 Basílio Felizardo Lima Neto1
Basílio Felizardo Lima Neto1 Jordania Oliveira Silva2
Jordania Oliveira Silva2 Felipe Boniedj Ventura Alvares1Samira Pereira Batista1
Felipe Boniedj Ventura Alvares1Samira Pereira Batista1 Geraldo Moreira Silva Filho1Gláucia Diojânia Azevedo Medeiros2Carla Monadeli Filgueira Rodrigues2Bruno Rafael Fermino3
Geraldo Moreira Silva Filho1Gláucia Diojânia Azevedo Medeiros2Carla Monadeli Filgueira Rodrigues2Bruno Rafael Fermino3 Vinícius Longo Ribeiro Vilela1,2*Thais Ferreira Feitosa1,2
Vinícius Longo Ribeiro Vilela1,2*Thais Ferreira Feitosa1,2- 1Postgraduate Program in Science and Animal Health, Federal University of Campina Grande, Patos, Brazil
- 2Department of Veterinary Medicine, Federal Institute of Paraíba, Sousa, Brazil
- 3Department of Veterinary Medicine, Federal Institute of Rondônia, Jaru, Brazil
This study aimed to estimate the prevalence and identify risk factors associated with Trypanosoma vivax infection in dairy cattle herds in the Brazilian Semiarid region, as well as report an outbreak of trypanosomiasis. A total of 390 cattle blood samples were collected from 61 farms in the states of Ceará e Paraíba, and a conventional PCR was performed for the diagnosis of T. vivax. Positive samples were bidirectionally sequenced. An epidemiological questionnaire was applied on each farm, and the resulting data were subjected to univariate and multivariate analyses to identify potential risk factors. An outbreak was detected in the municipality of Orós, Ceará, where cases of abortion and estrus repetition were reported among cows. Of the 390 samples analyzed, 1.02% (4/390) tested positive for T. vivax. No statistically significant risk factors were identified. All positive samples originated from the farm in Orós, where 80% (4/5) of the tested animals were infected. Similarity analysis using BLAST revealed 100% identity with T. vivax isolates previously reported in donkeys, sheep, and cattle from the Brazilian semi-arid region, and high similarity with isolates from South American countries and West Africa. In the phylogenetic analysis, sequences TviBrCe13 and TviBrCe14 clustered with isolates from the Brazilian Semiarid region, as well as with South American and West African strains. It was concluded that the prevalence of T. vivax in the Brazilian Semiarid region is low, although the area remains susceptible to outbreak events.
1 Introduction
Trypanosoma spp. is a genus of pathogenic protozoa of significant economic importance to the African continent, particularly in regions where the tsetse fly (Glossina spp.), its biological vector, is present (Gardiner, 1989). Within this genus, Trypanosoma (Dutonella) vivax (Ziemann, 1905, as cited in Silva, 1996) stands out as a major species affecting cattle, causing substantial impacts on herd health and both productive and reproductive performance (Couto et al., 2024). Through its adaptation to mechanical transmission by other hematophagous insects such as tabanids, Stomoxys calcitrans, and via contaminated fomites like needles and syringes, it has spread to South America (Levine, 1973; Melo Junior et al., 2022). Trypanosomiasis caused by T. vivax has been expanding and is currently responsible for significant economic losses in dairy farms across Brazil (Reis, 2021).
Several techniques can be used for the diagnosis of Trypanosoma vivax, classified into direct and indirect methods. Among the direct methods, the most notable are blood smear examination, the microhematocrit centrifugation technique (Woo’s technique), and Polymerase Chain Reaction (PCR). As an indirect method, the Enzyme-Linked Immunosorbent Assay (ELISA) stands. Direct parasitological examinations exhibit low sensitivity in cases of chronic infection due to the reduced number of circulating parasites in the bloodstream out (Alcindo et al., 2022). Serological tests, such as IFA and ELISA, are useful as screening tools to identify herd exposure to infection but are unable to distinguish active infections from post-treatment immune responses. On the other hand, molecular methods, such as conventional PCR (cPCR), are considered highly sensitive and specific tools, especially during the patent phase of the disease. The combination of molecular and serological tests allows for more effective diagnosis, particularly in animals that remain infected even after treatment (Fidelis Junior et al., 2019). The use of highly sensitive diagnostic methods is crucial, as T. vivax often induces low parasitemia (Cadioli et al., 2015). In this context, the PCR assay based on a T. vivax-like CatL-specific sequence (TviCatL-PCR) has been shown to be species-specific, highly sensitive, and suitable for the diagnosis of T. vivax (Cortez et al., 2009; Rodrigues et al., 2015).
In the Brazilian Semiarid region, T. vivax is not endemic. Due to prolonged periods of drought and high temperatures, mechanical vectors are unable to develop properly throughout the entire year, resulting in limited and inconsistent exposure of animals to the parasite and, consequently, the lack of active immunity development. When the rainy season begins, vector populations resume development, and outbreaks are observed with greater frequency (Batista et al., 2007, 2008; Rodrigues et al., 2015). The main clinical signs observed in animals infected with T. vivax during outbreaks in Brazil include progressive weight loss, a significant drop in milk production, anemia, abortion, anestrus, and perinatal mortality (Batista et al., 2022).
