 Margherita Greco1
Margherita Greco1 Annalaura Lopez1*
Annalaura Lopez1* Vittorio Maria Moretti1
Vittorio Maria Moretti1 Silvana Mattiello2
Silvana Mattiello2 Beatrice Marcolli1
Beatrice Marcolli1 Stefania Celozzi2
Stefania Celozzi2 Federica Bellagamba1
Federica Bellagamba1- 1Department of Veterinary Medicine and Animal Science, Università degli Studi di Milano, Via dell’Università, Lodi, Italy
- 2Department of Agricultural and Environmental Sciences, Università degli Studi di Milano, Via Celoria, Milan, Italy
This study evaluated milking performance and compositional characteristics of milk from ten Ciuta ewes maintaining concurrent lamb suckling throughout an 11-week experimental period. Daily milk yield averaged 345 ± 115g per animal during single milking sessions, with peak individual performances reaching 698g, while lambs continued partial milk consumption. Compositional analysis revealed lactose concentrations of 4.8 ± 0.2%, protein levels of 5.8 ± 0.5%, and fat content of 4.6 ± 1.2%. Somatic cell count remained consistently below 250,000 cells/mL, indicating a good mammary health status. (mean 100 ± 103 cells/mL) The fatty acid profile demonstrated nutritionally favourable characteristics, including an optimal n-6/n-3 ratio of 1.43, alongside significant proportions of branched-chain fatty acids (2.94 ± 0.4%) and conjugated linoleic acid (1.24 ± 0.3%). Observations on udder morphology indicate the potential for favourable udder traits that require further investigation. Behavioural assessment during milking procedures showed progressive adaptation of Ciuta ewes, with discomfort signs declining across the trial duration. Individual variation in milking temperament suggested potential for selection and improvement for adaptation to the milking routine. The outcomes from this study provided original knowledge, suggesting a promising integration of Ciuta sheep into milking while maintaining their traditional rusticity and resilience.
1 Introduction
The conservation and promotion of local breeds are part of a multi-dimensional approach to sustainable development, where biodiversity represents one of the key pillars for ensuring sustainability, food security and nutritional quality (FAO, 2019). Aside from their ecological role, local breeds also have cultural and economic relevance. Indeed, they are a valuable resource for preservation of genetic variability and socio-economic development of their native territories through their association with cultural heritage and traditional products (Lauvie et al., 2023). As a matter of fact, the demand for niche and quality products from local breeds has been constantly growing, promoting the valorisation of rural landscapes and economic opportunities for local producers and tourism. In line with this trend, in the last years consumers’ willingness to pay for products that adhere to sustainable production practices has been growing, driven by awareness about health, environmental sustainability and animal welfare (Bojovic and McGregor, 2023; Madududu et al., 2024). However, only a comprehensive characterization of local breeds and authentication of their products through certification and labelling can lead to positive outcomes (Skordos et al., 2024).
Sheep production represents a sustainable resource with possibilities of economic profitability and demographic stability, of special importance for arid, semi-arid and other unbendable regions (FAO, 2009). Even in the mountainous or semi-mountainous areas of Italy, autochthonous sheep breeds are reared under extensive or semi-extensive management regimes that are valuable in preserving genetic variability. In this context, autochthonous dairy sheep can play a strategic role in generating dairy products with strong territorial identity due to a long history of contributing to the resilience of marginal areas, such as alpine landscapes (Benedetti del Rio et al., 2024; Claps et al., 2020). Thanks to their adaptability and co-evolution with the territories, these breeds can use pastures in marginal areas more efficiently than other livestock, thanks to their low requirements for external feed input. Also, they contribute to environmental valorisation by supporting traditional transhumant and extensive grazing practices while providing high-quality food that adds value to the whole supply chain of Alpine products (Santillo et al., 2025; Sturaro et al., 2013).
However, many local breeds are still poorly documented, limiting our understanding of their farming system, their role in the environment and their potential for high-quality and sustainable products. A notable example is the Ciuta sheep, an indigenous sheep breed originating from Valtellina and Alto Lario, whose milk quality and milking aptitude remain largely uncharacterized. The Ciuta has been preserved in marginal areas such as Val Màsino, a small side valley of Valtellina (Central Italian Alps), where agriculture followed a subsistence rather than a commercial rationale. Recently rescued from extinction, the Ciuta sheep represents a valuable genetic resource in the pre-Alpine area and a strategic asset for the exploitation of territorial identity and preservation of traditional agro-pastoral heritage. Currently, 577 animals are registered in the herdbook held by ASSONAPA (Italian National Sheep Breeders’ Association).
