 Longfei Yan
Longfei Yan Jinling Hua
Jinling Hua Wenwen Lu
Wenwen Lu Huwei Zhao1,2
Huwei Zhao1,2- 1College of Animal Science, Anhui Science and Technology University, Chuzhou, China
- 2Anhui Provincial Department of Science and Technology, Anhui Provincial Key Laboratory of Animal Nutritional Regulation and Health, Anhui Science and Technology University, Chuzhou, China
In this study, we examined the impact of various maize to maize varieties on the performance and digestibility of hu sheep. A total of thirty-six healthy male hu sheep, all of similar weight, were selected and randomly assigned to one of three groups, each comprising 12 sheep. The groups were designated as CS1 (fed HanDan 777), CS2 (fed DengHai 605), and CS3 (fed NongDa 367). The study included a pre-feeding period of 10 days followed by an experimental period of 80 days. The findings indicated that the total weight gain and average daily gain in the CS2 group were higher than those in the CS1 group (p < 0.05), and the feed-to-weight ratio in the CS2 group was lower than those in the CS1 and CS3 groups (p < 0.05). The pre-slaughter live weight in the CS2 group was higher than in the CS1 and CS3 groups. Additionally, the L* value of the CS3 group was higher than those of the CS1 and CS2 groups (p < 0.05). The shear force of the CS3 group was greater than that of the CS2 group (p < 0.05). Compared with the CS1 group, both the CS2 and CS3 groups exhibited higher water loss rates (p < 0.05). Moreover, the cooking loss of the CS2 group was higher than those of the CS1 and CS3 groups (p < 0.05). There was a significant effect of the total weight gain and average daily weight gain in the CS2 group compared to the CS1 group (p < 0.05). These findings suggest that the DengHai 605 corn variety (CS2) is a suitable option for use as silage in Hu sheep diets due to its positive impact on growth performance.
1 Introduction
In recent times, Whole-plant maize silage has become a key feed for ruminants (e.g., dairy cows, beef cattle, and sheep) due to its palatability, storability, and weather resilience (Cheli et al., 2013). Over the past 25 years, its adoption has grown significantly, supporting efficient ruminant production worldwide (Ferraretto et al., 2018). As the second-largest maize producer globally, China has substantial potential to utilize maize for silage, offering farmers a cost-effective way to enhance profitability through optimized variety selection.
Maize silage improves ruminant growth performance and meat quality while reducing nutrient losses compared to fresh forage (ZoBell et al., 2002). Its economic and preservation advantages make it a practical choice for livestock operations. Additionally, repurposing crop by-products like maize for silage aligns with circular economy principles, promoting sustainable resource use (Ranum et al., 2014). By integrating silage into farming systems, producers can achieve both economic and environmental benefits, ensuring long-term agricultural sustainability.
Demiral et al. studied the nutritive value of silage from 12 maize varieties and found that the nutritive value of silage varies between varieties (Demirel et al., 2011). The choice of maize variety is crucial, as different varieties differ in nutritive value, harvesting time, and other factors. Additionally, the genetic background of maize varieties can affect both yield and quality. Liu et al. found that the selection of maize hybrids can influence several aspects of maize silage, including harvest yield, grain quantity at harvest time, and digestibility (Liu et al., 2021). Nazli et al. observed wide variation among maize varieties, with sweet maize typically having the lowest yields but slightly superior nutritional quality, while grain maize varieties tend to have higher yields but poorer quality (Nazli et al., 2018). Acharya and Casper found that different silage maize varieties did not affect milk production in ruminants but did result in varying economic benefits (Acharya and Casper, 2020). In contrast, Akins and Shaver (2014), in their study on the effect of maize silage hybrids on lactation performance in ruminants, found that feeding nutrient-dense silage maize increased both feed intake and milk production (Akins and Shaver, 2014). Zaralis and his colleagues demonstrated that the daily weight gain of beef cattle exclusively fed with whole-plant maize silage was notably higher than that of those fed a combination of whole-plant maize silage and forage silage (Zaralis et al., 2014). Moreover, whole-plant maize silage has the remarkable ability to effectively boost milk production, enhance the quality of milk, cut down on breeding expenses, and elevate economic returns (Hameleers, 1998).
Domestic studies have shown that, compared to maize stover silage, the inclusion of whole-plant maize silage in feed rations can improve the production performance and economic benefits of ruminants (Zhang et al., 2021; Sha et al., 2024). In light of this, the present experiment was conducted using Hu sheep as the research subjects, with whole-plant maize silage as the roughage source. We hypothesized that different varieties of maize silage would have varying effects on the growth performance, slaughter performance, and meat quality of mutton sheep. The results demonstrate that the Denghai 605 (CS2) whole-plant corn silage variety significantly enhances production performance and meat quality parameters in Hu sheep compared to other varieties.
1.1 Experimental design and animals
The experimental site was situated at Xinqun Herd, Songji Town, Linquan County, Anhui Province, China. Thirty-six weaned Hu sheep (2-month-old, body weight: 17.44 ± 1.51 kg) were selected and randomly allocated into three groups (CS1, CS2, and CS3). Each group comprised three replicate pens, with four sheep per pen. A 10-day pre-feeding period was conducted, followed by an 80-day formal feeding trial. Throughout the experiment, all sheep were managed under identical feeding conditions with ad libitum access to feed and water.
1.2 Experimental rations
The three types of maize were planted on June 7, 2023, and harvested on September 4, 2023, and the maize stover was crushed into 2–3 cm pieces, compacted with a compactor and then loaded into fermentation bags, and used for feeding experiments after 60 d of anaerobic fermentation. After 60 days of silage fermentation, the silage feed in 10 bags was randomly selected for sampling. The experimental ration was sampled using the quartering method, and the determination method referred to the approach described in “Feed Analysis and Quality Testing Technology” (Zhang Liying, Fourth Edition).
The analytical methods for feed nutritional components are as follows:
1. Dry matter (DM).
● Method: AOAC 934.01.
● Procedure: Weigh 2 g of the sample, dry it at 103 ± 2°C for 24 hours in a forced-air oven (Memmert UN110).DM (%)=W1(W1−W2)×100.
where W1 = initial weight, W2 = weight after drying.
2. Crude protein (CP).
● Method: AOAC 928.08.
● Apparatus: Shandong Haineng k9840 automatic Kjeldahl nitrogen analyzer.
● Protocol: Weigh 0.5g of feed sample, digest it at a temperature of about 350-420°C. After digestion and cooling, use K9840 for determination.CP (%)=Nitrogen content×6.25.
3. Ether extract (EE).
● Method: AOAC 960.39.
● Solvent: ether (boiling point 40–60°C).
● Procedure: Extract 2 g of the sample with the solvent for 6 hours, evaporate the solvent, and weigh the residue.
4. Ash.
● Method: AOAC 942.05.
● Procedure: Incinerate 2 g of the sample in a muffle furnace (Nabertherm L3/11) at 550°C for 6 hours.
5. Neutral detergent fiber (NDF).
● Method: AOAC 973.18.
● Reagents: Sodium Dodecyl Sulfate Solution.
● Protocol: Reflux 0.5 g of the sample in neutral detergent solution for 1 hour(Ankom 220). Wash the residue with acetone and dry at 105°C.
6. Acid detergent fiber (ADF).
● Method: AOAC 973.18.
● Reagent: Cetyltrimethylammonium bromide solution.
● Procedure: Reflux for 1 hour, filter through filter bags (Ankom 220), and dry at 105°C.
7. Quality control.
● All analyses are performed in triplicate.
● Certified reference materials (e.g., NIST SRM 1547) are included.
● Blank corrections are applied to fiber analyses.
● Results are expressed on a dry matter (DM) basis.
The composition and nutrient content of the rations are shown in the table below.
1.3 Measurement indicators and methods
1.3.1 Growth performance
At both the start and end of the experiment, the sheep were fasted for 12 hours (with free access to water) and then weighed. These measurements were recorded as the initial body weight (IBW) and final body weight (FBW), respectively. The experimental diet was provided twice daily at 8:00 and 18:00, and feed intake as well as leftover amounts were meticulously recorded throughout the entire experimental period.
The growth performance indices were calculated as follows (Equations 1–3):
1.3.2 Slaughtering performance
At the end of the experiment, five sheep were selected from each group, fasted for 24 hours, and slaughtered. The live weight before slaughter was recorded, blood was drained, and the carcass weight was determined by removing the skin, head, hooves, offal, and testes. After removing the bones, the net meat weight was measured, and the slaughter rate and net meat rate were calculated. The eye muscle between the 12th and 13th ribs on the back was taken, and the area of the eye muscle was calculated using sulfuric acid graph paper (Equations 4, 5).
1.3.3 Measurement of flesh color
The brightness (L*), redness (a*), and yellowness (b*) of the longest dorsal muscle were measured using a CR-400 colorimeter(Konica Minolta, Tokyo, Japan). The values were averaged based on three measurements taken at different positions.
1.3.4 Drip loss and shear force
Drip Loss: A sample of the longissimus dorsi (LD) muscle measuring 2 cm × 3 cm × 5 cm was taken, and the fascia was removed. The sample was weighed, hung with a wire, placed in a sealed plastic bag, and stored at 4°C for 24 hours. After 24 hours, the sample was weighed again, and the drip loss was calculated.
Shear Force: A sample of the LD muscle was heated in a water bath at 80°C until the center of the meat reached 70°C. After cooling, the shear force was measured using an RH-N50 instrument.
1.3.5 Cooking loss and water holding capacity
Cooking Loss: A sample of the longissimus dorsi (LD) muscle was trimmed to 2 cm × 2 cm × 3 cm, and the fascia was removed. The sample was weighed, placed in a sealed bag, and heated in a water bath for 30 minutes. After heating, the sample was placed indoors in a cool place to dry for 15 minutes and then weighed again to calculate the cooking loss. The cooking loss (%) was calculated using the following formula: Cooking loss (%)=(Pre-steaming meat weight−Post-steaming meat weight)/Pre-steaming meat weight×100%(6).
Water holding capacity: The meat sample was placed between 16 layers of filter paper (8 layers on top and 8 layers on the bottom) and positioned on a water loss rate plate. The RH-100 instrument was used to apply a pressure of 35 kg for 5 minutes. The sample was weighed immediately after pressing, and the water loss rate was calculated. The formula for water loss rate was as follows: Water holding capacity (%)=(Weight of meat before pressing−Weight of meat after pressing)/Weight of meat before pressing×100%(7).