In addition to climatic factors, anthropogenic aspects—such as the movement of animals without sanitary control, the shared use of piercing and cutting instruments (such as needles and syringes), and deficiencies in sanitary surveillance programs—also contribute significantly to the emergence and spread of cattle trypanosomiasis outbreaks (Bastos et al., 2020). The high density of mechanical vectors during the rainy season, combined with inadequate sanitary management practices in dairy herds, facilitates the maintenance of the T. vivax epidemiological cycle (Batista et al., 2008), resulting in substantial economic losses for producers and posing a threat to animal health and survival (Couto et al., 2024). Given the scarcity of updated data on the epidemiology of this parasite in the Brazilian Semiarid region, this study aimed to estimate the prevalence and identify risk factors associated with T. vivax infection in cattle from dairy farms in the region, as well as to describe the occurrence of a trypanosomiasis outbreak in an affected property.
2 Method
2.1 Sampling
To determine the number of samples to be evaluated, a simple random sampling method was used, according to Thrusfield (2007).
Where:
N: required sample size;
Z: standard normal distribution value for a 95% confidence level;
p: expected prevalence of 50%;
d: sampling error of 5%.
The local population was adjusted using the following formula:
Where:
najus: final number of selected farms;
n: initial number of selected farms;
N: total number of farms in the region.
The minimum number of samples to be evaluated, as determined by the sample size calculation, was 385. However, due to producer participation, a total of 390 samples were collected. To enhance spatial distribution, a maximum of six animals per farm were selected.
2.2 Study area
The state of Ceará has a herd of 2.772,173 cattle, while the state of Paraíba has a herd of 1.412,480 (IBGE 2023). A total of 390 blood samples were collected from the median caudal vein of cattle in six municipalities of the microregions Cariri, Iguatu, Lavras da Mangabeira and Sul Cearense of the state of Ceará and 17 municipalities of the microregions Brejo Paraibano, Cajazeiras, Campina Grande, Catolé do Rocha, Curimataú Ocidental, Esperança, João Pessoa, Patos, Piancó and Sousa of the state of Paraíba, located in the semi-arid region of Brazil (Figure 1). In the state of Ceará, 125 samples were collected from 17 farms, while in the state of Paraíba, 265 samples were collected from 44 farms. The samples were collected between September 2023 and December 2024 using Vacutainer® tubes containing EDTA as an anticoagulant, through reusable holders. Samples were collected from farms with semi-intensive and extensive dairy production systems, comprising male and female animals aged 0 to over 5 years. All samples were properly labeled and transported in isothermal boxes for processing at the Laboratory of Molecular Biology (LabMol) of the Federal Institute of Paraíba.
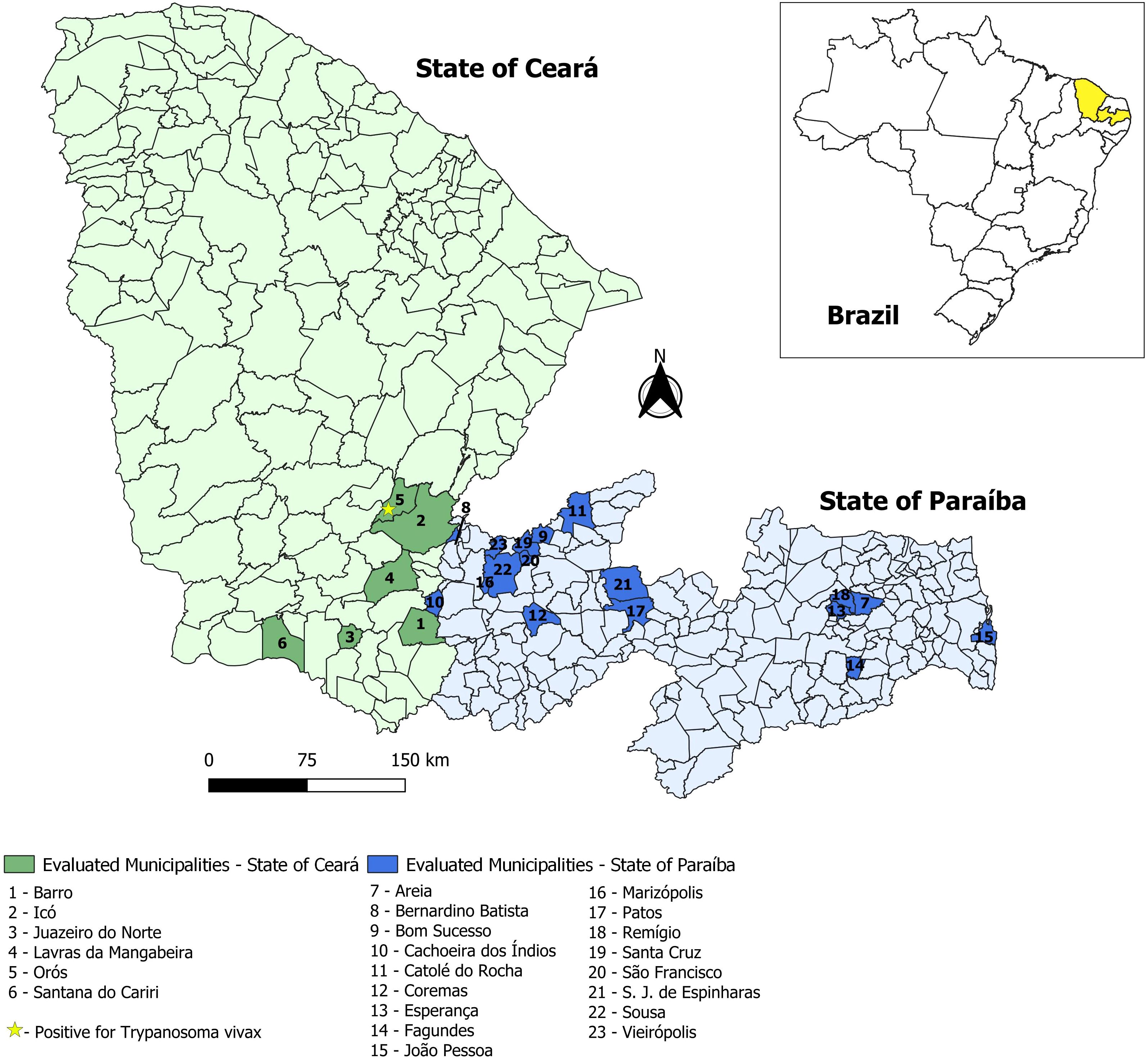
Figure 1. Distribution of municipalities where cattle blood sample collections were conducted in the states of Ceará and Paraíba, Northeast Brazil.
