 Maria Nikoloric
Maria Nikoloric Gamaliel Simanungkalit
Gamaliel Simanungkalit Malcolm Possell
Malcolm Possell Luciano Adrián González1,2
Luciano Adrián González1,2 Alex V. Chaves
Alex V. Chaves- 1Centre for Carbon Water and Food, The University of Sydney, Camden, NSW, Australia
- 2School of Life and Environmental Sciences, Faculty of Science, The University of Sydney, Sydney, NSW, Australia
Methane (CH4) emissions from ruminants significantly contribute to agricultural greenhouse gases. Strategies to reduce these emissions without compromising productivity are urgently needed. This study evaluated the effectiveness of a novel product incorporated into molasses containing additives and grain-based pellets to reduce CH4 emissions and improve productivity in beef cattle. Four products were screened in vitro for their efficacy in reducing CH4 production. The most effective (Product 3) was then incorporated into grain-based pellets and offered to steers via GreenFeed units at target intakes of 0, 100, and 200 g/head per day. Dry matter intake (DMI), CH4 production, CH4 yield (g CH4/kg DMI), CH4 intensity, water intake, and average daily gain (ADG) were measured. Product 3 reduced in vitro CH4 production by over 90% at a 6% inclusion rate (P < 0.05). In vivo study, steers supplemented with product-containing pellets tended (P=0.07) to reduce CH4 production by 10.7% compared to the control. Methane yield was reduced (P < 0.01) by 11.7% and 7.5% for the 100 and 200 g/day supplementation levels, respectively. Methane intensity tended (P=0.06) to be 16.8% lower in supplemented groups. Product intake was negatively correlated with CH4 yield and intensity and positively associated with water intake and ADG (P < 0.05). The inclusion of Product 3 in supplements beef cattle fed high forage diet significantly reduced methane yield and tended to reduce CH4 production and intensity, without negatively impacting performance. Product 3 shows potential as a viable CH4 mitigation strategy for high forage diet systems. However, further research is needed to assess the long-term effects and potential microbial adaptation associated with higher or prolonged dosing.
1 Introduction
Agriculture is responsible for 19.1% of Australia’s total greenhouse gas (GHG) emissions, with livestock methane (CH4) emissions contributing to 57% of the agriculture sector, approximately 10% of the country’s overall emissions (Department of Climate Change, 2022) The vast Australian landscape spans 769 million hectares that vary in soil and plant types as well as temperature zones. This diverse environment supports the grazing of approximately 97% of Australia’s beef cattle. Due to these environmental differences, there are variations in feed quality and availability across regions, which affect animal performance throughout the year. For instance, in certain regions, forages lack energy, crude protein, vitamins, and/or minerals (Dixon et al., 2020), leading to inefficient nutrient utilization (Leng, 1990) and, eventually, low animal performance. To address these challenges, improving nutritional management through efficient forage use has been identified as a viable strategy for reducing GHG emissions (Eugène et al., 2021). Improving the quality of the pasture is one of the most common approaches, however, it is not always easy or economically feasible to achieve. This context underscores a strategy for reducing the amount of CH4 produced per unit of feed consumed, thereby mitigating GHG emissions through dietary supplementation of grazing animals. Numerous studies have explored the supplementation of additives to ruminant diets, aiming to decrease enteric CH4 production. For instance, 3-NOP supplementation has been shown to reduce ruminant CH4 emissions ranging from 18 to 39% (Almeida et al., 2021). Asparagopsis included from 0.05%-3% of OM intake) could reduce methane emissions between 9 and 98% (Wasson and Hristov, 2022). However, the implementation of additives as a mitigation strategy is currently challenged by multiple factors including consistency of results, lack of information of responses in different production systems, cost- benefit, price of additives, and policies and legislation, amongst others. For example, inconsistencies in CH4 reduction across a range of studies, negative impacts on other fermentation parameters or DMI and challenges in integrating them into practical feeding regimes, particularly in grazing systems (Patra et al., 2017; O’Reilly et al., 2021).
Research has highlighted the potential of essential oils (EO) to modify rumen microbial fermentation and boost ruminant feed efficiency (Calsamiglia et al., 2007). Those are typically obtained from plants through steam distillation or solvent extraction techniques (Simon et al., 1990; Greathead, 2003) although the plant material can also be fed and included in diets (Ku-Vera et al., 2020). Essential oils have anti-nutritional and antimicrobial characteristics to protect plants, prevent herbivory and attract pollinators, and can also assist in modifying the rumen microbiome to reduce the production of CH4 (Patra et al., 2017; Garcia et al., 2020; Honan et al., 2021). In addition, the combinations of various EO may also increase microbial efficacy due to additive or synergistic effects between EO components (Benchaar and Greathead, 2011). Despite these promising findings, integrating EO into grazing systems remains a challenge. Lick blocks (LB) technology could deliver anti-methanogenic additives in grazing animals, which is traditionally used as a livestock strategic supplementation around the world to provide energy, protein, minerals, or therapeutic substances to improve animal performance (Makkar et al., 2007). The LB is practical to deliver, easy to manufacture and store, capable of containing feed additives with anti-methanogenic properties, and economically feasible (Imaz et al., 2019). The objective of the present study was to evaluate LB formulation to reduce CH4 emissions and enhance the performance of beef cattle fed high forage diet. An important aspect of LB is that the ingredients can be tailored for different regions, animal requirements, seasons, and production systems. Four proprietary formulations were initially evaluated in vitro, and the best candidate was tested in vivo.
2 Material and methods
2.1 Feed formulation
The products that have been used in the experiments of the present study comprised proprietary mixes of natural products and essential oils sourced from the Myrtaceae, Rutaceae, Poaceae, Amaryllidaceae, and Lamiaceae. The formulation is proprietary protected (Australian Provisional Patent Application No. 2023902943). All compounds are used in the human food chain as ingredients and are generally regarded as safe and the rate of feeding does not differ from concentrations used in food manufacturing. Free ranging cattle and sheep browse plants that contain the bioactive compounds identified in the products from the Myrtaceae, Rutaceae and Poaceae. The product is manufactured in accordance with excluded nutritional or digestive (ENDS) products and the Australian Stockfeed Regulation (Authority, 2024).
2.2 In vitro study
The in vitro trial was conducted to screen potential products for reducing CH4 emissions. A commercial LB manufacturing company (AgCoTech Holdings Pty. Ltd., Crestmead, QLD, Australia) provided four different proprietary products that were assessed at different concentrations. The products evaluated in this project contain natural plant ingredients, including vegetable oil, essential oils, condensed tannins, and tea saponins. The DM and ash results of the 4 products is presented in Table 1.
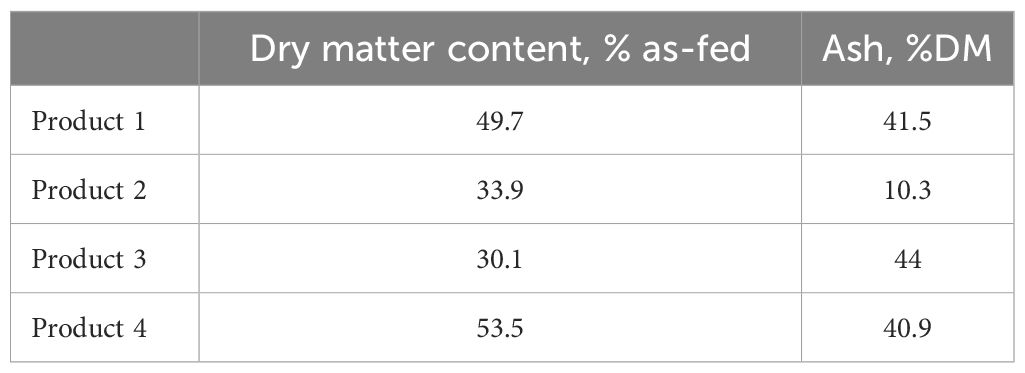
Table 1. Dry matter and ash content for products 1, 2, 3 and 4 used to manufacture molasses products containing additives assessed in vitro for their anti-methanogenic properties.
All animals were cared for in accordance with the guidelines of the University of Sydney Research Integrity Animal Ethics Committee (approval #2022/2180).
Three cannulated adult non-lactating crossbred beef cows fed with grass hay (13% CP; 60% aNDF) at maintenance level were employed for rumen fluid collection. These collections took place between two and three hours after feeding in the morning. Rumen fluid samples were taken from the dorsal, anterior ventral, medium ventral, posterior dorsal, and posterior ventral rumen areas and put into a prewarmed thermos filled to the lid to eliminate air in the headspace and immediately transported to the laboratory. The fluid was not purged with CO2 because there was no headspace after closing the lid. Rumen fluid was anaerobically transferred into a 2-L flask mixed with two parts of a phosphate buffer and kept at 39°C until in vitro incubation (Forwood et al., 2019).
The incubations design was a completely randomized design (CRD) with three incubation bottles (replicates) per treatment per run and two incubation runs. The base substrate for the incubation was a ratio of 100% grass hay (60% aNDF). Products 1, 2, and 4 were dried in a forced air oven at 55°C for 72h and grounded to 1-mm. Product 1 and 2 powders were added into F57 ANKOM filter bags (Ankom Technology, Macedon, NY USA) in 0, 5, 25, 50, 10 and 200 mg, corresponding to concentrations of 0, 1, 5, 10, 20, and 40% respectively, replacing the substrate to make up 0.5g DM in total. The product 4 powder was added in 0, 50, 100, and 200 mg, corresponding to concentrations of 0, 10, 20, and 40%, respectively, replacing the substrate to make up 0.5g DM. Product 3 was evaluated in the original liquid form and was added to the substrate bag containing 0.5g DM before the addition of rumen inoculum in 0, 0.5,1.0, and 1.5 mL corresponding to 0, 2, 4, and 6% concentrations of total volume (rumen fluid and buffer). All treatments were incubated in triplicate bottles using the batch fermentation method described in (Meale et al., 2012). Each run also contained three blank bottles with no substrate or treatments. On the incubation day, amber serum vials were warmed in the incubator at 39°C for 60 minutes and gassed with CO2 before adding 25 mL of a 2:1 mixture of buffer saturated with CO2: rumen fluid (Forwoord et al., 2019), after that process, the bottles were immediately closed with rubber stoppers and placed in an incubator set at 39°C with a rotatory shaker at 120 oscillations per min.
