resources, Writing – original draft, Methodology, Data curation, Visualization, Investigation. DB: Data curation, Writing – review & editing, Methodology. JC: Writing – review & editing, Data curation, Methodology, Investigation. KC: Data curation, Methodology, Conceptualization, Project administration, Supervision, Resources, Writing – review & editing, Investigation, Funding acquisition.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This study was funded by the University of Wisconsin-River Falls Undergraduate Research, Scholarly, and Creative Activity Undergraduate Stipends and Expenses Grant.
Acknowledgments
We would like to thank all the students and staff at the University of Wisconsin-River Falls for making this project possible including Hannah Olsen, Shawna Sigl, Andrea Clark, and Matthew Fischer.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Bernard R. A. (1964). An electrophysiological study of taste reception in peripheral nerves of the calf. Am. J. Physiology-Legacy. Content. 206, 827–835. doi: 10.1152/ajplegacy.1964.206.4.827
Berry D. P. (2021). Invited review: Beef-on-dairy—The generation of crossbred beef × dairy cattle. J. Dairy. Sci 104, 3789–3819. doi: 10.3168/jds.2020-19519
Borderas T. F., de Passillé A. M. B., and Rushen J. (2009). Feeding behavior of calves fed small or large amounts of milk. J. Dairy. Sci 92, 2843–2852. doi: 10.3168/jds.2008-1886
Churakov M., Eriksson H. K., Agenäs S., and Ferneborg S. (2023). Proposed methods for estimating loss of saleable milk in a cow-calf contact system with automatic milking. J. Dairy. Sci 106, 8835–8846. doi: 10.3168/jds.2022-23099
Congress, U. S (2023). 49 USC 80502: transportation of animals (United States Congress United States Statutes at Large, Washington D.C.).
Contreras R. J., Carson C. A., and Pierce C. E. (1995). A novel psychophysical procedure for bitter taste assessment in rats. Chem. Senses. 20, 305–312. doi: 10.1093/chemse/20.3.305
Costa J. H., Neave H. W., Weary D. M., and von Keyserlingk M. A. (2020). Use of a food neophobia test to characterize personality traits of dairy calves. Sci. Rep. 10, 1–11. doi: 10.1038/s41598-020-63930-8
Crabbe J. C. (2012). Translational behaviour-genetic studies of alcohol: are we there yet? Genes. Brain Behav. 11, 375–386. doi: 10.1111/j.1601-183X.2012.00798.x
Crabbe J. C. (2014). Rodent models of genetic contributions to motivation to abuse alcohol. Nebraska. Symp. Motiv. 61, 5–29. doi: 10.1007/978-1-4939-0653-6_2
Curtis G., McGregor Argo C., Jones D., and Grove-White D. (2018). The impact of early life nutrition and housing on growth and reproduction in dairy cattle. PloS One 13, e0191687. doi: 10.1371/journal.pone.0191687
D'Eath R. B., Tolkamp B. J., Kyriazakis I., and Lawrence A. B. (2009). ‘Freedom from hunger’ and preventing obesity: the animal welfare implications of reducing food quantity or quality. Anim. Behav. 77, 275–288. doi: 10.1016/j.anbehav.2008.10.028
Dawkins M. S. (1990). From an animal's point of view: Motivation, fitness, and animal welfare. Behav. Brain Sci. 13, 1–9. doi: 10.1017/S0140525X00077104
De Passillé A., Borderas T., and Rushen J. (2011). Weaning age of calves fed a high milk allowance by automated feeders: Effects on feed, water, and energy intake, behavioral signs of hunger, and weight gains. J. Dairy. Sci 94, 1401–1408. doi: 10.3168/jds.2010-3441
de Passillé A. M. B. and Rushen J. (2006). Calves’ behaviour during nursing is affected by feeding motivation and milk availability. Appl. Anim. Behav. Sci 101, 264–275. doi: 10.1016/j.applanim.2006.02.007
De Paula Vieira A., Guesdon V., de Passillé A. M., von Keyserlingk M. A. G., and Weary D. M. (2008). Behavioural indicators of hunger in dairy calves. Appl. Anim. Behav. Sci 109, 180–189. doi: 10.1016/j.applanim.2007.03.006
Eckert E., Brown H., Leslie K., DeVries T., and Steele M. (2015). Weaning age affects growth, feed intake, gastrointestinal development, and behavior in Holstein calves fed an elevated plane of nutrition during the preweaning stage. J. Dairy. Sci 98, 6315–6326. doi: 10.3168/jds.2014-9062
Franchi G. A., Herskin M. S., Tucker C. B., Larsen M., and Jensen M. B. (2021). Assessing effects of dietary and milking frequency changes and injection of cabergoline during dry-off on hunger in dairy cows using 2 feed-thwarting tests. J. Dairy. Sci 104, 10203–10216. doi: 10.3168/jds.2020-20046
