 Bjarne Gjerde1*†
Bjarne Gjerde1*† Solomon Antwi Boison2
Solomon Antwi Boison2 David Hazlerigg3
David Hazlerigg3 Trine Ytrestøyl4
Trine Ytrestøyl4 Turid Mørkøre5
Turid Mørkøre5 Even Jørgensen3
Even Jørgensen3 Anja Striberny3,6
Anja Striberny3,6 Simen Rød Sandve5
Simen Rød Sandve5- 1Department of Breeding and Genetics, Nofima AS, Ås, Norway
- 2Mowi Genetics AS, Bergen, Norway
- 3Department of Arctic and Marine Biology UiT, The Arctic University of Norway, Tromsø, Norway
- 4Department of Nutrition and Feed Technology, Nofima AS, Sunndalsøra, Norway
- 5Department of Animal and Aquacultural Sciences, Norwegian University of Life Sciences, Ås, Norway
- 6Department of Production Biology, Nofima AS, Tromsø, Norway
Introduction: This study investigates the impact of three different light regimes prior to seawater transfer on growth, survival, wounds, and losers in Atlantic salmon during a ten-month period in a net-cage in the sea.
Methods: A total of 3,000 Atlantic salmon pre-smolts from 100 families were tagged with passive integrated transponders (PIT) and distributed across six tanks, with duplicated tanks exposed to a different light regime: 6 weeks on 8-hour light (L)/16-hour dark (D), 12L:12D, and 24L:0D, followed by 6 weeks on 24L:0D. One week prior to seawater transfer in June 2021, body weights and external smolt status were recorded. Body weights, wounds, and losses were recorded 135 days after seawater transfer in November 2021, and body weights were recorded again 300 days after seawater transfer in April 2022. A binary survival trait was defined for the seawater periods: Period 1 (day 0 to day 135) and Period 2 (day 136 to day 300), along with the time to death for the fish that were collected as dead.
Results: Body weight in June 2021 and time to death in Period 1 were the only traits significantly affected by the light regime. Fish on the 24L:0D regime were, on average, 10.0% and 12.2% heavier than those on the 12L:12D and 8L:16D regimes, respectively. They also died, on average, 15.7 and 17.7 days earlier than those on the 12L:12D and 8L:16D regimes. During the first two months in seawater, mortality was higher for fish on the 24L:0D regime, with an accumulated mortality of 8.8%, compared to 0.7% and 1.0% for those on the 8L:16D and 12L:12D regimes, respectively. Mortality in Period 2 was similar across the three light regimes. Most traits had moderate heritability, but heritability was low for healthy fish and fish with wounds in November 2021, as well as for survival and time to death in Period 1. There was negligible genotype-by-light regime interaction. Genetic correlations between survival and the other traits were low to medium, and not significantly different from zero.
Discussion: While light regimes did not significantly affect growth during the seawater period, they did influence survival, particularly during the first two months in seawater. The estimated genetic parameters suggest that direct selection for increased growth and survival during the early seawater phase may be more effective for improving seawater performance than indirect selection based on the studied smolt-indicator traits.
Conclusion: The findings indicate that direct selection for increased growth and survival during the early seawater period offers promising prospects for enhancing seawater performance.
1 Introduction
Atlantic salmon (Salmo salar) is the major aquaculture commodity in Norway. In 2023, the production was 1.542 million tons (94% of Norway’s total aquaculture production), valued to 107.5 billion NOKs (https://www.fiskeridir.no) or 48 billion EUR. Wild salmon are anadromous fish adapted to spend their first years in freshwater, migrate to sea as “smolts,” grow to reach sexual maturity, and finally return to their river of birth to spawn. Hence, the production of farmed salmon involves two phases: first, a freshwater phase from the time of egg fertilization up to the smolt stage (12–18 months), when they are moved to floating seawater net cages where they grow for another 12–18 months to the desired harvest body weight. In 2023, 420 million smolts were transferred to sea cages in Norway. However, with a median mortality of 18.8%, a staggering 62.8 million fish died before harvesting, with delousing injuries, complex gill diseases, and winter ulcers as the three overall main heath issues (Sommerset et al., 2023).
Based on a survey from fish health personnel, inadequate physiological maladaptation and smoltification and the development of runt syndrome are still problems due to complex reasons, including the increasingly diverse methods of producing hatchery fish, both in terms of new production technology and smoltification protocols (Sommerset et al., 2023). In some of the production areas, these issues were the causes of more than 40%–50% of the losses (Sommerset et al., 2023). A significant part of the mortality takes place during the first months after seawater transfer and is attributed to suboptimal smolt development and physiology (Aunsmo et al., 2008).
Mortality obviously represents economic loss for farmers due to both reduced revenue and the higher cost per kilogram fish slaughtered mainly due to a reduced feed efficiency and, thus, the high feed cost, but also a huge animal welfare problem. A pressing challenge in salmon aquaculture research is therefore to better understand the smolt production factors that reduce mortality and increase the feed resource efficiency and animal welfare post-seawater transfer (Kolarevic et al., 2014; Ytrestøl et al., 2022; Khaw et al., 2021; Striberny et al., 2021).
The developmental process turning a freshwater salmon (parr) into a saltwater-adapted fish (smolt) is termed “smoltification.” This encompasses a multitude of behavioral, morphological, and physiological preparatory changes that help the fish cope with the seawater environment. The skin color changes from a darker tone with parr marks (i.e., spots) to a silvery appearance and the body shape becomes longer and thinner, resulting in a reduction in the condition factor (Folmar and Dickhoff, 1980; Hoar, 1988; Sigholt et al., 1998; McCormic et al., 2007; Nichols et al., 2008; Piironen et al., 2013; Khaw et al., 2021).
At the molecular level, a number of organs change functional properties (Takvam et al., 2021), and many of these changes are related to the change from a freshwater to a seawater rearing environment. In full-strength seawater, the osmolarity of the fish internal fluids is around one-third of the surrounding water (Evans et al., 2005); hence, they become hypoosmotic relative to the external seawater (McCormick et al., 2013). To cope with this, a hypo-osmoregulatory ability is developed during smoltification through changes in the kidney function (Takvam, 2020; Takvam et al., 2021), enhanced water permeability in the intestine (Boeuf, 1993; Ura et al., 1997; Duarte et al, 2023), and remodeling of the gill cell populations (McCormick et al., 2013), resulting in a more efficient salt excretion capacity.
In nature, smoltification is triggered by the increased day length in the spring, making the salmon prepared for migration to sea in the early summer (May–June in the Northern Hemisphere). The production of smolts in aquaculture has therefore traditionally been achieved by mimicking a natural photoperiodic regime. Typically, these production protocols use continuous light in the parr (pre-smolt) stage, interrupted by 6–8 weeks with short days (≤12 h of daylight), followed by 68 weeks on 24-h daylight. Based on observations of growth in seawater, it has been shown that the duration of exposure to short days should be at least 6 weeks for the following long days to induce proper smolt development (Duncan and Bromage, 1998). However, exposure to short days reduces the growth rate (Sigholt et al., 1998; Handeland et al., 2013). Therefore, motivated by growth maximization (Bjornsson et al., 1989; Myklatun et al., 2023) and practical considerations, the salmon farming industry has more recently explored alternative smolt production protocols. These protocols include rearing the smolts to a much larger body weight (200–500 g compared with 75–200 g) due to the positive correlation between size and salinity tolerance (Handeland et al., 2013; Sigholt et al., 1995; Strand et al., 2018) and using feeds with added salt prior to seawater transfer, which is known to increase seawater tolerance (Basulto, 1976; Salman and Eddy, 1988; Staurnes and Finstad, 2000; Duarte et al., 2023; Myklatun et al., 2023; 2025). A few recent trials have evaluated smolt performance under different rearing protocols.
The water temperature during the regime has been found to affect the rate of smolt development, but is of minor importance on the quality of the smoltification process itself (Sigholt et al., 1998). This is in agreement with the results of a recent paper, which found that the use of a reduced “winter” temperature treatment (8°C vs. 14°C) in combination with short day length “winter” photoperiods did not enhance the smolt quality of Atlantic salmon when compared with the use of short day length “winter” photoperiods alone [12-h light (L)/12-h dark (D) and 08L:16D vs. 24L:0D] (Montgomery et al., 2025).
Ytrestøyl et al. (2022) compared the growth of three groups of Atlantic salmon in recirculating aquaculture systems (RAS) that were exposed to either a conventional 12L:12D or a 24L:0D light smoltification regime prior to transfer to net cages in seawater at average body weights of 100, 200, and 600 g. At the first sampling, 8 weeks after seawater transfer, the group exposed to the 24L:0D regime was bigger than the 200-g group exposed to the 12L:12D regime due to the higher growth rate prior to seawater transfer. However, the best-performing group (with the largest average harvest weight, low mortality, and no sexual mature fish) was the 100-g group smoltified under the 12L:12D regime. Harvest weight was lowest in fish transferred to sea at 600 g, despite having the highest day-degree sum during their life span. This is in agreement with Myklatun et al. (2025), who found that the harvest yield of sexually immature fish was approximately 1.8 times higher in groups transferred directly to sea cages as smolts compared with those that remained on land during early post-smolt rearing. These results strongly indicate that smolts reared under constant light experience difficulties adjusting to the new saline environment.
In another trial, Striberny et al. (2021) compared the performance of smolts exposed to either a short day (i.e., 7L:14D) or raised under constant light (i.e., 24L:0D), in combination with transition feed. Here, the authors confirmed that salt-spiked transition feed increases the ability to hypo-osmoregulate, but that the combination of short-day treatment and transition feed resulted in the best-performing smolts (based on growth in seawater). Taken together; smolt production protocols matter, and these protocols benefit from leveraging seasonal signals that salmon are naturally adapted to respond to.
In addition to environmental factors, genetic variation is also known to play a major role in quantitative trait variation. Nevertheless, genetics has mostly been neglected in the research on smolt development and rearing protocols. Direct selection for improved smolt characteristics can only be obtained if the actual trait(s) show genetic variation, and their genetic correlation with other important production traits (e.g., growth and survival) should preferably not be unfavourable as this could result in less or unfavourable genetic gains for these other traits directly or indirectly selected for. Little is known about the magnitude of genetic variation in smolt development characteristics and their genetic and environmental correlation with other traits. In addition, no estimate is available regarding the extent of families ranking differently with respect to important smolt traits and other production traits when sibs of the same families are smoltified on different light regimes, i.e., the degree of genotype-by-light regime interaction. In a rare and pioneering study on smolt genetics, Khaw et al. (2021) found significant heritability of a molecular marker for smoltification status [the Na+/K+-ATPase (NKA) seawater/freshwater (SW/FW) ratio, see Takvam et al., 2023), indicating that selection could achieve a more synchronized smoltification process. However, this study was limited in that it did not contrast the production protocols with the different day lengths and did not track individual fish from the freshwater stage to their performance (e.g., growth, survival) in seawater.
The objectives of the present study were therefore to estimate the impact of three different light smoltification regimes on the a) growth, survival, and animal welfare traits in the seawater phase of Atlantic salmon; b) the magnitude of the genetic variation in the traits; and c) the magnitude of the genetic and environmental correlations among the traits.
2 Materials and methods
2.1 Fish
The fish material originated from a total of 4,500 eyed eggs of the Mowi strain from 100 full-sib families (50 eggs per family that were pooled at the eyed-egg stage). The families were produced over a 3-week period in November 2019, but were temperature-controlled to the same sum day-degree at the eyed-egg stage in January 2020. Of the 100 families, 94 were the offspring of 47 sires and 94 dams produced using a nested one sire/two dam mating ratio, while the remaining six families were produced using a one sire/one dam mating ratio. A family was produced by stripping a male and a female and externally mixing the eggs in a bucket. Thus, the parents of each family were known.
2.2 Freshwater phase
An outline of the experimental design with the dates of the different activities and trait recordings is shown in Appendix 1.
