 Rannyelle Gomes Souza1
Rannyelle Gomes Souza1 Maria Isabelly Leite Maia2
Maria Isabelly Leite Maia2 Luiz Arthur dos Anjos Lima2Larissa Kellen da Cunha Morais2Severino Guilherme Caetano Gonçalves dos Santos3Tarsys Noan Silva Veríssimo4José Danrley Cavalcante dos Santos2Pavlos Vinícius do Nascimento2Kilmer Oliveira Soares5Fernando Guilherme Perazzo Costa6Ricardo Romão Guerra7
Luiz Arthur dos Anjos Lima2Larissa Kellen da Cunha Morais2Severino Guilherme Caetano Gonçalves dos Santos3Tarsys Noan Silva Veríssimo4José Danrley Cavalcante dos Santos2Pavlos Vinícius do Nascimento2Kilmer Oliveira Soares5Fernando Guilherme Perazzo Costa6Ricardo Romão Guerra7 Lucas Rannier Ribeiro Antonino Carvalho8*
Lucas Rannier Ribeiro Antonino Carvalho8* Edilson Paes Saraiva2
Edilson Paes Saraiva2- 1Integrated Graduate Program in Tropical Animal Science, Federal University of Northern Tocantins, Araguaína, Tocantins, Brazil
- 2Research Group in Bioclimatology, Ethology and Animal Welfare (BioEt), Department of Animal Science, Federal University of Paraíba, Areia, Paraíba, Brazil
- 3Department of Animal Science, Center for Agricultural Sciences, State University of Maranhão, São Luís, Maranhão, Brazil
- 4University Center UNIFIP, Patos, Paraíba, Brazil
- 5Technological Education Center Institute, Marco, Ceará, Brazil
- 6Department of Animal Science, Federal University of Paraíba, Areia, Paraíba, Brazil
- 7Department of Veterinary Sciences, Federal University of Paraíba, Areia, Paraíba, Brazil
- 8Department of Physiology and Pharmacology, Karolinska Institutet, Stockholm, Sweden
Heat stress is one of the major constraints on quail production, compromising feed intake, egg quality, and overall welfare under tropical conditions. This study aimed to evaluate whether dietary methionine + cystine (Met+Cys) supplementation could mitigate the adverse effects of thermal stress on laying Coturnix japonica during peak egg production. A total of 504 female quails were distributed across 12 treatments in a 3 × 4 factorial design, combining three Met+Cys levels (100%, 110%, and 120% of NRC recommendations) and four ambient temperatures (20, 24, 28, and 32 °C). Over an 84-day trial, productive performance, egg quality, physiological markers (cloacal temperature, respiratory rate), and tissue morphology (duodenum, jejuno, hepatic, uterine) were evaluated. Heat stress markedly impaired reproductive performance and egg quality, particularly at 32 °C, with no significant interaction between temperature and supplementation level. Principal Component Analysis revealed structural degradation in intestinal, hepatic, and uterine tissues under elevated temperatures, partially alleviated by 120% Met+Cys at 24 °C. However, increased supplementation failed to reverse thermal-induced production losses under severe heat. These findings demonstrate that ambient temperature exerts a stronger influence than methionine and cysteine supplementation on overall quail performance. While nutritional interventions may help preserve tissue architecture under moderate heat stress (up to 28 °C), strategic thermal environment management remains the primary tool to safeguard welfare and productivity. This integrative approach underscores the value of multidimensional analysis in poultry science and supports refined thermal-nutritional management in quail production systems.
1 Introduction
Japanese quail (Coturnix japonica) farming has gained prominence as a fast-growing segment of the Brazilian poultry industry, driven by nutritional innovations and advances in genetic selection and environmental adaptation. According to the Brazilian Institute of Geography and Statistics (IBGE, 2024), national production reached 250.3 million dozen quail eggs, reflecting a 5.4% increase over the previous year. This upward trend highlights the intensification of production systems and the pursuit of greater zootechnical efficiency under tropical conditions.
However, high ambient temperatures remain one of the major constraints on performance in intensive systems. Quails are physiologically sensitive to thermal stress, with narrow thermoneutral zones. When exposed to elevated temperatures, especially under high humidity, birds experience reduced feed intake, impaired nutrient absorption, and altered metabolic energy partitioning, favoring maintenance over productivity and compromising both egg yield and animal welfare (Soares et al., 2019; Matos Júnior et al., 2023).
To mitigate these effects, dietary strategies have emerged as practical tools to enhance resilience under heat stress. One such approach involves the supplementation of DL-methionine, an amino acid added to diets to meet graded requirements for methionine plus cystine (Met + Cys). These sulfur-containing amino acids are metabolically interconvertible and play key roles in protein synthesis, egg formation, lipid metabolism, antioxidant defense via glutathione production, and epithelial tissue integrity (Harper et al., 1970; Mitchell and Robbins, 1984; Geraert et al., 1996). Recent findings demonstrated that methionine supplementation can enhance growth performance, productivity, and health parameters in Japanese quails, reinforcing its potential role in nutritional strategies under challenging environments (Aziz et al., 2025). In addition, other studies have shown that DL-methionine supplementation contributes to improved breast muscle yield and antioxidant defense, while maintaining meat quality in Japanese quails (Mahmoud et al., 2023).