Friard O. and Gamba M. (2016). BORIS: a free, versatile open-source event-logging software for video/audio coding and live observations. Methods Ecol. Evol. 7, 1325–1330. doi: 10.1111/2041-210X.12584
Ginane C., Baumont R., and Favreau-Peigné A. (2011). Perception and hedonic value of basic tastes in domestic ruminants. Physiol. Behav. 104, 666–674. doi: 10.1016/j.physbeh.2011.07.011
Goatcher W. and Church D. (1970). Taste responses in ruminants. IV. Reactions of pygmy goats, normal goats, sheep and cattle to acetic acid and quinine hydrochloride. J. Anim. Sci 31, 373–382. doi: 10.2527/jas1970.312373x
Grill H. J. and Norgren R. (1978). The taste reactivity test. I. Mimetic responses to gustatory stimuli in neurologically normal rats. Brain Res. 143, 263–279. doi: 10.1016/0006-8993(78)90568-1
Henslee D., Yelich J., Taylor J. B., and Ellison M. (2019). Avoidance of phenylthiocarbamide in mature Targhee and Rambouillet rams. Trans. Anim. Sci 3, 1194–1204. doi: 10.1093/tas/txz125
Herskin M. S., Skjøth F., and Jensen M. B. (2010). Effects of hunger level and tube diameter on the feeding behavior of teat-fed dairy calves. J. Dairy. Sci 93, 2053–2059. doi: 10.3168/jds.2009-2554
Hopf F. W., Chang S. J., Sparta D. R., Bowers M. S., and Bonci A. (2010). Motivation for alcohol becomes resistant to quinine adulteration after 3 to 4 months of intermittent alcohol self-administration. Alcohol.: Clin. Exp. Res. 34, 1565–1573. doi: 10.1111/j.1530-0277.2010.01241.x
Jensen M. B. and Pedersen L. J. (2008). Using motivation tests to assess ethological needs and preferences. Appl. Anim. Behav. Sci 113, 340–356. doi: 10.1016/j.applanim.2008.02.001
Jongman E. C., Conley M. J., Borg S., Butler K. L., and Fisher A. D. (2020). The effect of milk quantity and feeding frequency on calf growth and behaviour. Anim. Prod. Sci 60, 944–952. doi: 10.1071/AN19049
Khan M. A., Weary D. M., and von Keyserlingk M. A. G. (2011). Invited review: Effects of milk ration on solid feed intake, weaning, and performance in dairy heifers. J. Dairy. Sci 94, 1071–1081. doi: 10.3168/jds.2010-3733
Kirkden R. D., Edwards J. S. S., and Broom D. M. (2003). A theoretical comparison of the consumer surplus and the elasticities of demand as measures of motivational strength. Anim. Behav. 65, 157–178. doi: 10.1006/anbe.2002.2035
Kirkden R. D. and Pajor E. A. (2006). Using preference, motivation and aversion tests to ask scientific questions about animals’ feelings. Appl. Anim. Behav. Sci 100, 29–47. doi: 10.1016/j.applanim.2006.04.009
Kour H., Corbet N. J., Patison K. P., and Swain D. L. (2021). Changes in the suckling behaviour of beef calves at 1 month and 4 months of age and effect on cow production variables. Appl. Anim. Behav. Sci 236, 105219. doi: 10.1016/j.applanim.2021.105219
Kremer L., van Reenen C. G., Engel B., Bokkers E. A. M., Schnabel S. K., van der Werf J. T. N., et al. (2022). Developing a feasible and sensitive judgement bias task in dairy cows. Anim. Cogn. 25, 425–445. doi: 10.1007/s10071-021-01563-8
Ladewig J., Sørensen D. B., Nielsen P. P., and Matthews L. R. (2002). The quantitative measurement of motivation: generation of demand functions under open versus closed economies. Appl. Anim. Behav. Sci 79, 325–331. doi: 10.1016/S0168-1591(02)00156-9
Lecorps B., Woodroffe R. E., von Keyserlingk M. A., and Weary D. M. (2023). Hunger affects cognitive performance of dairy calves. Biol. Lett. 19, 20220475. doi: 10.1098/rsbl.2022.0475
Lesscher H. M., Van Kerkhof L. W., and Vanderschuren L. J. (2010). Inflexible and indifferent alcohol drinking in male mice. Alcohol.: Clin. Exp. Res. 34, 1219–1225. doi: 10.1111/j.1530-0277.2010.01199.x
MacPherson J., Meale S. J., Macmillan K., Haisan J., Bench C. J., Oba M., et al. (2019). Effects of feeding frequency of an elevated plane of milk replacer and calf age on behavior, and glucose and insulin kinetics in male Holstein calves. Animal 13, 1385–1393. doi: 10.1017/S175173111800294X
Morita S., Sugita S., Yamamoto M., Hoshiba S., and Uemura T. (1999). Behavioral investigation of group rearing calves in automatic milk replacer feeding system. J. Japanese. Soc. Anim. Sci 70, 542–546. doi: 10.2508/chikusan.70.542
National Academies of Sciences, E., and Medicine (2021). Nutrient Requirements of Dairy Cattle: Eighth Revised Edition (Washington, D.C: The National Academic Press).