2.2.1 Rearing conditions prior to treatment start
The eyed eggs were shipped from Mowi Genetics AS, Bergen, to the Aquaculture Research Station in Tromsø (ARST) on January 28, 2020, and incubated in one 200-L freshwater tank at 1.8°C, which was increased gradually to 7.9°C within the first 4 days. Hatching started on February 11, 2020, and was completed after 1 week. The hatchlings were start-fed on April 1, 2020, at which the water temperature was gradually increased to 10–12°C and the light regime changed from 24-h darkness to 24-h light. From May 19, 2020, the water temperature was changed to ambient temperature (4–12°C) until September 2020, when ambient temperature naturally decreased and kept constant at 8°C.
Malformed juveniles were constantly removed, and on July 22, 2020, a total of 4,090 fish were transferred into a 1,200-L tank. On December 15, 2020, lower mode fish smaller than 10 g were sorted out and the remaining 3,200 fish (mean body weight = 31.4 g, SD = 9.7 g) redistributed among four 1,200-L freshwater tanks. In early February, the fish were vaccinated with ALPHA JECT micro 6 (PHARMAQ®, Bergen, Norway) against Aeromonas salmonicida, Listonella anguillarum O1 and O2a, Vibrio salmonicida, Moritella viscosa, and infectious pancreatic necrosis (IPN) virus strain Sp.
The fish were fed ad libitum using automatic feeders with the commercial extruded pellet feed Nutra from Skretting AS, Stavanger Norway; Nutra Sprint during the early period and thereafter Nutra Olympic.
2.2.2 Tagging and tissue sampling
On March 15–22, 2021, a random sample of 3,000 fish (approximately 50 g) were individually tagged with a passive integrated transponder (PIT) tag, fin-clipped, and distributed among six circular 1,200-L tanks, with 500 fish per tank, on surplus flow-through water to secure optimal rearing conditions. Their body weights at tagging were not recorded. Prior to this, the fish were fasted for 24 h and anesthetized with benzocaine (60 ppm). The tags (ID-100A; www.trovan.com) were surgically implanted into the intraperitoneal cavity using syringes preloaded with transponders, and a small incision was made with a scalpel. Subsequently, a biopsy of the adipose fin was taken with scissors and stored in 96-tube racks consisting of 1-ml barcode-labeled tubes containing 750 μl of 100% ethanol. Finally, the fish were released into a wake-up bath and transferred into one of six tanks.
The biopsies of the fish, along with the biopsies of their parents, were stored at room temperature until shipping for genotyping (using a customized 56K Affymetrix Axiom SNP Array) for the purpose of parental assignment. The genotypes of the parental sires and dams and their offspring were quality controlled with Plink 1.9 software (Chang et al., 2015) using the following criteria. SNPs with a call rate<95%, a minor allele frequency<1%, and a Hardy–Weinberg p-value (Fisher’s exact test)<10−25 were removed. Lastly, samples with a call rate >95% were discarded. The parental assignments were based on the opposite homozygous (OH) method (Hayes, 2011; Calus et al., 2011; Ferdosi and Boerner, 2014) with 51,425 SNP markers after the quality control. The results using the OH method were conditional on the known matings. The sex of each fish was determined by the genetic markers located within the sdY gene (Houston et al., 2014).
2.2.3 The three light regimes
For a period of close to 6 weeks, from March 22 to May 3, 2021, the fish in duplicate tanks with freshwater were subjected to three different photoperiodic light regimes (i.e., 8L:16D, 12L:12D, and 24L:0D) on a constant water temperature of 8°C (312 degree-days) followed by 6 weeks (from May 4 to June 16, 2021) on a 24L:0D light regime, all at a light intensity of 200 lx (lumens per square meter) at water surface. The 12L:12D regime is much in use by the industry and the 8L:16D regime more closely resembles a real winter signal, while 24L:0D was chosen as a contrast to the two other regimes, but is also used by the industry particularly when producing large smolts. The fish in all tanks were fed ad libitum extruded salmon pellet feed (Nutra Supreme, Skretting AS) for 8 h of light using automatic feeders.
2.2.4 Recording of the smolt traits
Recording of the external smolt characteristics prior to seawater transfer was carried out from June 7 to 14, 2021. Fish were fasted 24 h, and the sampling of fish was randomized and carried out tank by tank. Fish were anesthetized with benzocaine (60 ppm) and the following traits recorded (see Table 1): body weight (to the nearest gram), body (fork) length (to the nearest 0.5 cm), and skin silveriness as a measure of smolt status (assessed visually according to the following criteria: 1 = parr marks clearly visible; 2 = silvery appearance, but still visible parr marks; or 3 = silvery skin, dark fin edges, and no parr marks visible). All traits were recorded electronically using FishReader and the software ZeusCapture (Trovan Ltd.). The proportion of fish with silvery skin was calculated as the ratio of the number of fish with score 3 and the total number of fish with scores 1, 2, and 3.
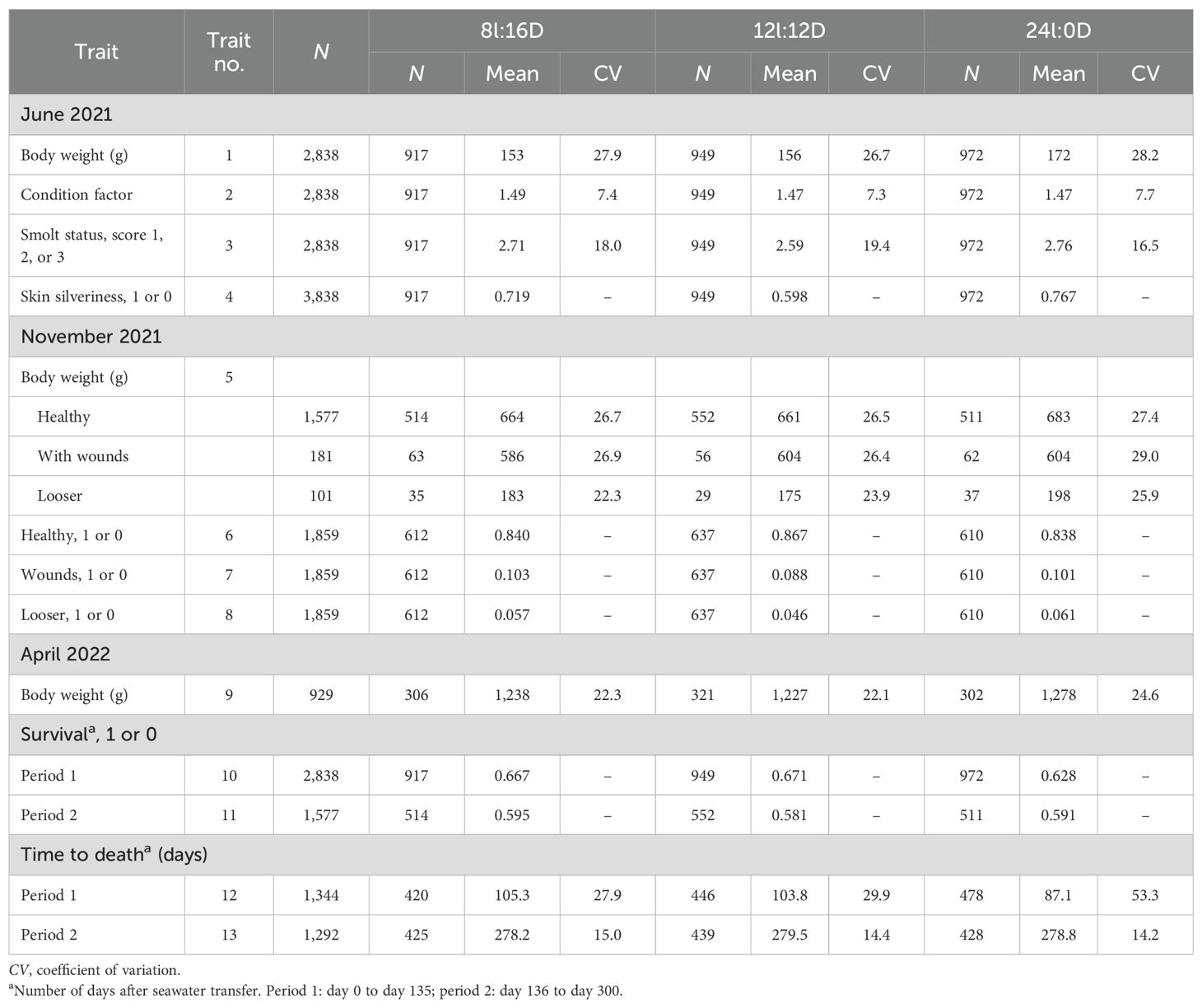
Table 1. Descriptive statistics for the studied traits for each light regime.
Condition factor was calculated as (Fulton, 1902; LeCren, 1951):
Gill biopsies from all fish were full transcriptome sequenced including the two alpha catalytic subunits (α1a and α1b) of NKA, the first one upregulated in freshwater and the second upregulated in saltwater (see Grønvold et al., 2024). In this study, the ratio of the transcriptomes of these two subunits (α1b/α1) was used as an indicator of whether the fish in the three light regime groups after the smoltification period were ready for seawater transfer.
2.3 Seawater phase
Appendix 1 shows the experimental design with the date of the different activities in the seawater phase. On June 23, 2021, a total of 2,910 1+ smolts were transferred in a transport tank to the seawater facility of ARST. Fish from the three light treatment groups were kept in one 5.5-m × 5.5-m × 5.5-m (~160 m3) sea net cage and were fed extruded salmon feed (Nutria Supreme, Skretting AS, with the pellet size adjusted according to fish growth) until the end of the experiment in April 2022.
At 8 weeks after seawater transfer, fish started to show signs of skin ulcers. Tissue from the fish sampled on August 25, 2021, showed the presence of Moritella vicosa, which can cause skin ulcers, and Vibrio wodanis (report from the Norwegian Veterinary Institute, 9480 Harstad), often found in ulcers (Sommerset et al., 2022). The condition worsened, and the mortality rate increased over the autumn. The prevalence of ulcers persisted throughout the winter, despite removal of the fish with wounds during the trait recordings of all fish in early November.
Mortality was monitored daily, and dead and moribund fish were removed from the sea cage, with the moribund fish euthanized with a lethal dose of benzocaine (150 ppm). Dead and euthanized fish were identified with a PIT tag reader and inspected visually for possible cause of death, after which their individual fork length and body weight were recorded.
Due to the high loss of fish and the associated welfare issues of alive fish, it was decided to end the experiment in April 2022, approximately 6 months ahead of schedule.
The traits recorded during the November 2021 and April 2022 recordings are shown in Table 1.
The thermal growth coefficient (TGC), as a measure of the overall growth rate, was calculated as:
where Bw is the body weight at the start (i) and the end (i − 1) of the actual period. The denominator (∑ d0) is the sum day-degrees of the seawater temperature over the period (Jobling, 2003). TGC was calculated for the period between stocking (June 2021) and the November 2021 recordings, as well as between the November 2021 recordings and the final April 2022 recordings.
2.3.1 November 2021 recordings
On November 2–5, 2022 (135 days after transfer to the net cage in the sea), batches of 20–30 fish were removed from the net pen with a dip net and were anesthetized in a seawater bath with benzocaine (50 ppm). The PIT tag of each fish was recorded along with their body weight, fork length, and health status, i.e., healthy, with wounds, and loosers without wounds (body weight<300 g). Those with wounds and the loosers were killed, while the healthy ones were put back into the cage.
2.3.2 April 2022 recordings
On April 19–22, 2022 (at the end of the 300-day seawater period), batches of 30 fish were removed from the net pen with a dip net and were euthanized in a seawater bath with a lethal dose of benzocaine (150 ppm) until gill ventilation had ceased. The PIT tag of each fish was recorded along with their body weight and fork length.
2.3.3 Survival in the seawater period
It was decided to record all the fish still alive one time in between their transfer to the net cage in the sea and their final harvest. Early November 2021 was chosen as the appropriate time point as the recordings could be performed at an acceptable seawater temperature (approximately 6°C) (Figure 1). Delaying this to, e.g., early December would imply handling of the fish at a lower water temperature (<5°C) that would invoke more stress on the fish.
