 Bjarne Gjerde1*‡
Bjarne Gjerde1*‡ Tale Marie Karlsson-Drangsholt1†
Tale Marie Karlsson-Drangsholt1† Jørgen Ødegård1†Frode Finne-Fridell2Ingunn Thorland3
Jørgen Ødegård1†Frode Finne-Fridell2Ingunn Thorland3 Kirsti Hjelde4
Kirsti Hjelde4- 1Department of Breeding and Genetics, Nofima AS, Ås, Norway
- 2PHARMAQ AS part of Zoetis, Oslo, Norway
- 3Benchmark Genetics Norway, Sunndalsøra, Norway
- 4Feed Development and Nutrition, Nofima AS, Sunndalsøra, Norway
Vaccination is an effective tool to prevent disease outbreaks in Atlantic salmon. However, oil-adjuvant vaccines, which may lead to negative side effects such as adhesions of abdominal organs and melanin deposits on abdominal organs and the abdominal wall, have been linked to vertebral deformities. The main objective of this study was to estimate both the genetic and environmental (residual) correlations between vaccine-induced side effects and vertebral deformities and the genetic correlation between vertebral deformities measured at different ages. The recorded fish were the offspring of 85 sires and 150 dams from SalmoBreed (year-class 2007) and included two groups of vaccinated fish: pre-smolts exposed to 17°C freshwater for 3 months (FW) and a group of post-smolts (SW) evaluated after 6 (SW6) and 12 (SW12) months in a net cage in the sea. Adhesions of abdominal organs and viscera (score 0–6) and melanin deposits on abdominal organs and the abdominal wall (score 0–3) were evaluated by trained personnel, and vertebral deformities were recorded using radiography. The frequency of vertebral deformities was 6.4%, 5.2%, and 6.4% for the FW, SW6, and SW12 groups, respectively. Fused vertebrae were the most common type of deformity in all groups, with fused vertebrae (4.2%) in the FW group and a low percentage of missing intervertebral space in the SW6 (0.5%) and SW12 (0.6%) groups. The mean adhesion scores for the FW, SW6, and SW12 groups were 2.06, 1.68, and 1.46, respectively, while the mean melanin scores were 0.95, 1.49, and 1.45. The heritability on the liability scale for fused vertebrae was of medium magnitude for the FW (0.28 ± 0.11) group as well as for the SW6 (0.20 ± 0.10) and the SW12 (0.20 ± 0.10) groups. Across the experimental groups, the genetic and residual correlations of fused vertebrae with adhesion and melanin scores were all close to and not significantly different from zero. Therefore, no evidence was found of a genetic or environmental association between the negative side effects of vaccination (adhesions and melanin deposits) and fused vertebrae in Atlantic salmon.
1 Introduction
Vaccination is an effective tool to prevent disease outbreaks in the aquaculture industry (Sommerset et al., 2005; Ramstad et al., 2007; Brudeseth et al., 2013). In Atlantic salmon (Salmo salar), oil-adjuvanted vaccines have been found to cause negative side effects like abdominal adhesions and melanin deposits (Midtlyng et al., 1996), which are found to develop over time (Mutoloki et al., 2004). Their severity may be caused by several factors like vaccine formulation, temperature, and fish size at vaccination as well as genetics (Gudding et al., 1999; Gjerde et al., 2005; Berg et al., 2006, 2007; Drangsholt et al., 2011). Focal dark melanin spots in Atlantic salmon have also been associated with rib abnormalities, as they are predominantly located at the cranio-ventral region of the abdominal wall (Jiménez-Guerrero et al., 2022), as well as to a sequence of events with the pivotal role of adiposity, hypoxia, and fat necrosis (Bjørgen et al., 2024).
Based on an online survey to the processing industry in Norway, Germany, and Poland, melanin spots together with pale fillet color represent the most significant quality challenge in Norwegian salmon farming with an estimated prevalence of approximately 16.4%, with vaccine and stress looked upon as the major causes to the problem, and with an average economic loss from 8% to 44% depending on the size of the melanin spots (Färber, 2017). Similar estimates of prevalence and economic loss due to deformities have not been found.
Vertebral deformities occur frequently in farmed Atlantic salmon (see review by Fjelldal et al., 2012) and can be affected by a number of factors like temperature during incubation (Takle et al., 2005), minerals in feed (Helland et al., 2005; Fjelldal et al., 2009; Baeverfjord et al., 2018), and genetics (McKay and Gjerde, 1986; Gjerde et al., 2005). Vertebral deformities are potentially also a side effect of vaccination as their frequency has been reported to be higher in vaccinated than in unvaccinated fish (Berg et al., 2006) and in fish vaccinated with different vaccines (Thorarinsson et al., 2023). A relatively strong positive association between vaccine-induced intra-abdominal lesions and prevalence of deformities has also been reported (Berg et al., 2006; Aunsmo et al., 2008). The likely reasons for such an association may be an additional direct side effect of the vaccine (such as adhesions and melanin deposits), an indirect effect induced by the abdominal vaccine side effects, or due to a correlation, environmentally and/or genetically, between deformity and side effects of the vaccine.
Some vertebral deformities are reported to develop over time, and both aggravation and stabilization of fused vertebrae have been observed (Witten et al., 2006; Fjelldal et al., 2007; Drábiková et al., 2022). Animals with low-mineralized vertebrae in freshwater caused by low dietary phosphorus have been found to recover in seawater and with no significant effect on the prevalence of deformities at harvest, whereas progressive vertebral fusions arising in freshwater that affect more than three vertebral bodies worsen over time (Drábiková et al., 2022).
For genetic variation in adhesion and melanin deposits, there are few reported estimates. In two year-classes of Atlantic salmon that were vaccinated intraperitoneally as pre-smolts (one year-class with Norvax Protect, the other with Biojec 1900, both with 0.2 mL/fish), Gjerde et al. (2009) reported significant genetic variation for adhesions (score 0 to 6) and melanin deposits (score 0 to 3) with a combined heritability estimate of 0.19 ± 0.03 for adhesions and 0.18 ± 0.03 for melanin deposits recorded after 17 and 13 months in a net cage in the sea, respectively. Similarly, in a year-class of Atlantic salmon that was vaccinated intraperitoneally as pre-smolts with a six-component vaccine, significant genetic variation was observed in adhesions (score 0 to 6) and melanin deposits (score 0 to 3) both recorded after 6 (heritability 0.19 ± 0.04 for adhesions and 0.28 ± 0.05 for melanin deposits) and 12 (0.16 ± 0.04 for adhesions and 0.30 ± 0.05 for melanin deposits) months in a net cage in the sea.
For vertebral deformities, the reported estimates of genetic variation are from one paper only (Gjerde et al., 2005). They scored Atlantic salmon externally for deformity (0 to 5) after 14–17 months in a net cage in the sea and found substantial genetic variation in four out of the five year-classes (heritability 0.22 ± 0.09, 0.25 ± 0.12, 0.36 ± 0.14, and 0.00 ± 0.00).
Consequently, the traits adhesions, melanin deposits, and vertebral deformities have all been reported to show genetic variation and have also been linked to vaccination. Berg et al. (2006) reported a strong negative phenotypic correlation (r2 = 0.85) between body size at vaccination and incidence of deformities. However, no estimate of the genetic correlation of deformity traits with growth or traits related to the negative side effects of vaccination is available. The primary objective of this study was therefore to obtain reliable estimates of the magnitude of both the genetic and environmental correlations of vertebral deformity traits with traits possibly associated with vaccine-induced side effects in farmed Atlantic salmon.