1.3.6 Apparent digestibility of nutrients
At the end of the experiment, a 7-day digestive metabolism test was conducted, including a 3-day acclimatization period and a 4-day fecal collection period. During this time, all hu sheep were transferred to individual metabolism pens equipped with plastic sieves and plastic film underneath for fecal and urine collection. Feces and urine were collected before feeding, and the collected fresh feces were treated with 10% sulfuric acid to prevent ammonia-nitrogen volatilization. All feed and fecal samples were dried at 65°C for 48 hours, filtered through a 1-mm mesh sieve, and stored in self-sealing bags for subsequent routine nutritional analysis. The formula for calculating the apparent digestibility of nutrients was as follows: Apparent digestibility (%) = [(nutrient intake - nutrient excretion in feces)/nutrient intake] × 100%. (8).
1.3.7 Rumen microbiota
At the end of the experiment, rumen fluid samples were collected from Hu sheep and aliquoted into 2 mL sterile cryovials. The samples were immediately flash-frozen in liquid nitrogen and subsequently stored at -80°C until 16S rRNA sequencing could be performed. The preserved samples.
At the conclusion of the experiment, rumen fluid samples were collected from Hu sheep and aliquoted into 2 mL sterile cryovials. The samples were immediately flash-frozen in liquid nitrogen and subsequently stored at -80°C until 16S rRNA sequencing could be performed. The preserved samples were transported on dry ice to Sangon Biotech (Shanghai) Co., Ltd., where sequencing was conducted following the company’s standardized operating procedures.
1. Total Microbial DNA Extraction.
Total microbial DNA was extracted from lamb rumen fluid samples using the OMEGA E.Z.N.A.® Mag-Bind Soil DNA Kit. Specific primers targeting the V3-V4 hypervariable region of the 16S rRNA gene were designed and synthesized to amplify the 16S rRNA gene from genomic DNA samples.
2. Sequencing and Data Processing.
Paired-end sequencing was performed on the Illumina NovaSeq platform to generate high-throughput sequence data. The sequencing reads contained barcode sequences, as well as primer and adapter sequences introduced during library preparation. Raw data were processed through the following standardized pipeline:
● Primer/Adapter Trimming: Sequences containing primer or adapter fragments were removed using Cutadapt software with a minimum match length of 10 bp.
● Read Merging: Paired-end reads were merged into single contigs using FLASH software, requiring a minimum overlap of 10 bp and a maximum mismatch rate of 0.2.
● Sample Demultiplexing: Samples were distinguished based on barcode tags, allowing for 1 bp mismatch in barcode sequences to ensure accurate partitioning.
● Quality Filtering: Reads with an average Phred score <20, single-end length <200 bp, or ambiguous bases were discarded. Each sample retained a minimum of 5,000 valid reads to ensure statistical reliability.
● Chimera Removal: Chimeric sequences were identified and eliminated using the UCHIME algorithm against the Silva 138 reference database.
3. Bioinformatics Analysis.
● OTU Clustering: High-quality sequences were clustered into operational taxonomic units (OTUs) at a 98% similarity threshold using Usearch software. OTUs with a minimum sequence count of 2 were retained, while singletons (OTUs with only 1 read) were filtered out.
● Alpha Diversity Analysis: Diversity indices (Chao1, Shannon, Simpson) were calculated using Mothur software with rarefaction analysis to assess community richness and evenness. Visualization of rarefaction curves and rank-abundance plots was performed using R software, with a requirement for ≥99% sequencing depth coverage to ensure comprehensive sampling.
● Beta Diversity Analysis: Differences in species abundance distribution between samples were quantified using Euclidean distance, Bray-Curtis distance, Unweighted UniFrac, and Weighted UniFrac metrics. Distance matrices were generated for principal coordinate analysis (PCoA) and non-metric multidimensional scaling (NMDS), with visualizations constructed using the R vegan package to illustrate inter-sample relationships.
All experimental and bioinformatics procedures were executed by Sangon Biotech (Shanghai) Co., Ltd. on the Illumina NovaSeq platform following established industry standards, ensuring rigorous quality control and reproducibility of microbial sequencing data.
Generated Statement: The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://www.ncbi.nlm.nih.gov/, PRJNA128884 https://www.ncbi.nlm.nih.gov/, SUB15355450.
1.3.8 Statistical analysis
The data were subjected to one-way ANOVA analysis using SPSS 27.0 statistical software. Results are presented as mean ± SEM, with statistical significance set at p < 0.05.
2 Result
2.1 Nutritional value of silage material
Table 1 shows that the CS3 group had a substantially greater dry matter (DM) content than the CS2 and CS1 groups (p < 0.05), and the CS2 group had a higher crude protein (CP) content than the CS1 groups (p < 0.05). Furthermore, the CS1 group had substantially greater amounts of acid detergent fiber (ADF) and neutral detergent fiber (NDF) than the CS2 and CS3 groups (p < 0.05).
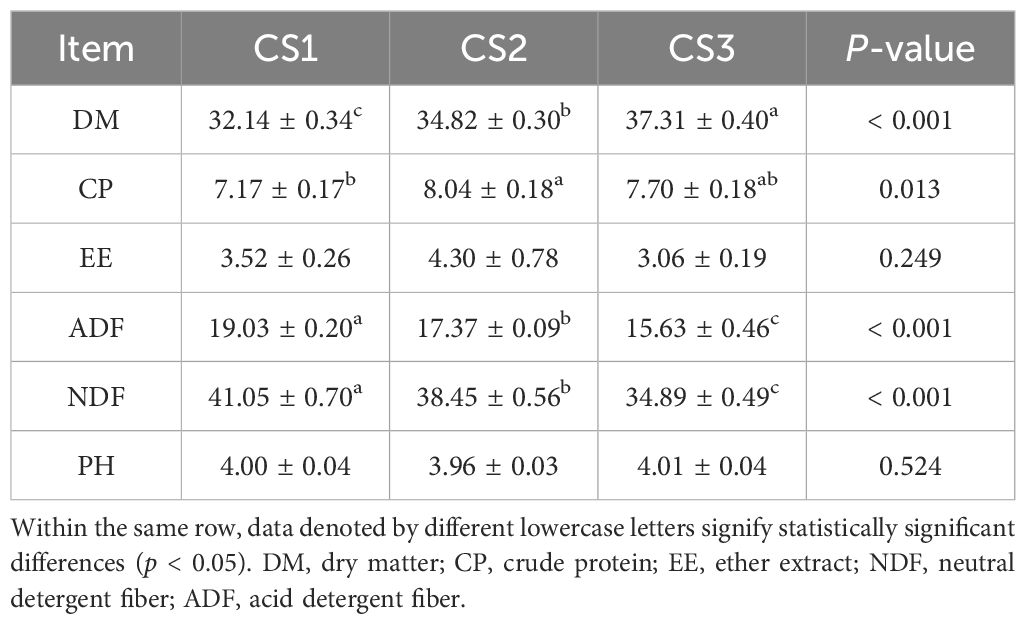
Table 1. The conventional nutritional value of different silage materials (%, except where otherwise stated).
2.2 Effect of different varieties of maize silage on the production performance of Hu sheep
As shown in Table 2, the CS2 group exhibited significantly higher final body weight (FBW) and average daily gain (ADG) compared to the CS1 group (p < 0.05), along with a significantly lower feed-to-gain ratio than both CS1 and CS3 groups (p < 0.05). The CS3 group demonstrated significantly greater dry matter intake (DMI) than both CS1 and CS2 groups (p < 0.05).
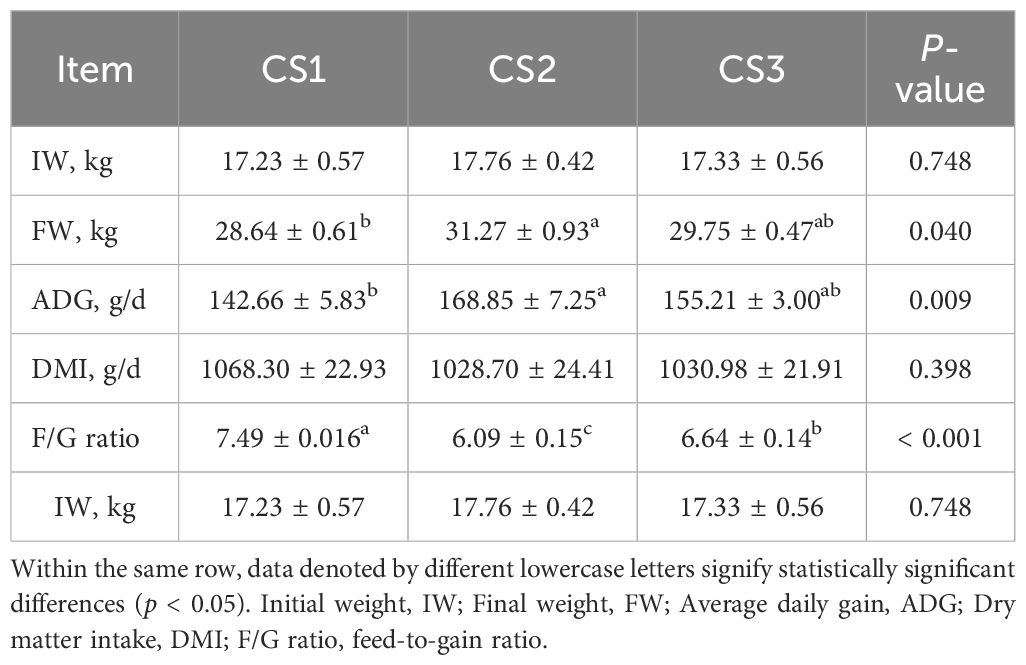
Table 2. Effect of different varieties of maize silage on the production performance of Hu sheep.
2.3 Effect of different varieties of maize silage on the quality of the longest muscle of the back of Hu sheep
The slaughter performance of the three treatment groups was compared, as indicated in Table 3. In comparison to the CS1 and CS2 groups, the CS3 group’s L* value was considerably greater (p < 0.05). The water holding capacity of the CS3 group was substantially higher than that of the CS1 group (p < 0.05), while the CS3 group showed a higher shear force value than the CS2 group (p < 0.05). Furthermore, the CS2 group saw a considerably larger cooking loss than both the CS1 and CS3 groups (p < 0.05).