While reference standards such as NRC (1994) and region-specific data (Silva and Costa, 2009) guide dietary formulations for quail, there is still limited understanding of how Met + Cys requirements may shift under thermal challenge. Beyond amino acid supplementation, functional minerals such as dietary zinc have also shown promising results in preserving bone strength and performance in quails under thermal stress, suggesting a synergistic role of nutritional components in adaptive responses (Lima et al., 2025).
Recent evidence indicates that elevated levels of sulfur-containing amino acids may contribute to preserving intestinal and reproductive tissue morphology, as well as modulating calcium transport proteins under heat stress conditions (Moraes et al., 2021). Moreover, (Wasti et al., 2020). highlighted that nutritional, environmental, and genetic strategies play a critical role in mitigating the physiological and productive impacts of heat stress in poultry systems. Despite these advances, few studies have simultaneously addressed the combined influence of ambient temperature and DL-methionine supplementation on the productive, physiological, and morphological responses of laying Japanese quails. This gap limits the optimization of nutritional strategies under climate stress scenarios.
Therefore, the present study aimed to evaluate whether different levels of DL-methionine supplementation, corresponding to 100%, 110%, and 120% of NRC recommendations for Met + Cys, could mitigate the negative effects of thermal stress on laying performance, egg quality, physiological parameters, and the histomorphology of key tissues (intestine, liver, and uterus) in Japanese quails during the peak laying period. We hypothesized that elevated temperatures would impair all measured variables, and that higher supplementation levels could provide protective effects under moderate thermal stress. By integrating productive, physiological, and histological data, this study seeks to provide a comprehensive understanding of nutrition–environment interactions and to contribute to the refinement of feeding strategies for quails raised in tropical environments.
2 Materials and methods
2.1 Experimental site, ethical approval, and experimental design
The trial was conducted at the Research Unit in Bioclimatology, Ethology, and Animal Welfare, Department of Animal Science, Federal University of Paraíba (UFPB), Areia, Brazil (6°57′02″ S, 35°41′04″ W). All procedures were approved by the Committee of Ethics in Animal Use of UFPB (protocol no. 097/2015) and followed the national and international guidelines for the care and use of animals in research.
A total of 504 female Japanese quails (Coturnix japonica), aged 56 days and weighing approximately 180 g, were used in the experiment. Before the beginning of the experiment, all quails were kept under standardized management at the Research unit, receiving balanced commercial ration and ad libitum water. When the birds had an average of 93% laying, were distributed in a completely randomized 3 × 4 factorial design. Treatments combined three dietary levels of methionine + cystine (Met+Cys: 100%, 110%, and 120% of NRC (1994) recommendations) and four ambient temperatures (20, 24, 28, and 32°C). Each treatment included six replicates, totaling 72 experimental units (3 dietary x 4 environments x 6 replicates), with six birds in each replicate.
2.2 Pre-experimental management and housing
Prior to the start of the experimental period, a two-week standardization phase was conducted to monitor egg production and ensure uniform laying performance across experimental units. Birds were then distributed based on average productivity, and each group was weighed and identified according to treatment and replicate.
The experiment began when the quails were 56 days old, with a baseline laying rate of approximately 93%. The trial lasted 84 days and was divided into four consecutive 21-day phases.
The study was carried out in four independent bioclimatic chambers, each measuring 5.40 m in length and 3.83 m in width. Each chamber contained galvanized wire cages (47 × 23.5 × 16 cm; length × width × height), equipped with trough-type feeders and nipple drinkers suitable for adult laying quails. Birds were housed in groups of seven per cage, with each cage representing one experimental unit (replicate). The stocking density was consistent across all treatments.
Feed was offered twice daily (at 07:00 and 16:00), and fresh water was provided ad libitum, being replaced twice daily to avoid overheating. Environmental control within the chambers was maintained using heaters with digital thermostats and air conditioning units with automatic temperature regulation. Air renewal and circulation were ensured by axial wall-mounted exhaust fans. Ambient temperature and relative humidity were monitored twice daily (morning and afternoon) using calibrated digital thermohygrometers, positioned at the birds’ center-of-mass height to accurately reflect the microclimate.
A fixed photoperiod of 17 hours of light and 7 hours of darkness was applied throughout the experimental period, following standard protocols for laying performance.
The protocols for the different thermal environments were established based on the studies of Sousa (2013). The birds were continuously exposed to different temperatures throughout the evaluated production cycles, including two within the thermal comfort zone (20 and 24°C) and two classified as heat stress (moderate, 28°C and severe, 32°C). The general layout and structural configuration of the bioclimatic chambers used in this study are illustrated in Figure 1.
