 Marina Galvez-Lopez1
Marina Galvez-Lopez1 Gema Romero1*
Gema Romero1* Carlos Alfredo Sandoval-Castro2
Carlos Alfredo Sandoval-Castro2 Elena Perez3Martín Rodriguez4Cristòfol Peris4Jose Ramón Díaz1
Elena Perez3Martín Rodriguez4Cristòfol Peris4Jose Ramón Díaz1- 1Institute of Agri-Food and Agro-Environmental Research (CIAGRO-UMH), Miguel Hernandez University of Elche, Orihuela, Spain
- 2Faculty of Veterinary Medicine and Animal Science, Autonomous University of Yucatan, Merida, Mexico
- 3Technical Support Service for Teaching and Research (SATDI-UMH), Miguel Hernandez University of Elche, Orihuela, Spain
- 4Institut de Ciència i Tecnologia Animal, Universitat Politècnica de València, València, Spain
Grape pomace is the winery’s main byproduct, with an estimated 260 million tons generated annually. It includes high concentrations of grape-derived bioactive compounds that may exert beneficial effects on oxidative and biochemical status, and productive performance in the animals, being an environmentally and economically sustainable alternative to conventional raw materials. The effect of incorporating different proportions (0, 5, 10, and 15% DM) of white grape pomace silage (WGP) into isoenergetic and isoproteic diets on the performance of dairy goats was evaluated. Eighty-eight Murciano-Granadina goats were selected and split into 8 homogeneous batches of 11 goats each, according to parity, body weight, milk yield and SCC. After a pre-experimental sampling, every diet was randomly assigned to two batches, and the feeding trial lasted 8 weeks. Biweekly body weight was recorded, and individual milk samples were collected to evaluate both yield and gross composition. The intake of dry and fresh matter and water was recorded weekly at the batch level over two consecutive days. Physiological and oxidative stress, assessed through hemogram, serum biochemical profile, and plasma antioxidant activity (ABTS, FRAP, SOD), were evaluated at the goat level on the pre-experimental and the final experimental samplings. Although a significant decrease in body weight was observed in the 15-WGP treatment, it was not translated into differences in milk yield or composition. Feed efficiency (fat and protein corrected milk/dry matter intake) was higher in WGP diets compared to the control. The 10-WGP and 15-WGP treatments consumed significantly less water than the other treatments. Hematocrit (HCT) increased more markedly in the control than in the WGP-diets treatments. At the last sampling, GLU concentrations in the 15-WGP treatment remained significantly lower than in the other treatments. None of the changes observed in the hemogram or serum biochemical profile were considered physiologically relevant. No dietary effects were detected on the oxidative stress of the animals. In conclusion, the ensiled white grape pomace inclusion in dairy goat diets at levels of up to 15% of dry matter appears to be feasible without inducing short-term adverse effects, provided the diets are nutritionally balanced. However, further research is recommended to confirm long-term safety and performance outcomes.

Graphical Abstract.
1 Introduction
Global demand for animal-sourced foods continues to rise, amid the depletion of agri-food sector inputs (FAO, 2023). Given the limited availability of arable land, this trend has intensified competition between livestock production and human food demand (Benoit and Mottet, 2023). Meanwhile, the agro-industrial sector is generating large quantities of fibrous residues and by-products, representing a considerable loss of high-value biomass and nutrients (Kilama et al., 2023), producing 57 million tons in the European Union (Malenica et al., 2023). Reintegrating plant-based by-products into livestock feeding systems offers a viable strategy to enhance the sustainability of the agri-food chain. Their inclusion in ruminant diets can reduce the environmental footprint of livestock systems, lower feed costs, and promote nutrient recycling, while also supplying bioactive compounds to animals. This strategy has gained increasing relevance in recent years, particularly within global and European efforts to develop sustainable, energy-efficient food systems aligned with circular economy principles. (Marques et al., 2022).
Grape pomace is the primary residue from winemaking, estimated to be around 25 kg of by-product for every 100 kg of grapes processed (Taladrid et al., 2019), leading to amounts that can reach 1200 tons per year in the main wine-producing countries (Bordiga et al., 2015). Globally, approximately 260 million tons of grape pomace are generated annually. This material is characterized by high concentrations of grape-derived bioactive compounds, such as flavonoids, tocopherols, antioxidants and polyunsaturated fatty acids (Galanakis, 2012; Tangolar et al., 2011; Ovcharova et al., 2016; Harbeoui et al., 2017). Due to its unique chemical profile, grape pomace has attracted interest as a functional component of ruminant diets. Several studies suggest that its inclusion may enhance rumen function by modulating the gut microbiota, reducing methane emissions and protein degradation, and increasing post-ruminal protein flow (Patra and Saxena, 2011). Moreover, the most well-known property of polyphenolic compounds is their antioxidant potential, which protects cells against oxidative stress. They also exhibit health-promoting effects such as anti-inflammatory, antibacterial, and anticoagulant activities, and contribute to improved vascular function (Nudda et al., 2015; Qiu et al., 2023).
Grape pomace (Martins Flores et al., 2021) and other agro-industrial by-products (Monllor et al., 2023; Dentinho et al., 2023) have demonstrated that, when diets are carefully formulated to meet nutritional requirements, such inclusion does not impair animal performance or product quality. Furthermore, multiple studies support the economic viability of these feeding strategies (Molosse et al., 2023; Baker et al., 2024). Ruminants are particularly adapted to diets rich in fibrous material and slowly digestible bioactive compounds, features common to plant-based by-products (Makkar, 2003; Frutos et al., 2004; Patra and Saxena, 2011). Goats, in particular, exhibit greater tolerance to tannins compared to sheep or cattle, attributed to salivary proteins with a high affinity for tannin binding (Ventura-Cordero et al., 2017). This mitigates the anti-nutritional effects observed in other species, such as intake depression or impaired rumen function (Correddu et al., 2020). Ruminants play a key role in closing nutrient loops within circular food systems by converting low-value biomass—such as agro-industrial by-products—into high-quality animal products. In addition to their ecological role, ruminants make a substantial contribution to global food security through the production of highly digestible animal protein with elevated biological value, particularly in regions with limited agricultural resources and vulnerable food systems (Mottet et al., 2017). The balanced essential amino acid profile of ruminant-derived milk and meat further underscores their nutritional value in human nutrition (Castillo et al., 2019).
In sheep, a more extensively studied species, the inclusion of 300 g/day of grape seed pomace did not affect feed intake (Correddu et al., 2015). Similarly, Manso et al. (2016) reported no impact on lactational performance or body condition when up to 10 g/100 g DM of the grape pomace was offered. Bennato et al. (2022) noted that supplementing 10% grape pomace had no effect on gross milk composition but increased the content of monounsaturated fatty acids (MUFA) in the milk of Assaf ewes intended for cheese production. In lambs, grape pomace silage caused a linear reduction in dry matter intake and, although a decrease in average daily gain was observed, feed conversion efficiency remained comparable to the control group (Martins Flores et al., 2020). Moreover, dietary supplementation with grape pomace has been shown to enhance antioxidant defenses in lambs, as evidenced by increased catalase and glutathione activity, along with reduced lipid and protein oxidation markers in blood and tissues (Kafantaris et al., 2016). In Saanen goats, the inclusion of 150 g/kg of dry grape by-products had no significant effects on feed intake, digestibility, blood metabolites, or milk fatty acid composition (Badiee Baghsiyah et al., 2023). However, few studies have evaluated the effects of grape pomace on dairy goats.
Based on the nutritional and environmental potential of grape pomace, the present study evaluated the effects of incorporating different levels of white grape pomace silage (WGP) into the diets of lactating dairy goats. The parameters assessed included individual milk yield, gross milk composition (including somatic cell count), body weight, hematobiochemical profile, and oxidative stress biomarkers. The aim was to identify an inclusion level that supports productive performance and physiological stability, while contributing to sustainable feeding strategies.
