 Jan Szmek
Jan Szmek Michaela Englmaierová2†
Michaela Englmaierová2†- 1Department of Microbiology, Nutrition and Dietetics, Faculty of Agrobiology, Food and Natural Resources, Czech University of Life Sciences Prague, Suchdol, Prague, Czechia
- 2Department of Nutrition Physiology and Animal Product Quality, Institute of Animal Science, Uhříněves, Prague, Czechia
Hemp seed (HS) and flax seed (FS) are natural sources of bioactive compounds, including n-3 polyunsaturated fatty acids (PUFAs). The aim of this study was to investigate the effects of HS from the non-psychoactive variety Futura, non-extruded FS from the variety Floral, and their combination on the performance of laying hens and the quality and nutritional profile of the eggs produced. A total of 240 Lohmann Brown laying hens were randomly allocated to 4 dietary treatments, with 6 replicate cages and 10 hens per cage. The first (I) control diet contained no seed supplementation. The second (II) and third (III) diets were supplemented with 40 g/kg FS and 40 g/kg HS, respectively. The fourth (IV) diet included a combination of HS and FS (both at 40 g/kg). The inclusion of FS significantly increased both egg production and egg mass (p < 0.001). The lowest feed conversion ratio was observed in group II, which received FS alone. The addition of HS increased feed consumption per egg (p = 0.005) and total egg mass (p < 0.001). All the seed-supplemented groups exhibited increased egg weights. HS supplementation markedly increased yolk γ-tocopherol concentrations (p < 0.001) and reduced total cholesterol (p = 0.042) and malondialdehyde levels in fresh yolks (p = 0.003), indicating a potential improvement in oxidative stability. The combined supplementation with HS and FS (IV) significantly increased the total PUFA content (p = 0.004), including both n-6 (p < 0.022) and n-3 (p = 0.001) PUFAs. Among the n-3 PUFAs, the levels of α-linolenic and eicosapentaenoic acids were elevated. These changes contributed to an improved n-6/n-3 ratio, which was significantly affected by both HS and FS supplementation (p < 0.001). All dietary treatments significantly improved the lipid quality indices (p < 0.001). These findings highlight the potential of HS and FS as functional feed ingredients, with FS mainly enhancing hen performance and HS improving the nutritional quality of eggs through bioactive compound enrichment.
1 Introduction
Over the years, interest has dramatically increased in using animal-derived food products to enhance human nutrition and health (Cuchillo-Hilario et al., 2024). The enrichment of animal products with bioactive compounds, particularly n-3 polyunsaturated fatty acids (PUFAs), has attracted growing attention, largely due to increasing consumer awareness of the relationship between diet and health (Palmquist, 2009; Mandal et al., 2014). The fatty acid (FA) composition of animal-derived foods, such as egg yolks, closely reflects the lipid profile and composition of the animals’ diet (Taaifi et al., 2023).
α-linolenic acid (ALA, C18:3, n-3), the main n-3 PUFA, is an essential nutrient for both animals and humans (Jing et al., 2017). ALA and its long-chain derivatives, eicosapentaenoic acid (EPA, C20:5, n-3) and docosahexaenoic acid (DHA, C22:6, n-3), are FAs required for normal development and optimal health maintenance (Farinon et al., 2020) and have been shown to reduce the risk of cardiovascular and allergic diseases (van den Elsen et al., 2012), as well as inflammatory conditions (Honda et al., 2015). Furthermore, the n-6/n-3 PUFA ratio remains a critical indicator of fat quality and is recommended to be below 4 (Simopoulos, 2008). The World Health Organization (WHO) recommended sufficient intake of n-3 PUFAs, particularly EPA and DHA, which are found primarily in fish and seafood products (Cardoso et al., 2016). However, this recommendation is not applicable in low-income countries, where fish consumption is low because of limited availability and high prices (Neori and Agami, 2024). In contrast, eggs are consumed more frequently in almost all countries around the world because of their availability and low cost (Taaifi et al., 2023). Therefore, enriching eggs with n-3 PUFA-rich ingredients represents a sustainable and practical strategy to support a healthy, well-balanced diet while simultaneously reducing pressure on marine resources (Kumar et al., 2019). Good sources of n-3 PUFAs are seeds and their oils (Jing et al., 2017), which may include hemp seed (HS) and flax seed (FS).
HS and FS are natural sources of various health-promoting bioactive compounds, and it is hypothesized that they may exert distinct effects on the performance of laying hens, as well as the quality and nutritional profile of the eggs produced.
Hemp (Cannabis sativa L.) has recently garnered attention in animal nutrition because of its rich nutraceutical properties (Incharoen et al., 2025). Industrial hemp cultivars bred to contain less than 0.2% Δ9-tetrahydrocannabinol (THC) have attracted renewed interest in the nutritional value of HS and its by-products (e.g., hemp seed oil – HO) as promising dietary ingredients for livestock, including poultry (Gakhar et al., 2012; Neijat et al., 2014; Farinon et al., 2020; Shariatmadari, 2023). Whole HS typically contains 25–35% lipids with a unique and well-balanced FA composition (Farinon et al., 2020). The high PUFA content of HO is characterized by a favorable ratio of approximately 3:1 between linoleic acid (LA, C18:2, n-6, ~60%) and ALA (~19%; Parker et al., 2003). This ratio makes HO a desirable ingredient for supporting development and health in both animals (Yao et al., 2012) and humans (Siano et al., 2019). Moreover, multiple studies have consistently shown that the inclusion of HS or HO alters the FA profile of egg yolks in a consistent manner: monounsaturated fatty acids (MUFAs) tend to decrease, whereas essential FAs and their long-chain PUFA derivatives tend to increase (Silversides and LefranÇois, 2005; Gakhar et al., 2012; Halle and Schöne, 2013; Neijat et al., 2016). These dose-dependent effects may have positive implications for human health (Fraeye et al., 2012). HS is also rich in tocopherols (Farinon et al., 2020). Tocopherols are commonly supplemented in laying hen diets to improve the oxidative stability of n-3 PUFA-enriched eggs (Ren et al., 2013). When HS is included in the diets of laying hens, both dietary α-tocopherol and γ-tocopherol are largely transferred to the egg yolks (Skřivan et al., 2019), further increasing their antioxidant potential. As comprehensively reviewed by Szmek et al. (2025), hemp and its products possess a unique and well-balanced composition of the aforementioned bioactive compounds, which are associated with remarkable antioxidant and health-promoting properties.
Flax (Linum usitatissimum L.) is another oilseed that has attracted considerable worldwide attention because of the high concentration of n-3 PUFAs, especially ALA, in its seeds (Shim et al., 2015). Whole FS typically contains 30–41% lipids (Coşkuner and Karababa, 2007). Flax seed oil (FO) is the most valuable fraction of FS. FS lipids consist largely of PUFAs, such as ALA (~55%) and LA (~14%), and a moderate proportion of MUFAs, primarily oleic acid (C18:1, n-9, ~18%; Bekhit et al., 2018). Numerous studies have shown that including FS in poultry diets can increase bird performance and improve product quality (Azcona et al., 2008; Mridula et al., 2014; Alagawany et al., 2019). Moreover, increasing dietary FO levels in laying hen diets allows dietary n-3 PUFAs to be efficiently absorbed, transferred, and deposited into the egg yolks (Ehr et al., 2017), thereby offering a route to deliver these FAs to humans. This approach is also consistent with the United Nations (UN) Sustainable Development Goals (SDG), particularly SDG 2, SDG 3, and SDG 12, by contributing to improved nutrition, healthier food products, and more sustainable use of agricultural resources (United Nations, 2015).
In this context, the present study aimed to investigate the dietary effects of HS from the non-psychoactive variety Futura, non-extruded FS from the variety Floral, and their combination on laying hen performance and egg characteristics. To comprehensively evaluate egg quality and nutritional composition, this study assessed physical parameters, egg yolk concentrations of lipophilic vitamins and β-carotene, FA profiles with related nutritional indices and cholesterol content, and the oxidative stability of egg yolk lipids, as indicated by malondialdehyde (MDA) levels. Unlike previous studies, which have mainly focused on either HS or FS supplementation alone, this work provides a direct comparison of their individual and combined dietary effects. Furthermore, by using specific seed varieties and evaluating a broad spectrum of nutritional and functional egg traits, this study provides a novel and more integrated perspective on the potential of these oilseeds in laying hen nutrition.
2 Materials and methods
All experimental procedures in this study complied with the guidelines of the Ethics Committee of the Central Commission for Animal Welfare at the Ministry of Agriculture of the Czech Republic (Prague, Czech Republic) and with Directive 2010/63/EU of the European Parliament and the Council on the protection of animals used for scientific purposes. The research protocol (code 03/2022) was reviewed and approved by the Ethical Committee of the Institute of Animal Science (Prague – Uhříněves, Czech Republic).
2.1 Experimental design, management of laying hens, and dietary treatments
The experiment was performed with a total of 240 Lohmann Brown laying hens aged 50 weeks. The laying hens were housed in three-tier enriched cages within the same air-conditioned facility and randomly allocated to 4 dietary treatments, with 6 replicate cages and 10 hens per cage. The 2 × 2 factorial experiment included two levels of non-psychoactive HS (Futura variety; 0 and 40 g/kg) and two levels of non-extruded FS (Floral variety; 0 and 40 g/kg) in the diet. The first (I) control diet consisted of basal wheat–maize–soybean meal mixed feed without HS and FS. The second (II) and third (III) experimental groups of laying hens were fed diets supplemented with 40 g/kg FS and 40 g/kg HS, respectively. The laying hens from the fourth (IV) experimental group received a diet containing a combination of the above seeds, specifically 40 g/kg HS and 40 g/kg FS. The experimental design and allocation of laying hens to dietary treatments are presented in Table 1. Each enriched cage had an area of 7 560 cm2. The stocking density, cage equipment (i.e., feeders, drinkers, nests, and perches), lighting, ventilation, and vaccination complied with the requirements of the European Council Directive 1999/74/EC (European Union, 1999). The technological and microclimatic housing conditions complied with the standard procedure for the Lohmann Brown hybrid, with an average temperature of 20–22 °C and humidity between 50 and 60%. The light cycle was 14 hours of light and 10 hours of darkness, with a light intensity of approximately 10 lux at the middle tier of the three-tier enriched cage system. The ingredients and chemical compositions of the experimental laying hen diets, HS, and FS are presented in Table 2 and Table 3. All the diets were formulated to achieve comparable levels of metabolizable energy (~11.50 MJ/kg) and crude protein (~162 g/kg). Feed and fresh water complying with hygiene standards were supplied ad libitum. The experiment lasted for 12 weeks, including a two-week preparation period. The health status of the laying hens and the number of eggs laid were monitored daily. The hen-day egg production (%), feed intake (g/hen/day and g/egg), and feed conversion ratio (FCR; kg/kg) were calculated weekly for each cage and group. Egg weights (g) were recorded three times per week for all eggs in each replicate. The eggs used for chemical analyses of the yolk were collected at the end of the experiment, when the laying hens were at 61 weeks of age. Three eggs from each replicate were homogenized to form one sample (n = 6).
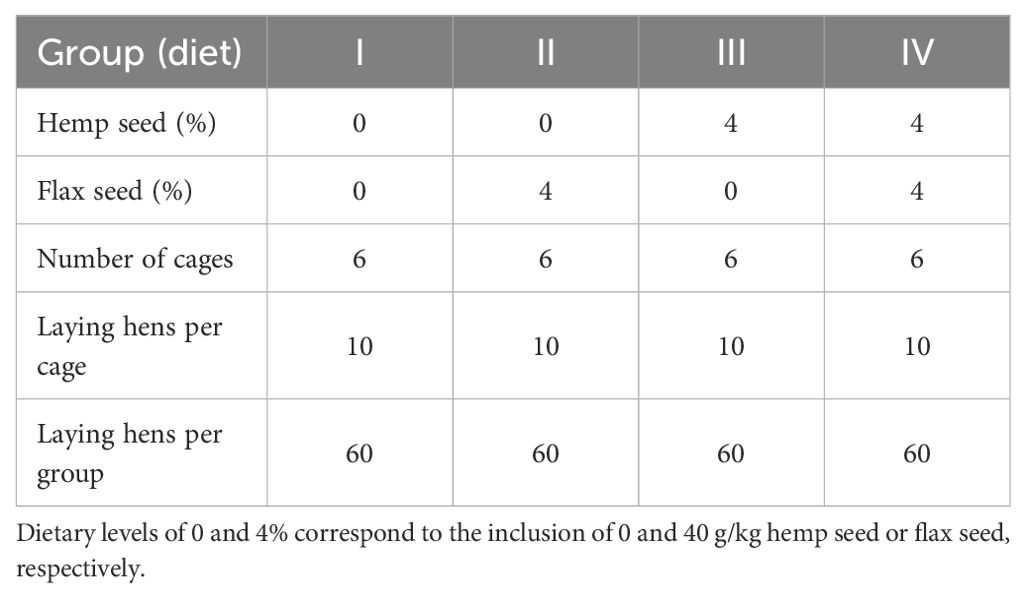
Table 1. Experimental design and allocation of laying hens to dietary treatments.
