 Daniel Serrano-Jara1*
Daniel Serrano-Jara1* Iván Agea1
Iván Agea1 Gema Romero1
Gema Romero1 María José Argente1
María José Argente1 María Antonia Santacreu2María de la Luz García1
María Antonia Santacreu2María de la Luz García1- 1Instituto de Investigación e Innovación Agroalimentaria y Agroambiental, CIAGRO-UMH, Orihuela, Spain
- 2Instituto de Ciencia y Tecnología Animal, Universidad Politécnica de Valencia, Valencia, Spain
Heat stress is a major challenge for rabbit production, due to the species’ limited thermoregulatory capacity. Two rabbit lines were divergently selected over 17 generations for environmental variance in litter size: the Low line, with greater resilience, and the High line, with less resilience. This study aimed to compare acute stress and inflammatory responses in males from both lines under heat stress (temperature–humidity index, THI ≥ 27.8) and thermoneutral (THI< 27.8) conditions. Forty males (20 per line) were evaluated for eyeball temperature via infra-red thermography before and after a semen collection stressor, and for plasma cortisol, C-reactive protein (CRP), and tumor necrosis factor-α (TNF-α) by ELISA. Bayesian methodology was used for statistical analysis. The Low line maintained lower eyeball temperatures than the High line under both thermal conditions (P > 90%), while retaining normal acute stress responses. Under heat stress, the Low line also showed lower TNF-α levels, and under both conditions, lower cortisol levels (P > 90%). CRP did not differ between lines or thermal conditions (P< 90%). These results indicate that selection for reduced litter size variability improves thermal regulation and attenuates stress-associated physiological responses, supporting its use to enhance resilience, welfare, and sustainability in rabbit production.
1 Introduction
The global rise in temperatures due to climate change is a challenge for livestock production. The Mediterranean region constitutes a hottest and large area of highly specialized and technically skilled rabbit production (Giorgi and Lionello, 2008; Cullere and Dalle-Zotte, 2018). Heat stress is one of the main factors directly and indirectly linked to environmental stress in rabbits (Oladimeji et al., 2022; Ebeid et al., 2023).
Rabbits are homeothermic animals that are very sensitive to high temperatures (Marai et al., 2002). Due to their thick fur and the absence of sweat glands, this species has a low thermoregulatory capacity (Marai et al., 2001; Yağci et al., 2006). The zone of thermoneutrality in rabbits is between 15 to 25 °C and 55-65% of humidity, the critical temperature from 27 to 30 °C, and 35 °C is the maximum temperature above which rabbits cannot regulate their body temperature (Marai et al., 2001; Cervera and Carmona, 2010; Nielsen et al., 2022; Oladimeji et al., 2022). Heat stress occurs when rabbits are unable to maintain equilibrium between the heat they produce and the heat they lose.
Heat stress negatively impacts average daily gain and feed conversion ratio (Jaén-Téllez et al., 2021; Liang et al., 2022), while also impairing reproductive performance. This includes reduced sperm quality in males (Pei et al., 2012; Huang et al., 2023) and decreased fertility, embryo implantation rates, embryo quality and birth weights in females (García and Argente, 2017; Liang et al., 2022; Ebeid et al., 2023). It also reduces the quality of the processed product (Zeferino et al., 2013) and increases the mortality rate (Yan and Li, 2008).
High ambient temperatures activate the hypothalamic-pituitary-adrenocortical (HPA) axis and the sympathetic adrenomedullary system, leading to increased plasma levels of cortisol and catecholamines, and body temperature (Hennessy, 1997; Möstl and Palme 2002). Infrared thermography (IRT) enables the detection of variations in body temperature (Agea et al., 2021; Jaén-Téllez et al., 2021), which serves as an indicator of heat stress and is less invasive than measuring plasma cortisol levels. In particular, the rabbit’s eyeball provides an optimal anatomical site for rapidly assessing the response to acute stress stimuli (Serrano-Jara et al., 2025).
Acute stress dysregulates the immune system, increasing susceptibility to diseases (Mormede et al., 2018). This homeostatic disruption, caused by factors like infections, tissue damage, or stress itself, triggers an acute phase response (Kushner, 1988). In turn, the acute phase response induces the release of cytokines, which alter the plasma concentration of acute phase proteins (Eckersall and Bell, 2010). Among these, TNF-α, a pro-inflammatory cytokine, and C-reactive protein (CRP), a positive acute phase protein, are widely used as biomarkers for assessing stress and animal welfare (Gutiérrez et al., 2009; Argente et al., 2019; Beloumi et al., 2020).
TNF-α is a central mediator in multiple physiological and disease-related processes. In humans, it contributes to inflammation by activating neutrophils and disrupting intestinal absorption (Deem et al., 1991; Suter et al., 1992). Studies in rodents have linked TNF-α to oxidative stress-induced cell damage in mice and regulation of the HPA axis in rats (Bohlinger et al., 1996; Takao et al., 1997). In rabbits, evidence suggests it impairs jejunal absorption of nutrients such as L-leucine and D-fructose (Abad et al., 2002; García-Herrera et al., 2004). CRP is synthesized in the liver and its serum concentrations increase in response to an inflammatory process. In rabbits, plasma CRP levels have been associated with reproductive, dental, and musculoskeletal diseases (Oohashi et al., 2019). In goats, it has been linked to metritis and mastitis, and in sows to infectious processes and stress situations (Saco and Bassols, 2022).
Resilience is the capacity of the animal to be minimally affected by a disturbance or to rapidly return to the physiological, behavioral, cognitive, health, affective and production states that pertained before exposure to a disturbance (Colditz and Hine, 2016). A divergent selection program was developed in rabbits with the aim of improving resilience. However, resilience is a trait that is difficult to measure directly. For this reason, the team proposed assessing it through the environmental variance in litter size within females. The Low line, which developed greater resilience as a result of the selection process, was selected for reduced litter size variability, whereas the High line, with comparatively less resilience, was selected for increased variability (Blasco et al., 2017).
