 Makenzie Harrison1
Makenzie Harrison1 John Odunayo Adebayo1Anijia Mills-Widemon1
John Odunayo Adebayo1Anijia Mills-Widemon1 Sola Ikuejamoye-Omotore1
Sola Ikuejamoye-Omotore1 Zaira M. Estrada-Reyes1,2*
Zaira M. Estrada-Reyes1,2* Ibukun M. Ogunade3
Ibukun M. Ogunade3 Andres A. Pech-Cervantes2
Andres A. Pech-Cervantes2 Thomas H. Terrill4
Thomas H. Terrill4 Aftab Siddique4Alexis Ruiz-González5
Aftab Siddique4Alexis Ruiz-González5- 1Department of Animal Science, North Carolina Agricultural and Technical State University, Greensboro, NC, United States
- 2College of Agriculture, Food and Natural Resources, Cooperative Agricultural Research Center, Prairie View A&M University, Prairie View, TX, United States
- 3Division of Animal and Nutritional Science, West Virginia University, Morgantown, WV, United States
- 4College of Agricultural, Family Sciences, and Technology, Fort Valley State University, Fort Valley, GA, United States
- 5Department of Animal Science, Laval University, Quebec, QC, Canada
The study aimed to investigate the effects of short-term heat stress on physiological, hematological, immunological, and metabolomic responses in Florida Cracker ewes. Fourteen Florida Cracker ewes were randomly assigned to either heat stress (HT) or cooling (control, CTL) conditions, respectively, for a duration of six weeks during the summer season. Ambient temperature and relative humidity data for HT and CTL ewes were recorded weekly to estimate temperature humidity index (THI). Respiration rate and rectal temperature were measured for each experimental ewe. Blood samples were collected from each ewe at baseline (week 0) and at week 1, 2, 3, 4, 5 and 6 post-exposure for hematology analysis and plasma extraction. Plasma samples were used for analysis of IgA level and to conduct targeted metabolomics. Targeted metabolomics analyzed a total of 50 metabolites, including organic acids, amino acids, hexoses, lipids, and carnitines. The THI for the HT and CTL groups ranged from 81.3 – 89.5 and from 73.5 – 75.6, respectively. The HT group exhibited a significant increase (P < 0.05) in respiratory rate compared to the CTL group. No significant changes were observed in rectal temperature or IgA levels between the experimental groups. Hematology analysis revealed a significant increase (P < 0.05) in neutrophil count and neutrophil percentage (%) and a decrease in lymphocyte percentage (%) at week 3 in the HT group. Targeted metabolomics analysis identified 19 differentially abundant metabolites between the HT and CTL groups. Pathway enrichment analysis showed upregulation of fatty acid biosynthesis, glutathione metabolism, primary bile biosynthesis, porphyrin metabolism, and glycine, serine, and threonine metabolism (P ≤ 0.05) in the HT group compared to CTL. Our findings highlight that short-term heat stress alters the immunological, hematological, and metabolomic profile of Florida Cracker ewes. Understanding these changes can contribute to developing effective management strategies to mitigate the detrimental impact of heat stress on sheep operations in the southern U.S.
1 Introduction
Heat stress is one of the major limiting factors for sustainable sheep production (Marai et al., 2007; Slimen et al., 2019). In sheep, exposure to heat stress causes reduction in feed intake and digestive capacity which compromises nutrient absorption and metabolism (Belhadj Slimen et al., 2016). Additionally, increased respiration rate and induction of oxidative stress responses has been observed in this species (Marai et al., 2007; Belhadj Slimen et al., 2016; Slimen et al., 2019). These alterations have a negative impact on body weight, body condition score, dry matter intake, and immune response (Marai et al., 2007) and vary depending on breed (Slimen et al., 2019). and U.S. region (Tadesse et al., 2019). For example, previous studies on St. Croix, Katahdin, and Dorper sheep have identified breed-specific differences in body weight under heat stress, with St. Croix sheep showing the lowest body weight, followed by Dorper and then Katahdin breeds (Tadesse et al., 2019). Similar results were also reported for body condition score among these sheep breeds. Additionally, these authors noted that sheep from the Southeast U.S exhibited lower body weights compared to those from the Midwest and Northwest regions (Tadesse et al., 2019). In beef and dairy cattle, previous research has shown that heat stress causes alterations of neutrophil and lymphocyte populations, making animals more susceptible to infections (Strong et al., 2015; Bagath et al., 2019; Dahl et al., 2020; Abduch et al., 2022).
Environmental parameters such as barn temperature and humidity are commonly used for predicting heat stress in livestock, however, they do not directly measure the metabolic changes in the animals due to heat stress. Additional physiological parameters, such as rectal temperature, respiratory rate and heart rate are integrated with environmental parameters to diagnose heat stress in livestock (Slimen et al., 2019). However, these indicators can vary significantly based on the animal´s physiological stage and health status. Indirect biomarkers of heat stress may provide diagnostic value and enhance our understanding of the associated metabolic and physiological changes. Plasma metabolites and immunological status serve as valuable biomarkers for diagnosing and identifying heat stress adaptation in farm animals because they could reflect the physiological and metabolic adjustment animals make in response to heat stress (Jorge-Smeding et al., 2024).
Utilization of thermotolerant livestock is one of the strategies to mitigate heat stress in sustainable operations. Adaptation to heat stress has been observed in some livestock species, such as cattle and sheep (Beatty et al., 2006; Fadare et al., 2012; Dikmen et al., 2014). Florida Cracker sheep is a heritage breed well-adapted to the subtropical climate of Florida and offers a unique opportunity to study the biological responses to heat stress. This breed is used for meat purposes and evolved under natural selection for more than five centuries in Florida, U.S. Despite their adaptation, the physiological, hematological, immunological, and metabolomic responses of Florida Cracker sheep to heat stress remain unknown. This study aims to fill these knowledge gaps by systematically investigating the effects of heat stress on the physiological, hematological, immunological, and metabolomic responses in Florida Cracker sheep. Therefore, the objective of this study was to determine the effect of short-term heat stress on physiological, hematological, immunological and metabolomic responses in Florida Cracker sheep.
2 Materials and methods
2.1 Animal population and collection of physiological measures
The research protocol was approved by the Institutional Animal Care and Use Committee (Approval number LA23-0019) of North Carolina Agricultural and Technical State University. A total of fourteen ewes (2 years old) were allocated to heat stress (HT, n = 7) or cooling (CTL, n = 7) conditions for six weeks during the summer season (July – August) of 2023. Experimental ewes were free of gastrointestinal parasites or other infectious diseases prior and during the experimental period. Treatment groups (HT and CTL) were grouped in concrete floor pens with similar dimensions and characteristics and had access to shelter, food, and freshwater ad libitum. The HT pen was exposed to regular ambient temperatures and the cooling pen was equipped with an evaporative cooling fan. Two data loggers were placed on both sides of the HT and CTL pens to record daily ambient temperature and relative humidity of HT and CTL pens at 1500 h and to estimate temperature humidity index (THI) using the following formula (National Research Council, 1971):
THI = (1.8 x T + 32) – ((0.55 – 0.0055 x RH) x (1.8 x T – 26.8)),
where T = dry bulb temperature (°C) and RH= relative humidity (percent).
The respiratory rate and rectal temperature were recorded daily from 1400 to 1500 h on each experimental ewe. The respiratory rate was measured by counting the number of flank movements in one minute with the help of a chronograph, and the rectal temperature was measured with a digital veterinary thermometer.