2 Materials and methods
2.1 Animals and facilities
The study was carried out on the dairy goat herd located at the teaching and experimental farm of Miguel Hernandez University, Spain. The experimental period lasted from late April (pre-experimental sampling) through the end of July. This farm houses a herd of Murciano-Granadina goats, housed in pens with straw bedding, providing each animal with 1.5 m² of usable individual space in a free-stall system, 35 cm of linear feeding space per animal, and ad libitum access to water. The animals were fed twice daily, at 9:00 h and 15:00 h, and milked once a day, prior to the morning feeding, using a Casse milking parlor (2 × 12 × 12, GEA, Germany), in accordance with standard regional practices. This study was approved by the Office of Responsible Research at Miguel Hernandez University (code UMH.DTA.JDS.04.22).
2.2 Experimental design
From an initial group of 120 goats, at five weeks postpartum stage of lactation, and feeding a conventional diet, 88 animals were selected based on parity, body weight (37.70 ± 5.48 kg), milk yield (2.89 ± 0.84 kg/day), and LSCC (2.96 ± 3.30–103 cells/mL). The animals were distributed into 8 homogeneous batches, ensuring balance across the selection criteria. Following a one-week adaptation period to their new groups, a pre-experimental sampling (sampling 0) was conducted, including all the variables analyzed. After the pre-experimental sampling, every treatment (or diet) was randomly assigned to 2 batches: Control (conventional diet without by-products), 5-WGP (diet including 5% WGP silage on a dry matter basis), 10-WGP (diet including 10% DM of WGP silage), and 15-WGP (diet including 15% DM of WGP silage). After a two-week dietary adaptation period, four biweekly samplings were performed to assess body weight, individual milk yield, and milk gross composition. Only one additional individual blood sampling was done at the end of the experiment. Feed and water intake were monitored weekly for two consecutive days at the batch level.
All diets were formulated according to the recommendations of Fernandez et al. (2005) for goats producing 2.5 kg of milk/day, ensuring isoenergetic and isoproteic rations tailored to their production level. Animals were fed fixed amounts twice daily, with no ad libitum access. Details regarding the ingredients, chemical composition, and daily feed offered are presented in Table 1. The WGP silage used in the study was sourced from the quality and stability trials of this agroindustrial by-product conducted by Galvez-Lopez et al. (2025).

Table 1. Ingredients and chemical composition of experimental diets.
2.3 Variables analyzed
The composition of the diets (Table 1) was determined according to Association of Official Analytical Chemists (1999) methods: dry matter (DM, g/kg; method 930.5), organic matter (OM, g/kg DM; method 942.05), ether extract (EE, g/kg DM; method 920.39), crude protein (CP, g/kg DM; method 984.13), and total sugars (g/kg DM; method 974.06). Neutral detergent fiber (NDF, g/kg DM), acid detergent fiber (ADF, g/kg DM), and acid detergent lignin (ADL, g/kg DM) were analyzed according to Van Soest et al. (1991). Starch content was determined using the polarimetric method of Ewers (ISO, 1997). Total polyphenol content (PT, g GAE eq./kg DM) was analyzed using the Folin-Ciocalteu method as described by Kim et al. (2003). The mineral content in the diets was determined using an Inductively Coupled Plasma Mass Spectrometer (ICP-MS; Agilent 7700x, Santa Clara, CA, USA) following acid digestion of the samples according to Gonzalez Arrojo et al. (2016). Apparent in vitro dry matter digestibility (IVDMD, g/kg DM) was assessed using the method of Menke and Steingass (1988), with slight modifications recommended by Yañez-Ruiz et al. (2016). The degradability of NDF (IVDNDF, g/kg DM) was determined based on digested material according to Van Soest et al. (1991). The proportion of volatile fatty acids (VFAs, g/kg DM) —acetate, propionate, butyrate, isobutyrate— was measured using the method proposed by Feng-Xia et al. (2013), employing high-performance liquid chromatography (HPLC 1200 Agilent, Santa Clara, CA, USA) with a Supelcogel C610H column (30 cm × 7.8 mm ID, Saint Louis, MO, USA).
During each sampling, animals were weighed using a precision scale attached to the handling chute (Accurex RX Fox, Gram Group, Barcelona, Spain) with an accuracy of 100 g, to assess body weight (kg) evolution throughout the experiment. Individual milk yield (kg/day) was recorded daily using a Lactocorder® device (Lactocorder, Balgach, Switzerland). Milk macrocomposition—including fat, crude protein, lactose, useful dry matter (UDM), non-fat dry matter (NFDM), and milk urea—was analyzed using mid-infrared spectroscopy (CombiFossTM 7 DC, Foss, Hillerød, Denmark). Somatic cell count (SCC, 10³ cells/mL) was determined via flow cytometry (CombiFossTM 7 DC, Foss, Hillerød, Denmark). Fat-Corrected Milk (FCM 3.5%) was calculated following Gravert (1987): FCM (3.5%) = 0.433 × milk yield (kg/day) + 16.218 × fat yield (kg/day). Fat and Protein-Corrected Milk (FPCM) was estimated using Schau and Fet (2008): FPCM = milk yield (kg/day) × (0.337 + 0.116 × Fat (%) + 0.06 × Protein (%)).
Feed intake (FMI and DMI) and water intake (WI) were recorded weekly at the batch level for two consecutive days each sampling week. Feed intake was calculated as the difference between the amount of feed offered and refused, expressed on both a fresh matter (FM) and dry matter (DM) basis. The DM content was determined by drying representative samples of offered and refused rations at 105 °C for 48 hours. Water intake was measured by difference using individual water meters (SV-RTK E4® Flow Systems, Gilmonde, Portugal) installed in each pen.
Fasting blood samples (8:00 am) were collected only at pre-experimental and final samplings. Samples were collected from the jugular vein using an Eclipse™ needle (BD Vacutainer, Franklin Lakes, NJ, USA) and stored in three test tubes (BD Vacutainer, Franklin Lakes, NJ, USA): EDTA K2 (4 mL) for hematological analysis, Serum CAT (10 mL) with clot activator for serum biochemical profile assessment, and LH Lithium Heparin (10 mL) for oxidative status evaluation. Serum and plasma obtained from samples collected in Serum CAT and LH tubes, respectively, were stored at −80°C and analyzed within one month of collection. Hematology was determined within one hour after blood sampling using an Element HT5 analyzer (SCIL VET, Tudela, Spain) calibrated for a female goat: white blood cells (WBC), lymphocytes (LYM), monocytes (MON), neutrofiles (NEU), red blood cells (RBC), hemoglobin (HGB), hematocrit (HCT) were analyzed, and the ratios neutrophils /lymphocytes (NLR), and monocytes /lymphocytes (MLR) were calculated. Serum biochemical profile was conducted using the rapid diagnostic disk “Diagnostic II” for Element RC3X (SCIL VET, Tudela, Spain); variables analyzed were: albumine (ALB), alanine aminotransferase (ALT), total bilirubin (TBIL), blood urea nitrogen (BUN), creatinine (CREA), total protein (TP), globuline (GLOB), glucose (GLU), cholesterol (CHOL), Ca, P, Na, K. Oxidative status was evaluated using total antioxidant capacity assay —FRAP Oyaizu (1986) and ABTS Leite et al. (2011)—. Specific antioxidant capacity was determined via superoxide dismutase (SOD) activity assay (DetectX® Superoxide Dismutase Activity Kit, Arbor Assays, Ann Arbor, MI, USA).