2 Materials and methods
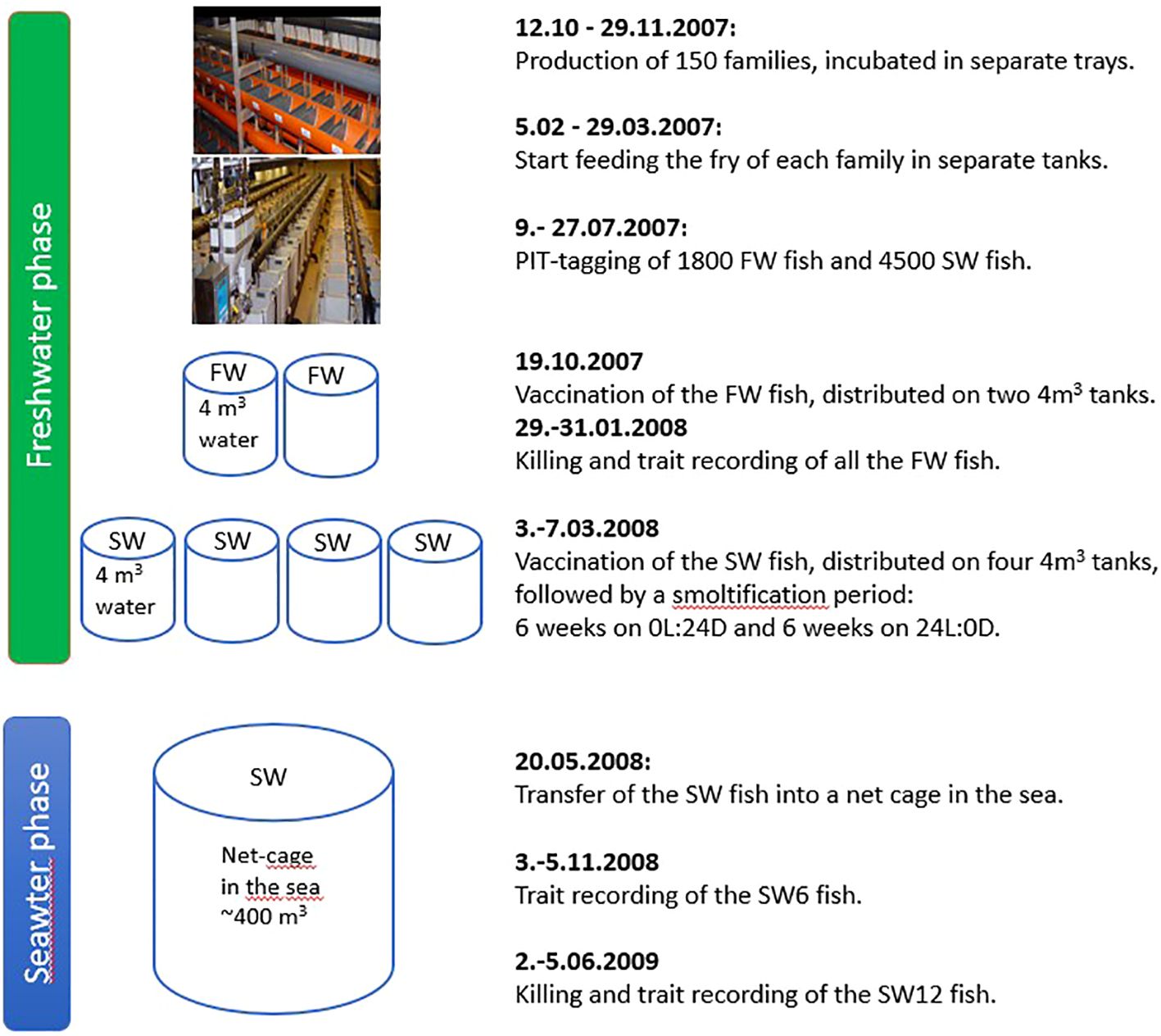
An outline of the experimental design with dates of the different activities and trait recordings is shown in Appendix 1.
2.1 Fish
The Atlantic salmon in this study were from 150 full-sib families, i.e., the offspring of 85 sires and 150 dams from the breeding nucleus of SalmoBreed AS (now Benchmark Genetics Norway). The parent fish had been selected for increased growth, reduced proportion of early sexual maturing fish, normal body shape and reduced incidence of deformities for several generations, increased fillet color, reduced fillet fat content, increased resistance to furunculosis (Aeromonas salmonicida) and infectious salmon anemia (ISA), increased cage survival, and increased yield for one generation.
Each sire was mated to one to three dams, and each dam to one sire only. The families were produced on 10 different dates from the 12th of October to the 29th of November 2006 at two different sites in southwest Norway: Bolaks AS, Eikelandsosen (28 families), and Erfjord Stamfisk AS (now Grieg Seafood Rogaland), Erfjord (122 families). The families were incubated in separate trays at Bolaks at a water temperature varying from 5.2°C to 8.0°C, on average 7.7°CC, or in separate compartments in a cylinder at Erfjord Stamfisk at a water temperature varying from 3.0°C to 8.0°C, on average 7.1°C. In December 2006 and January 2007, eyed-eggs were transported to Nofima, Sunndalsøra, where they were reared further in separate trays at an average water temperature varying from 6.5°C to 7.3°C.
The first feeding of the families took place from the 5th of February to the 29th of March 2007 in separate 0.1 m3 tanks. All tanks were supplied with water from the same source and with an average temperature of 10.8°C (varying from 9.8°C to 13.7°C) from the 5th of February to the 30th of April. During the following 3 months until PIT tagging, the average temperature in the tanks was 13.2°C (varying from 12.0°C to 14.2°C). From first feeding, the fish were kept on a 24L:0D light regime until the start of the smoltification regime.
2.2 Two experimental groups
There were two experimental groups of vaccinated fish randomly sampled from the same 150 families: the FW group from which the records of the studied traits were obtained after 3 months on an elevated temperature (17°C) in freshwater (FW) and the SW group from which records of the studied traits of a sample of the fish were obtained after 6 months (SW6) and of the remaining fish after 12 months (SW12) in a net cage in the sea, thus resulting in a total of three experimental groups for the analyses of the recorded data.
The FW and SW groups were individually tagged over a 3-week period from the 9th to the 27th of July 2007. For the FW group, a total of 1,800 fish were tagged (a random sample of 12 per family) with an average body weight of 11.5 g (SD 3.5), while for the SW group, a total of 4,500 fish were tagged (a random sample of 30 per family) with an average body weight of 11.5 g (SD 3.4).
In the freshwater phase, the oxygen saturation level of the water in the rearing tanks was kept >80%, which was monitored twice a week. Both in the freshwater phase and the seawater phase, the fish were fed a commercial dry pelleted feed from Skretting AS.
2.2.1 The vaccine
The FW (see 2.2.2) and SW (see 2.2.3) groups received a commercial six-component vaccine (ALPHA JECT® 6–2 produced by PHARMAQ AS, Oslo, Norway) with an injection volume of 0.1 mL per fish of an oil-in-water-adjuvant (mineral oil) emulsion containing antigens of Aeromonas salmonicida, Vibrio anguillarum, Vibrio salmonicida, Moritella viscosa, and infectious pancreatic necrosis virus (IPNV), which protects against furunculosis, classical vibriosis, cold water vibriosis, winter ulcer, and IPN, respectively.
2.2.2 The FW group
The FW group was kept in a separate ~4 m3 water volume tank with freshwater (average 11.6°C, varying from 8° to 13°) until the fish were vaccinated on the 19th of October 2007. On the 28th of October 2007, the fish were randomly distributed into two replicated tanks, each with ~6 m3 of freshwater, which over 12 days was gradually increased to 17°C and kept at approximately this temperature for 3 months (see Appendix 2). This elevated temperature regime had previously been shown to be a stable laboratory model to obtain the worst-case scenario with respect to vaccine-induced side effects observed, such as adhesions of organs in the abdominal cavity and melanin spots on internal organs and the abdominal wall (PHARMAQ AS, pers. comm.). Unfortunately, there were several short-lasting drops in the water temperature due to technical problems with the water pump (Appendix 2).
2.2.3 The SW group
The SW group was kept in a common tank with freshwater (water volume of ~10 m3, average 9.4°C, SD = 2.3, varying from 6.5°C to 13.6°C) until the fish were vaccinated on the 3rd to the 7th of March 2008 and distributed on four tanks each with ~4 m3 of water and, therefore, on average, approximately 58 kg/m3 (including approximately 1,800 fish that were vaccinated with a reduced 0.05 mL/fish injection volume, but not recorded for deformity and therefore not included in this study; see Drangsholt et al., 2011). Prior to their transfer to seawater, the fish were smoltified for 6 weeks (269 day-degrees) on a 0L:24D light regime followed by 6 weeks (287 day-degrees) on a 24L:0D light regime.
On the 20th of May 2008, two and a half months (460 day-degrees) after vaccination, the SW fish were transferred to a common net cage (400 m3) in the sea at Nofima Averøy, Norway, where they were reared until the studied trait records were obtained on a random sample of the fish in early November 2008 (SW6) and of the remaining fish in early June 2009 (SW12).
At the November 2008 sampling, the biomass in the cage was ~15 kg/m3 (including approximately 1,800 fish mentioned above), and at the June 2009 slaughter, approximately 10 kg/m3, both well below the recommended biomass (<25 kg/m3) to secure the fish sufficient access to oxygen under the seawater current levels at the Averøy locality. The average weekly seawater temperature during the 12-month rearing period at Averøy is shown in Appendix 3.