2.2 Blood and IgA analysis
Two blood samples were collected by jugular venipuncture using EDTA vacutainer tubes and 21-gauge needles from each ewe at baseline (week 0) and at week 1, 2, 3, 4, 5 and 6 post-exposure. Blood samples were placed in an ice bucket and transported to the laboratory for further analysis. The first blood sample was used to perform a full hematology analysis using the Vetscan HM5 (Zoetis, Parsippany, New Jersey). The second blood sample was centrifuged at 3,600 rpm for 15 minutes for plasma separation. Then, two aliquots of plasma were stored at -80°C for analysis of IgA levels and targeted metabolomics. For IgA analysis, plasma samples were thawed over night at 4°C prior to assessment, and a commercial kit (Biotang Inc, Sheep IgA ELISA kit) was used following the manufacturer’s instructions. A total of 50 μL of plasma was added to each well of the ELISA plate (96 wells). Then, HRP-conjugate (50 μL) was added to each well and the plate incubated for 30 minutes at 37°C. The wells were aspirated washed five times with Wash Buffer using a squirt bottle. A total of 90 μL of TMB Substrate was added to each well and incubated for 20 minutes at 37°C. For this step, plates were protected from light to avoid degradation of the substrate. To stop the reaction, 50 μL of Stop Solution was added to each well. Optical density of each well was determined within five minutes using a microplate reader set to 450 nm.
2.3 Targeted metabolomics
The second aliquot of plasma was used for targeted metabolomics. Targeted metabolomics was performed by Nuclear Magnetic Resonance (NMR) spectroscopy to quantify a total of 50 metabolites, including organic acids, amino acids, hexoses, lipids, and carnitines (Supplementary Table 1). The procedures for plasma sample preparation and the NMR spectral analysis followed the methods used by Ogunade et al. (2018). Plasma samples collected at weeks 1, 2, 3, 4, 5 and 6 were pooled within each group and deproteinized using ultrafiltration and the modified protocol of Psychogios et al. (2011). Deproteinization was used to remove proteins and lipoproteins from plasma samples. Then, a total of 160 μL of plasma sample was mixed with 40 μL of a standard buffer solution (54% D2O and 46% 250 mM KH2PO4 at pH 7.0). The mixture was transferred into a 3 mm SampleJet NMR tube for further spectral analysis. The 1H-NMR spectra were obtained utilizing a 700 MHz Avance III spectrometer. The processing of the 1H-NMR spectra was performed using Bayesil (automated analysis software).
2.4 Statistical analysis
For the statistical analysis of the physiological, hematological, and immunological parameters, a mixed model with repeated measures was used. Fixed effects included treatment (HT or CTL), week (1 - 6), and the treatment by week interaction. Animal was fitted as a random effect and the baseline data (week 0) was used as a covariate. The variance-covariance structure with the best fit was unstructured (showed the lowest Bayesian information criterion (BIC), Akaike information criterion (AIC), and the Akaike information criterion corrected (AICc). Data from targeted metabolomics was analyzed using Metaboanalyst 5.0 software (Xia and Wishart, 2016). Prior to the statistical analysis, the data were log-transformed and auto-pareto scaled. A PLS-DA scores plot was used to evaluate the level of separation between HT and CTL groups of animals using the first two principal components. Then, differentially abundance analysis was performed and a VIP scores plot was generated. To determine the metabolic pathways that were differentially (FDR ≤ 0.05) altered in the HT group compared to CTL, pathway enrichment analysis using KEGG (Kyoto Encyclopedia of Genes and Genomes) database was performed.
3 Results
3.1 Temperature humidity index and physiological, hematological and immunological measures
The THI between the HT and CTL groups was compared. These results are presented in Figure 1. The THI for the HT and CTL groups ranged from 81.3 – 87.9 and from 73.5 – 75.6, respectively. For the HT group, the highest THI (87.9) was observed in week 3 followed by week 2 (85.8) and week 5 (84.8).
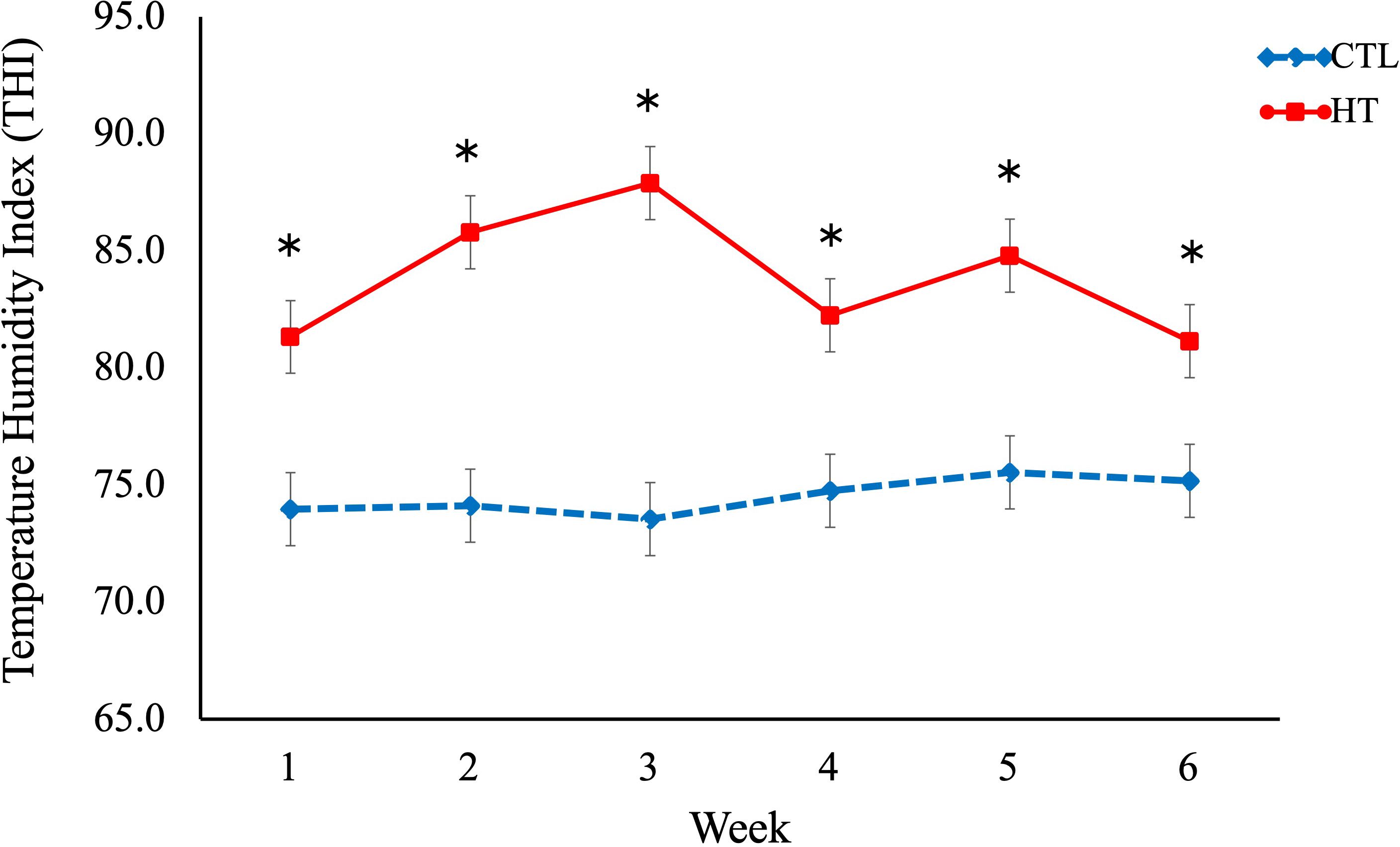
Figure 1. Temperature humidity index (THI) in heat stress (HT) and control (CTL) Florida Cracker ewes. * = p-value < 0.05.
No significant differences in rectal temperature or IgA levels were observed among the experimental groups (Table 1). However, a significant difference in respiratory rate was found between the HT and CTL groups. The respiratory rate was greater (P < 0.05) in the HT group (Table 1) on week 2, 3, 4, 5 and 6 when compared to the CTL group (Figure 2). For hematological data (Table 1), a significant (P < 0.05) increase of neutrophil % and a decrease in lymphocyte % was observed for the HT group relative to CTL at week 3 (Figures 3, 4).