2.4 Statistical analysis
The SCC values were transformed into base ten logarithms for statistical analysis. The statistical evaluation of variables measured at the individual level, excepting those analyzed on blood/plasma/serum samples, was conducted using a mixed linear model (PROC GLIMMIX, SAS v9.4), considering the effect of the covariate (pre-experimental sampling), diet, samplings, the interaction (diet × sampling), and batch nested within diet, according to the following equation:
where Y is the dependent variable, µ is the intercept, Di represents the fixed effect of diet (i = Control, 5-WGP, 10-WGP, 15-WGP), Sj is the fixed effect of sampling (j = 1, 2, 3, 4), Di × Sj represents the interaction between diet and sampling (16 levels), covYo accounts for the effect of Y in the pre-experimental sampling, Bk(Di) is the batch nested within diet, and e is the residual error.
For variables analyzed on blood/plasma/serum samples, the model included the fixed effects of diet (i = 4 levels), sampling (2 levels, j = 0, 4), their interaction (8 levels), and batch (diet).
For both scenarios, the animal was considered a random effect (Ak), and e represents the residual error. A compound symmetry covariance structure was employed to account for the correlation of repeated measures, based on the lowest AIC and BIC values.
For variables measured at the batch level (WI, DMI, FMI), a general linear model (PROC GLM, SAS v9.4, 2022) was used, considering the effect of diet, sampling, and the interaction, according to the following equation:
where Y is the dependent variable, μ is the intercept, Di represents the fixed effect of diet (i = Control, 5-WGP, 10-WGP, 15-WGP), Tj is the fixed effect of sampling (k = 0, 1, 2, 3, 4), Di × Tk represents the interaction between both effects (20 levels), and e is the residual error.
Least squares means were calculated to interpret differences between levels of the fixed effects. The null hypothesis was rejected at P< 0.05, indicating statistically significant differences between levels.
3 Results
3.1 Feed intake, body weight, and milk yield
The inclusion of WGP silage did not result in significant differences in most variables associated with productive performance (Table 2). The evolution of DMI throughout feeding trial showed no significant differences among diets excepting at the final sampling point, when DMI in the 15-WGP group (1.99 ± 0.03 kg DM/day) was significantly lower than in the Control (2.05 ± 0.03 kg DM/day) and 5-WGP (2.10 ± 0.03 kg DM/day) groups (Figure 1). In contrast, FMI was significantly higher in the 15-WGP group than the Control group from sampling 1 through the end of the trial, with mean values of 2.59 ± 0.01 vs. 2.32 ± 0.01 kg FM/day (Figure 1). From the beginning of the treatments, water intake exhibited a divergent pattern compared to FMI. It was significantly lower in samplings 2 and 3 for 10-WGP (5.53 ± 0.17 L/day) and 15-WGP (5.60 ± 0.17 L/day) diets, and in sampling 4 for the 15-WGP group (7.33 ± 0.17 L/day), compared to 0-WGP and 10-WGP diets (Figure 1). Diet had a significant effect (P = 0.02) on BW, with a significantly lower average in the 15-WGP group (36.44 ± 0.47 kg) compared to animals fed the other three diets (Table 2). BW in animals receiving the 15-WGP diet decreased significantly from sampling 1 (39.53 vs. 36.10 kg). This difference among diets persisted until the end of the trial (Figure 1).

Table 2. Statistical analysis results (F and Pvalue, andd Least Square Means ± SEM) for main productive performance traits by tested diets.

Figure 1. Dry matter intake (A), fresh matter intake (B), body weight (C), and water intake (D) recorded throughout the experiment (Least Square Means ± SE, dotted lines indicate the adaptation period).
Regarding milk yield (MY), no significant differences were detected among diets, although a sampling effect was observed (P = 0.001) (Table 2). Milk yield (Figure 2), along with Fat-Corrected Milk (FCM) and Fat and Protein-Corrected Milk (FPCM) (Figures 2), declined as the experimental period progressed. In contrast, feed efficiency (FPCM/DMI) only declined significantly (P< 0.001) in the Control group toward the end of the experimental period (Figure 2).

Figure 2. Milk yield (A), Fat corrected milk (B), Fat and Protein corrected milk (C), and feed efficiency (FPCM/DMI) (D) recorded throughout the experiment (Least Square Means ± SE; dotted lines indicate the adaptation period).
As illustrated in Figure 3, variations in milk composition across dietary treatments remained minimal during the trial period and did not reach statistical significance for the majority of measured parameters. The concentration of main milk components fluctuated throughout the experiment, with sampling consistently showing significant effects (P< 0.001); thus, the diet × sampling interaction did not show significant effects. The concentrations of fat, crude protein, lactose, NFDM, and UDM were slightly lower at the end of the trial for all diets (Figure 3). Only milk urea was significantly affected by diet (P = 0.02). The 10-WGP diet resulted in significant lower milk urea concentrations compared to the other diets, although the differences were not considered relevant (Table 2). No significant effects of diet or sampling were observed for LSCC (Table 2).

Figure 3. Milk composition—Fat (A); Crude protein (B); Lactose (C); No-fat dry matter (NFDM) (D); Useful dry matter (UDM) (E)—recorded throughout the experiment (Least Square Means ± SE).
3.2 Hemogram, serum biochemical profile, and oxidative stress
Table 3 presents the evolution of the hemogram throughout the feeding trial, illustrating the interaction between diet and sampling time. Neither diet nor sampling time exerted a significant effect on white blood cell profile (WBC, LYM, MON, NEU, NLR, MLR). The overall mean WBC count across diets was 15.27 ± 0.96 × 109/L (control), 15.31 ± 0.96 × 109/L (5-WGP), 15.62 ± 0.96 × 109/L (10-WGP), and 15.10 ± 0.96 × 109/L (15-WGP). The highest NLR (neutrophil-lymphocyte ratio) and MLR (monocyte-lymphocyte ratio) values were observed in the 15-WGP (2.95 ± 0.29 and 0.05 ± 0.01, respectively), while the lowest corresponded to the Control treatment (2.41 ± 0.29 and 0.04 ± 0.0). Diet had a significant effect on HCT, while sampling time also influenced HCT, as well as RBC and HGB levels. Mean RBC counts by diet were as follows: 16.85 ± 0.38 × 10¹²/L (control), 16.30 ± 0.38 × 10¹²/L (5-WGP), 17.37 ± 0.38 × 10¹²/L (10-WGP), and 16.83 ± 0.38 × 10¹²/L (15-WGP). The significant dietary effect on HCT was already evident at sampling 0, with the 10-WGP group showing higher HCT values (25.52 ± 0.63%) compared to the remaining groups. At sampling 4, Control HCT (25.48 ± 0.63%) was significantly higher than its baseline value (23.70 ± 0.63%, sampling 0), thereby reducing the difference with the 10-WGP group, which exhibited no significant change over time (25.52 ± 0.63% at sampling 4) as the other WGP treatments.