2.3 Data recording
The FW fish in the two replicated tanks were all killed and recorded for the studied traits on the 29th to the 31st of January 2008. A random sample of the SW fish within each family (a maximum of 12 fish per family) was recorded on the 3rd to the 5th of November 2008 (8 months after vaccination, 6 months after sea transfer), while the remaining SW fish were recorded on the 2nd to the 5th of June 2009 (15 months after vaccination, 12 months after their transfer to the net cage in the sea).
At each of the three recording samplings or slaughter events (of the FW, SW6, and SW12 fish), a dip net was used to catch fish from the tanks or the cage. At the FW and SW6 samplings, the fish were anesthetized with ethyl 3-aminobenzoate methanesulfonate (FINQUEL®, SCANAQUA), while at the SW12 sampling, the fish were anesthetized with CO2 gas before being killed by cutting the gills and bled in running water. For each fish, their ID, body weight, and sex (male, female, or unknown based on gonad inspection after gutting the fish) were recorded. Furthermore, the fish were examined for vaccine-induced side effects and vertebral deformities as described below. Sexually maturing or mature fish were excluded from the data (in total, 21 fish at the FW sampling, 12 at the SW6 sampling, and none at the SW12 sampling).
2.3.1 Evaluation of vaccine-induced side effects
Adhesions of internal organs in three separate regions of the abdominal cavity (the anterior, dorsal and posterior, and ventral part of the abdominal cavity, see Figure 1) were subjectively scored using a scale (Speilberg Skala) from 0 to 6 with 0.5 score intervals (0 = no adhesions, 6 = extremely severe adhesions including granuloma; Table 1 in Midtlyng et al., 1996). Melanin deposits on internal organs and abdominal wall were scored using a four-point scale from 0 to 3 (0 = no visible melanin, 1 = some faint melanin on some organs and/or peritoneum, 2 = moderate amount of melanin on some organs or on the peritoneum that may cause downgrading of the final product, 3 = severe melanin spots on the viscera or the peritoneum resulting in significant downgrading of the final product).
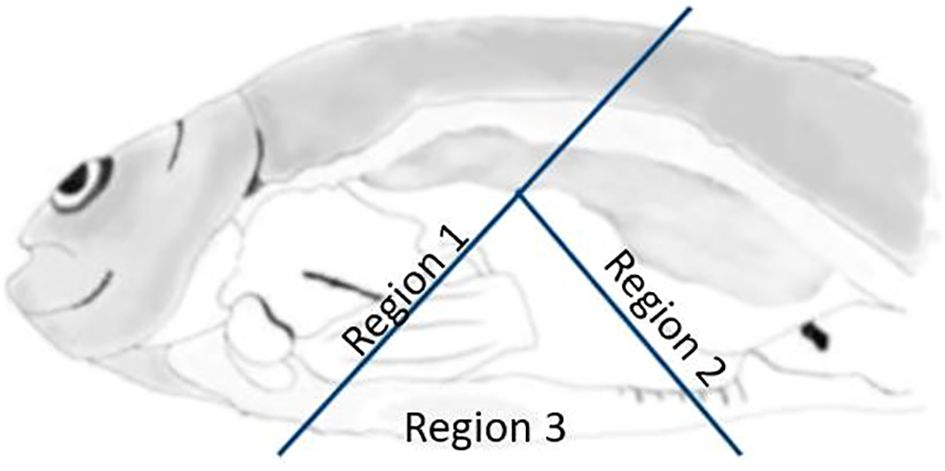
Figure 1. The three separate regions of the abdominal cavity scored for adhesions of organs and viscera. Region 1: anterior and anterior-dorsal of the cavity including the esophagus, liver, and anterior parts of the swim bladder; region 2: posterior and posterior-dorsal parts of the cavity including the hind gut; and region 3: ventral region of the cavity (PHARMAQ AS).
The scored traits were evaluated by two (for the FW and the SW12 groups) or four (for the SW6 group) trained persons from PHARMAQ. Two of the total four persons scored fish at all three samplings, while the other two persons took part only at the SW6 sampling. Within each of the three samplings, each fish was evaluated by one person only. At the time of scoring, information about the experimental group and family was not known to the scoring personnel.
2.3.2 Radiography
Evaluation of radiographs was used to score each fish for vertebral deformities. For the FW group, this was obtained using a high-resolution semi-digital system (IMS Giotto mammography X-ray source, serial number 61/01/08 2005/03, Model number 6020/3, dosage 22 kV and 100 mAs). The image reader was an FCR Profect delivered from Fuji Medical, linked to a computer (Fuji CR Console) with digital software delivered from Fujifilm. This allowed us to pre-set the best possible adjustments for each fish group to get images of high quality for our needs. The resolution of the image is 20 pixels/mm2 and there were one to six fish in each image, depending on their size. Images were stored full size in the original DICOM format.
The images of fish in the SW6 and SW12 groups were obtained by a transportable radiation source (Sedecal portable X-ray unit, dosage 40 kV and 40 mAs). The image plate reader was a Fuji FCR XG1, the computer a Fuji CR Console, and the image resolution 10 pixels/mm2, and there were one to four fish in each image, depending on their size.
The image of the vertebrae of each fish was subjectively scored by a trained veterinarian (the last author of the paper) using a diagnostic, high-resolution Eizo LCD screen. The four main diagnostic categories in the material were non-deformed spine, fused vertebrae, “missing intervertebral space,” and hyperdense vertebrae.
The number of affected or deformed vertebrae and the type of deformity were recorded for each fish. The number of affected fish with more than two fused vertebrae was very low, which made it impossible to explore the magnitude of genetic variation in the number or severity of fused vertebrae in this study. Consequently, each of the three types of deformity categories was defined as a binary trait (1 = fused, missing intervertebral space or hyperdense; 0 = no deformed vertebra). In addition, a binary trait called deformity was defined as 1 = one or more vertebrae with any deformity (fused, missing intervertebral space) and 0 = no deformed vertebrae of any type. The term deformity in this setting includes even minor vertebral deviations that would not be considered a deformity on the slaughter line or be detectable by external evaluation.
2.4 Statistical analyses
Estimates of (co)variances for the random effects of the studied traits were obtained using a multitrait animal model or a multitrait sire and dam threshold model and the ASReml software (Gilmour et al., 2009). The two binary traits, missing intervertebral space and hyperdense vertebrae, were not included in these analyses due to their very low frequency in all three experimental groups.
The animal model, with a separate analysis for each of the three experimental groups as well as across these groups, included the five traits deformity (Def, i.e., deformity or fused vertebrae, one at a time), adhesions of organs (Adh), melanin deposits on organs and abdominal wall (Mel), whole body weight at vaccination (Bwv), and sampling (Bws) and can be written in matrix notation as (model 1):
where
yDef,yAdh, yMel, yBwv, and yBws are vectors of observations for the abovementioned traits; βDef, βAdh, βMel, βBwv, and βBws are vectors of the combined fixed effects of tank-sex for the Def, Bwv, and Bws traits of the FW data; tank-sex-person for the Adh and Mel traits of the FW data; sex for the Def and Bwv and Bws traits of the SW data; and sex-person for the Adh and Mel traits of the SW data and of the regression coefficient of the actual trait on the age of the fish (from first feeding); X is the incidence matrix for the fixed effects; Z is the incidence matrix for the random animal additive genetic effect; Wis the incidence matrix for the random effect common to full sibs; is a vector of animal additive genetic effects; cBwvand eBws are vectors of the random effects common to full sibs for the two body weight traits; and eDef, eAdh, eMel, eBwv, and eBwv are vectors of the random residual for each of the five traits.
Further , where u and e are as defined above; A is the additive genetic relationship matrix among the animals and their parents, I is an identity matrix of appropriate size, G is the additive genetic (co)variance matrix, and R is the residual variance–covariance matrix among the traits.
Drangsholt et al. (2011), using the same data as in this study, found high positive genetic correlations between melanin score on organs and abdominal wall (on average 0.84; range 0.68–0.99) and high positive genetic correlations between adhesion scores in the three regions of the abdominal cavity (on average 0.78; range 0.56–0.99). Therefore, for the trait melanin score and adhesion score for each fish in this study, the average melanin score (across organs and abdominal wall) and the average adhesion score (across the three regions) were used.
Sex had three levels: males, females, or unknown. The unknown were fish in the FW (159/1,631 = 9.7%) and SW12 (74/1,565 = 4.7%) groups for which sex could not be determined upon inspection of the gonads after being gutted, and for some surplus randomly sampled SW6 fish (325/1,897 = 17.1%) for which only their whole body weights were recorded and were left for rearing for another 6 months until the SW12 recording.