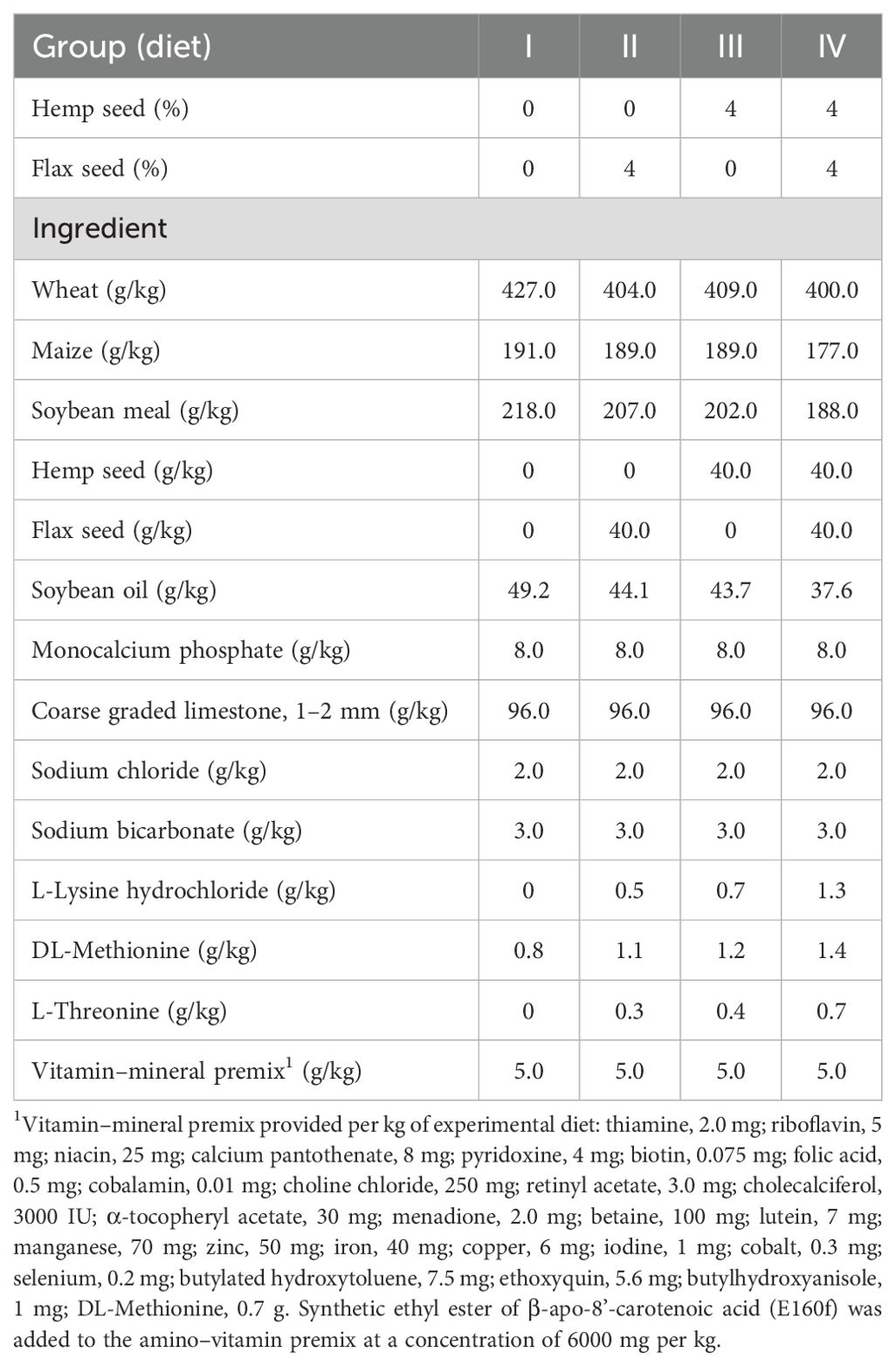
Table 2. Ingredient composition of the experimental diets of the laying hens.
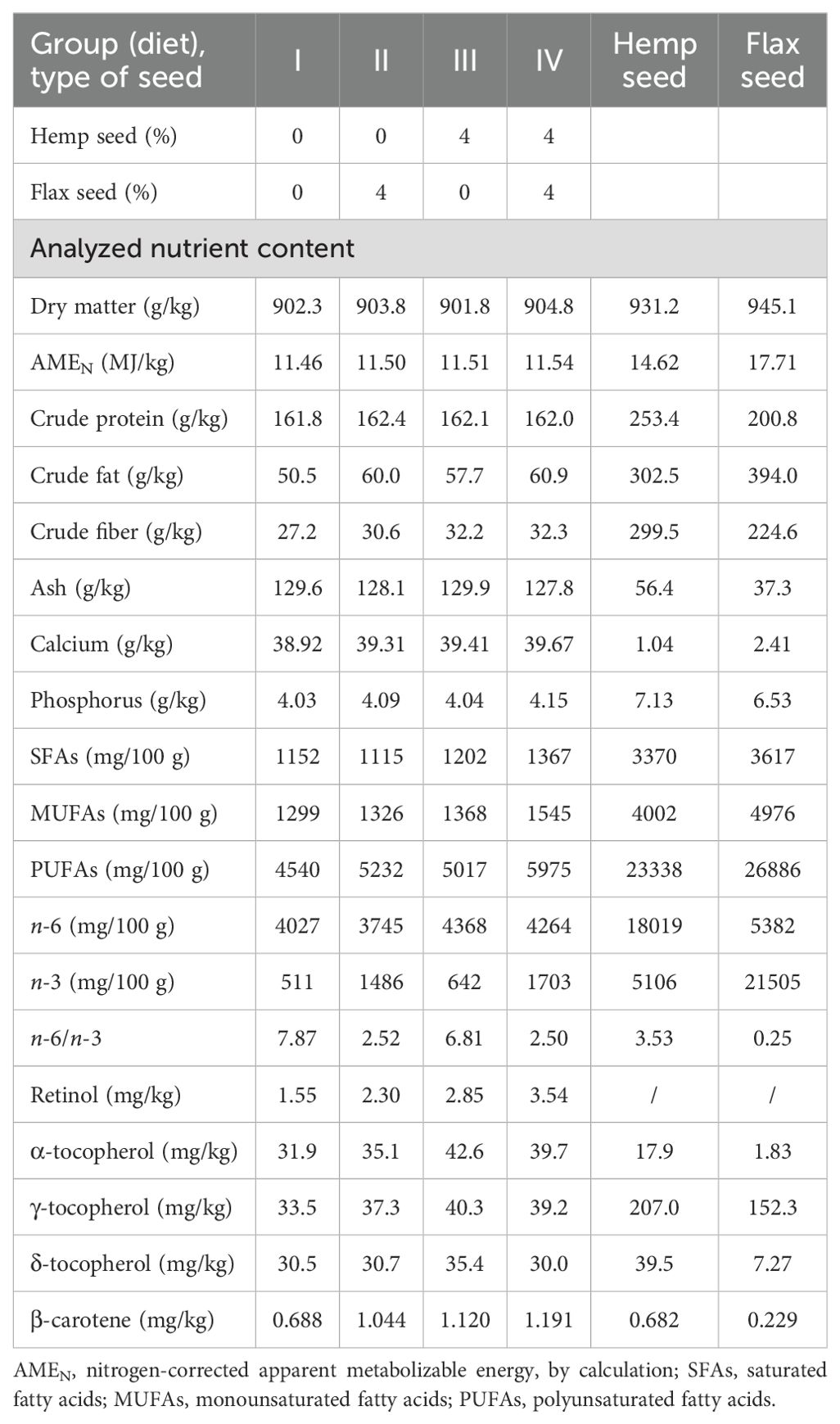
Table 3. Chemical composition of the experimental diets of the laying hens, hemp seed, and flax seed.
2.2 Physical analysis of eggs
For the analysis of physical parameters, a whole day of egg production was analyzed in the laying hens at 61 weeks of age. A total of 219 eggs were analyzed. The values were averaged per cage (n = 6). The percentages of albumen, yolk, and shell were determined on the basis of the individual weight of each egg (g) and the corresponding weights of its components (g). The yolk was weighed after the chalazae were removed. All eggs and their components were weighed using a digital scale (KERN & SOHN GmbH, Balingen, Germany) with an accuracy of 0.1 g.
The Haugh units (HU), which assess the quality of albumen, were calculated on the basis of the method of Haugh (1937). The formula for calculating HU was as follows:
where H represents the height of the albumen (mm) and W represents the weight of the egg (g).
The color of the yolk was determined with a DSM yolk color fan (DSM Nutritional Products, Basel, Switzerland) ranging from 1–15 and expressed as the a* (redness) and b* (yellowness) values obtained with a Minolta CR-300 colorimeter (Konica Minolta, Osaka, Japan). On the CIELAB color space diagram, the a* and b* values indicate the red-green and yellow-blue components, respectively, of a color (CIE, International Commission on Illumination).
The breaking strength of the shell (g/cm2) was measured along the vertical axis with an Instron 3360 apparatus (Instron, Norwood, MA, USA). After the shell membranes were removed, the shell thickness (μm) was determined by averaging three measurements taken from the sharp and blunt ends and the shell equator using a digital micrometer (Schut Geometrical Metrology, Groningen, The Netherlands).
2.3 Laboratory analyses
Nutrient analyses of the experimental laying hen diets, HS, and FS were conducted according to the methods of the Association of Official Analytical Chemists (AOAC, 2005). The dry matter content (g/kg) of the diets, HS, and FS was determined by drying to a constant weight at 105 °C in an oven (Memmert ULM 500; Memmert, Schwabach, Germany). The crude protein content (g/kg) of the diets, HS, and FS was measured with a Kjeltec Auto 1030 instrument (Tecator, Höganäs, Sweden). The dry homogenized diets, HS, and FS were heated to 550 °C in a muffle furnace (LMH 11/12 with Ht40 AL temperature controller; LAC Asia Limited, Hong Kong, China), and the mineral ash was dissolved in 3 M hydrochloric acid (HCl). The calcium (Ca) concentration (g/kg) in the HCl extract was measured by atomic absorption spectrometry with a Solar M6 instrument (TJA Solutions, Cambridge, UK). The total phosphorus (P) concentration (g/kg) was determined in the HCl solution with a vanadate-molybdate reagent (AOAC, 2005; method No. 965.17).
The FA profiles (mg/100 g fresh weight) of the experimental laying hen diets, HS, FS, and egg yolks (n = 6) were determined following chloroform–methanol extraction of total lipids (Folch et al., 1957). The FAs underwent alkaline trans-methylation as described by Raes et al. (2003). The resulting FA methyl esters (FAME) were analyzed by gas chromatography (GC) with an HP 6890 chromatograph (Agilent Technologies, Inc., Santa Clara, CA, USA) equipped with a 60 m DB-23 capillary column (150–230 °C) and a flame ionization detector (FID), with split injections carried out with an Agilent autosampler. One-microliter FAME samples in hexane were injected in split mode (split ratio 1:40) with nitrogen (N2) as the carrier gas at a total flow rate of 35.2 mL/min, using hydrogen (H2) and air for flame ionization detection. The column temperature program was as follows: initially held at 60 °C for 7 minutes, then increased by 20 °C/min to 110 °C, which was held for 4 minutes, followed by a 10 °C/min increase to 120 °C, which was held for 4 minutes, then increased by 15 °C/min to 170 °C, followed by a 2 °C/min increase to 210 °C, which was held for 13.5 minutes, and finally, the temperature was increased by 40 °C/min to 230 °C and maintained for 7 minutes. FAs were identified by comparing their retention times with those of reference standards PUFA No.1 (cat. no. 47033), PUFA No.2 (cat. no. 47015-U), PUFA No.3 (cat. no. 47085-U), and a 37-component FAME mixture (Supelco, Bellefonte, PA, USA). For quantification, nonadecanoic acid (C19:0; cat. no. N5252, Sigma-Aldrich, Prague, Czech Republic) was used as an internal standard.
The concentrations of retinol, α-tocopherol, γ-tocopherol, δ-tocopherol, and β-carotene (mg/kg) in the experimental laying hen diets, HS, FS, and lyophilized egg yolks (n = 6) were determined with a Shimadzu high-performance liquid chromatography (HPLC) system (VP series; Shimadzu, Kyoto, Japan) equipped with a diode array detector (HPLC-DAD). Samples underwent alkaline saponification with 60% aqueous potassium hydroxide (KOH) solution and diethyl ether extraction, following the EN 12822-1 (2000), EN 12823-1 (2000), and EN 12823-2 (2000) European standards. Chromatographic analyses were performed with a Phenomenex Synergi 4 μm Fusion-RP 80 Å column (150 × 4.6 mm, 4 μm; P/No. 00F-4424-E0, Torrance, CA, USA) using methanol as the mobile phase. The method employed a gradient flow program, where the solvent composition remained constant and the flow rate was adjusted from 0.6 mL/min to 1.5 mL/min and back to 0.6 mL/min. A 50 μL sample injection volume was used. Quantification was performed using individual calibration curves for each compound. Detection wavelengths (λ) were set as 325 nm for retinol, 292 nm for α-tocopherol, 296 nm for γ-tocopherol, 296 nm for δ-tocopherol, and 450 nm for β-carotene. The concentrations of lipophilic vitamins and β-carotene in the experimental laying hen diets, HS, and FS were expressed as mg/kg fresh weight, whereas in egg yolks they were expressed on a dry matter basis (i.e., mg/kg DM).
The cholesterol concentration (g/kg egg yolk) in egg yolks (n = 6) was determined after lipid saponification and extraction of the unsaponified fraction with diethyl ether, following ISO 3596:2011. Silyl derivatives were prepared with trimethylchlorosilane (TMCS) and hexamethyldisilazane (HMDS) silylation reagents (Sigma-Aldrich, Prague, Czech Republic) and quantified by GC with a SAC-5 capillary column (Supelco, Bellefonte, PA, USA) operated isothermally at 285 °C. One-microliter samples were injected in split mode (split ratio 15:1) with N2 as the carrier gas at a total flow rate of 19.001 mL/min, using H2 and air for flame ionization detection (FID).