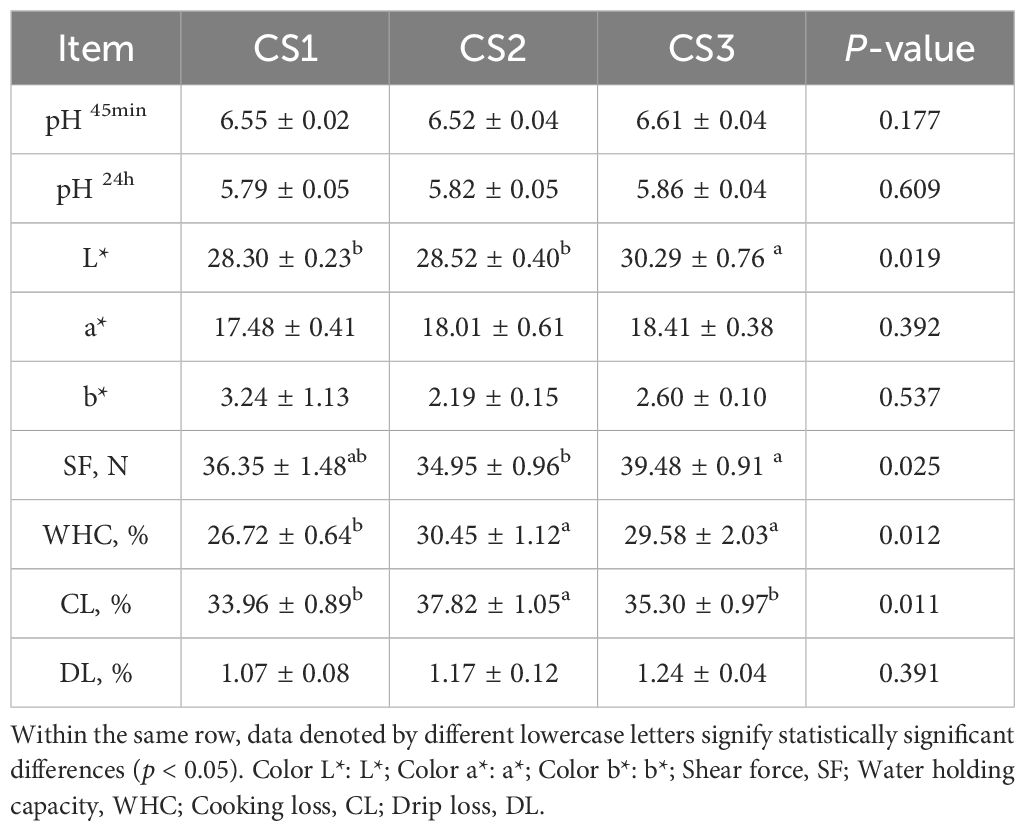
Table 3. Effect of different varieties of maize silage on the quality of the longissimus dorsi (LD) quality of Hu sheep.
2.4 Effect of different varieties of maize silage on slaughter performance of Hu sheep
The pre-slaughter live weight of the CS2 group was greater than that of the CS1 group, as seen in Table 4. In terms of slaughter performance, the CS2 group’s carcass weight was considerably higher than the CS3 group’s (p < 0.05). Furthermore, there was a significant difference (p < 0.05) in the slaughter rate between the CS1 and CS3 groups. However, the eye muscle area did not differ significantly among the groups (p > 0.05).
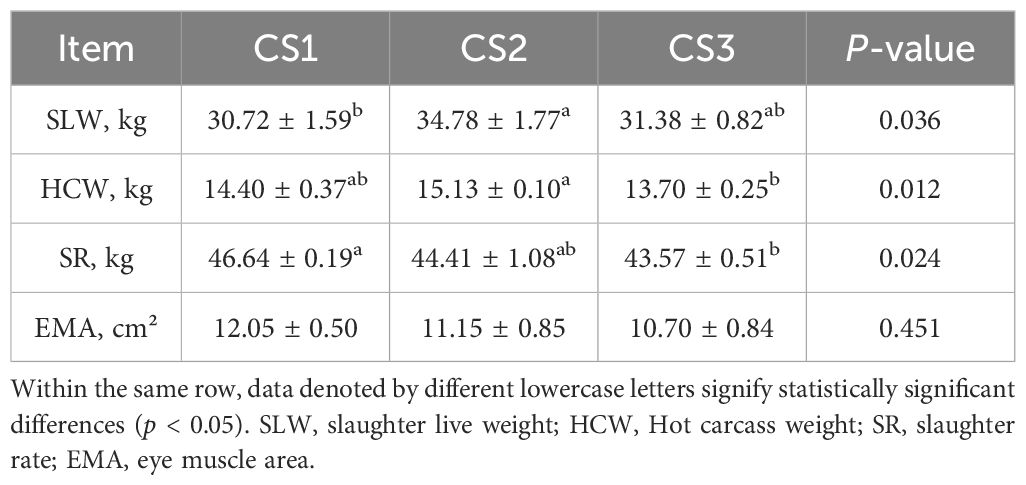
Table 4. Effect of different varieties of maize silage on slaughter performance of Hu sheep.
2.5 Effect of different varieties of maize silage on the digestibility of nutrients in Hu sheep
The apparent nutritional digestibility of diets in hu sheep fed various types of maize silage was assessed, as indicated in Table 5. The digestibility of dry matter (DM), organic matter (OM), crude protein (CP), ether extract (EE), neutral detergent fiber (NDF), and acid detergent fiber (ADF) did not differ (p > 0.05) between the CS1, CS2, and CS3 groups.
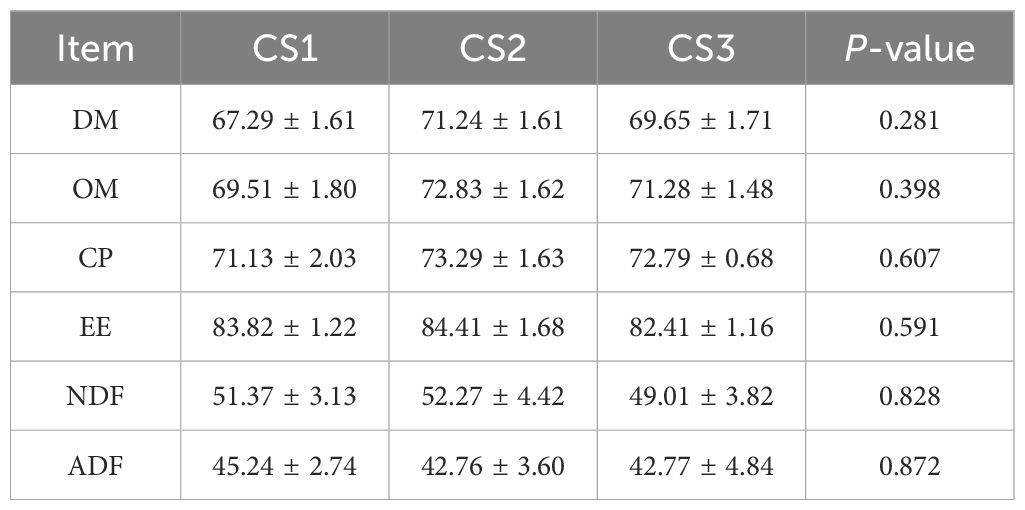
Table 5. Effect of different varieties of maize silage on nutrient digestibility of Hu sheep (%, except where otherwise stated).
2.6 Effect of different varieties of maize silage on nitrogen utilization in Hu sheep
As shown in Table 6, the effect of whole-plant maize silage on nitrogen utilization in hu sheep is presented. There were no significant differences (p > 0.05) among the CS1, CS2, and CS3 groups in terms of ingested nitrogen, total excreted nitrogen, deposited nitrogen, nitrogen utilization, nitrogen apparent absorption, and biological value of nitrogen.
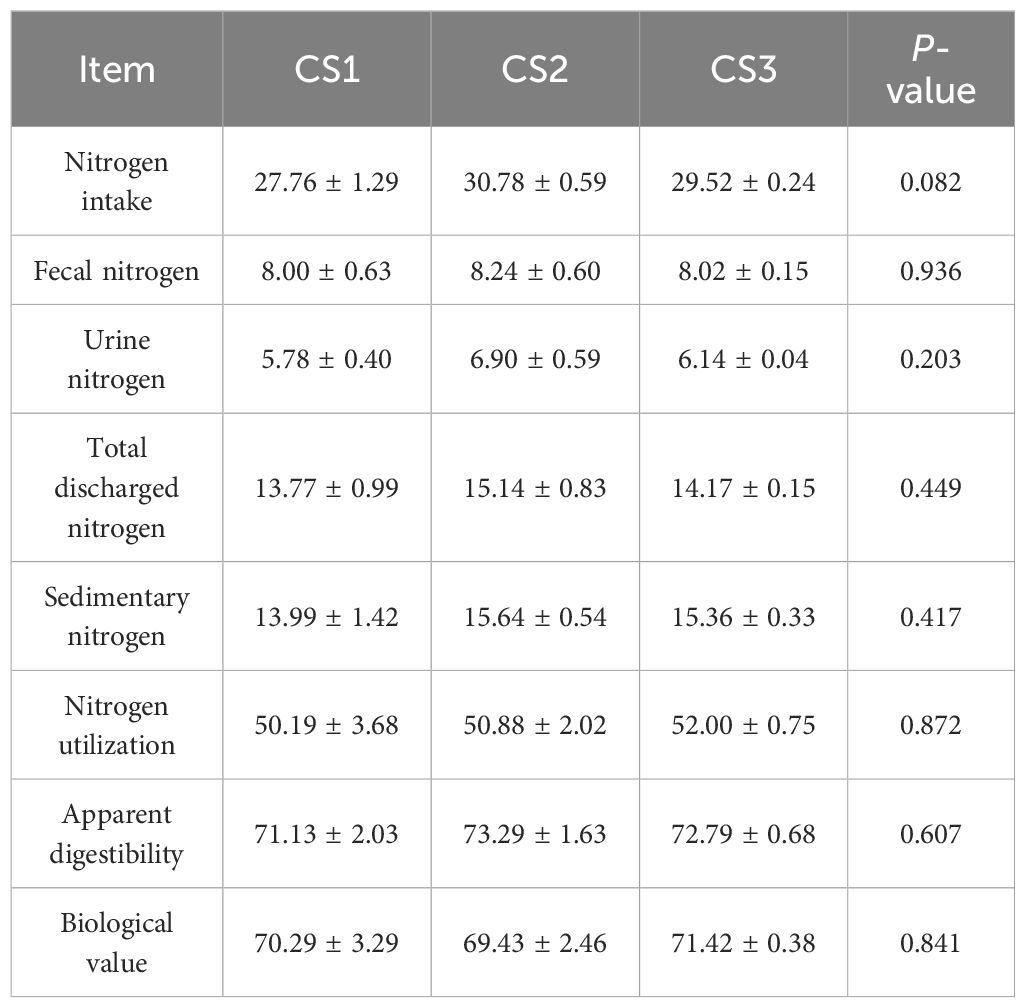
Table 6. Effect of different varieties of maize silage on nitrogen utilization in Hu sheep (%).