The effect common to full sibs was only included for the body weight traits. When this effect was included for the other traits, the log-likelihood of the model did not converge. Consequently, this effect had to be omitted from the model for these traits.
Estimates of genetic correlations between the same deformity trait (deformity or fused vertebrae, one at a time), recorded at each sampling (FW, SW6, and SW12), were obtained from a multitrait sire and dam threshold model assuming a normally distributed underlying liability variable, lijk, which determines the outcome of the binary trait, such that lijk ≤ 0 corresponds with Yijk = 0, and lijk > 0 corresponds with Yijk = 1. In matrix notation, the model for each trait can be written as (model 2):
where k represents an offspring of sire i and dam j; is the cumulative standard normal distribution, Zs and Zd are the incidence matrices for the random effect of sire and dam, respectively; X is the incidence matrix defined in model 1; usd is the random effect of sire or dam; and the other parameters are as described above. The effect common to full sibs was omitted from the model for all traits, as the log-likelihood of the model did not converge. The estimated variance component for sires was set equal to the variance component for dams, and equal to one quarter of the additive genetic variance (); and where the residual variance of l, , was set equal to 1.0. The heritability for each trait was calculated as:
3 Results
3.1 The four scored spine categories
Figure 2 shows a radiographic image of four fish, each with one of the four scored vertebral categories.

Figure 2. Radiographic images of (a) normal spine (fish of 3,480 g, SW12 group), (b) fused vertebrae (fish of 2,605 g from the SW12 group), (c) hyperdense vertebrae (fish of 143 g, FW group), and (d) missing intervertebral space (fish of 1,700 g, SW12 group). Numbers denote the vertebral number counted from the cranium.
Fused vertebrae include all proper fusions and incomplete fusions that we knew from experience would develop into complete fusions as the fish grows (Witten et al., 2006) (Figure 2B). Some fused vertebrae will stabilize and not cause problems, but radiography is not sensitive enough as a tool for deciding which processes are finished and which are still active. In this material, we chose not to include the last four tail vertebrae because they have a large normal variation in morphology due to tail development (Kryvi et al., 2020). This includes twin vertebral units, a single vertebra with two neural or hemal spines that were never separated in the notochord stage. Based on our observations and experience from this and earlier materials, these twin vertebral units seem to be of little or no clinical importance to the fish. Vertebral deformities are of less clinical importance in the tail area and are hard to separate from the normal variation in their morphology, and hence are not included in the material.
Hyperdense vertebrae (Helland et al., 2006; Baeverfjord et al., 2018) are seen in smaller fish, seldom after sea transfer, and linked to low phosphorus levels. They are single vertebrae with increased radiodensity, appearing more radiopaque than the others on radiographs (Figure 2C). Sometimes, they affect their neighboring vertebrae, leading to higher radiopacity in the vertebral end-plate closest to the affected vertebrae. Some of these hyperdense vertebrae normalize as the fish grows, while others develop into fused vertebrae (Baeverfjord et al., 2009).
Missing intervertebral space (type 1 deformity described by Witten et al., 2009) is a diagnosis seen in, for example, 3–5 kg fish (Holm et al., 2020), and with a less-known etiology since there are few deformity trials from the grow-out phase in the sea. In missing intervertebral space, the vertebrae appear to lose their intervertebral space, and sometimes, a dorsoventral shift in position is observed.
Diagnosis of vertebral defects is still not classified properly due to a lack of knowledge on causes and possible correlations between different types of deformities. Therefore, a descriptive term was chosen in the classification of the observed changes.
Relatively similar deformities classified as platyspondyly (Kvellestad et al., 2000), decreased intervertebral space (Witten et al., 2009), compressions (Witten et al., 2009; Baeverfjord et al., 2018), or cross-stitch vertebrae (Holm et al., 2020; Thorarinsson et al., 2023) may be related to or the same as the deformity observed in this material (Figure 2D), with the early stages of “cross-stitch vertebrae” being the ones appearing most similar. Classification of vertebral deformities is complicated by pathology that overlaps or is very similar but with different causes and at different developmental stages, so a differentiation into major groups is necessary to draw relevant information out of the material.
3.2 Descriptive statistics
In the FW group, 9.4% of the fish died during the 3 months from tagging (mid-July 2007) to vaccination (mid-October 2008). In the SW group, 11.5% of the fish died during the 7 months from tagging to vaccination, 19.0% died during the 8 months from vaccination (first week of March 2008) to the SW6 sampling (first week of November 2008), while 3.6% died during the 7 months from the SW6 sampling to the SW12 slaughter (June 2009).
Descriptive statistics for the traits recorded for the FW, SW6, and SW12 groups are shown in Table 1. The frequency of all types of deformities was 6.4% in the FW group, while the incidence increased significantly (P < 0.001), rising from 5.2% at the SW6 recording to 6.4% at SW12. Fused vertebrae were the most dominant deformity in all groups, with significantly higher frequency (P < 0.05) in the SW12 group (5.7%) than in the FW group (4.2%). The frequency of hyperdense vertebrae was 2.2% in the FW group, but as expected, it was very low (SW6) or non-existent (SW12) in the two other groups as this type of deformity is seldom seen after sea transfer. The frequency of missing intervertebral space was very low in the SW6 and SW12 groups and did not occur in the FW group.
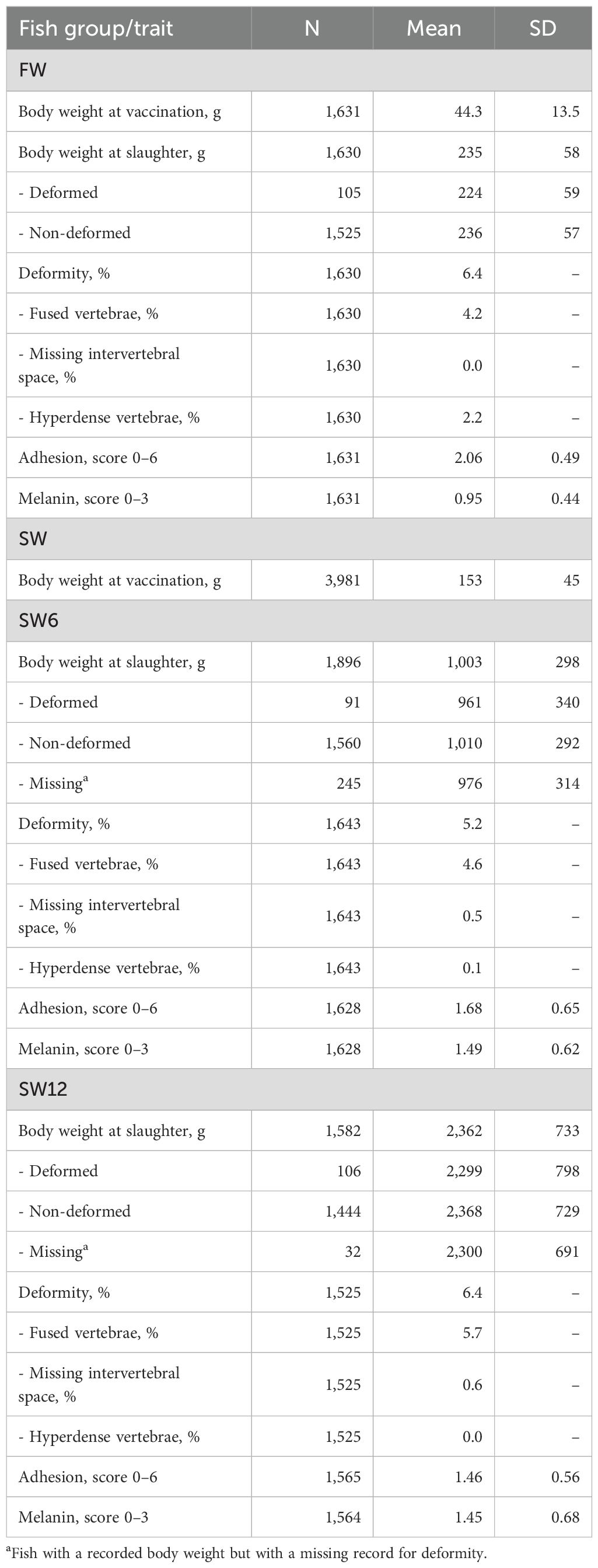
Table 1. Descriptive statistics of the recorded traits (N, number of recorded fish; SD, standard deviations) for fish reared 3 months on 17°C freshwater (FW) and for 6 (SW6) and 12 (SW12) months in a net cage in the sea.