Lipid peroxidation levels in the egg yolks (n = 6) of fresh eggs and eggs stored for 28 days at 18 °C and 50–55% relative humidity on paper trays were measured by a modified method based on that outlined by Czauderna et al. (2011). For derivatization of malondialdehyde (MDA), egg yolk hydrolysates were acidified and reacted with 2,4-dinitrophenylhydrazine (DNPH) to hydrazone (i.e., MDA-DNPH), as described by Czauderna et al. (2011). Chromatographic analysis was performed with a Phenomenex C18 Synergi 2.5 μm Hydro-RP 100 Å column (100 × 3 mm, 2.5 μm; P/No. 00D-4387-Y0, Torrance, CA, USA) coupled with a Shimadzu HPLC system (VP series; Shimadzu, Kyoto, Japan) and a diode array detector (HPLC-DAD). The mobile phase consisted of solvent A (water-acetonitrile, 95:5) and solvent B (acetonitrile), using a gradient program from 90% A/10% B to 1% A/99% B within 23.5 min at a flow rate increasing from 1.10 to 1.40 mL/min, followed by column re-equilibration. The total run time was 26 min. Detection of MDA-DNPH derivatives was performed at λ = 303 nm. The oxidative stability of egg yolk lipids was expressed as the amount of MDA in milligrams per kilogram of fresh egg yolk (i.e., mg/kg egg yolk), determined in both fresh and stored eggs.
2.4 Calculations of nutritional indices
On the basis of the FA profiles determined, nutritional indices (i.e., index of atherogenicity – IA, index of thrombogenicity – IT, hypocholesterolemic/hypercholesterolemic ratio – HH, and index of peroxidizability – IP) related to the nutritional value of egg yolk lipids were calculated with the formulas provided below.
The formula for calculating the index of atherogenicity (IA) was as follows:
where lauric acid (C12:0), myristic acid (C14:0), and palmitic acid (C16:0) represent the main classes of saturated fatty acids (SFAs), and ΣUFA denotes the sum of unsaturated fatty acids (UFAs), including MUFAs and PUFAs.
The formula for calculating the index of thrombogenicity (IT) was as follows:
where myristic acid (C14:0), palmitic acid (C16:0), and stearic acid (C18:0) represent the main classes of SFAs; ΣMUFA represents the sum of MUFAs; and Σn-6 PUFA and Σn-3 PUFA denote the sums of n-6 and n-3 PUFAs, respectively.
The formula for calculating the hypocholesterolemic/hypercholesterolemic ratio (HH) was as follows:
where MUFAs are represented by oleic acid in the cis configuration (cis-C18:1), ΣPUFA denotes the sum of PUFAs, and lauric acid (C12:0), myristic acid (C14:0), and palmitic acid (C16:0) represent the main classes of SFAs.
The formula for calculating the index of peroxidizability (IP) was as follows:
where the peroxidizability of FAs increases with the number of double bonds, with highly UFAs being the most susceptible to oxidation.
2.5 Statistical analysis
The experimental data obtained were analysed with a two-way analysis of variance (ANOVA) with the general linear model (GLM) procedure in the Statistical Analysis System (SAS, 2013). The main effects were the level of HS in the diet, the level of FS in the diet, and the interaction effect between these factors (i.e., HS*FS). Differences between the groups were tested with Duncan’s multiple range test. The experimental unit was a cage (n = 6). The results in the tables are presented as the means (a,b,c,d), the standard error of the mean (SEM), and the probability (p) values. All differences were considered significant at p < 0.05.
Testing of significant differences was carried out according to the following mathematical statistical model:
where Yijk represents the trait value; μ represents the overall mean; αi represents the effect of HS content in the diet (i = 1 and 2; 0 and 4%, respectively); βj represents the effect of FS content in the diet (j = 1 and 2; 0 and 4%, respectively); γij represents the interaction effect between these factors (i.e., HS*FS); and eijk represents the random residual error.
3 Results
All the results of the experimental data evaluated were summarized and presented in the following tables. The ingredient compositions of the experimental laying hen diets are shown in Table 2. As shown in Table 3, both feed ingredients, HS and FS, demonstrated a nutritionally favorable composition. HS exhibited a notably higher crude protein content (253.4 g/kg) compared to that of FS (200.8 g/kg). In terms of the lipid profile, FS contained greater total PUFA content (26886 mg/100 g) than HS did (23338 mg/100 g), mainly because of its high concentration of n-3 PUFAs (21505 mg/100 g), which subsequently improved the lipid profile of both the diets and the egg yolks. However, HS had a more balanced n-6/n-3 PUFA ratio (3.53) compared to that of FS (0.25), which may be beneficial for achieving an optimal dietary FA balance. Moreover, HS was particularly rich in γ-tocopherol (207.0 mg/kg), an antioxidant isomer form of vitamin E, which was considerably more abundant than in FS (152.3 mg/kg), potentially increasing the oxidative stability of yolk lipids.
3.1 Performance characteristics of the laying hens
The performance characteristics of the laying hens are presented in Table 4. The inclusion of FS (40 g/kg) in the experimental laying hen diets significantly increased both egg production (p < 0.001) and egg mass (p < 0.001). In contrast, HS supplementation had no significant effect on egg production. The addition of HS (40 g/kg) to the diets resulted in increased feed consumption per egg produced (p = 0.005) and a greater total egg mass (p < 0.001). The highest overall feed intake (128.3 g/hen/day) was observed in laying hens whose diet (IV) was supplemented with both types of seeds, with a significant increase (HS*FS; p = 0.002) compared with that of the other groups. Additionally, laying hens fed the FS-enriched diet (II) presented a lower (p < 0.001) FCR (2.09 kg/kg) than those fed the combination of HS and FS (IV; 2.14 kg/kg) or the control diet (I; 2.17 kg/kg).
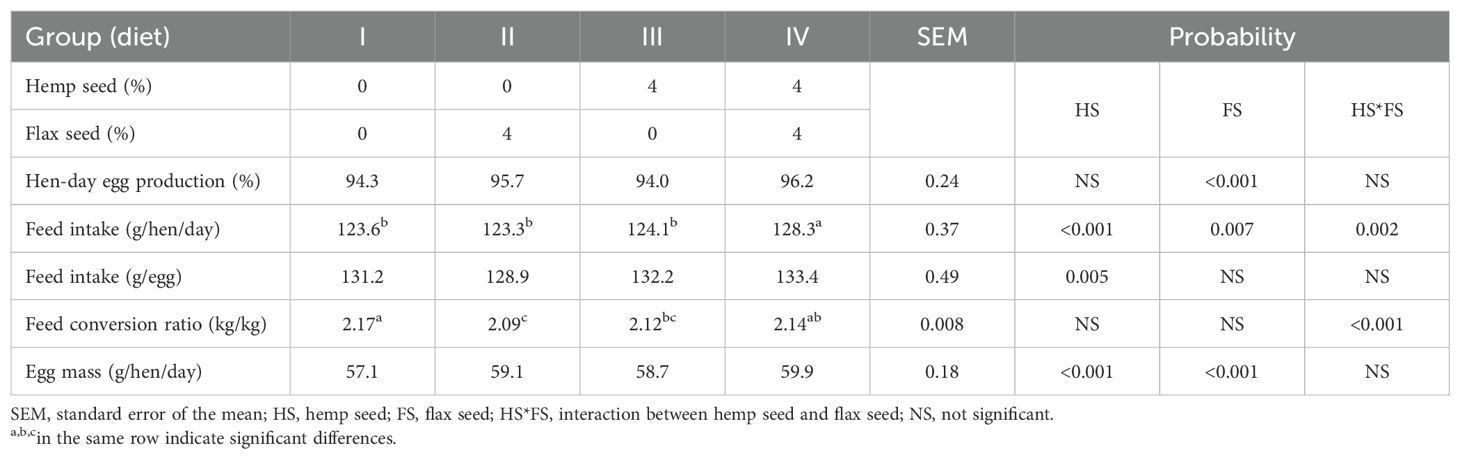
Table 4. Performance characteristics of the laying hens.
3.2 Physical characteristics of egg quality
As shown in Table 5, the mixed feed containing both HS and FS (IV) significantly increased egg weight (HS*FS; p = 0.002) by almost 2% compared with that of the control group (I). The highest egg weight (62.4 g) was observed in laying hens fed the diet (III) with HS supplementation alone, followed by the combination of HS and FS (IV; 62.3 g) and FS supplementation alone (II; 61.7 g), compared with that of the control group (I; 60.5 g). The experimental laying hen diets containing FS (II), HS (III), and a combination of HS and FS (IV) exhibited a marked increase in the percentage of egg albumen, which was significantly affected by the HS*FS interaction (p < 0.001). As a direct trade-off, the egg yolk proportion was reduced, with the highest egg yolk percentage (28.0%) observed in the control group (I) without the addition of seeds (p = 0.001). Compared with the control group (I), all the seed-supplemented groups (i.e., II, III, and IV) showed a reduction in egg yolk yellowness (b* values), with significant effects of HS (p = 0.001) and the HS*FS interaction (p = 0.028). On the other hand, the combined supplementation with HS (40 g/kg) and FS (40 g/kg) in the diet (IV) resulted in a significantly higher shell percentage (p = 0.036). Shell thickness and breaking strength, however, remained unaffected.
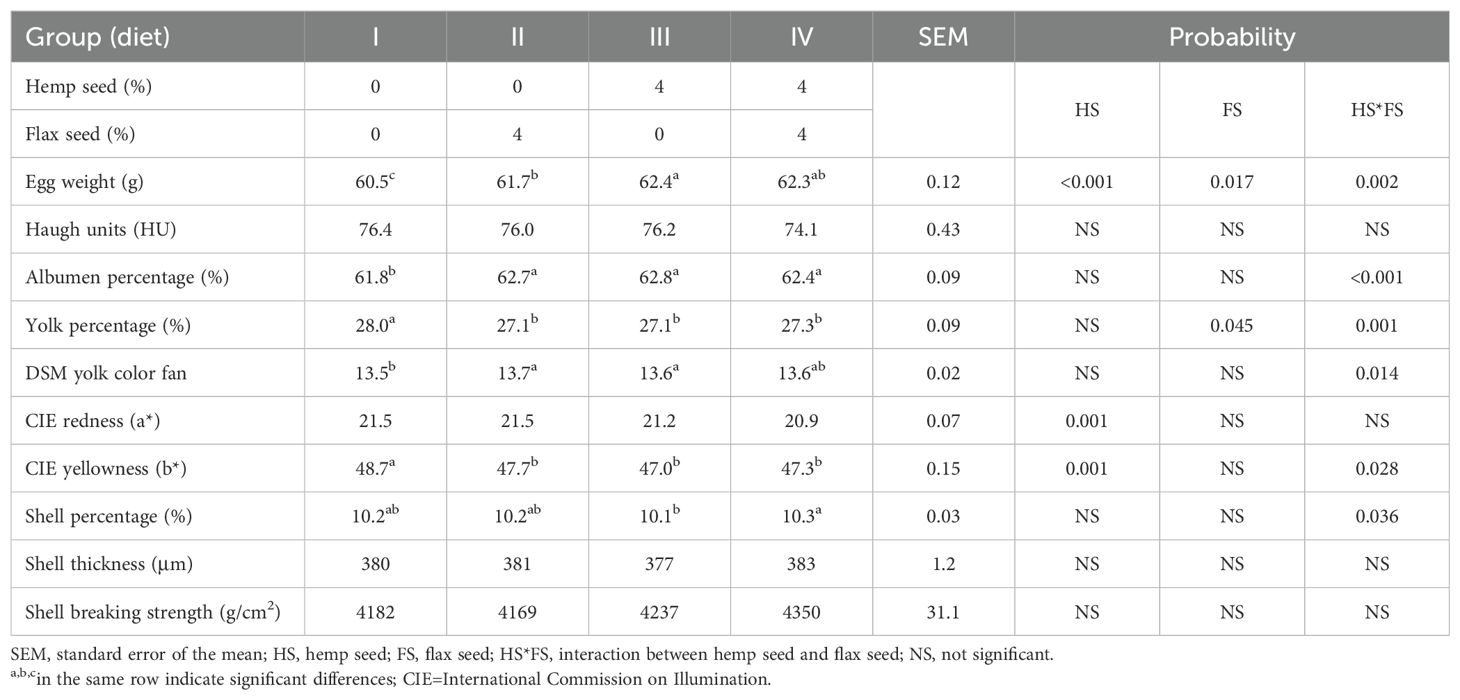
Table 5. Physical characteristics of egg quality.
3.3 Lipophilic vitamin and β-carotene concentrations in the egg yolks
The concentrations of lipophilic vitamins and β-carotene in egg yolks are summarized in Table 6. The inclusion of 40 g/kg HS in the experimental diets of laying hens resulted in elevated levels of γ-2tocopherol (p < 0.001). As shown in Table 3, HS was the dominant source of this vitamin E isomer. The concentration of δ-tocopherol in egg yolks was negatively affected by both HS (p = 0.006) and FS (p < 0.001) supplementation. A significant HS*FS interaction (p = 0.029) was detected in the β-carotene content of egg yolks. The control group (I) showed the highest β-carotene level (0.387 mg/kg DM), whereas the lowest (0.312 mg/kg DM) was recorded in group III, which received HS alone. No significant effects of dietary treatments were observed for the retinol or α-tocopherol concentrations.
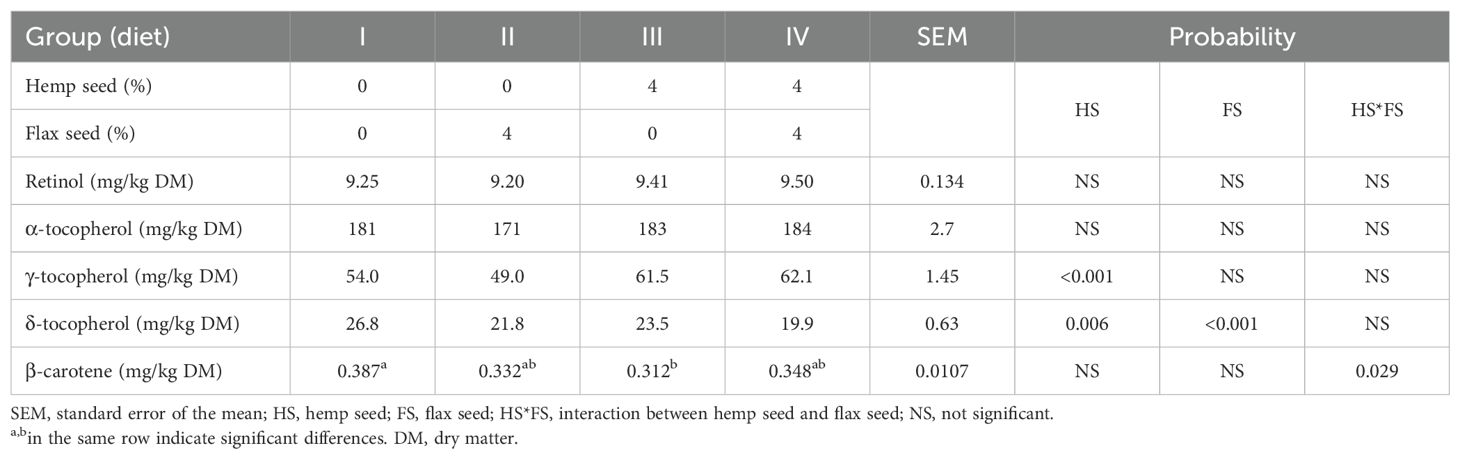
Table 6. Lipophilic vitamin and β-carotene concentrations in the egg yolks.