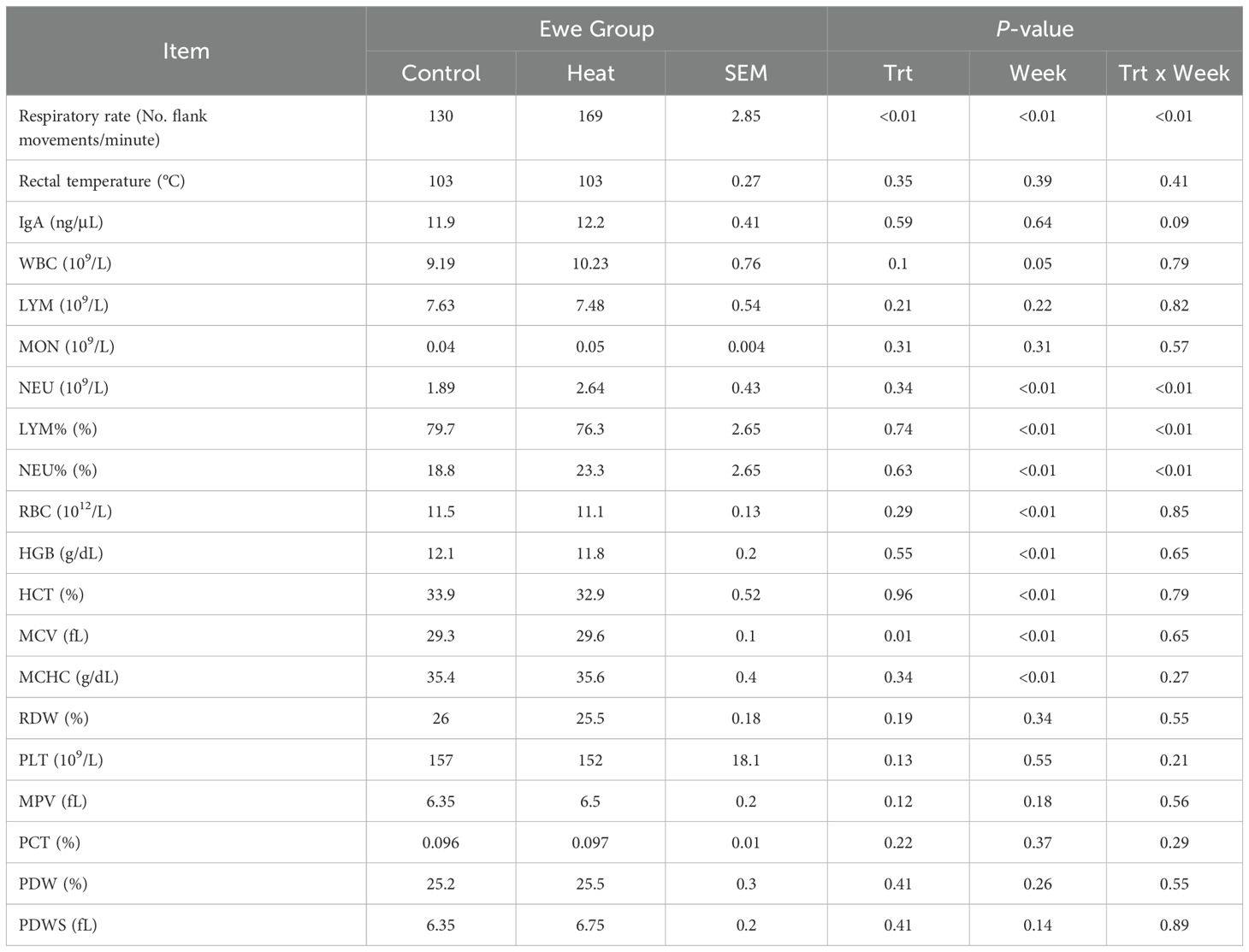
Table 1. Effects of short-term heat stress on respiratory rate (No. flank movements/minute), rectal temperature (°C), white blood cell count (WBC, 109/L), lymphocyte count (LYM, 109/L), monocyte count (MON, 109/L), neutrophil count (NEU, 109/L), lymphocyte percentage (LYM%, %), neutrophil percentage (NEU%, %), red blood cell count (RBC, 1012/L), hemoglobin level (HGB, g/dL), hematocrit (HCT, %), mean corpuscular volume (MCV, fL), mean corpuscular hemoglobin concentration (MCHC, g/dL), erythrocyte distribution width (RDW, %), platelet count (PLT, 109/L), mean platelet volume (MPV, fL), procalcitonin (PCT, %), and platelet distribution width (PDW, %) in Florida Cracker ewes.
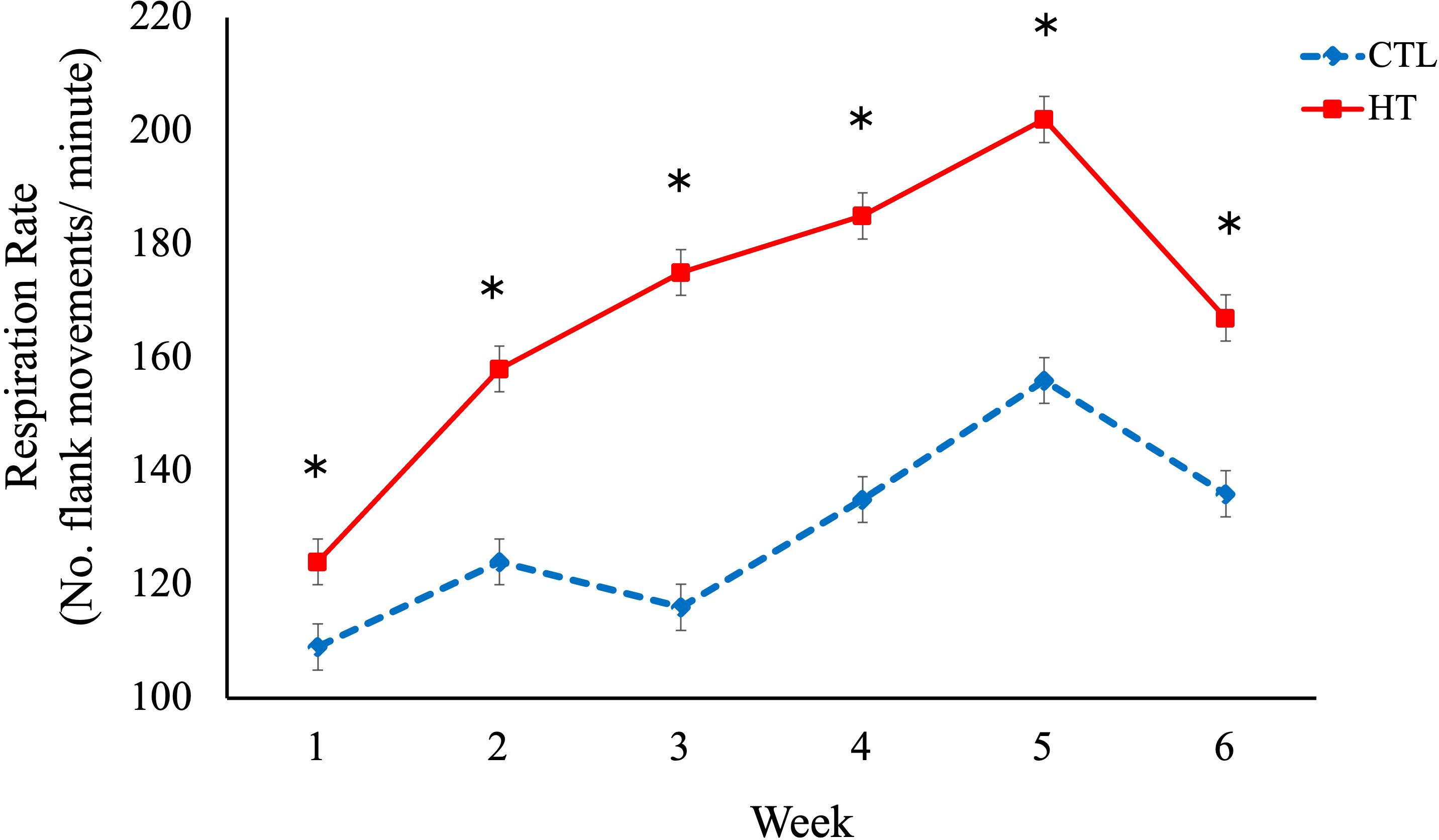
Figure 2. Respiration rate in heat stress (HT) and control (CTL) Florida Cracker ewes. * = p-value < 0.05.