2.7 Effects of different varieties of maize silage on the alpha-diversity of rumen microorganisms in Hu sheep
To explore the impact of whole-plant maize silage on rumen microbiota in hu sheep, 16S rRNA gene sequencing was conducted on rumen fluid samples from all experimental groups. The rarefaction curve, which leveled off with an increasing number of sequences, demonstrated that the sequencing depth was adequate to capture the abundance and diversity of the majority of microbial communities present (Figure 1).
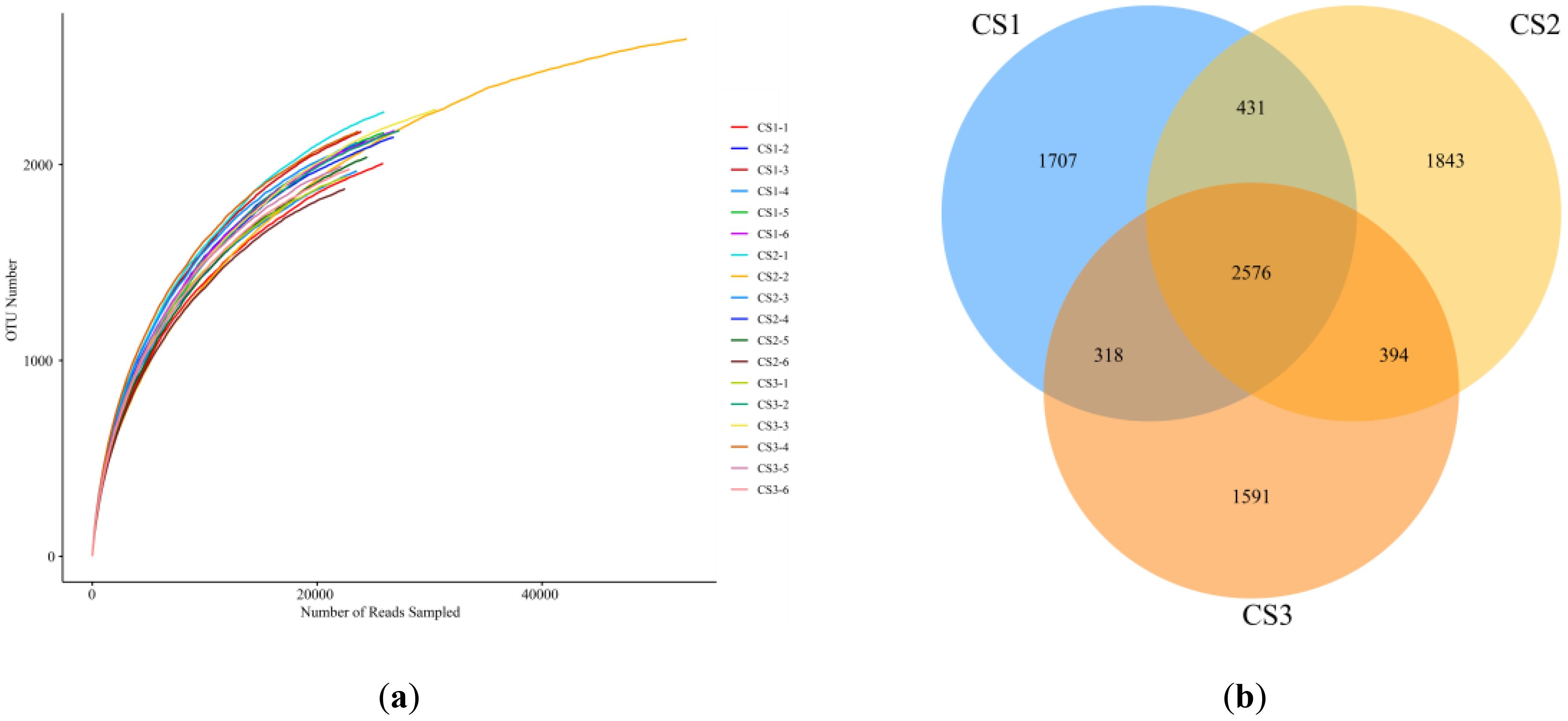
Figure 1. The effect of using different whole-plant maize silages on the rumen microbiota of hu sheep was investigated. (a) Microbial dilution curves based on observational metrics were used to assess the depth of coverage of each sample (samples were distinguished by different colored lines). (b) Venn diagrams showing the composition of rumen microbial operational taxonomic units (OTUs) for the CS1, CS2, and CS3 groups of whole-plant maize silage. Before the end of the experiment, specimens were collected from 6 sheep per group.
Based on 97% similarity, Venn diagrams showed 2,600 common OTUs and 1,477, 1,869, and 1,852 unique OTUs in the CS I, CS II, and CS III groups, respectively.
2.8 Comparison of microbial communities (beta-diversity)
Beta diversity results are shown in Figure 2. Only a small number of samples showed deviations, while the rest of the samples exhibited relative proximity and lacked any recognizable segregation. These results indicate that there were no substantial differences in the structural composition of the microorganisms among the three groups, suggesting that the use of different silage maize did not alter the composition of the rumen microorganisms in hu sheep.
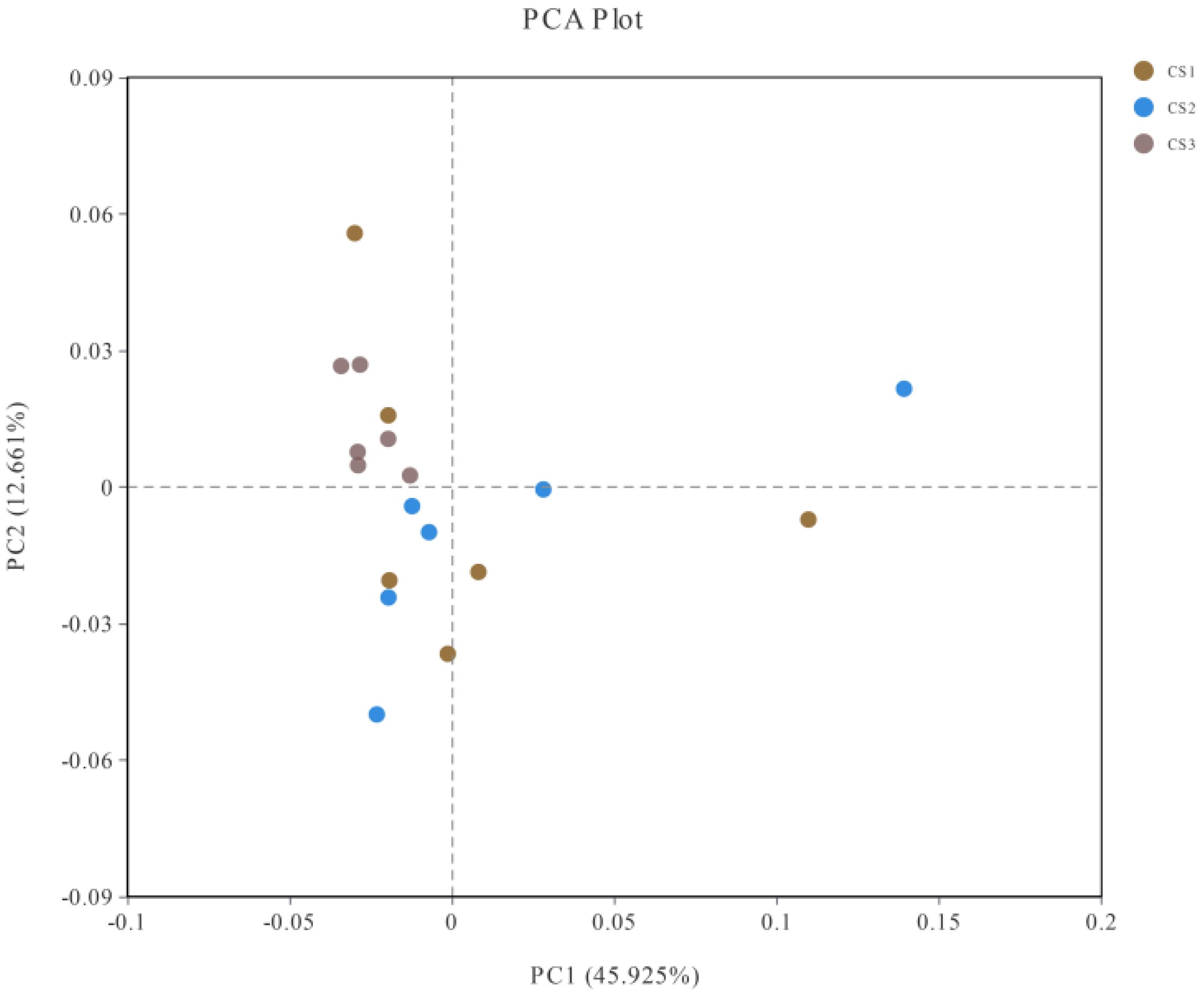
Figure 2. Effect of using different silage maize on Beta diversity of rumen microorganisms.
2.9 Effect of different varieties of maize silage on the composition of rumen microflora in Hu sheep
Table 7 shows the taxonomic composition of the top ten gastrointestinal microbial communities at the phylum level. Bacteroidota was the most abundant phylum, followed by Firmicutes. Notably, the relative abundance of Verrucomicrobia in the CS1 group was higher than that in the CS2 and CS3 groups (p < 0.05). Figure 3 depicts the distribution of dominant phyla at the phylum level.