2.3 Initial sample processing and DNA extraction
The samples were centrifuged, and 200 µL of the buffy coat was collected and transferred to sterile, 1.5 mL microtubes that were free of DNases, RNases, pyrogens, dyes, and heavy metals. DNA was extracted using the Cellco® Blood-Animal-Plant DNA Preparation Kit (Cellco®, São Carlos, SP, Brazil), following the manufacturer’s instructions. After extraction, DNA and RNA quantification was performed using a microvolume spectrophotometer (L-Quant III, Loccus, Brazil) to assess extraction efficiency. 50 µL of extracted DNA was stored in 1.5 mL microtubes and kept at −20°C until use in Polymerase Chain Reaction (PCR) assays.
2.4 Polymerase chain reaction
For a final volume of 20 µL, 2 µL of DNA (the DNA concentration range from 50 to 92ng/µL) were added to 3.15 µL of Platinum™ Taq DNA Polymerase mix (cat. 10966-030, Invitrogen, Carlsbad, CA, USA), 12.85 µL of ultrapure water (dH2O), and 1.0 µL of each primer (10 µM). The PCR assay employed the primers TviCatL1-F (5′ GCCATCGCCAGTACCTCGCGA 3′) and DTO155-R (5′ TTAAAGCTCCAGGAGATTCTTGATGATCCAGTA 3′), using the thermal cycling conditions described by Cortez et al. (2009).
The PCR products were subjected to electrophoresis in 1.5% agarose gel at 80 V for 40 minutes and visualized under UV light. Positive samples amplified a 177 base pairs (bp) fragment, consistent with the band size described for T. vivax (Cortez et al., 2009). Amplicon sizes were determined using a 100 bp molecular weight marker (GeneRuler DNA Ladder Mix™, Invitrogen®, Carlsbad, CA, USA).
2.5 Sequencing and phylogenetic analysis
The CATL-amplified DNA sequences were analyzed using Geneious v4.8.5 (http://www.geneious.com/). The sequences were aligned using the MUSCLE algorithm (Edgar, 2004), implemented in MEGA X (Kumar et al., 2018). The alignment, which included Trypanosoma vivax sequences from Brazil, Venezuela, Ecuador, The Gambia, Burkina Faso, Nigeria, Mozambique, Kenya, and Zambia, was used to construct a Maximum Likelihood (ML) phylogenetic tree.
The ML analysis was performed with RAxML v8.2 (Stamatakis, 2014), employing the GTRGAMMA model with a proportion of invariable sites. Model parameters were estimated during tree searches, and branch support values were obtained with 500 bootstrap replicates, following the methodology previously described by Cortez et al. (2009). Importantly, the use of the TviCatL-PCR assay (Cortez et al., 2009), which targets repeated gene sequences encoding the cathepsin L-like enzyme, was intended exclusively for species confirmation of T. vivax through genetic sequencing.
2.6 Epidemiological questionnaire
At the time of sampling, an epidemiological questionnaire was administered at each farm, addressing the following items: herd size, production system, breed, type of farming activity, grazing area, main health problems, use of the same needle on multiple animals, presence of flies, pasture sharing with other farms, bull rotation, and contact of cows with bulls from other properties.
2.7 Statistical analysis
To identify factors associated with the prevalence of T. vivax, data from the epidemiological questionnaires were analyzed in two steps: univariate and multivariate analyses. In the univariate analysis, each independent variable was correlated with the dependent variable (positivity), and those with a p-value ≤ 0.2, according to the chi-square test (Zar, 1999), were included in a multivariate analysis using multiple logistic regression (Hosmer and Lemeshow, 2000), with a significance level of 5%. To detect possible collinearity among variables, a correlation test was applied; if the correlation coefficient exceeded 0.9, one of the variables was excluded based on biological plausibility. Model fit was assessed using the chi-square statistic and the Omnibus test. The results were analyzed using GraphPad Prism software, version 8.0.1.
2.8 Monitoring of a trypanosomiasis outbreak in the State of Ceará
During the study, an outbreak was identified on a farm located in the municipality of Orós, State of Ceará, where successive abortions and repeated estrus cycles were being observed among breeding cows.
3 Results
3.1 Prevalence and risk factors associated with T. vivax infection
Among the 390 blood samples collected in the states of Ceará and Paraíba, 1.02% (4/390) tested positive for T. vivax (Table 1). No statistically significant risk factors were associated with T. vivax infection.