The genetic program proved successful: after 12 generations of selection, litter size variability was 2.5 kits² in the Low line and 5.5 kits² in the High line. Females from the Low line showed a lower inflammatory response to infectious challenges and greater disease resistance compared to those from the High line. This was reflected in lower female mortality at parturition, reduced litter mortality at birth and weaning, and greater uniformity in litter weight at weaning (Argente et al., 2017). Moreover, females from the High line exhibited higher stress responses, lower disease resistance, and smaller litter sizes than those from the Low line (Argente et al., 2017, 2019; Beloumi et al., 2020), confirming that the Low line is the more resilient than the High line.
The study aims to assess the acute stress and inflammatory correlated response in two divergent selected rabbit lines for litter size variability, under both with and without environmental heat stress, by monitoring body temperature and plasma levels of CRP, TNF-α, and cortisol.
2 Material and methods
The experimental procedures with animals were approved by the General Directorate of Agriculture, Livestock and Fisheries of the Generalitat Valenciana with code 2022/VSC/PEA/0226.
2.1 Animals and experimental design
The research was carried out on the farm at Miguel Hernandez University in Orihuela (38.06747347849708, -0.9821394614708082), Alicante (Spain). The facilities included equipped with forced ventilation and cooling, and a photoperiod of 16 hours of light and 8 hours of darkness. Animals were housed in individual cages, fed with a commercial diet (16.1% of crude protein; 3.522 kcal/kg of digestible energy) and provided water ad libitum.
In the selection process for environmental variability of litter size, each line includes approximately 125 ± 5 females and 25 males per generation. Selection is based on the phenotypic variance of litter size within each female, after correcting litter size for year-season effects and parity-lactation status. Once systematic effects are corrected, the intra-doe phenotypic variance reflects environmental variability. The selection intensity for each generation is 30%. To reduce inbreeding, each male is mated with five females, and one male offspring from the best female is selected to breed the next generation (see more details in Blasco et al., 2017). Each generation lasts 12 months.
A total of 40 males, 20 each from the 17th generation of the High (heterogeneous) and Low (homogeneous) lines, were used in this study. The age range was between 5 and 10 months on both lines. The average weight was 3.53 kg.
From ambient temperature and relative humidity, the temperature/humidity index (THI) was calculated: THI = t - [(0.31 - 0.31 × rh) × (t - 14.4)] where t = average temperature of the farm from the time of infra-red imaging until 24 hours before and rh = relative humidity/100, collected in the same way as temperature (Marai et al., 2001). The weeks in which data were collected were classified according to the THI into heat stressed (THI ≥ 27.8) and non-heat stressed (THI< 27.8) (Marai et al., 2001). According to the THI, the groups were classified into High Stressed Line (HS), High Non-Stressed Line (HNS), Low Stressed Line (LS) and Low Non-Stressed Line (LNS).
2.2 Temperature records
A total of 120 body temperature records were taken from the High and Low lines (60 per line; 30 per group). Body temperature emissivity was measured using IRT on the eyeball. The images were obtained using a ®FLIR SC660 thermal imaging camera and were processed with the ®ThermaCAM Researcher Pro 2.10 software to obtain the temperature record. The camera was calibrated according to temperature, relative humidity, emissivity (98%), and distance from the subject (0.7 m).
Data collection was carried out for 12 weeks, between June 2023 and February 2024. Body temperature measurements at the eyeball were taken at three moments: minute 0 (corresponding to basal temperature), minute 1 (one minute after the application of the acute stressor), and minute 5 (five minutes after the application of the acute stressor). Between 0 and 1 minute the rabbit was subjected to an acute stress stimulus based on semen collection (Serrano-Jara et al., 2025).
For semen collection, males started the training period at 150 days of age. The training sessions were conducted once per week over a period of 2–3 weeks (Lavara et al., 2011). After training, the males entered the production period.
2.3 Blood parameters
Eighty blood samples were collected from the central auricular artery in two conditions: under heat stress (THI ≥ 27.8) and non-heat stress (THI< 27.8). The samples were collected in 13 x 75 mm tubes with a 3 mL capacity containing potassium salt K3EDTA (ethylenediaminetetraacetic acid) as anticoagulant. Blood samples were collected using a needle with a diameter of 0.6 mm (23G). Blood was centrifuged at 4000 rpm for 15 minutes to obtain plasma and proceed with the determination of CRP, TNF-α, and cortisol. The samples were frozen at -80°C until analysis.
TNF-α. TNF-α was measured by commercial ELISA test (Elabscience Bionovation Inc, Catalog Number: E-EL-RB0011, Houston, TX, USA). The distribution of the samples was as follows: 20 (LS), 20 (HS), 20 (LNS) and (20 HNS).
CRP. CRP was measured by commercial ELISA test (Life Diagnostics Inc, Catalog Number: CRP-10-N, West Chester, PA, USA). A total of 61 samples were recorded: 17 (LS), 16 (HS), 16 (LNS) and 12 (HNS). The rest of the samples were not recorded in the ELISA test.
Cortisol. Cortisol was measured by commercial ELISA test (Cusabio Technology LLC, Catalog Number: CSB-E06956Rb, Houston, TX, USA). A total of 38 samples were recorded: 18 (LS), 12 (HS), 5 (LNS) and 5 (HNS). The rest of the samples were not recorded in the ELISA test.
2.4 Statistical analysis
The correlated response was estimated as the difference between the High and Low lines. Temperature (yijk) was analyzed with the model:
where μ is the overall mean, TLSi is the time-line-stress effect with twelve levels (time: minute 0, 1 or 5; line: High or Low; stress: THI < 27.8 or THI ≥ 27.8), b is the regression coefficient, Wij is the covariate weight, mij is the random effect of male and eijk is the residual term.