Batch culture bottles were removed from the incubator after 24h. To determine CH4 concentration at that timepoint, 25 mL of fermentation gas was collected from the headspace of bottle using a syringe and transferred into a 12 mL evacuated exetainer for later measurement by gas chromatography. A water displacement apparatus was used to measure the remaining gas in the bottle. Total gas production was calculated by adding the 25 mL of CH4 sample to the reading from the water displacement instrument (Fedorah and Hrudey, 1983). Then serum bottles were opened and immediately placed on ice to stop microbial fermentation, and pH was immediately tested (Activon Model 209, Gladesville, NSW, Australia). A 1.5 mL subsample of each bottle culture was placed into a 2 mL Eppendorf tube with 300 mL of metaphosphoric acid (20% w/w) and refrigerated at -20°C for subsequent volatile fatty acid (VFA) measurement following procedures described by (Wang et al., 2000; Chaves et al., 2006). ANKOM bags were removed from bottles, washed twice using a washing machine at short cycle (10 minutes), and cleaned to remove culture media, then dried for 24h at 60°C for the estimation of in vitro dry matter digestibility (IVDMD).
2.3 In vivo study
Animal ethics approval was obtained from the Animal Ethics Committee of The University of Sydney (Approval # 2293). This animal feeding trial was carried out with the product that had the greatest reduction in CH4 production from all products tested in the in vitro trials.
2.3.1 Animal feeding
For 70-d period, a completely randomized design (CRD) experiment was conducted using 45 Angus steers with an initial live weight (LW) of 233 ± 41.3kg (mean ± SD). The cohort included 36 Angus and 9 Angus × Charolais steers. The 45 steers were stratified into three LW strata (15 steers each), and within each stratum, steers were randomly assigned to three groups of 15 steers each. Upon arrival, all steers were vaccinated against BVD virus, major clostridial diseases (Ultravac® 5 in 1, Zoetis Australia Pty Ltd., Rhodes, New South Wales, Australia), and BRD (Bovilis® MH + IBR, Intervet Australia Pty Ltd., East Bendigo, Victoria, Australia). The animals were fed an oaten hay-based diet for two weeks before the treatments were introduced (as an adaptation period) and throughout the entire experiment. Each of the 3 treatments had 15 animals assigned to it: 1) control group (supplemented with pellets without product), 2) pellets containing the product for a target intake of 100g of product per head per day and, 3) pellets containing the product for a target intake of 200g of product per head per day treatment of the product incorporated into pellets. All pelleted sources were delivered through the GreenFeed® system (C-Lock, 2023).
The three treatment groups were split into two pens (4m length × 20m width) groups, with one pen occupied by control and a second pen occupied by 100g and 200g treatments. The control pen was equipped with 4 and the treatment pen had 6 electronic feeders (Intergado® Ltd, Contagem, Minas Gerais, Brazil), to deliver the hay and measure individual feed intake. Each pen had a water through with two cattle weighing scales measuring individual LW and estimating water intake from the change in weight of the animals while drinking water (Intergado® Ltd, Contagem, Minas Gerais, Brazil) and a GreenFeed® system with dual hoppers (C-Lock, 2023) to measure individual daily CH4, H2, and CO2 production rate (g/d) and O2 consumption rate (g/d), and to deliver the target dose of pellet per animal. Electronic feeders are mounted on load cells to measure individual feed intake and are equipped with a radio-frequency identification (RFID) reader to detect the presence of an animal by recording its electronic identification (EID) number. The electronic feeders automatically assigned the amount of feed consumed to the unique EID number present at the feeder when the feed weight decreased (Chizzotti et al., 2015; Oliveira et al., 2018). Samples of the offered feeds were collected weekly and dried at 70°C for 24h for the determination of DM content. Animals were taken to the yard and weighed at the start of the experiment, at 14-d intervals throughout the trial, and on the last day of the experiment. The average daily gain (ADG) was calculated by dividing weight gain (Final– Initial weight) by 56 (days of the experiment).
2.3.2 Pellet manufacturing process
Pellets were manufactured in a low-temperature process to preserve the active ingredients of the product. Rice bran (66.7%) and canola meal (33.3%) were finely ground and proportionally blended with the liquid product to achieve the designated inclusion levels (0, 6.28%, and 11.74%) without adding the settling agent. Mechanical compression was used to form pellets via a 4.4mm × 12mm die with the maximum temperature maintained below 54°C. Following this, the pellets were air-dried and cooled under ambient conditions to reduce moisture content, improve pellet durability, and minimize volatilization of the active ingredients.
2.3.3 Product supplementation
The target intake was 0, 100, and 200g of product/d into a pellet amount of 1.5 kg/head.d. One ton of pellets was proportionally mixed with 0, 67, and 133kg of the liquid product, respectively. These pellets were fed at 1.5kg (0g product), 1.6kg (100g product), and 1.7kg (200g product) per head per day.
The GreenFeed system was configured to provide the target amount of pellets to entice the animals to visit the GreenFeed multiple times per day (C-Lock, 2023). The double hopper GreenFeed in pen with the two treatments provided the two doses of product incorporated into the pellets depending on the treatment allocated to the RFID identified by the system. The pellets were dropped into a tray in a semi-enclosed hood when the RFID tag of an animal was present within the hood of the GreenFeed. The visit frequency (visits/d), number of feed drops (drops/visit), and the interval between visits were adjusted to ensure that each steer across all groups received the targeted intake as follows:
Control: visit frequency = 6 visits/d (spaced at a minimum of 120min intervals); number of cups dropped/visit = 5 (at 40 s intervals); average cup drop = 42g.
100 g per head per day: frequency = 4 visits/day (120min interval); feed drops/visit = 6 (40 s interval); average cup drop = 70g.
200 g per head per day: visit frequency = 6 visits/d (spaced at a minimum of 120min intervals); number of cups dropped/visit = 7 (40 s interval); average cup drop = 41g.
2.3.4 Quantification of CH4 emissions
Individual CH4 emissions measured by the GreenFeed system are expressed as daily CH4 production (g CH4/d). Visit duration and the number of records per individual are critical for CH4 measurement because CH4 is typically belched at 40–120 s intervals (Hammond et al., 2016). The GreenFeed operation commences when the steer places its head inside the shroud (Hammond et al., 2015). Following this, the proximity sensor in the shroud will monitor the head position of the animal during each visit, which will also be used to dismiss all measures where animals stepped out from the GreenFeed. Air is continuously drawn through the shroud and past the neck of the animal at a precisely measured rate, and the CH4 and carbon dioxide (CO2) concentrations are quantified in the exhaust air stream (Velazco et al., 2016). As the GreenFeed system provides multiple short-term breath measures, 30 measurements over a two-week period (Koning et al., 2024) with a minimum of three min durations per visit are needed to achieve a minimal variance in CH4 production rate per animal (Arthur et al., 2017). Data are logged and transmitted into the C-Lock Inc. data management system and can be downloaded through the C-Lock Inc. website interface (https://greenfeed.c-lockinc.com).
The chemical composition of the hay and treatment pellets is presented in Table 2, and the ingredients composition of pellets is shown in Table 3. Dry matter (DM) analysis was determined by oven-drying samples at 70°C for 48h, followed by hot weighing (Cunniff and Washington, 1995). Pellets were sent to be analyzed using AOAC methods of chemical analysis (Cunniff and Washington, 1995). Unless otherwise stated, all results are reported on a DM basis (Cunniff and Washington, 1995).