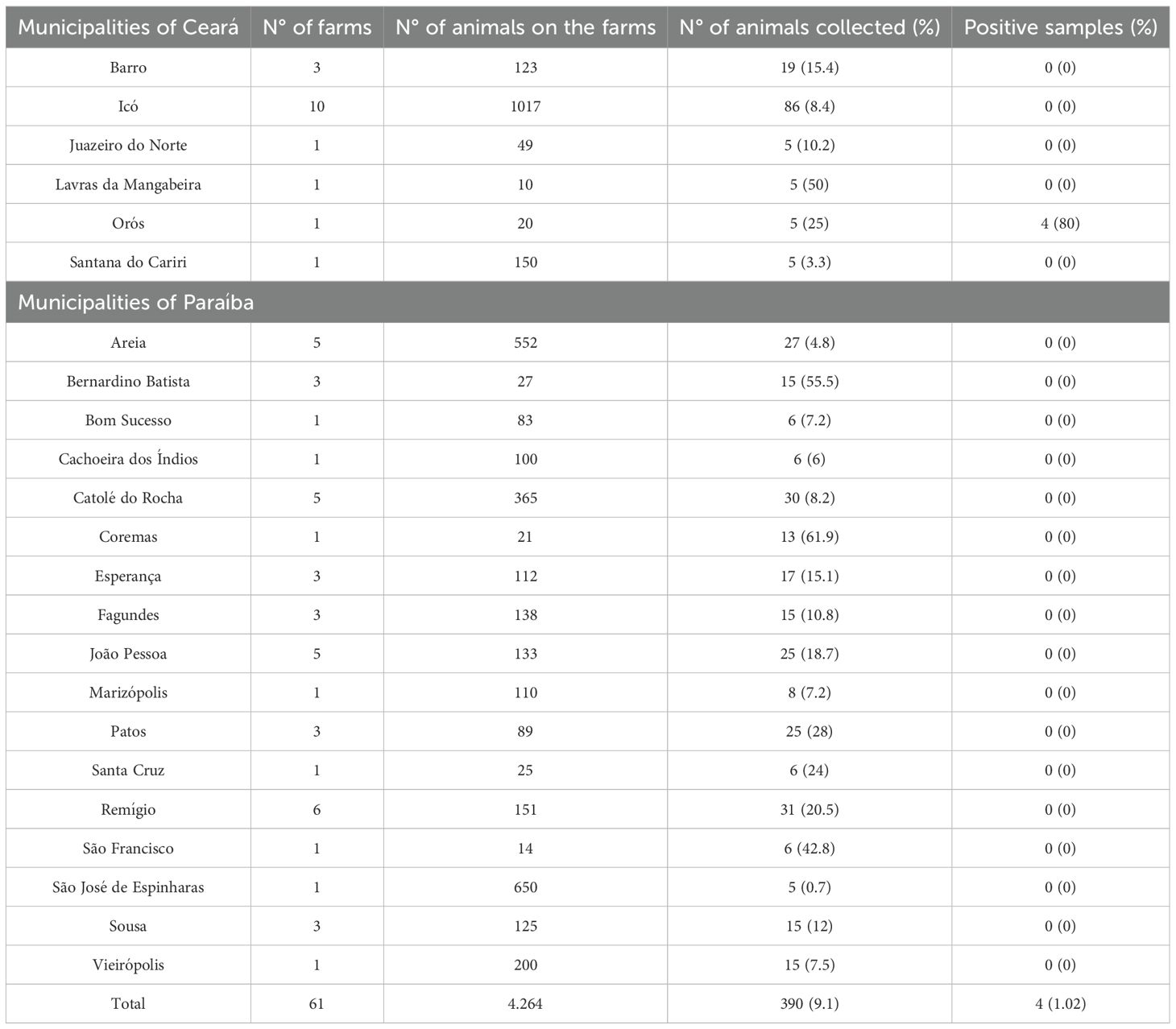
Table 1. Distribution of farms and cattle blood samples collected by municipality in the states of Ceará and Paraíba, Northeast Brazil.
3.2 Outbreak of trypanosomiasis in the municipality of Orós, State of Ceará
All positive samples in the present study originated from a single farm located in the municipality of Orós, in the state of Ceará, where T. vivax infection was detected in 80% (4/5) of the sampled animals from the herd (Table 1). The outbreak occurred among the dairy cows on the property, which exhibited cases of successive abortions and repeated estrus cycles. The outbreak took place in September 2023, during the dry season, in the absence of mechanical vectors. The animals were raised under a semi-extensive production system and had no contact with animals from other farms. It was reported that, in the months prior to sampling, animals had been acquired from another property in the region. There was no history of reproductive problems in the herd before the outbreak. Additionally, it was reported that the farm practiced sharing of syringes and needles among animals for the administration of medications and exogenous oxytocin.
3.3 Sequencing and phylogenetic analysis
The samples positive for T. vivax were subjected to Cathepsin L gene sequencing, resulting in two sequences from cattle, named TviBrCe13 (PX262614) and TviBrCe14 (PX262615). The similarity analysis conducted using BLAST revealed that both sequences shared 100% identity with previously reported isolates from the Brazilian Semiarid region, obtained from donkeys, sheep, and cattle, and also showed high similarity with isolates from other South American countries and West Africa.
In the phylogenetic analysis, sequences TviBrCe13 and TviBrCe14 consistently clustered within the same clade as isolates from the Brazilian Semiarid region and were closely related to other Brazilian isolates, as well as those from South American countries and West Africa (Figure 2). This clustering confirms the low genetic diversity observed among South American isolates and reinforces the evolutionary connection between these genotypes and those originally described in West Africa.