For blood parameters (CRP, TNF-α, and cortisol; yijk) the following model was used:
where μ is the overall mean, LSi is the line-stress effect with four levels (High or Low; stress: THI< 27.8 or THI ≥ 27.8), b is the regression coefficient, Wij is the covariate weight, mij is the random effect of male and eijk is the residual term.
All analyses were performed using Bayesian methodology. Bounded uniform priors were used for all effects except for the male effect, considered normally distributed with mean 0 and variance Iσ2p. Residuals were a priori normally distributed with mean 0 and variance Iσ2e. The priors for the variance were also bounded uniform. Features of the marginal posterior distributions for all unknowns were estimated using Gibbs sampling. Inferences were derived from the marginal posterior distributions. Means, standard errors and actual probability (P) were provided. P refers to the probability that the absolute value of the difference between two levels of a fixed effect will be greater than zero (Blasco et al., 2017). P indicates the probability that the differences are above zero when H–L > 0, or below zero when H–L< 0. Within a Bayesian framework, the notion of “statistical significance” in the classical sense does not apply; instead, the model directly estimates the probability of the differences being positive or negative. Notably, these probabilities may reach or even surpass 0.90, even in cases where the 95% credible interval still encompasses zero. We consider that there are relevant differences when the P is greater than or equal to 90%. The Rabbit software program developed by the Institute for Animal Science and Technology (Valencia, Spain) was used for all procedures. We used a chain of 60,000 samples, with a burn-in period of 10,000. Only one out of every 10 samples was saved for inferences. Convergence was tested using the Z criterion of Geweke (Sorensen and Gianola, 2002) and Monte Carlo sampling errors were computed using time-series procedures (Geyer, 1992).
3 Results
3.1 Temperature
Figure 1 shows the progression (at minutes 0, 1, and 5) of eyeball temperature both conditions (heat stress and non-heat stress). For all the moments, the Low line showed lower temperatures than the High line, both in the absence and presence environmental heat stress (P = 100%; P: probability that the difference between the temperatures of both lines at each moment is different from 0).
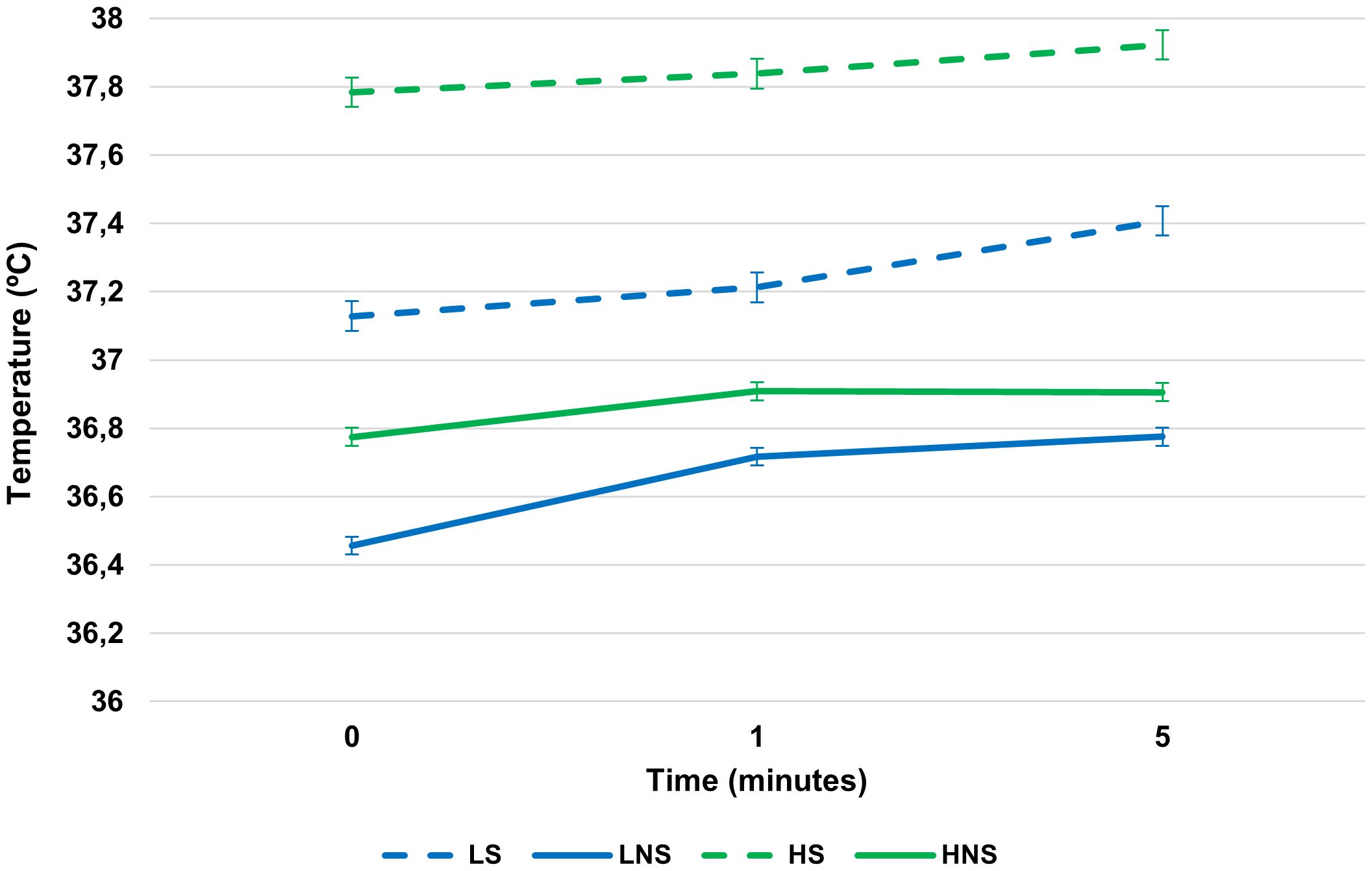
Figure 1. Means and standard errors (bars) of the marginal posterior distributions for eyeball temperature, recorded with infra-red thermography, in two rabbit lines with and without heat stress. HS, High stressed line; LS, Low stressed line; HNS, High non-stressed line; LHS, Low non-stressed line. The vertical bars represent the standard error.