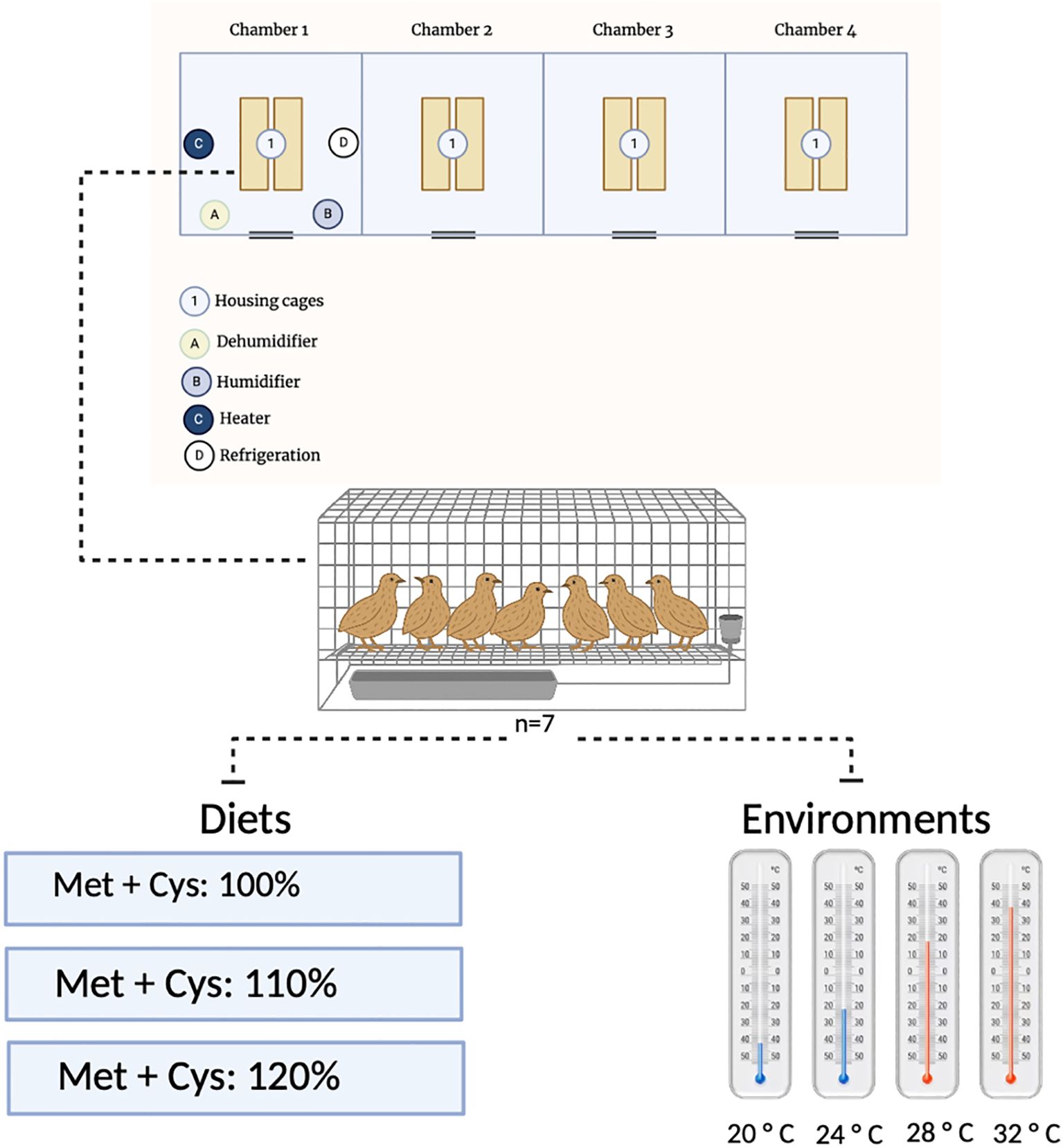
Figure 1. Schematic layout of the climate chambers and equipment used for environmental control during the experiment.
2.3 Diets and nutritional composition
Experimental diets were formulated to meet the nutritional requirements of laying quails according to the NRC (1994), with adjustments for the levels of methionine plus cystine (Met + Cys). All diets were isocaloric and isonitrogenous, differing only in the total Met + Cys content, which was adjusted by the inclusion of DL-methionine (99% purity; Evonik Industries, Hanau, Germany) and inert filler.
The three experimental levels were:
100% NRC: 0.70% Met + Cys.
110% NRC: 0.77% Met + Cys.
120% NRC: 0.84% Met + Cys.
Diets were prepared using commercially available ingredients (corn, soybean meal, soybean oil, dicalcium phosphate, limestone, premixes, and additives), ground and homogenized in a mechanical mixer at the UFPB feed mill. Representative feed samples were collected and analyzed for proximate composition (dry matter, crude protein, ether extract, crude fiber, ash, calcium, and phosphorus) according to AOAC (2005) methods, while amino acid contents were calculated based on tabular values and formulation software.
The ingredient composition and calculated nutrient levels of the basal diet are presented in Table 1.

Table 1. Composition of experimental diets with three methionine + cystine levels for laying Japanese quails under four ambient temperatures.
2.4 Performance measurements
The performance characteristics evaluated in the experimental trial were: feed intake, egg laying percentage, whole egg percentage, average egg weight, egg mass, feed conversion per dozen and per egg mass produced. Data on the number of whole eggs were recorded daily on specific forms. The number of broken eggs was also collected for later calculation of the egg laying percentage and whole eggs.
Feed consumption was determined for each period by the difference between the amount of feed provided and the remaining feed at the end of each twenty-one-day period. The result was divided by the average number of birds in each plot and expressed in grams per bird per day. The feed corresponding to each experimental unit was weighed and stored in 3 kg plastic buckets, and identified for each treatment and repetition. In the case of birds that died during the period, the average consumption of the plot was corrected, obtaining the true average consumption for the experimental unit.
For performance analyses, egg-laying percentages were also measured by each experimental group, relative to total egg production. Egg-laying percentage was obtained by dividing the total number of eggs laid per cage per week by the average number of birds, multiplied by seven, and the result multiplied by 100. The percentage of healthy eggs was obtained by dividing the total number of healthy eggs produced per week by the average number of birds, multiplied by seven, and the result multiplied by 100.