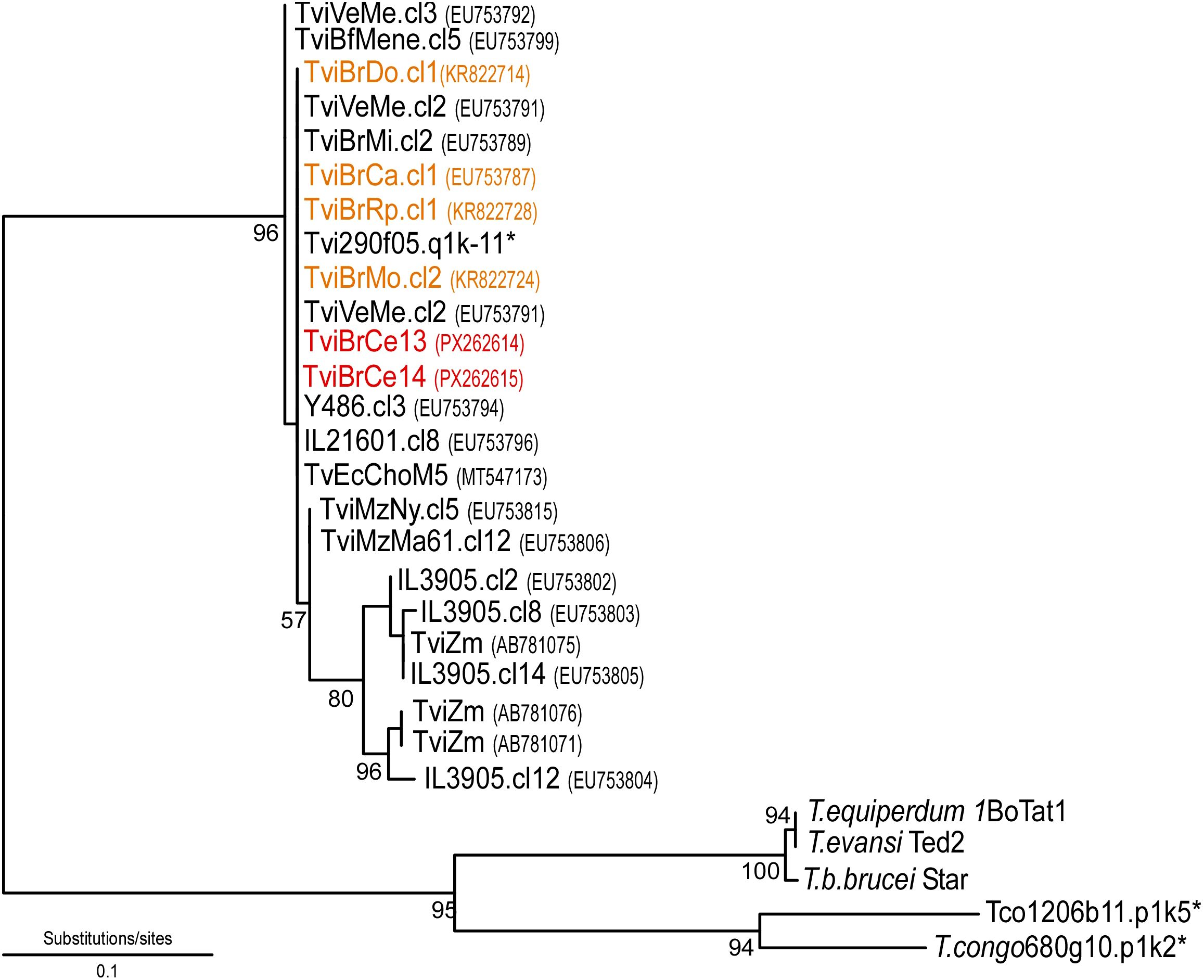
Figure 2. Neighbor-joining phylogenetic tree displaying the evolutionary relationships among the sequences obtained in this study and related GenBank entries, constructed using MEGA software (version 11). Bootstrap percentages are shown next to the main branches, indicating the level of statistical confidence for each cluster. The isolates from this research (TviBrCe13 and TviBrCe14) are highlighted in red and positioned within a strongly supported clade. Black sequences, derived from various hosts and geographic regions, were incorporated to demonstrate the genetic variability and evolutionary connections among the isolates. The sequences highlighted in orange represent isolates previously reported in the Semiarid region of Northeast Brazil. Sequences marked with an asterisk were retrieved from genome projects (Sanger Institute; https://www.sanger.ac.uk/science/data).
4 Discussion
This is the first report in the state of Ceará that includes not only molecular detection but also sequencing-based characterization and phylogenetic analysis of T. vivax, complementing previous reports that were limited to molecular diagnosis without in-depth genetic characterization (Rodrigues et al., 2024). This finding expands the understanding of the genetic diversity of the parasite in the Northeast region of Brazil and provides essential data to support control strategies and epidemiological surveillance in the region.
Studies estimating the prevalence of T. vivax in the Brazilian Semiarid region are scarce, with most research focused on isolated outbreaks. Four animals tested positive for T. vivax, and these cases were associated with an outbreak of trypanosomiasis in the municipality of Orós, State of Ceará. The low prevalence found in the present study is consistent with other reports from the Brazilian Semiarid. Rodrigues et al. (2024) reported a prevalence of 0.4% (1/246) for T. vivax in cattle in the state of Ceará. However, their research was limited to a single microregion, the Sertão Central, with five municipalities, whereas the present study was conducted across 23 municipalities of different microregions of the states of Ceará and Paraíba, covering a broader geographical area. Seroprevalence surveys carried out in the state of Paraíba similarly found no animals positive for T. vivax, as observed in the present work (Costa et al., 2013). In contrast, other regions of Brazil that are endemic for T. vivax exhibit higher prevalence rates. Bastos et al. (2020) reported a prevalence of 8.4% in cattle from 24 dairy farms in the state of Goiás, a known endemic area for T. vivax, which differs from the findings of the present study.