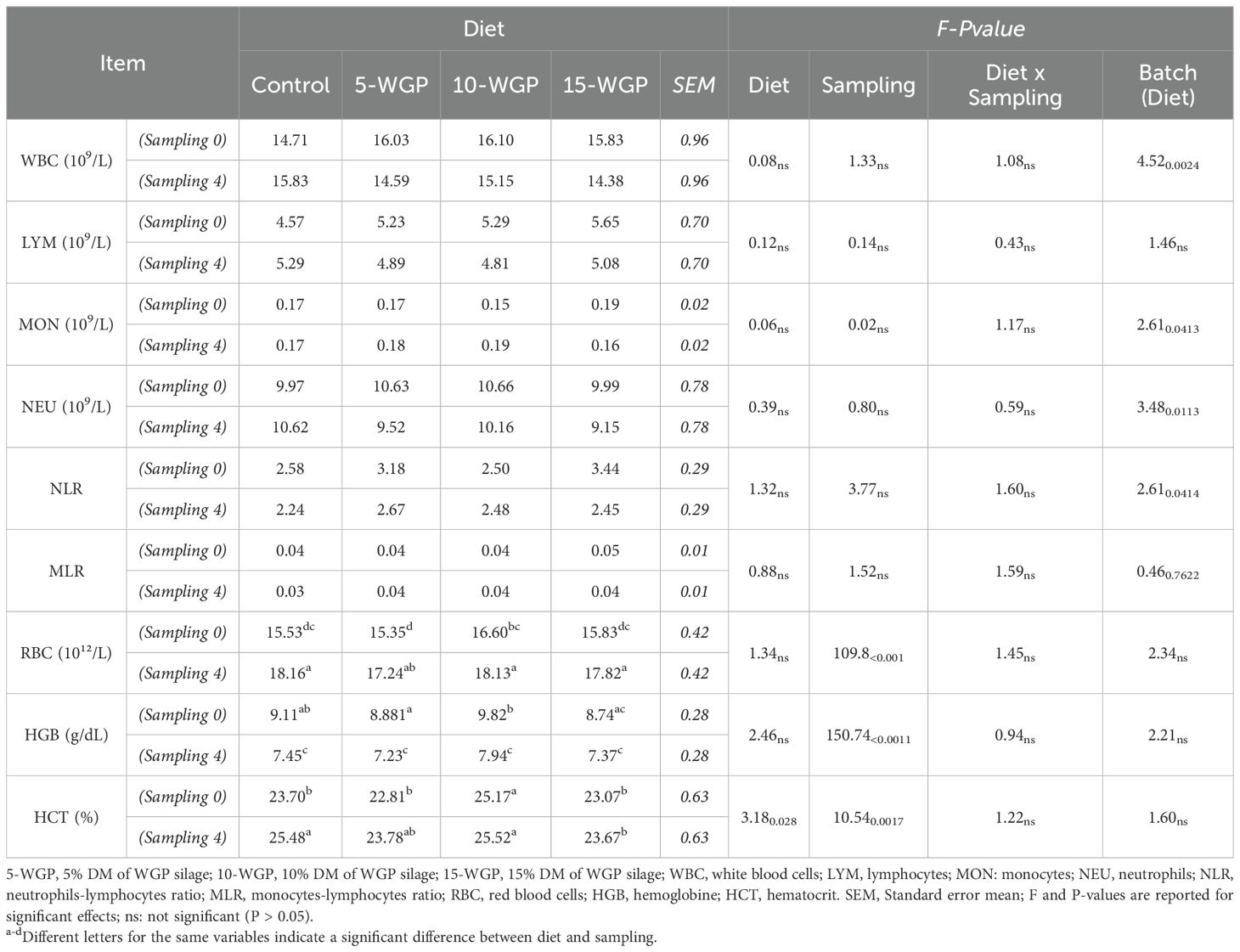
Table 3. Hemogram evolution throughout the experiment: comparison of mean values across diets and sampling (Least Square Means ± SEM), and statistical effects of diet, sampling time, and their interaction (F and P-value).
Regarding the serum biochemical profile, slight differences were observed among groups (Table 4). Diet had a significant effect on BUN and GLU. Sampling time significantly influenced ALB, ALT, TBIL, CREA, BUN and GLU. In addition, a significant interaction between diet and sampling time was observed for GLU. Accordingly, BUN and GLU were plasma metabolites with a significant effect of Diet, although dietary differences on BUN were already evident at sampling 0, before the treatments started, and at sampling 4, BUN decreased significantly across all dietary groups, revealing the same pattern of differences seen at sampling 0. At sampling 0, fasting glucose concentrations were significantly higher in the Control and 5-WGP treatments than in the 10-WGP and 15-WGP treatments. By sampling 4, GLU increased significantly across all treatments relative to baseline. Nonetheless, the 10-WGP increased significantly GLU (difference of 19.22 mg/dL) in higher concentrations than the others, whereas 15-WGP increased (difference of 10.80 mg/dL) in similar proportions as 0-WGP (difference of 7.73 mg/dL) and 5-WGP (difference of 7.63 mg/dL). At sampling 4 GLU concentrations in the 15-WGP group were significantly lower than the others.

Table 4. Serum biochemical profile evolution throughout the experiment: comparison of mean values (Least Square Means ± SEM) across diets and samplings, and statistical effects of diet, sampling time, and their interaction (F and P-value).
No significant differences in oxidative status were observed by diet, as evaluated by reduction power using the FRAP and ABTS assays. Similarly, the diet had no significant effect on specific antioxidant capacity, as evaluated by the SOD activity assay. However, sampling had a significant effect on all antioxidant assays, with antioxidant capacity at sampling 4 consistently higher than observed at sampling 0 values, without significant interaction with diet (Table 5).

Table 5. Evaluation of oxidative stress throughout the feeding trial: comparison of mean values across diets (Least Square Means ± SEM), and statistical effects of diet, sampling time, and their interaction (F and P-value).
4 Discussion
Dry matter intake (DMI) of the diets containing WGP silage aligned with the expected values for lactating Murciano-Granadina goats five weeks postpartum, supporting an average milk yield of 2.89 L/day—within the standard range of 2.50–2.70 L/day (Jarrige et al., 1981). The DMI was similar to the consumption reported for this breed under comparable physiological conditions (Fernandez et al., 2005; Ibañez et al., 2016; Monllor et al., 2020). Although Martins Flores et al. (2020) reported a dose-dependent intake response when increasing the inclusion levels (20%, 37.5%, and 50% DM) of grape pomace silage in lambs, our findings suggest that a moderate inclusion of WGP silage does not adversely affect dry matter intake in lactating goats. The observed reduction in water intake among goats receiving 10% and 15% WGP diets may be attributed to the higher forage moisture intake associated with the inclusion of silage. As described by Van Soest (1994a), moist feeds such as silages or fresh forages can partially satisfy free water requirements under temperate environmental conditions, an aspect particularly relevant in thermal stress scenarios or when access to water is limited (Van Soest, 1994b). Moreover, the reduction of approximately 0.5 L of water per animal per day represents a meaningful saving of this resource, which is especially valuable in dry-climate regions where water availability is a limiting factor for livestock production. This consideration becomes even more relevant given that livestock farming increasingly competes with agriculture for access to water, highlighting the importance of efficient resource use in integrated production systems (Wisser et al., 2024).
Despite a significant reduction in body weight observed in the 15-WGP goats, no detrimental effects were detected on milk yield performance (MY, UDM, FCM, FPCM), milk composition, or feed conversion ratios (FPCM/DMI). This response might be supported by the improved in vitro NDF digestibility (IVNDFD) and the more glucogenic fermentation profile -in the in vitro assay- which likely enhanced energy supply for milk synthesis (Van Soest, 1994b; Chilliard et al., 2003), without increasing feed intake. Still, the physiological prioritization of lactation (homeorhesis) in ruminants may have constrained energy allocation to body tissue maintenance (Chilliard et al., 2000), particularly in heavier animals—such as those in the 15-WGP group—with inherently greater maintenance requirements. The moderate energy deficit suggested by the decline in body weight could thus reflect nutrient partitioning favoring milk production, supported by rumen efficiency and mobilization of body reserves (Aguerre et al., 2011), showcasing the high adaptability of the Murciano-Granadina breed to intensive milk production systems.