The Ciuta is considered the smallest sheep breed in the Alpine area: adult males have a withers height of 45–50 cm and a body weight of 35–40 kg, while females measure 40–45 cm at withers and weigh 30–35 kg (Bigi and Zanon, 2020). A typical feature of this breed is the presence of horns in both sexes: in males, the horns are spiral-shaped, while in females, when present, they are smaller and resemble those of goats. The Ciuta is resilient and frugal, well adapted to a harsh environment: for most of the year, the animals graze on steep pastures at elevations ranging from 500 to 2,700 m a.s.l.; only in the winter months they are fed with hay.
This breed is primarily suited for meat production, although writings from the late 1800s recall it as a triple-purpose animal. In Val Masino, a side valley of the Valtellina region in northern Italy, its milk was historically used, alongside cow and goat milk, to produce a “three-milk” cheese (Turri, 2025). The fleece is mostly straw-white coloured, though other colour variants exist, including brown, black, and spotted. Wool production is about 1.4 kg per animal; the wool is ordinary (fibre average diameter 30 µm), but suitable for felting (Lopez et al., 2024).
The milking aptitude and milk quality of Ciuta sheep remain largely unexplored. This lack of data limits both the breed’s genetic preservation and its productive integration into certified quality chains, as these traits are essential to assess the productive potential in a small-scale or artisanal production context. Furthermore, on-farm milk availability and cheese production could offer farmers an economic advantage, particularly when marketed and consumed directly through agritourism activities.
The objective of this study was to provide the first comprehensive characterization of the milk quality and milking potential of the Ciuta sheep, including the morphological evaluation of the udder and sheep’s adaptation to milking routine with the broader aim of highlighting the importance of this local breed in maintaining pastureland health, enhancing biodiversity, and supporting rural livelihoods.
2 Materials and methods
All the procedures described in the following paragraphs have been approved by the Animal Welfare Organization of the University of Milan as non-experimental protocol with the number 36_2024 in date 03/05/2024.
2.1 Animal management
The experiment was conducted in a farm located in Valtellina using 10 Ciuta ewes, part of a flock of 51 animals, with an average body weight of 36.4 kg. Two ewes were primiparous, three were in their second parity, and the others were in their third to sixth parity. The size of the experimental group was constrained by the limited availability of animals within the flock, which did not allow a balanced experimental design in terms of parity and stage of lactation.
The ewes lambed between 13th December 2023 and 16th January 2024 and suckled their offspring for 35 to 69 days before the beginning of the trial. The study lasted for 11 weeks, from March to April 2024. Animals were managed under a partial suckling regime: ewes and lambs grazed daily on farm grassland for 7 hours (11:00 - 18:00), then they were housed indoors for 17 hours (18:00-11:00), with ewes and lambs kept in separated pens. To limit stress and support maternal behaviour, visual contact between ewes and lambs was always maintained, and they were rejoined after the morning milking.
Milking was carried out manually once per day, using a raised wooden stanchion equipped with two stalls, which allowed the simultaneous milking of two ewes. During milking, each ewe received 200 g of concentrate feed, while local hay was provided ad libitum throughout the trial. The chemical composition of the hay and concentrate can be found in Table 1.