The absence of significant risk factors in this study may be attributed to the low prevalence observed. Nevertheless, the introduction of T. vivax-infected animals and the administration of exogenous oxytocin using shared syringes and needles in dairy cows are identified as causes of trypanosomiasis transmission. Bastos et al. (2020) has previously highlighted the importance of this risk factors and transmission method in Brazil. On this farm, the lack of contact with animals from other herds and the absence of mechanical vectors at the time of the outbreak indicate that the introduction of cattle not subjected to quarantine was the most plausible source of infection. The unrecorded history of previous reproductive problems reinforces the hypothesis that T. vivax was introduced with the newly arrived animals. Supporting this finding, Rodrigues et al. (2024) reported that the only T. vivax-positive animal identified in their study originated from a farm with a known history of trypanosomiasis, low incidence of Stomoxys calcitrans and tabanids, and that six months prior to sampling, new animals from external sources had been introduced into the herd. In areas of enzootic instability, where animals are not regularly exposed to the parasite, the introduction of animals infected may lead to trypanosomiasis outbreaks (Batista et al., 2008). Therefore, the absence of quarantine in dairy farms, combined with the introduction of new animals, represents an important risk factor for the occurrence of T. vivax outbreaks, particularly in areas of enzootic instability, reinforcing the need for preventive management practices and sanitary surveillance measures.
The occurrence of the outbreak in the absence of mechanical vectors suggests that management practices, such as the shared use of needles and syringes, may play a considerable role in the transmission of T. vivax in dairy herds. This underscores the importance of considering iatrogenic routes of transmission, particularly when newly introduced animals may serve as asymptomatic carriers. Studies have demonstrated the viability of T. vivax in 109 injectable veterinary products following contamination during administration, revealing that in 44% (48/109) of the samples, T. vivax remained viable regardless of time. Among the drug classes evaluated, the parasite was found to be viable in 100% (7/7) of oxytocin-based hormonal products (Melo Junior et al., 2022). The survival of the parasite in injectable fomites enables the transmission of T. vivax from an infected animal to a healthy one during the administration of medications or hormones. The persistence of T. vivax in injectable products, raises significant concerns regarding routine veterinary practices, emphasizing the urgent need for training and awareness among farmers and veterinarians to mitigate iatrogenic transmission.
The phylogenetic analysis of the isolates from this study reveals a pattern consistent with that previously observed in T. vivax outbreaks in South America, including the Brazilian Semiarid region. The sequences TviBrCe13 and TviBrCe14, obtained from cattle during an outbreak in the municipality of Orós, Ceará, showed 100% identity with isolates previously described in the Semiarid region, originating from donkeys, sheep, and cattle (Batista et al., 2007; Rodrigues et al., 2015). This finding demonstrates the low genetic diversity of the CatL gen in the T. vivax population circulating in the region, in accordance with previous studies, and supports the hypothesis that T. vivax was introduced into South America from West Africa, possibly through the transatlantic movement of animals during the colonial period (Cortez et al., 2009; Rodrigues et al., 2015). Rodrigues et al. (2015) previously identified that Brazilian isolates of T. vivax, similar to those in the present study, are genetically related to isolates from other regions of South America and West Africa, while differing from isolates from East and South-Central Africa.
Studies conducted in the Brazilian Semiarid region have demonstrated that the circulating genotypes of T. vivax are highly conserved and are shared among different host species such as donkeys, cattle, and sheep, regardless of clinical severity (Batista et al., 2007; Rodrigues et al., 2015). Previous studies have shown that sequences from isolates obtained from donkeys (TviBrDo) were identical or highly similar to sequences from isolates associated with acute outbreaks in cattle (TviBrCa and TviBrMo) and sheep (TviBrRp) in the same region, as well as similar to those from asymptomatic animals in endemic areas. The severity of clinical manifestations is influenced by previous contact with T. vivax and by the tolerance levels of different cattle species and breeds, rather than being associated with a specific parasite genotype (Rodrigues et al., 2015). This same pattern was observed in the present study, in which the genotypes of T. vivax isolates obtained from cattle affected by the outbreak were indistinguishable from those previously described in the Brazilian Semiarid region. Furthermore, outbreaks of trypanosomiasis in the Brazilian semiarid region are not restricted to cattle, having also been reported in goats and sheep (Batista et al., 2008, 2022), species of great importance for livestock production in this area. The monitoring of the disease’s occurrence in cattle, the preferred hosts of T. vivax, serves as an important indicator for epidemiological monitoring, enabling the implementation of measures to prevent the spread of the parasite among different species and to minimize the economic impacts of the disease on herds in the semiarid region.