Egg mass was obtained by multiplying the average egg weight of each plot by the egg laying percentage of the same plot and dividing the result by 100 and expressing it in grams of eggs per bird per day. All individual eggs of the day were weighed on the last three days of each cycle of each experimental plot.
2.5 Egg quality assessment
Egg quality was assessed on the last three days of each 21-day period. The following quality characteristics were assessed: specific gravity, percentages of yolk, albumen and shell, shell thickness, shell weight, Haugh unit and yolk color. All eggs from each plot were weighed on an analytical balance with a precision of 0.01 g. Shell thickness was measured using a digital caliper (Mitutoyo, Japan), and yolk color was determined using a DSM YolkFan™ colorimetric scale with integrated light source.
The specific gravity of the eggs was calculated according to the methodology described by Stadelman and Cotterill (1995). All collected intact eggs were immersed in NaCl solutions with densities ranging from 1.005 to 1.100 g/cm³.
The yolk percentage was determined by dividing the yolk weight by the egg weight and multiplying the result by 100, and the albumen percentage was determined by dividing the albumen weight by the egg weight and multiplying the result by 100. To calculate the shell percentage, the shells were initially dried in a forced ventilation oven at 65°C for three days. Afterwards, the room temperature and the shell temperature were allowed to equilibrate before weighing them.
The percentage of shell was then obtained by dividing the weight of the shell by the weight of the egg, and the result multiplied by 100. To obtain the thickness of the shell, the same shells used in the previous item were used. Two measurements were taken in the equatorial region of the egg, and the average of these measurements was then calculated, and the result expressed in millimeters.
Subsequently, the three most homogeneous eggs, close to the average weight, selected from each plot, were identified and broken for measurements of the other variables such as albumen height. The albumen height was considered the measurement taken between the surface where the egg was broken and the intersection between the albumen and the yolk, with the aid of a caliper on a flat glass surface.
The yolk color was estimated using DSM’s digital fan, the YolkFan™, which allows precise color measurement through a highly sensitive device that evaluates the color of the egg yolk having its own light source, meaning that the reading is made without any external interference such as light or the different perceptions of the human eye and judgment.
To calculate the Haugh unit (HU), the albumen height and egg weight were substituted into the formula proposed by Haugh (1937):
HU = 100 × log(H – 1.7W^0.37 + 7.57) Where H is the albumen height in millimeters and W is the egg weight in grams.
The yolks of the three eggs from each plot were weighed individually on a digital scale to determine the percentage of yolk in relation to the egg weight. For the percentages of yolk and shell, the calculation was made in relation to the egg weight. However, for the percentage of albumen, it was determined by the difference of the constituents that made up the egg weight: 100 – (% yolk + % shell).
2.6 Physiological and histomorphological assessments
At the end of the 84-day trial, two birds per replicate (n = 12 per treatment) were randomly selected and euthanized by cervical dislocation, following ethical guidelines for poultry research. Samples of the duodenum, jejunum, liver, and uterus were immediately collected, fixed in 10% neutral buffered formalin, processed, and stained with hematoxylin and eosin. For each bird, at least two slides per tissue were prepared, and histomorphometric measurements were performed on 10 well-oriented villi and crypts per slide. Villus height, crypt depth, villus-to-crypt ratio, epithelial integrity, and structural morphology of liver and uterine tissues were evaluated using an Olympus BX53F microscope (Tokyo, Japan) equipped with a DP73 digital camera. Digital images were analyzed with CellSens Dimension® software (Olympus, Tokyo, Japan) at 20× magnification, with measurements taken in randomly selected microscopic fields.
Physiological parameters were measured in two birds per replicate at 13:00 on the final day of each cycle. Respiratory rate (RR) was determined by counting thoracic movements per minute. Cloacal temperature (CT) was measured using a digital clinical thermometer inserted 2 cm into the cloaca for approximately 30 seconds, until stabilization.
2.7 Statistical analysis
All statistical analyses were performed using SAS® software, version 6.50 (SAS Institute Inc., Cary, NC, USA) (SAS Institute Inc, 1999). Data were analyzed using a two-way analysis of variance (ANOVA), with dietary Met + Cys level, ambient temperature, and their interaction as fixed effects. The experimental unit was the cage for productive performance and egg quality traits, and the individual bird for physiological and histological measurements. Data were first checked for normality (Shapiro–Wilk test) and homogeneity of variances (Levene’s test). Means were compared using Tukey’s test at a significance level of p < 0.05.
Principal Component Analysis (PCA) was conducted using the PROC PRINCOMP procedure of SAS® (SAS Institute Inc, 1999), following methodological recommendations for animal science studies (Khanikar et al., 2024) and poultry immunology (Varmuzova et al., 2016). Two models were constructed:
PCA 1: based on intestinal morphometric parameters (villus height, crypt depth, villus:crypt ratio).
PCA 2: integrating physiological (respiratory rate, cloacal temperature) and histological variables (preserved hepatic area and uterine area).
Eigenvalues greater than 1 were used as the criterion for component retention, and biplots were generated to visualize treatment clustering.