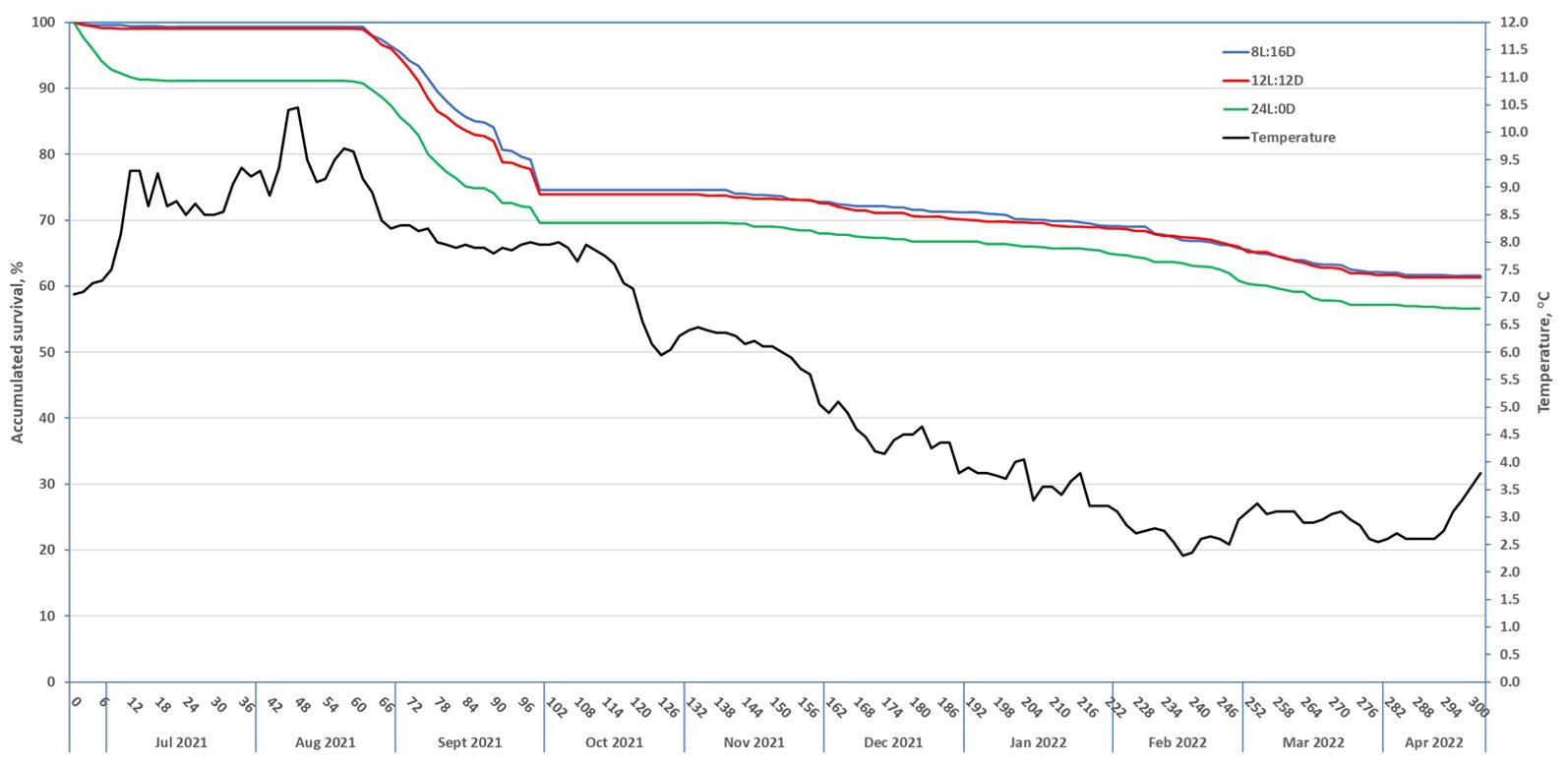
Figure 1. Survival (in percent) curves in the net cage in the sea for fish that received three different light regimes during the smoltification period [calculated based on the collected dead fish with readable passive integrated transponder (PIT) tags].
Three survival traits were defined based on whether a fish with a recorded PIT tag was recorded as dead (coded as 0) during two different periods in the net cage in the sea or alive (coded as 1) at the start or the end of the period.
- Period 1: Coded as alive on day 135 (November 5, 2021); otherwise, coded as dead (but not necessarily collected and thus recorded as dead) during days 0–135
- Period 2: Coded as alive on day 300 (April 19, 2022); otherwise, coded as dead (but not necessarily collected and recorded as dead) during days 136–300
In addition, another period, i.e., period 1a, was defined for fish coded as alive on day 0 (June 23, 2021) and that were collected and recorded as dead until day 58 (August 20, 2021), i.e., the period before the ulcers started.
2.4 Statistical analyses
2.4.1 Effect of light regime on the recorded traits
To estimate the effect of light regime on the 13 studied traits (Table 1), the animal (normally distributed traits) or the sire and dam (binary traits) model was used in the ASReml software (Gilmour et al., 2021). The model also accounted for the fixed effect of sex and the random tank effect.
Firstly, a bivariate linear mixed animal model was used for body weight in June 2021 and for each of the other continuous traits in Table 1, one at a time.
where y1 is a column vector for body weight in June 2021 and y2 is a column vector for trait 2, 3, 5, 9, 12, or 13; X is an incidence matrix that assigns each trait record to the appropriate level of the fixed light regime and sex; Z1 and Z2 are incidence matrices that assign each trait record to the appropriate level of the random effects of animal and tank, respectively; b is a vector of the fixed effects of light regime (with three levels) and sex (with two levels); u is a vector of random additive genetic values for animal with u ∼ N(0, Aσa2), where σa2 is the additive genetic variance and A is the additive genetic numerator relationship matrix; t is the vector of the random effect of tank with two levels within each of the three light regimes with t ∼ N(0, Iσt2), where σt2 is the tank variance and I an identity matrix; and e is a vector of random residual effects with e ∼ N(0, Iσe2), where σe2 is the residual variance.
For the binary traits in Table 1, a bivariate linear mixed sire and dam model was used for body weight in June 2021 as one of the traits (y1) and a mixed sire and dam threshold model for each of the binary traits (y2), i.e., trait 4, 6, 7, 8, 10, or 11 in Table 1, one at a time.
where Φ(·) is the cumulative standard normal distribution function assuming a normal underlying liability variable, l, determining the categorical outcome of the binary trait such that l ≤ 0 corresponds with y2 = 0 and l > 0 with y2 = 1). us and ud are random additive genetic values for sires and dams, respectively, with us = ud ∼ N(0, A σs2) and σs2 = σd2 = 0.25σa2; e is a vector of the random residual effect with e ∼ N(0, Iσe2); and σe2 is the residual variance including half of the additive genetic or the Mendelian sampling variance. The other parameters and matrices are as described in Model 1. For each of the bivariate traits, σe2 = 1.0. The least squares estimates of sex and light regime in the above model were transformed back to the observed binary scale.
In both Models 1 and 2, the body weight in June 2021 was included as a trait to account for possible bias in the estimates due to mortality that may be genetically correlated with each of the other traits.
For each of the studied traits, the importance of the random tank effect was expressed as: a) the ratio of its estimated variance component relative to the total variance (σtotal2 = σa2 + σt2 + σe2 for the continuous traits and σtotal2 = 2σe2 + σt2 + σe2 for the binary traits); b) the ratio of its estimated variance component and its standard error (σt2/see) (an output in the *asr file in ASREML); c) the ratio of the estimated variance component of the error of the reduced model (after omitting the tank effect) and the full model (for each of the continuous traits); and d) the ratio of the estimated variance component of the sire effect of the reduced model (after omitting the tank effect from the model) and the full model (for each of the binary traits as the error variance for each of the binary traits was set equal to unity both in the full and the reduced model).
The effect of light regime on each trait was examined using the Wald F-test statistics with two (numerator) and three (denominator) degrees of freedom. The random effect of tank was calculated using a log-likelihood ratio test (LRT; χ2 variate with 1 degree of freedom) (Wald, 1943; Lynch and Walsh, 1997) at a 5% critical value of 2.71 for continuous traits. For the binary traits, the random tank effect was determined using a Wald chi-square test (WT; χ2 variate with 1 degree of freedom) (Wald, 1943) as LRT is not an appropriate test statistic for binary traits.
An additional random effect common to full sibs other than additive genetics was also included in the above models, i.e., non-additive genetic effects. However, for all traits, this effect was not significantly different from zero and was therefore omitted from the models.
2.4.2 Genetic correlation between the same traits recorded under different light regimes
Estimates of the genetic correlations between the same traits for fish that received different light regimes were obtained from a three-trait linear mixed animal model for each of the continuous traits (Model 3) and from a mixed sire and dam threshold model for each of the binary traits (Model 4).
where the y for each of the three light regimes is a column vector of the actual trait at the June 2021, November 2021, and April 2022 recordings. The other parameters and matrices are as described in Models 1 and 2. In Models 3 and 4, the residual correlations between the same traits at the three different light regimes were set equal to zero as the trait was recorded on different animals. For the traits recorded in November 2021 and April 2022, the body weight in June 2021 was included as a fourth trait to account for possible bias in the estimates due to mortality that may be genetically correlated with the other traits.
For some of the traits (nos. 6, 7, 8, 12, and 13 in Table 1), the correlation structure was not positive-definite, or the log-likelihood of the model did not converge. For some traits (nos. 1, 2, 3, 4, 5, and 9 in Table 1), the log-likelihood converged, but the parameters did not converge, in most cases due to one, two, or all three of the estimates of the genetic correlations at the upper boundary (unity) of the parameter space. For those cases, one or two of the three genetic correlations were fixed close to the upper boundary (0.99) and at which both the log-likelihood and the parameters converged.
2.4.3 Estimates of genetic correlations between traits
For the traits in Model 3 for which the log-likelihood and the parameters converged and with a heritability significantly different from zero, the genetic correlations between the same traits for fish that received different light regimes were found to be high or very high.
Consequently, for these traits, the same traits for the three different light regimes at each of the three recordings could be looked upon as the same trait, thus reducing the number of traits in Table 1 from 13 to 5 traits in Table 2. Skin silveriness was not included as its genetic correlation with smolt status was unity.