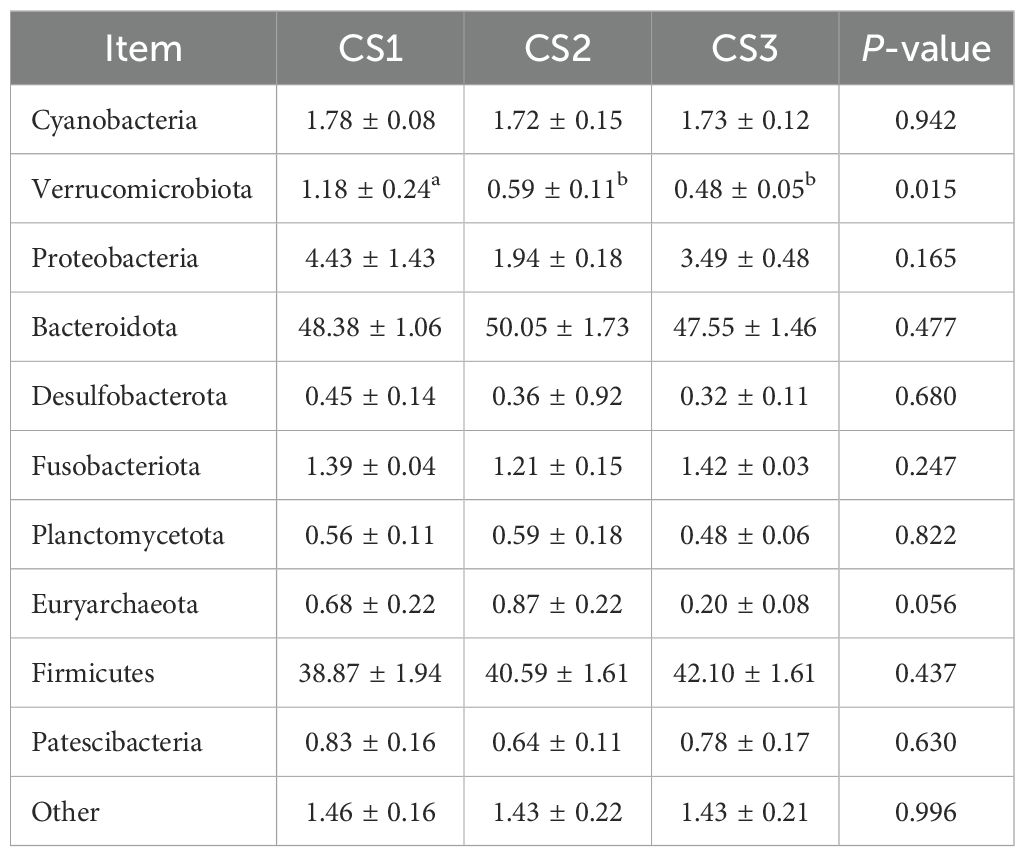
Table 7. Effect of different varieties of maize silage on the level of rumen flora phylum in Hu sheep.
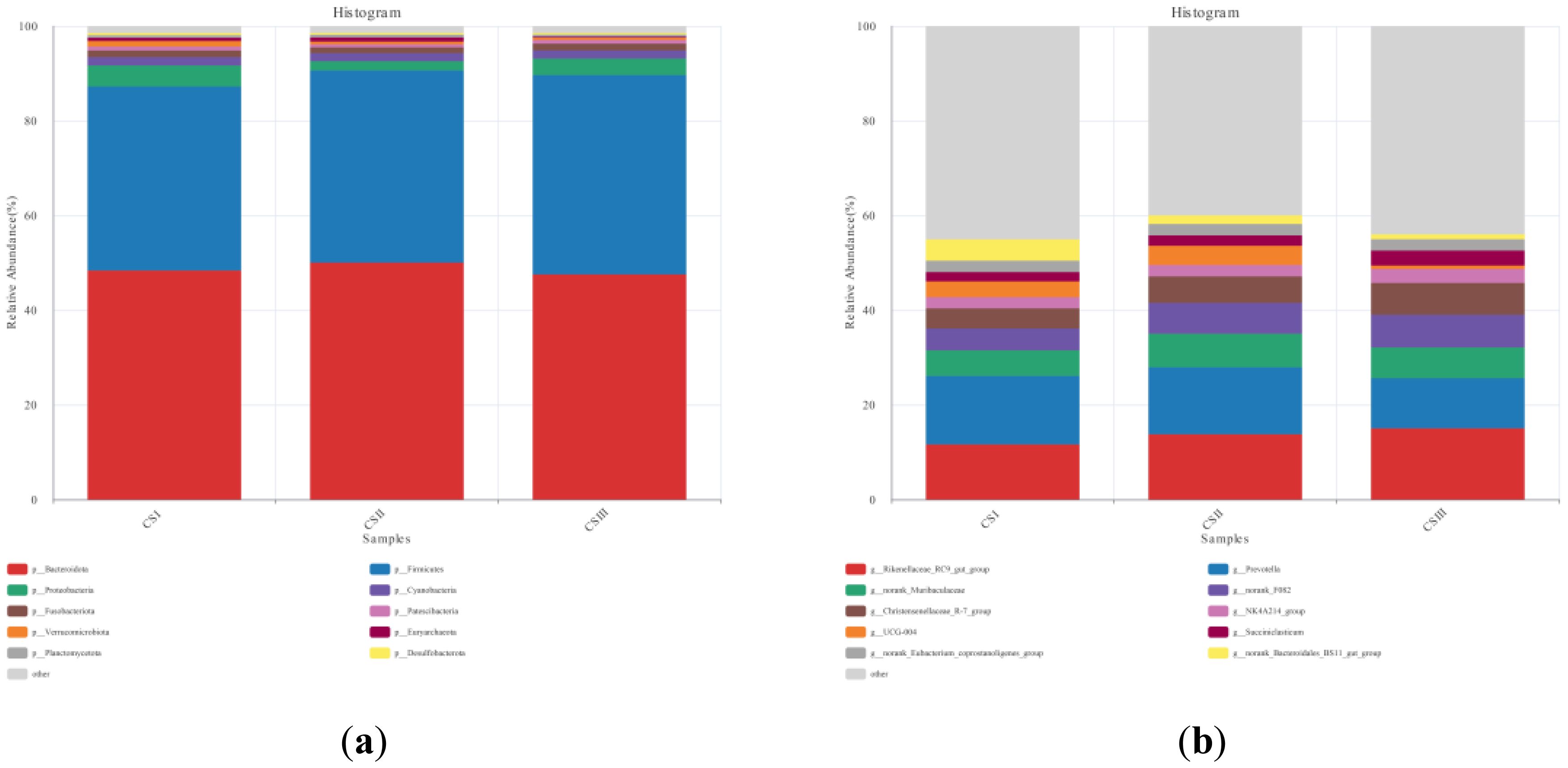
Figure 3. Relative abundance of each group of bacteria (a) phylum level (b) genus level.
The dominant microorganisms at the genus level are depicted in Figure 3. The taxonomic composition of the top 10 intestinal microbial communities at the genus level is detailed in Table 8 Prevotella spp. showed the highest relative abundance, followed by Rikenellaceae. The relative abundance of Christensenellaceae in the CS3 group was higher than that in the CS1 group. Similarly, the relative abundance of Bacteroidales in the CS1 group was greater than that in the CS2 and CS3 groups.
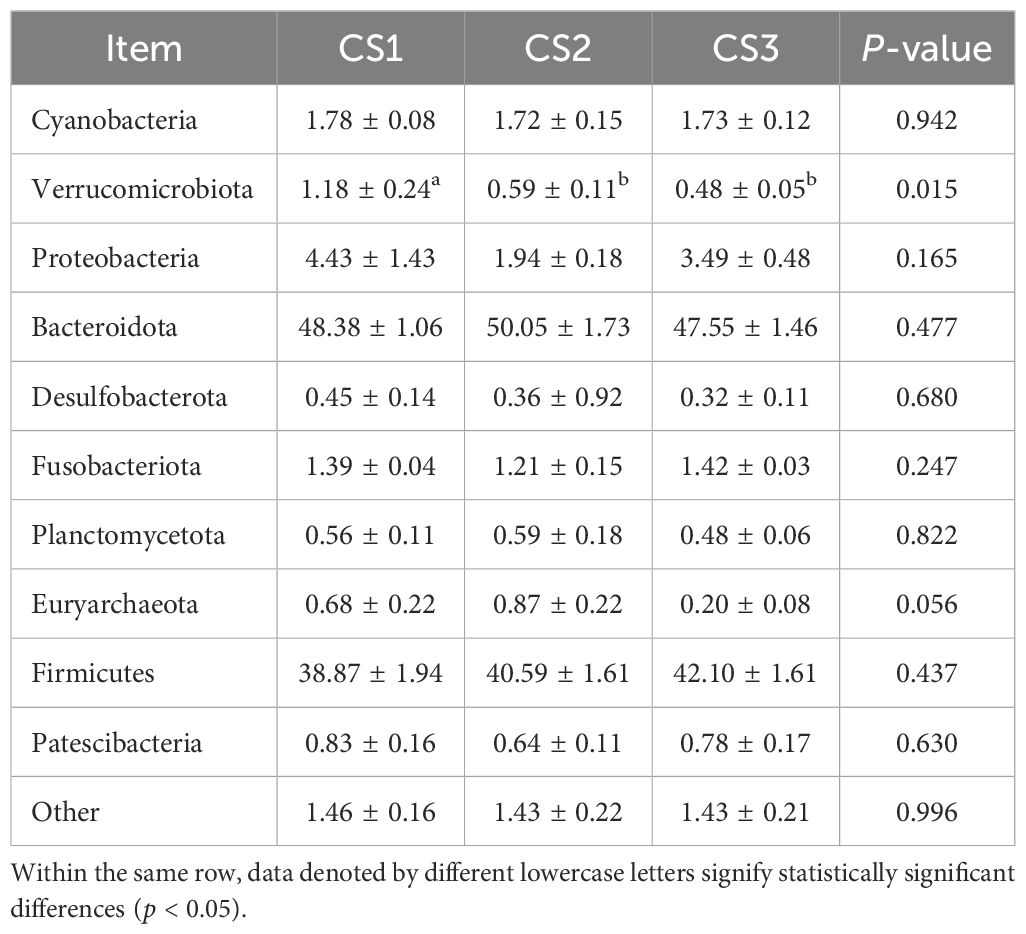
Table 8. Effect of different varieties of maize silage on the level of rumen flora genus in Hu sheep.
3 Discuss
Maize silage is commonly used in ruminant production, and studies have shown that harvest date and variety of maize are the two main factors affecting the yield and quality of maize silage, in addition to climatic differences (Nazli et al., 2019; Wang X. et al., 2020). Terler studied the nutritional value of nine different maize silages over a three-year period and concluded that variety and harvest date had a significant effect on CP and NDF (Terler et al., 2019b). Terler et al. demonstrated that even when the nutrient composition of different maize varieties does not differ significantly, incorporating distinct maize varieties into the diet significantly influences ruminal degradability (Terler et al., 2019a). Liu et al. found that dual-purpose maize silage had higher MT(milk-per-acre) and energy values, while silage-specific maize silage had lower yields per unit of DM and MA(milk-per-ton index), probably because dual-purpose maize silage has higher yields per unit of DM and energy values (Liu et al., 2021). We observed that in this experiment, according to Table 9, there were differences in DM, CP, ADF, and NDF of different maize silages, and the planting time and harvesting time were the same. Some studies have shown that none of the energy values were consistent among different maize varieties, while in this experiment, the differences may be due to the differences in varieties.