3 Results
3.1 Productive performance
There was no significant interaction between dietary Met+Cys levels and ambient temperature for any performance parameter (p > 0.05). However, temperature alone significantly affected all metrics. As shown in Table 2, feed intake decreased progressively with rising temperature: quails at 20°C consumed 26.11 g/bird/day, while those at 32°C consumed only 20.21 g/bird/day (p < 0.0001). Egg production also declined with temperature, reaching 75.26% at 32°C compared to 90.69% at 20°C (p < 0.0001). Egg mass followed the same trend, dropping from 10.90 g to 8.60 g/bird/day (p < 0.0001).

Table 2. Performance of laying Japanese quails fed different methionine + cystine levels under four ambient temperatures.
Feed conversion ratio per kilogram of egg mass increased under heat stress, reaching 2.32 at 32°C, while the best efficiency was recorded at 28°C (2.14) (p = 0.0074). Feed conversion per dozen eggs was also significantly higher at 32°C (p = 0.0123). Across all supplementation levels, Met+Cys did not significantly influence any performance variables, with values remaining statistically similar among the 100%, 110%, and 120% groups.
3.2 Egg quality parameters
Temperature had a significant impact on egg quality, as detailed in Table 3. Egg weight was lowest at 32°C (11.83 g), with yolk weight also decreasing to 3.59 g in this treatment (p < 0.001). Albumen height decreased from 4.86 mm at 24°C to 4.43 mm at 32°C, and Haugh units dropped accordingly (p < 0.0001). Shell thickness was significantly affected by heat, measuring 0.24 mm at 32°C versus 0.25 mm at milder temperatures (p = 0.0111).

Table 3. Egg quality of Japanese quails fed different methionine + cystine levels under four ambient temperatures.
Despite these changes, eggs remained within “AA” commercial classification standards. Supplementation with Met+Cys showed no significant effect on egg weight, shell integrity, or internal quality (p > 0.05). Yolk color varied slightly among groups, but the differences were not meaningful in the context of quail egg commercialization.
3.3 Physiological and histomorphological
Principal Component Analysis (PCA) based on intestinal morphometry (villus height, crypt depth, and villus:crypt ratio) accounted for 72.9% of total variance (PC1: 46.5%; PC2: 26.4%). As shown in Figure 2, treatments at 20°C and 24°C, particularly those with 120% Met+Cys, were clustered in the positive PC1 quadrant, associated with better mucosal integrity. In contrast, treatments at 28°C and 32°C, especially with lower supplementation levels, clustered in the negative region, indicating epithelial disruption.

Figure 2. Principal component analysis (PCA) of intestinal morphometric parameters (villus height, crypt depth, and villus:crypt ratio) of Japanese quails under different temperatures.
A second PCA model integrating physiological and histological data (cloacal temperature, respiratory rate, preserved hepatic area, and functional uterine area) explained 95.5% of variance (PC1: 79.4%; PC2: 16.1%) (Figure 3). Treatments under heat stress (28°C and 32°C) were grouped on the positive PC1 axis, characterized by increased respiratory effort, elevated body temperature, and reduced tissue preservation. Conversely, birds at 20°C and 24°C with 120% Met+Cys supplementation were distributed on the negative PC1 axis, reflecting physiological stability and structural tissue integrity.

Figure 3. Principal component analysis (PCA) integrating physiological and histological variables (cloacal temperature, respiratory rate, preserved hepatic area, and functional uterine area).
4 Discussion
4.1 Productive performance
Ambient temperature was the most influential factor affecting the productive performance of laying Japanese quails. As temperature increased, birds reduced feed intake, likely as a thermoregulatory mechanism to limit metabolic heat production, resulting in significant declines in egg production and mass. These findings are consistent with Sahin et al (Sahin et al., 2002), who reported that chronic heat exposure disrupts metabolic and endocrine homeostasis, thereby impairing performance.
Feed intake was significantly higher at 20°C compared to 24°C and 28°C, although both 20 °C and 24°C are generally considered within the thermoneutral zone for quails. This difference likely reflects metabolic adjustments, as quails at 20°C may increase feed consumption to maintain body temperature, whereas at 24°C and 28°C, thermal conditions impose lower energetic demands. No significant difference in intake was observed between 24°C and 28°C, suggesting that the variation represents a physiological adaptation rather than a detrimental thermal effect (Santos et al., 2019).
Notably, methionine + cystine supplementation did not improve productivity under severe heat stress (32°C), corroborating results by Lima et al (Lima et al., 2024), who found limited efficacy of amino acid supplementation under intense thermal challenge. However, recent studies demonstrated that methionine supplementation can enhance growth performance, productivity, and health parameters in Japanese quails (Aziz et al., 2025), indicating that its effectiveness may be more pronounced under moderate heat stress conditions. Similarly, DL-methionine supplementation improved growth, breast muscle yield, and antioxidant defense in Japanese quails (Mahmoud et al., 2023), reinforcing its importance as a nutritional tool even when productive benefits are not evident under extreme thermal load. In addition, chromium–methionine supplementation improved performance, egg quality, and blood antioxidant status in heat-stressed quails (Khalifah et al., 2022), highlighting the potential of combined nutritional approaches to enhance resilience in hot climates.
These findings support the notion that while amino acid supplementation may contribute to sustaining performance under moderate temperatures, it is insufficient as a standalone solution when environmental temperatures surpass the birds’ thermotolerance threshold. This threshold, influenced by species-specific metabolic characteristics, has been previously described by Lara and Rostagno (2013) in various poultry systems. Previous studies also showed that methionine and lysine levels influence the productive performance of pigs under thermoneutral conditions (Ferreira et al., 2005). These findings, although species-specific, support the principle that amino acid-based strategies can be metabolically efficient under well-controlled environments.