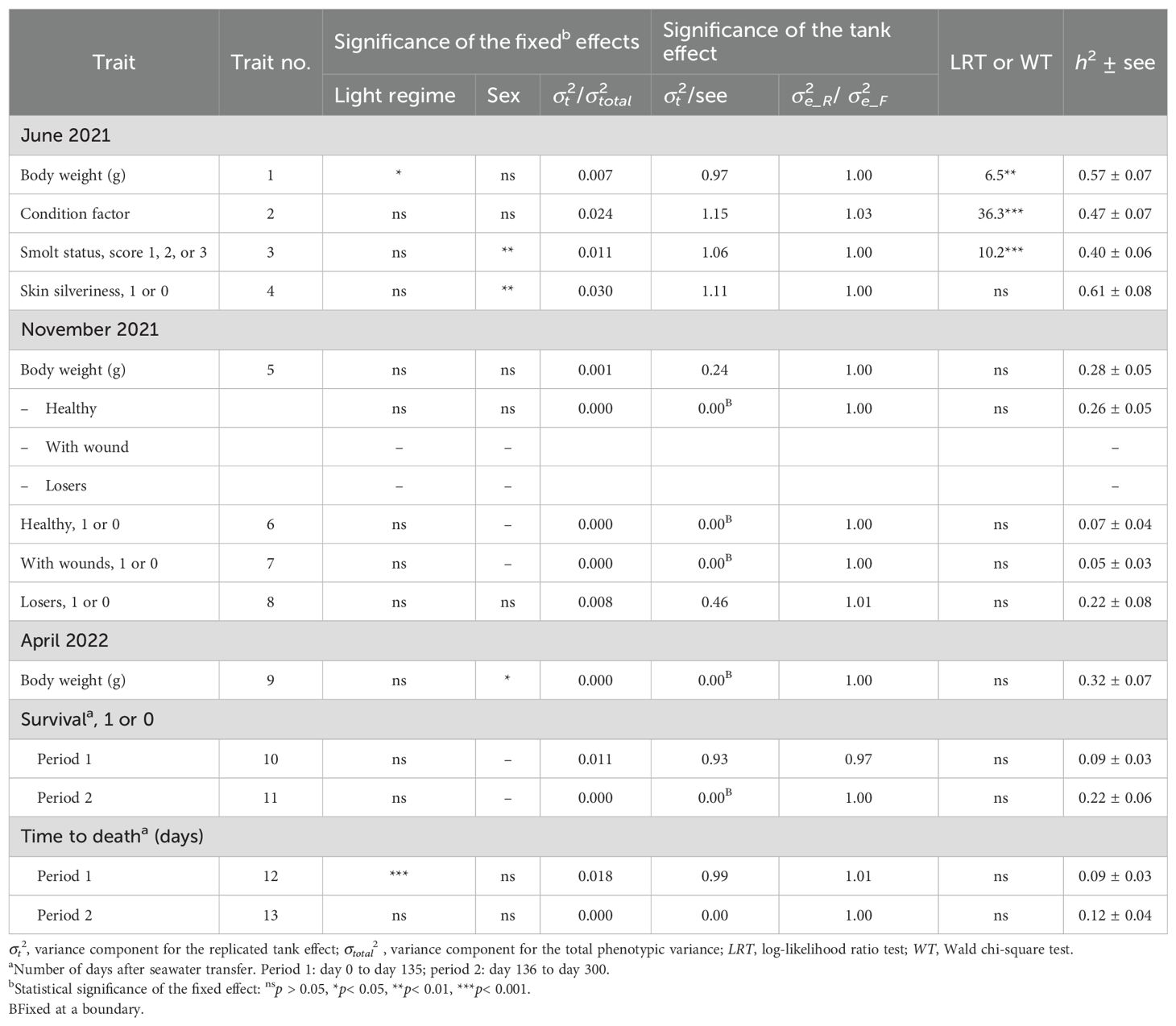
Table 2. Statistical significance of the two fixed effects and of the random tank effect and the estimated heritability (h2) for each trait.
Estimates of the (co)variances among these traits were obtained from the following multi-trait mixed animal model:
where y1, y2, y3, y5, and y9 are vectors of the animal trait records for each of the five traits (see Table 1); Xb is as described in Model 1 and was assigned to each trait; u is the additive genetic effect, where u = MVN[0, A ⨂ G]; A is the same as in Model 1; and G is the additive genetic (co)variance matrix between traits. Residual covariances were assumed to follow a multivariate normal distribution as follows MVN[0, I ⨂ R], where R is the residual covariance matrix among traits and I an identity matrix.
For each of the two binary survival and the two time-to-death traits in Table 1, covariances with the five traits in Model 5 were obtained from the multi-trait sire and dam models similar to Model 2, with the above-mentioned survival traits included in the model, one at a time, in addition to the five traits in Model 5.
2.4.4 Calculation of heritability and genetic correlations
The heritability estimate was reported from Models 1 and 2, while the genetic correlations were reported from Models 3, 4, and 5. For the traits in Model 2, the sire component of variance was set equal to the dam component using the model function and (dam,1) in the ASREML software. For the binary traits on the underlying liability scale, the residual variance (σs2) was set equal to unity; hence, only the variance components for sire (σs2) and full sib (σf2) need to be estimated.
The heritability for each trait from Model 1 was calculated as , and that for the sire and dam models (Model 2) as .
The relative magnitude of the tank effect was expressed as , where the total variance in the denominator was σa2 + σt2 + σe2 for the normally distributed traits and 2σs2 + σt2 + σe2 for the binary traits.
3 Results
3.1 Overall means
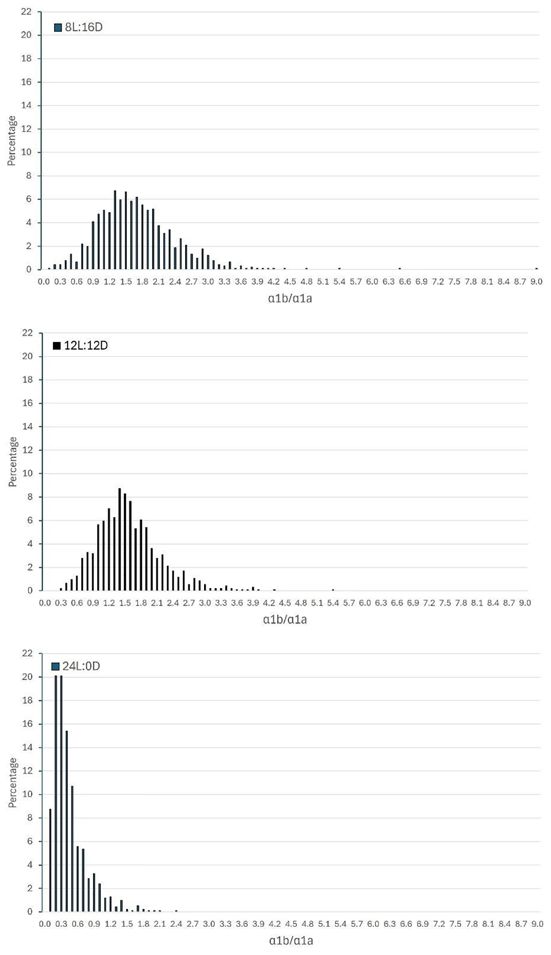
Just after the smoltification period, the ratio between the gene expressions of the two subunits (α1a/α1b) of Na+, K+ATPase (NKA) were much higher in the 8L:16D (mean 1.71, CV 44.1) and 12L:12D (mean 1.57, CV 38.9) groups and with a much lower CV compared to the 24L:0D group (mean 0.44, CV 71.6) (see Appendix 2).
During the 12-week smoltification period, only five fish died, one in five of the six tanks.
The descriptive statistics of the traits recorded during the three recording events are shown in Table 1. At the June 2021 recordings, the fish on the two short-day regimes had, on average, lower body weights, but a higher proportion of fish with silvery skin than those on the long-day regime, while the fish on the three light regimes had very similar condition factor. At the November 2021 recordings, 84%–85% of the fish were classified as healthy, 9%–10% with wounds, and 5%–6% as loosers, with relatively small differences between the three light regimes (10.3%, 8.8%, and 10.2% with wounds and 5.7%, 4.6%, and 6.1% as loosers). At the April 2022 recordings, the average body weights of the fish at the three light regimes were very similar.
As only a fraction of the dead fish could be collected from the cage (see Section 3.2), survival data could only be obtained for periods 1 and 2 based on the number of fish stocked in June 2021, the number of fish recorded in November 2021, and the number of fish slaughtered in April 2022.
The fish started developing ulcers relatively early during the trial. With the increased mortality in period 2—and at the end of this period driven by the continuous culling of fish with severe welfare problems due to wounds—the experiment was terminated after 300 days in the net cage, i.e., 3–4 months earlier than the original plan. In period 2, the survival rates of the fish on the three light regimes were very similar (58%–60%). The overall survival in the entire seawater period was 34.8% [929/(2,938 − 181 − 89)]. During the seawater period, the person responsible for the trial followed local fish welfare regulations.
The overall mean TGC was 2.80 (SD = 0.85) in period 1 and was 3.02 (SD = 0.89) in period 2 for fish with recorded body weights at the November 2021 and April 2022 recordings, respectively. In period 1, the mean TGC was 2.96 (SD = 0.65) for those classified as healthy, 2.61 (SD = 0.60) for those with wounds, and 0.26 (SD = 0.56) for the loosers. At the November 2021 recordings, the mean condition factor of the loosers (1.11 ± 0.02) [coefficient of variation (CV) = 0.160, min = 0.86, max = 1.78] was significantly lower (p< 0.001) than that of those with wounds (1.33 ± 0.01; CV = 0.084, min = 0.97, max = 1.59) and of those classified as healthy (1.39 ± 0.003; CV = 0.081, min = 0.93, max = 1.79).
3.2 Survival curves
During the entire seawater period, a total of 1,141 fish with readable PIT tags were collected: 101 in period 1a, 778 in period 1, and 363 in period 2. In period 1, the 778 dead fish accounted for 79.5% [=778/(2,838 − 1,859)] of the dead fish in this period; in period 2, the 363 dead fish accounted for 56.0% [=363/(1,577 − 929)] of the dead fish in this period. For the entire seawater period, 70.0% [(778 + 363)/(2,838 − 181 − 89 − 929)] of the dead fish were recorded as dead with readable PIT tags.
Of the 101 dead fish collected in period 1a, six was from the 8L:16D light regime, nine from 12L:12D, and 86 from 24L:0D, with average number of 6.8, 3.9, and 5.6 days to death, respectively. Due to the low number of dead fish in two of the three light regimes, survival in period 1a was not considered a reliable binary trait or a time-to-death trait.
Based on the collected dead fish with PIT tags, the survival curves in the entire seawater period were calculated for each of the three light regimes (Figure 1), with a substantially higher accumulated survival (63.6%, 64.9%, and 58.4% for groups 8L:16D, 12L:12D, and 24L:0D, respectively) than the overall “true survival” of 34.8% reported above. The fish in group 24L:0D showed increased mortality from day 2 after seawater transfer, with an accumulated mortality of 8.9% during period 1a, while groups 12L:12D and 8L:16D showed mortality rates of 1.0% and 0.7%, respectively. From day 59 onwards until harvest on day 300 in April, the mortality rates were very similar for the fish on the three light regime groups. These survival curves are overestimated as only 79.5% and 56.0% of the dead fish were collected and thus recorded as dead in periods 1 and 2, respectively.
3.3 Effect of light regime
The body weight in June 2021 and the time to death in period 1 were the only traits significantly affected by light regime (Table 3). The fish on the 24L:0D regime were, on average, 15.0 ± 4.0 g (10.0%) and 18.0 ± 4.0 g (12.2%) heavier than those on light regimes 12L:12D and 8L:16D, respectively (Figure 2), but died earlier, on average 15.7 ± 3.4 and 17.7 ± 3.4 less days to death than the fish on the 12L:12D and 8L:16D regimes, respectively (p< 0.001).