Towards the end of the experimental period, the upward trends in MY and FPCM/DMI observed in WGP diets, along with signs of weight gain recovery in the 15-WGP group (during the latter half of the study period), suggest that long-term dietary inclusion of WGP silage may enhance performance in lactating goats. Previous studies have already demonstrated the positive influence of grape pomace on productive performance in small ruminants when included at 2% of daily intake, either in the form of grape pomace meal (Alba et al., 2019) or grape pomace cake (Antunovic et al., 2024). WGP silage inclusion did not significantly alter the gross composition of milk (Fat, CP, NFDM, UDM), nor LSCC, indicating that milk from WGP silage diets maintained a quality profile comparable to the Control diet. In the present study, the inclusion of WGP silage in the diet was tightly controlled (up to 15%), resulting in a limited impact on diet composition—specifically, an increase of only 0.50 g/100 g in fat content of diet—and no significant effect on FCM was observed. This finding is consistent with Nudda et al. (2015) and Manso et al. (2016), who also observed no increase in milk fat content when 300 g/day of grape seeds and up to 100 g/kg of grape pomace were incorporated, respectively, into diets for lactating sheep. In contrast, Alba et al. (2019) included 2% of grape pomace meal in the daily intake, which contained 26 g/kg DM of fat, and reported an increase of 5.8 g/kg in milk fat content in sheep. Lower milk urea concentrations in the 10-WGP group do not represent a relevant effect and could be attributed to enhanced nitrogen utilization, likely due to improved synchrony between degradable protein and fermentable carbohydrates in the rumen. This group exhibited the lowest initial body weight —no significant— and the greatest weight gain during the experiment, suggesting lower maintenance requirements (National Research Council, 2007) and an anabolic status favoring tissue nitrogen retention over excretion (Raggio et al., 2006). Consequently, hepatic urea synthesis and subsequent milk urea secretion may have decreased, not due to protein deficiency, but to more efficient nitrogen partitioning (Chilliard et al., 2003; Cannas et al., 2007). Importantly, all productive parameters observed were in line with those reported by MAPA (2022) for this breed under comparable conditions.
The hemogram and serum biochemical profile measured throughout the study remained within the physiological reference ranges established for dairy goat (Kaneko et al, 1997). The minor fluctuations observed between the beginning and end of the experimental period were consistent with expected physiological adaptations associated with the progression of lactation. The differences detected between dietary treatments cannot be attributed solely to the direct effect of the diets themselves, but rather to the interplay between nutrient intake, fermentative composition of the diet and the transient metabolic adjustments required to sustain homeostasis under evolving physiological demands.
Notably, GLU increased significantly in the 10-WGP diet, suggesting elevated propionate production and enhanced hepatic gluconeogenesis (Wang et al., 2024), though its magnitude might also hint at temporary dysregulation of peripheral insulin sensitivity under a high energy demand context, such as reaching peak lactation in the postpartum period (Chavez et al., 2009). Conversely, the 15-WGP group showed a more stable GLU profile with lower final GLU concentrations, which may reflect a more efficient utilization of ruminal propionate (Chilliard et al., 2003; Cai et al., 2018). This observation further supports the previously mentioned hypothesis of a fermentative profile with a predominant glucogenic orientation in among animals with greater WGP intake. Parallel reductions in blood urea nitrogen (BUN) across all groups suggest improved nitrogen use efficiency as lactation advanced, likely due to greater synchrony between dietary nitrogen and fermentable carbohydrates (Donkin, 2016). These synergistic relationships between nitrogen and fermentable carbohydrates were further supported by the in vitro ruminal fermentation results, where WGP diets enhanced fiber digestibility (IVNDFD) and increased total VFA production, particularly propionate, reflecting more efficient fermentation and a greater availability of glycogenic substrates (Zhen et al., 2023; Shi et al., 2025). A similar enteric VFA fermentation profile was also reported by Correddu et al. (2015) during in vitro digestion of diets containing grape seed. In vivo digestibility trials in wethers (Juraček et al., 2021) and lactating goats (Do Nascimento et al., 2025) likewise reported an increase in NDF degradability in diets supplemented with dehydrated grape pomace. Overall, moderate to high WGP inclusion—especially at 15%—appeared to support more efficient nutrient partitioning, lower hepatic nitrogen load, and improved metabolic regulation in mid-lactation goats. In addition, the higher total phenol content observed in WGP diets may have contributed to enhanced fiber digestibility and nitrogen utilization. Studies with other vegetables rich in phenolic compounds (Singh et al., 2022; Huang et al., 2023) have already demonstrated that these bioactive compounds modulate ruminal microbial activity, promoting better NDF degradability and reducing nitrogen excretion, thereby reinforcing the metabolic benefits associated with WGP inclusion.
No diet-related effects were detected for blood cell counts (WBC, RBC) or hemoglobin (HGB) levels, consistent with previous reports in dairy goats fed with 10% DM of grape seed cake (Antunovic et al., 2024). Similarly, no alterations in hematological profiles were observed in lambs fed with ensiled grape pomace (Martins Flores et al., 2020), nor in sheep receiving grape pomace flour (Alba et al., 2019). However, hematocrit (HCT) showed a different pattern, although initial differences between diets were already present, the maintenance of its concentration in WGP diets—compared to the increase observed in the control diet—could be attributed to a dietary effect. Given that RBC increased across all treatments, the HCT rise in control animals likely reflects plasma volume contraction—suggesting a relative dehydration—which did not occur in WGP groups. Since HCT is considered an indirect marker of hydration status (Silanikove, 1994), the absence of such a rise in WGP-fed goats reinforces the notion that diets rich in moisture can positively contribute to fluid homeostasis (Van Soest, 1994a). Given the geographical location (Spanish Mediterranean region) and the timing of the experiment—coinciding with a seasonal rise in ambient temperatures compared to the first quarter of the year—this hypothesis gains relevance.
Finally, no significant effects of WGP silage inclusion were observed on oxidative stress biomarkers in plasma. The relatively short experimental period and moderate inclusion levels may have limited the expression of antioxidant responses potentially linked to the bioactive compounds present in WGP (Harbeoui et al., 2017). Nonetheless, the slight upward trend observed toward the end of the experiment—particularly in ABTS and SOD levels at higher WGP diet inclusions—appears promising for reassessing the long-term antioxidant potential of WGP. Antunovic et al. (2024) reported increased SOD activity in dairy goats fed diets containing 5% and 10% grape seed cake over a comparable study duration. A similar effect had previously been observed in sheep supplemented with 20 g/kg of grape pomace flour for 15 days (Alba et al., 2019).
As evidenced by the studies referenced throughout this discussion, the dietary inclusion formats of grape byproducts in ruminant nutrition are diverse. There is a clear predominance of studies including dry grape pomace presentations—such as flours and extracts—over silage applications. However, it is also evident that inclusion levels for flour-based forms are substantially lower than those achieved with grape pomace silage. While both formats may provide access to the nutraceutical properties of this encouraging feed ingredient, the use of silage allows for higher inclusion rates and greater utilization of the overabundant by-product, thereby offering a more effective strategy for valorizing agro-industrial byproducts. This aligns more closely with the principles of sustainable production systems, particularly in the context of circular economy approaches to agri-food production.
The main findings in the present research indicate that the inclusion of WGP silage in dairy goats diets did not elicit adverse effects on productive performance, physiological or oxidative status. Encouragingly, the results suggest a favorable potential for enhancing dietary efficiency when WGP is incorporated into well-balanced formulations aligned with the animals’ nutritional requirements.
The absence of relevant effects in productive performance and metabolic status among diets supports the feasibility of including white grape pomace silage in the diets of Murciano-Granadina dairy goats at the tested inclusion levels. The findings highlight a promising outlook regarding the potential effects of sustained inclusion rates at least between 10% and 15% of dietary dry matter across a complete lactation period. These findings support the inclusion of white grape pomace silage as a dietary ingredient for ruminants, specifically dairy goats, provided that its incorporation in nutritionally balanced diets and aligned with the energy requirements dictated by the animal's physiological state. Such an approach offers a sustainable strategy for managing winery by-products and closing nutrient loops within circular economy systems by converting low-value biomass into high-quality animal products. Nevertheless, additional studies involving longer-term experiments are needed to substantiate these results.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The animal studies were approved by Office of Responsible Research at Miguel Hernandez University (code UMH.DTA.JDS.04.22. The studies were conducted in accordance with the local legislation and institutional requirements. Written informed consent was obtained from the owners for the participation of their animals in this study.