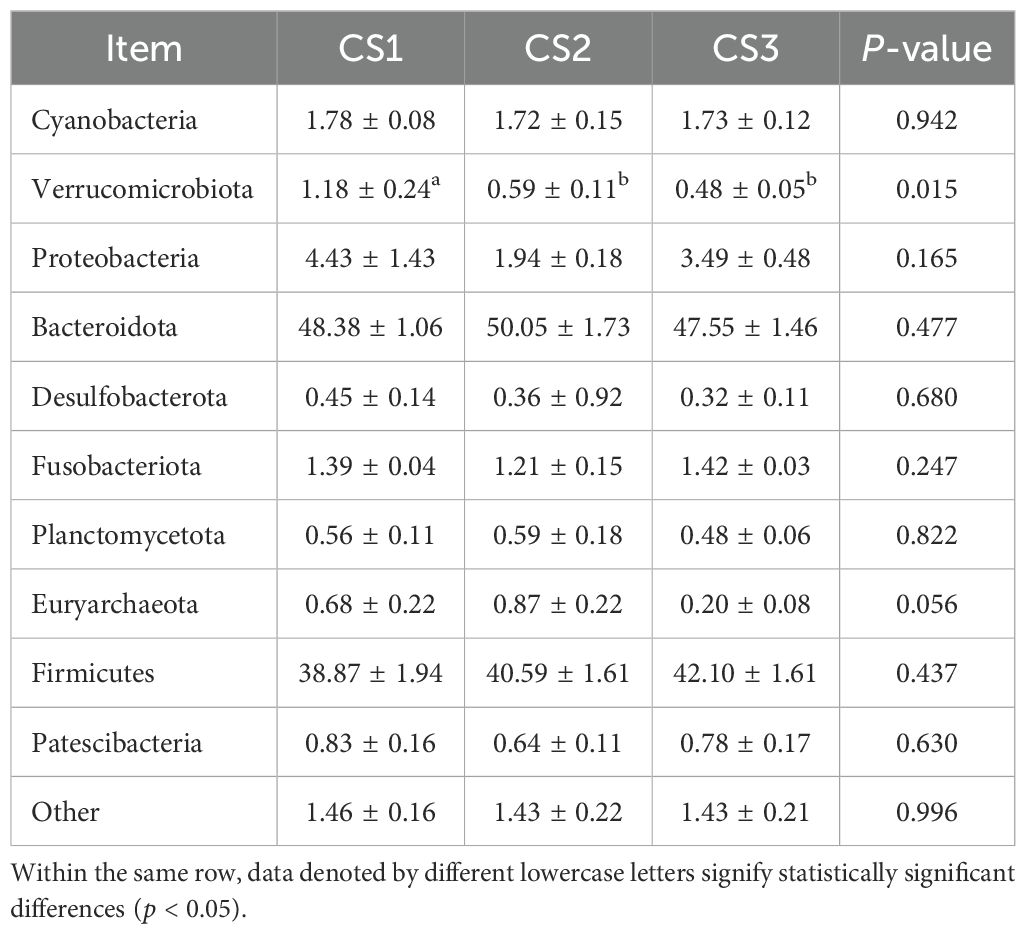
Table 9. Nutritional composition and nutrient levels of diets (% DM, except where otherwise stated).
Body weight, daily weight gain, and dry matter intake in ruminants are positively correlated with the CP content in feed (Devincenzi et al., 2019). Studies have demonstrated that as the energy level of the diet increases, daily weight gain and dry matter intake significantly rise, while the feed-to-meat ratio significantly decreases (Wang et al., 2021). In this experiment, the using different maize silage varieties resulted in significant variations in ADG of Hu sheep. This phenomenon can be primarily attributed to the divergent nutrient compositions that emerged after silage fermentation among various maize varieties, which in turn influenced the growth and development of Hu sheep. Previous research has shown that the protein content of feed can significantly influence dietary intake, health status, and growth performance of livestock (Wang X. et al., 2020; Lee et al., 2021). Moreover, ADG and total weight gain are critical indicators of animal growth performance, reflecting the growth rate and nutrient utilization efficiency of animals. The ADG and total weight gain of the CS2 group were significantly higher than those of the CS1 and CS3 groups, and the feed-to-weight ratio was significantly lower than those of the CS1 and CS3 groups. Daily weight gain serves as the most direct measure of the digestive and absorptive efficiency of feeds (Li et al., 2024). Most previous experiments have verified that protein and energy within a certain range jointly influence animal metabolism, growth, and development. However, there are also experiments showing that the primary factor affecting animal weight gain is the varied protein-to-fat ratios of raw materials in the diet (Arriola Apelo et al., 2014). The daily weight gain of the CS2 group was significantly higher than that of the CS1 and CS3 groups, which may be because silage raw materials in the CS2 group had the highest content of crude protein. The use of silage maize from the CS2 group had a promotional effect on the production performance of the hu sheep in the present experiment, making it the most suitable additive.
Carcass traits are key indicators of ideal livestock production in animal husbandry (Luo et al., 2019; Silva et al., 2019) and are closely related to feed composition (Francisco et al., 2020). The results showed that feeding different varieties of maize silage had a greater effect on the pre-slaughter live weight, carcass weight, and slaughter rate of hu sheep. Su et al. showed that the use of silage could increase the level of protein in the muscle (Su et al., 2020). Wang et al. showed that with the increase in the level of protein in the diet, the daily weight gain and dry matter intake increased dramatically, the feed conversion ratio significantly decreased, and the pre-slaughter live weight and carcass weight significantly increased (Wang Q. et al., 2020). It has been suggested that higher carcass weights reflect higher feed consumption, promoting muscle growth (Atsbha et al., 2021). In line with the present experiment, there was a trend of higher protein intake in the CS2 group than in the CS1 and CS3 groups, while the pre-slaughter live weight and daily weight gain in the CS2 group were significantly higher than those in the CS1 group, which is consistent with the results discussed above. It has been confirmed that muscle fiber diameter and density are closely related to muscle tenderness and tension force (Mandell et al., 1997). In this experiment, the CS2 group had the highest cooking loss, the lowest shear force, and ultimately the highest water loss rate. The results were similar to those of Mandell et al. The smaller the muscle diameter, the greater the density of muscle fibers, indicating more intramuscular fat accumulation and the more tender the meat. In contrast, when the muscle diameter is larger, the meat becomes tougher (Fahey et al., 2005). In this experiment, there were many factors that affected the shear force, as reported by Hocquette et al (Hocquette et al., 2010). Intramuscular fat can indirectly affect meat tenderness, and factors such as pH, genetic factors, and the age of the animal can also affect shear force (Hopkins et al., 2006; de Rezende et al., 2020). Some studies have shown that the increase in lamb meat tenderness may be a result of the decrease in fiber content, the decrease in connective tissue strength, and the decomposition of myofibrillar proteins and connective tissues (Jama et al., 2008; Qi et al., 2019). An et al. demonstrated that the addition of fermented feed to the diets of hu sheep was able to reduce the diameter and area of the muscle fibers, suggesting that the addition of fermented feeds can reduce the diameter of muscle fibers, making the meat more tender (An et al., 2021). Muscle tenderness was positively correlated with the amount of fermented feed added. In this experiment, it was found that the shear force of the CS2 group was lower than that of the CS1 group and significantly lower than that of the CS3 group. This may be due to the high proportion of fermented feed added in this experiment and the significant differences in nutrient composition between maize varieties, which influenced muscle characteristics and intramuscular fat deposition. The above experimental results showed that the addition of silage maize from the CS2 group improved the meat quality of the hu lambs.
Meat color, represented by parameters such as L*, a*, and b*, is a crucial visual indicator of meat quality (Devincenzi et al., 2019; Salim et al., 2022; Li et al., 2022c). It directly influences consumers’ assessment of meat quality and their purchasing intent. The L*, a*, and b* values are the key metrics used to characterize meat color. Within an appropriate range, a higher a* value indicates better meat quality and freshness. Moreover, a higher L* value of the meat color implies that the meat appears glossier and lighter in shade. Notably, the a* value has a positive correlation with meat quality, whereas the b* and L* values exhibit an inverse relationship with it (Li et al., 2016). The CS3 group significantly increased the luminosity (L*) value of the meat without having any impact on the hue (a*) and saturation (b*) levels, as well as the dripping loss. Typically, for small ruminants, the cooking loss values of their meat fall within the range of 14-41% (Ayeb et al., 2016).In the present experiment, the values obtained by observing the changes in luminosity were similar to those in the study by Ocak et al (Ocak et al., 2016). In this experiment, the L* values of the CS3 group were significantly higher than those of the CS2 and CS1 groups. This finding may be attributed to the different intramuscular fat content among the three groups, which was confirmed in the literature (Wu et al., 2021). In addition, Ledward et al. reported that the meat pH value at 45 min may also be another reason influencing the L* of meat (Ledward et al., 1986), but the exact mechanism underlying this effect needs to be explored in depth.
Variation in nutrient digestibility in ruminants depends on the chemical composition of their diet. Li et al. found that when different roughages were added to ruminant diets, only the digestibility of dry matter and organic matter differed significantly, while other aspects did not (Li et al., 2022a). Brask et al. found that the use of maize silage instead of grass clover silage had little effect on the apparent digestibility of nutrients in ruminants (Brask-Pedersen et al., 2023). Protein utilization and amino acid deposition by the ruminant organism can be reflected by nitrogen metabolism (Zeitz et al., 2018). When feed enters the rumen, proteins are initially broken down by extracellular proteases secreted by pepsin and protein-hydrolyzing bacteria, resulting in the production of peptides and amino acids (El-Shazly, 1952). These compounds are further degraded into ammonia, which is utilized by rumen microorganisms to synthesize microbial proteins, providing the animal with a high-quality nitrogen source. However, a significant portion of the ammonia enters the liver via blood circulation, where it is converted into endogenous urea (Getahun et al., 2019). This urea can then re-enter the rumen, where it is hydrolyzed back into NH3, serving as a nitrogen source for microbial protein synthesis and contributing to the animal’s energy supply (Jin et al., 2018).In this experiment, the use of three different varieties of silage maize had almost no effect on the apparent digestibility of nutrients and nitrogen utilization in hu sheep, and only the CS2 group had a trend toward higher protein intake, fecal nitrogen, deposited nitrogen, and higher digestibility of CP or apparent absorption of N than the CS1 and CS3 groups. This may be partly because our roughage source, harvesting, and production methods were consistent in terms of source, harvesting, and production methods, and t this may also suggest that different roughages have minimal influence on nutrient apparent digestibility on the apparent digestibility of nutrients. Some studies have shown that different roughages have minimal effect on the apparent digestibility of nutrients in ruminants, and since we used the same type of roughage, we hypothesize that this is the main reason for the minimal effect on the digestibility.