3.4 Fatty acid composition, nutritional indices, and cholesterol concentrations in the egg yolks
As reported in Table 7, the dietary inclusion of FS (II), HS (III), or their combination (IV) had a significant effect on the FA profile, nutritional indices, and cholesterol content of egg yolks.
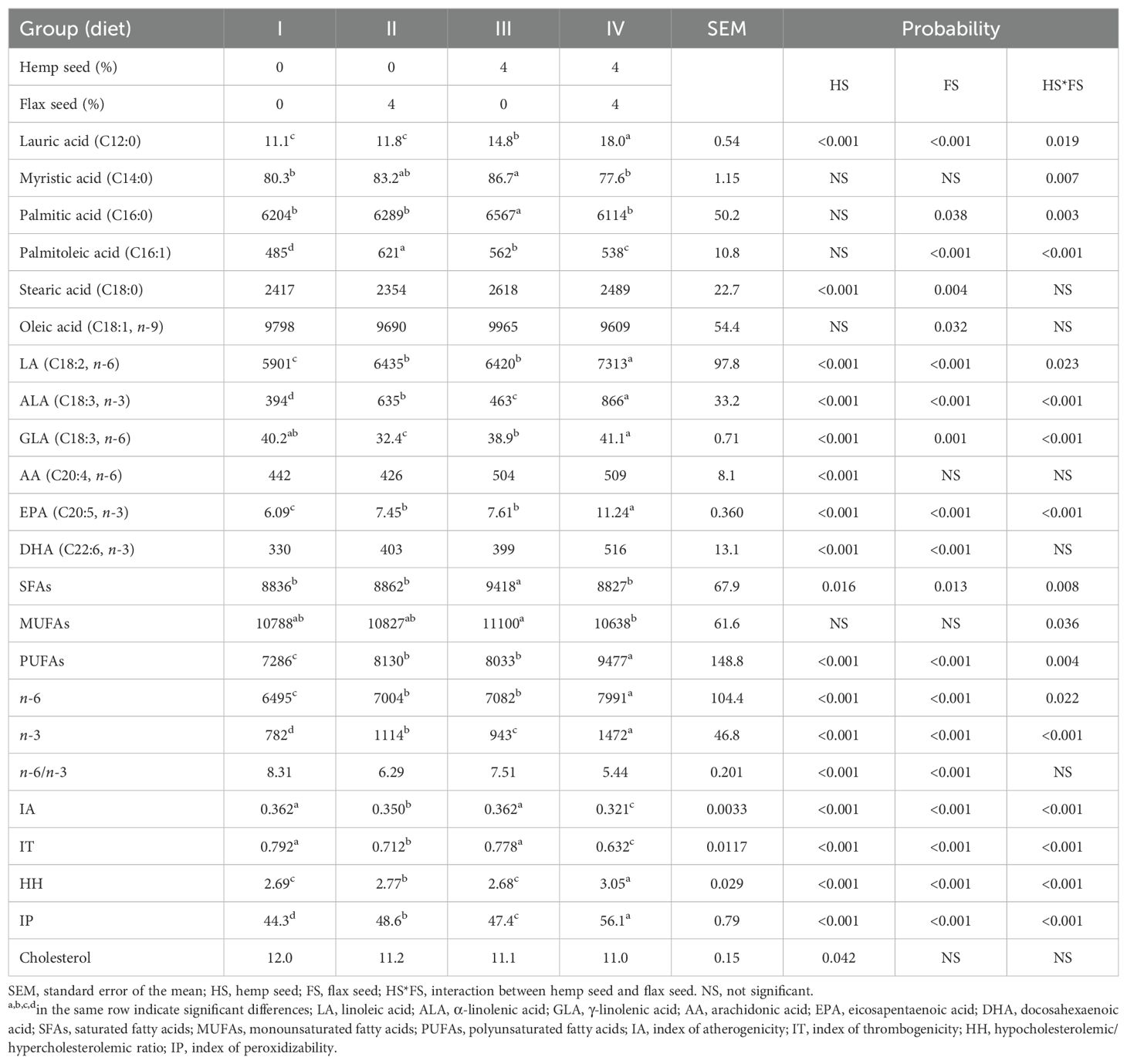
Table 7. Fatty acid composition (mg/100 g), nutritional indices, and cholesterol concentrations (g/kg) in the egg yolks.
The addition of HS (40 g/kg) and FS (40 g/kg) to the diet (IV) of laying hens significantly reduced (HS*FS; p = 0.008) the total SFA content (8827 mg/100 g) compared with that of the diet (III) with HS supplementation alone (9418 mg/100 g). More specifically, both myristic and palmitic acid concentrations were significantly higher in group III than in the other groups. The combined supplementation with HS and FS (IV) mitigated this increase (HS*FS; p = 0.007 and p = 0.003, respectively), resulting in levels comparable to those of the control group (I). A similar trend (HS*FS; p = 0.036) was observed for the total MUFA content, which was lower in group IV (10638 mg/100 g) than in group III (11100 mg/100 g), which received HS alone. The total PUFA levels were significantly increased in all the seed-supplemented groups (i.e., II, III, and IV) compared to the control group (I), with the highest concentration found in group IV (9477 mg/100 g), which received the combination of HS and FS (HS*FS; p = 0.004). The increase was primarily due to a substantial increase in ALA, with the highest level in group IV (866 mg/100 g), reflecting the strong n-3 enrichment effect of FS (p < 0.001). As shown in Table 3, FS contained a 4.2-fold higher concentration of n-3 PUFAs than did HS. Furthermore, supplementation with HS and FS significantly increased (p < 0.001) the levels of EPA and DHA. In the yolk fat of laying hens fed the combination of both seeds (IV), the concentrations of ALA, EPA, and DHA were 2.2-, 1.8-, and 1.6-fold higher, respectively, than those in the control group (I). These results contributed to an improved n-6/n-3 ratio, which was significantly affected by both HS (p < 0.001) and FS (p < 0.001) supplementation. The ratio of pro-inflammatory n-6 PUFAs to anti-inflammatory n-3 PUFAs in egg yolks decreased from 8.31 (I) to 5.44 (IV).
All dietary treatments also affected lipid quality indices. Both the IA and IT were significantly reduced (p < 0.001) in the FS-supplemented groups (i.e., II and IV), indicating improved nutritional quality of yolk lipids. The lowest IA (0.321) and IT (0.632) values were observed in laying hens fed a diet (IV) containing 40 g/kg HS and 40 g/kg FS. The HH ratio was significantly increased in group IV (p < 0.001), indicating a more favorable lipid profile in terms of cholesterol metabolism. In contrast, the IP was also significantly elevated in this group (IV; p < 0.001), reflecting a higher PUFA content and thus greater susceptibility of yolk lipids to oxidative degradation.
Finally, supplementation with 40 g/kg HS significantly reduced (p = 0.042) the total cholesterol content in egg yolks.
3.5 Oxidative stability of the egg yolks
The oxidative stability of egg yolks, expressed as the MDA concentration (mg/kg egg yolk), is shown in Table 8. In fresh eggs, the inclusion of HS (40 g/kg) in the diets of laying hens significantly reduced MDA levels (p = 0.003), indicating improved oxidative stability of yolk lipids. However, no significant differences were detected among the groups after 28 days of storage.
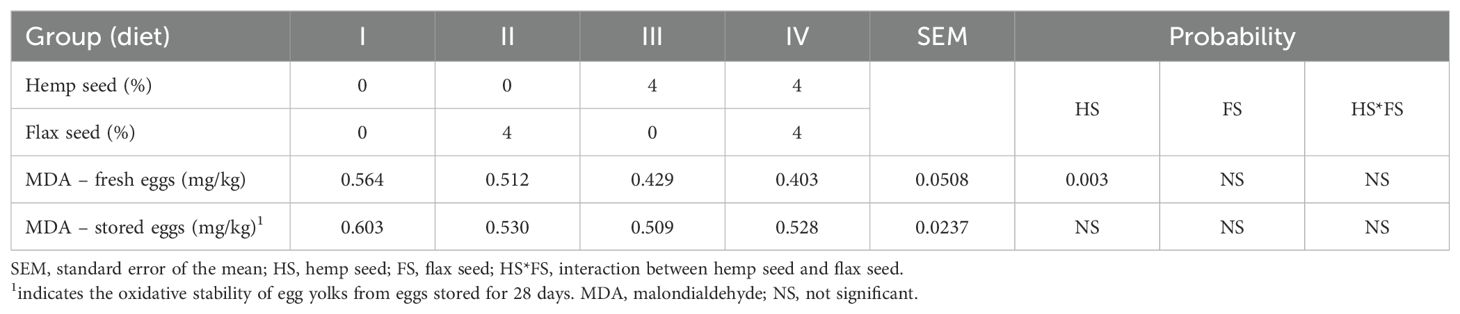
Table 8. Oxidative stability of the egg yolks expressed as malondialdehyde values.
4 Discussion
Eggs are an affordable and highly nutritious food that provides an excellent source of high-quality proteins and essential fats (Réhault-Godbert et al., 2019). They are also widely recognized as a valuable source of several fat-soluble and water-soluble vitamins (Adeoye et al., 2023). Owing to their rich nutritional profile, eggs are considered an ideal medium for delivering additional health-promoting bioactive compounds, the presence of which can be significantly modulated by the diet of laying hens. In particular, growing interest in n-3 PUFAs in relation to human health and well-being has driven research aimed at enriching the n-3 PUFA content of eggs through dietary supplementation with n-3 PUFA-rich ingredients (Usturoi et al., 2025). In the present study, HS from the non-psychoactive variety Futura, non-extruded FS from the variety Floral, and their combination were tested as supplementary feed ingredients in laying hen diets.
As shown in Table 3, both types of seeds (i.e., HS and FS) exhibited favorable nutritional profiles, particularly in terms of their crude protein content, the presence of specific PUFA groups, and the concentrations of individual tocopherol isomers. Numerous studies have evaluated the nutritional quality of HS and its by-products (e.g., hemp seed meal – HM), which have identified them as excellent sources of protein and energy (Gakhar et al., 2012; Neijat et al., 2014; Farinon et al., 2020; Shariatmadari, 2023). In addition to their nutritional profile, hemp by-products and extracts have gained popularity because of their high contents of PUFAs, proteins and cannabinoids, which offer considerable benefits for dietary applications and possess antimicrobial properties that are effective against bacterial and fungal pathogens (Ostapczuk et al., 2021; Meffo Kemda et al., 2024; Paulová et al., 2025). The crude protein content of HS, as determined by our laboratory analysis, aligns with previous findings (Vonapartis et al., 2015; Lan et al., 2019; Mohamed et al., 2024). According to Farinon et al. (2020), whole HS typically contains 20–25% proteins, which are highly digestible and rich in essential amino acids (EAAs). With respect to protein quality, lysine is considered the first limiting EAA in HS proteins for most animal species (House et al., 2010). The formulation of laying hen diets with up to 20% HM demonstrated that the protein and energy provided by HM can support the production performance of laying hens over a 4-week study period (Silversides and LefranÇois, 2005). Similarly, Gakhar et al. (2012) confirmed the efficacy of whole HS and HO in supporting egg production in laying hens over a 12-week feeding period. In line with these findings, Neijat et al. (2014) reported no significant differences in overall laying hen performance when graded levels of either HS (up to 30%) or HO (up to 9%) were included in the diet, compared with laying hens fed the control diet. However, Skřivan et al. (2019) reported reduced performance in laying hens when the HS concentration exceeded 30 g/kg. In the present study, the inclusion of HS did not increase overall hen-day egg production. However, it led to elevated feed consumption per egg produced, greater total egg mass (Table 4), and greater egg weight (Table 5). On the other hand, the inclusion of FS in the experimental diets of laying hens significantly increased both egg production and egg mass. In addition, the diet (II) with FS supplementation alone resulted in the lowest FCR among all dietary treatments (Table 4). FS is rich in lipids and provides high levels of n-3 PUFAs, particularly ALA (Shim et al., 2015), which increases the energy density of the feed. This may lead to reduced feed intake while sustaining production levels, thereby directly improving FCR. However, the experimental results concerning the effects of FS on the performance characteristics of laying hens are not consistent. Bean and Leeson (2003) reported that laying hens can tolerate up to 15% flax seed meal (FM) without any negative effects on egg production, egg weight, or feed efficiency, in both brown- and white-egg laying hens. Similarly, Attia et al. (2022) reported that diets containing up to 12% soaked FM did not negatively affect the laying rate, egg weight or mass, or FCR. In contrast, other studies have shown that FS inclusion at levels up to 15% can impair the production performance of laying hens (Aymond and van Elswyk, 1995; Antruejo et al., 2011; Halle and Schöne, 2013). Long-term FS supplementation has also been associated with reductions in egg production and impaired shell quality (Jia et al., 2008). Furthermore, dietary inclusion of extruded FS at levels above 20% has been shown to reduce laying performance, despite extrusion significantly lowering the anti-nutritional compound content (Imran et al., 2015). Anti-nutritional compounds represent a limiting factor in the use of oilseeds in poultry diets, with HS being no exception (Galasso et al., 2016). However, as noted by Skřivan et al. (2020), the negative effects of these compounds appear to be more pronounced in FS than in HS. Despite its nutritional benefits, FS contains several anti-nutritional compounds, such as phytic acid, oxalates, cyanogenic glycosides, pyridoxine antagonists (e.g., linatine), trypsin inhibitors, condensed tannins, soluble and insoluble non-starch polysaccharides, and mucilage (Katare et al., 2012; Shim et al., 2015; Russo and Reggiani, 2016; Bekhit et al., 2018), which may adversely affect nutrient availability and digestibility (Leeson et al., 2000; Rodríguez et al., 2001). These factors must be carefully considered when FS is used in animal feed or as a food additive in the food industry.