Two fused vertebrae were most common in all groups, but a few fish had up to 16 affected vertebrae. Hyperdense vertebrae were mostly seen in the FW group, with one to two affected vertebrae as the most common (Figure 3). The number of vertebrae diagnosed as missing intervertebral space varied from 2 to 9 in the SW6 group and from 1 to 7 in the SW12 group.
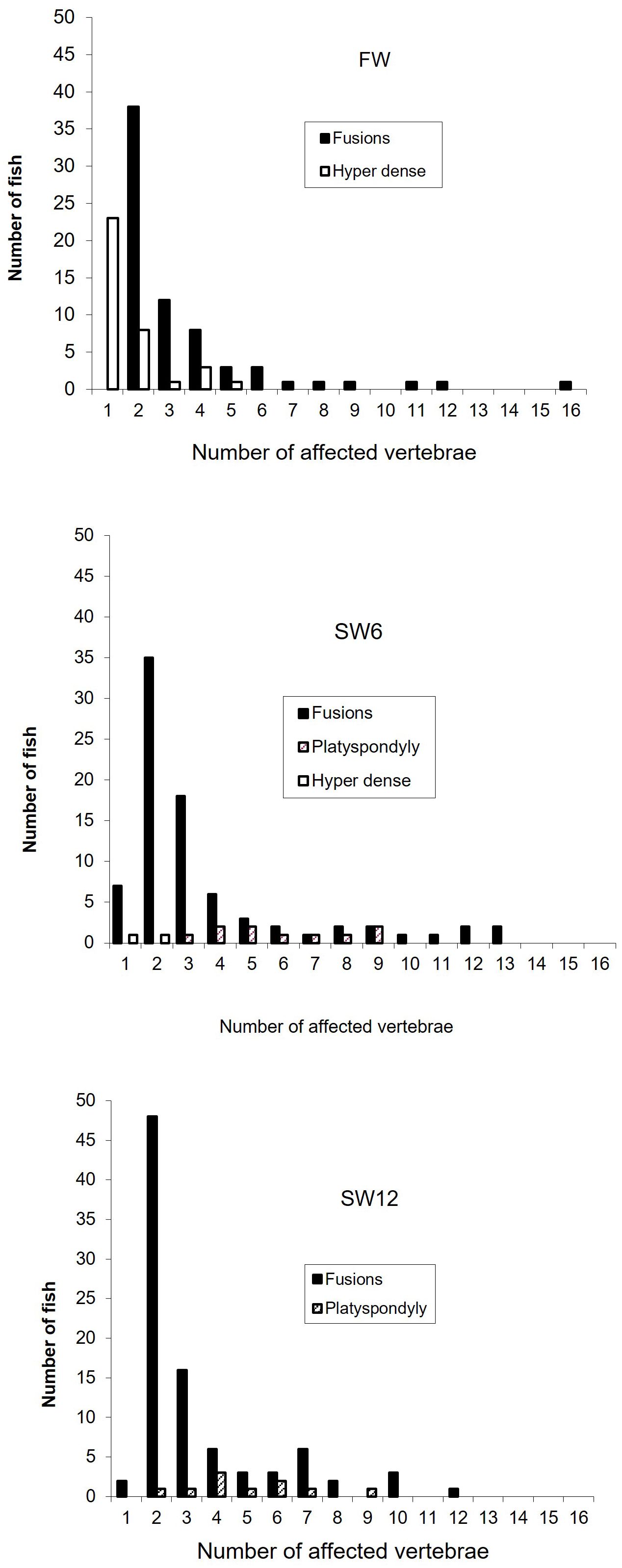
Figure 3. The distribution of the number of deformed vertebrae per fish (fish with no deformities excluded) of the vaccinated fish recorded in freshwater (FW) and after 6 (SW6) and 12 (SW12) months in a net cage in the sea.
The average body weight of the deformed fish was 4.8% (FW), 4.9% (SW6), and 2.9% (SW12) lower than the non-deformed (Table 1), in accordance with an earlier report (Gjerde et al., 2005), but none of these differences were significantly different from zero (P > 0.05).
Adhesion score was higher in the FW group than in the SW6 and SW12 groups, while the opposite was the case for melanin deposits. In the SW6 and SW12 groups, the average adhesion and melanin scores were very similar.
3.3 Heritability
For the FW and SW6 groups, the (underlying) heritability estimate for fused vertebrae was of medium magnitude and higher than for deformity. However, for the SW12 group, heritability was low for both deformity and fused vertebrae (Table 2). On the observed scale, heritability across the three experimental groups was 0.07 ± 0.02 for deformity and 0.08 ± 0.02 for fused vertebrae.
The heritability estimates for adhesion, melanin deposits, and body weight were of medium magnitude (Table 2) and, as expected, very similar to the estimates obtained from the same fish material (Drangsholt et al., 2011; Drangsholt et al., 2012).
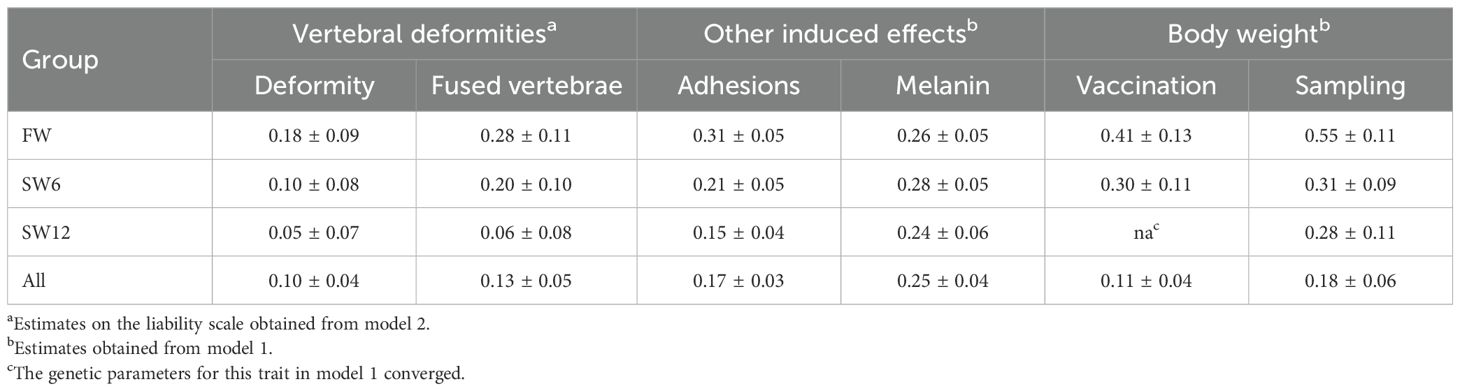
Table 2. Estimates of heritability with standard errors for the studied traits recorded after 3 months on 17°C freshwater (FW) and after 6 (SW6) and 12 (SW12) months in a net cage in seawater.
3.4 Effect common to full sibs
For the trait body weight at vaccination, the effect common to full sibs did not converge for the SW12 group. For the two other groups (FW, SW6), the estimates of this effect, as a proportion of the total variance, were low for both body weight at vaccination (<0.08) and body weight at the SW6 sampling (<0.03). Across the three experimental groups, the effect common to full sibs was low for both body weight at vaccination (0.02 ± 0.02) and body weight at the SW samplings (0.03 ± 0.02).
3.5 Genetic correlations of deformities across groups
The genetic correlation between the FW and SW6 groups was very high for both deformity and fused vertebrae, while the genetic correlations of FW and SW6 with SW12 were intermediate for deformity but lower for fused vertebrae, albeit uncertain as indicated by their large standard errors (Table 3).