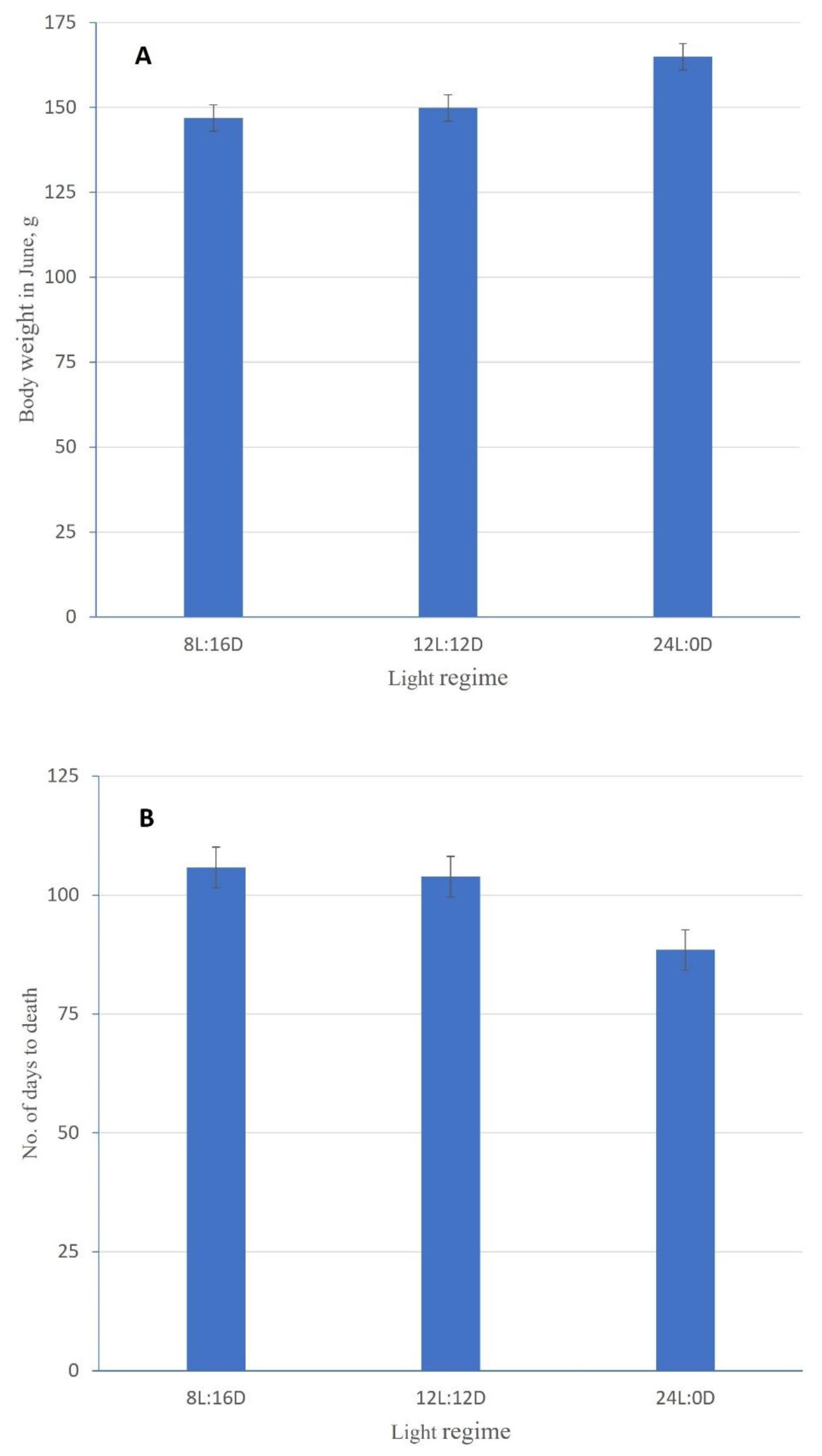
Figure 2. Least square means estimates (±see) for the body weight in June 2021 (A) and the time to death (B) in period 1 for each of the three light regimes.
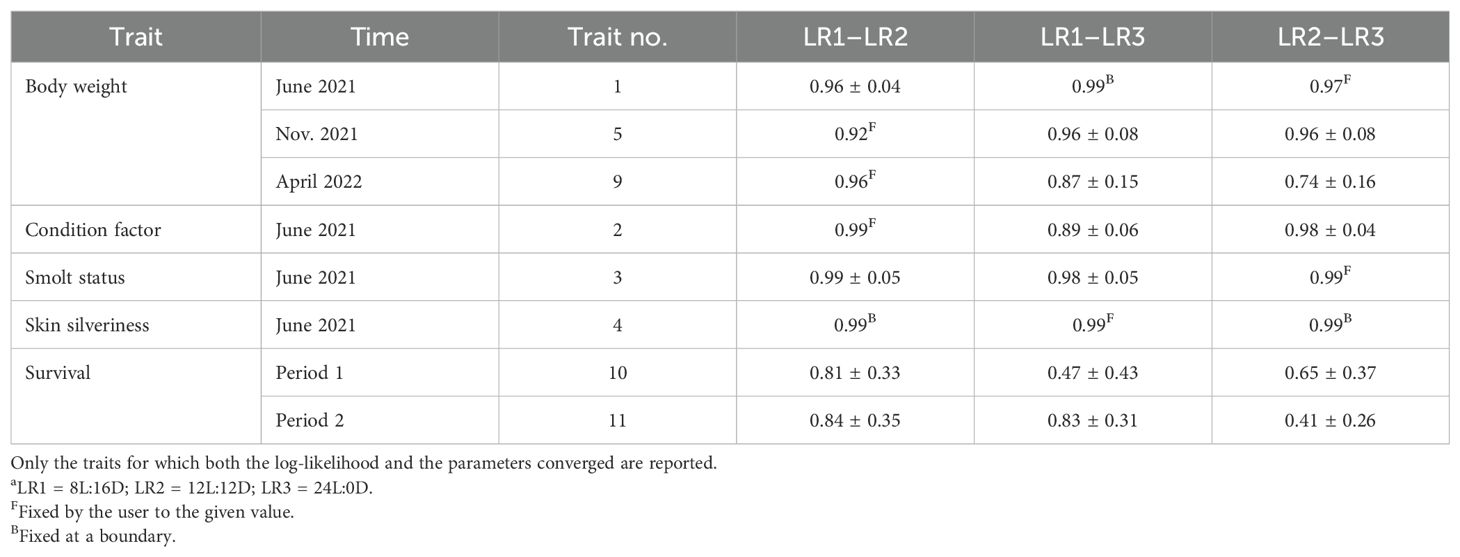
Table 3. Estimates of the genetic correlations between the same traits recorded for the different light regimesa..
Figure 3 shows that, prior to seawater transfer, the survivors from all three light regimes in period 1a, period 1, and period 2 had more favorable values for the smolt indicator traits than the fish that died during these periods, i.e., lower condition factor, higher smolt status score, and higher skin silveriness proportion. In addition, the survivors from all three regimes had a higher body weight in periods 1a and 2, but not in period 1. Significant differences between light regimes for the differences between the survivors and the dead were observed for all traits in period 1a and for the condition factor in period 1. In period 2, no significant difference between the survivors and the dead between light regimes was found for any of the four traits recorded prior to seawater transfer.
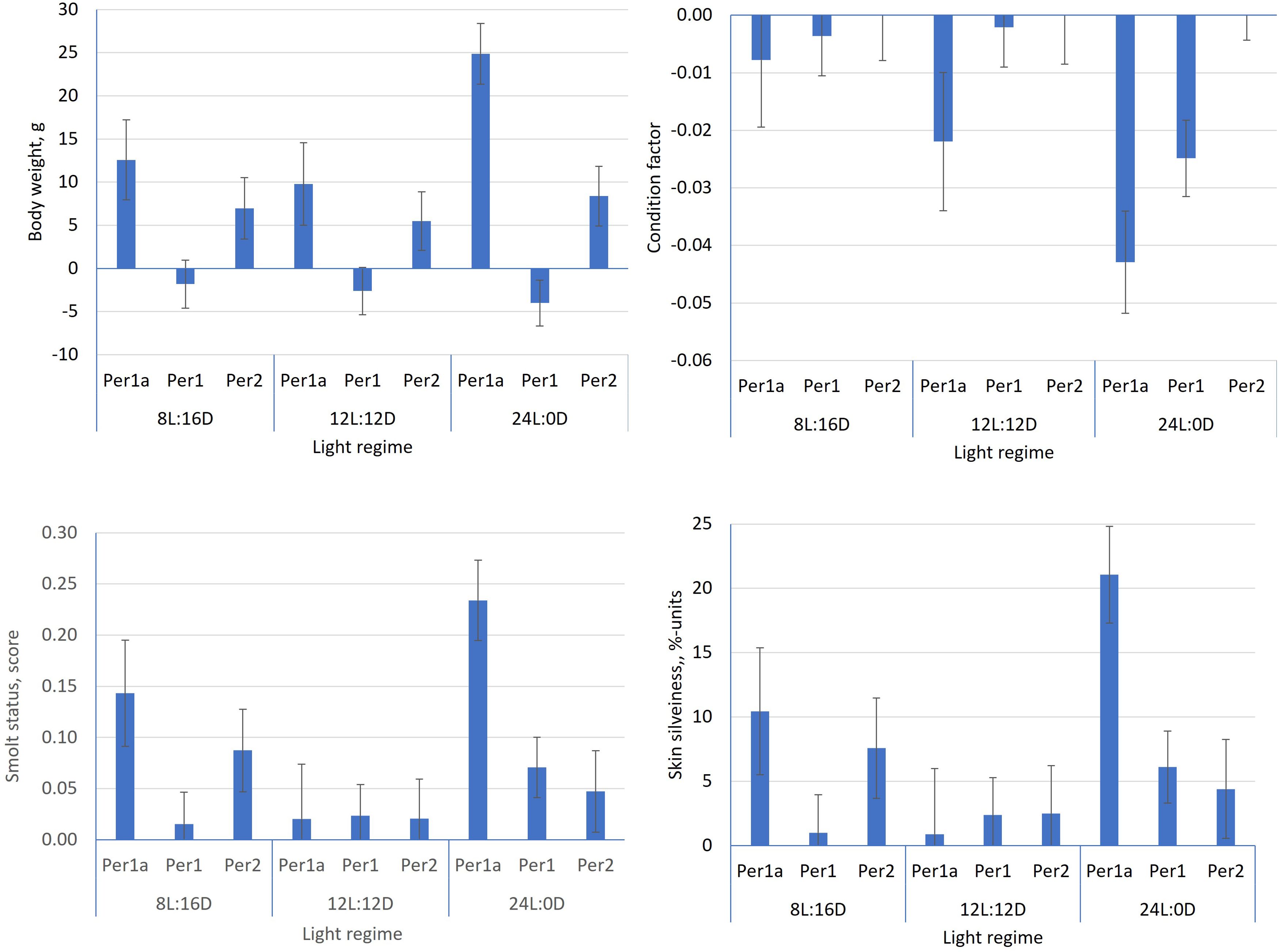
Figure 3. Estimates of the differences (±see) between the survivors and the dead (trait of survivor minus trait of dead) in period 1a, period 1, and period 2 for the traits recorded in June 2021 for each of the three light regimes.
3.4 Effect of sex
Table 3 shows that sex had a significant effect on only three of the studied traits, i.e., smolt status score and skin silveriness in June 2021 and body weight in April 2022. In June 2021, the males had a significantly higher score for smolt status (0.044 ± 0.016, 1.6%, p< 0.01), a significantly higher liability threshold for skin silveriness (0.151 ± 0.050, 4.8% units on the observed scale, p< 0.01), and, in April 2022, a significantly higher body weight (34.9 ± 15.8 g or 2.9%, p< 0.05).
3.5 Effect of tank within light regime
The body weight, condition factor, and smolt status score in June 2021 were the only traits for which the tank effect was significantly different from zero (p< 0.05) (Table 3). Of the total variance of each trait, the tank effect accounted for 0.7% for body weight, 2.4% for condition factor, and 1.1% for the smolt status score. For majority of the other traits, the tank effect accounted for an even smaller proportion of the total variance.
For survival and time to death in period 1, the magnitude of the tank effect is illustrated in Figure 4. For survival, the estimated tank solutions for each of the six tanks are shown both on the liability scale and the back-transformed observed scale. On the observed scale, the percentage estimated survival rates for the fish from the two replicated tanks were 91.6 and 90.0 for the 08L:16D group, 92.2 and 91.1 for the 12L:12D group, and 87.9 and 78.8 for the 24L:0D group. The power of detecting the observed survival difference in period 1 between the 24L:0D regime (83.4%) and the mean survival of the 8L:16D and 12L:12D regimes [0.5 × (90.9 + 91.8) = 91.4%], a 8.0% unit difference, was found to be only 0.70 (two-sided test with a 0.05 type 1 error), i.e., when calculated on the observed binary scale based on the estimated variance component of the tank effect within light regime as a proportion of the sum of the variance component of the tank effect and the variance component within tank . On the liability scale, this ratio was substantially higher .