4.2 Egg quality parameters
Egg quality deteriorated progressively with increasing temperature, particularly at 32°C, where reductions in egg and yolk weight, albumen height, and Haugh Unit values were observed. These effects are likely linked to impaired protein synthesis, mineral metabolism, and hepatic function under thermal stress, as reported by (Ebeid et al., 2012). and Abdulkadir and Reddy (2023). Additionally, respiratory alkalosis and reduced carbonic anhydrase activity at high temperatures can inhibit calcium carbonate deposition in the shell gland, weakening shell formation (Yahav et al., 2000). Consistently, this study observed that eggshell thickness was significantly reduced at temperatures of 28–32°C compared to 20–24°C, confirming that heat stress impairs calcium carbonate deposition and results in thinner shells.
Despite these physiological disruptions, all egg quality traits remained within commercial standards, with eggs classified as “AA.” Methionine + cystine supplementation did not significantly influence internal or external egg quality. Similar to our findings, (Castro and Kim, 2020). and (Kwak et al., 2024). also reported limited direct effects of methionine supplementation on egg quality traits. However, evidence from chromium–methionine supplementation indicates that combined nutritional approaches may exert beneficial effects on egg quality in quails exposed to high temperatures, improving shell thickness, albumen height, and blood antioxidant profile (Khalifah et al., 2022). These results suggest that while methionine alone may not alter egg quality, its integration with other nutritional strategies could enhance resilience against thermal stress.
4.3 Physiological and morphological responses
Thermal stress triggered classical physiological responses in quails, including elevated respiratory rate and cloacal temperature, consistent with the findings of (Ramnath et al., 2008). and Tinoco and Gates (2011). These changes reflect increased evaporative heat loss but also indicate systemic stress (Mashaly et al., 2004), which can compromise welfare and productivity.
Histomorphological evaluations revealed significant degeneration in the intestinal epithelium and reduced hepatic and uterine tissue preservation at higher temperatures, corroborating studies by Abdulkadir and Reddy (2023) and Maiorano et al (Matos Júnior et al., 2023). These structural impairments suggest a loss of absorptive capacity, epithelial integrity, and reproductive efficiency.
Interestingly, at 24°C, the group receiving 120% Met+Cys maintained mucosal integrity and physiological homeostasis, suggesting a protective effect under moderate heat stress. Similar protective effects of methionine supplementation against heat-induced mucosal injury have been reported, demonstrating its role in maintaining epithelial integrity (Feng et al., 2024). Moreover, maternal dietary methionine supplementation was shown to improve lipid metabolism and intestinal development in both laying quails and their progeny (Santana et al., 2025), indicating that its benefits may extend across generations. Complementarily, studies in breeder Japanese quails have demonstrated that methionine supplementation also enhances reproductive performance, antioxidant status, immunity, and maternal antibody transmission under heat stress (Kalvandi et al., 2019), reinforcing the relevance of methionine in sustaining physiological resilience and reproductive success under challenging environments.
These findings align with those reported by Moraes et al (Moraes et al., 2021), who demonstrated that methionine supplementation modulates intestinal, hepatic, and uterine histomorphology in laying quails under both thermoneutral and heat stress conditions. Their study further highlighted the upregulation of epithelial calcium carriers (Calbindin-D28k and TRPV6), suggesting a mechanistic role of sulfur amino acids in maintaining calcium homeostasis and tissue integrity under thermal stress.
The integrated PCA approach synthesized the complex relationships among productive, physiological, and morphological responses. Birds under moderate temperatures and high Met+Cys supplementation clustered on the negative PC1 axis, characterized by stable physiology and preserved tissues. In contrast, heat-stressed birds showed greater physiological activation and histological damage. These results confirm that cloacal temperature, respiratory rate, and tissue preservation are effective biomarkers of thermal stress. The use of PCA to visualize and discriminate treatment effects in this study underscores its value in integrating multidimensional biological responses in poultry under stress (Varmuzova et al., 2016).
5 Conclusions
Ambient temperature was the main factor influencing the productive, physiological, and histological responses of laying Japanese quails. Supplementation with up to 120% of the recommended levels of methionine + cystine did not prevent performance losses under severe heat stress (32°C), but under moderate thermal conditions (up to 28°C) it was associated with some preservation of intestinal, hepatic, and uterine structures.
These findings indicate that nutritional strategies based on sulfur amino acids should not be considered in isolation, but rather in association with environmental management practices. The integration of dietary interventions with adequate environmental control appears to be essential for sustaining performance and maintaining tissue integrity under challenging conditions.