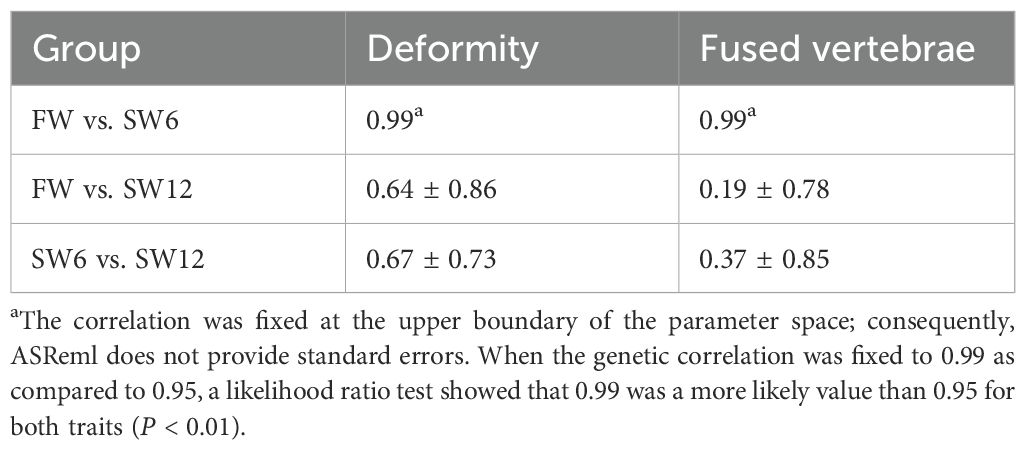
Table 3. Estimates of genetic correlations with standard errors between the same trait (deformity or fused vertebra) recorded after 3 months on 17°C freshwater (FW) and after 6 (SW6) and 12 (SW12) months in a net cage in seawater (model 2).
3.6 Genetic and residual correlations of fused vertebrae with the other traits
Across the three experimental groups (FW, SW6, and SW12), the genetic and residual correlations of fused vertebrae with the other studied traits were all close to and not significantly different from zero (P > 0.05) (Table 4). Within each of the experimental groups, the genetic correlations were more variable and with larger standard errors (0.18 to 0.24) than for those across groups, and none were significantly different from zero (P > 0.05). The same pattern was seen for the correlations of deformity with each of the other traits (results not shown).
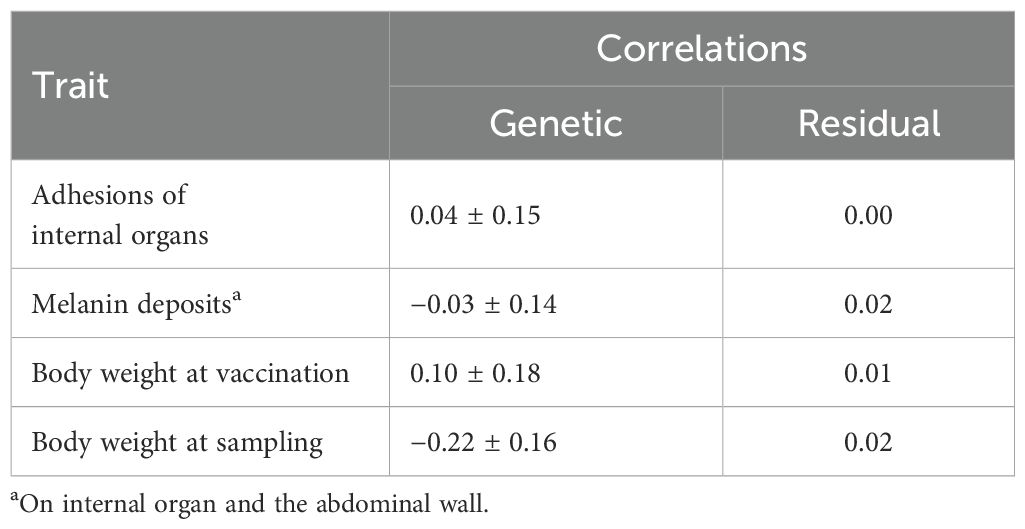
Table 4. Estimates of genetic and residual (environmental) correlations with standard errors of the trait fused vertebra with the other mentioned traits across the three experimental groups: FW, SW6, and SW12 (model 1).
4 Discussion
The magnitude of the estimated genetic and residual correlations of fused vertebrae, the most dominant deformity in the FW, SW6, and SW12 groups, with adhesions and melanin deposits, was close to and not significantly different from zero, which strongly indicates that vaccine-induced side effects and vertebral deformities are caused by different genetic and environmental factors. This agrees with Grini et al. (2011), who reported no effect of vaccination on deformity, and with Bjørgen et al. (2024), who found evidence that melanized focal changes in the fillet of Atlantic salmon are due to a sequence of events with the pivotal role of adiposity, hypoxia, and fat necrosis, but in contrast to two earlier studies that reported a strong phenotypic association between these traits (Berg et al., 2006; Aunsmo et al., 2008) and a recent study that observed a significant difference in the percentage of deformed fish between fish that received two different vaccines (Thorarinsson et al., 2023). The incidence rates of deformities in this study (6.4%, 5.2%, and 6.4% at the FW, SW6, and SW12 recording, respectively) were, in general, lower than in other studies in which the fish were examined 6–13 months after sea transfer (Witten et al., 2006; Aunsmo et al., 2008; Fjelldal et al., 2009; Grini et al., 2011; Thorarinsson et al., 2023). Therefore, it cannot be ruled out that the estimated genetic correlations may be different at higher deformity incidence rates.
The close to unity genetic correlation between the same deformity trait (deformity or fused vertebrae) in the FW and SW6 groups showed that fused vertebrae, the dominant deformity trait in this study, has a strong common genetic base (Table 3). The reason for the much lower genetic correlations between fused vertebrae in the FW and SW12 groups, as well as in the FW6 and the SW12 groups, may be the lower heritability of fused vertebrae in the SW12 than in the SW6 group (Table 2).
Similarly, a significant genetic variation in adhesion score and melanin score was supported by Drangsholt et al. (2011) who, using the same data as in this study, found a medium genetic correlation of adhesion score in the FW group with adhesion score in the SW6 (0.62 ± 0.12) and SW12 (0.48 ± 0.14) groups; a medium to relatively high genetic correlation of melanin score in the FW group with melanin score in the SW6 (0.84 ± 0.08) and SW12 (0.61 ± 0.11) groups; and a high genetic correlation between adhesion scores in the SW6 and SW12 groups (0.92 ± 0.11) and between melanin score in the two groups (0.89 ± 0.07).
In this study, as observed from the SW6 to the SW12 recording, an increase in the frequency of deformity as fish grow was also reported both prior to (Witten et al., 2006) and after (Witten et al., 2006; Grini et al., 2011) smoltification and may have increased to a higher level if the fish were reared to a larger marketing body size of, for example, 4–5 kg. Factors that may have contributed to these differences are different temperatures during egg incubation (Takle et al., 2005; Gjerde et al., 2005), the use of yearling smolt (+1) as compared to under yearling smolt (0+), and differences in time and body weight at vaccination (Vågsholm, 1998; Berg et al., 2006).
The very low number of affected fish with more than two fused vertebrae made it impossible to explore the magnitude of genetic variation in the number or severity of fused vertebrae in this study. As expected, hyperdense vertebrae were mostly seen in the FW group as this deformity is typically seen in small fish before sea transfer (Helland et al., 2005), while missing intervertebral space, normally observed in larger fish (Witten et al., 2006; Thorarinsson et al., 2023), was observed at a very low frequency at the two seawater samplings in this study. In Atlantic salmon, reduced growth was first observed in fish with more than 10 affected vertebrae (Hansen et al., 2010). Therefore, the deformity in this study was most likely a minor welfare problem for the fish due to the low frequency of fish with deformed vertebrae and the very low number of affected vertebrae.
The lower melanin score in the FW group than in the SW6 and SW12 groups indicates that vaccine-induced intra-abdominal lesions take a longer time to develop than adhesions. The very similar adhesion and melanin scores of the vaccinated SW6 and SW12 fish show that their severity did not develop over time as the vertebrae deformity did. A similar trend was observed by Grini et al. (2011) with a slightly decreasing lesion score from the time of seawater transfer until 26 and 36 months in seawater for fish that were reared at 10°C both in the freshwater and seawater phase, but with a higher lesion score for fish that had been reared at a higher fresh and seawater temperature. The lack of further development of the vaccine-induced side effects from the SW6 to the SW12 recordings may therefore be due to the relatively low seawater temperature during this period.
The intermediate heritability estimates for fused vertebrae in the FW and SW6 groups indicate a significant additive genetic component for this type of deformity in Atlantic salmon. The lower heritability for the composite trait deformity (fused vertebrae, missing intervertebral space, and hyperdense vertebrae) may indicate that these types of deformities have a somewhat different etiology. However, due to the low frequency of missing intervertebral spaces and hyperdense vertebrae and the relatively large standard errors of the heritability estimates, this explanation should be interpreted with caution. That vertebral deformities in Atlantic salmon has a genetic component is in agreement with Gjerde et al. (2005) who, based on visual scores of external deformities of fish from three different year-classes with relatively low to intermediate deformity (9.5%, 7.6%, and 21.5%), found medium heritability estimates (on the liability scale: 0.22, 0.25, and 0.36, respectively) but zero heritability in a four year-class with a much lower deformity (2.3%).