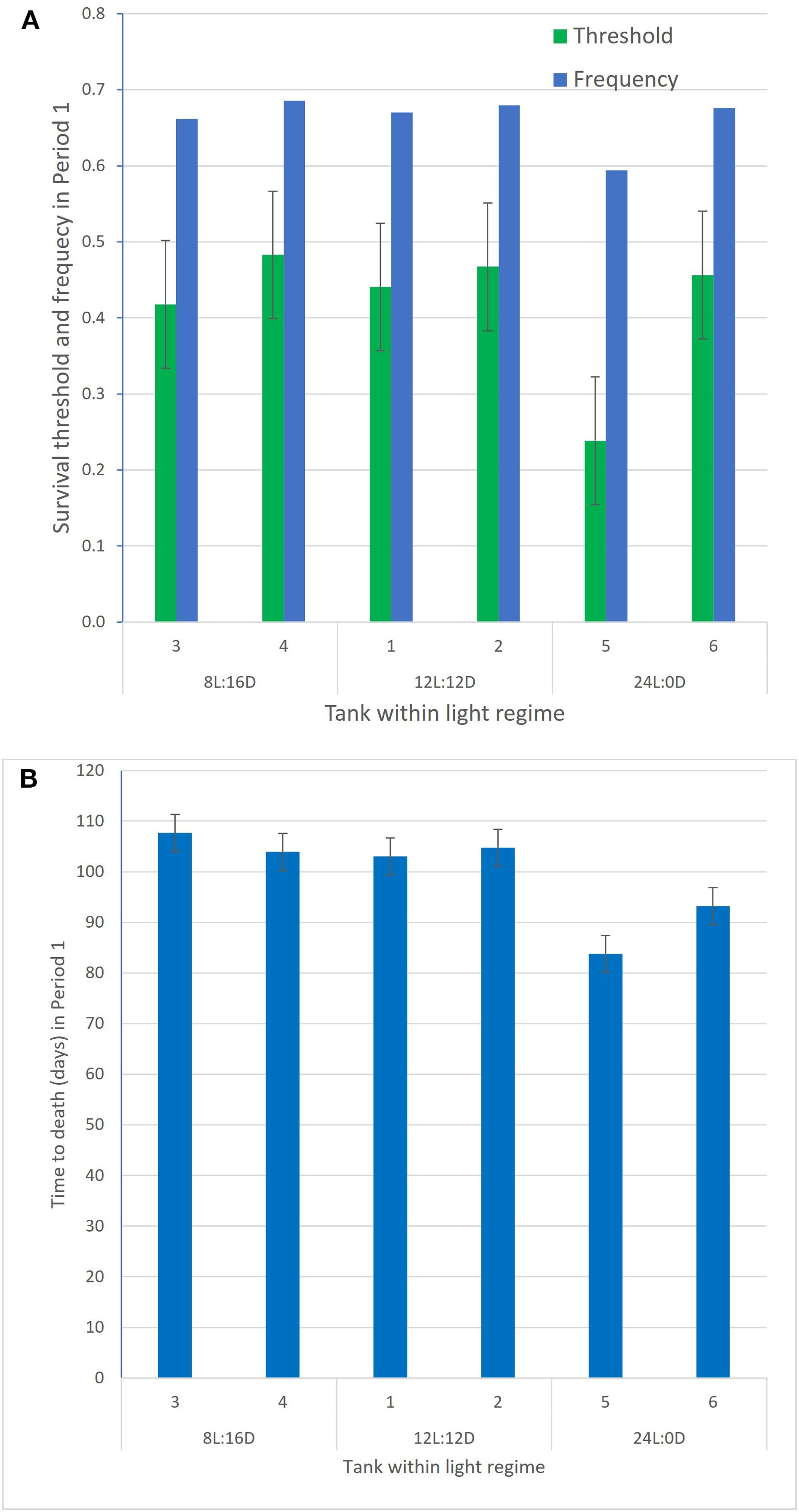
Figure 4. Estimated liability thresholds for survival and their corresponding back-transformed survival frequency values (A) and time to death (B) in period 1 for each of the two tanks within each of the three light regimes (data from Grønvold et al., 2024).
3.6 The same traits at the different light regimes as different traits
For 8 of the 13 studied traits, the genetic correlations between the same traits in the three different light regimes were generally high to very high (Table 4). However, for the three binary traits—healthy, wounds, and loosers—the correlation structure was not positive-definite; for the three time-to-death traits, the log-likelihood and the parameters did not converge, most likely due to the relatively low number of observations per light regime for these traits (Table 1).
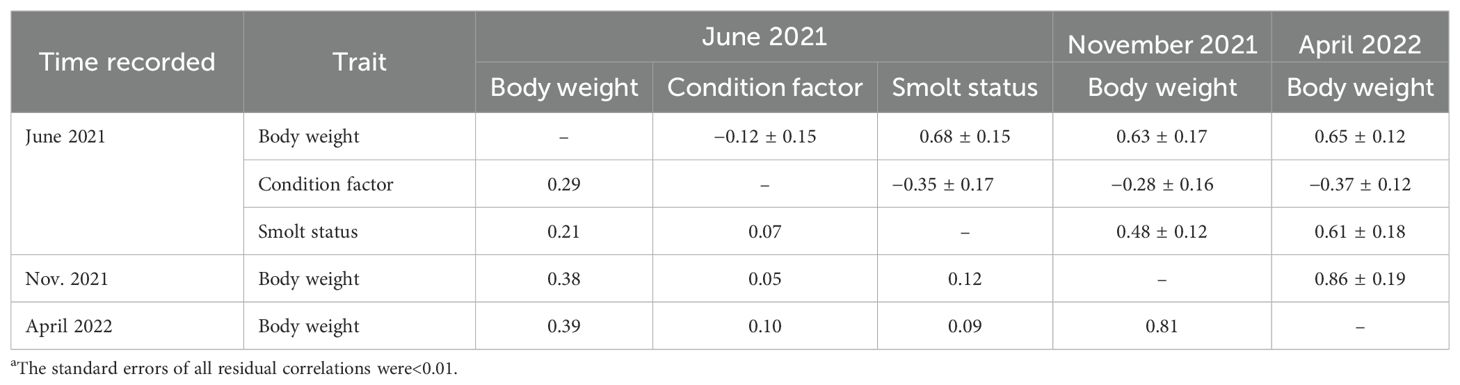
Table 4. Estimates of the genetic (above diagonal) and residuala (below diagonal) correlations between the growth-related traits measured during the freshwater and seawater production periods.
Consequently, eight of the traits at each light regime could be looked upon as the same trait and the estimates of heritability and genetic correlations based on the trait records from all three light regimes. From the total number of records in Table 1, it can be calculated that, for the normally distributed traits recorded in June 2021, November 2021, and April 2022, the average numbers of fish/family were 28.4, 18.6, and 9.3, respectively; for the two binary survival traits, these were 28.4 in period 1 and 15.9 in period 2. For the two time-to-death traits, the average number of fish/family was 13.4 in period 1 and was 12.9 in period 2. Thus, for all traits, the number of records per family is close to or larger than 10, which should result in quite reliable genetic parameters.
3.7 Heritability
The heritability estimates for the external indicator traits for smolt development (condition factor, smolt status score, and skin silveriness) were moderate, as were the heritability estimates for loosers in November 2021 and the survival and time to death in period 2 (Table 3). The heritability estimates for healthy fish and wounds in November 2021, and for both survival and time to death in period 1, were all low (Table 3).
3.8 Genetic and residual correlations among the traits
Estimates of the genetic correlation of the binary traits healthy, wounds, and loosers in November with the other traits could not be obtained as none of the (co)variance components converged. The genetic correlation between skin silveriness (in percent) and smolt status (score) was close to unity (0.9985); consequently, only smolt status was included in further analyses.
Estimates of the genetic and residual correlations among the five traits included in Model 5 are given in Table 2. The genetic correlations of the smolt status in June 2021 with the body weights in June 2021, November 2021, and April 2022 were all moderately positive and thus favorable. In June 2021, the genetic correlation between smolt status and condition factor was negative, as were the genetic correlation of the smolt status in June with the body weights in November 2021 and April 2022.
The genetic correlations between the body weights in June, November, and April were all positive, and those of June with November 2021 (0.63) and April 2022 (0.65) were lower than those between November and April (0.86). The magnitudes of the correlations between these traits, and of their heritability estimates (Table 3), were similar to earlier published estimates in Atlantic salmon (Gjerde et al., 1994). The remaining genetic and residual correlations in Table 2 were all close to zero.
3.9 Genetic correlations of the survival traits with the other traits
The genetic correlations of the survival traits with the other traits recorded in June 2021, November 2021, and April 2022 (Table 5) were all low to medium in magnitude, with large standard errors, and were not significantly different from zero (p > 0.05).
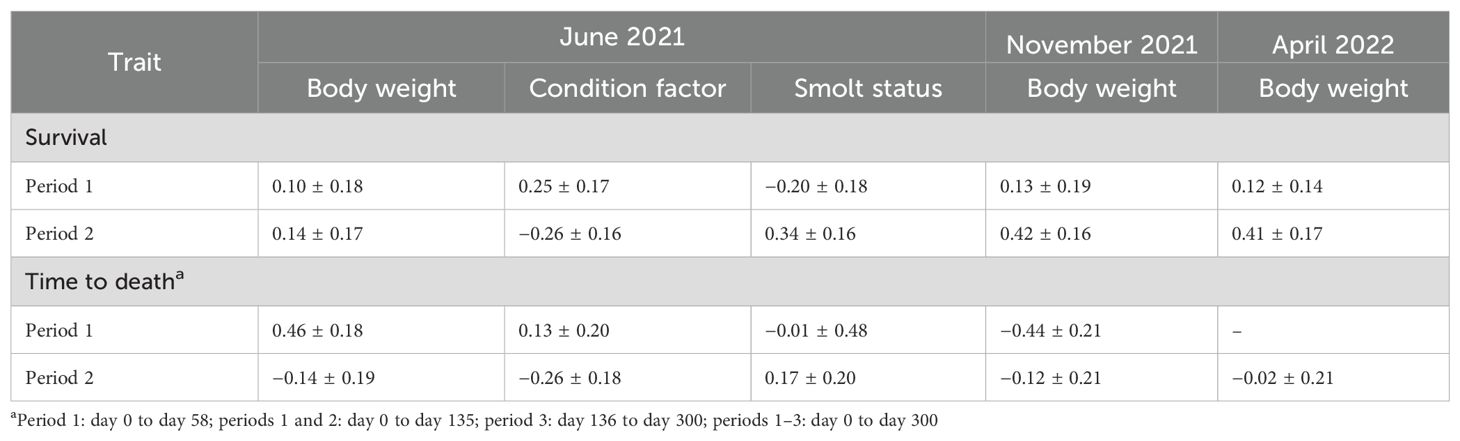
Table 5. Estimates of the genetic correlations of the growth-related traits with the two binary survival and time-to-death traits.
4 Discussion
The objective of this study was to assess the impact of three light smoltification regimes and quantitative genetics, as well as their interactions, in Atlantic salmon on the development of external smolt characteristics and on the survival, growth, and general fish welfare characteristics in the seawater phase.
4.1 Short-day regimes give more synchronous smolt development
In line with similar experiments (Striberny et al., 2021; Ytrestøyl et al., 2022), we found that at the end of the 12-week light regimes, about 2 weeks prior to seawater transfer, fish on the two short-day regimes (i.e., 8L:16D and 12L:12D) had significant lower body weights compared with fish on the continuous light regime (i.e., 24L:0D), likely an effect of the shorter day length on appetite (Bjørnsson et al., 1989). After 5 and 10 months in the sea, however, there was no longer a significant effect of light regime on body weight. This “catching up in sea” effect is also in agreement with previous studies (Striberny et al., 2021; Montgomery et al., 2025) where fish with reduced growth during short-daylight regimes grow better in seawater compared with fish kept on continuous light.
Atlantic salmon on a short-day regime are expected to have a reduced condition factor at the end of the light regime compared with salmon on a continuous long-day regime (Sigholt et al., 1998). This was also found for fish transferred to seawater at an average body weight of 100 or 200 g (Ytrestøyl et al., 2022), but not observed in this study, as was the case for smolt transferred to seawater at an average body weight of 600 g (Ytrestøyl et al., 2022). In this study, a non-significant effect of light regime was also found on the condition factor, smolt status, and skin silveriness. Myklatun et al. (2023) also reported no significant effect of light regime (12L:12D vs. 24L:0D) on the condition factor prior to seawater transfer.