Author contributions
MG-L: Data curation, Formal analysis, Investigation, Methodology, Writing – original draft, Writing – review & editing. GR: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Project administration, Supervision, Writing – review & editing. CS-C: Methodology, Writing – review & editing. EP: Investigation, Writing – review & editing. MR: Methodology, Writing – review & editing. CP: Funding acquisition, Project administration, Writing – review & editing. J-RD: Conceptualization, Funding acquisition, Investigation, Methodology, Project administration, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research is part of the AGROALNEXT program, supported by the Ministry of Science and Innovation (MCIN), with funding from the European Union through NextGeneration EU (PRTR-C17.I1) and the Generalitat Valenciana (AGROALNEXT/2022/062). The funders had no role in the design of the study, in the collection, analyses or interpretation of data, in the writing of the manuscript, or in the decision to publish the results.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abbreviations
ABTS: 2.2′-azino-bis(3-ethylbenzothiazoline-6-sulfonate)
ADF: Acid Detergent Fiber
ADL: Acid Detergent Lignin
ALB: Albumin
ALP: Alkaline Phosphatase
ALT: Alanine Aminotransferase
BW: Body Weight
BUN: Blood Urea Nitrogen
CHOL: Cholesterol
CP: Crude Protein
CREA: Creatinine
DM: Dry Matter
DMI: Dry Matter Intake
EE: Ether Extract
FCM: Fat-Corrected Milk
FPCM: Fat and Protein-Corrected Milk
FRAP: Ferric Reducing Antioxidant Power
GLOB: Globulin
GLU: Glucose
HCT: Hematocrit
HGB: Hemoglobin
IVDMD: In Vitro Dry Matter Digestibility
IVNDFD: In Vitro Neutral Detergent Fiber Digestibility
IVOMD: In Vitro Organic Matter Digestibility
LSCC: Log10 Somatic Cell Count
LYM: Lymphocytes
ME: Metabolizable Energy
MLR: Monocytes-Lymphocytes Ratio
NDF: Neutral Detergent Fiber
NFDM: Non-Fat Dry Matter
NLR: Neutrophils-Lymphocytes Ratio
OM: Organic Matter
RBC: Red Blood Cells
SCC: Somatic Cell Count
SEM: Standard Error of the Mean
SOD: Superoxide Dismutase
TBIL: Total Bilirubin
TP: Total Protein
UDM: Useful Dry Matter
VFA: Volatile Fatty Acids
WBC: White Blood Cells
WI: Water Intake
WGP: White Grape Pomace
References
Aguerre M. J., Wattiaux M. A., Powell J. M., Broderick G. A., and Arndt C. (2011). Effect of forage-to-concentrate ratio in dairy cow diets on emission of greenhouse gases, total tract digestibility, and energy partitioning. J. Dairy Sci. 94, 3081–3093. doi: 10.3168/jds.2010-4011
Alba D. F., Campigotto G., Cazarotto C. J., Dos Santos D. S., Gebert R. R., Reis J. H., et al. (2019). Use of grape residue flour in lactating dairy sheep in heat stress: Effects on health, milk production and quality. J. Thermal Biol. 82, 197–205. doi: 10.1016/j.jtherbio.2019.04.007
Antunovic Z., Novoselec J., Klir Šalavardić Ž., Steiner Z., Drenjančević M., Pavić V., et al. (2024). The effect of grape seed cake as a dietary supplement rich in polyphenols on the quantity and quality of milk, metabolic profile of blood, and antioxidative status of lactating dairy goats. Agriculture 14, 479. doi: 10.3390/agriculture14030479
Association of Official Analytical Chemists (1999). Official methods of analysis. 16th (Washington, DC, USA: Association of Official Analytical Chemists).
Badiee Baghsiyah M., Bashtani M., Farhangfar S. H., and Sarir H. (2023). Effect of grape by-products inclusion on ruminal fermentation, blood metabolites, and milk fatty acid composition in lactating Saanen goats. Iranian J. Appl. Anim. Sci. 13, 731–742.
Baker L., Bender J., Ferguson J., Rassler S., Pitta D., Chann S., et al. (2024). Leveraging dairy cattle to upcycle culled citrus fruit for emission mitigation and resource co-benefits: A case study. Resources Conserv. Recycling 203, 107452. doi: 10.1016/j.resconrec.2024.107452
Bennato F., Ianni A., Grotta L., and Martino G. (2022). Evaluation of chemical-nutritional characteristics of whey and ricotta obtained by ewes fed red grape pomace dietary supplementation. Food Sci. Anim. Resour. 42, 504–516. doi: 10.5851/kosfa.2022.e20
Benoit M. and Mottet A. (2023). Energy scarcity and rising cost: Towards a paradigm shift for livestock. Agric. Syst. 205, 103585. doi: 10.1016/j.agsy.2022.103585
Bordiga M., Travaglia F., Locatelli M., Arlorio M., and Coïsson J. D. (2015). Spent grape pomace as a still potential by-product. Int. J. Food Sci. Technol. 50, 2022–2031. doi: 10.1111/ijfs.12853
Cai Y., Cheng Y., and Wan Q. (2018). Local mammary glucose supply regulates availability and intracellular pathways in dairy goats. Front. Physiol. 9. doi: 10.3389/fphys.2018.01467
Cannas A., Tedeschi L. O., Fox D. G., Pell A. N., and Van Soest P. J. (2007). A mechanistic model for predicting the nutrient requirements and feed biological values for sheep. J. Anim. Sci. 85, 171–186. doi: 10.2527/2004.821149x
Castillo C., Abuelo A., and Hernandez J. (2019). “Ruminant (bovine, caprine, and ovine) milk and meat production: The challenge of food quality and sustainability through the use of plant extracts,” in Encyclopedia of food security and sustainability. Eds. Ferranti P., Berry E. M., and Anderson J. R. (Amsterdam, Netherlands: Elsevier), 25–42. doi: 10.1016/B978-0-08-100596-5.21834-4
Chavez J., Bernal G., Rodríguez A., Mark K., Díaz E., Aguilera A., et al. (2009). Influence of pregnancy and lactation on glucose metabolism of Nubian goats. Trop. Subtropical Agroecosystems 11, 225–232. Available online at: https://www.redalyc.org/pdf/939/93913000046.pdf.