In the absence of environmental heat stress, at minute 0, the Low line showed a lower eyeball temperature than the High line (36.45 °C vs 36.77°C; P = 92%). At minute 1, temperature tended to be lower in the Low line than in the High line (90% > P > 80%). No differences were found between the lines at minute 5 (P = 71%).
After seminal collection, between minute 0 and 1, both lines increased in temperature (Low line: 36.71°C and P = 99%; High line: 36.91°C and P = 90%). Between minute 1 and 5, both lines increased their temperatures (Low line: 36.77°C and P = 71%; High line: 36.91°C and P = 51%).
In the presence of environmental heat stress the Low line presented a lower temperature than the High line at minute 0 (37.12°C vs 37.78°C; P = 100%). After seminal collection, between minute 0 and 1, none of the lines increased their temperature during the first minute (Low line: 37.21°C and P = 72%; High line: 37.84°C and P = 65%). The difference between the lines was maintained at minute 1 (P = 99%). Between minute 1 and 5, only the Low line increased its temperature (37.41°C; P = 92%) in comparison to the High line (37.92°C; P = 72.2%). The difference between the temperature of the two lines remained present (P = 97%).
3.2 Blood parameters (CRP, TNF-α, cortisol)
Table 1 shows the differences of correlated response between the lines in CRP, TNF-α, and cortisol levels under conditions with and without heat stress. CRP levels showed no differences between the lines (P< 80%). The Low line showed lower TNF-α levels in the presence of environmental heat stress (P > 90%), but only a trend (90% > P > 80%) in the absence of environmental heat stress. Regarding plasma cortisol levels, the Low line showed lower levels than the High line (P > 90%) both in the presence and absence of environmental heat stress.
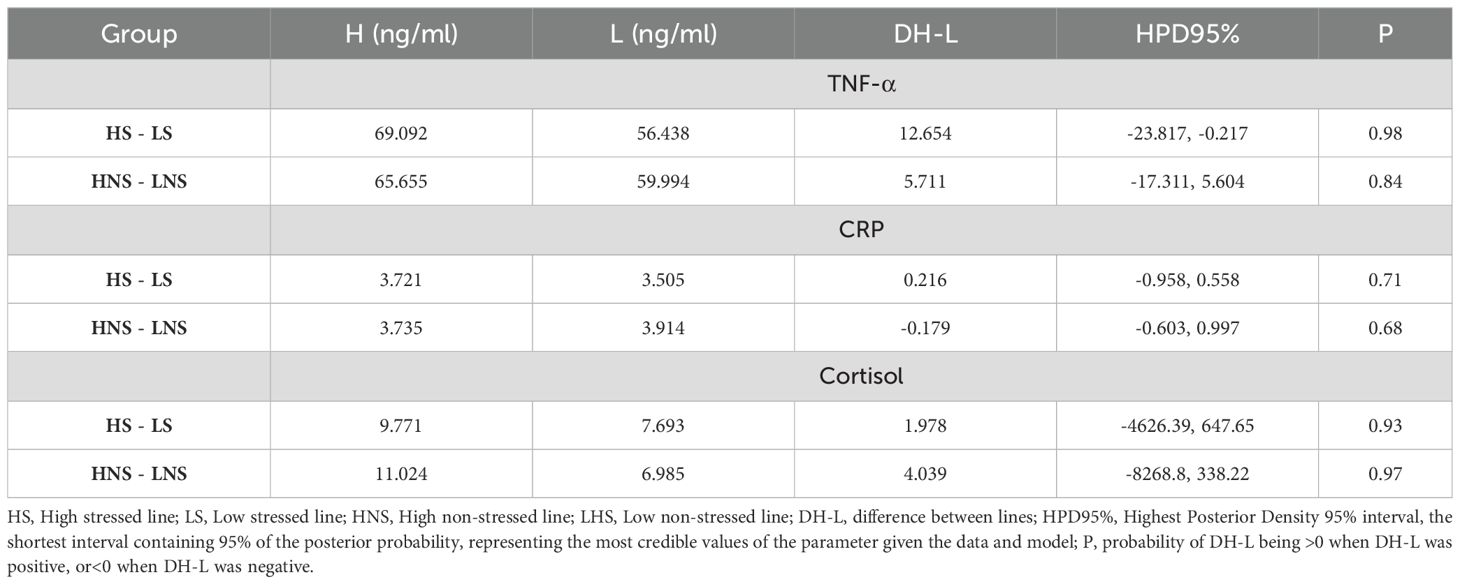
Table 1. Correlated response to selection for resilience in plasma TNF-α, CRP, and cortisol levels in two rabbit lines.
Table 2 shows a comparison of the results obtained under heat stress and thermal comfort conditions within the same lines. TNF-α showed an upward trend in the presence of environmental heat stress in the High line (90% > P > 80%). As for CRP, only in the absence of environmental heat stress in the Low line was there a tendency for its concentration to increase (90% > P > 80%). Cortisol levels did not vary between the presence and absence of environmental heat stress in any of the lines (P< 80%).