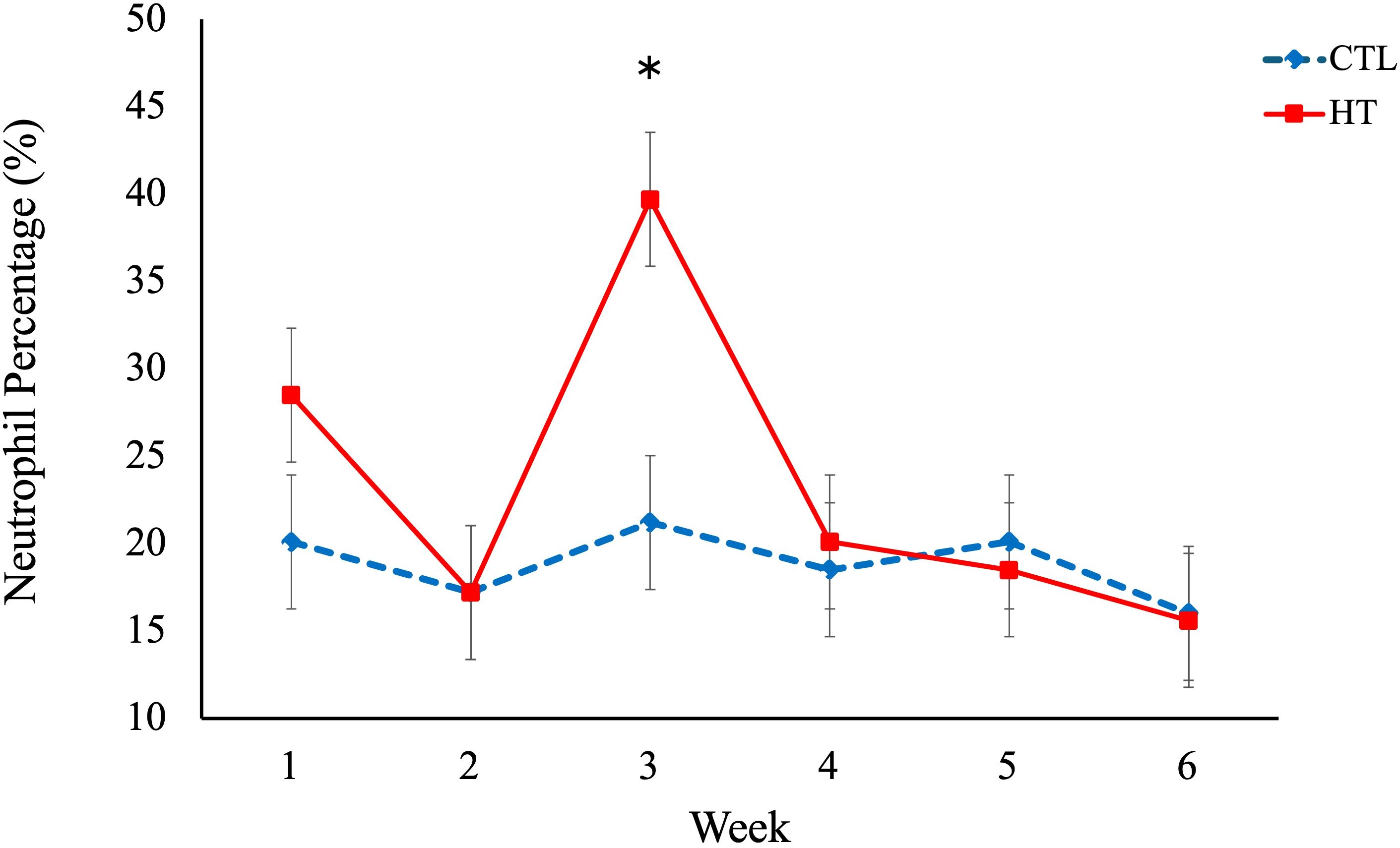
Figure 3. Neutrophil percentage (%) in heat stress (HT) and control (CTL) Florida Cracker ewes. * = p-value < 0.05.

Figure 4. Lymphocyte percentage (%) in heat stress (HT) and control (CTL) Florida Cracker ewes. * = p-value < 0.05.
3.2 Targeted metabolomics
Targeted metabolomics was performed to detect and quantify total of 50 metabolites in plasma samples of experimental groups (Supplementary file 1). The PLS-DA plot showed a slight separation between HT and CTL groups. The first and second principal components explained 48.7% and 12.1% of the total variance observed between groups, respectively (Figure 5). The results of the VIP scores plot analysis revealed a total of 19 differentially abundant (VIP > 1) metabolites between CTL and HT animals (Figure 6). Compared to the control, plasma concentrations of 18 metabolites, such as malonate, betaine, glycine, ornithine, citric acid, glucose, and sarcosine were greater in HT ewes, whereas the plasma concentration of lactic acid was lower in HT sheep (Figure 6). To determine the metabolic pathways that were altered in the HT group compared to CTL, a pathway enrichment analysis using KEGG was performed. Results of the pathway analysis of all metabolites revealed that fatty acid biosynthesis, glutathione metabolism, primary bile biosynthesis, porphyrin metabolism, and glycine, serine, and threonine metabolism were upregulated (P ≤ 0.05) in HT compared to CTL sheep (Figure 7). The Figure 8 shows the overall changes observed in peripheral blood and plasma, and in respiratory rate in Florida Cracker ewes exposed to heat stress.
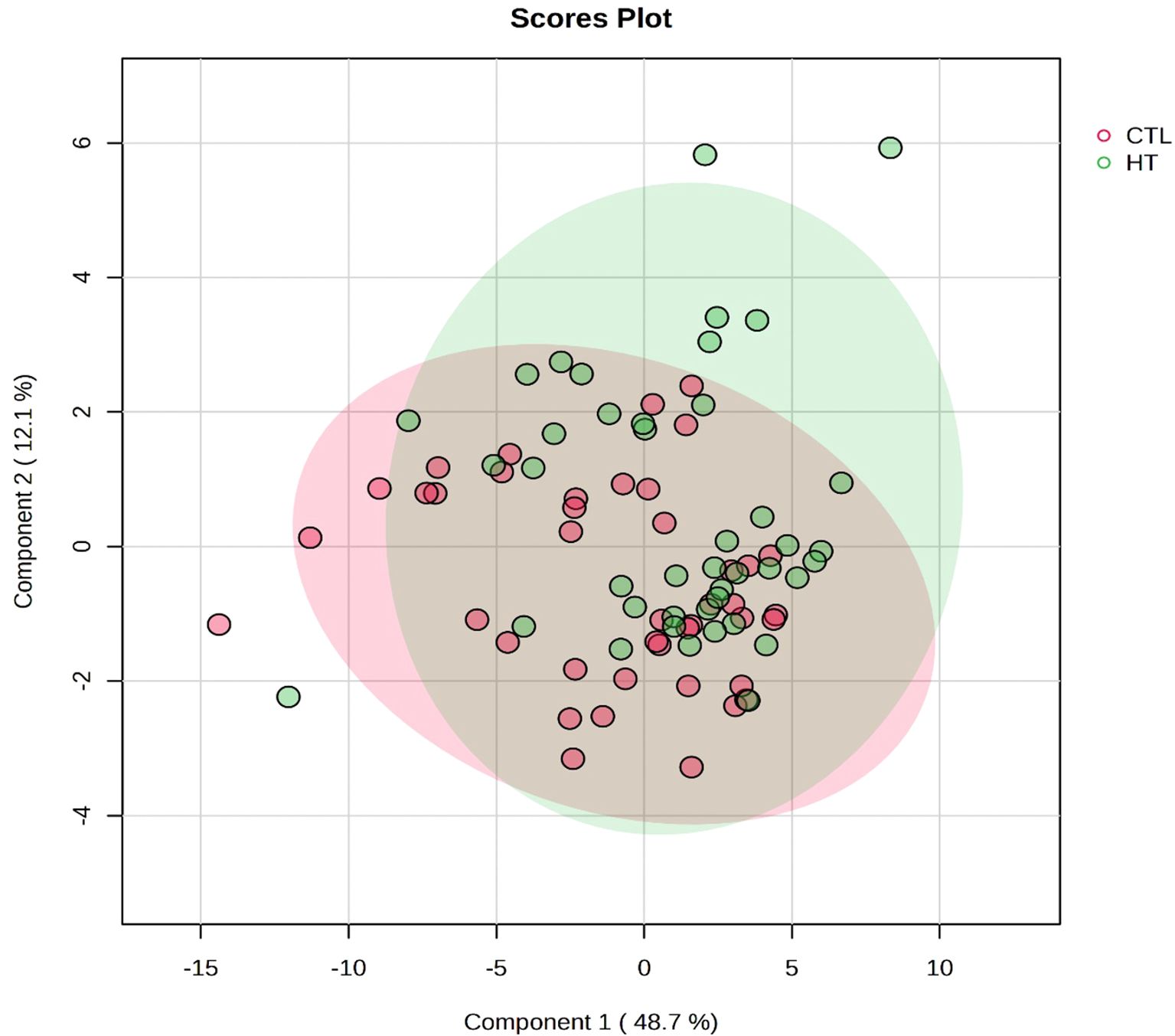
Figure 5. PLS-DA scores plot of plasma metabolome of heat stress (HT) and control (CTL) Florida Cracker ewes. The PLS-DA scores plot shows a slight separation between HT and CTL ewes.