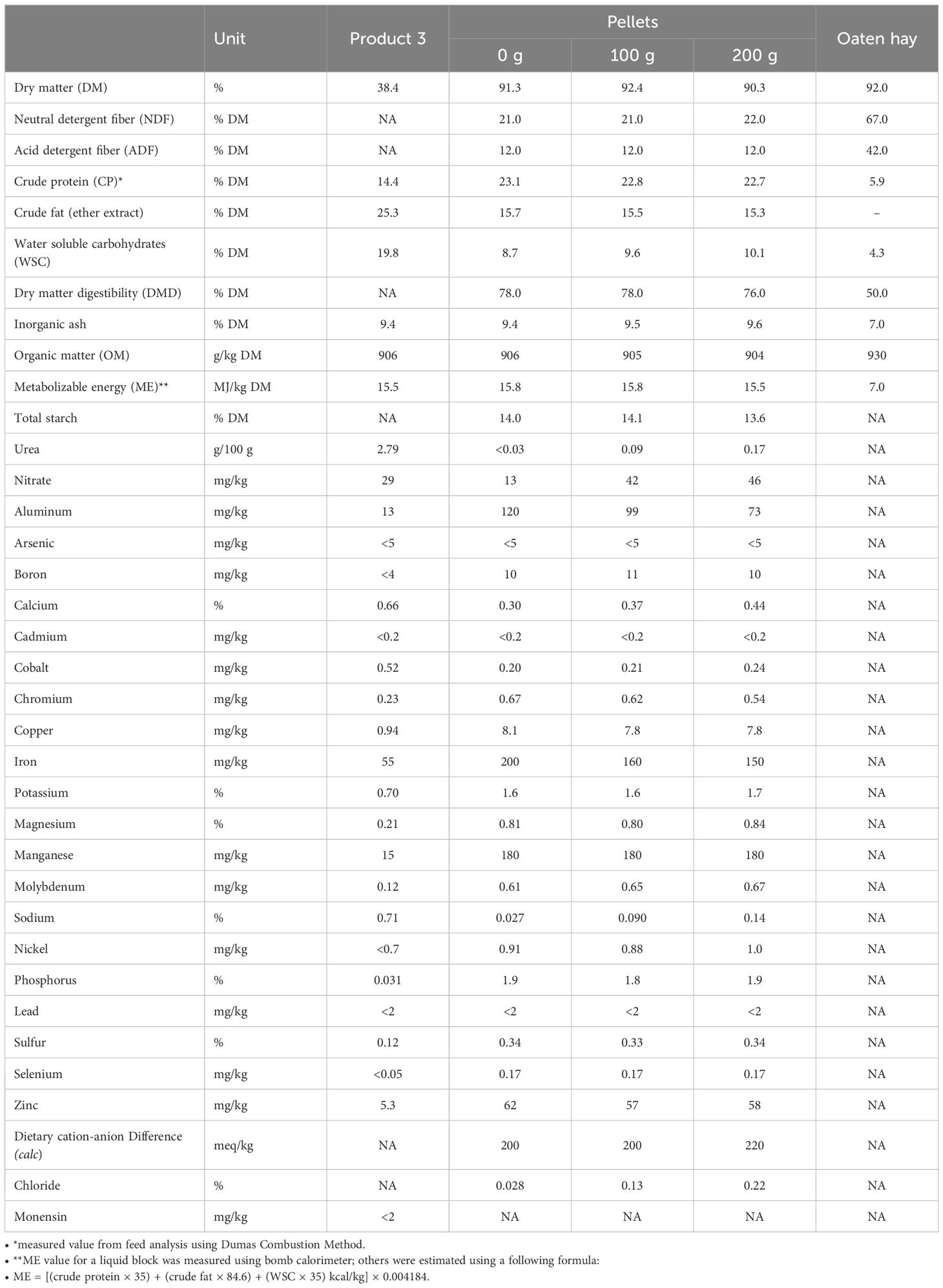
Table 2. Chemical composition of Product 3 used to manufacture molasses products containing additives treatment pellets and oaten hay fed to growing steers during the in vivo trial.
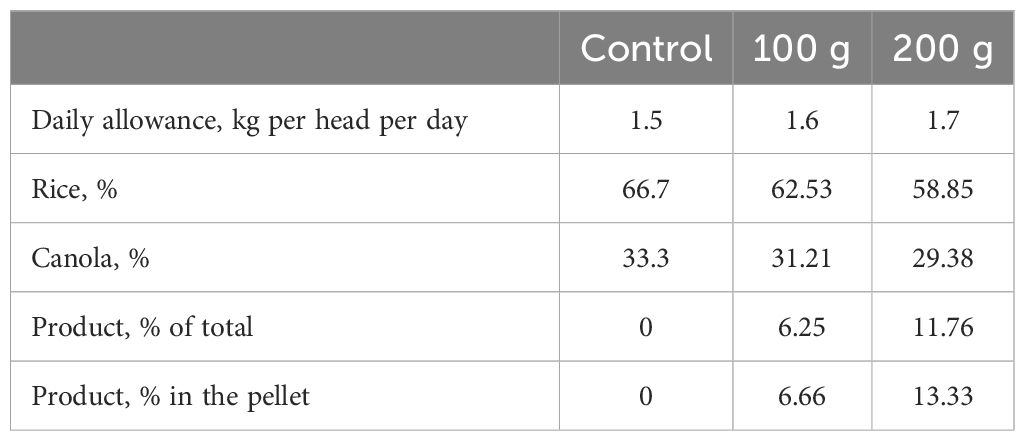
Table 3. Ingredients composition of control and treatment pellets used to manufacture molasses products containing additives fed to growing steers.
2.4 Compound odor analysis
Products compounds were identified through gas chromatography mass spectrometry (GC-MS) analysis of volatile compounds using a Gerstel Thermal Desorption Unit (TDU; Gerstel, Mülheim an der Ruhr, Germany). The samples were analyzed after drying for products 1, 2, and 4, and as received for product 4 as used for the in vitro studies. Milligram masses of each product were weighed into 200 µL micro vials, secured in glass thermal desorption liners and inserted into the TDU for analysis. When inserted in the TDU, the samples were purged with ultra-high purity helium (BOC Ltd, North Ryde, NSW, Australia) at 20°C for 1min to eliminate air from the sample and inlet. Samples were heated to 150°C by the TDU at a rate of 12°C/s with a helium flow of 75 mL/min. Thermal desorption (TD) products were carried by the helium through to a programmed temperature vaporization (PTV) inlet (CIS-4; Gerstel) installed in an Agilent 7890 GC (Agilent Technologies Pty Ltd, Mulgrave, Australia), which was used in solvent mode during the TD. The PTV inlet, containing a glass liner filled with Tenax TA, was held at 20°C during the TD using liquid CO2 (BOC Ltd) as the cryogen. After 1min of TD, the CIS-4 was heated at 12°C/s to 300°C and held at that temperature for 5 minutes while the TD products were injected into the GC at either a 1:10 or 1:500 split ratio. TD products were separated on a HP5-ms capillary column (30m × 0.25mm × 0.25 µm film thickness; Agilent) for analysis by a mass selective detector (Model 5975C; Agilent). Ultra-high purity helium was used as carrier gas (flow rate through the HP5-ms column was 2.3 mL min–1. The initial oven temperature of the GC was 35°C, held for 5 minutes, then heated at a rate of 5°C/min to 160°C, then heated at a rate of 25°C/min to 300°C and held isothermally for 4.4min. The temperature of the GC-MS interface was 280°C, the MS ion source 230°C and the quadrupole 150°C. The detector, in electron impact mode (70 eV), scanned the range of 35–300 m/z. Operation of the GC-MS was controlled by Agilent Chemstation (version E.02.01.117) and the TDU by Maestro (version 1.4.36.16; Gerstel).
The choice of 1:10 or 1:500 split ratio for a sample was determined by preliminary method testing the four products used for the in vitro study (Product 1, 2, 3 and 4), exhibiting significant quantities of VOCs. Hence, 2–5 mg of sample was used with a 1:500 split ratio. The remaining samples were injected at a 1:10 split ratio from 12–16 mg of sample.
Post-processing of the GC-MS data was performed as described in Lawson et al. (2020) except compound identification was made using the NIST14 database (Mikaia et al., 2014), and results were expressed as “present” or “not present”.
2.5 Statistical analysis
2.5.1 In vitro study
Data obtained from in vitro fermentations were analyzed as a completely randomized design using PROC MIXED of SAS software (9.4 version, SAS Institute Inc., Cary, NC), using treatment, and dose as fixed effects whereas the random effects were defined as the incubation run (experimental unit). The incubation run was employed as an error term to assess treatment effects on CH4, rumen fermentation parameters, and in vitro dry matter digestibility (IVDMD). Treatment effects were determined by comparing treatment dose replicates means with control, using least squares mean linear hypothesis test (LSMEANS/DIFF) with Dunnett adjustment, declaring significance at P ≤ 0.05 and tendency at 0.05 < P ≤ 0.10.
2.5.2 In vivo study
Data from the GreenFeed system and Intergado® electronic feeders were statistically processed using R (R Core Team, 2023). Intake data from feeders was deleted if DMI that were above 3.5% of individual LW from that period. Methane emissions, LW and feed intake data were used to calculate average values for each animal throughout the 56d of the treatment period and data from the 30-d adaptation period were not used for analysis. There were 15, 13, and 14 steers for treatment 0, 100, and 200 g/d that had more than 30 measurements for the statistical analysis. Pearson correlation analyses were performed among CH4 production and feed and product intake after the data was averaged for each across the treatment period (Charmley et al., 2015). Average daily product intake was calculated by multiplying the pellet intake measured by GreenFeed by the concentration of product in the pellet according to treatment. A CRD with ANOVA was used with the treatment group as the fixed effect and 45 animals as the experimental units. Differences between treatments across the groups were considered significant when P ≤ 0.05 and tendency at 0.05 < P ≤ 0.10.
3 Results
The results from odor analysis of the four products (Table 4) are consistent with those commonly found in blends of plant secondary metabolites such as essential oils (EO). Product 1 contained 3-carene, eucalyptol, D-limonene, and p-cymene. Product 2 contained 2,3-butanediol, 3-carene, eucalyptol, D-limonene, β-longipinene, (1aR,7R,7aS,7bR)-1,1,4,7-tetramethyl-1a,2,3,5,6,7,7a,7b-octahydro-1H-cyclopropa[e]azulene, and 2,3-dimethyl-nonadecane. Product 3 analysis found the presence of 2,3-butanediol, D-limonene, and trans-1-methyl-4-(1-methylethyl)- 2-cyclohexen-1-ol. Finally, the compounds found in product 4 were 3-carene, eucalyptol, and [1S, cis]-naphthalene,1,2,3,5,6,8a-hexahydro-4,7-dimethyl-1-[1-methylethyl].