Future studies should focus on exploring combined mitigation approaches, particularly the synergistic effects of nutrition and environmental management, to enhance the thermal resilience of laying quails.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
All experimental procedures were approved by the Animal Use Ethics Committee of UFPB (protocol no. 097/2015) and complied with national and international standards for animal experimentation. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
RS: Formal Analysis, Investigation, Methodology, Writing – original draft, Writing – review & editing. MM: Formal Analysis, Investigation, Methodology, Writing – review & editing. LA: Formal Analysis, Investigation, Methodology, Validation, Writing – review & editing. LM: Formal Analysis, Investigation, Methodology, Writing – review & editing. SS: Formal Analysis, Investigation, Methodology, Validation, Writing – review & editing. TV: Formal Analysis, Investigation, Methodology, Validation, Writing – review & editing. JS: Formal Analysis, Investigation, Methodology, Validation, Writing – review & editing. PN: Formal Analysis, Investigation, Methodology, Validation, Writing – review & editing. KS: Data curation, Formal Analysis, Investigation, Methodology, Writing – review & editing. FC: Formal Analysis, Investigation, Methodology, Supervision, Visualization, Writing – review & editing. RG: Formal Analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Writing – review & editing. LC: Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Writing – original draft, Writing – review & editing. ES: Conceptualization, Funding acquisition, Investigation, Project administration, Resources, Supervision, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This work was supported by Karolinska Institutet through an institutional publication agreement. The funding covered the open access publication fees.
Acknowledgments
The authors would like to thank the Coordination for the Improvement of Higher Education Personnel (CAPES), the National Council for Scientific and Technological Development (CNPq), and the Federal University of Paraíba (UFPB) for providing scholarships and research infrastructure. We also acknowledge the support from the Research Unit in Bioclimatology, Ethology, and Animal Welfare at UFPB for hosting the experimental procedures.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abdulkadir A. and Reddy P. G. (2023). Effect of heat stress on poultry health and performance. J. Appl. Anim. Res. 51, 23–31.
AOAC (2005). Official methods of analysis. 18th ed (Gaithersburg, MD: Association of Official Analytical Chemists).
Aziz T., Tariq M., and Ullah N. (2025). Effects of Methionine Supplementation on Growth, Productivity, and Health Parameters in Japanese Quails (Coturnix coturnix japonica). J. Anim. Plant Res. 2(3), 6–9.
Castro de F. L. S. and Kim W. K. (2020). Secondary functions of arginine and sulfur amino acids in poultry health: Review. Animals 10(11), 2106. doi: 10.3390/ani10112106
Ebeid T. A., Suzuki T., and Sugiyama T. (2012). High temperature affects eggshell quality and calbindin-D28k expression in laying hens. Poult Sci. 91, 2282–2287. doi: 10.3382/ps.2011-01898
Feng S., Zeng J., Li J., Yuan S., and Wu B. (2024). Alleviating effect of methionine on intestinal mucosal injury induced by heat stress. J. Therm Biol. 123, 103935. doi: 10.1016/j.jtherbio.2024.103935
Ferreira R. A., Oliveira R. F. M., Donzele J. L., Araújo C. V., Silva F. C. O., Fontes D. O., et al. (2005). Redução do nível de proteína bruta e suplementação de aminoácidos em rações para suínos machos castrados mantidos em ambiente termoneutro dos 30 aos 60 kg. Rev. Bras. Zootec 34, 548–556. doi: 10.1590/S1516-35982005000200024
Geraert P. A., Padilha J. C. F., and Guillaumin S. (1996). Metabolic and endocrine changes induced by chronic heat exposure in broiler chickens: growth performance, body composition and energy retention. Br. J. Nutr. 75, 195–204. doi: 10.1017/BJN19960124
Harper A. E., Benevenga N. J., and Wohlhueter R. M. (1970). Effects of ingestion of disproportionate amounts of amino acids. Physiol. Rev. 50, 428–458. doi: 10.1152/physrev.1970.50.3.428
IBGE (2024). Indicadores pecuários. Available online at: https://www.ibge.gov.br/ (Accessed August 15, 2025).
Kalvandi O., Sadeghi A., and Karimi A. (2019). Methionine supplementation improves reproductive performance, antioxidant status, immunity and maternal antibody transmission in breeder Japanese quail under heat stress conditions. Arch. Anim. Breed. 62, 275–286. doi: 10.5194/aab-62-275-2019
Khalifah A. M., El-Naggar K., Ismail I. E., Alagawany M., Taha A. E., Laudadio V., et al. (2022). Leverage of chromium–methionine supplementation in laying Japanese quails’ diets on performance, egg quality and blood traits challenged by heat stress. Adv. Anim. Vet. Sci. 10, 676–684. doi: 10.17582/journal.aavs/2022/10.3.676.684
Khanikar D., Phookan A., Das E., and Tolenkhomba T. C. (2024). Principal component analysis and its role in animal science: a review. Int. J. Vet. Sci. Anim. Husbandry 9, 264–268.