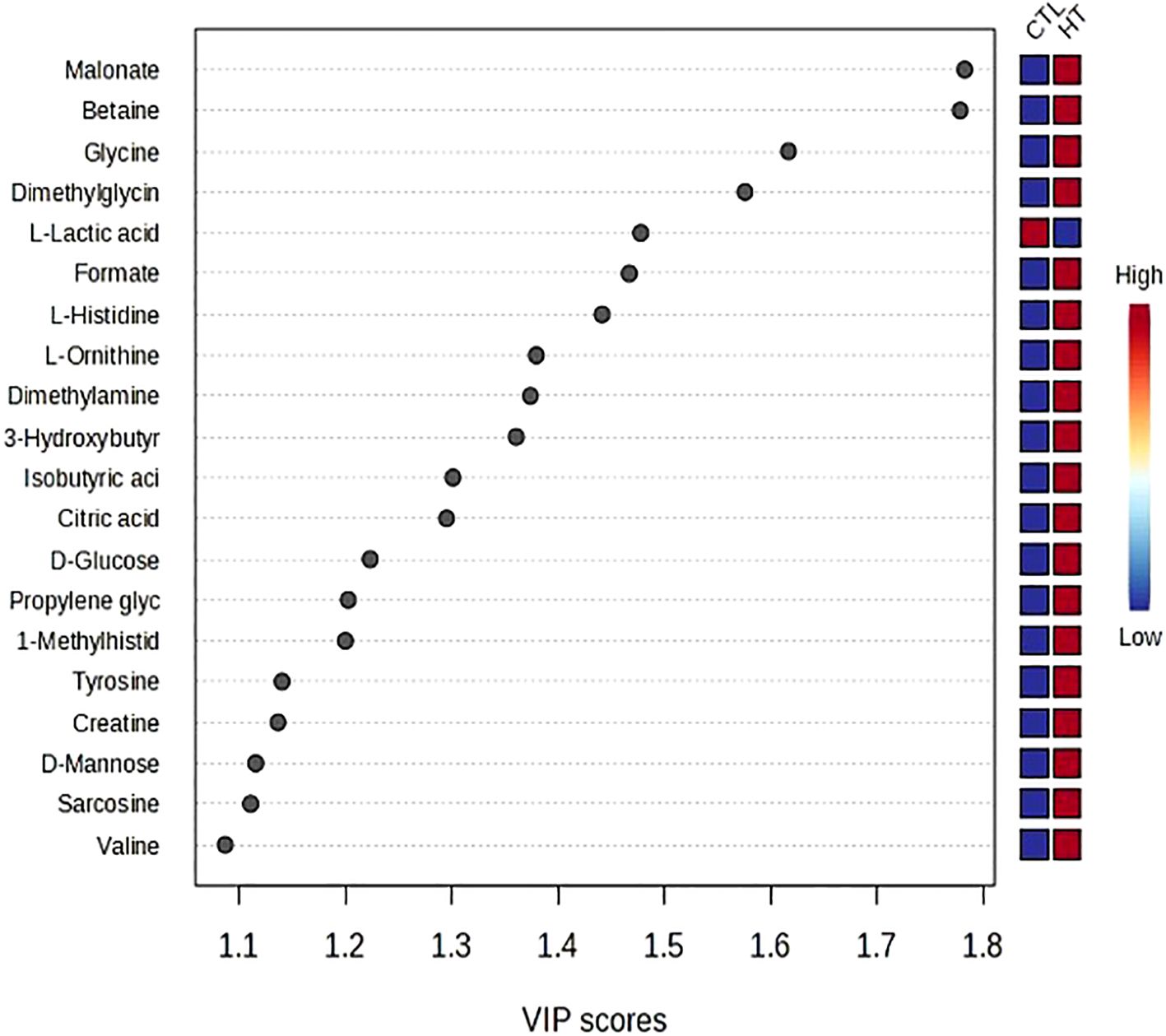
Figure 6. VIP scores plot of the differentially abundant metabolites between heat stress (HT) and control (CTL) Florida Cracker ewes.
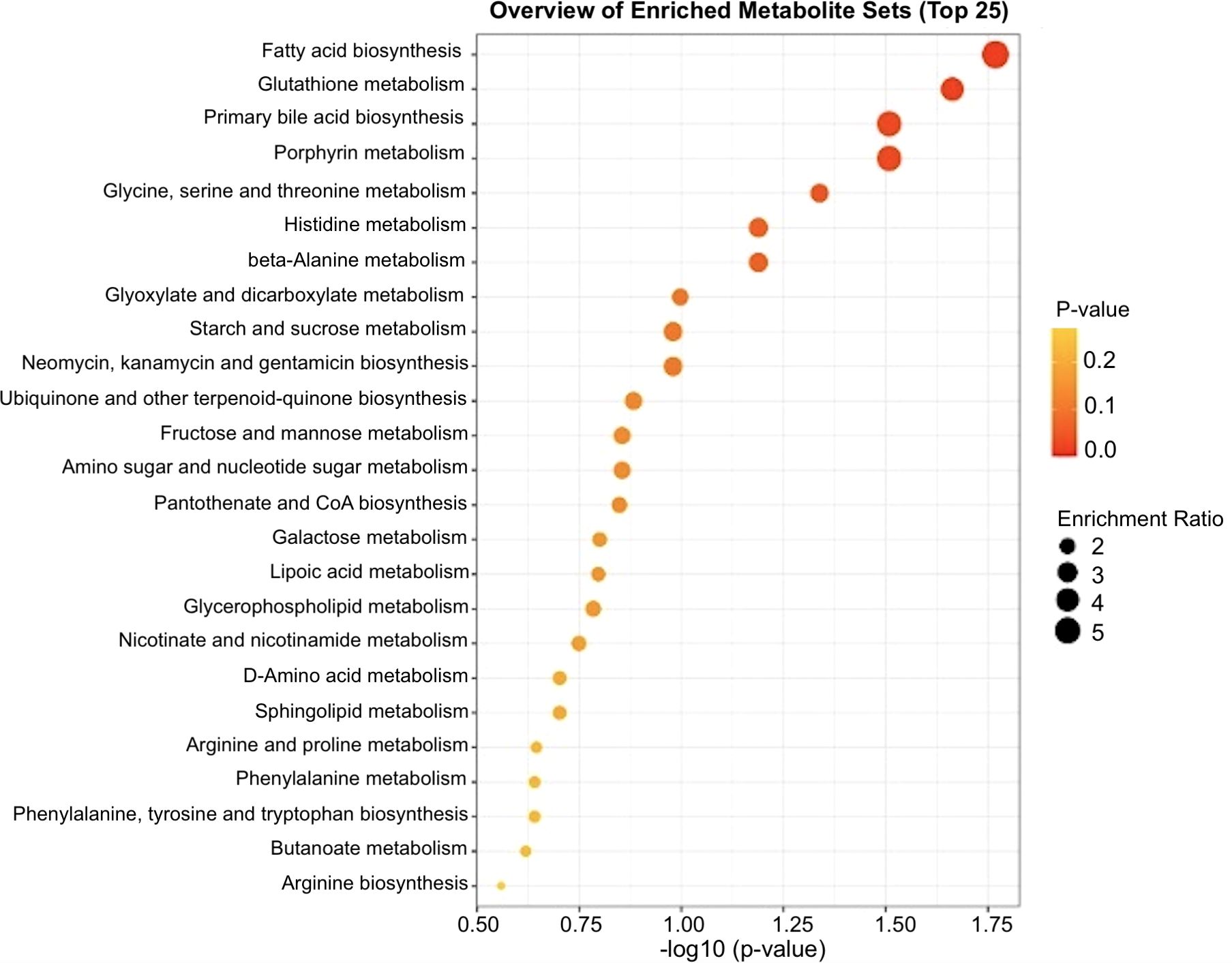
Figure 7. Pathway enrichment analysis of the differentially abundant metabolites between heat stress (HT) and control (CTL) Florida Cracker ewes. Pathways with -log10(P-value) > 1.3 are considered significant.