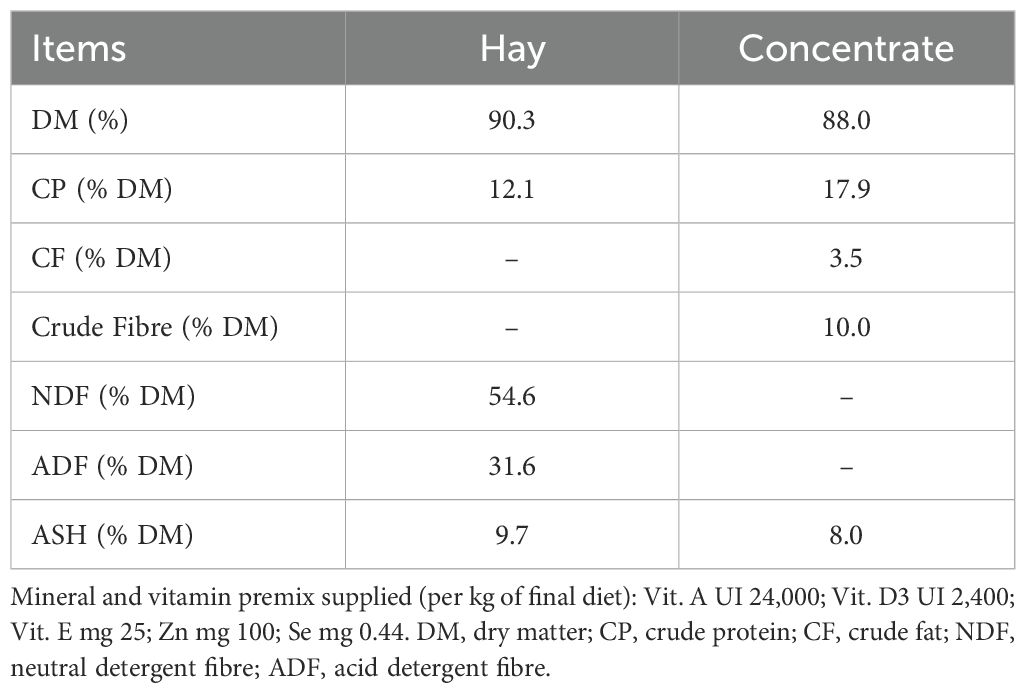
Table 1. Chemical composition of the concentrate and hay offered to ewes.
2.2 Milk sampling
Individual milk samples were collected twice a week, on Monday and Wednesday, from 19th February to 27th March, corresponding to experimental days 1, 3, 8, 10, 15, 17, 22, 24, 29, 31, 36, 38, 43, and 45. Starting from day 45, sampling was reduced to once a week and continued until April 29th. Seventeen sampling sessions were carried out, and a total of 170 milk samples were collected. Individual milk production was measured by manual weighing. The milk samples were divided into two aliquots of approximately 50 mL each. The first was transported under refrigerated conditions (+4 °C) to the Department laboratories and stored at −20 °C until fatty acid analysis. The second aliquot was poured in a 50 mL tube and promptly delivered to the laboratory of the Regional Breeders Association of Lombardy (ARAL) for compositional analysis.
2.3 Milk composition and fatty acid profile
Milk samples were analysed for fat, protein, casein, lactose, urea, and somatic cell count (SCC) using a CombiFoss™ 7 analytical system (FOSS, Hillerød, Denmark). This integrated unit combines the MilkoScan™ 7 RM, which applies Fourier Transform Infrared (FTIR) spectroscopy, and the Fossomatic™ 7, which uses flow cytometry for SCC analysis. All analyses were conducted at room temperature.
To determine the fatty acids (FAs) composition of milk, weekly samples from each ewe were combined. Pooling was adopted based on the assumption that, under consistent management conditions, within-milking variation in fatty acid composition is minimal (Rico et al., 2014), and short-term pooling does not compromise the representativeness of the samples. FAs methylation was performed without lipid extraction, with a modified dried milk method from Liu et al. (2020). In summary, a 5 mL glass vial was filled with 150 μL of raw milk (corresponding approximately to 7.5–10 mg of lipids) and, after drying the sample under N2 in a heating block at 40 °C, milk fat was processed to trans-esterification in cold alkaline environment, following the method described by Christie (2003). Fatty acid methyl esters (FAMEs) were analysed by gas-chromatography coupled to flame-ionisation detection (GC-FID) using a Thermo Fisher gas-chromatograph (model TRACE™1300; Thermo Fisher Scientific, Waltham, MA, USA) equipped with an autosampler (model AI 1310; Thermo Fisher Scientific). A 120 m fused silica capillary column (TRACE TR-FAME 0.25μL film thickness, 0.25mm I.D.; Thermo Fisher Scientific) and helium as carrier gas (1.0mL/min flow rate) were used. The injector was set at 250 °C in split operating mode (1:20 split ratio). The oven temperature program started with an isotherm of 8 min at 50 °C, then the temperature increased at the rate of 12 °C/min until 173 °C and kept at this temperature for 47 min. Afterwards the temperature was increased at 4 °C/min rate until reaching the 220 °C, which was hold for 20 min. Data was acquired through the software Chromeleon (Thermo Fisher Scientific, Waltham, MA, US). Peaks of individual FAMEs were identified by retention times comparison with standard mixture (Supelco FAME 37 Mix, Bellafonte, PA, USA, plus several individual FAMEs) and expressed as percentage over total FAMEs. Individual FAs were summed to obtain the following categories saturated fatty acids (SFAs), monounsaturated fatty acids (MUFAs), polyunsaturated fatty acids (PUFAs), branched chain fatty acids (BCFAs), odd chain fatty acids (OCFAs), n-3 and n-6 PUFAs and n-6/n-3 PUFA ratio.