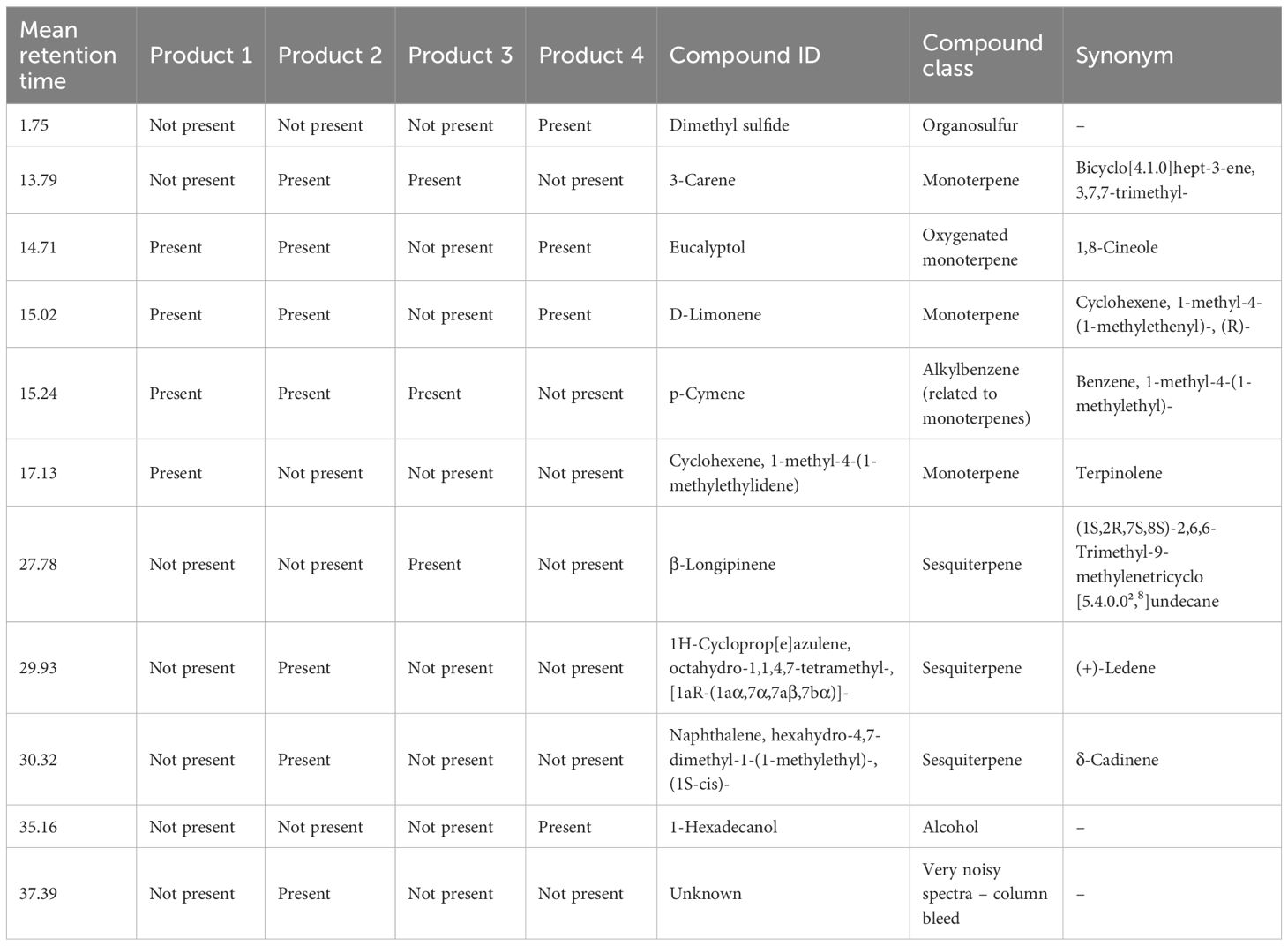
Table 4. Gas chromatography mass spectrometry (GC-MS) analysis of products 1–4 for odor compounds potentially reducing methane emissions in cattle.
3.1 In vitro study
The results of in vitro trial evaluating the four products are shown in Table 5. All variables were affected by treatment (P < 0.05). Product 1 improved IVDMD at concentrations of 5% and above (P < 0.05) and did not affect other in vitro fermentation parameters (P > 0.10). Product 2 increased IVDMD by 10% and above whereas gas production increased, and pH decreased by 40% of the product only (P < 0.05). Product 3 was the most effective in all in vitro fermentation parameters, reducing CH4 production by over 90% at 6% dose of the product compared to the control (P < 0.01), and decreasing pH at 2% to 6% concentrations. In addition, 6% concentration of product 3 also reduced VFA concentration and IVDMD (P < 0.05). The lowest dose of product 3 enhanced fermentation due to a 27% numerical increase of total VFA concentration and 55% (P <0.01) increase of gas production compared to the control treatment. A dose of 4% v/v of product 3 reduced methane production (P < 0.01) without negatively affecting fermentation as suggested by no differences in gas production and VFA against the control (P > 0.05). The molar percentage of acetate was drastically reduced with high doses of product 3 whereas propionate and butyrate were increased (P < 0.01). Product 4 improved IVDMD at all the concentrations assessed (P < 0.05), but all the other paraments were not affected (P > 0.05). Butyrate molar proportion increased with all doses of product 3 whereas branch-chained volatile fatty acids (BCVFA) increased by 6% of product 3 compared to the control treatment (P < 0.05; Table 5). None of the other treatments affected butyrate or BCVFA (P > 0.05).
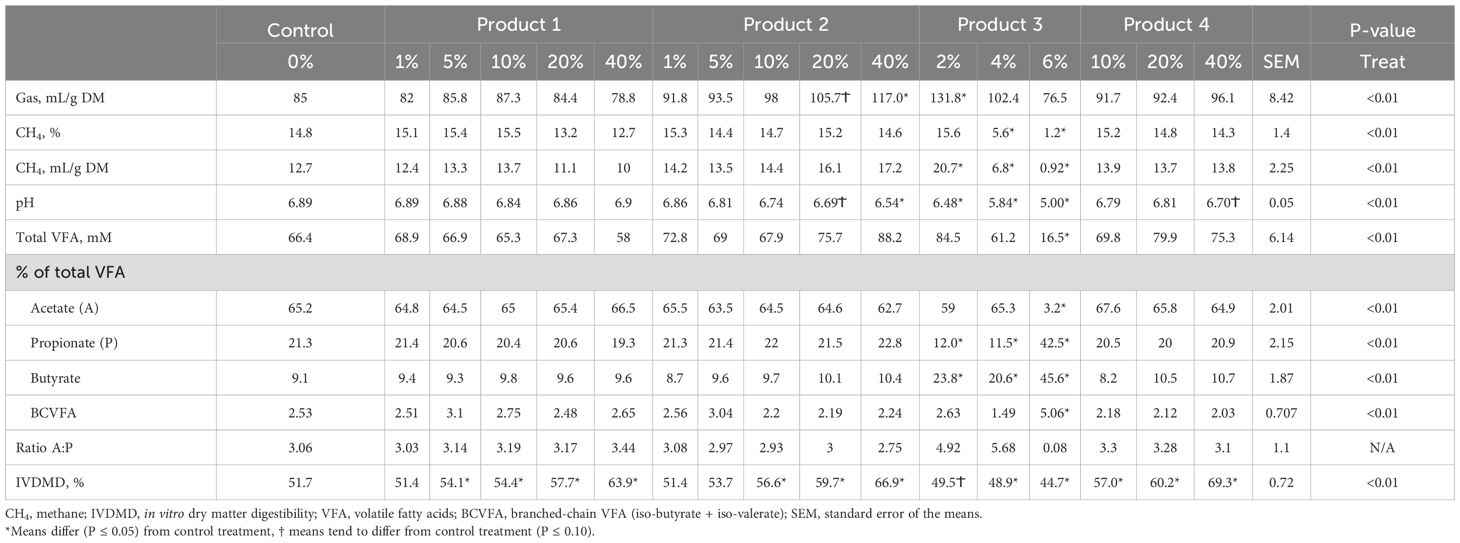
Table 5. In vitro gas production and fermentation parameters of four different candidate products for the manufacturing of lick blocks.
3.2 In vivo study
Table 6 presents the performance and CH4 emissions of growing beef steers over the 56-d experiment. Total feed, hay, and pellet intakes were similar among treatments (P ≥ 0.28, Table 6). In addition, ADG was similar for all dietary treatments (P=0.35). Product intake (g/d) was significantly different across the groups, with the greatest intake in the 200 g/d group (P < 0.01).