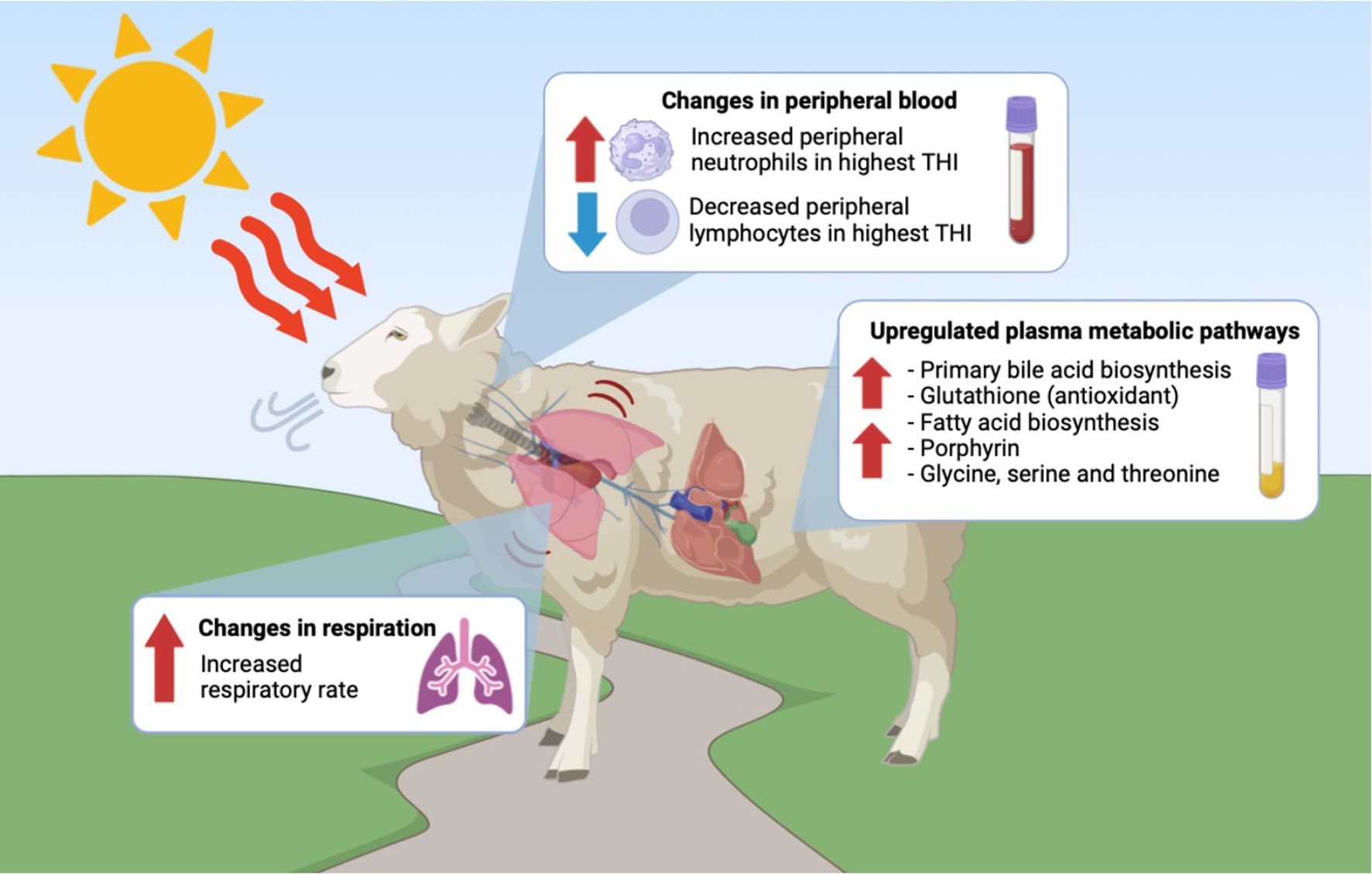
Figure 8. Alterations in respiration rate, peripheral blood cells and plasma metabolomic pathways observed in Florida Cracker ewes exposed to heat stress. This image was generated with BioRender.
4 Discussion
In this study, the exposure of ewes to short-term heat stress caused changes in respiration rate, hematological parameters, and plasma metabolites. The heat stressed group exhibited higher respiratory rate when compared to the control group. Previous studies in sheep have observed a similar increase in this physiological variable in response to heat stress (Marai et al., 2007; Slimen et al., 2019). Respiration rate has also been proposed as a trait to evaluate thermotolerance in sheep adapted to humid and hot environments (Castanheira et al., 2010). Rectal temperature is another physiological measure that has been used small ruminants for assessing heat stress, and studies have reported variations in rectal temperature in response to THI fluctuations (Hooper et al., 2018; Slimen et al., 2019; Tadesse et al., 2019). However, it is important to highlight that our results showed no significant changes on rectal temperature between heat stressed and control ewes. Similar findings in rectal temperature have been reported for Omani sheep, a sheep breed naturally adapted to hot environmental conditions. This breed can tolerate THI values up to 82, without any physiological sign of heat stress (Srikandakumar et al., 2003). Similarly, Florida Cracker sheep is a heritage sheep breed that was developed under natural selection for almost 500 years in Florida, U.S. (https://livestockconservancy.org/heritage-breeds/conservation-priority-list/). The state of Florida is characterized for its high humidity and ambient temperatures, especially during the summer months, reaching THI estimates greater than 70 (Dikmen and Hansen, 2009; Ouellet et al., 2021). As such, it is possible that Florida Cracker sheep incorporated mechanisms of adaptation to these environmental conditions to be able to survive, regulating their body temperature. Although the THI values for the HT group of ewes indicated moderate heat stress, the absence of significant differences in rectal temperature and some plasma metabolites between the HT and CLT groups suggests that Florida Cracker sheep may possess adaptive metabolic mechanisms to tolerate heat stress. Further research is needed to elucidate these adaptive processes.
One of the changes observed in week 3 of the current study was an increase in peripheral blood neutrophil percentage in the HT group compared to the CLT group. This increase may be attributed to several factors associated with heat stress, such as the activation of the hypothalamic-pituitary-adrenal (HPA) axis. Heat stress triggers the HPA axis, leading to elevated cortisol levels, a stress hormone that mobilizes neutrophils from the bone marrow into the bloodstream (Caroprese et al., 2022). Increased neutrophil concentration has also been associated with inflammatory responses. Heat stress can cause cellular damage and systemic inflammation, as previously investigated in dairy cows (Ruiz-González et al., 2023). Additionally, the activation of the immune system during heat stress can alter immune function, leading to changes in the distribution and concentration of various immune cells, including neutrophils (Tüfekci and Sejian, 2023). Also, the increase in neutrophils may be linked to the elevated metabolic rate of sheep exposed to heat stress. This heightened metabolic activity can lead to a greater demand for oxygen and nutrients, thereby stimulating the production and release of neutrophils (Dunshea et al., 2017). Nevertheless, the observed decrease in peripheral blood lymphocyte % in the HT group during week 3 may be associated with increased apoptosis (programmed cell death), a phenomenon that has been evaluated in mice under stress (Domínguez-Gerpe and Rey-Méndez, 2001). Similar observation has been reported in heat stressed cattle (Gu et al., 2021; Dahl et al., 2020; Bagath et al., 2019). The observation that these changes were only present in week 3 and not noted in weeks 4 to 6 suggests that Florida Cracker sheep may have adapted to heat episodes by developing metabolic mechanisms to cope with these changes. However, further research is necessary to understand the underlying processes involved.