Furthermore, in order to characterize the nutritional quality and value of milk lipids, Atherogenic (AI) and Thrombogenic (TI) Index were calculated through the method described by Ulbricht and Southgate (1991), according to the following formulas: equation 1: AI= (C12:0+C14:0+C16:0)/(MUFAs+PUFAs); equation 2: TI= (C14:0+C16:0+C18:0)/(0.5*MUFAs+0.5*PUFAs n-6+PUFAs n-3 + n-3/n-6)
These two indices represent concise and reliable tools for assessing the potential impact of dietary fats on cardiovascular health. Their determination adds value to the nutritional characterization of traditional or niche products, particularly within sustainable and health-oriented production chains, such as those in Alpine regions.
2.4 Milk pH and colour assessment
Milk pH was measured using a pH/mV/Temp bench meter (model XS pH50 + DHS; Giorgio Bormac, Carpi, MO, Italy) equipped with pH probe (XS Sensor 201T; Giorgio Bormac). The instrument was calibrated using a two-point procedure with standard buffer solutions at pH 4.00 ± 0.02 and pH 7.00 ± 0.02, in accordance with the manufacturer’s guidelines.
The colour evaluation of the milk was carried out according to Kiełczewska et al. (2021). Samples were poured into standard Petri dishes to a uniform depth of approximately 1 cm and analysed at room temperature (20–22 °C) with controlled lighting conditions following homogenization using an ULTRA-Turrax for 30 seconds to minimize fat separation and environmental variability. Surface colour was assessed using a colorimeter CR-400 Chroma Meter (Konica Minolta Sensing Europe, Inc., Nieuwegein, UT, Netherlands). The device operates with an 8 mm aperture and D65 illuminant (colour temperature 6500K, pulsed xenon lamp). To ensure data consistency, the instrument was calibrated with the manufacturer-supplied white reference tile (Y = 84.7, x= 0.3177 and y= 0.3336). Colorimetric data were recorded in the CIE Lab* colour space (L*a*b*) according with default data acquirement, where L* indicates lightness, a* the red-green axis, b* the yellow-blue axis. In addition, the parameters chroma or saturation (C*) and hue (h*) were calculated to assess the cylindrical representation of the CIELAB system following the formula described by Milovanovic et al. (2020):equation 3: C*= (a*2+b*2)0.5; equation 4: H*= tan-1(b*/a*). Furthermore, to characterize the visual appearance of the milk, the Whiteness (WI) and Yellowness (YI) index were calculated following the equations reported by Milovanovic et al. (2021): equation 5: WI=100-[(100-L*)2+a*2+b*2)0.5; equation 6: YI= 142.86 (b*/L*).
The WI indicates the degree of whiteness through the combination of lightness and yellow-blue into a single term. YI indicates the degree of yellowness related to lightness. These indices provide a more integrated and perceptually relevant representation of colour differences among milk samples in a way that aligns with human visual perception and food quality assessment.
2.5 Udder morphological appraisal
The udder appraisal was carried out using the method of Casu et al. (2006). This method considers four udder traits, each scored with a 9-point linear scale: teat placement (TP), udder depth (UD), degree of separation of the halves (DS) and degree of suspension of the udder (SU).
For teat placement, the distance between the teats and the lower surface of the udder is taken into account. Scores range from 1, for teats placed on the lowest part of the udder (vertical), to 9, for teats located above the area with the maximum radius of curvature (horizontal).
Udder depth is the distance between the udder lower profile and the abdominal wall, taking as a reference point the line joining the hocks. It is scored from 1, for deep udders near to the ground, to 5, when udder profile is at the hock level, to 9, for shallow udders close to the abdominal wall.
The degree of separation scores the strength of the suspensor ligament, from 1, with no separation between the left and right glands, to 9, for udders clearly divided into two halves.