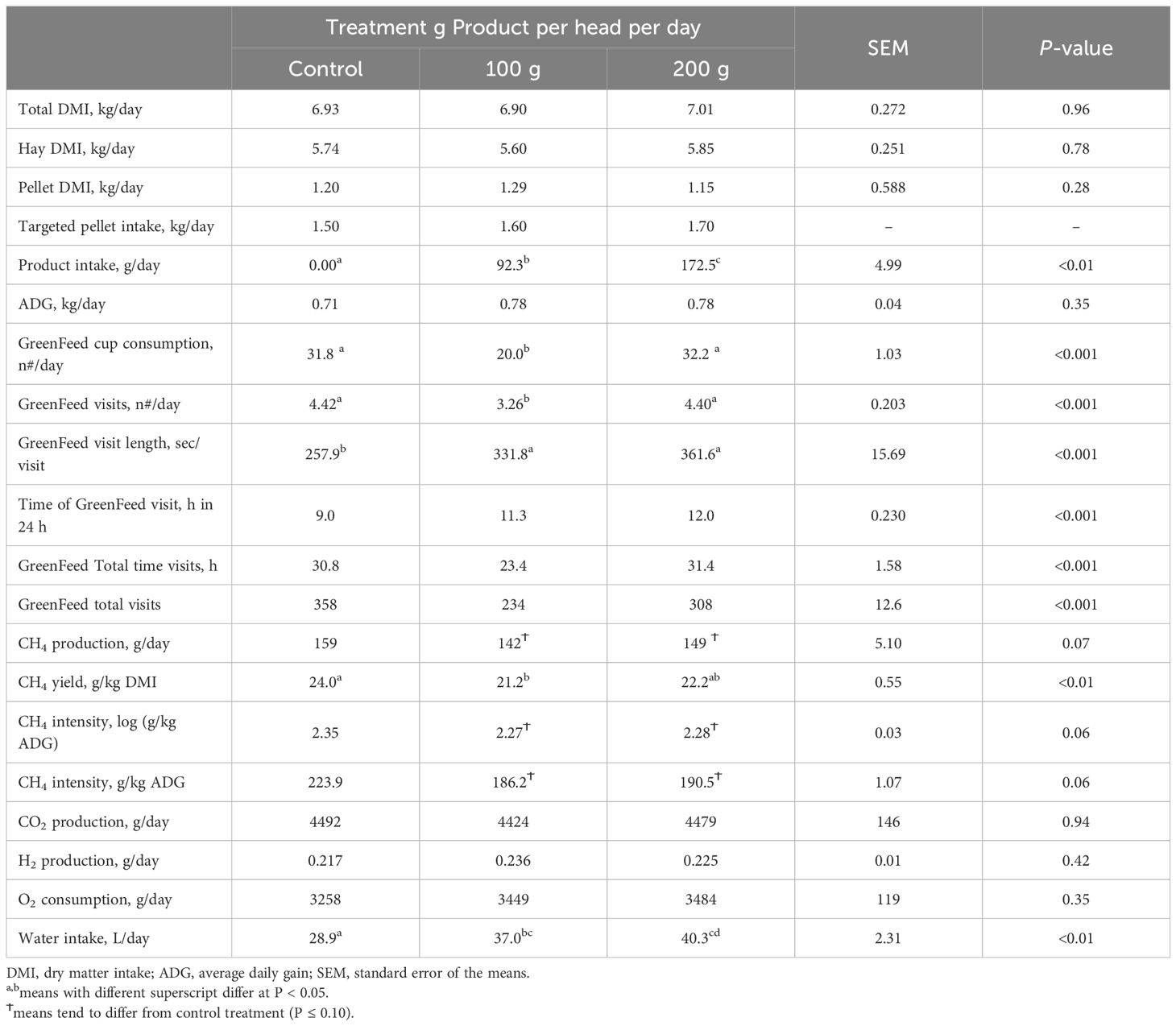
Table 6. Animal performance and greenhouse gases (GHG) emissions of growing beef steers after being offered pelleted with product 3 for 56 days of the experiment.
The number of cups delivered was the fewest for steers fed 100 g/d, whereas the GreenFeed average visit duration was shortest for steers in the control treatment (P < 0.05; Table 6). This resulted in no differences in total daily time with good data in the GreenFeed between 0 and 100 g/d (P > 0.05), but the 200 g/d had the longest daily GreenFeed data compared to the other treatments (P < 0.05; data not shown).
Results indicated a tendency for CH4 production to decrease in the 100 and 200 g/d groups compared to the control treatment, with reductions of 10.7% and 6.3%, respectively (P=0.07; Table 6). Methane yield (g CH4/kg DMI) was lower (P < 0.01) in steers fed 100 and 200 g/d treatments, showing reductions of 11.7% and 7.5% compared to the control group (Table 6). Methane intensity (g CH4/kg ADG) also tended to decrease in the 100 and 200 g/d groups by 16.8% and 14.9%, respectively (P=0.06) compared to the control. No significant differences were observed between the 100g and 200 g/d treatments for any variable (P > 0.10). Additionally, there were no differences among groups (P ≥ 0.35) in CO2 and H2 production (g/d) or oxygen consumption (g/d) as measured by the GreenFeed unit.
Daily water intake increased with the incremental dose of product supplementation, showing 28% and 39% increases on the steers treated with the 100 and 200 g/d treatments compared to the control steers, respectively (P < 0.01).
The correlation matrix among selected variables is illustrated in Figure 1. Average product intake throughout the trial was positively correlated with ADG (P < 0.05) and water intake (P< 0.001). Importantly, the average product intake was negatively correlated with CH4 intensity (P < 0.01) and CH4 yield (P < 0.05) and positive with O2 consumption (P < 0.05).
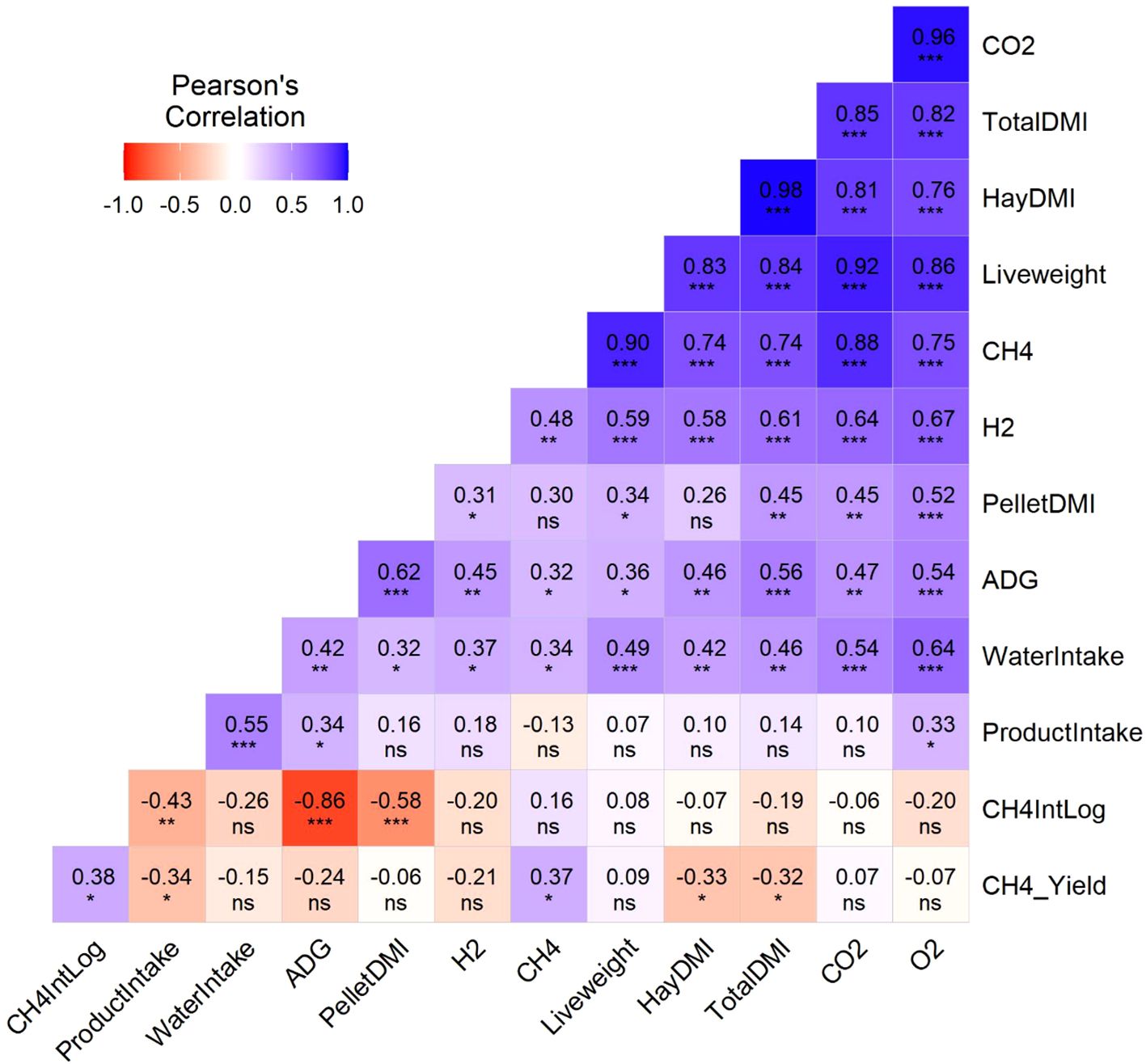
Figure 1. Correlation matrix of the performance and greenhouse gases (GHG) emissions of beef steers offered product supplement used to manufacture pellets with molasses containing additive during 56d of the experiment. ns P ≥ 0.05; *P ≤ 0.05; **P ≤ 0.01; and ***P ≤ 0.001. TotalDMI, Total Dry Matter Intake; HayDMI, Total Dry Matter Intake from Hay; PelletDMI, Dry matter Intake from Pellets; ADG, Average Daily Gain.
4 Discussion
The objective of the in vitro studies was to assess different formulations of products containing a range of essential oils. Some of these products are volatile as demonstrated in the odor analysis, and others can bind with sugar and lipid moieties. It is important to note that products 1, 2, and 4 were dried at 55°C to improve dose accuracy. In contrast, product 3 was used ‘as received’ because it could not be dried. Drying may have affected the activity or concentration of some essential oils, which prevents us from making a direct comparison between the products. However, the objective of the in vitro trial was to select the product with the greatest effect on CH4 production. It is important to note that the analysis of volatile compounds of each product was done after drying products 1, 2, and 4 as used in vitro. In addition, both pellet and lick block manufacturing (where the products are added) also involve increased temperatures. Furthermore, the ash content values obtained in this study are higher than typically expected for organic feed products. The exceptionally high ash content, particularly in Product 3 (44% DM), raises concerns about the composition of the material. While Mix 3 is described as a liquid-solid admixture, the ash content suggests a significant presence of inorganic material, potentially making it similar in composition to a clay slurry or mineral-rich mixture. Given this, further investigation into the mineral composition of the products may be warranted to confirm the nature of the ash content and its potential impact on fermentation dynamics.