The discrepancy between our results of no evidence of an association between vaccine-induced side effects and deformities in Atlantic salmon and a positive association as referred to in the abovementioned studies (Berg et al., 2006, 2007; Aunsmo et al., 2008) may be due to a number of reasons. First, the level of adhesion and melanin deposits was slightly higher in the other studies, which can be due to differences in both the vaccine formulation and volume injected, as a new formulated vaccine with a smaller injection volume (0.05 mL vs. 0.1 mL) resulted in lower adhesion and melanin scores (Drangsholt et al., 2011). Second, considering the different etiologies of different types of vertebral deformities, fused vertebrae were the most frequent deformity in our study, whereas platyspondyly was the most prominent in Berg et al. (2006) and platyspondyly and fused vertebrae were the most common in Aunsmo et al. (2008). Third, the use of yearling smolt (1+) in our study, such as under yearling smolts (0+), showed a higher risk of deformities (Vågsholm, 1998). Fourth, it has been proposed that both the time of vaccination and body weight at vaccination can influence the development of both adhesions and vertebral deformities (Berg et al., 2006). However, in this study, the effect of body weight on deformity was not likely, as both the genetic and residual correlations of body weight at vaccination with adhesions, melanin deposits, and deformity were all close to zero.
The etiology of deformities in vaccinated Atlantic salmon is not fully understood, but periodic mineral deficiency due to starvation prior to vaccination and reduced feed intake due to vaccination have been proposed as explanatory factors (Aunsmo et al., 2008). However, Berg et al. (2012) found no long-term effects of starvation on bone mineralization or deformity incidence in Atlantic salmon. The development of a structural weakness in the vertebrae at the time of vaccination is shown in Holm et al. (2020), leading to the development of “cross-stitch vertebrae.” This particular type of deformity tends to develop near slaughter size and is not easily observed in early life stages.
Missing intervertebral space, as observed at low frequency in the SW6 and SW12 groups in this material, may be an early stage of cross-stitch vertebrae (Thorarinsson et al., 2023) or reduced mineralization, resulting in compressed vertebrae/platyspondylia (Kvellestad et al., 2020). Fish with more severe vaccine-induced side effects may exhibit poorer feed intake and/or mineral absorption; however, these effects may be difficult to observe when the side effects of vaccination are mild, or the high availability of minerals in the feed compensates for the reduced feed intake and absorption. However, this is a less likely reason for the missing intervertebral space deformity in this material, as it was only observed after 6 and 12 months in seawater, at which point animals with low-mineralized vertebrae caused by low dietary phosphorus in the freshwater period may have recovered during the seawater period (Drábiková et al., 2022).
The close to zero genetic and residual correlations of the deformity trait fused vertebrae with body weight at vaccination and body weight of the fish both at the FW, SW6, and SW12 samplings (Table 4) indicate that high growth rate is a minor, if any, cause of the fused vertebrae.
To conclude, no evidence was found of a genetic association between the trait of fused vertebrae and adhesions of internal organs or melanin deposits on internal organs and the abdominal wall in Atlantic salmon. The reliability of this finding is supported by the significant genetic variation in both the deformity trait and the adhesions and melanin trait. That the genetic correlation of fused vertebra with adhesions and/or melanin deposits should be significantly different from zero in another material with, for example, a higher proportion of deformed fish cannot be ruled out, but not very likely.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
Ethical approval was not required for the studies involving animals in accordance with the local legislation and institutional requirements. Written informed consent was obtained from the owners for the participation of their animals in this study.
Author contributions
BG: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. TK-D: Conceptualization, Data curation, Methodology, Writing – review & editing. JØ: Writing – review & editing. FF-F: Conceptualization, Data curation, Methodology, Supervision, Writing – review & editing. IT: Resources, Writing – review & editing. KH: Data curation, Methodology, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. The study was funded by grant 179009/S40 from the Research Council of Norway and The Fishery and Aquaculture Industry Research Fund and PHARMAQ AS.
Acknowledgments
Fish used in the project were given free of charge from SalmoBreed AS (now Benchmark Genetics). Kjellrun Gannestad and other staff at Nofima Research Station at Sunndalsøra and Averøy provided skillful assistance during the data collection.
Conflict of interest
Author FF-F was employed by company PHARMAQ AS. Author IT was employed by company Benchmark Genetics Norway.
The authors declare that this study received funding from PHARMAQ AS. This funder had the following involvement in the study: produced the vaccine used and scoring of adhesion and melanin.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Aunsmo A., Guttvik A., Midtlyng P. J., Larssen R. B., Evensen Ø., and Skjerve E. (2008). Association of spinal deformity and vaccine-induced abdominal lesions in harvest-sized Atlantic salmon, Salmo salar L. J. Fish Dis. 31, 515–524. doi: 10.1111/j.1365-2761.2007.00899.x
Baeverfjord G., Hjelde K., Helland S., and Refstie S. (2009). “The effect of minerals on juvenile Atlantic salmon,” in Control of alformations in fish aquaculture: science and practice, federation of european aquaculture producers. Eds. Baeverfjord G., Helland S., and Hough C. (RapidPress, Luxembourgh), 67–71.
Baeverfjord G., Prabhu P. A. J., Fjelldal P. G., Albrektsen S., Hatlen B., Denstadli V., et al. (2018). Mineral nutrition and bone health in salmonids. Rev. Aquaculture. 11, 740–765. doi: 10.1111/raq.12255
Berg A., Rødseth O. M., and Hansen T. (2007). Fish size at vaccination influence the development of side-effects in Atlantic salmon (Salmo Salar L.). Aquaculture 265, 9–15. doi: 10.1016/j.aquaculture.2007.02.014
Berg A., Rødseth O. M., Tangerås A., and Hansen T. (2006). Time of vaccination influences development of adhesions, growth and spinal deformities in Atlantic salmon Salmo salar. Dis. Aquat. Organisms 69, 239–248. doi: 10.3354/dao069239
Berg A., Yurtseva A., Hansen T., Lajus D., and Fjelldal P. G. (2012). Vaccinated farmed Atlantic salmon are susceptible to spinal and skull deformities. J. Appl. Ichthyol. 28 (3), 446–452. doi: 10.1111/j.1439-0426.2012.01988.x
Bjørgen H., Brimsholm M., Asserson C. F., Skaar K., Knutsen G. M., Oaland Ø., et al. (2024). Deciphering the pathogenesis of melanized focalchanges in the white skeletal muscle of farmed Atlantic salmon (Salmo salar). J. Fish Dis. 47, e13988. doi: 10.1111/jfd.13988
Brudeseth B. E., Wiulsrod R., Fredriksen B. N., Lindmo K., Lokling K. E., Bordevik M., et al. (2013). Status and future perspectives of vaccines for industrialised fin-fish farming. Fish Shellfish Immunol. 35, 1759–1768. doi: 10.1016/j.fsi.2013.05.029
Drábiková L., Fjelldal P. G., De Clercq A., Yousaf M. N., Morken T., McGurk C., et al. (2022). What will happen to my smolt at harvest? Individually tagged Atlantic salmon help to understand possible progression and regression of vertebral deformities. Aquaculture 559. doi: 10.1016/j.aquaculture.2022.738430
Drangsholt T. M. K., Gjerde B., Ødegård J., Fridell F., and Bentsen H. B. (2011). Quantitative genetics of vaccine-induced side effects in farmed Atlantic salmon (Salmo salar). Aquaculture 318, 316–324. doi: 10.1016/j.aquaculture.2011.05.044
Drangsholt T. M. K., Gjerde B., Ødegård J., Finne-Fridell F., Evensen Ø., Bentsen H. B., et al. (2012). Genetic correlations between disease resistance, vaccine induced side effects and harvest body weight in Atlantic salmon. Aquaculture. 325, 324–325. doi: 10.1016/j.aquaculture.2011.11.007
Färber F. (2017). Melanin spots in Atlantic salmon fillets – An investigation of the general problem, the frequency and the economic implication based on an online survey (Norway: Department of Animal and Aquaculture Sciences, Norwegian University of Life Sciences (NMBU), 82.