As previously mentioned, the highest egg weight observed in our experiment was recorded in laying hens fed the diet (III) supplemented exclusively with HS (Table 5). An increase in egg weight is generally associated with an age-related increase in egg yolk weight, often at the expense of other egg components (Johnston and Gous, 2007). Interior egg quality, particularly yolk weight, can also be influenced by the dietary lipid content (Ayerza and Coates, 1999). Although the inclusion of oilseeds in the experimental laying hen diets did not positively affect the egg yolk percentage, a noticeable increase in the egg albumen percentage was observed in all the seed-supplemented groups (i.e., II, III, and IV). Neijat et al. (2014) reported that a 30% inclusion rate of HS, which partially replaced soybean meal and corn oil in the diet of laying hens, substantially lowered egg weight compared with 10% and 20% inclusion levels, although total egg production and feed intake remained unaffected. Nevertheless, several other studies have provided evidence of a positive effect of HS and its by-products on egg weight (Gakhar et al., 2012; Halle and Schöne, 2013; Skřivan et al., 2019) and shell thickness (Konca et al., 2019). However, although the highest dietary concentration of HS (90 g/kg) used in the study by Skřivan et al. (2019) led to a reduction in shell thickness, no statistically significant effect was observed in our experiment, where shell thickness and breaking strength remained unaffected across all dietary treatments. Interestingly, a significantly higher shell percentage was observed in group IV, which received the combination of HS and FS, than in group III, which received HS alone. However, Panaite et al. (2021) reported no effect on shell percentage or breaking strength with 6% FM supplementation, although shell thickness increased significantly. With respect to yolk color, only a modest decrease in yellowness (b* values) was observed in all the seed-supplemented groups (i.e., II, III, and IV), with significant effects attributed to HS and the HS*FS interaction (Table 5). Yolk lipids contain carotenoids, which are key pigments responsible for yolk color intensity (Goldberg et al., 2012). The observed reduction in yellowness (b* values) may have been caused by increased utilization of carotenoids as powerful natural antioxidants to protect the UFAs present in the supplemented seeds. Specifically, lutein, zeaxanthin, and β-carotene have been shown to decrease lipid peroxidation in UFA-rich membranes and egg yolks, thereby improving oxidative stability and reducing MDA formation (Panaite et al., 2021; Dansou et al., 2023). However, in contrast to our findings, Goldberg et al. (2012); Konca et al. (2019), and Skřivan et al. (2019) reported increases in both redness (a*) and yellowness (b*) values following increased inclusion of HS and its by-products. In line with these observations, Halle and Schöne (2013) reported that FM increased yolk color more markedly than rapeseed meal or HM did.
HPLC-DAD analysis of tocopherols (i.e., vitamin E isomers) revealed that the main tocopherol with the highest concentration detected in both HS and FS corresponded to the γ-isomer (Table 3). According to Kriese et al. (2004), the predominant tocopherol isomer in HS is indeed γ-tocopherol (~22.41 mg/100 g), followed by α-tocopherol (~1.88 mg/100 g), δ-tocopherol (~1.50 mg/100 g), and β-tocopherol (~0.18 mg/100 g). While egg yolks are a valuable source of α-tocopherol (Skřivan et al., 2019), the amount of γ-tocopherol present is negligible (Guinaz et al., 2009). In our experiment, the inclusion of HS in the experimental laying hen diet (III) resulted in 61.5 mg/kg DM of γ-tocopherol in egg yolks (Table 6). In contrast, in the study by Skřivan et al. (2019), a considerably higher dietary inclusion level of HS (90 g/kg) resulted in a γ-tocopherol content of only 43.3 mg/kg DM in egg yolks. In our study, the vitamin–mineral premix provided to the experimental animals contained only α-tocopheryl acetate, which had no influence on the γ-tocopherol concentration. According to Wang et al. (2010), the main factors influencing the tocopherol content of egg yolks are nutrition, age, and laying hen variety or genotype. As noted by Skřivan et al. (2019), tocopherols in the diet originate exclusively from dietary ingredients, primarily from dietary fat sources such as rapeseed oil, which was used in their study. In our experimental laying hen diets, soybean oil served as the primary fat source. Data in the literature indicate that the γ-tocopherol concentration in soybean oil can reach approximately 273.3 ± 11.1 mg/kg (Grilo et al., 2014). Given that the experimental diet with HS (III) contained 43.7 g/kg soybean oil (Table 2), the theoretical contribution of γ-tocopherol from this fat source would be approximately 12 mg/kg of the feed. However, the actual γ-tocopherol concentration detected in the same diet (III) was 40.3 mg/kg, suggesting that the majority of this compound originated from the inclusion of HS (207.0 mg/kg; Table 3). This calculation supports the assumption that HS is a potent source of γ-tocopherol in poultry diets and can markedly increase its content in both feed and egg yolks. On the other hand, as the HS concentration in the study by Skřivan et al. (2019) increased in the feed of laying hens, the amounts of both α-tocopherol and γ-tocopherol increased in the egg yolks. Interestingly, the laying hens from this study also presented evidence of improved bone-breaking strength, as well as increased Ca concentrations in the tibia when HS was included in the diet at a rate of 9%. The role of tocopherols in bone metabolism further underscores the potential of HS and its by-products for improving the skeletal health of laying hens housed in both cage and non-cage production systems (Szmek et al., 2025). Moreover, a significant increase in total tocopherol content depending on the dose of HS incorporated was observed after 84 days of treatment varying from ~281.45 µg/g (control) to ~327.03 µg/g (HS 30%) in egg yolks (Taaifi et al., 2023). Tocopherols have well-known strong antioxidant properties, which can contribute to increasing the antioxidant potential and thus oxidative stability of the produced eggs. Among all isomers, γ-tocopherol is considered the most active antioxidant in lipophilic matrices (Porto et al., 2014). In contrast, α-tocopherol is considered the only tocopherol isomer with vitamin E activity in the human body (EFSA NDA Panel, 2015). Chen et al. (1998) demonstrated that α-tocopherol exerts a positive antioxidant effect on egg yolk stability at concentrations of 25, 45, and 50 µg/g yolk; however, at 75 µg/g and above, a prooxidant effect may occur. These effects, however, depend not only on dietary concentrations but also on how individual tocopherol isomers are absorbed and utilized by the organism. Although all tocopherol isomers are absorbed in the intestinal mucosa, excessive enrichment of α-tocopherol in the diet may reduce the absorption efficiency of other isomers (Wagner et al., 2004). Several studies have reported preferential retention of the α-isomer in both mammals (Behrens and Madère, 1986; Hayes et al., 1993) and poultry (Cherian et al., 1996). This phenomenon is attributed primarily to the presence of a specific hepatic α-tocopherol transfer protein (α-TTP), which selectively binds α-tocopherol and facilitates its incorporation into low-density lipoproteins (LDL), thereby increasing its distribution to peripheral tissues. Additionally, the existence of metabolic or transport mechanisms discriminating against the uptake and accumulation of γ-tocopherol may also contribute to this observed preference (Taaifi et al., 2023). Nevertheless, despite its relative low accumulation, γ-tocopherol has distinct physiological functions that are not shared with its α-isomer. In addition to its strong antioxidant properties, γ-tocopherol is also characterized by anti-inflammatory activity, particularly through the inhibition of prostaglandin-endoperoxide synthase (i.e., cyclooxygenase) activity (Jiang et al., 2001; Wagner et al., 2004; Singh et al., 2005). Finally, the increased concentration of γ-tocopherol in egg yolks from laying hens fed the HS-enriched diets observed in our experiment may be attributed to the antioxidant properties of carotenoids, which protect this lipophilic compound against oxidative degradation in the upper gastrointestinal tract (Mortensen et al., 2001). This may explain why the lowest recorded concentration of β-carotene occurred in group III (Table 6), suggesting that a greater proportion of this carotenoid was utilized for protective antioxidant functions.
From a nutritional standpoint, a highly significant and beneficial change was observed in the lipid profile of egg yolks (Table 7). Primarily, the simultaneous inclusion of HS and FS in the experimental laying hen diet (IV) resulted in a significant reduction in the total SFA content in egg yolks, particularly myristic and palmitic acids, compared with the diet (III) with HS supplementation alone. This reduction was most likely driven by the high ALA content in FS, which shifted the FA profile toward n-3 PUFAs and concomitantly decreased SFAs. This finding is supported by the results of Omri et al. (2019), who reported that the dietary inclusion of FS significantly reduced the egg yolk content of palmitic acid from 25.41% (control) to 23.43% (FS 4.5%) and that of stearic acid from 14.75% (control) to 12.53% (FS 4.5%). A similar trend in palmitic acid reduction was observed by Taaifi et al. (2023), who reported a slight decrease from 24.38% (control) to 22.46% in egg yolks from laying hens fed a diet containing 30% HS. However, the incorporation of non-industrial HS into the diet of laying hens did not significantly affect the total SFA rate (Taaifi et al., 2023). A similar trend to that observed for the total SFA content was also noted for the total MUFA content, with a reduction in group IV compared with that in group III. Significant reductions in total and individual MUFA levels have been reported in several studies evaluating the potential use of HS alone in laying hen diets (Shahid et al., 2015; Konca et al., 2019; Taaifi et al., 2023; Bošković Cabrol et al., 2025; Rbah et al., 2025). Nevertheless, in our case, these results were observed in the group (IV) of laying hens fed a diet enriched not only with HS but also with FS, which makes direct comparisons with the abovementioned studies somewhat limited. According to the GC analysis, the total PUFA content was greater in FS than in HS. This was mainly due to the high concentration of n-3 PUFAs in FS compared with that in HS (Table 3). The predominant n-3 PUFA was ALA, which is an essential nutrient in both birds and mammals, as it cannot be synthesized de novo and must be obtained from the diet (Whittle et al., 2024). Through desaturation and elongation processes, ALA can be converted into long-chain FAs such as EPA and DHA (Oliveira et al., 2010). These bioactive forms of FAs are crucial for numerous physiological functions and play a vital role in maintaining optimal health in both animals and humans (Jing et al., 2017; Farinon et al., 2020). Both the individual seed supplements (i.e., HS and FS) and their combination led to an increased content of n-6 and n-3 PUFAs in the egg yolks. The highest total PUFA content was found in group IV, which received the combination of HS and FS. More specifically, this increase was primarily due to a substantial increase in ALA, with the highest level in group IV, reflecting the strong n-3 enrichment effect of FS. The production of eggs enriched with n-3 PUFAs through dietary supplementation with FS as well as HS has been extensively studied in poultry. In particular, the inclusion of FS (Cherian and Quezada, 2016) and HS (Rbah et al., 2025) in laying hen diets led to increased deposition of the health-promoting ALA in the egg yolks. Moreover, according to Mattioli et al. (2017) and Omri et al. (2019), dietary inclusion of 10% and 4.5% FS, respectively, was shown to alter the FA composition of egg yolks and significantly increase the levels of EPA and DHA in eggs. In agreement with these findings, our study demonstrated that combined dietary supplementation with HS and FS (IV) substantially increased the deposition of n-3 PUFAs in egg yolks, with ALA, EPA, and DHA levels being 2.2-, 1.8-, and 1.6-fold higher, respectively, than those in the control group (I). The long-chain PUFA metabolism in animals allows the conversion of ALA to EPA and DHA only if the ratio of n-6 to n-3 is not too high; otherwise, the pathway converting LA to arachidonic acid (AA, C20:4, n-6) will be promoted. In general, n-3-derived lipid mediators are less bioactive and more anti-inflammatory, including more pro-resolving species, whereas n-6-derived lipid mediators tend to be more bioactive and more pro-inflammatory (Watkins et al., 2003; de Batlle et al., 2012). Therefore, beyond the absolute concentrations of PUFAs, the evaluation of the overall n-6/n-3 ratio is crucial, as it represents a key indicator of nutritional quality and plays an important role in the prevention of chronic degenerative diseases (Simopoulos, 2008). In our study, the ratio of pro-inflammatory n-6 PUFAs to anti-inflammatory n-3 PUFAs in egg yolks decreased from 8.31 (I) to 5.44 (IV). Comparable reductions in the n-6/n-3 ratio have also been reported in previous studies evaluating the individual supplementation of either HS or FS. For example, Al-Nasser et al. (2011) reported a significant decrease in this ratio in eggs from laying hens fed FS-enriched diets (5, 7.5, and 10%) after a 4-week feeding period. Likewise, Taaifi et al. (2023) reported a progressive decline in the n-6/n-3 ratio from 8.19 (control) to 4.89 in the group receiving 30% HS by day 84 of the experiment. Similarly, the findings of other studies consistently indicate that dietary supplementation with FS (Aguillón-Páez et al., 2020) and HS (Bošković Cabrol et al., 2025) can beneficially modulate the lipid profile of egg yolks and enrich them with n-3 PUFAs, respectively, which aligns with current nutritional recommendations.