The degree of suspension is the ratio between the width of the rear udder attachment and the udder height. It is scored from 1, for udders with an attachment width smaller than the udder height, to 7, for square udders, to 9, for udders with attachment width larger than the udder height. Scoring was performed just prior to milking by two operators.
Figure 1 shows a representative photo of the udder of one ewe involved in the trial with the four udder traits evaluated.
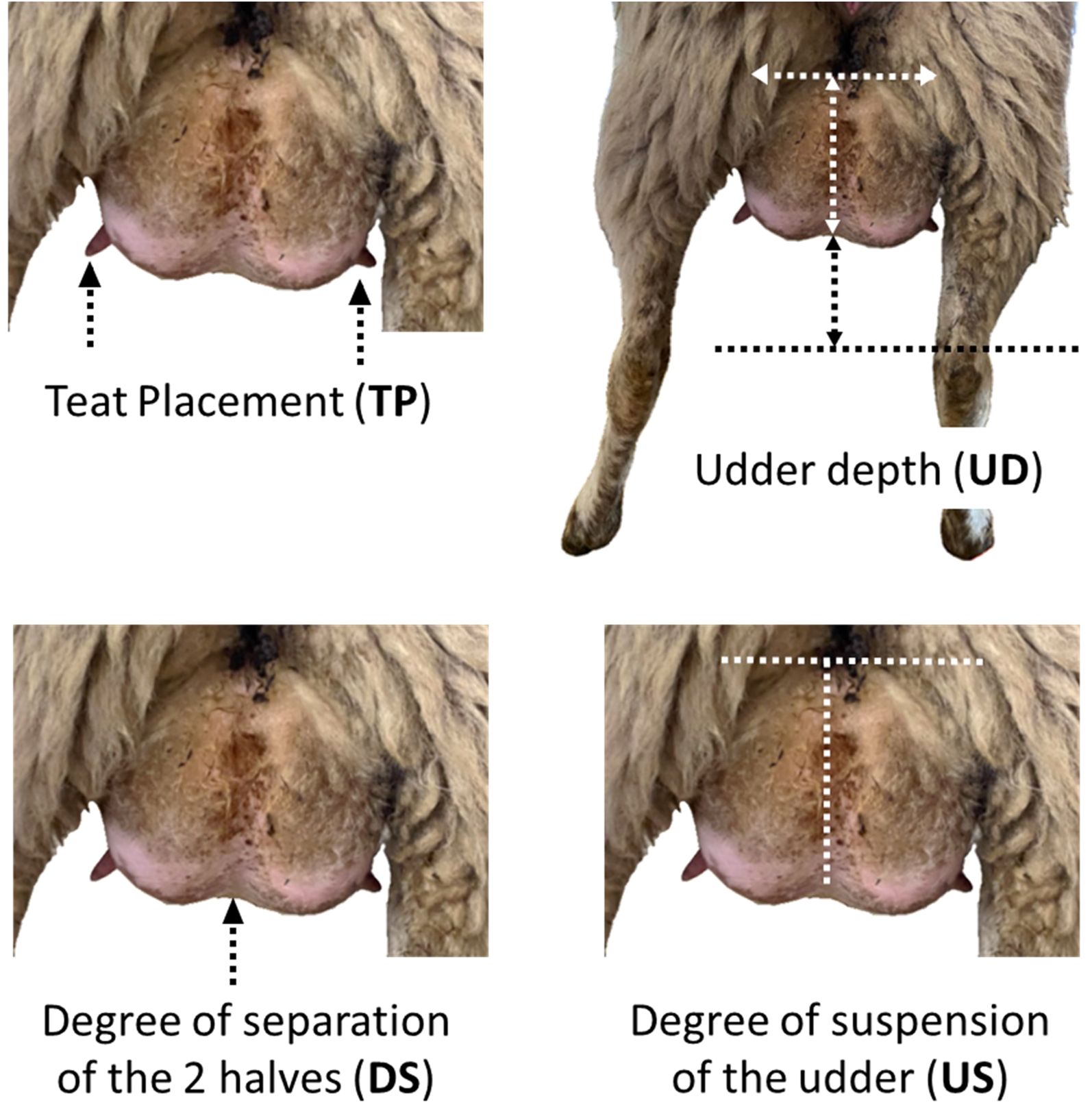
Figure 1. Representative photo of the udder appraisal on one Ciuta ewe involved in the trial as indicated by Casu et al. (2006).