Kwak M. J., Kang A., Eor J. Y., Ryu S., Choi Y., Heo J. M., et al. (2024). Protective effect of methionine supplementation on heat stress-induced intestinal injury in broilers. J. Anim. Sci. Technol. 66, 193–200. doi: 10.1186/s42523-024-00303-w
Lara L. J. and Rostagno M. H. (2013). Impact of heat stress on poultry production. Animals. 3, 356–369. doi: 10.3390/ani3020356
Lima dos L. A. A., Maia M. I. L., Morais L. K. C., Veríssimo T. N. S., dos Santos J. D. C., da Silva A. L., et al. (2024). How does the adoption of nutritional plans influence the performance of meat quail during the growth phase in hot environments? Front. Vet. Sci. 11. doi: 10.3389/fvets.2024.1469516
Lima L. A. A., Moraes T. A., Morais L. K. C., de Amorim MLCCM, Veríssimo T. N. S., dos Santos J. D. C., et al. (2025). Effect of zinc level and the thermal environment on the zootechnical performance and tibia-breaking strength of Japanese quails. Front. Vet. Sci. 11. doi: 10.3389/fvets.2024.1467487
Mahmoud R. E., Ghanem H. M., and Salem S. M. (2023). Effect of DL-methionine supplementation in Japanese quail diets on growth, meat quality and health. Alexandria J. Veterinary Sci. 77, 64–75. doi: 10.5455/ajvs.147511
Mashaly M. M., Hendricks G. L., Kalama M. A., Gehad A. E., Abbas A. O., and Patterson P. H. (2004). Effect of heat stress on production parameters and immune responses of commercial laying hens. Poult Sci. 83, 889–894. doi: 10.1093/ps/83.6.889
Matos Júnior J. J. L., Furtado D. A., Ribeiro N. L., Marques J. I., Leite P. G.., do Nascimento J. W. B., et al. (2023). Productive performance, egg quality and the morphometry of the organs of Japanese quails (Coturnix coturnix japonica) kept at different temperatures. Food Sci. Technol. 43, e117822. doi: 10.1590/fst.117822
Mitchell N. S. and Robbins K. R. (1984). Effect of dietary energy level on total sulfur amino acid requirement of growing broilers. Tenn Farm Home Sci. 6), 6–10.
de Moraes L. R., Delicato M. E. A., da Silva Cruz A., Pereira da Silva H. T. F. N., de Vasconcelos Alves C. V. B., Campos D. B., et al. (2021). Methionine supplementing effects on intestine, liver and uterus morphology in laying quail under thermal stress. PloS One 16, e0245615. doi: 10.1371/journal.pone.0245615
Ramnath V., Rekha P. S., and Sujatha K. S. (2008). Amelioration of heat stress-induced disturbances of antioxidant defense in chicken by Brahma Rasayana. Evid Based Complement Alternat Med. 5, 77–84. doi: 10.1093/ecam/nel116
Sahin K., Sahin N., and Küçük O. (2002). Effects of dietary chromium and ascorbic acid on nutrient digestion and antioxidant status in laying hens reared at low ambient temperature. Biol. Trace Elem Res. 87, 113–124. doi: 10.1385/BTER:87:1-3:113
Santana T. P., Costa L. S., Silva L. M., Almeida H. R., Rocha J. B., Oliveira C. F., et al. (2025). Maternal dietary methionine supplementation enhances lipid metabolism and intestinal development of Japanese quails and their progeny. Anim. Sci. J. 96, e70044. doi: 10.1111/asj.70044
Santos T. C., Silva J. H., Vieira F. C. S., Rocha J. H. B., Nascimento R. R., Furtado D. A., et al. (2019). Productive performance and surface temperatures of Japanese quail exposed to different environment conditions at start of lay. Poultry Sci 98, 2830–2839. doi: 10.3382/ps/pez068
Silva J. H. V. and Costa F. G. P. (2009). Table for Japanese and European quails. 2nd ed (Jaboticabal: FUNEP).
Soares K. O., Saraiva E. P., Santos J. D. C., Ferreira R. A., Furtado D. A., Farias A. L. N., et al. (2019). Effect of ambient temperature on production parameters and egg quality of Japanese quail (Coturnix japonica). Biol. Rhythm. Res. 52, 1130–1137. doi: 10.1080/09291016.2019.1619128
Sousa M. S. (2013). Determination of thermal comfort ranges for meat quails of different ages. Universidade Federal de Viçosa, Viçosa. Available online at: https://www.locus.ufv.br/handle/123456789/752 (Accessed August 15, 2025).
Stadelman W. J. and Cotterill O. J. (1995). Egg science and technology. 4th ed (New York: Food Products Press).
Tinoco I. F. F. and Gates R. S. (2011). “Environmental influences on poultry production,” in Environmental stress in poultry (CAB International, Wallingford), 1–22.
Varmuzova K., Matiasovic J., and Faldyna M. (2016). Principal component analysis in poultry immunology. Vet. Immunol. Immunopathol. 171, 59–66.
Wasti S., Sah N., and Mishra B. (2020). Impact of heat stress on poultry health and performances, and potential mitigation strategies. Animals. 10, 1266. doi: 10.3390/ani10081266
Keywords: animal welfare, egg quality parameters, heat stress, intestinal integrity, methionine metabolism, thermoregulation, tissue morphology
Citation: Souza RG, Maia MIL, Anjos Lima LAd, Morais LKdC, Santos SGCGd, Veríssimo TNS, Santos JDCd, Nascimento PVd, Soares KO, Costa FGP, Guerra RR, Carvalho LRRA and Saraiva EP (2025) DL-methionine supplementation in Japanese quails under different thermal conditions. Front. Anim. Sci. 6:1671042. doi: 10.3389/fanim.2025.1671042
Received: 22 July 2025; Accepted: 11 September 2025;
Published: 25 September 2025.
Edited by:
Assar Ali Shah, Jiangsu University, ChinaReviewed by:
Ali Hassan Nawaz, Nanjing Agricultural University, ChinaSadid Al Amaz, University of Georgia, United States
Abdurrahman Abdulkadir, Federal University, Nigeria
Copyright © 2025 Souza, Maia, Anjos Lima, Morais, Santos, Veríssimo, Santos, Nascimento, Soares, Costa, Guerra, Carvalho and Saraiva. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lucas Rannier Ribeiro Antonino Carvalho, bHVjYXMuY2FydmFsaG9Aa2kuc2U=