Pereira et al. (2018) stated that trypanosomiasis presents clinical signs similar to other diseases, such as anaplasmosis and babesiosis, which are widely distributed throughout the Semiarid region. Furthermore, several studies conducted in the Brazilian Semiarid have shown that the disease occurs in the form of sporadic outbreaks and is not endemic to the region. A study conducted in Uganda found higher positivity for T. vivax using rapid diagnostic tests compared to PCR, which was attributed to the fact that the rapid test detects antibodies, whereas PCR detects parasite DNA. The authors further emphasized that this limits the use of antibody detection methods in guiding treatment decisions, as compared to antigen-based methods. Therefore, antigen detection methods are considered more relevant for supporting field veterinarians in treatment decision-making (Kizza et al., 2024). Consequently, caution is advised when interpreting results from rapid tests as a diagnostic tool for trypanosomiasis in the Brazilian Semiarid, since these tests are highly specific but have low sensitivity, which may result in false positives and lead to unnecessary treatment expenditures.
Based on these findings, it can be concluded that the prevalence of T. vivax in cattle in the Brazilian semiarid is low and that the region is predisposed to experiencing trypanosomiasis as outbreak events. Therefore, the disease should not be underestimated in this area.
Data availability statement
The data presented in the study are deposited in the GenBank repository, accession numbers PX262614 and PX262615.
Ethics statement
The animal studies were approved by Comitê de Ética no Uso de Animais - CEUA/Instituto Federal da Paraíba - IFPB. The studies were conducted in accordance with the local legislation and institutional requirements. Written informed consent was obtained from the owners for the participation of their animals in this study.
Author contributions
BL: Conceptualization, Data curation, Investigation, Methodology, Writing – original draft, Writing – review & editing. JS: Investigation, Methodology, Writing – review & editing. FA: Investigation, Methodology, Writing – review & editing. SB: Investigation, Methodology, Writing – review & editing. GS: Investigation, Methodology, Writing – review & editing. GM: Investigation, Methodology, Writing – review & editing. CR: Investigation, Methodology, Writing – review & editing. BF: Investigation, Methodology, Writing – review & editing. VV: Conceptualization, Formal Analysis, Investigation, Methodology, Supervision, Writing – review & editing. TF: Conceptualization, Formal Analysis, Investigation, Methodology, Project administration, Resources, Supervision, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This work was supported by the Paraíba State Research Foundation (FAPESQ-PB 19/2022, N outorga 067/2023). The National Council for Scientific and Technological Development (CNPq/Brazil) provided scholarships to VV.
Acknowledgments
The authors would like to thank the Paraíba State Research Foundation (FAPESQ-PB), the National Council for Scientific and Technological Development (CNPq/Brazil), and the Federal Institute of Education, Science and Technology of Paraíba (IFPB) for their support in conducting this research.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Alcindo J. F., Vieira M. C. G., Rocha T. V. P., Cardinot C. B., Deschk M., Amaral G. G., et al. (2022). Evaluation of techniques for diagnosis of Trypanosoma vivax infections in naturally infected cattle in the Zona da Mata Mineira. Rev. Bras. Parasitol. Vet. 31, e018021. doi: 10.1590/S1984-29612022007
Bastos T. S. A., Faria A. M., Couto L. F. M., Nicaretta J. E., Cavalcante A. S. A., Zapa D. M. B., et al. (2020). Epidemiological and molecular identification of Trypanosoma vivax diagnosed in cattle during outbreaks in central Brazil. Parasitology 147, 1313–1319. doi: 10.1017/S0031182020001006
Batista J. S., Bezerra F. S. B., Lira R. A., Carvalho J. R. G., Rosado Neto A. M., Petri A. A., et al. (2008). Clinical, epidemiological and pathological signs of natural infection in cattle by Trypanosoma vivax in Paraíba, Brazil. Pesq. Vet. Bras. 28, 63–69. doi: 10.1590/S0100-736X2008000100010
Batista J. S., Riet-Correa F., Teixeira M. M. G., Madruga C. R., Simões S. D. V., and Maia T. F. (2007). Trypanosomiasis by Trypanosoma vivax in cattle in the Brazilian semiarid: Description of an outbreak and lesions in the nervous system. Vet. Parasitol. 143, 174–181. doi: 10.1016/j.vetpar.2006.08.017
Batista J. S., Santos W. L. A., de Sousa A. C. F. C., Silva Teófilo T., Bezerra A. C. D. S., Rodrigues V. H. V., et al. (2022). Abortion and congenital transmission of Trypanosoma vivax in goats and ewes in semiarid northeastern Brazil. Res. Vet. Sci. 149, 125–127. doi: 10.1016/j.rvsc.2022.06.009
Brazilian Institute of Geography and Statistics (2023). Cattle herd (Bulls and Cows). Available online at: https://www.ibge.gov.br/explica/producao-agropecuaria/bovinos/br (Accessed September 06, 2025).