Nutritional indices (i.e., IA, IT, HH, and IP) represent other indicators of fat quality, specifically egg yolk lipids, in terms of human health. The IA reflects the atherogenic potential of FAs, indicating their ability to promote the formation of fatty plaques in arteries (Ulbricht and Southgate, 1991). The IT characterizes the thrombogenic potential of FAs, indicating the tendency to form clots in blood vessels and describing the contributions of different FAs, particularly the balance between pro-thrombogenic FAs (i.e., SFAs) and anti-thrombogenic FAs (i.e., MUFAs and PUFAs; Ulbricht and Southgate, 1991). The consumption of foods or products with lower values of these indices can reduce total and LDL cholesterol levels in human blood plasma (Yurchenko et al., 2018) and thus contribute to better cardiovascular health (Chen and Liu, 2020). Although the IA and IT values in all our groups remained below the recommended thresholds (0.5 and 1, respectively), their further reduction in the FS-supplemented groups (i.e., II and IV) indicates an improvement in the nutritional quality of yolk lipids (Table 7). The lowest values observed in group IV may suggest a synergistic effect of combined HS and FS supplementation. In line with this finding, Omri et al. (2019) reported that feeding FS-supplemented diets significantly reduced the IT. Similarly, Mierliţă (2019) observed that the inclusion of HS and HM in the diet of laying hens decreased the value of IT, although it did not significantly affect the IA value or the HH ratio in egg yolks. The HH ratio, first proposed by Santos-Silva et al. (2002) and later optimized by Mierliţă (2018), is an index used to assess the effect of FA composition on cholesterol levels. In our experiment, the HH ratio was significantly increased in group IV, reflecting a more favorable balance between hypocholesterolemic FAs (i.e., cis-C18:1 and PUFAs) and hypercholesterolemic FAs, which is considered beneficial for cholesterol metabolism and cardiovascular health (Chen and Liu, 2020). The IP, developed by Holman (1954) and later refined by Laganiere and Yu (2004), assesses the susceptibility of egg yolk lipids to oxidation on the basis of their FA composition. In our study, the IP of the eggs was significantly elevated in the group (IV) of laying hens fed the diet supplemented with both types of seeds (i.e., HS and FS), although in this case, the effect was considered negative. This increase was likely attributable to the higher PUFA content in HS and particularly in FS (Table 3), as the degree of FA unsaturation directly influences the susceptibility of yolk lipids to peroxidation (Hulbert et al., 2007).
Finally, supplementation with HS significantly reduced the total cholesterol content in egg yolks (Table 7). This finding is consistent with previous studies by Shahid et al. (2015) and Skřivan et al. (2019), who reported reductions in egg yolk cholesterol from 19.27 mg/g (control) to 11.65 mg/g (HS 20%) and from 12.2 g/kg (control) to 10.6 g/kg (HS 9%), respectively. According to Rbah et al. (2024), the cholesterol-lowering effect is likely attributable to a combination of factors associated with the chemical composition of HS, particularly the presence of phytosterols, proteins, and insoluble fibers. Notably, phytosterols, especially β-sitosterol, are known for their hypocholesterolemic effects, primarily through the inhibition of intestinal cholesterol absorption (Mattson et al., 1982; Malini and Vanithakumari, 1990). They may also reduce endogenous cholesterol biosynthesis in the liver by modulating the activity of HMG-CoA reductase, thereby limiting cholesterol formation (Asem and Hertelendy, 1987). However, effectively reducing egg cholesterol through dietary modification, genetic selection, non-nutritive factors, or pharmacological agents remains challenging, as most experimental approaches have resulted in only minor reductions (< 10%; Puertas and Vázquez, 2018). This may be because laying hens typically meet their cholesterol requirements entirely through de novo synthesis (Elkin, 2006).
Lipid peroxidation, a process that compromises the nutritional and sensory quality of eggs, was assessed by determining MDA concentrations in egg yolks by HPLC-DAD analysis. MDA, a major oxidation product of PUFAs, is widely used as a marker of lipid peroxidation in food products. In fresh eggs, the inclusion of HS in the diets of laying hens significantly reduced MDA levels, indicating improved oxidative stability of yolk lipids (Table 8). A similar effect was reported by Mierliţă (2019), who reported the lowest MDA concentrations in egg yolks from laying hens fed HS (0.22 mg/kg in fresh eggs and 0.35 mg/kg in eggs after 30 days of storage). According to the author, the unexpectedly low lipid oxidation despite the elevated n-3 PUFA content in these eggs may be attributed to the relatively high concentration of tocopherols, particularly α-tocopherol, in the egg yolks. Other studies have also demonstrated that the oxidative stability of eggs is significantly influenced by the type and level of PUFAs, the α-tocopherol content, and the duration of storage (Chen et al., 1998; Galobart et al., 2001; Ren et al., 2013). Nevertheless, the explanation offered by Mierliţă (2019) may also apply to our findings, although the HS used in our experiment led specifically to a marked increase in γ-tocopherol levels in the egg yolks (Table 6). This tocopherol isomer is the predominant form of vitamin E in HS (Kriese et al., 2004) and is considered the most potent antioxidant present (Porto et al., 2014), capable of preventing peroxidation of n-3 PUFAs and thereby contributing to the increased oxidative stability of the produced eggs.
While numerous studies have evaluated the effects of either HS or FS supplementation alone, the present findings demonstrate that their combination produces distinct outcomes. Specifically, the simultaneous inclusion of HS and FS markedly enhanced the yolk deposition of n-3 PUFAs, particularly ALA and EPA, beyond the levels observed with either HS or FS alone. Moreover, the combined diet improved nutritional indices and reduced the n-6/n-3 ratio of the eggs, highlighting the synergistic effect between these two seeds. These results suggest that dual supplementation can overcome some of the limitations and inconsistencies previously reported in single-seed studies, thereby providing a novel dietary strategy for the enrichment of eggs with bioactive compounds. Nevertheless, certain limitations of the present study must be acknowledged. Only a single inclusion level of HS and FS was evaluated, and detailed egg quality and yolk lipid analyses were restricted to one time point (61 weeks of age). This inevitably constrains the generalizability of the findings across different production stages and supplementation regimens. In addition, while our results are broadly consistent with recent investigations into oilseed supplementation in laying hens, such as those by Bošković Cabrol et al. (2025); Rbah et al. (2025), and Saengsuwan et al. (2025) for HS and by Ehr et al. (2017); Mattioli et al. (2017), and Omri et al. (2019) for FS, direct comparisons remain limited due to differences in seed type, inclusion level, and experimental design.
5 Conclusion
The findings of this study indicate that HS and FS are promising supplementary feed ingredients, especially because of their beneficial effects on the performance of laying hens and their enriching influence on egg quality and nutritional value. Supplementation with HS (40 g/kg) led to the production of eggs with the highest recorded weight, significantly reduced yolk cholesterol and MDA levels, and markedly increased γ-tocopherol concentrations, thereby improving the oxidative stability and nutritional quality of the eggs. The comparable benefits observed in laying hens fed FS (40 g/kg) were further supported by the lowest FCR among all dietary treatments, along with improved egg production and greater egg mass. The combined dietary inclusion of HS and FS (both at 40 g/kg) yielded the most favorable outcomes, including a higher shell percentage and increased yolk deposition of n-3 PUFAs, particularly ALA and EPA, resulting in a more balanced n-6/n-3 ratio and improved nutritional indices of the eggs. The inclusion of HS and FS in laying hen diets may contribute to increasing the deposition of various bioactive compounds with potential health benefits for humans. Further research should investigate the optimal inclusion levels and potential synergistic effects of HS and FS under different alternative housing systems, in which the beneficial concentrations of tocopherols and n-3 PUFAs provided by these supplements may not only increase egg quality and nutritional value but also contribute to improving the health and welfare of laying hens facing new environmental challenges.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The animal study was approved by the Ethical Committee of the Institute of Animal Science (Prague – Uhříněves, Czech Republic) under protocol code 03/2022. All procedures involving animals followed the guidelines of the Ethics Committee of the Central Commission for Animal Welfare at the Ministry of Agriculture of the Czech Republic (Prague, Czech Republic) and complied with Directive 2010/63/EU of the European Parliament and the Council on the protection of animals used for scientific purposes.
Author contributions
JS: Visualization, Formal analysis, Software, Writing – original draft, Resources, Investigation, Conceptualization, Writing – review & editing. ME: Supervision, Conceptualization, Data curation, Methodology, Investigation, Funding acquisition, Project administration, Validation, Writing – review & editing. MS: Conceptualization, Writing – review & editing, Supervision, Project administration, Methodology. EP: Conceptualization, Supervision, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This research was supported by the Ministry of Agriculture of the Czech Republic under grant number MZE-RO0723. The funder had no role in the study design, data collection and analysis, data interpretation, decision to publish, or manuscript preparation. The author(s) also acknowledge the financial support of the Department of Microbiology, Nutrition and Dietetics of the Faculty of Agrobiology, Food and Natural Resources of the Czech University of Life Sciences Prague.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as potential conflicts of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Adeoye A. A., Oyeleye O. O., Olorunsola R. A., Udoh J. E., and Oladepo A. D. (2023). Table egg quality and nutritional composition assessments of different breeds and ages of laying hens. Slovak J. Anim. Sci. 56, 38–45. doi: 10.36547/sjas.792
Aguillón-Páez Y. J., Romero L. A., and Diaz G. J. (2020). Effect of full-fat sunflower or flaxseed seeds dietary inclusion on performance, egg yolk fatty acid profile and egg quality in laying hens. Anim. Nutr. 6, 179–184. doi: 10.1016/j.aninu.2019.12.005, PMID: 32542198
Alagawany M., Elnesr S. S., Farag M. R., Abd El-Hack M. E., Khafaga A. F., Taha A. E., et al. (2019). Omega-3 and omega-6 fatty acids in poultry nutrition: effect on production performance and health. Animals 9, 573. doi: 10.3390/ani9080573, PMID: 31426600
Al-Nasser A. Y., Al-Saff A., Abdulla F., Al-Baho M., Ragheb G., and Mashaly M. (2011). Effect of adding flaxseed in the diet of laying hens on both production of omega-3 enriched eggs and on production performance. Int. J. Poultry Sci. 10, 825–831. doi: 10.3923/ijps.2011.825.831
Antruejo A., Azcona J. O., Garcia P. T., Gallinger C., Rosmini M., Ayerza R., et al. (2011). Omega-3 enriched egg production α-linolenic ω-3 fatty acid sources on laying hen performance and yolk lipid content and fatty acid composition: the effect of α-linolenic ω-3 fatty acid sources on laying hen performance and yolk lipid content and fatty acid composition. Br. Poultry Sci. 52, 750–760. doi: 10.1080/00071668.2011.638621, PMID: 22221241
AOAC (2005). Official methods of analysis (Gaithersburg, MD, USA: AOAC International). Available online at: https://www.researchgate.net/publication/292783651 (Accessed September 10, 2025).
Asem E. K. and Hertelendy F. (1987). Influence of follicular maturation on 3-hydroxy-methylglutaryl coenzyme A reductase activity in hen granulosa cells. Reproduction 79, 215–221. doi: 10.1530/jrf.0.0790215, PMID: 3820173
Attia Y. A., Al-Harthi M. A., Sagan A. A. A., Abdulsalam N. M., Hussein E. O. S., and Olal M. J. (2022). Egg production and quality, lipid metabolites, antioxidant status and immune response of laying hens fed diets with various levels of soaked flax seed meal. Agriculture 12, 1402. doi: 10.3390/agriculture12091402
Ayerza R. and Coates W. (1999). An ω-3 fatty acid enriched chia diet: Influence on egg fatty acid composition, cholesterol and oil content. Can. J. Anim. Sci. 79, 53–58. doi: 10.4141/A98-048
Aymond W. M. and van Elswyk M. E. (1995). Yolk thiobarbituric acid reactive substances and n-3 fatty acids in response to whole and ground flaxseed. Poultry Sci. 74, 1388–1394. doi: 10.3382/ps.0741388, PMID: 7479519
Azcona J. O., Schang M. J., Garcia P. T., Gallinger C., Ayerza R. Jr., and Coates W. (2008). Omega-3 enriched broiler meat: The influence of dietary α-linolenic-ω-3 fatty acid sources on growth, performance and meat fatty acid composition. Can. J. Anim. Sci. 88, 257–269. doi: 10.4141/CJAS07081
Bean L. D. and Leeson S. (2003). Long-term effects of feeding flaxseed on performance and egg fatty acid composition of brown and white hens. Poultry Sci. 82, 388–394. doi: 10.1093/ps/82.3.388, PMID: 12705398
Behrens W. A. and Madère R. (1986). Alpha- and gamma tocopherol concentrations in human serum. J. Am. Coll. Nutr. 5, 91–96. doi: 10.1080/07315724.1986.10720116, PMID: 3700885
Bekhit A. E.-D. A., Shavandi A., Jodjaja T., Birch J., Teh S., Mohamed Ahmed I. A., et al. (2018). Flaxseed: Composition, detoxification, utilization, and opportunities. Biocatalysis Agric. Biotechnol. 13, 129–152. doi: 10.1016/j.bcab.2017.11.017
Bošković Cabrol M., Pravato M., Bordignon F., Xiccato G., Ciarelli C., Bailoni L., et al. (2025). Whole hempseed as a feeding enrichment for laying hens: effects on egg quality, nutritional profile and sensory attributes. Poultry Sci. 104, 105483. doi: 10.1016/j.psj.2025.105483, PMID: 40609326
Cardoso C., Afonso C., and Bandarra N. M. (2016). Seafood lipids and cardiovascular health. Nutrire 41, 1–10. doi: 10.1186/s41110-016-0008-8
Chen J. Y., Latshaw J. D., Lee H. O., and Min D. B. (1998). α-tocopherol content and oxidative stability of egg yolk as related to dietary α-tocopherol. J. Food Sci. 63, 919–922. doi: 10.1111/j.1365-2621.1998.tb17927.x
Chen J. and Liu H. (2020). Nutritional indices for assessing fatty acids: A mini-review. Int. J. Mol. Sci. 21, 5695. doi: 10.3390/ijms21165695, PMID: 32784511
Cherian G. and Quezada N. (2016). Egg quality, fatty acid composition and immunoglobulin Y content in eggs from laying hens fed full fat camelina or flax seed. J. Anim. Sci. Biotechnol. 7, 1–8. doi: 10.1186/s40104-016-0075-y, PMID: 26949531
Cherian G., Wolfe F. W., and Sim J. S. (1996). Dietary oils with added tocopherols: effects on egg or tissue tocopherols, fatty acids, and oxidative stability. Poultry Sci. 75, 423–431. doi: 10.3382/ps.0750423, PMID: 8778738
Coşkuner Y. and Karababa E. (2007). Some physical properties of flaxseed (Linum usitatissimum L.). J. Food Eng. 78, 1067–1073. doi: 10.1016/j.jfoodeng.2005.12.017
Cuchillo-Hilario M., Fournier-Ramírezm M.-I., Díaz Martínez M., Montaño Benavides S., Calvo-Carrillo M.-C., Carrillo Domínguez S., et al. (2024). Animal food products to support human nutrition and to boost human health: the potential of feedstuffs resources and their metabolites as health-promoters. Metabolites 14, 496. doi: 10.3390/metabo14090496, PMID: 39330503
Czauderna M., Kowalczyk J., and Marounek M. (2011). The simple and sensitive measurement of malondialdehyde in selected specimens of biological origin and some feed by reversed phase high performance liquid chromatography. J. Chromatogr. B 879, 2251–2258. doi: 10.1016/j.jchromb.2011.06.008, PMID: 21727037
Dansou D. M., Zhang H., Yu Y., Wang H., Tang C., Zhao Q., et al. (2023). Carotenoid enrichment in eggs: From biochemistry perspective. Anim. Nutr. 14, 315–333. doi: 10.1016/j.aninu.2023.05.012, PMID: 37635928
de Batlle J., Sauleda J., Balcells E., Gómez F. P., Méndez M., Rodriguez E., et al. (2012). Association between Ω3 and Ω6 fatty acid intakes and serum inflammatory markers in COPD. J. Nutr. Biochem. 23, 817–821. doi: 10.1016/j.jnutbio.2011.04.005, PMID: 21889886
EFSA NDA Panel (2015). Scientific opinion on dietary reference values for vitamin E as α-tocopherol. EFSA J. 13, 4149. doi: 10.2903/j.efsa.2015.4149
Ehr I. J., Persia M. E., and Bobeck E. A. (2017). Comparative omega-3 fatty acid enrichment of egg yolks from first-cycle laying hens fed flaxseed oil or ground flaxseed. Poultry Sci. 96, 1791–1799. doi: 10.3382/ps/pew462, PMID: 28108729
Elkin R. G. (2006). Reducing shell egg cholesterol content. I. Overview, genetic approaches, and nutritional strategies. World’s Poultry Sci. J. 62, 665–687. doi: 10.1017/S0043933906001206
EN 12822-1 (2000). Foodstuffs – determination of vitamin E by high performance liquid chromatography – measurement of α-, β-, γ- and δ-tocopherols (Brussels, Belgium: European Committee for Standardization).