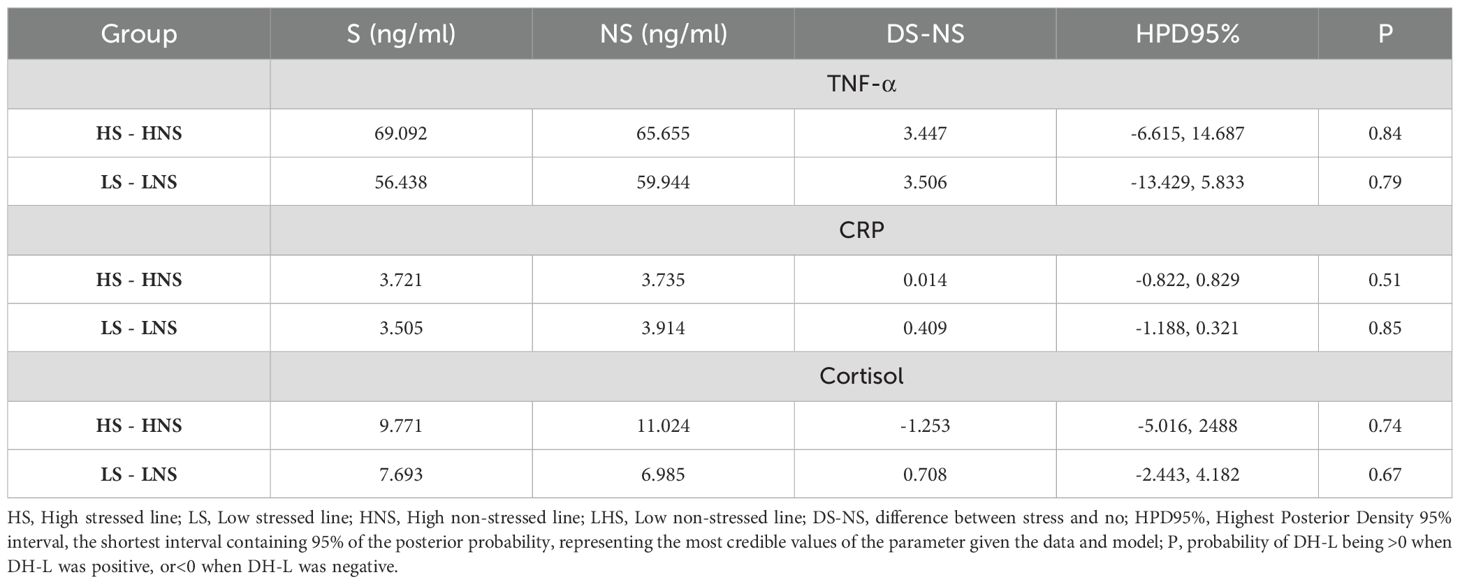
Table 2. Comparison of plasma TNF-α, CRP, and cortisol levels between stressed and non-stressed groups in two rabbit lines.
4 Discussion
Selecting animals primarily for productivity has been associated with negative side effects in their health, fertility, and immune responses (Rauw et al., 1998). Nowadays, the fight against climate change and the improvement of animal welfare are related challenges that affect livestock farming both socially and scientifically.
Over the course of 17 generations, two rabbit lines have been divergently selected based on the environmental variability of litter size (i.e. phenotypic variance of litter size within each female, after correcting litter size for year-season effects and parity-lactation status). This selection strategy has been proposed as a direct approach to improving resilience traits (Blasco et al., 2017). Several studies have demonstrated that females from the Low line exhibit superior adaptive capacity under adverse environmental conditions (García et al., 2016; García et al., 2019; Argente et al., 2019).
Our results indicate that, the Low line maintains a lower eyeball temperature compared to the High line, while retaining a normal physiological response to the sperm collection stimulus. Furthermore, the findings show that, in non-heat-stress condition the response to the acute stressor is rapid in both lines, consistent with observations reported in the cited literature (Jaén-Téllez et al., 2021; Serrano-Jara et al., 2025).
Under heat stress, as also reported by De Lima et al. (2013) and Jaén-Téllez et al. (2020), the body temperature of both lines increased. The lower temperature observed in the Low line reflects its superior ability to adapt to adverse environments compared to the High line. In contrast, the lack of a thermal response to semen collection in the High line may be due to the presence of a chronic stress state, which could diminish, delay, or impair its adaptive capacity (Moberg, 2000; McEwen and Wingfield, 2003; Koolhaas et al., 2011).
The improved regulation of body temperature observed in the Low line, selected for resilience, may reflect an enhanced capacity to maintain homeostasis under challenging conditions. This thermoregulatory advantage could be mediated by more efficient neuroendocrine responses to stress, in line with observations in other robustness-selected lines (Biada et al., 2024).
The analysis of stress-associated biomarkers complements the study of temperature by providing insight into the animals’ chronic stress status. The consistently lower cortisol levels in the Low line, regardless of environmental conditions, suggest a more adaptive HPA axis response, which may prevent the detrimental effects of prolonged glucocorticoid release. In contrast, the High line appears to exhibit higher baseline activation of the HPA axis, consistent with Argente et al. (2019), which could indicate a reduced threshold for stress activation and a chronic stress profile.
Differences in TNF-α concentrations reinforce this interpretation. Under heat stress, the Low line displayed lower TNF-α levels, suggesting a more controlled inflammatory response. Elevated cortisol levels in the High line may compromise immune competence by suppressing lymphoid activity and impairing the balance between pro- and anti-inflammatory cytokines (Ayyat and Marai, 1997; Liang et al., 2022). This imbalance has been described in other livestock species, where heat stress enhances systemic inflammation and increases susceptibility to infections (Lara and Rostagno, 2013; Lemal et al., 2023). Thus, the interaction between glucocorticoids and cytokines could explain the parallel differences observed in both stress physiology and thermoregulation.
Regarding CRP, no differences were found between lines in our study. The discrepancies with Argente et al. (2019) and Beloumi et al. (2020), where the Low line exhibited lower CRP levels, might be related to differences in sex, physiological stage, or timing of sampling, as has been observed in rats (Nunomura, 1992; Balog et al., 2015). It is also possible that CRP, unlike other acute-phase proteins such as haptoglobin, is less sensitive to heat stress in rabbits, as also reported by Argente et al. (2014).