Neave H. W., Costa J. H., Benetton J., Weary D. M., and von Keyserlingk M. A. (2019). Individual characteristics in early life relate to variability in weaning age, feeding behavior, and weight gain of dairy calves automatically weaned based on solid feed intake. J. Dairy. Sci 102 (11, 10250–10265. doi: 10.3168/jds.2019-16438
Occhiuto F., Vázquez-Diosdado J. A., King A. J., and Kaler J. (2023). Evidence of personality-dependent plasticity in dairy calf movement behaviours derived from automated data collection. Sci. Rep. 13, 18243. doi: 10.1038/s41598-023-44957-z
Rodenburg T. B., Zimmerman P. H., and Koene P. (2002). Reaction to frustration in high and low feather pecking laying hens. Behav. Proc. 59, 121–129. doi: 10.1016/S0376-6357(02)00086-4
Rosenberger K., Costa J., Neave H., Von Keyserlingk M., and Weary D. (2017). The effect of milk allowance on behavior and weight gains in dairy calves. J. Dairy. Sci 100, 504–512. doi: 10.3168/jds.2016-11195
Steele M. A., Doelman J. H., Leal L. N., Soberon F., Carson M., and Metcalf J. A. (2017). Abrupt weaning reduces postweaning growth and is associated with alterations in gastrointestinal markers of development in dairy calves fed an elevated plane of nutrition during the preweaning period. J. Dairy. Sci 100, 5390–5399. doi: 10.3168/jds.2016-12310
Steiner J. E., Glaser D., Hawilo M. E., and Berridge K. C. (2001). Comparative expression of hedonic impact: affective reactions to taste by human infants and other primates. Neurosci. Biobehav. Rev. 25, 53–74. doi: 10.1016/S0149-7634(00)00051-8
Terré M., Tortadès M., Genís S., Cresci R., Frongia A., Verdú M., et al. (2022). Short communication: A milk replacer aversion model in calves to test flavour-masking effects. Livestock. Sci 256, 104830. doi: 10.1016/j.livsci.2022.104830
Todd C. G., Millman S. T., Leslie K. E., Anderson N. G., Sargeant J. M., and DeVries T. J. (2018). Effects of milk replacer acidification and free-access feeding on early life feeding, oral, and lying behavior of dairy calves. J. Dairy. Sci 101, 8236–8247. doi: 10.3168/jds.2018-14487
USDA (2016). Dairy Cattle Management Practices in the United Stated 2014 Vol. 692. Ed. Fort Collins C. O. (USDA–APHIS–VS–CEAH–NAHMS).
Vestergaard M., Jørgensen K., Çakmakçı C., Kargo M., Therkildsen M., Munk A., et al. (2019). Performance and carcass quality of crossbred beef x Holstein bull and heifer calves in comparison with purebred Holstein bull calves slaughtered at 17 months of age in an organic production system. Livestock. Sci 223, 184–192. doi: 10.1016/j.livsci.2019.03.018
Wolffgramm J., Galli G., Thimm F., and Heyne A. (2000). Animal models of addiction: models for therapeutic strategies? J. Neural Transm. 107, 649–668. doi: 10.1007/s007020070067
Woodrum Setser M. M., Neave H. W., and Costa J. H. C. (2024a). Are you ready for a challenge? Personality traits influence dairy calves' responses to disease, pain, and nutritional challenges. J. Dairy. Sci. doi: 10.3168/jds.2023-24514
Woodrum Setser M. M., Neave H. W., and Costa J. H. C. (2024b). Can individual variation in food neophobia explain variability in dairy calf feeding behavior patterns and performance? Appl. Anim. Behav. Sci 274, 106266. doi: 10.1016/j.applanim.2024.106266
Keywords: aversion test, feed withholding, hunger motivation, quinine, welfare
Citation: Woodrum Setser M, Brown D, Costa JHC and Creutzinger K (2025) Taste aversion as a motivation test of hunger in dairy calves. Front. Anim. Sci. 6:1602011. doi: 10.3389/fanim.2025.1602011
Received: 28 March 2025; Accepted: 30 September 2025;
Published: 24 October 2025.
Edited by:
Leonie Jacobs, Virginia Tech, United StatesReviewed by:
Francisco Ceacero, Czech University of Life Sciences Prague, CzechiaThomas Ede, University of Pennsylvania, United States
Ghazanfar Ali Chishti, University of Veterinary and Animal Sciences, Pakistan
Copyright © 2025 Woodrum Setser, Brown, Costa and Creutzinger. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Katherine Creutzinger, a2F0ZS5jcmV1dHppbmdlckB1dm0uZWR1
†Present address: Megan Woodrum Setser, Department of Animal Science, University of Connecticut, Storrs, CT, United States