EN 12823-1 (2000). Foodstuffs – determination of vitamin A by high performance liquid chromatography – part 1: measurement of all-trans-retinol and 13-cis-retinol (Brussels, Belgium: European Committee for Standardization).
EN 12823-2 (2000). Foodstuffs – determination of vitamin A by high performance liquid chromatography – part 2: measurement of β-carotene (Brussels, Belgium: European Committee for Standardization).
European Union (1999). Council Directive 1999/74/EC of 19 July 1999 laying down minimum standards for the protection of laying hens. Off. J. Eur. Communities L 203, 53–57.
Farinon B., Molinari R., Costantini L., and Merendino N. (2020). The seed of industrial hemp (Cannabis sativa L.): nutritional quality and potential functionality for human health and nutrition. Nutrients 12, 1935. doi: 10.3390/nu12071935, PMID: 32610691
Folch J., Lees M., and Stanley G. H. S. (1957). A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 226, 497–509. doi: 10.1016/S0021-9258(18)64849-5, PMID: 13428781
Fraeye I., Bruneel C., Lemahieu C., Buyse J., Muylaert K., and Foubert I. (2012). Dietary enrichment of eggs with omega-3 fatty acids: A review. Food Res. Int. 48, 961–969. doi: 10.1016/j.foodres.2012.03.014
Gakhar N., Goldberg E., Jing M., Gibson R., and House J. D. (2012). Effect of feeding hemp seed and hemp seed oil on laying hen performance and egg yolk fatty acid content: Evidence of their safety and efficacy for laying hen diets. Poultry Sci. 91, 701–711. doi: 10.3382/ps.2011-01825, PMID: 22334746
Galasso I., Russo R., Mapelli S., Ponzoni E., Brambilla I. M., Battelli G., et al. (2016). Variability in seed traits in a collection of cannabis sativa L. Genotypes. Front. Plant Sci. 7. doi: 10.3389/fpls.2016.00688, PMID: 27242881
Galobart J., Barroeta A. C., Baucells M. D., Cortinas L., and Guardiola F. (2001). α-tocopherol transfer efficiency and lipid oxidation in fresh and spray-dried eggs enriched with ω3-polyunsaturated fatty acids. Poultry Sci. 80, 1496–1505. doi: 10.1093/ps/80.10.1496, PMID: 11599710
Goldberg E. M., Gakhar N., Ryland D., Aliani M., Gibson R. A., and House J. D. (2012). Fatty acid profile and sensory characteristics of table eggs from laying hens fed hempseed and hempseed oil. J. Food Sci. 77, 153–160. doi: 10.1111/j.1750-3841.2012.02626.x, PMID: 22429187
Grilo E. C., Costa P. N., Gurgel C. S. S., Beserra A.F. de L., Almeida F.N. de S., and Dimenstein R. (2014). Alpha-tocopherol and gamma-tocopherol concentration in vegetable oils. Food Sci. Technol. 34, 379–385. doi: 10.1590/S0101-20612014005000031
Guinaz M., Milagres R. C. R. M., Pinheiro-Sant´Ana H. M., and Chaves J. B. P. (2009). Tocopherols and tocotrienols in vegetable oils and eggs. Química Nova 32, 2098–2103. doi: 10.1590/S0100-40422009000800021
Halle I. and Schöne F. (2013). Influence of rapeseed cake, linseed cake and hemp seed cake on laying performance of hens and fatty acid composition of egg yolk. J. für Verbraucherschutz und Lebensmittelsicherheit 8, 185–193. doi: 10.1007/s00003-013-0822-3
Haugh H. (1937). The haugh unit for measuring egg quality. U.S. Egg Poultry Magazine 43, 552–555, 572-573.
Hayes K. C., Pronczuk A., and Liang J. S. (1993). Differences in the plasma transport and tissue concentrations of tocopherols and tocotrienols: observations in humans and hamsters. Exp. Biol. Med. 202, 353–359. doi: 10.3181/00379727-202-43546, PMID: 8437992
Holman R. T. (1954). Autoxidation of fats and related substances. Prog. Chem. Fats Other Lipids 2, 51–98. doi: 10.1016/0079-6832(54)90004-X
Honda K. L., Lamon-Fava S., Matthan N. R., Wu D., and Lichtenstein A. H. (2015). EPA and DHA Exposure Alters the Inflammatory Response but not the Surface Expression of Toll-like Receptor 4 in Macrophages. Lipids 50, 121–129. doi: 10.1007/s11745-014-3971-y, PMID: 25408476
House J. D., Neufeld J., and Leson G. (2010). Evaluating the Quality of Protein from Hemp Seed (Cannabis sativa L.) Products Through the use of the Protein Digestibility-Corrected Amino Acid Score Method. J. Agric. Food Chem. 58, 11801–11807. doi: 10.1021/jf102636b, PMID: 20977230
Hulbert A. J., Pamplona R., Buffenstein R., and Buttemer W. A. (2007). Life and death: metabolic rate, membrane composition, and life span of animals. Physiol. Rev. 87, 1175–1213. doi: 10.1152/physrev.00047.2006, PMID: 17928583
Imran M., Anjum F. M., Nadeem M., Ahmad N., Khan M. K., Mushtaq Z., et al. (2015). Production of Bio-omega-3 eggs through the supplementation of extruded flaxseed meal in hen diet. Lipids Health Dis. 14, 1–9. doi: 10.1186/s12944-015-0127-x, PMID: 26453429
Incharoen T., Nopparatmaitree M., Kongkeaw A., Soisuwan K., Likittrakulwong W., Thongnum A., et al. (2025). Dietary micronized hemp fiber enhances in vitro nutrient digestibility and cecal fermentation, antioxidant enzyme, lysosomal activity, and productivity in finisher broilers reared under thermal stress. Front. Anim. Sci. 6. doi: 10.3389/fanim.2025.1553829
Jia W., Slominski B. A., Guenter W., Humphreys A., and Jones O. (2008). The effect of enzyme supplementation on egg production parameters and omega-3 fatty acid deposition in laying hens fed flaxseed and canola seed. Poultry Sci. 87, 2005–2014. doi: 10.3382/ps.2007-00474, PMID: 18809863
Jiang Q., Christen S., Shigenaga M. K., and Ames B. N. (2001). γ-Tocopherol, the major form of vitamin E in the US diet, deserves more attention. Am. J. Clin. Nutr. 74, 714–722. doi: 10.1093/ajcn/74.6.714, PMID: 11722951
Jing M., Zhao S., and House J. D. (2017). Performance and tissue fatty acid profile of broiler chickens and laying hens fed hemp oil and HempOmegaTM. Poultry Sci. 96, 1809–1819. doi: 10.3382/ps/pew476, PMID: 28160006
Johnston S. A. and Gous R. M. (2007). Modelling the changes in the proportions of the egg components during a laying cycle. Br. Poultry Sci. 48, 347–353. doi: 10.1080/00071660701381134, PMID: 17578698
Katare C., Saxena S., Agrawal S., and Prasad G. B. K. S. (2012). Flax seed: A potential medicinal food. J. Nutr. Food Sci. 2, 1–8. doi: 10.4172/2155-9600.1000120
Konca Y., Yuksel T., Yalcin H., Beyzi S. B., and Kaliber M. (2019). Effects of heat-treated hempseed supplementation on performance, egg quality, sensory evaluation and antioxidant activity of laying hens. Br. Poultry Sci. 60, 39–46. doi: 10.1080/00071668.2018.1547360, PMID: 30421987
Kriese U., Schumann E., Weber W. E., Beyer M., Brühl L., and Matthäus B. (2004). Oil content, tocopherol composition and fatty acid patterns of the seeds of 51 Cannabis sativa L. genotypes. Euphytica 137, 339–351. doi: 10.1023/B:EUPH.0000040473.23941.76
Kumar N. G., Contaifer D., Madurantakam P., Carbone S., Price E. T., Van Tassell B., et al. (2019). Dietary bioactive fatty acids as modulators of immune function: implications on human health. Nutrients 11, 2974. doi: 10.3390/nu11122974, PMID: 31817430
Laganiere S. and Yu B. P. (2004). Modulation of membrane phospholipid fatty acid composition by age and food restriction. Gerontology 39, 7–18. doi: 10.1159/000213509, PMID: 8440492
Lan Y., Zha F., Peckrul A., Hanson B., Johnson B., Rao J., et al. (2019). Genotype x Environmental Effects on Yielding Ability and Seed Chemical Composition of Industrial Hemp (Cannabis sativa L.) Varieties Grown in North Dakota, USA. J. Am. Oil Chemists’ Soc. 96, 1417–1425. doi: 10.1002/aocs.12291
Leeson S., Summers J. D., and Caston L. J. (2000). Response of layers to dietary flaxseed according to body weight classification at maturity. J. Appl. Poultry Res. 9, 297–302. doi: 10.1093/japr/9.3.297
Malini T. and Vanithakumari G. (1990). Rat toxicity studies with β-sitosterol. J. Ethnopharmacology 28, 221–234. doi: 10.1016/0378-8741(90)90032-O, PMID: 2329812
Mandal G. P., Ghosh T. K., and Patra A. K. (2014). Effect of Different Dietary n-6 to n-3 Fatty Acid Ratios on the Performance and Fatty Acid Composition in Muscles of Broiler Chickens. Asian-Australasian J. Anim. Sci. 27, 1608–1614. doi: 10.5713/ajas.2014.14013, PMID: 25358321
Mattioli S., Ruggeri S., Sebastiani B., Brecchia G., Dal Bosco A., Cartoni Mancinelli A., et al. (2017). Performance and egg quality of laying hens fed flaxseed: highlights on n-3 fatty acids, cholesterol, lignans and isoflavones. Animal 11, 705–712. doi: 10.1017/S175173111600207X, PMID: 27819218
Mattson F. H., Grundy S. M., and Crouse J. R. (1982). Optimizing the effect of plant sterols on cholesterol absorption in man. Am. J. Clin. Nutr. 35, 697–700. doi: 10.1093/ajcn/35.4.697, PMID: 7072622
Meffo Kemda M., Marchi M., Neri E., Marchettini N., and Niccolucci V. (2024). Environmental impact assessment of hemp cultivation and its seed-based food products. Front. Environ. Sci. 12. doi: 10.3389/fenvs.2024.1342330
Mierliţă D. (2018). Effects of diets containing hemp seeds or hemp cake on fatty acid composition and oxidative stability of sheep milk. South Afr. J. Anim. Sci. 48, 504–515. doi: 10.4314/sajas.v48i3.11
Mierliţă D. (2019). Fatty acids profile and oxidative stability of eggs from laying hens fed diets containing hemp seed or hempseed cake. South Afr. J. Anim. Sci. 49, 310–321. doi: 10.4314/sajas.v49i2.11
Mohamed N., Slaski J. J., Shwaluk C., and House J. D. (2024). Chemical characterization of hemp (Cannabis sativa L.)-derived products and potential for animal feed. ACS Food Sci. Technol. 4, 88–103. doi: 10.1021/acsfoodscitech.3c00391
Mortensen A., Skibsted L. H., and Truscott T. G. (2001). The interaction of dietary carotenoids with radical species. Arch. Biochem. Biophysics 385, 13–19. doi: 10.1006/abbi.2000.2172, PMID: 11361009
Mridula D., Kaur D., Nagra S. S., Barnwal P., Gurumayum S., and Singh K. K. (2014). Growth performance and quality characteristics of flaxseed-fed broiler chicks. J. Appl. Anim. Res. 43, 345–351. doi: 10.1080/09712119.2014.978773
Neijat M., Gakhar N., Neufeld J., and House J. D. (2014). Performance, egg quality, and blood plasma chemistry of laying hens fed hempseed and hempseed oil. Poultry Sci. 93, 2827–2840. doi: 10.3382/ps.2014-03936, PMID: 25239534
Neijat M., Suh M., Neufeld J., and House J. D. (2016). Hempseed products fed to hens effectively increased n-3 polyunsaturated fatty acids in total lipids, triacylglycerol and phospholipid of egg yolk. Lipids 51, 601–614. doi: 10.1007/s11745-015-4088-7, PMID: 26515300
Neori A. and Agami M. (2024). Low-income fish consumers’ Subsidies to the fish reduction industry: the case of forage fish. World 5, 769–788. doi: 10.3382/ps.2009-00522, PMID: 20952712
Oliveira D. D., Baião N. C., Cançado S. V., Grimaldi R., Souza M. R., Lara L. J.C., et al. (2010). Effects of lipid sources in the diet of laying hens on the fatty acid profiles of egg yolks. Poultry Science 89 (11), 2484–2490. doi: 10.3382/ps.2009-00522, PMID: 20952712