Altogether, these results suggest that resilience-selected animals (Low line) may mitigate the negative effects of heat stress by combining better thermoregulatory efficiency with a more adaptive HPA axis and a moderated inflammatory response. This integrative response could represent a physiological mechanism underpinning resilience, similar to findings reported in other species exposed to heat stress (Laghouaouta et al., 2024; Sejian et al., 2018).
In this research, the assessment of stress relied on three biomarkers (CRP, TNF-α, and cortisol). These markers provide valuable insights into immune and endocrine responses, but the inclusion of additional indicators such as haptoglobin, oxidative stress parameters, or corticosterone, the primary glucocorticoid in rabbits (Bush, 1953), would allow for a more comprehensive understanding of the physiological mechanisms underlying resilience to heat stress. Although cortisol remains a relevant and widely used hormone for evaluating stress in rabbits (Szeto et al., 2004; Verga et al., 2007), complementing its measurement with corticosterone could further refine the interpretation of the stress response.
5 Conclusions
The biomarkers evaluated in this study, together with infrared thermography, represent useful tools for assessing acute stress responses both under heat stress and comfort conditions.
These results highlight the value of selecting for functional traits such as resilience in improving animals’ adaptive responses to heat stress. Thus, incorporating these criteria, specifically selection for litter size homogeneity, into breeding programs contribute to both enhanced animal welfare and sustainable livestock production.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The animal study was approved by General Directorate of Agriculture, Livestock and Fisheries of the Generalitat Valenciana with code 2022/VSC/PEA/0226. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
DS-J: Data curation, Formal analysis, Investigation, Methodology, Writing – original draft. IA: Conceptualization, Methodology, Writing – review & editing. GR: Conceptualization, Methodology, Writing – review & editing. MA: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Supervision, Writing – review & editing. MS: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Supervision, Writing – review & editing. MG: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Supervision, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This study forms part of the AGROALNEXT programme (AGROALNEZT/2022/037) and was supported by MCIN with funding from European Union NextGenerationEU (PRTR-C17.I1) and by Generalitat Valenciana.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abad B., Mesonero J., Salvador M., García-Herrera J., and Rodríguez-Yoldi M. (2002). Tumor necrosis factor-α mediates inhibitory effect of lipopolyaccharide on L-leucine intestinal uptake. Dig Dis. Sci. 47, 1316–1322. doi: 10.1177/096805190200800206
Agea I., García M. L., and Argente M. L. (2021). Preliminary study of body temperature emissivity in rabbits selected for litter size residual variability. Agriculture 11, 604. doi: 10.3390/agriculture11070604
Argente M. J., Calle E. W., García M. L., and Blasco A. (2017). Correlated response in litter size components in rabbits selected for litter size variability. J. Anim. Breed. Genet. 134, 505–511. doi: 10.1111/jbg.12283
Argente M. J., García M. L., Birlanga V., and Muelas R. (2014). Relationship between cortisol and acute phase protein concentrations in female rabbits. Vet. J. 202, 172–175. doi: 10.1016/j.tvjl.2014.07.020
Argente M. J., García M. L., Zbyňovská K., Petruška P., Capcarová M., and Blasco A. (2019). Correlated response to selection for litter size environmental variability in rabbits’ resilience. Animal 13, 2348–2355. doi: 10.1017/S1751731119000302
Ayyat M. and Marai I. (1997). Effects of heat stress on growth, carcass traits and blood components of New Zealand White rabbits fed various dietary energy-fibre levels, under Egyptian conditions. J. Arid Environ. 37, 557–568. doi: 10.1006/jare.1997.0308
Balog M., Mlinaveric D., Seric V., Miljanovic M., Blazekovic R., Degmecic I. V., et al. (2015). Plasma content of glucosa, c-reactive protein, uric acid, and cholesterol in male, female, and ovariectomized rats upon acute and chronic stress – a path for development of cardiovascular diseases. Coll. Antropol 39, 385–392.
Beloumi D., Blasco A., Muelas R., Santacreu M., García M. L., and Argente M. J. (2020). Inflammatory correlated response in two lines of rabbit selected divergently for litter size environmental variability. Animals 10, 1540. doi: 10.3390/ani10091540
Biada I., Serrano-Jara D., Argente M. J., Ibañez-Escriche N., García M. L., and Santacreu M. A. (2024). “Heat stress influence on haematological status and body temperature in two maternal rabbit lines with different longevities,” in Proceedings of the 13th World Rabbit Congress (Ed: World Rabbit Science Association: Tarragona, Spain), 49–52.
Blasco A., Martínez-Álvaro M., García M., Ibáñez-Escriche N., and Argente M. (2017). Selection for environmental variance of litter size in rabbits. Genet. Sel. Evol. 49, 48. doi: 10.1186/s12711-017-0323-4
Bohlinger I., Leist M., Gantner F., Angermüller S., Tiegs G., and Wendel A. (1996). DNA fragmentation in mouse organs during endotoxic shock. Am. J. Pathol. 149, 1381.
Bush I. E. (1953). “Species differences and other factors influencing adrenocortical secretion in rats and rabbits,” in Synthesis and Metabolism of Adrenocortical Steroids. Eds. Klyne W., Wolstenholme G. E. W., and Cameron M. P. (Ciba Foundation Colloquia on Endocrinology, London), 210–232.