Chilliard Y., Ferlay A., Faulconnier Y., Bonnet M., Rouel J., and Bocquier F. (2000). Adipose tissue metabolism and its role in adaptations to undernutrition in ruminants. Proc. Nutr. Soc. 59, 127–134. doi: 10.1017/S002966510000015X
Chilliard Y., Ferlay A., Rouel J., and Lamberet G. (2003). A review of nutritional and physiological factors affecting goat milk lipid synthesis and lipolysis. J. Dairy Sci. 86, 1751–1770. doi: 10.3168/jds.S0022-0302(03)73761-8
Correddu F., Lunesu M. F., Buffa G., Atzori A. S., Nudda A., Battacone G., et al. (2020). Can agro-industrial by-products rich in polyphenols be advantageously used in the feeding and nutrition of dairy small ruminants? Animals 10, 131. doi: 10.3390/ani10010131
Correddu F., Nudda A., Battacone G., Boe R., Francesconi A. H. D., and Pulina G. (2015). Effects of grape seed supplementation, alone or associated with linseed, on ruminal metabolism in Sarda dairy sheep. Anim. Feed Sci. Technol. 199, 61–72. doi: 10.1016/j.anifeedsci.2014.11.002
Dentinho M. T. P., Paulos K., Costa C., Costa J., Fialho L., and Cachucho L. (2023). Silages of agro-industrial by-products in lamb diets: Effect on growth performance, carcass, meat quality and in vitro methane emissions. Anim. Feed Sci. Technol. 298, 115603. doi: 10.1016/j.anifeedsci.2023.115603
Do Nascimento E. M., Silva T. M., Garcez Neto A. F., Reis F. B., Santos E. B. L., Silva V. A., et al. (2025). Ingestive behavior, intake and digestibility of nutrients, and water and nitrogen balance of lactating goats fed dehydrated grape pomace. Livestock Sci. 275, 105726. doi: 10.1016/j.livsci.2025.105726
Donkin S. S. (2016). “Control of hepatic gluconeogenesis during the transition period,” in Ruminant nutrition symposium (Gainesville, FL, USA: Purdue University Extension), 71–80. Available online at: https://animal.ifas.ufl.edu/apps/dairymedia/rns/2016/11.%20Donkin.pdf.
FAO (Food and Agriculture Organization of the United Nations) (2023). “The state of food security and nutrition in the world 2023,” in Urbanization, agrifood systems transformation, and healthy diets across the rural-urban continuum (Rome, Italy: FAO). Available online at: https://openknowledge.fao.org/items/445c9d27-b396-4126-96c9-50b335364d01 (Accessed March 28, 2025).
Feng-Xia L., Shu-Fang F., Xiu-Fang B., Fang C., Xiao-Jun L., Xiao-Song H., et al. (2013). Physico-chemical and antioxidant properties of four mango (Mangifera indica L.) cultivars in China. Food Chem. 138, 396–405.
Fernandez C., Sanchez-Seiquer P., Navarro M. J., and Garces C. (2005). “Modelling the voluntary dry matter intake in Murciano-Granadina dairy goats,” in Sustainable grazing, nutritional utilization and quality of sheep and goat products. Eds. Alcaide M. E., Salem B. H., Biala K., and Morand-Fehr P. (Zaragoza, Spain: CIHEAM), 395–399.
Frutos P., Hervas G., Giraldez F. J., and Mantecón A. R. (2004). Review: Tannins and ruminant nutrition. Spanish J. Agric. Res. 2, 191–202. doi: 10.5424/sjar/2004022-73
Galanakis C. M. (2012). Recovery of high added-value components from food wastes: Conventional, emerging technologies, and commercialized applications. Trends Food Sci. Technol. 26, 68–87. doi: 10.1016/j.tifs.2012.03.003
Galvez-Lopez M., Navarro A., Muelas R., Roca A., Peris C., Romero G., et al. (2025). Potential of baled silage to preserve white grape pomace for ruminant feeding. Agriculture 15, 974. doi: 10.3390/agriculture15090974
Harbeoui H., Rebey I. B., Ouerghemmi I., Wannes W. A., Zemni H., Zoghlami N., et al. (2017). Biochemical characterization and antioxidant activity of grape (Vitis vinifera L.) seed oils from nine Tunisian varieties. J. Food Biochem. 42, e12595. doi: 10.1111/jfbc.12595
Huang R., Romero P., Belanche A., Ungerfeld E. M., Yañez-Ruiz D., Morgavi D. P., et al. (2023). Evaluating the effect of phenolic compounds as hydrogen acceptors when ruminal methanogenesis is inhibited in vitro – Part 1. Dairy cows. Anim. 17, 100788. doi: 10.1016/j.animal.2023.100788
Ibañez C., Criscioni P., Arriaga H., Merino P., Espinós F. J., and Fernandez C. (2016). Murciano-Granadina goat performance and methane emission after replacing barley grain with fibrous by-products. PloS One 11, e0151215. doi: 10.1371/journal.pone.0151215
ISO (1997). ISO 10520: Native starch—Determination of starch content—Ewers polarimetric method (Geneva, Switzerland: International Organization for Standardization).
Jarrige R., De Blas J. C., and Fraga M. J. (Eds.) (1981). Alimentación en rumiantes (Mundi-Prensa: INRA).
Juraček M., Vašekova P., Massanyi P., Kovačik A., Bíro D., Šimko M., et al. (2021). The effect of dried grape pomace feeding on nutrients digestibility and serum biochemical profile of wethers. Agriculture 11, 1194. doi: 10.3390/agriculture11121194
Kafantaris I., Kotsampasi B., Christodoulou V., Kokka E., Kouka P., Terzopoulou Z., et al. (2016). Grape pomace improves antioxidant capacity and faecal microflora of lambs. J. Anim. Physiol. Anim. Nutr. 101, 1096–1108. doi: 10.1111/jpn.12569
Kaneko J. J., Harvey J. W., and Bruss M. L. (1997). “Appendixes,” in Clinical biochemistry of domestic animals, 5th. Eds. Kaneko J. J., Harvey J. W., and Bruss M. L. (Academic Press), 885–905.
Kilama J., Yakir Y., Shaani Y., Adin G., Kaadan S., Wagali P., et al. (2023). Chemical composition, in vitro digestibility, and storability of selected agro-industrial by-products: Alternative ruminant feed ingredients in Israel. Heliyon 9, e14581. doi: 10.1016/j.heliyon.2023.e14581
Kim D., Jeong S. W., and Lee C. Y. (2003). Antioxidant capacity of phenolic phytochemicals from various cultivars of plums. Food Chem. 81, 321–326. doi: 10.1016/S0308-8146(02)00423-5
Leite A. V., Malta L. G., Riccio M. F., Eberlin M. N., Pastore G. M., and Maróstica Júnior M. R. (2011). Antioxidant potential of rat plasma by administration of freeze-dried jaboticaba peel (Myrciaria jaboticaba Vell Berg). J. Agric. Food Chem. 59, 2277–2283. doi: 10.1021/jf103181x
Makkar H. P. S. (2003). Effects and fate of tannins in ruminant animals, adaptation to tannins, and strategies to overcome detrimental effects of feeding tannin-rich feeds. Small Ruminant Res. 49, 241–256. doi: 10.1016/S0921-4488(03)00142-1
Malenica D., Kass M., and Bhat R. (2023). Sustainable management and valorization of agri-food industrial wastes and by-products as animal feed: For ruminants, non-ruminants and as poultry feed. Sustainability 15, 117. doi: 10.3390/su15010117
Manso T., Gallardo B., Salva A., Guerra-Rivas C., Mantecón a.R., Lavín P., et al. (2016). Influence of dietary grape pomace combined with linseed oil on fatty acid profile and milk composition. J. Dairy Sci. 99, 1111–1120. doi: 10.3168/jds.2015-9981
Marques J. G. O., De Oliveira Silva R., Barioni L. G., Hall J. A. J., Fossaert C., Tedeschi L. O., et al. (2022). Evaluating environmental and economic trade-offs in cattle feed strategies using multiobjective optimization. Agric. Syst. 195, 103308. doi: 10.1016/j.agsy.2021.103308
Martins Flores D. R., Da Fonseca P. A. F., Schmitt J., Tonetto C. J., Rosado Junior A. G., Hammerschmitt R. K., et al. (2020). Lambs fed with increasing levels of grape pomace silage: Effects on productive performance, carcass characteristics, and blood parameters. Livestock Sci. 240, 104169. doi: 10.1016/j.livsci.2020.104169
Martins Flores D. R., Patrícia Da Fonseca A. F., Schmitt J., Tonetto C. J., Rosado Junior A. G., and Hammerschmitt R. K. (2021). Lambs fed with increasing levels of grape pomace silage: Effects on meat quality. Small Ruminant Res. 195, 106234. doi: 10.1016/j.smallrumres.2020.106234
Menke K. H. and Steingass H. (1988). Estimation of the energetic feed value obtained from chemical analysis and in vitro gas production using rumen fluid. Anim. Res. Dev. 28, 7–55.