Product 3 emerged as the standout performer in reducing CH4 emissions in vitro, demonstrating a remarkable efficacy across all evaluated in vitro parameters. Notably, at a 6% dosage of product 3, it reduced CH4 production by over 90% in comparison to the control group, highlighting its potent effect albeit the significant reduction in IVDMD. This product was then selected for the in vivo trial, where results indicated that steers treated with the product tended to have up to 10.7% reduction in CH4 production. Moreover, CH4 yield from steers treated with the product were up to 11.7% lower than those without the product in the pellet. Furthermore, CH4 intensity tended to be 16.8% lower for the animals fed Product 3 compared with the control, reflecting the efficiency of the product in reducing emissions.
Results from the volatile analysis showed compounds present in each product that are consistent with those commonly found in blends of plant secondary metabolites such as EO. Dimethyl sulfide was one of the compounds found to be present in Product 4, belonging to the organosulfur chemical group, which are common constituents of many plant species including those used in the present study and garlic oil (Honan et al., 2021). Organosulfur compounds have been suggested to reduce enteric CH4 due to their toxic effects on methanogenic archaea (Soliva et al., 2011). In a study by Soliva et al. (2011), garlic oil was added to a mix of hay and soybean meal concentrate in vitro at a dose of 300 mg/L of incubation liquid and reported a 91% reduction in CH4 production. Pawar et al. (2014) also screened garlic oil in vitro in a concentrate diet and reported reductions in CH4 of up to 71.5% when included in a dose of 30 µL/mL of liquid incubation, though there was also a significant negative impact on overall fermentation noted. Patra and Yu (2015) used garlic oil in a forage-based diet in vitro and also reported a reduction in CH4 but only by 29%, indicating that dimethyl sulfide may be more effective for use in concentrate diets compared to forage-based diets as used in the present study. However, Product 4 did not have a significant reduction in CH4 and IVDMD had a significant increase in the present study. These differences may be due to low concentration of the active ingredients in the present compared to previous studies, but this cannot be confirmed because dimethyl sulfide concentration was not measured, and it is also unclear whether this or other compounds present in garlic oil may be responsible for a reduction of CH4 production.
Compounds belonging to the monoterpene and oxygenated monoterpene chemical groups were identified in all four products screened in the present study. As classes of secondary plant metabolites commonly found in EO, some oxygenated monoterpenes have been found to inhibit the metabolism and growth of microbes in the rumen (Garcia et al., 2020). Eucalyptol, an oxygenated monoterpene was identified in product 1, 2, and 4. Results from in vitro analysis of those products did not indicate any significant change in CH4, pH and VFA in the present study which aligns with the results found in the literature. Eucalyptol oil was screened in vitro in a study by Colombini et al. (2021). It was added to a total mixed ration substrate in a dose of 200 mg/L of inoculum and incubated with measurements taken at 24 and 48h time points. The study found that there was no observable impact on CH4 production at either time point, or on CH4 concentration after 24h. However, there was a 7% reduction in CH4 concentration after 48h (Colombini et al., 2021). The latter study did not observe any impact on pH or total volatile fatty acids (VFA) production, despite the changes in the relative abundance of rumen microbes. In particular, the Firmicutes-to-Bacteroidetes ratio was higher when eucalyptol was added compared to the control, a trend that has been previously associated with reduced feed conversion (Jami et al., 2014; Colombini et al., 2021). Colombini et al. (2021) did not measure IVDMD, but the reduced fermentation may have been a result of impaired microbial proportions and explained the reduction of CH4 at a later time point.
Comparatively, the hydrocarbon monoterpenes, such as 3-carene or D-limonene identified in products 1, 2, 3, and 4, have been reported to increase rumen microbial activity in some cases (Kahvand and Malecky, 2018), which aligns with results from our experiments assessing product 3. For instance, the in vitro evaluation of product 3 showed a significant reduction of CH4 at 4 and 6% v/v concentration, but no significant changes at 2% concentration. This could be attributed to an insufficient dose. Furthermore, feeding product 3 in vivo confirmed that CH4 yield decreased and CH4 production and intensity tended to decrease with product 3. In contrast, no change was observed on CH4 production when product 2 was assessed in vitro, but there was an increase in gas production and IVDMD. It has been suggested that monoterpenes may be broken down and utilized as a source of carbon by rumen microbes (Benchaar et al., 2008; Garcia et al., 2020). This may explain the significant increase in IVDMD with increasing dose of products 1, 2, and 4 in the present study, all of which contained monoterpenes. Nonetheless, Product 3 also contained monoterpenes but showed a significant decrease in IVDMD especially when a greater dose was added. This decrease could be attributed to the reduction in pH which will be further explained in this discussion.
Sesquiterpene compounds were also found in Products 2 and 3. (Nunes et al., 2023) evaluated the effects of three EO from exotic plants at different levels, with Hedychium gardnerianum containing essential oils from the sesquiterpene family. Results from that study indicated that both gas and CH4 production decreased significantly compared to the control treatment after incubation at medium and high but not at the lowest concentration. Similarly, a significant reduction in CH4 production was observed at medium and high concentration of product 3, but gas production was not affected at medium and high concentrations and increased at the lowest concentration of product compared to the control treatment. Product 2 increased gas production by 28.9 and 37% at 20 and 40% of the substrate, although CH4% was not affected. The variation in outcomes observed from differing treatments and concentrations indicates an interaction between the type of EO and its quantity, which affects both total gas and CH4 production. The presence of active compounds within each EO, and their combination, can lead to distinct synergistic, antagonistic, or additive impacts on the results (Burt, 2004). Also, possible differences exist in rates at which microbial populations degrade these secondary metabolites (Broudiscou et al., 2007). Given these factors, additional studies are required to ascertain the degree to which the essential oils used in the present project can be degraded or metabolized within the digestive system of ruminants.
Previous research that has demonstrated successful CH4 mitigation from EO supplementation suggests that a trade-off exists between CH4 inhibition and overall fermentation (Durmic et al., 2014; Jahani-Azizabadi et al., 2019; Parra et al., 2023).
Although modifications in VFA profiles are often deemed beneficial, EO can sometimes lead to undesirable shifts in individual VFA proportions. For instance, Castillejos et al. (2006) observed that eugenol at 500 mg/L diminished propionate proportion without altering total VFA concentration. Moreover, Cardozo et al. (2005) demonstrated that the impact of EO on VFA profiles is contingent upon pH. The 6% concentration of Product 3 decreased the medium’s pH to 5, seemingly at odds with the lower VFA concentration. These pH values are of particular concern as pH < 5.5 is considered within the threshold for subacute ruminal acidosis, and pH < 5.0 is considered acute ruminal acidosis (Penner et al., 2007). However, this can be attributed to the product’s inherent acidity which was measured at 4.82 (data not shown), indicating that the low ruminal pH may be due to the increasing concentration of Product 3 instead of a result of fermentation products. It is important to point out that the dose rate used in the present in vitro study may not yield a significant pH change in vivo because the animal produces bicarbonate buffer and the dose is low compared to total rumen turnover and volumes. In contrast, 4% of product 3 reduced CH4% by 62% without negatively impacting fermentation, and medium pH was 5.84. This underscores the significance of product 3 dose optimization and suggests that as an optimum concentration.
In the in vivo study, water intake increased with product 3 supplementation, potentially due to its higher mineral content, prompting animals to drink more to sustain high pellet intake. This phenomenon has been well-documented: cattle consuming salt-enhanced supplements or saline water show increased water intake and enhanced renal diuresis (e.g., heifers consuming salty water exhibited water intake rising from 21.8 to 41.5 kg/day with increasing salt concentrations) (Visscher et al., 2013). Additionally, studies investigating the broader impacts of salt supplementation to low-quality forage diets found that increasing salt levels can alter forage intake, rumen fill, liquid kinetics, and fermentation patterns, further reflecting the physiological consequences of elevated salt intake (White et al., 2024). Despite this rise in water intake, no significant changes were noted in ADG or feed intake. This observation aligns with findings from various in vivo studies with EO fed to beef cattle where no effect was demonstrated on feed intake (Beauchemin and McGinn, 2006; Yang et al., 2010), suggesting no changes in rumen methanogenesis or adaptation of the microbes to EO. Cardozo et al. (2006) found that while a high dose of EO (a blend of 600 mg/day of cinnamaldehyde and 300 mg/d of eugenol) did not affect DMI, a lower dose (180 mg/d of cinnamaldehyde and 90 mg/d of eugenol) decreased DMI in beef heifers fed a high-concentrate diet. Similarly, Yang et al. (2010) reported that eugenol supplementation in growing heifers did not influence DMI. Conversely, Benchaar et al. (2006) observed that supplementation of 2–4 g/day of an EO mixture containing eugenol increased DMI in a digestion study, but this effect was not replicated in a growth study with beef cattle on a high-forage diet. Nonetheless, product 3 supplementation showed a trend for a reduction in CH4 production and intensity, especially noticeable in 100 g/head.d and a significant reduction in CH4 yield compared to the control group. Additionally, a negative correlation exists between the log of CH4 intensity and ADG. This correlation may stem from the fact that Product 3 could be enhancing feed conversion, resulting in better weight gain without a proportional increase in methane production. Therefore, while the cattle gain more weight, the methane per unit of weight gain is reduced. These results indicate the potential for product 3 to contribute to lower CH4 emissions from beef steers fed high-forage diets. The specificity of Product 3 in reducing CH4 without affecting CO2 and H2 production further supports its targeted action, as corroborated in the literature on plant-based compounds, including EO, saponins, and tannins (Francis et al., 2002; Benchaar and Greathead, 2011; Wang et al., 2012; Honan et al., 2021).