Fjelldal P. G., Hansen T. J., and Berg A. E. (2007). A radiological study on the development of vertebral deformities in cultured Atlantic salmon (Salmo salar L.). Aquaculture 273, 721–728. doi: 10.1016/j.aquaculture.2007.07.009
Fjelldal P. G., Hansen T., Breck O., Ørnsrud. R. E.-J., Lock E.-J., Waagbø R., et al. (2012). Vertebral deformities in farmed Atlantic salmon (Salmo salar L.) – etiology and pathology. J. Appl. Ichthyol. 28, 433–440. doi: 10.1111/j.1439-0426.2012.01980.x
Fjelldal P. G., Hansen T., Breck O., Sandvik R., Waagbø R., Berg A., et al. (2009). Supplementation of dietary minerals during the early seawater phase increase vertebral strength and reduce the prevalence of vertebral deformities in fast-growing under-yearling Atlantic salmon (Salmo salar L.) smolt. Aquaculture Nutr. 15, 366–378. doi: 10.1111/j.1365-2095.2008.00601.x
Gilmour A. R., Gogel B. J., Cullis B. R., Welham S. J., and Thompson R. (2009). ASReml user guide release 3.0 (Hemel Hempstead, UK: VSN International Ltd).
Gjerde B., Evensen Ø., Bentsen H. B., and Storset A. (2009). Genetic (co)variation of vaccine injuries and innate resistance to furunculosis (Aeromonas salmonicida) and infectious salmon anaemia (ISA) in Atlantic salmon (Salmo salar). Aquaculture 287, 52–58. doi: 10.1016/j.aquaculture.2008.10.028
Gjerde B., Pante M. J. R., and Baeverfjord G. (2005). Genetic variation for a vertebral deformity in Atlantic salmon (Salmo salar). Aquaculture 244, 77–87. doi: 10.1016/j.aquaculture.2004.12.002
Grini A., Hansen T., Berg A., Wargelius A., and Fjelldal P. G. (2011). The effect of water temperature on vertebral deformities and vaccine-induced abdominal lesions in Atlantic salmon, Salmo salar L. J. Fish Dis. 34, 531–546. doi: 10.1111/j.1365-2761.2011.01265.x
Gudding R., Lillehaug A., and Evensen O. (1999). Recent developments in fish vaccinology. Veterinary Immunol. Immunopathology 72, 203–212. doi: 10.1016/S0165-2427(99)00133-6
Hansen T., Fjelldal P. G., Yurtseva A., and Berg A. (2010). A possible relation between growth and number of deformed vertebrae in Atlantic salmon (Salmo salar L.). J. Appl. Ichthyol. 26, 355–359. doi: 10.1111/j.1439-0426.2010.01434.x
Helland S., Refstie S., Espmark Å., Hjelde K., and Baeverfjord G. (2005). Mineral balance and bone formation in fast-growing Atlantic salmon parr (Salmo salar) in response to dissolved metabolic carbon dioxide and restricted dietary phosphorus supply. Aquaculture 250, 364–376. doi: 10.1016/j.aquaculture.2005.03.032
Helland S., Vegard Denstadli V., Witten P. E., Hjelde K., Storebakken T., Skrede A., et al. (2006). Hyper dense vertebrae and mineral content in Atlantic salmon (Salmo salar L.) fed diets with graded levels of phytic acid. Aquaculture 261, 603–614. doi: 10.1016/j.aquaculture.2006.08.027
Holm H., Ytterborg E., Høst V., Reed A. K., Dalum A. S., and Baeverfjord G. (2020). A pathomorphological description of cross-stitch vertebrae in farmed Atlantic salmon (Salmo salar L.). Aquaculture 526, 735382. doi: 10.1016/j.aquaculture.2020.735382
Jiménez-Guerrero R., Baeverfjord G., Evensen Ø., Hamre K., Larsson T., Dessen J.-E., et al. (2022). Rib abnormalities and their association with focal dark spots in Atlantic salmon fillets. Aquaculture 561. doi: 10.1016/j.aquaculture.2022.738697
Kryvi H., Nordvik K., Fjelldal P. G., Eilertsen M., Helvik J. V., Støren E. N., et al. (2020). Heads and tails: The notochord develops differently in the cranium and caudal fin of Atlantic Salmon (Salmo salar, L.). Anat Rec. 304, 1629–1649. doi: 10.1002/ar.24562
Kvellestad A., Høie S., Thorud K., Tørud B., and Lyngøy. A. (2000). Platyspondyly and shortness of vertebral column in farmed Atlantic salmon Salmo salar in Norway - description and interpretation of pathologic changes. Dis. Aquat Org 39, 97–108. doi: 10.3354/dao039097
McKay L. R. and Gjerde B. (1986). Genetic variation for a spinal deformity in Atlantic salmon. Aquaculture 52, 263272. doi: 10.1016/0044-8486(86)90369-8
Midtlyng P. J., Reitan L. J., and Speilberg L. (1996). Experimental studies on the efficacy and side-effects of intraperitoneal vaccination of Atlantic salmon (Salmo salar L.) against furunculosis. Fish Shellfish Immunol. 6, 335–350. doi: 10.1006/fsim.1996.0034
Mutoloki S., Alexandersen S., and Evensen Ø. (2004). Sequential study of antigen persistence and concomitant inflammatory reactions relative to side-effects and growth of Atlantic salmon (Salmo salar L.) following intraperitoneal injection with oil-adjuvanted vaccines. Fish Shellfish Immunol. 16, 633–644. doi: 10.1016/j.fsi.2003.10.002
Ramstad A., Romstad A. B., Knappskog D. H., and Midtlyng P. J. (2007). Field validation of experimental challenge models for IPN vaccines. J. Fish Dis. 30, 723–731. doi: 10.1111/j.1365-2761.2007.00858.x
Sommerset I., Krossøy B., Biering E., and Frost P. (2005). Vaccines for fish in aquaculture. Expert Rev. Vaccines 4, 89–101. doi: 10.1586/14760584.4.1.89
Takle H., Baeverfjord G., Lunde M., Kolstad K., and Andersen Ø. (2005). The effect of heat and cold exposure on HSP70 expression and development of deformities during embryogenesis of Atlantic salmon (Salmo salar). Aquaculture 249, 515–524. doi: 10.1016/j.aquaculture.2005.04.043
Thorarinsson R., Negaard P., Baeverfjord G., Peña P., and Skjerve E. (2023). The effect of vaccine-associated cross-stitch vertebrae pathology on growth of farmed Atlantic salmon. J. World Aquaculture Soc. 55, 240–256. doi: 10.1111/jwas.1297
Vågsholm D. (1998). Risk factors for spinal deformities in Atlantic salmon, Salmo salar L. J. Fish Dis. 21, 47–53. doi: 10.1046/j.1365-2761.1998.00069.x
Witten P. E., Gil-Martens L., Huysseune A., Takle H., and Hjelde K. (2009). Towards a classification and an understanding of developmental relationships of vertebral body malformations in Atlantic salmon (Salmo salar L.). Aquaculture 295, 6–14. doi: 10.1016/j.aquaculture.2009.06.037
Witten P. E., Obach A., Huysseune A., and Baeverfjord G. (2006). Vertebrae fusion in Atlantic salmon (Salmo salar): Development, aggravation and pathways of containment. Aquaculture 258, 164–172. doi: 10.1016/j.aquaculture.2006.05.005
Appendix 1. The experimental design with dates of the different activities and trait recordings.
.
Appendix 2. Freshwater temperature during the 3-month test period of the FW group.
.
Appendix 3. Seawater temperature during the seawater period of the SW group.
.
Keywords: Atlantic salmon, Salmo salar, vaccine, adhesions, melanin, deformity, heritability, genetic correlations
Citation: Gjerde B, Karlsson-Drangsholt TM, Ødegård J, Finne-Fridell F, Thorland I and Hjelde K (2025) Estimates of genetic and environmental (co)variances for vertebral deformities and vaccine-induced side effects in Atlantic salmon. Front. Anim. Sci. 6:1609372. doi: 10.3389/fanim.2025.1609372
Received: 10 April 2025; Accepted: 31 July 2025;
Published: 09 October 2025.
Edited by:
Gregorio Miguel Ferreira De Camargo, Federal University of Bahia (UFBA), BrazilReviewed by:
Mario Caruffo, University of Chile, ChileLuciana Shiotsuki, Brazilian Agricultural Research Corporation - Fisheries and Aquaculture, Brazil
Copyright © 2025 Gjerde, Karlsson-Drangsholt, Ødegård, Finne-Fridell, Thorland and Hjelde. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Bjarne Gjerde, Ymphcm5lLmdqZXJkZUBub2ZpbWEubm8=
†Present addresses: Tale Marie Karlsson-Drangsholt, The Research Council of Norway, Norway
Jørgen Ødegård, Department of Animal and Aquacultural Sciences, Norwegian University of Life Sciences, Aas, Norway
‡ORCID: Bjarne Gjerde, orcid.org/0000-0002-6827-7407