Targeted plasma metabolomics analysis revealed an upregulation in fatty acid biosynthesis, glutathione metabolism, primary bile acid biosynthesis, glycine, serine, and threonine metabolism, and porphyrin metabolism. Notably, the upregulation of plasma fatty acid biosynthesis observed in the HT group may be attributed to increased energy demands during heat stress. This heightened energy requirement can lead to enhanced muscle and lipid catabolism, where muscle proteins and stored fats are broken down to meet energy needs (Rhoads et al., 2009; Ruiz-González et al., 2023). Additionally, the observed changes in glutathione metabolism in the current study may be also attributed to increased metabolic demands during heat stress (Dunshea et al., 2017). In sheep, heat stress induces oxidative stress and favors the production of reactive oxygen species, disrupting normal cellular function (Slimen et al., 2019). Glutathione is a crucial antioxidant that helps mitigate the production of reactive oxygen species (ROS) and functions as a cofactor for enzymes such as peroxidases and transferases (Foster et al., 2009). Thus, it is possible that the upregulation of glutathione metabolism serves as a strategy to alleviate the negative effects of ROS and oxidative stress induced by summer heat stress in Florida Cracker sheep.
In our study, primary bile biosynthesis was upregulated in heat stressed ewes. Bile acids are synthetized by oxidation of cholesterol in the hepatocytes. These are stored in the gallbladder and secreted into the duodenum to promote digestion and absorption of dietary lipids. About 95% of bile acids are reabsorbed from the ileum back to the liver through enterohepatic circulation (Chiang, 2013). Enterohepatic circulation of bile acids plays an important role in absorption of nutrients, xenobiotic disposal, and metabolic homeostasis (Houten et al., 2006). In normal conditions, the concentration of bile acids is usually low in the peripheral blood (Chiang and Ferrell, 2018). Our results showed an increase of primary bile biosynthesis in serum, which could be linked to liver changes in heat stressed ewes and possible mechanisms of adaptation to high ambient temperatures. Similar results have been observed in thermotolerant buffaloes that were kept under tropical environmental conditions, as Gu et al. (2018) reported an increase in primary bile biosynthesis and amino acid synthesis in heat-stressed buffaloes. They demonstrated that buffaloes adapt to heat stress by utilizing the metabolism of branched-chain amino acids, ketogenic amino acids, and gluconeogenesis as part of their adaptive strategy.
The possible alteration in liver functions due to heat stress could also be the consequence of the affection to porphyrin metabolism, leading to disruptions in the synthesis and breakdown of porphyrins, which has been reported in pigs exposed to heat stress (Ross et al., 2017). The observed changes in plasma amino acids (glycine, serine, and threonine) during heat stress may primarily be attributed to metabolic alterations, such as hormonal changes, like hyperinsulinemia (Ruiz-González et al., 2023). These changes may prioritize muscle breakdown to meet the increased energy demands under these conditions. Additionally, altered nutrient absorption, potentially due to a leaky gut, may result in inefficient amino acid absorption, compounded by reduced feed intake (Loor et al., 2023).
5 Conclusion
In conclusion, our study demonstrates that short-term heat stress significantly impacts the physiological, hematological, immunological, and metabolomic profiles of Florida Cracker ewes. The observed increase in respiratory rate and changes in blood parameters, such as elevated neutrophil counts and decreased lymphocyte percentages, indicate a stress response to elevated ambient temperatures. Metabolomic analysis further revealed alterations in key metabolic pathways, including fatty acid biosynthesis, glutathione metabolism, and primary bile biosynthesis, which may play crucial roles in the ewes’ adaptation to heat stress. These findings underscore the importance of developing effective management strategies to mitigate the adverse effects of heat stress on sheep, particularly in regions with hot climates like the southern US. By enhancing our understanding of the biological responses to heat stress, we can better support the health and productivity of sheep under challenging environmental conditions.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Ethics statement
The animal study was approved by Institutional Animal Care and Use Committee of North Carolina Agricultural and Technical State University (Approval number LA23-0019). The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
MH: Formal analysis, Investigation, Methodology, Writing – original draft. JA: Formal analysis, Investigation, Methodology, Writing – original draft. AM: Formal analysis, Investigation, Methodology, Writing – original draft. SI-O: Formal analysis, Investigation, Methodology, Writing – original draft. ZE: Formal analysis, Investigation, Methodology, Writing – original draft, Conceptualization, Data curation, Funding acquisition, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – review & editing. IO: Data curation, Formal analysis, Methodology, Supervision, Writing – review & editing, Writing – original draft. AP: Data curation, Formal analysis, Methodology, Supervision, Writing – review & editing, Writing – original draft. TT: Funding acquisition, Investigation, Methodology, Project administration, Supervision, Validation, Writing – original draft, Writing – review & editing. AS: Funding acquisition, Investigation, Project administration, Supervision, Writing – original draft, Writing – review & editing. AR: Investigation, Methodology, Supervision, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This work was funded by 1890 Universities Foundation and USDA, Grant # 2022-38427-37307.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fanim.2025.1631531/full#supplementary-material
References
Abduch N. G., Pires B. V., Souza L. L., Vicentini R. R., Zadra L. E. F., Fragomeni B. O., et al. (2022). Effect of thermal stress on thermoregulation, hematological and hormonal characteristics of Caracu beef cattle. Animals 12, 3473. doi: 10.3390/ani12243473
Bagath M., Krishnan G., Devaraj C., Rashamol V. P., Pragna P., Lees A. M., et al. (2019). The impact of heat stress on the immune system in dairy cattle: A review. Res. veterinary Sci. 126, 94–102. doi: 10.1016/j.rvsc.2019.08.011
Beatty D. T., Barnes A., Taylor E., Pethick D., McCarthy M., and Maloney S. K. (2006). Physiological responses of Bos taurus and Bos indicus cattle to prolonged, continuous heat and humidity. J. Anim. Sci. 84, 972–985. doi: 10.2527/2006.844972x
Belhadj Slimen I., Najar T., Ghram A., and Abdrrabba M. J. O. A. P. (2016). Heat stress effects on livestock: molecular, cellular and metabolic aspects, a review. J. Anim. Physiol. Anim. Nutr. 100, 401–412. doi: 10.1111/jpn.12379
Caroprese M., Ciliberti M. G., Albenzio M., and Sevi A. (2022). “Heat stress associated changes in the immune system related responses in sheep,” in Climate change and livestock production: recent advances and future perspectives (Springer Singapore, Singapore), 49–58).