5 Conclusion
This study evaluated four novel products in vitro, with Product 3 showing the strongest methane-reducing effect. At 6% (v/v), CH4 production was reduced by over 90%, although digestibility was also compromised. At a lower inclusion level (4%), methane emissions were reduced by 62% without negatively affecting digestibility, emphasizing the importance of dose optimization.
In vivo, incorporation of Product 3 into beef steer diets modestly reduced methane yield (−11.7%) without affecting feed intake or growth performance. While these results are promising, they should be interpreted cautiously given the short duration and scale of the trial.
Further research at higher dosages, over extended periods, and with incorporation into lick blocks is needed to clarify the balance between methane abatement, digestibility, and overall animal performance.
Data availability statement
The raw data supporting the conclusions of this article are not publicly available because they are subject to confidentiality and contractual obligations. In accordance with the governing agreement, such data cannot be disclosed or distributed without the prior written consent of both Meat & Livestock Australia (MLA) and Agcotech. Requests to access the datasets should be directed to the corresponding author/s.
Ethics statement
The experimental protocols and use of animals were approved by the Animal Ethics Committee of the University of Sydney for both the in vitro (2022/2180) and in vivo (ARA No. 2023/2293) trials. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
MN: Data curation, Formal analysis, Investigation, Methodology, Writing – original draft. GS: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Project administration, Supervision, Validation, Writing – review & editing. MP: Formal Analysis, Investigation, Methodology, Writing – review & editing, Data curation. LG: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Supervision, Writing – review & editing. AC: Formal analysis, Investigation, Methodology, Supervision, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This paper and the research behind it would not have been possible without the financial support from grant PPSH.1379 from Meat & Livestock Australia Donor Company, Commonwealth Government of Australia, The University of Sydney, and AgCoTech. The funders were not involved in the study design, collection, analysis, interpretation of data, the writing of this article, or the decision to submit it for publication.
Acknowledgments
We would like to thank Paul and Jeanette the managers at Pye Farm where the pivotal in vivo study took place. We are equally indebted to Milad, Mulisa and Alessio, whose commitment and enthusiasm help during the experiment set up played a crucial role in fruition of this study. Without their help and passion, this study would not have been possible. We also thank AgCoTech for providing the novel products evaluated in the present paper, inclusive of Products 1–4 as well as molasses products. Products 1–4 are protected under Australian Provisional Patent Application No. 2023902943.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Almeida A. K., Hegarty R. S., and Cowie A. (2021). Meta-analysis quantifying the potential of dietary additives and rumen modifiers for methane mitigation in ruminant production systems. Anim. Nutr. 7, 1219–1230. doi: 10.1016/j.aninu.2021.09.005
Arthur P., Barchia I., Weber C., BirdGardiner T., Donoghue K., Herd R., et al. (2017). Optimizing test procedures for estimating daily methane and carbon dioxide emissions in cattle using short-term breath measures. J. Anim. Sci. 95, 645–656. doi: 10.2527/jas.2016.0700
Authority, A.P.V.M.A (2024). Stockfeed and pet food regulation (Australian Government: Australian Pesticides and Veterinary Medicines Authority).
Beauchemin K. and McGinn S. (2006). Methane emissions from beef cattle: Effects of fumaric acid, essential oil, and canola oil. J. Anim. Sci. 84, 1489–1496. doi: 10.2527/2006.8461489x
Benchaar C. and Greathead H. (2011). Essential oils and opportunities to mitigate enteric methane emissions from ruminants. Anim. Feed Sci. Technol. 166–167, 338–355. doi: 10.1016/j.anifeedsci.2011.04.024
Benchaar C., Calsamiglia S., Chaves A. V., Fraser G., Colombatto D., McAllister T. A., et al. (2008). A review of plant-derived essential oils in ruminant nutrition and production. Anim. Feed Sci. Technol. 145, 209–228. doi: 10.1016/j.anifeedsci.2007.04.014
Benchaar C., Duynisveld J., and Charmley E. (2006). Effects of monensin and increasing dose levels of a mixture of essential oil compounds on intake, digestion and growth performance of beef cattle. Can. J. Anim. Sci. 86 (1), 91–96. doi: 10.4141/A05-027
Broudiscou L. P., Cornu A., and Rouzeau A. (2007). In vitro degradation of 10 mono- and sesquiterpenes of plant origin by caprine rumen micro-organisms. J. Sci. Food Agric. 87, 1653–1658. doi: 10.1002/jsfa.2863
Burt S. (2004). Essential oils: Their antibacterial properties and potential applications in foods—a review. Int. J. Food Microbiol. 94, 223–253. doi: 10.1016/j.ijfoodmicro.2004.03.022
Calsamiglia S., Busquet M., Cardozo P., Castillejos L., and Ferret A. (2007). Invited review: essential oils as modifiers of rumen microbial fermentation. J. Dairy Sci. 90, 2580–2595. doi: 10.3168/jds.2006-644
Cardozo P. W., Calsamiglia S., Ferret A., and Kamel C. (2005). Screening for the effects of natural plant extracts at different pH on in vitro rumen microbial fermentation of a high-concentrate diet for beef cattle. J. Anim. Sci. 83 (11), 2572–2579. doi: 10.2527/2005.83112572x
Cardozo P., Calsamiglia S., Ferret A., and Kamel C. (2006). Effects of alfalfa extract, anise, capsicum, and a mixture of cinnamaldehyde and eugenol on ruminal fermentation and protein degradation in beef heifers fed a high-concentrate diet. J. Anim. Sci. 84, 2801–2808. doi: 10.2527/jas.2005-593
Castillejos L., Calsamiglia S., and Ferret A. (2006). Effect of essential oil active compounds on rumen microbial fermentation and nutrient flow in in vitro systems. J. Dairy Sci. 89 (7), 2649–2658. doi: 10.3168/jds.s0022-0302(06)72341-4
Charmley E., Williams S. R. O., Moate P. J., Hegarty R. S., Herd R. M., Oddy V. H., et al. (2015). A universal equation to predict methane production of forage-fed cattle in Australia. Anim. Prod. Sci. 56, 169–180. doi: 10.1071/AN15365
Chaves A. V., Thompson L. C., Iwaasa A. D., Scott S. L., Olson M. E., Benchaar C., et al. (2006). Effect of pasture type (alfalfa vs. grass) on methane and carbon dioxide production by yearling beef heifers. Can. J. Anim. Sci. 86, 409–418. doi: 10.4141/A05-081
Chizzotti M., MaChado F., Valente E., Pereira L., Campos M., Tomich T., et al. (2015). Validation of a system for monitoring individual feeding behavior and individual feed intake in dairy cattle. J. Dairy Sci. 98, 3438–3442. doi: 10.3168/jds.2014-8760
C-Lock. (2023). GreenFeed system: User manual. (USA: C-Lock Inc.). Available online at: https://www.c-lockinc.com.
Colombini S., Graziosi A. R., Parma P., Iriti M., Vitalini S., Sarnataro C., et al. (2021). Evaluation of dietary addition of 2 essential oils from Achillea moschata, or their components on in vitro ruminal fermentation and microbial community composition. Anim. Nutr. 7, 224–231. doi: 10.1016/j.aninu.2020.11.001
Cunniff P. and Washington D. (1995). AOAC, Official methods of analysis of AOAC international (Arlington, VA: AOAC).
Department of Climate Change, E., The Environment and Water (2022). Quarterly Update of Australia’s National Greenhouse Gas Inventory: September 2022 (Canberra, Australia: Department of Climate Change, Energy, the Environment and Water).
Dixon R., Fletcher M., Anderson S., Kidd L., Benvenutti M., Mayer B., et al. (2020). Improved Management of Cattle Phosphorus Status through Applied Physiology (North Sydney, NSW Australia: Meat and Livestock Australia).