The fish classified as loosers at the November 2021 recordings were, on average, much smaller (72.2%) than those classified as healthy (186 vs. 669 g). Their mean condition factor (1.11; min = 0.86, max = 1.78) was much higher than the Atlantic salmon in Figenschou et al. (2025) that 69 days post seawater transfer were classified as stunted or runts (0.77; mean body weight = 78.4 g, CV = 0.154) compared with their normal fish (1.16; mean body weight = 353.6 g, CV = 0.150). In their study, stunted fish were identified by their distinct smaller and leaner appearance, with a condition factor below 0.9, as defined in previous studies (Stradmeyer, 1994; Van Rijn et al., 2021). Consequently, at the November 2021 recordings, the slow-growing fish in period 1, without wound but with a condition factor not typical for runts, were classified as loosers. That their proportions did not differ between the three light regimes indicates that their much lower growth rate may be caused by being affected by, e.g., the skin ulcer that occurred 2 months after their transfer into the net cage in the sea rather than being poorly smoltified.
Similarly, the similar proportion of fish with wound among the three light regimes, and with a condition factor marginally lower than that of the healthy fish, indicates that their lower growth rate compared with healthy fish may also be caused by being affected by, e.g., the skin ulcer rather than being poorly smoltified.
For wild salmon, circannual changes in day length are important cues that regulate the timing of smoltification and the subsequent migration to sea (Hoar, 1988). Experimental day length manipulations have shown that both a simulated natural increase in day length in the spring and an abrupt increase in day length to mimic the transition from a winter to a spring photoperiod impact hormonal regulation and the development of smolt physiology and seawater adaptations (McCormick et al., 2007). It is therefore possible that the fish on constant light in our study were less “synchronized” in their smolt development. This corresponds with several studies reporting a lack of synchronization in smolt development when Atlantic salmon are kept on a long photoperiod (Björnsson et al., 1995; Sigholt et al., 1995; Duncan and Bromage, 1998).
Several results from this study support this notion. Firstly, over the first 58 days in the net cage in the sea (period 1a), mortality was, to a large degree, confined to fish raised on the 24L:0D regime (Figure 1). Secondly, the differences in the trait values measured prior to seawater transfer between fish that survived and those that died in period 1 were largest in the 24L:0D group (Figure 3). Majority of these differences were small relative to their trait means and large standard errors, but support higher group-level uniformity and better smolt physiology at the time of seawater transfer for fish on 8L:16D and 12L:12D compared with fish on 24:0D. More optimal smolt physiology in short-day-exposed fish is also backed up by RNA sequencing (RNAseq) data from the same experiment recently published in Grønvold et al. (2024). Thirdly, the much higher average α1a/α1b ratio in the 8L:16D and 12L:12D groups with a much lower CV as compared to the 24L:0D group (see Appendix 2) indicate both a better and more uniform smolt quality of the fish in the two first mentioned groups (Khaw et al., 2021; Takvam et al., 2023), were much higher (i.e., better smolt physiology) in 8L:16D (mean = 1.71, CV = 44.1) and 12L:12D (mean = 1.57, CV = 38.9) compared with that in 24L:0D (mean = 0.44, CV = 71.6) (see Appendix 2). The ratio (α1b/α1a) values for the 8L:16D and 12L:12D groups were similarly to the same ratio values reported by Khaw et al. (2021) from the gill biopsies of 0+ smolts smoltified under a 12L:12D–24L:0D light regime and sampled at their considered optimum time of seawater transfer (mean = 1.86, CV = 59.6), while lower mean ratios and higher variations among the fish were found from the gill biopsies of 1+ smolts (mean = 1.09, CV = 77.5), also smoltified under a 12L:12D–24L:0D light regime and sampled at their considered optimum time of seawater transfer.
The effect of light regime on the time-to-death trait in period 1 was highly significant in fish on the long-day regime experiencing less days to death than those on the two short-day regimes. However, for the binary survival trait during the same period, the effect of light regime was non-significant, despite the much higher number of dead fish in the 24L:0D group than in the 8L:16D and 12L:12D groups during the first 58 days (period 1a) in the sea. This could be due to the binary nature of this trait and the very low statistical power of the experiment (see Section 3.5). Increasing the total number of tanks in the freshwater phase to 9 or 12 tanks, instead of six as in this experiment, would imply a relative marginal increase in the experimental cost as the fish had to be kept in these tanks for 12 weeks only, or only 6 weeks if the fish were PIT-tagged after the first 6 weeks and could be pooled and reared in a common tank on the 24L:0D regime, a strategy that may also have reduced the magnitude of the tank effect and increased the power of the experimental design, and thus the probability to detect also a smaller true difference between the light regimes for the studied traits.
A positive effect of increased body size at seawater transfer on survival in both periods 1 and 2 was observed for all three light regimes (Figure 3), and most likely one of the reasons for smolt producers tending to produce larger smolts. However, Ytrestøyl et al. (2022) showed that the choice of optimal smolt size and smoltifying strategy is not always a straightforward procedure as larger smolts (600 g) have been found to perform worse in the sea compared with fish transferred at a smaller size (100 and 200 g). Within each size group, the positive effect of a short-day regime (12L:12D) was seen as a slightly higher TGC in seawater than for fish on the continuous light regime (24L:0D), but with no effect of light regime on survival in seawater.
4.2 Can we breed for a more robust smolt?
A robust and good quality smolt can be defined as young salmon that shows both high survival and growth after being transferred from freshwater to seawater. However, we are aware of only one study on improving smolt quality through selective breeding (Khaw et al., 2024).
In this study, it was found that the condition factor and the smolt status score, both considered as external indicator traits for smolt development, have a significant genetic component and thus can be improved through selection. A significant genetic variation in condition factor was also reported by Khaw et al. (2021) in both 0+ and 1+ smolts. Significant genetic variation was also found for survival and time to death in both periods 1 and 2, as well as for growth measured as body weight until seawater transfer and after 135 and 300 days after seawater transfer. The reliability of these heritability estimates was confirmed by the high genetic correlations between the same traits recorded on siblings exposed to the three different light regimes. Consequently, groups of siblings from the same families smoltified under different light regimes are expected to rank similarly for these traits. It is therefore quite irrelevant under which light regime the test fish and the breeding candidates in a nucleus breeding population, or the commercially fish produced from selected breeders in a breeding program, are being smoltified.
Therefore, selection for improved smolt status through selection for, e.g., a lower condition factor and/or a higher smolt status score should result in a favorable selection response in the traits irrespective of which light regime the fish are being smoltified. However, the low genetic correlations of these smolt indictor traits with survival and time to death limits their value as indirect selection criterion traits and are therefore not likely to produce any significant favorable correlated genetic gain in survival in the seawater period (Falconer and McKay, 1989), which should be the main breeding objectives for improved smolt development and quality. To what extent the above genetic parameters are also valid for fish of an average larger body size at the time of seawater transfer than the fish in this study needs further research.
A more effective strategy to improving both the survival and growth in the seawater phase might be to perform direct selection for increased survival and growth during the first months in the sea. This would require recording the individual body weights of all fish just before or after the smoltification regime, as well as the body weights of all the survivors after these, e.g., 4–6 months after seawater transfer. The use of genomic selection (Meuwissen et al., 2016; Boudry et al., 2021) would increase the selection response for the survival traits, particularly through higher accuracy and intensity of selection through both higher accuracy and selection intensity.
Finally, the effect of the observed skin ulcers with increased mortality and skin wounds, particularly in period 2, may have masked differences in the studied trait values between the three studied light regimes, as well as the magnitude of the genetic correlation of the survival traits and the other traits. However, to what extent this may have affected the reported results can only be inferred upon in a follow-up study.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The experiment was performed according to EU regulations concerning the protection of experimental animals (Directive 2010/63/EU). Appropriate measures were taken to minimize pain and discomfort. The experiment was approved by the Norwegian Food and Safety Authority (FOTS id. number 25658). The studies were conducted in accordance with the local legislation and institutional requirements. Written informed consent was obtained from the owners for the participation of their animals in this study.
Author contributions
BG: Conceptualization, Methodology, Writing – review & editing, Writing – original draft, Project administration, Data curation. SB: Writing – review & editing. DH: Data curation, Writing – review & editing, Conceptualization, Project administration. TY: Writing – review & editing, Data curation. TM: Data curation, Writing – review & editing. EJ: Writing – review & editing. AS: Data curation, Writing – review & editing. SS: Conceptualization, Writing – review & editing, Project administration.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. Fund was obtained from FHF – Norwegian Seafood Research Fund through the project “Production protocols and breeding strategies for synchronized smoltification”, project no. 901589, January 1, 2020 to March 31, 2024.
Conflict of interest
Author SB is employed by MOWI Genetics ASA.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Aunsmo A., Bruheim T., Sandberg M., Skjerve E., Romstad S., and Larssen R. B. (2008). Methods for investigating patterns of mortality and quantifying cause-specific mortality in sea-farmed Atlantic salmon Salmo salar. Dis. Aquat. Organ 81, 99–107. doi: 10.3354/dao01954
Basulto S. (1976). Induced saltwater tolerance in connection with inorganic salts in the feeding of Atlantic salmon (Salmo salar L.). Aquaculture 8, 45–55. doi: 10.1016/0044-8486(76)90018-1
Bjornsson B. T., Thorarensen H., Hirano T., Ogasawara T., and Kristinsson J. B. (1989). Photoperiod and temperature affect plasma growth hormone levels, growth, condition factor and hypoosmoregulatory ability of juvenile Atlantic salmon (Salmo salar) during parr-smolt transformation. Aquaculture 82, 77–91. doi: 10.1016/0044-8486(89)90397-9
Björnsson B. T., Stefansson S. O., and Hansen T. (1995). Photoperiod regulation of plasma growth hormone levels during parr-smolt transformation of Atlantic salmon: implications for hypoosmoregulatory ability and growth. Gen. Comp. Endocrinol. 100, 73–82. doi: 10.1006/gcen.1995.1135
Boeuf G. (1993). “Salmonid smolting: A pre-adaptation to the oceanic environment,” in Fish ecophysiology. Eds. Rankin J. C. and Jensen F. B. (Chapman and Hall, London), 105–135.
Boudry P., Allal F., Aslam M. L., Bargelloni L., Bean T. P., Brard-Fudulea S., et al. (2021). Current status and potential of genomic selection to improve selective breeding in the main aquaculture species of International Council for the Exploration of the Sea (ICES) member countries. Aquaculture Rep. 20. doi: 10.1016/j.aqrep.2021.10070
Calus M. P. L., Mulder H. A., and Bastiansen J. W. M. (2011). Identification of Mendelian inconsistencies between SNP and pedigree information of sibs. Genet. Selection Evol. 43, 44. doi: 10.1186/1297-9686-43-34
Chang C. C., Chow C. C., Tellier L. C. A. M., Vattikuti S., Purcell S. M., and Lee J. J. (2015). Second-generation PLINK: rising to the challenge of larger and richer datasets. GigaScience 4, 7. doi: 10.1186/s13742-015-0047-8
Duarte V., Gaetano P., Striberny A., Hazlerigg D., Jørgensen E. H., Fuentes J., et al. (2023). Modulation of intestinal growth and differentiation by photoperiod and dietary treatment during smoltification in Atlantic salmon (Salmo salar, L.). Aquaculture 566, 739164. doi: 10.1016/j.aquaculture.2022.739164
Duncan N. J. and Bromage N. (1998). The effect of different periods of constant short days on smoltification in juvenile Atlantic salmon (Salmo salar). Aquaculture 168, 369–386. doi: 10.1016/S0044-8486(98)00363-9
Evans D. H., Piermarini P. M., and Choe K. P. (2005). The multifunctional fish gill: Dominant site of gas exchange, osmoregulation, acid-base regulation, and excretion of nitrogenous waste. Physiol. Rev. 85, 97–177. doi: 10.1152/physrev.00050.2003
Falconer D. S. and McKay T. (1989). Introduction to quantitative genetics. 3 (Harlow, Essex, UK/New York: Longmans Green/John Wiley & Sons).