The rumen, a unique digestive organ in ruminants, harbors a vast and diverse population of microorganisms, including bacteria, fungi, archaea, protozoa, and others, with anaerobic bacteria being the dominant group (Pitta et al., 2016). Research has demonstrated that rumen microorganisms are closely linked to the production performance and meat quality of ruminants (Matthews et al., 2019). When feed enters the rumen, it undergoes fermentation and decomposition by these microorganisms, enabling efficient nutrient absorption by the animal. Studies have shown that feed is the primary factor influencing the composition of the rumen microbial community, which in turn affects nutrient digestion, absorption, and energy supply (Henderson et al., 2015). Moreover, the structure of the rumen microbial community is critical for enhancing feed digestion and absorption, as well as improving animal performance. In this experiment, we observed that Bacteroidota and Firmicutes were the dominant phyla in the rumen of ruminants, consistent with findings from previous studies (Maga et al., 2014; Zhou et al., 2017; Bi et al., 2018). Bacteroidota plays a significant role in the degradation of non-fibrous materials, while Firmicutes are primarily involved in the breakdown of fibrous materials (Evans et al., 2011). In this study, we observed that the use of different varieties of whole-plant maize silage exerted no significant effects on the relative abundances of Bacteroidetes and Firmicutes at the phylum level in the rumen microbiota of Hu sheep. Huang et al. found that feeding fermented feeds can promote the growth and proliferation of Bacteroides and Firmicutes in the rumen of Hu sheep, which is conducive to the degradation of fibrous materials in the diets (Huang et al., 2023). This phenomenon likely contributes to the non-significant differences in nutrient apparent digestibility observed in our trial. Furthermore, we found that feeding Hu sheep with different varieties of maize silage exerted no significant effects on either α-diversity or β-diversity of the rumen microbiota. The underlying mechanism may be attributed to the similar nutritional matrix of the feed ingredients: even though the nutritional values differed considerably across varieties, such differences did not fundamentally alter the nature of the roughage, resulting in negligible impacts on microbial α- and β-diversity.
In this study, we found that using different varieties of maize silage fed to Hu sheep had no significant effect on rumen microbial α- or β-diversity. We hypothesized that this was because all the maize used was silage, which did not change the nature of the feed although the varieties differed in their nutritional value, ultimately leading to non-significant differences in rumen microbial populations. The dominant genera in the rumen of hu sheep were Prevotella, Rikenellaceae, Christensenellaceae, Muribaculaceae, and Bacteroidales_BS11. This is not entirely consistent with previous studies, and it is hypothesized that it may be due to the differences in nutrient composition between different maize varieties (Li et al., 2022b). The highest relative abundance of Prevotella 1 in the rumen of hu sheep was consistent with previous results, but the difference between the groups was not significant, which was presumed to be due to the fact that all the maize silage used were similar feed types, and there was not much difference in the feeds between the groups, ultimately leading to the non-significant difference between the groups (Jiao et al., 2025).
4 Conclusions
In the present experiment, after undergoing 60 days of silage fermentation, the three distinct maize varieties showed notable differences in nutritional composition. The incorporation of silage maize into the animal diet led to a notable improvement in the microbial flora in the animals’ digestive tracts. Specifically, based on comprehensive evaluations of growth performance (final body weight, average daily gain, and feed-to-gain ratio), slaughter traits, meat quality parameters, apparent nutrient digestibility, and nitrogen utilization efficiency—all of which showed significant improvements—DengHai 605 (CS2) exhibited superior performance compared to other varieties under the same feeding conditions. These findings suggest that the CS2 corn variety presents potential for cultivation and practical application in practical livestock production.
Data availability statement
The data presented in the study are deposited in the SRA repository, accession number PRJNA1268884.
Ethics statement
The full name and affiliation of the ethics committee/institutional review board are: Anhui University of Science and Technology Experimental Animal Ethics Committee Affiliation: Anhui University of Science and Technology, No. 9 Donghua Road, Fucheng Town, Fengyang County, Chuzhou City, Anhui Province, China. The studies were conducted in accordance with the local legislation and institutional requirements. Written informed consent was obtained from the owners for the participation of their animals in this study.
Author contributions
LFY: Conceptualization, Writing – original draft. JH: Funding acquisition, Writing – review & editing, Methodology. WL: Writing – review & editing, Software. HZ: Writing – review & editing, Software. CC: Project administration, Writing – review & editing. LY: Writing – review & editing, Formal Analysis. XL: Writing – review & editing, Visualization.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This study was supported by the Science and Technology Innovation Team for the Utilization of Herbivorous Livestock Resources and Their Health (Grant Number: 2023AH010061) and the Creation and Application of Green, Low-Carbon and High-Efficiency Sheep Farming Technologies (Grant Number: tzy202211). We would like to acknowledge the financial support from the following projects. Additionally, special thanks are extended to Anhui University of Science and Technology for its institutional support.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Acharya I. P. and Casper D. P. (2020). Lactational response of early-lactation Holstein cows fed starch or floury corn silage. J. Dairy. Sci. 103, 5118–5130. doi: 10.3168/jds.2019-16767
Akins M. S. and Shaver R. D. (2014). Influence of corn silage hybrid type on lactation performance by Holstein dairy cows. J. Dairy. Sci. 97, 7811–7820. doi: 10.3168/jds.2014-8400
An X., Zhang S., Li T., Chen N., Wang X., Zhang B., et al. (2021). Transcriptomics analysis reveals the effect of Broussonetia papyrifera L. fermented feed on meat quality traits in fattening lamb. PeerJ 9, e11295.
Arriola Apelo S. I., Bell A. L., Estes K., Ropelewski J., de Veth M. J., and Hanigan M. D. (2014). Effects of reduced dietary protein and supplemental rumen-protected essential amino acids on the nitrogen efficiency of dairy cows. J. Dairy. Sci. 97, 5688–5699. doi: 10.3168/jds.2013-7833
Atsbha K., Gebremariam T., and Aregawi T. (2021). Slaughter performance and meat quality of Begait breed lambs fattened under different diets. Heliyon 7 (5), e06935. doi: 10.1016/j.heliyon.2021.e06935
Ayeb N., Ghrab A., Barmat A., and Khorchani T. (2016). Chemical and tissue composition of meat from carcass cuts of localgoats affected by different feeding in Tunisian arid lands. Turk. J. Vet. Anim. Sci. 40, 95–101. doi: 10.3906/vet-1412-76
Bi Y., Zeng S., Zhang R., Diao Q., and Tu Y. (2018). Effects of dietary energy levels on rumen bacterial community composition in Holstein heifers under the same forage to concentrate ratio condition. BMC Microbiol. 18, 1–11. doi: 10.1186/s12866-018-1213-9
Brask-Pedersen D. N., Lamminen M., Mogensen L., Hellwing A., Johansen M., Lund P., et al. (2023). Effect of substituting grass-clover silage with maize silage for dairy cows on nutrient digestibility, rumen metabolism, enteric methane emission and total carbon footprint. Livest. Sci. 274, 105273. doi: 10.1016/j.livsci.2023.105273
Cheli F., Campagnoli A., and Dell Orto V. (2013). Fungal populations and mycotoxins in silages: From occurrence to analysis. Anim. Feed. Sci. Technol. 183, 1–16. doi: 10.1016/j.anifeedsci.2013.01.013
Demirel R., Akdemir F., Saruhan V., Demirel D. S., Akinci C., and Aydin F. (2011). The determination of qualities in different whole-plant silages among hybrid maize cultivars. Afr. J. Agric. Res. 6, 5469–5474.
de Rezende M. P. G., Figueiredo G. C., Araujo J. I. M., Campos B. M., Moretti R., Bozzi R., et al. (2020). Growth curve, carcass traits and Kleiber ratio of Dorper crossbreed with hairless native Brazilian sheep breeds. Small. Ruminant. Res. 192, 106190. doi: 10.1016/j.smallrumres.2020.106190
Devincenzi T., Prunier A., Meteau K., and Prache S. (2019). How does barley supplementation in lambs grazing alfalfa affect meat sensory quality and authentication? Animal 13, 427–434. doi: 10.1017/S1751731118001477
El-Shazly K. (1952). Degradation of protein in the rumen of the sheep. 2. The action of rumen micro-organisms on amino-acids. Biochem. J. 51, 647.
Evans N. J., Brown J. M., Murray R. D., Getty B., Birtles R. J., Hart C. A., et al. (2011). Characterization of novel bovine gastrointestinal tract Treponema isolates and comparison with bovine digital dermatitis treponemes. Appl. Environ. Microbiol. 77, 138–147. doi: 10.1128/AEM.00993-10
Fahey A. J., Brameld J. M., Parr T., and Buttery P. J. (2005). The effect of maternal undernutrition before muscle differentiation on the muscle fiber development of the newborn lamb. J. Anim. Sci. 83, 2564–2571. doi: 10.2527/2005.83112564x
Ferraretto L. F., Shaver R. D., and Luck B. D. (2018). Silage review: Recent advances and future technologies for whole-plant and fractionated corn silage harvesting. J. Dairy. Sci. 101, 3937–3951. doi: 10.3168/jds.2017-13728
Francisco A. E., Janíček M., Dentinho T., Portugal A. P., Almeida J. M., Alves S. P., et al. (2020). Effects of alfalfa particle size and starch content in diets on feeding behaviour, intake, rumen parameters, animal performance and meat quality of growing lambs. Meat. Sci. 161, 107964. doi: 10.1016/j.meatsci.2019.107964
Getahun D., Alemneh T., Akeberegn D., Getabalew M., and Zewdie D. (2019). Urea metabolism and recycling in ruminants. BioMed. J. Sci. Tech. Res. 20, 14790–14796.
Hameleers A. (1998). The effects of the inclusion of either maize silage, fermented whole crop wheat or ure-treated whole crop wheat in a diet based on a high-quality grass silage on the performance of dairy cows. Grass. Forage. Sci. 53, 157–163. doi: 10.1046/j.1365-2494.1998.5320157.x
Henderson G., Cox F., Ganesh S., Jonker A., Young W., and Janssen P. H. (2015). Rumen microbial community composition varies with diet and host, but a core microbiome is found across a wide geographical range. Sci. Rep. 5, 14567. doi: 10.1038/srep14567
Hocquette J. F., Gondret F., Baéza E., Médale F., Jurie C., and Pethick D. W. (2010). Intramuscular fat content in meat-producing animals: development, genetic and nutritional control, and identification of putative markers. Animal 4, 303–319. doi: 10.1017/S1751731109991091
Hopkins D. L., Hegarty R. S., Walker P. J., and Pethick D. W. (2006). Relationship between animal age, intramuscular fat, cooking loss, pH, shear force and eating quality of aged meat from sheep. Aust. J. Exp. Agric. 46, 879–884. doi: 10.1071/EA05311
Huang X., Zhou L., You X., Han H., Chen X., and Huang X. (2023). Production performance and rumen bacterial community structure of Hu sheep fed fermented spent mushroom substrate from Pleurotus eryngii. Sci. Rep. 13, 8696. doi: 10.1038/s41598-023-35828-8
Jama N., Muchenje V., Chimonyo M., Strydom P. E., Dzama K., and Raats J. G. (2008). Cooking loss components of beef from Nguni, Bonsmara and Angus steers. Afr. J. Agric. Res. 3, 416–420.