Cadioli F. A., Fidelis Júnior O. L., Sampaio P. H., dos Santos G. N., André M. R., Castilho K. J., et al. (2015). Detection of Trypanosoma vivax using PCR and LAMP during aparasitemic periods. Vet. Parasitol. 214, 174–177. doi: 10.1016/j.vetpar.2015.09.001
Cortez A. P., Rodrigues A. C., Garcia H. A., Neves L., Batista J. S., Bengaly Z., et al. (2009). Cathepsin L-like genes of Trypanosoma vivax from Africa and South America – characterization, relationships and diagnostic implications. Mol. Cell Probes 23, 44–51. doi: 10.1016/j.mcp.2008.11.003
Costa V. M., Ribeiro M. F., Duarte A. L., Mangueira J. M., Pessoa A. F., Azevedo S. S., et al. (2013). Seroprevalence and risk factors for cattle anaplasmosis, babesiosis, and trypanosomiasis in a Brazilian semiarid region. Rev. Bras. Parasitol. Vet. 22, 207–213. doi: 10.1590/S1984-29612013005000022
Couto L. F. M., Bastos T. S. A., Morais I. M. L., Salvador V. F., Leal L. L. L. L., Falavigna R. B., et al. (2024). Reproductive, productive and financial consequences of chronic Trypanosoma vivax infection in a dairy cattle herd in a region without a cyclic vector. Vet. Parasitol. 330, 110221. doi: 10.1016/j.vetpar.2024.110221
Edgar R. C. (2004). MUSCLE: multiple sequence alignment with improved accuracy and speed. Proc. IEEE Comput. Syst. Bioinf. Conf. (CSB 2004), 728–729. doi: 10.1109/CSB.2004.1332560
Fidelis Junior O. L., Sampaio P. H., Gonçalves L. R., André M. R., MaChado R. Z., Wijffels G., et al. (2019). Comparison of conventional and molecular techniques for Trypanosoma vivax diagnosis in experimentally infected cattle. Rev. Bras. Parasitol. Vet. 28, 203–209. doi: 10.1590/S1984-29612019034
Gardiner P. R. (1989). Recent studies of the biology of Trypanosoma vivax. Adv. Parasitol. 28, 229–317. doi: 10.1016/s0065-308x(08)60334-6
Hosmer D. W. and Lemeshow S. (2000). Applied Logistic Regression. 2nd (New York: John Wiley & Sons).
Kizza D., Azuba R., Wampande E., Okwasiimire R., Owembabazi L., Mandela W., et al. (2024). Comparative evaluation of rapid diagnostic test and PCR-based diagnostic assay for identification of trypanosomes in cattle of Apac and Kiryandongo districts, Uganda: A cross sectional study. BMC Vet. Res. 20, 570. doi: 10.1186/s12917-024-04436-7
Kumar S., Stecher G., Li M., Knyaz C., and Tamura K. (2018). MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 35, 1547–1549. doi: 10.1093/molbev/msy096
Levine N. D. (1973). “The hemoflagellates,” in Protozoan parasites of domestic animals and of man, 2nd. Ed. Levine N. D. (Burgess Publishing, Minneapolis), 36–78.
Melo Junior R. D., Azeredo Bastos T. S., Heller L. M., Couto L. F. M., Zapa D. M. B., de Assis Cavalcante A. S., et al. (2022). How many cattle can be infected by Trypanosoma vivax by reusing the same needle and syringe, and what is the viability time of this protozoan in injectable veterinary products? Parasitology 149, 270–282. doi: 10.1017/S003118202100175X
Pereira H. D., Simões S. V. D., Souza F. A. L., Silveira J. A. G., Ribeiro M. F. B., Cadioli F. A., et al. (2018). Clinical and epidemiological aspects and diagnosis of Trypanosoma vivax infection in a cattle herd, state of Maranhão, Brazil. Pesq. Vet. Bras. 38, 896–901. doi: 10.1590/1678-5150-PVB-5303
Reis M. O. (2021). Economic losses associated with Trypanosoma vivax infection in cattle in Minas Gerais (Lavras (MG: Federal University of Lavras).
Rodrigues C. M., Batista J. S., Lima J. M., Freitas F. J., Barros I. O., Garcia H. A., et al. (2015). Field and experimental symptomless infections support wandering donkeys as healthy carriers of Trypanosoma vivax in the Brazilian Semiarid, a region of outbreaks of high mortality in cattle and sheep. Parasit Vectors 8, 564. doi: 10.1186/s13071-015-1169-7
Rodrigues G. P., Fernandes B. D., Araújo B. V. S., Costa J. O. J., Silva M. M., Medeiros A. M., et al. (2024). Molecular diagnosis of the main hemoparasites of dairy cows in the state of Ceará. Genes (Basel) 15, 1369. doi: 10.3390/genes15111369
Silva R. A., Silva J. A., Schneider R. C., Freitas J., Mesquita D., Mesquita T., et al. (1996). Outbreak of trypanosomiasis due to Trypanosoma vivax (Ziemann 1905) in bovines of the Pantanal, Brazil. Mem. Inst. Oswaldo Cruz 91, 561–562. doi: 10.1590/s0074-02761996000500005
Stamatakis A. (2014). RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 30, 1312–1313. doi: 10.1093/bioinformatics/btu033
Keywords: trypanosomiasis, Brazilian northeast, epidemiological survey, molecular diagnosis, phylogenetic analysis
Citation: Lima Neto BF, Silva JO, Alvares FBV, Batista SP, Silva Filho GM, Medeiros GDA, Rodrigues CMF, Fermino BR, Vilela VLR and Feitosa TF (2025) Epidemiological assessment and outbreak report of Trypanosoma vivax in dairy cattle from the Brazilian Semiarid region. Front. Anim. Sci. 6:1679658. doi: 10.3389/fanim.2025.1679658
Received: 04 August 2025; Accepted: 25 September 2025;
Published: 20 October 2025.
Edited by:
Francesca Conte, University of Messina, ItalyReviewed by:
Armando Reyna-Bello, Universidad De las Fuerzas Armadas ESPE, EcuadorGilderlandio Rodrigues, Doctor Leão Sampaio University Center, Brazil
Copyright © 2025 Lima Neto, Silva, Alvares, Batista, Silva Filho, Medeiros, Rodrigues, Fermino, Vilela and Feitosa. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Vinícius Longo Ribeiro Vilela, dmluaWNpdXMudmlsZWxhQGlmcGIuZWR1LmJy