Ferdosi M. H. and Boerner V. (2014). A fast method for evaluating opposing homozygosity in large SNP data sets. Livestock Sci 166, 35–37. doi: 10.1016/j.livsci.2014.05.026
Figenschou A. J. H., Lund H., Mayer I., Svendby M. B., Nørstebø S. F., Olofsson C. K., et al. (2025). Characterization of the smoltification process and post-sea transfer stunts of Atlantic salmon (Salmo salar) in full-scale intensive production. Aquaculture 609. doi: 10.1016/j.aquaculture.2025.742815
Folmar L. C. and Dickhoff W. W. (1980). The parr-smolt transformation (smoltification) and seawater adaptations in salmonids. Aquaculture 21, 1–37. doi: 10.1016/0044-8486(80)90123-4
Gilmore A. R., Gogel B. J., Cullis B. R., Welham S. J., and Thompson R. (2021). ASReml user guide release 4.2 functional specification, VSN international ltd, hemel hempstead, HP2 4TP, UK.
Gjerde B., Simianer H., and Refstie T. (1994). Estimates of genetic and phenotypic parameters for body weight, growth rate and sexual maturity in Atlantic salmon. Livest. Prod. Sci. 38, 133–143. doi: 10.1016/0301-6226(94)90057-4
Grønvold L., van Dalum M. J., Striberny A., Manousi D., Ytrestøyl T., Mørkøre T., et al. (2024). Transcriptomic profiling of gill biopsies to define predictive markers for seawater survival in farmed Atlantic salmon. bioRxiv, 2024.08.20.608748. doi: 10.1101/2024.08.20.608748
Handeland S. O., Imsland A. K., Bjornsson B. T., and Stefansson S. O. (2013). Long-term effects of photoperiod, temperature and their interaction on growth, gill Na+, K+- ATPase activity, seawater tolerance and plasma growth-hormone levels in Atlantic salmon Salmo salar. J. Fish Biol. 83, 1197–1209. doi: 10.1111/jfb.12215
Hayes B. J. (2011). Technical note: Efficient parentage assignment and pedigree reconstruction with dense single nucleotide polymorphism data. J. Dairy Sci. 94, 2114–2117. doi: 10.3168/jds.2010-3896
Hoar W. S. (1988). “The physiology of smolting salmonids,” in In fish physiology, vol. Vol XIB . Eds. Hoar W. S. and Randall D. (Academic Press, New York), 275–343.
Houston R. D., Taggart J. B., Cézard T., Cézard T., Bekaert M., Lowe N. R., et al. (2014). Development and validation of a high density SNP genotyping array for Atlantic salmon (Salmo salar). BMC Genomics 15, 90. doi: 10.1186/1471-2164-15-90
Jobling M. (2003). The thermal growth coefficient (TGC) model of fish growth: a cautionary note. Aquaculture Res. 34, 581–584. doi: 10.1046/j.1365-2109.2003.00859.x
Khaw H. L., Gjerde B., Boison S. A., Hjelle E., and Difford G. F. (2021). Quantitative Genetics of Smoltification Status at the Time of Seawater Transfer in Atlantic Salmon (Salmo salar). Front. Genet. 12. doi: 10.3389/fgene.2021.696893
Kolarevic J., Baeverfjord G., Ytteborg E., Megård Reiten B. K., Nergård S., and Terjesen B. F. (2014). Performance and welfare of Atlantic salmon smolt reared in recirculating or flow through aquaculture systems. Aquaculture 432, 15–25. doi: 10.1016/j.aquaculture.2014.03.033
Le Cren C. D. (1951). The Length-Weight Relationship and Seasonal Cycle in Gonad Weight and Condition in Perch, Perca fluviatilis. J. Animal Ecol. 20, 201–219. doi: 10.2307/1540
Lynch M. and Walsh B. (1997). Genetics and analysis of quantitative traits (Sunderland, MA, USA: Sinauer Associates).
McCormick S. D., Regish A. M., Christensen A. K., and Björnsson B. T (2013). Differential regulation of sodium–potassium pump isoforms during smolt development and seawater exposure of Atlantic salmon. J. Exp. Biol. 216(7), 1142–1151
McCormick S. D., Shrimpton J. M., Moriyama S., and Björnsson B. T. (2007). Differential hormonal responses of Atlantic salmon parr and smolt to increased daylength: a possible developmental basis for smolting. Aquaculture 273, 337–344. doi: 10.1016/j.aquaculture.2007.10.015
Meuwissen T., Hayes B., and Goddard M. (2016). Genomic selection: a paradigm shift in animal breeding. Anim. Front. 6, 6–14. doi: 10.2527/af.2016-0002
Montgomery D. W., Negrete B. Jr., Hong Gam L. T., Izutsu A., Culbert B. M., Bernier N. J., et al. (2025). Producing a better smolt: Can ‘winter’ treatments combining low temperature and short daylength photoperiods enhance the smolt quality of Atlantic salmon? Aquaculture 596. doi: 10.1016/j.aquaculture.2024.741699
Myklatun L. E., Fraser T. W. K., and Hansen T. J. (2023). Long term effects of smolt production strategy and early seawater phase rearing environment on mortality, growth, sexual maturation, and vertebra deformities in farmed Atlantic salmon (Salmo salar L.). Aquaculture 569. doi: 10.1016/j.aquaculture.2023.739346
Myklatun M., Madaro M., Philip A. J. P., Pedersen A.Ø., Remø S., Hansen T. J., et al. (2025). Long term effects of smolt and post-smolt production strategy on mortality, growth, sexual maturation and melanized focal changes in farmed Atlantic salmon (Salmo salar L.). Aquaculture 602. doi: 10.1016/j.aquaculture.2025.742371
Nichols K. M., Edo A. F., Wheeler P. A., and Thorgaard G. H. (2008). The genetic basis of smoltification – related tratis in Oncorhynchus mykiss. Genetics 179, 1559–1575. doi: 10.1534/genetics.107.084251
Piironen J., Kiiskinen P., Huuskonen H., Heikura-Ovaskainen M., and Vornanrn M. (2013). Comparison of smoltification in Atlantic salmon (Salmo salar) from anadromous and landlocked populations under common garden conditions. Annales Zoologici Fennici 50, 1–15. doi: 10.5735/086.050.0101
Salman N. A. and Eddy F. B. (1988). Effect of dietary sodium chloride on growth, food intake and conversion efficiency in rainbow trout (Salmo gairdneri Richardson). Aquaculture 70, 131–144. doi: 10.1016/0044-8486(88)90012-9
Sigholt T., Åsgård T., and Staurnes M. (1998). Timing of parr-smolt transformation in Atlantic salmon (Salmo salar): effects of changes in temperature and photoperiod. Aquaculture 160, 129–144. doi: 10.1016/S0044-8486(97)00220-2
Sigholt T., Staurnes M., Jakobsen H. J., and Åsgard T. (1995). Effects of continuous light and short-day photoperiod on smolting, seawater survival and growth in Atlantic salmon (Salmo salar). Aquaculture 130, 373–388. doi: 10.1016/0044-8486(94)00349-S
Sommerset I., Wiik-Nielsen J., Moldal T., Oliveira V. H. S., Svendsen J. C., Haukaas A., et al. (2023). “Norwegian fish health report 2023,” in Norwegian veterinary institute report, series 8b/2024, 270. (Ås, Norway: Norwegian Veterinary Institute).
Sommerset I., Wiik-Nielsen J., Oliveira V. H. S., Moldal T., Bornø G., Haukaas, et al. (2022). “Norwegian fish health report 2022,” in Norwegian veterinary institute report, series 2a/2022, 219. (Ås, Norway: Norwegian Veterinary Institute).
Staurnes M. and Finstad B. (2000). The effects of dietary NaCl supplement on hypoosmoregulatory ability and seawater performance of Arctic charr (Salvelinus alpinus L.) smolts. Aquac. Res. 31, 737–743. doi: 10.1046/j.1365-2109.2000.00495.x
Stradmeyer L. (1994). Survival, growth and feeding of Atlantic salmon (Salmo salar) smolts after transfer to seawater in relation to the failed smolt syndrome. Aquaculture Res. 25, 103 –1112. doi: 10.1111/j.1365-2109.1994.tb00670.x
Strand J. E. T., Hazlerigg D., and Jorgensen E. H. (2018). Photoperiod revisited: is there a critical day length for triggering a complete parr–smolt transformation in Atlantic salmon Salmo salar? J. Fish Biol. 93, 440–448. doi: 10.1111/jfb.13760
Striberny A., Lauritzen D. E., Marco J. F., Campinho A., Gaetano P., Duarte V., et al. (2021). More than one way to smoltify a salmon? Effects of dietary and light treatment on smolt development and seawater growth performance in Atlantic salmon. Aquaculture 532, 1–16. doi: 10.1016/j.aquaculture.2020.736044
Takvam M. (2020). Development of Na+, K+ ATPase enzyme activity and expression patterns of sulfate transporters in gills, intestine and kidney during smoltification and SW acclimation in Atlantic salmon (Salmo salar L.) (Master Degree in Aquaculture BiologyUniversity of Bergen, Department of biological sciences). Available online at: https://hdl.handle.net/1956/2321.
Takvam M., Sundell K., Sundh H., Gharbi N., Kryvi H., and Nilsen T. O. (2023). New wine in old bottles: Modification of the Na+/K+-ATPase enzyme activity assayand its application in salmonid aquaculture. Rev. Aquac 16, 1087–1098. doi: 10.1111/raq.128871098
Takvam T., Denker E., Gharbi N., Kryvi H., and Nilse. T. O. (2021). Sulfate homeostasis in Atlantic salmon is associated with differential regulation of salmonid-specific paralogs in gill and kidney. Physiol. Rep. 9, e15059. doi: 10.14814/phy2.1505
Ura K., Mizuno S., Okubo T., Chida Y., Misaka N., Adachi S., et al. (1997). Immunohistochemical study on changes in gill Na+/K+-ATPase α-subunit during smoltification in the wild masu salmon, Oncorhynchus masou. Fish Physiol. Biochem. 17, 397–403. doi: 10.1023/A:1007755008204
Van Rijn C. A., Jones P. L., Evans B. S., and Afonso L. O. B. (2021). Physiological and growth responses of juvenile Atlantic salmon (Salmo salar) transferred to seawater during different stages of smolt development. Aquaculture . 538. doi: 10.1016/j.aquaculture.2021.736527
Wald A. (1943). Tests of statistical hypotheses concerning several parameters when the 873 number of parameters is large. Trans. Am. Math. Soc 54, 426–482. doi: 10.1090/S0002-9947-1943-0012401-3
Ytrestøyl T., Hjelle E., Kolarevic J., Takle H., Rebl A., Afanasyev S., et al. (2022). Photoperiod in recirculation aquaculture systems and timing of seawater transfer affect seawater growth performance of Atlantic salmon (Salmo salar). J. World Aquaculture Soc. 54(1), 73–95. doi: 10.1111/jwas.12880
Appendix 1. The experimental design with the dates of the different activities and trait recordings.

Appendix 2. Distribution of the ratio between the relative mRNA expression values of two alpha catalytic subunits of two variants (α1b/α1a) of the Na+,K+-ATPase (NKA) expressed in the gill tissue samples of all the fish in each of the three light regimes about two weeks prior to seawater transfer.
Keywords: A. salmon, light regimes, smoltification, growth, survival, heritability, genetic correlations
Citation: Gjerde B, Boison SA, Hazlerigg D, Ytrestøyl T, Mørkøre T, Jørgensen E, Striberny A and Sandve SR (2025) Impact of three light smoltification regimes on the performance and genetic parameters of traits in Atlantic salmon. Front. Anim. Sci. 6:1615804. doi: 10.3389/fanim.2025.1615804
Received: 21 April 2025; Accepted: 29 August 2025;
Published: 06 October 2025.
Edited by:
Yuliaxis Ramayo Caldas, Institute of Agrifood Research and Technology (IRTA), SpainReviewed by:
Sebastian Escobar-Aguirre, Pontificia Universidad Católica de Chile, ChileThiruvenkadan Aranganoor Kannan, Tamil Nadu Veterinary and Animal Sciences University, India
Copyright © 2025 Gjerde, Boison, Hazlerigg, Ytrestøyl, Mørkøre, Jørgensen, Striberny and Sandve. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Bjarne Gjerde, Ymphcm5lLmdqZXJkZUBub2ZpbWEubm8=
†ORCID: Bjarne Gjerde, orcid.org/0000-0001-5011-1516