Jiao J., Ma S., Jiao T., Shi S., Gao Y., Zhang X., et al. (2025). Effects of corn variety in whole-plant corn silage on dry matter intake, average daily gain and gastrointestinal tract bacteria and metabolites in Hu lambs. Front. Microbiol. 16, 1465078. doi: 10.3389/fmicb.2025.1465078
Jin D., Zhao S. G., Zheng N., Bu D. P., Beckers Y., and Wang J. Q. (2018). Urea nitrogen induces changes in rumen microbial and host metabolic profiles in dairy cows. Livest. Sci. 210, 104–110. doi: 10.1016/j.livsci.2018.02.011
Ledward D. A., Dickinson R. F., Powell V. H., and Shorthose W. R. (1986). The colour and colour stability of beef longissimus dorsi and semimembranosus muscles after effective electrical stimulation. Meat. Sci. 16, 245–265. doi: 10.1016/0309-1740(86)90037-9
Lee M. R. F., McAuliffe G. A., Tweed J. K. S., Griffith B. A., Morgan S. A., Rivero M. J., et al. (2021). Nutritional value of suckler beef from temperate pasture systems. Animal 15, 100257. doi: 10.1016/j.animal.2021.100257
Li J., Lian H., Zheng A., Zhang J., Dai P., Niu Y., et al. (2022a). Effects of different roughages on growth performance, nutrient digestibility, ruminal fermentation, and microbial community in weaned Holstein calves. Front. Vet. Sci. 9, 864320. doi: 10.3389/fvets.2022.864320
Li X., Liu X., Song P., Zhao J., Zhang J., and Zhao J. (2022c). Skeletal muscle mass, meat quality and antioxidant status in growing lambs supplemented with guanidinoacetic acid. Meat. Sci. 192, 108906. doi: 10.1016/j.meatsci.2022.108906
Li X., Wang J., Wang C., Zhang C., Li X., Tang C., et al. (2016). Effect of dietary phosphorus levels on meat quality and lipid metabolism in broiler chickens. Food Chem. 205, 289–296. doi: 10.1016/j.foodchem.2016.02.133
Li Q., Xu G., Yang D., Tu Y., Zhang J., Ma T., et al. (2024). Effects of feed ingredients with different protein-to-fat ratios on growth, slaughter performance and fat deposition of small-tail han lambs. Animals 14, 859. doi: 10.3390/ani14060859
Li J., Yan H., Chen J., Duan C., Guo Y., Liu Y., et al. (2022b). Correlation of ruminal fermentation parameters and rumen bacterial community by comparing those of the goat, sheep, and cow in vitro. FERMENTATION 8, 427. doi: 10.3390/fermentation8090427
Liu Y., Wang G., Wu H., Meng Q., Khan M. Z., and Zhou Z. (2021). Effect of hybrid type on fermentation and nutritional parameters of whole plant corn silage. Animals 11, 1587. doi: 10.3390/ani11061587
Luo J., Zeng D., Cheng L., Mao X., Yu J., Yu B., et al. (2019). Dietary β-glucan supplementation improves growth performance, carcass traits and meat quality of finishing pigs. Anim. Nutr. 5, 380–385. doi: 10.1016/j.aninu.2019.06.006
Maga E. A., Weimer B. C., and Murray J. D. (2014). Dissecting the role of milk components on gut microbiota composition. Gut. Microbes 4, 136–139. doi: 10.4161/gmic.23188
Mandell I. B., Gullett E. A., Buchanan-Smith J. G., and Campbell C. P. (1997). Effects of diet and slaughter endpoint on carcass composition and beef quality in Charolais cross steers. Can. Vet. J. 77, 403–414. doi: 10.4141/A97-006
Matthews C., Crispie F., Lewis E., Reid M., O Toole P. W., and Cotter P. D. (2019). The rumen microbiome: a crucial consideration when optimising milk and meat production and nitrogen utilisation efficiency. Gut. Microbes 10, 115–132. doi: 10.1080/19490976.2018.1505176
Nazli M. H., Halim R. A., Abdullah A. M., Hussin G., and Samsudin A. A. (2018). Potential of four corn varieties at different harvest stages for silage production in Malaysia. Asian Australas. J. Anim. Sci. 32, 224. doi: 10.5713/ajas.18.0175
Nazli M. H., Halim R. A., Abdullah A. M., Hussin G., and Samsudin A. A. (2019). Potential of four corn varieties at different harvest stages for silage production in Malaysia. Asian Australas. J. Anim. Sci. 32, 224–232. doi: 10.5713/ajas.18.0175
Ocak S., Ogun S., and Yilmaz O. (2016). Dorper sheep utilizing feed resources efficiently: a Mediterranean case study. Rev. Bras. Zootecn. 45, 489–498. doi: 10.1590/S1806-92902016000800010
Pitta D. W., Indugu N., Kumar S., Vecchiarelli B., Sinha R., Baker L. D., et al. (2016). Metagenomic assessment of the functional potential of the rumen microbiome in Holstein dairy cows. Anaerobe 38, 50–60. doi: 10.1016/j.anaerobe.2015.12.003
Qi K., Men X., Wu J., and Xu Z. (2019). Rearing pattern alters porcine myofiber type, fat deposition, associated microbial communities and functional capacity. BMC Microbiol. 19, 1–12. doi: 10.1186/s12866-019-1556-x
Ranum P., Peña Rosas J. P., and Garcia Casal M. N. (2014). Global maize production, utilization, and consumption. Ann. N. Y. Acad. Sci. 1312, 105–112. doi: 10.1111/nyas.12396
Salim A., Ferrari R. G., Monteiro M., and Mano S. B. (2022). Effect of different feeding systems on color of longissimus muscle from Bos cattle: A systematic review and meta-analysis. Meat. Sci. 192, 108871. doi: 10.1016/j.meatsci.2022.108871
Sha Y., Yu J., Xia D., Zhang Y., Liu J., and Wang H. (2024). Remodeling of intestinal bacterial community and metabolome of Dezhou donkey induced by corn silage. Sci. Rep. 14. doi: 10.1038/s41598-024-67869-y
Silva L. H., Assis D. E., Estrada M. M., Assis G. J., Zamudio G. D., Carneiro G. B., et al. (2019). Carcass and meat quality traits of Nellore young bulls and steers throughout fattening. Livest. Sci. 229, 28–36. doi: 10.1016/j.livsci.2019.09.012
Su Y., Chen G., Cai Y., Gao B., Zhi X., and Chang F. (2020). Effects ofBroussonetia papyrifera -fermented feed on the growth performance and muscle quality of Hu sheep. Can. J. Anim. Sci. 100, 771–780. doi: 10.1139/cjas-2018-0167
Terler G., Gruber L., and Knaus W. (2019a). Effects of ruminal degradability of ensiled whole crop maize varieties on feed intake and milk production of dairy cows. Animal 13, 1917–1926. doi: 10.1017/S1751731119000028
Terler G., Gruber L., and Knaus W. F. (2019b). Nutritive value of ensiled maize stover from nine different varieties harvested at three different stages of maturity. Grass. Forage. Sci. 74, 53–64. doi: 10.1111/gfs.12390
Wang Y., Wang Q., Dai C., Li J., Huang P., Li Y., et al. (2021). Effect of dietary protein level on growth, carcass characteristics, serum biochemical index, and meat quality of Hu male lambs. Small. Ruminant. Res. 194, 106294. doi: 10.1016/j.smallrumres.2020.106294
Wang Q., Wang Y., Hussain T., Dai C., Li J., Huang P., et al. (2020). Effects of dietary energy level on growth performance, blood parameters and meat quality in fattening male Hu lambs. J. Anim. Physiol. Anim. Nutr. 104, 418–430. doi: 10.1111/jpn.13278
Wang X., Xu T., Zhang X., Geng Y., Kang S., and Xu S. (2020). Effects of dietary protein levels on growth performance, carcass traits, serum metabolites, and meat composition of tibetan sheep during the cold season on the qinghai-tibetan plateau. Animals 10, 801. doi: 10.3390/ani10050801
Wu P., Fu X., Wang H., Hou M., and Shang Z. (2021). Effect of silage diet (sweet sorghum vs. whole-crop corn) and breed on growth performance, carcass traits, and meat quality of lambs. Animals 11, 3120.
Zaralis K., Nørgaard P., Helander C., Murphy M., Weisbjerg M. R., and Nadeau E. (2014). Effects of maize maturity at harvest and dietary proportion of maize silage on intake and performance of growing/finishing bulls. Livest. Sci. 168, 89–93. doi: 10.1016/j.livsci.2014.07.013
Zeitz J. O., Weber A., Most E., Windisch W., Bolduan C., Geyer J., et al. (2018). Effects of supplementing rumen-protected niacin on fiber composition and metabolism of skeletal muscle in dairy cows during early lactation. J. Dairy. Sci. 101, 8004–8020. doi: 10.3168/jds.2018-14490
Zhang H., Zhang L., Xue X., Zhang X., Wang H., Gao T., et al. (2021). Effect of feeding a diet comprised of various corn silages inclusion with peanut vine or wheat straw on performance, digestion, serum parameters and meat nutrients in finishing beef cattle. Anim. Biosci. 35 (1), 29–38 doi: 10.5713/ab.21.0088
Zhou Z., Fang L., Meng Q., Li S., Chai S., Liu S., et al. (2017). Assessment of ruminal bacterial and archaeal community structure in yak (Bos grunniens). Front. Microbiol. 8, 179. doi: 10.3389/fmicb.2017.00179
Keywords: maize silage, assortment, growth performance, digestibility, meat quality
Citation: Yan L, Hua J, Lu W, Zhao H, Chao C, Yan L and Lv X (2025) Effects of silage from different maize varieties on growth traits, meat quality, slaughter performance, apparent digestibility of nutrients, and rumen microbiota in Hu sheep. Front. Anim. Sci. 6:1640756. doi: 10.3389/fanim.2025.1640756
Received: 04 June 2025; Accepted: 26 June 2025;
Published: 30 July 2025.
Edited by:
Yafeng Huang, Anhui Agricultural University, ChinaReviewed by:
Wen Zhu, Anhui Agricultural University, ChinaYuwen Dong, University of Pennsylvania Division of Gastroenterology and Hepatology, United States
Liyang Zhang, Henan Agricultural University, China
Copyright © 2025 Yan, Hua, Lu, Zhao, Chao, Yan and Lv. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jinling Hua, aHVhamxAYWhzdHUuZWR1LmNu