Omri B., Chalghoumi R., Izzo L., Ritieni A., Lucarini M., Durazzo A., et al. (2019). Effect of dietary incorporation of linseed alone or together with tomato-red pepper mix on laying hens’ Egg yolk fatty acids profile and health lipid indexes. Nutrients 11, 813. doi: 10.3390/nu11040813, PMID: 30974860
Ostapczuk K., Apori S. O., Estrada G., and Tian F. (2021). Hemp growth factors and extraction methods effect on antimicrobial activity of hemp seed oil: A systematic review. Separations 8, 183. doi: 10.3390/separations8100183
Palmquist D. L. (2009). Omega-3 fatty acids in metabolism, health, and nutrition and for modified animal product foods. Prof. Anim. Scientist 25, 207–249. doi: 10.15232/S1080-7446(15)30713-0
Panaite T. D., Nour V., Saracila M., Turcu R. P., Untea A. E., and Vlaicu P. A. (2021). Effects of linseed meal and carotenoids from different sources on egg characteristics, yolk fatty acid and carotenoid profile and lipid peroxidation. Foods 10, 1246. doi: 10.3390/foods10061246, PMID: 34072662
Parker T. D., Adams D. A., Zhou K., Harris M., and Yu L. (2003). Fatty acid composition and oxidative stability of cold-pressed edible seed oils. J. Food Sci. 68, 1240–1243. doi: 10.1111/j.1365-2621.2003.tb09632.x
Paulová T., Malíková L., Lanzoni D., Taubner T., Malík M., Houdková M., et al. (2025). Inhibitory potential of cannabis biomass extracts on livestock-associated staphylococcal and streptococcal pathogens. Microorganisms 13, 432. doi: 10.3390/microorganisms13020432, PMID: 40005797
Porto C. D., Decorti D., and Natolino A. (2014). Potential oil yield, fatty acid composition, and oxidation stability of the hempseed oil from four cannabis sativa L. Cultivars. J. Dietary Suppl. 12, 1–10. doi: 10.3109/19390211.2014.887601, PMID: 24552275
Puertas G. and Vázquez M. (2018). Advances in techniques for reducing cholesterol in egg yolk: A review. Crit. Rev. Food Sci. Nutr. 59, 2276–2286. doi: 10.1080/10408398.2018.1448357, PMID: 29513557
Raes K., De Smet S., Balcaen A., Claeys E., and Demeyer D. (2003). Effect of diets rich in n-3 polyunsatured fatty acids on muscle lipids and fatty acids in Belgian Blue double-muscled young bulls. Reproduction nutrition Dev. 43, 331–345. doi: 10.1051/rnd:2003029, PMID: 14971825
Rbah Y., Taaifi Y., Allay A., Belhaj K., Melhaoui R., Houmy N., et al. (2024). A comprehensive exploration of the fatty acids profile, cholesterol, and tocopherols levels in liver from laying hens fed diets containing nonindustrial hemp seed. Scientifica 2024, 1–11. doi: 10.1155/2024/8848436, PMID: 38222849
Rbah Y., Taaifi Y., Allay A., Mansouri F., Belhaj K., Houmy N., et al. (2025). Optimization of hemp seed supplementation with phytobiotics in laying hen feed to improve egg yolk fatty acids, tocopherols and cholesterol using response surface models. Front. Sustain. Food Syst. 9. doi: 10.3389/fsufs.2025.1566447
Réhault-Godbert S., Guyot N., and Nys Y. (2019). The golden egg: nutritional value, bioactivities, and emerging benefits for human health. Nutrients 11, 684. doi: 10.3390/nu11030684, PMID: 30909449
Ren Y., Perez T. I., Zuidhof M. J., Renema R. A., and Wu J. (2013). Oxidative stability of omega-3 polyunsaturated fatty acids enriched eggs. J. Agric. Food Chem. 61, 11595–11602. doi: 10.1021/jf403039m, PMID: 24164329
Rodríguez M. L., Alzueta C., Rebolé A., Ortiz L. T., Centeno C., and Treviño J. (2001). Effect of inclusion level of linseed on the nutrient utilisation of diets for growing broiler chickens. Br. Poultry Sci. 42, 368–375. doi: 10.1080/00071660120055359, PMID: 11469558
Russo R. and Reggiani R. (2016). Evaluation of protein and antinutritional compounds content in meal from seven flax varieties. J. Global Agric. Ecol. 6, 182–188.
Saengsuwan H., Bunchasak C., Rakangthong C., and Poungpong K. (2025). Hemp inflorescence meal as a novel feed ingredient in laying hens: Safety assessment, nutritional characterization, and effects on egg quality. Veterinary World 18, 2406–2413. doi: 10.14202/vetworld.2025.2406-2413, PMID: 41064825
Santos-Silva J., Bessa R. J. B., and Santos-Silva F. (2002). Effect of genotype, feeding system and slaughter weight on the quality of light lambs. Livestock Production Sci. 77, 187–194. doi: 10.1016/S0301-6226(02)00059-3
Shahid S., Chand N., Khan R. U., Suhail S. M., and Khan N. A. (2015). Alternations in Cholesterol and Fatty Acids Composition in Egg Yolk of Rhode Island Red x Fyoumi Hens Fed with Hemp Seeds (Cannabis sativa L.). J. Chem. 2015, 1–6. doi: 10.1155/2015/362936
Shariatmadari F. (2023). Emergence of hemp as feed for poultry. World’s Poultry Sci. J. 79, 769–782. doi: 10.1080/00439339.2023.2234871
Shim Y. Y., Gui B., Wang Y., and Reaney M. J. T. (2015). Flaxseed (Linum usitatissimum L.) oil processing and selected products. Trends Food Sci. Technol. 43, 162–177. doi: 10.1016/j.tifs.2015.03.001
Siano F., Moccia S., Picariello G., Russo G. L., Sorrentino G., Di Stasio M., et al. (2019). Comparative study of chemical, biochemical characteristic and ATR-FTIR analysis of seeds, oil and flour of the edible fedora cultivar hemp (Cannabis sativa L.). Molecules 24, 83. doi: 10.3390/molecules24010083, PMID: 30591638
Silversides F. G. and LefranÇois M. R. (2005). The effect of feeding hemp seed meal to laying hens. Br. Poultry Sci. 46, 231–235. doi: 10.1080/0071660500066183, PMID: 15957445
Simopoulos A. P. (2008). The importance of the omega-6/omega-3 fatty acid ratio in cardiovascular disease and other chronic diseases. Exp. Biol. Med. 233, 674–688. doi: 10.3181/0711-MR-311, PMID: 18408140
Singh U., Devaraj S., Jialal I., and Vitamin E. (2005). Oxidative stress, and inflammation. Annu. Rev. Nutr. 25, 151–174. doi: 10.1146/annurev.nutr.24.012003.132446, PMID: 16011463
Skřivan M., Englmaierová M., Taubner T., and Skřivanová E. (2020). Effects of dietary hemp seed and flaxseed on growth performance, meat fatty acid compositions, liver tocopherol concentration and bone strength of cockerels. Animals 10, 458. doi: 10.3390/ani10030458, PMID: 32164238
Skřivan M., Englmaierová M., Vít T., and Skřivanová E. (2019). Hempseed increases gamma-tocopherol in egg yolks and the breaking strength of tibias in laying hens. PloS One 14, 1–9. doi: 10.1371/journal.pone.0217509, PMID: 31136626
Szmek J., Englmaierová M., Skřivan M., and Pěchoučková E. (2025). Skeletal disorders in laying hens: a systematic review with a focus on non-cage housing systems and hemp-based dietary interventions for bone health. Br. Poultry Sci., 1–30. doi: 10.1080/00071668.2025.2489059, PMID: 40331968
Taaifi Y., Belhaj K., Mansouri F., Rbah Y., Melhaoui R., Houmy N., et al. (2023). The effect of feeding laying hens with nonindustrial hemp seed on the fatty acid profile, cholesterol level, and tocopherol composition of egg yolk. Int. J. Food Sci. 2023, 1–15. doi: 10.1155/2023/1360276, PMID: 37273530
Ulbricht T. L. V. and Southgate D. A. T. (1991). Coronary heart disease: seven dietary factors. Lancet 338, 985–992. doi: 10.1016/0140-6736(91)91846-M, PMID: 1681350
United Nations (2015). Transforming our world: the 2030 Agenda for Sustainable Development. Resolution adapted by the General Assembly on 25 September 2015 (New York: United Nations). Available online at: https://sdgs.un.org/goals (Accessed September 14, 2025).
Usturoi M. G., Rațu R. N., Crivei I. C., Veleșcu I. D., Usturoi A., Stoica F., et al. (2025). Unlocking the power of eggs: nutritional insights, bioactive compounds, and the advantages of omega-3 and omega-6 enriched varieties. Agriculture 15, 242. doi: 10.3390/agriculture15030242
van den Elsen L., Garssen J., and Willemsen L. (2012). Long chain n-3 polyunsaturated fatty acids in the prevention of allergic and cardiovascular disease. Curr. Pharm. Design 18, 2375–2392. doi: 10.2174/138161212800165960, PMID: 22390701
Vonapartis E., Aubin M.-P., Seguin P., Mustafa A. F., and Charron J.-B. (2015). Seed composition of ten industrial hemp cultivars approved for production in Canada. J. Food Composition Anal. 39, 8–12. doi: 10.1016/j.jfca.2014.11.004
Wagner K.-H., Kamal-Eldin A., and Elmadfa I. (2004). Gamma-tocopherol – an underestimated vitamin? Ann. Nutr. Metab. 48, 169–188. doi: 10.1159/000079555, PMID: 15256801
Wang Z. G., Pan X. J., Zhang W. Q., Peng Z. Q., Zhao R. Q., and Zhou G. H. (2010). Methionine and selenium yeast supplementation of the maternal diets affects antioxidant activity of breeding eggs. Poultry Sci. 89, 931–937. doi: 10.3382/ps.2009-00268, PMID: 20371845
Watkins B. A., Li Y., Lippman H. E., and Feng S. (2003). Modulatory effect of omega-3 polyunsaturated fatty acids on osteoblast function and bone metabolism. Prostaglandins Leukotrienes Essential Fatty Acids 68, 387–398. doi: 10.1016/S0952-3278(03)00063-2, PMID: 12798659
Whittle R. H., Kiarie E. G., and Widowski T. M. (2024). The effect of feeding flaxseed as a source of omega-3 fatty acids to broiler and layer breeders during rearing and lay on body weight, reproductive performance, and hatchability performance indices. Can. J. Anim. Sci. 104, 313–323. doi: 10.1139/cjas-2023-0118
Yao W., Li J., Wang J. J., Zhou W., Wang Q., Zhu R., et al. (2012). Effects of dietary ratio of n-6 to n-3 polyunsaturated fatty acids on immunoglobulins, cytokines, fatty acid composition, and performance of lactating sows and suckling piglets. J. Anim. Sci. Biotechnol. 3, 1–8. doi: 10.1186/2049-1891-3-43, PMID: 23270637
Keywords: oilseeds, bioactive compounds, tocopherols, α-linolenic acid, eicosapentaenoic acid, feed conversion ratio, cholesterol, nutritional indices
Citation: Szmek J, Englmaierová M, Skřivan M and Pěchoučková E (2025) Evaluation of hemp seeds (Cannabis sativa L.) and flax seeds (Linum usitatissimum L.) as feed ingredients in laying hen diets: effects on the performance, egg quality, and n-3 fatty acid composition of egg yolks. Front. Anim. Sci. 6:1685765. doi: 10.3389/fanim.2025.1685765
Received: 14 August 2025; Accepted: 22 September 2025;
Published: 15 October 2025.
Edited by:
Femi Fawole, University of Ilorin, NigeriaReviewed by:
Youssef Rbah, Mohamed Premier University, MoroccoKazeem Adeyemi, University of Ilorin, Nigeria
Copyright © 2025 Szmek, Englmaierová, Skřivan and Pěchoučková. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jan Szmek, c3ptZWtAYWYuY3p1LmN6
†ORCID: Jan Szmek, orcid.org/0009-0003-3487-921X
Michaela Englmaierová, orcid.org/0000-0003-2752-3170
Miloš Skřivan, orcid.org/0000-0001-9023-8124
Eva Pěchoučková, orcid.org/0000-0001-9336-5436