Durmic Z., Moate P. J., Eckard R., Revell D. K., Williams R., and Vercoe P. E. (2014). In vitro screening of selected feed additives, plant essential oils and plant extracts for rumen methane mitigation. J. Sci. Food Agric. 94, 1191–1196. doi: 10.1002/jsfa.6396
Eugène M., Klumpp K., and Sauvant D. (2021). Methane mitigating options with forages fed to ruminants. Grass Forage Sci. 76, 196–204. doi: 10.1111/gfs.12540
Fedorah P. M. and Hrudey S. E. (1983). A simple apparatus for measuring gas production by methanogenic cultures in serum bottles. Environ. Technol. 4, 425–432. doi: 10.1080/09593338309384228
Forwood D. L., Hooker K., Caro E., Huo Y., Holman D. B., Meale S. J., et al. (2019). Crop sorghum ensiled with unsalable vegetables increases silage microbial diversity. Front. Microbiol. 10, 2599. doi: 10.3389/fmicb.2019.02599
Francis G., Kerem Z., Makkar H. P., and Becker K. (2002). The biological action of saponins in animal systems: a review. Br. J. Nutr. 88, 587–605. doi: 10.1079/bjn2002725
Garcia F., Colombatto D., Brunetti M. A., Martínez M. J., Moreno M. V., Scorcione Turcato M. C., et al. (2020). The reduction of methane production in the in vitro ruminal fermentation. Animals 10, 786. doi: 10.3390/ani10050786
Greathead H. (2003). Plants and plant extracts for improving animal productivity. Proc. Nutr. Soc 62, 279–290. doi: 10.1079/pns2002197
Hammond K., Humphries D., Crompton L., and Green C. (2015). Methane emissions from cattle: Estimates from short-term measurements using a GreenFeed system. Anim. Feed Sci. Technol. 203, 41–52. doi: 10.1016/j.anifeedsci.2015.02.003
Hammond K., Waghorn G., and Hegarty R. (2016). The GreenFeed system for measurement of enteric methane emission from cattle. Anim. Prod. Sci. 56, 181–189. doi: 10.1071/AN15316
Honan M., Feng X., Tricarico J., and Kebreab E. (2021). Feed additives as a strategic approach to reduce enteric methane production in cattle: modes of action, effectiveness and safety. Anim. Prod. Sci. 61, 1153–1163. doi: 10.1071/AN20295
Imaz J. A., García S., and González L. A. (2019). Real-time monitoring of self-fed supplement intake, feeding behaviour, and growth rate as affected by forage quantity and quality of rotationally grazed beef cattle. Animals 9, 1129. doi: 10.3390/ani9121129
Jahani-Azizabadi H., Durmic Z., Vadhanabhuti J., and Vercoe P. (2019). Effect of some Australian native shrubs essential oils on in vitro rumen microbial fermentation of a high-concentrate diet. J. Anim. Plant Sci. 29, 8–15. doi: 10.36899/JAPS.2019.1.003
Jami E., White B. A., and Mizrahi I. (2014). Potential role of the bovine rumen microbiome in modulating milk composition and feed efficiency. PLoS One 9, e85423. doi: 10.1371/journal.pone.0085423
Kahvand M. and Malecky M. (2018). Dose-response effects of sage (Salvia officinalis) and yarrow (A. millefolium) essential oils on rumen fermentation in vitro. Ann. Anim. Sci. 18, 125–142. doi: 10.2478/aoas-2018-0009
Koning L., van Gastelen S., Burgers E. E. A., Klootwijk C. W., Bannink A., and Dijkstra J. (2024). Minimum number of spot measurements required for quantifying enteric methane production in dairy cattle. In: Why grasslands? Proceedings of the 30th General Meeting of the European Grassland Federation. Klootwijk C. W., Bruinenberg M. H., Cougnon M., Hoekstra N. J., Ripoll-Bosch R., Schelfhout S., et al (eds.), (Leeuwarden, Netherlands: Grassland Science in Europe) 29, 465–467.
Ku-Vera J.C., Jiménez-Ocampo R., Valencia-Salazar S.S., Montoya-Flores M.D., Molina-Botero I.C., Arango J., et al. (2020). Role of secondary plant metabolites on enteric methane mitigation in ruminants. Front. Vet. Sci. 7, 584. doi: 10.3389/fvets.2020.00584
Lawson C. A., Seymour J. R., Possell M., Suggett D. J., and Raina J.-B. (2020). The volatilomes of Symbiodiniaceae-associated bacteria are influenced by chemicals derived from their algal partner. Front. Mar. Sci. 7, 106. doi: 10.3389/fmars.2020.00106
Leng R. (1990). Factors affecting the utilization of ‘poor-quality’ forages by ruminants particularly under tropical conditions. Nutr. Res. Rev. 3, 277–305. doi: 10.1079/NRR19900018
Makkar H. P., Sánchez M., and Speedy A. W. (2007). Feed supplementation blocks: urea-molasses multinutrient blocks: simple and effective feed supplement technology for ruminant agriculture (Rome, Italy: Food & Agriculture Org).Available online at: https://www.fao.org/4/a0242e/a0242e00.htm.
Meale S. J., Chaves A. V., Baah J., and McAllister T. A. (2012). Methane production of different forages in in vitro ruminal fermentation. AsianAustralas. J. Anim. Sci. 25, 86–91. doi: 10.5713/ajas.2011.11249
Mikaia A., White E. V., Zaikin V., Zhu D., Sparkman O. D., Neta P., et al. (2014). NIST standard reference database 1A: NIST/EPA/NIH mass spectral library (NIST 14) and NIST mass spectral search program (version 2.2). National Institute of Standards and Technology.
Nunes H., Dias C. M., and Borba A. (2023). Bioprospecting essential oils of exotic species as potential mitigations of ruminant enteric methanogenesis. Heliyon 9, e12786. doi: 10.1016/j.heliyon.2022.e12786
O’Reilly G. C., Huo Y., Meale S. J., and Chaves A. V. (2021). Dose response of biochar and wood vinegar on in vitro batch culture ruminal fermentation using contrasting feed substrates. Transl. Anim. Sci. 5, txab107. doi: 10.1093/tas/txab107
Oliveira B. Jr., Ribas M., MaChado F., Lima J., Cavalcanti L., and Chizzotti M. (2018). Validation of a system for monitoring individual feeding and drinking behavior and intake in young cattle. Animal 12, 634–639. doi: 10.1017/S1751731117002002
Parra M. C., Forwood D. L., Chaves A. V., and Meale S. J. (2023). In vitro screening of anti-methanogenic additives for use in Australian grazing systems. Front. Anim. Sci. 4. doi: 10.3389/fanim.2023.1123532
Patra A. K. and Yu Z. (2015). Effects of adaptation of in vitro rumen culture to garlic oil, nitrate, and saponin and their combinations on methanogenesis, fermentation, and abundances and diversity of microbial populations. J. Appl. Microbiol. 6, 1434. doi: 10.3389/fmicb.2015.01434
Patra A., Park T., Kim M., and Yu Z. (2017). Rumen methanogens and mitigation of methane emission by anti-methanogenic compounds and substances. J. Anim. Sci. Biotechnol. 8, 1–18. doi: 10.1186/s40104-017-0145-9
Pawar M. M., Kamra D. N., Agarwal N., and Chaudhary L. C. (2014). Effects of essential oils on in vitro methanogenesis and feed fermentation with buffalo rumen liquor. Agric. Res. 3 (1), 67–74. doi: 10.1007/s40003-014-0092-z
Penner G., Beauchemin K., and Mutsvangwa T. (2007). Severity of ruminal acidosis in primiparous Holstein cows during the periparturient period. J. Dairy Sci. 90, 365–375. doi: 10.3168/jds.S0022-0302(07)72638-3
R Core Team (2023). R: A Language and Environment for Statistical Computing. (Vienna, Austria: R Foundation for Statistical Computing). Available online at: https://www.R-project.org/.
Simon J. E., Quinn J., and Murray R. G. (1990). Basil: a source of essential oils. Adv. New Crops 1, 484–489. doi: 10.1007/BF02907824
Soliva C. R., Amelchanka S. L., Duval S. M., and Kreuzer M. (2011). Ruminal methane inhibition potential of various pure compounds in comparison with garlic oil as determined with a rumen simulation technique (Rusitec). Br. J. Nutr. 106, 114–122. doi: 10.3168/jds.S0022-0302(07)72638-3
Velazco J., Mayer D., Zimmerman S., and Hegarty R. (2016). Use of short-term breath measures to estimate daily methane production by cattle. Animal 10, 25–33. doi: 10.1017/S1751731115001603
Visscher C. F., Witzmann S., Beyerbach M., and Kamphues J. (2013). Watering cattle (young bulls) with brackish water–a hazard due to its salt content? Tierarztl. Prax. Ausg. G: Grosstiere – Nutztiere 41, 363–370. doi: 10.1055/s-0038-1623198
Wang Y., McAllister T. A., Yanke L. J., Xu Z. J., Cheeke P. R., and Cheng K. J. (2000). In vitro effects of steroidal saponins from Yucca schidigera extract on rumen microbial protein synthesis and ruminal fermentation. J. Sci. Food Agric. 80, 2114–2122. doi: 10.1016/S0377-8401(98)00137-0
Wang J. K., Ye J. A., and Liu J. X. (2012). Effects of tea saponins on rumen microbiota, rumen fermentation, methane production and growth performance—a review. Trop. Anim. Health Prod. 44, 697–706. doi: 10.1007/s11250-011-9960-8
Wasson D. E. and Hristov A. N. (2022). Enteric methane mitigation through Asparagopsis taxiformis supplementation and potential algal alternatives. Front. Anim. Sci. 3. doi: 10.3389/fanim.2022.999338
White H. C., Davis N. G., Van Emon M. L., DelCurto-Wyffels H. M., Wyffels S. A., and DelCurto T. (2024). Impacts of increasing levels of salt on intake, digestion, and rumen fermentation with beef cattle consuming low-quality forages. J. Anim. Sci. 102, skae284. doi: 10.1093/jas/skae284
Keywords: enteric methane, greenhouse gas, cattle, molasses, pellet supplementation
Citation: Nikoloric M, Simanungkalit G, Possell M, González LA and Chaves AV (2025) Assessment of molasses products containing additives to reduce methane emissions from beef cattle. Front. Anim. Sci. 6:1666818. doi: 10.3389/fanim.2025.1666818
Received: 15 July 2025; Accepted: 16 September 2025;
Published: 23 October 2025.
Edited by:
David L. Harmon, University of Kentucky, United StatesReviewed by:
Arda Yıldırım, Gaziosmanpaşa University, TürkiyeOlegario Hernández, National Institute of Agricultural Technology (INTA), Argentina
Copyright © 2025 Nikoloric, Simanungkalit, Possell, González and Chaves. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alex V. Chaves, YWxleC5jaGF2ZXNAc3lkbmV5LmVkdS5hdQ==