Molosse V. L., Deolindo G. L., Lago R. V. P., Cecere B. G. O., Zotti C. A., Vedovato M., et al. (2023). Effects of the inclusion of ensiled and dehydrated grape pomace in beef cattle diet: Growth performance, health, and economic viability. Anim. Feed Sci. Technol. 302, 115671. doi: 10.1016/j.anifeedsci.2023.115671
Monllor P., Romero G., Sendra E., Atzori A. S., and Díaz J. R. (2020). Short-term effect of the inclusion of silage artichoke by-products in diets of dairy goats on milk quality. Animals 10, 339. doi: 10.3390/ani10020339
Monllor P., Zemzmi J., Muelas R., Roca A., Sendra E., Romero G., et al. (2023). Long-term feeding of dairy goats with 40% artichoke by-product silage preserves milk yield, nutritional composition and animal health status. Animals 13, 3585. doi: 10.3390/ani13223585
Mottet A., De Haan C., Falcucci A., Tempio G., Opio C., and Gerber P. (2017). Livestock: On our plates or eating at our table? A new analysis of the feed/food debate. Global Food Secur. 14, 1–8. doi: 10.1016/j.gfs.2017.01.001
NRC (National Research Council) (2007). Nutrient requirements of small ruminants: Sheep, goats, cervids, and new world camelids (Chs. 4–5) (Washington, DC, USA: The National Academies Press). doi: 10.17226/11654
Nudda A., Correddu F., Marzano A., Battacone G., Nicolussi P., Bonelli P., et al. (2015). Effects of diets containing grape seed, linseed, or both on milk production, liver and kidney activity, and immunity in lactating dairy ewes. J. Dairy Sci. 98, 1157–1166. doi: 10.3168/jds.2014-8492
Ovcharova T., Zlatanov M., and Dimitrova R. (2016). Chemical composition of seeds of four Bulgarian grape varieties. J. Viticulture Enology 31, 31–40. doi: 10.1051/ctv/20163101031
Oyaizu M. (1986). Studies on products of browning reaction: Antioxidative activities of products of browning reaction prepared from glucosamine. Japanese J. Nutr. Dietetics 44, 307–315. doi: 10.5264/eiyogakuzashi.44.307
Patra A. K. and Saxena J. (2011). Exploitation of dietary tannins to improve rumen metabolism and ruminant nutrition. J. Sci. Food Agric. 91, 24–37. doi: 10.1002/jsfa.4152
Qiu Y., Zhao H., He X., Zhu F., Zhang F., Liu B., et al. (2023). Effects of fermented feed of Pennisetum giganteum on growth performance, oxidative stress, immunity, and gastrointestinal microflora of Boer goats under thermal stress. Front. Microbiol. 13. doi: 10.3389/fmicb.2022.1030262
Raggio G., Lobley G. E., Lemosquet S., Rulquin H., and Lapierre H. (2006). Effect of casein and propionate supply on milk yield and nitrogen utilization in dairy cows. J. Dairy Sci. 89, 4340–4351. doi: 10.3168/jds.S0022-0302(06)72481-X
Schau E. M. and Fet A. M. (2008). LCA studies of food products as background for environmental product declarations. Int. J. Life Cycle Assess. 13, 255–265. doi: 10.1065/lca2007.12.372
Shi Q., Zhang D., and Tan Z. (2025). Pan-genomic insights into rumen microbiome-mediated SCFA production in ruminants. Microorganisms 13, 1175. doi: 10.3390/microorganisms13061175
Silanikove N. (1994). The struggle to maintain hydration and osmoregulation in animals experiencing severe dehydration and rapid rehydration: The story of ruminants. Exp. Physiol. 79, 281–300. doi: 10.1113/expphysiol.1994.sp003764
Singh S., Hundal J. S., Patra A. K., Sethi R. S., and Sharma A. (2022). A composite polyphenol-rich extract improved growth performance, ruminal fermentation and immunity, while decreasing methanogenesis and excretion of nitrogen and phosphorus in growing buffaloes. Environ. Sci. pollut. Res. 29, 24757–24773. doi: 10.1007/s11356-021-17674-1
Taladrid D., Laguna L., Bartolome B., and Moreno-Arribas M. V. (2019). Applications and new uses of winemaking byproducts. Intercompany 22, 42–45.
Tangolar S. G., Özogul F., Tangolar S., and Yağmur C. (2011). Tocopherol content in fifteen grape varieties obtained using a rapid HPLC method. J. Food Composition Anal. 24, 481–486. doi: 10.1016/j.jfca.2010.08.003
Van Soest P. J. (1994a). Nutritional ecology of the ruminant (2nd ed., Chapter 20: Intake (Ithaca, NY, USA: Cornell University Press), 337–353.
Van Soest P. J. (1994b). Nutritional ecology of the ruminant (2nd ed., Section: Water Balance, in Chapter 20: Intake (Ithaca, NY, USA: Cornell University Press), 342–344.
Van Soest P. J., Robertson J. B., and Lewis B. A. (1991). Methods for dietary neutral detergent fibre and non-starch polysaccharides in relation to animal nutrition. J. Dairy Sci. 74, 3583–3597. doi: 10.3168/jds.S0022-0302(91)78551-2
Ventura-Cordero J., Sandoval-Castro C. A., Torres-Acosta J. F. J., Hoste H., Chan-Perez J. I., and Aguilar-Caballero A. J. (2017). Foliage from tropical deciduous forest as a nutraceutical feed and its anthelmintic potential in small ruminants [in Spanish. Rev. la Asociación Interamericana Protección Vegetal 37, 4–15.
Wang J., Yin X., and Sun X. (2024). Hepatic gluconeogenesis and regulatory mechanisms in lactating ruminants: A literature review. Anim. Sci. J. 95, e14080. doi: 10.1002/aro2.80
Wisser D., Grogan D. S., Lanzoni L., Tempio G., Cinardi G., Prusevich A., et al. (2024). Water use in livestock agri-food systems and its contribution to local water scarcity: A spatially distributed global analysis. Water 16, 1681. doi: 10.3390/w16121681
Yañez-Ruiz D. R., Bannink A., Dijkstra J., Kebreab E., Morgavi D. P., O’Kiely P., et al. (2016). Design, implementation and interpretation of in vitro batch culture experiments to assess enteric methane mitigation in ruminants—a review. Anim. Feed Sci. Technol. 216, 1–18. doi: 10.1016/j.anifeedsci.2016.03.016
Keywords: circular economy, by-products, animal nutrition, feedstuffs, functional properties, oxidative stress
Citation: Galvez-Lopez M, Romero G, Sandoval-Castro CA, Perez E, Rodriguez M, Peris C and Díaz JR (2025) Short-term feeding of different levels of white grape pomace silage in dairy goats: impacts on milk yield, nutritional efficiency, and metabolic responses. Front. Anim. Sci. 6:1675085. doi: 10.3389/fanim.2025.1675085
Received: 28 July 2025; Accepted: 06 October 2025;
Published: 10 December 2025.
Edited by:
Vincenzo Lopreiato, University of Messina, ItalyReviewed by:
Annalisa Amato, University of Messina, ItalyAttapol Tiantong, Silpakorn University, Thailand
Copyright © 2025 Galvez-Lopez, Romero, Sandoval-Castro, Perez, Rodriguez, Peris and Díaz. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Gema Romero, Z2VtYXJvbWVyb0B1bWguZXM=