Cervera C. and Carmona J. (2010). “Nutrition and the climatic environment,” in Nutrition of the Rabbit. Eds. de Blas C. and Wiseman J. (CABI Publishing, Wallingford), 267–284. doi: 10.1079/9781845936693.0267
Colditz I. G. and Hine B. C. (2016). Resilience in farm animals: biology, management, breeding and implications for animal welfare. Anim. Prod Sci. 56, 1961–1983. doi: 10.1071/AN15297
Cullere M. and Dalle-Zotte A. (2018). Rabbit meat production and consumption: State of knowledge and future perspectives. Meat Sci. 143, 137–146. doi: 10.1016/j.meatsci.2018.04.029
Deem R., Shanahan F., and Targan S. (1991). Triggered human mucosal T cells release tumour necrosis factor-alpha and interferon-gamma which kill human colonic epithelial cells. Clin. Exp. Immunol. 83, 79–84. doi: 10.1111/j.1365-2249.1991.tb05592.x
De Lima V., Piles M., Rafel O., López-Béjar M., Ramón J., Velarde A., et al. (2013). Use of infrared thermography to assess the influence of high environmental temperature on rabbits. Res. Vet. Sci. 95, 802–810. doi: 10.1016/j.rvsc.2013.04.012
Ebeid T., Aljabelli H., Al-Homidan I., Volek Z., and Barakat H. (2023). Ramifications of heat stress on rabbit production and role of nutraceuticals in alleviating its negative impacts: an updated review. Antioxidants 12, 1407. doi: 10.3390/antiox12071407
Eckersall P. and Bell R. (2010). Acute phase proteins: Biomarkers of infection and inflammation in veterinary science. Vet. J. 185, 23–27. doi: 10.1016/j.tvjl.2010.04.009
García M. L. and Argente M. J. (2017). Exposure to high ambient temperatures alters embryology in rabbits. Int. J. Biometeorol 61, 1555–1560. doi: 10.1007/s00484-017-1334-0
García M. L., Blasco A., and Argente M. J. (2016). Embryologic changes in rabbit lines selected for litter size variability. Theriogenology 86, 1247–1250. doi: 10.1016/j.theriogenology.2016.04.065
García M. L., Blasco A., García M. E., and Argente M. J. (2019). Correlated response in body condition and energy mobilisation in rabbits selected for litter size variability. Animal 13, 784–789. doi: 10.1017/S1751731118002203
García-Herrera J., Navarro M., Marca M., de la Osada J., and Rodríguez-Yoldi M. (2004). The effect of tumor necrosis factor-α on D-fructose intestinal transport in rabbits. Cytokine 25, 21–30. doi: 10.3748/wjg.v16.i9.1057
Geyer C. M. (1992). Practical Markov chain Monte Carlo (with discussion). Stat. Sci. 7, 467–511. doi: 10.1214/ss/1177011137
Giorgi F. and Lionello P. (2008). Climate change projections for the Mediterranean region. Glob. Planet. Change 63, 90–104. doi: 10.1016/j.gloplacha.2007.09.005
Gutiérrez A., Martínez-Subiela S., Eckersall P., and Cerón J. (2009). C-reactive protein quantification in porcine saliva: a minimally invasive test for pig health monitoring. Vet. J. 181, 261–265. doi: 10.1016/j.tvjl.2008.03.021
Hennessy M. (1997). Hypothalamic-pituitary-adrenal responses to brief social separation. Neurosci. Biobehav. Rev. 21, 11–29. doi: 10.1016/s0149-7634(96)00013-9
Huang D., Cai J., Zhang C., Jin R., Bai S., Yao F., et al. (2023). Semen quality and seminal plasma metabolites in male rabbits (Oryctolagus cuniculus) under heat stress. PeerJ Life Environ. 11, e15112. doi: 10.7717/peerj.15112
Jaén-Téllez J., Sánchez-Guerrero M., López-Campos J., Valera M., and González-Redondo P. (2020). Acute stress assessment using infrared thermography in fattening rabbits reacting to handling under winter and summer conditions. Span. J. Agric. Res. 18, e0502. doi: 10.5424/sjar/2020182-15706
Jaén-Téllez J., Sánchez-Guerrero M., Valera M., and González-Redondo P. (2021). Influence of stress assessed through infrared thermography and environmental parameters on the performance of fattening rabbits. Animals 11, 1747. doi: 10.3390/ani11061747
Koolhaas J. M., Bartolomucci A., Buwalda B., de Boer S. F., Flügge G., Korte S. M., et al. (2011). Stress revisited: a critical evaluation of the stress concept. Neurosci. Biobehav. Rev. 35, 1291–1301. doi: 10.1016/j.neubiorev.2011.02.003
Kushner I. (1988). The acute phase response: An overview. Methods Enzymol. 163, 373–383. doi: 10.1016/0076-6879(88)63037-0
Laghouaouta H., Fraile L. J., and Pena R. N. (2024). Selection for resilience in livestock production systems. Int. J. Mol. Sci. 25, 13109. doi: 10.3390/ijms252313109
Lara L. J. and Rostagno M. H. (2013). Impact of heat stress on poultry production. Animals 3, 356–369. doi: 10.3390/ani3020356
Lavara R., Vicente J., and Baselga M. (2011). Genetic parameter estimates for semen production traits and growth rate of a paternal rabbit line. J. J. Anim. Breed. Genet. 128, 44–51. doi: 10.1111/j.1439-0388.2010.00889
Lemal P., May K., König S., Schroyen M., and Gengler N. (2023). Invited review: From heat stress to disease—Immune response and candidate genes involved in cattle thermotolerance. J. Dairy Sci. 106, 4751–4771. doi: 10.3168/jds.2022-22727
Liang Z., Chen F., Park S., Balasubramanian B., and Liu W. (2022). Impacts of heat stress on rabbit immune function, endocrine, blood biochemical changes, antioxidant capacity and production performance, and the potential mitigation strategies of nutritional intervention. Front. Vet. Sci. 9. doi: 10.3389/fvets.2022.906084
Marai I., Ayyat M., and Abd El-Monem U. (2001). Growth performance and reproductive traits at first parity of New Zealand White female rabbits as affected by heat stress and its alleviation under Egyptian conditions. Trop. Anim. Health Prod 33, 451–462. doi: 10.1023/a:1012772311177
Marai I., Habeeb A., and Gad A. (2002). Rabbits’ producive, reproductive and physiological performance traits as affected by heat stress: a review. Livest. Prod. Sci. 78, 71–90. doi: 10.1016/S0301-6226(02)00091-X
McEwen B. S. and Wingfield J. C. (2003). The concept of allostasis in biology and biomedicine. Horm. Behav. 43, 2–15. doi: 10.1016/s0018-506x(02)00024-7
Moberg G. P. (2000). “Biological response to stress: Implications for animal welfare,” in The biology of animal stress: Basic principles and implications for animal welfare. Eds. Moberg G. P. and Mench J. A. (CABI Publishing, Wallingford), 1–21. doi: 10.1079/9780851993591.0001
Mormede P., Boisseau-Sowinski L., Chiron J., Diederich C., Eddison J., Guichet J., et al. (2018). Bien-être animal: context, definition, évaluation. INRAE Prod. Anim. 31, 145–162. doi: 10.20870/productions-animales.2018.31.2.2299
Möstl E. and Palme R. (2002). Hormones as indicators of stress. Domest. Anim. Endocrinol. 23, 67–74. doi: 10.1016/s0739-7240(02)00146-7
Nielsen S., Álvarez J., Bicout D., Calistri P., Canali E., Drewe J., et al. (2022). Welfare of domestic birds an rabbits transported in containers. EFSA J. 20, e07441. doi: 10.2903/j.efsa.2022.7441
Nunomura W. (1992). C-reactive protein (CRP) in animals: its chemical properties and biological functions. Zool. Sci. 9, 499–513.