Castanheira M., Paiva S. R., Louvandini H., Landim A., Fiorvanti M. C. S., Dallago B. S., et al. (2010). Use of heat tolerance traits in discriminating between groups of sheep in central Brazil. Trop. Anim. Health Production 42, 1821–1828. doi: 10.1007/s11250-010-9643-x
Chiang J. Y. (2013). Bile acid metabolism and signaling. Compr. Physiol. 3, 1191. doi: 10.1002/j.2040-4603.2013.tb00517.x
Chiang J. Y. and Ferrell J. M. (2018). Bile acid metabolism in liver pathobiology. Gene Expression 18, 71. doi: 10.3727/105221618X15156018385515
Dahl G. E., Tao S., and Laporta J. (2020). Heat stress impacts immune status in cows across the life cycle. Front. veterinary Sci. 7, 116. doi: 10.3389/fvets.2020.00116
Dikmen S. E. R. D. A. L. and Hansen P. J. (2009). Is the temperature-humidity index the best indicator of heat stress in lactating dairy cows in a subtropical environment? J. dairy Sci. 92, 109–116. doi: 10.3168/jds.2008-1370
Dikmen S. E. R. D. A. L., Khan F. A., Huson H. J., Sonstegard T. S., Moss J. I., Dahl G. E., et al. (2014). The SLICK hair locus derived from Senepol cattle confers thermotolerance to intensively managed lactating Holstein cows. J. dairy Sci. 97, 5508–5520. doi: 10.3168/jds.2014-8087
Domínguez-Gerpe L. and Rey-Méndez M. (2001). Alterations induced by chronic stress in lymphocyte subsets of blood and primary and secondary immune organs of mice. BMC Immunol. 2, 7. doi: 10.1186/1471-2172-2-7
Dunshea F. R., Gonzalez-Rivas P. A., Hung A. T., DiGiacomo K., Chauhan S. S., Leury B. J., et al. (2017). Nutritional strategies to alleviate heat stress in sheep. Sheep production adapting to Climate Change (Singapore, Singapore: Springer), 371–388. doi: 10.1007/978-981-10-4714-5_18
Fadare A. O., Peters S. O., Yakubu A., Sonibare A. O., Adeleke M. A., Ozoje M. O., et al. (2012). Physiological and haematological indices suggest superior heat tolerance of white-coloured West African Dwarf sheep in the hot humid tropics. Trop. Anim. Health production 45, 157–165. doi: 10.1007/s11250-012-0187-0
Foster M. W., Hess D. T., and Stamler J. S. (2009). Protein S-nitrosylation in health and disease: a current perspective. Trends Mol. Med. 15 (9), 391–404. doi: 10.1016/j.molmed.2009.06.007
Gu B. H., Park Y. J., Joo S. S., Lee S. S., Kim S. H., Kim E. T., et al. (2021). Dynamic changes in blood immune cell composition and function in Holstein and Jersey steers in response to heat stress. Cell Stress Chaperones 26, 705–720. doi: 10.1007/s12192-021-01216-2
Gu Z., Li L., Tang S., Liu C., Fu X., Shi Z., et al. (2018). Metabolomics reveals that crossbred dairy buffaloes are more thermotolerant than Holstein cows under chronic heat stress. J. Agric. Food Chem. 66, 12889–12897. doi: 10.1021/acs.jafc.8b02862
Hooper H. B., dos Santos Silva P., de Oliveira S. A., Merighe G. K. F., and Negrão J. A. (2018). Acute heat stress induces changes in physiological and cellular responses in Saanen goats. Int. J. Biometeorology 62, 2257–2265. doi: 10.1007/s00484-018-1630-3
Houten S. M., Watanabe M., and Auwerx J. (2006). Endocrine functions of bile acids. EMBO J. 25, 1419–1425. doi: 10.1038/sj.emboj.7601049
Jorge-Smeding E., Leung Y. H., Ruiz-González A., Xu W., Astessiano A. L., Trujillo A. I., et al. (2024). Plasma and milk metabolomics revealed changes in amino acid metabolism in Holstein dairy cows under heat stress. animal 18, 101049. doi: 10.1016/j.animal.2023.101049
Loor J. J., Lopreiato V., Palombo V., and D’Andrea M. (2023). Physiological impact of amino acids during heat stress in ruminants. Anim. Front. 13, 69–80. doi: 10.1093/af/vfad052
Marai I. F. M., El-Darawany A. A., Fadiel A., and Abdel-Hafez M. A. M. (2007). Physiological traits as affected by heat stress in sheep—a review. Small Ruminant Res. 71, 1–12. doi: 10.1016/j.smallrumres.2006.10.003
National Research Council (1971). A Guide to Environmental Research on Animals. Natl. Acad. Sci. (Washington, DC).
Ogunade I., Yun J., James A., Andre O., Diwakar V., and Adegbola A. (2018). Biomarker of aflatoxin ingestion: 1H NMR-based plasma metabolomics of dairy cows fed aflatoxin B1 with or without sequestering agents. Toxins. 10, 545. doi: 10.3390/toxins10120545
Ouellet V., Toledo I. M., Dado-Senn B., Dahl G. E., and Laporta J. (2021). Critical temperature-humidity index thresholds for dry cows in a subtropical climate. Front. Anim. Sci. 2, 706636. doi: 10.3389/fanim.2021.706636
Psychogios N., Hau D. D., Peng J., Guo A. C., Mandal R., and Bouatra S. (2011). The human serum metabolome. PloS One 6, e16957. doi: 10.1371/journal.pone.0016957
Rhoads M. L., Rhoads R. P., VanBaale M. J., Collier R. J., Sanders S. R., Weber W. J., et al. (2009). Effects of heat stress and plane of nutrition on lactating Holstein cows: I. Production, metabolism, and aspects of circulating somatotropin. J. dairy Sci. 92, 1986–1997. doi: 10.3168/jds.2008-1641
Ross J. W., Hale B. J., Seibert J. T., Romoser M. R., Adur M. K., Keating A. F., et al. (2017). Physiological mechanisms through which heat stress compromises reproduction in pigs. Mol. Reprod. Dev. 84, 934–945. doi: 10.1002/mrd.22859
Ruiz-González A., Suissi W., Baumgard L. H., Martel-Kennes Y., Chouinard P. Y., Gervais R., et al. (2023). Increased dietary vitamin D3 and calcium partially alleviate heat stress symptoms and inflammation in lactating Holstein cows independent of dietary concentrations of vitamin E and selenium. J. Dairy Sci. 106, 3984–4001. doi: 10.3168/jds.2022-22345
Slimen I. B., Chniter M., Najar T., and Ghram A. (2019). Meta-analysis of some physiologic, metabolic, and oxidative responses of sheep exposed to environmental heat stress. Livestock Sci. 229, 179–187. doi: 10.1016/j.livsci.2019.09.026
Srikandakumar A., Johnson E. H., and Mahgoub O. (2003). Effect of heat stress on respiratory rate, rectal temperature and blood chemistry in Omani and Australian Merino sheep. Small Ruminant Res. 49, 193–198. doi: 10.1016/S0921-4488(03)00097-X
Strong R. A., Silva E. B., Cheng H. W., and Eicher S. D. (2015). Acute brief heat stress in late gestation alters neonatal calf innate immune functions. J. dairy Sci. 98, 7771–7783. doi: 10.3168/jds.2015-9591
Tadesse D., Puchala R., Gipson T. A., and Goetsch A. L. (2019). Effects of high heat load conditions on body weight, feed intake, temperature, and respiration of Dorper, Katahdin, and St. Croix sheep. J. Appl. Anim. Res. 47, 492–505. doi: 10.1080/09712119.2019.1674658
Tüfekci H. and Sejian V. (2023). Stress factors and their effects on productivity in sheep. Animals 13 (17), 2769. doi: 10.3390/ani13172769
Keywords: Florida Cracker sheep, heat stress, plasma metabolome, adaptation, thermotolerance
Citation: Harrison M, Adebayo JO, Mills-Widemon A, Ikuejamoye-Omotore S, Estrada-Reyes ZM, Ogunade IM, Pech-Cervantes AA, Terrill TH, Siddique A and Ruiz-González A (2025) Short term heat stress alters the hematological, immunological, and plasma metabolomic responses of Florida Cracker sheep. Front. Anim. Sci. 6:1631531. doi: 10.3389/fanim.2025.1631531
Received: 19 May 2025; Accepted: 23 July 2025;
Published: 01 September 2025.
Edited by:
Vishal Suthar, Gujarat Biotechnology University, IndiaReviewed by:
Weinan Zhou, Louisiana Tech University, United StatesHalima Sultana, University of Florida, United States
Copyright © 2025 Harrison, Adebayo, Mills-Widemon, Ikuejamoye-Omotore, Estrada-Reyes, Ogunade, Pech-Cervantes, Terrill, Siddique and Ruiz-González. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Zaira M. Estrada-Reyes, emVyZXllc0BwdmFtdS5lZHU=