Oladimeji A., Johnson T., Metwally K., Farghly M., and Mahrose K. (2022). Environmental heat stress in rabbits: implications and ameliorations. Int. J. Biometeorol 66, 1–11. doi: 10.1007/s00484-021-02191-0
Oohashi E., Kimura Y., and Matsumoto K. (2019). Pilot study on serum C-reactive protein in pet rabbits: clinical usefulness. Vet. Rec. Open 6, e000272. doi: 10.1136/vetreco-2017-000272
Pei Y., Wu Y., Cao J., and Qin Y. (2012). Effects of chronic heat stress on the reproductive capacity of male Rex rabbits. Livest. Sci. 146, 13–21. doi: 10.1016/j.livsci.2012.02.015
Rauw W., Kanis E., Noordhurizen-Stassen E., and Grommers F. (1998). Underisable side effects of selection for high production efficiency in farm animals: a review. Livest. Prod. Sci. 56, 15–33. doi: 10.1016/S0301-6226(98)00147-X
Saco Y. and Bassols A. (2022). Acute phase proteins in cattle and swine: A review. Vet. Clin. Pathol. 52, 50–63. doi: 10.1111/vcp.13220
Sejian V., Bhatta R., Gaughan J. B., Dunshea F. R., and Lacetera N. (2018). Review: Adaptation of animals to heat stress. Animal 12, s431–s444. doi: 10.1017/S1751731118001945
Serrano-Jara D., Agea I., Díaz J., Argente M., and García M. (2025). Stress analysis due to semen collection using infra-red thermography in rabbits. Ital. J. Anim. Sci. 24, 996–1007. doi: 10.1080/1828051X.2025.2491753
Sorensen D. and Gianola D. (2002). Likelihood, Bayesian and MCMC methods in quantitative genetics (New York: Springer). doi: 10.1007/b98952
Suter P., Suter S., Girardin E., Roux-Lombard P., Grau G., and Dayer J. (1992). High bronchoalveolar levels of tumor necrosis factor and its inhibitors interleukin-1, interferon, and elastase, in patients with adult respiratory distress syndrome after trauma, shock, or sepsis. Am. Rev. Respir. Dis. 145, 1016–1022. doi: 10.1164/ajrccm/145.5.1016
Szeto A., Gonzales J., Spitzer S., Levine J., Zaias J., Saab P., et al. (2004). Circulating levels of glucocorticoid hormones in WHHL and NZW rabbits: circadian cycle and response to repeated social encounter. Psychoneuroendocrinology 29, 861–866. doi: 10.1016/S0306-4530(03)00153-7
Takao T., Nanamiya W., Takemura T., Nishiyama M., Asaba K., Makino S., et al. (1997). Endotoxin induced increases in rat plasma pituitary-adrenocortical hormones are better reflected by alterations in tumor necrosis factor α than interleukin-1β. Life Sci. 61, PL263–PL268. doi: 10.1016/S0024-3205(97)00780-7
Verga M., Luzi F., and Carenzi C. (2007). Effects of husbrandy and management systems on physiology and behaviour of farmed and laboratory rabbits. Horm. Behav. 52, 122–120. doi: 10.1016/j.yhbeh.2007.03.024
Yağci A., Zik B., Uguz C., and Altunbas K. (2006). Histology and morphometry of white New Zealand rabbit skin. Indian Vet. J. 83, 876–880.
Yan Y. and Li M. (2008). “Feeding management and technology of breeding rabbit in hot climate,” in Proceeding of the MEKARN Workshop 2008: Organic rabbit production from forages (Can Tho City, Vietnam: Ed: MEKARN Project, Cantho University), 25–27.
Keywords: resilience, heat stress, infra-red thermography, litter size variability, stress biomarkers, CRP, TNF-α, cortisol
Citation: Serrano-Jara D, Agea I, Romero G, Argente MJ, Santacreu MA and García MdlL (2025) Genetic correlated responses to selection for resilience in key inflammatory and stress biomarkers under heat stress of rabbits. Front. Anim. Sci. 6:1694508. doi: 10.3389/fanim.2025.1694508
Received: 28 August 2025; Accepted: 08 October 2025;
Published: 28 October 2025.
Edited by:
Francisco Javier Navas González, University of Cordoba, SpainReviewed by:
Hui Wen, Purdue University, United StatesArnulfo Montero Pardo, Universidad Autonoma de Sinaloa, Mexico
Copyright © 2025 Serrano-Jara, Agea, Romero, Argente, Santacreu and García. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Daniel Serrano-Jara, ZC5zZXJyYW5vQHVtaC5lcw==