2.6 Milking behaviour
During each milk sampling, sheep behaviour was recorded using a stationary camera (Panasonic HC-VX1) placed at 2.5 meters from the milking area and at 1 meter height to provide a complete view. The camera was placed behind the milking area and the video included both the two ewes that were milked at the same time. Video recordings were acquired with the following technical specifications: a frame rate of 25.00 frame per second, 3840 × 2160-pixel resolution, 60’808 kbps total bitrate. Recordings began as the ewes entered the milking station and ended upon reunion with the lamb. Each session was divided into pre-milking (including moving the sheep from the pen to the milking place, and preliminary manipulation and dipping), milking (from the start to the end of the milk extraction from the udder), and post-milking (including post-dipping and return to the home pen) phases. Environmental factors, such as lighting and ambient noise, were kept consistent to minimize variability across sessions. Three of the originally recorded sessions were excluded from the analysis due to file corruption, resulting in 14 usable sessions for behavioural analysis. Overall, 34 hours of video material were analysed.
Continuous observations were performed by a trained observer (blind to most of the information about ewes, e.g., delivery number, age, week of milking) for each milking session, defined as the time interval between the end of pre-milking procedures (teats cleaning and disinfection) and the start of post-dipping procedures (application of a soothing and disinfectant barrier on the teat). The Behavioral Observation Research Interactive Software (BORIS) (Friard and Gamba, 2016) was used to allow for standardized and time-aligned event coding. A structured ethogram was developed to identify potential indicators of discomfort or stress, including: turning head - the ewe turns its head backward, typically toward the udder or milker; kicking - the ewe performs a backward leg movement; turning body - the ewe shifts its entire body axis, altering its orientation within the milking area; moving - the ewe is stepping, shifting its weight between limbs, or is flinching, exhibiting restlessness in place; and tail moving – the ewe is changing the position of its tail. Kicking and body movements (turning body and moving/flinching) were chosen as discomfort indicators based on the EFSA Scientific Opinion on the welfare risks related to the farming of sheep for wool, meat and milk production (EFSA AHAW Panel, 2014); tail movements were included based on previous research in sheep (Reefmann et al., 2009); finally, we also included head moving, that had previously been used as discomfort indicator in goats (Andrioli et al., 2024).
Behaviours were recorded as the number of events over the duration of the milking phase, and their frequency per minute was then calculated. To quantify the general behavioural reaction of the animals to the milking procedures, an overall discomfort score (ODS) was calculated by summing the frequencies of all the above-mentioned discomfort-related behaviours and calculating the frequency per minute of milking phase. The individual duration of the milking phase within each milking session was also recorded and was expressed in minutes.
2.7 Statistical analysis
All statistical analyses were performed using JMP® Pro18 (SAS Institute) and IBM SPSS Statistics. Descriptive statistics (mean, standard deviation, standard error of the mean and range) were computed for all continuous variables, including milk yield, chemical composition, FAs profile, pH, and colour coordinates (CIE L*, a*, b*, h*, C*). To assess milking aptitude (milking duration and occurrence of discomfort behaviours), a preliminary square root transformation (y= √x+0.5) was applied to the selected behavioural response variables to reduce right-skewness in their distributions.
All variables were analysed using a linear mixed model (LMM) including the “day of sampling” as fixed factor to account for the progression of milking adaptation throughout lactation. To account for variability among ewes and the non-independence of repeated observations on the same animal, the ewe was included in the model as a random effect. When the fixed effect was statistically significant (p < 0.05), Tukey HSD post hoc test was employed for pairwise comparisons for the observations made on the different days of sampling.
3 Results
Descriptive statistics of the average yield and chemical parameters of milk are shown in Table 2. On average, a total of 3.5 kg of milk was collected per milking session from the group of 10 ewes included in the trial. The average milk yield recorded during the daily manual milking session was 345 g per ewe, with a statistically significant individual variation (p<0.0001), from a minimum of 109 g to a maximum of 698 g observed in two different ewes. The average chemical composition of milk was characterized by 4.6% fat, 5.76% protein, 4.80% lactose, 4.31% casein. Milk urea content (MUC) mean value was 38.5 mg/dl (maximum value of 63.2 mg/dl), and the average SCC was equal to 100,000 cells/mL.
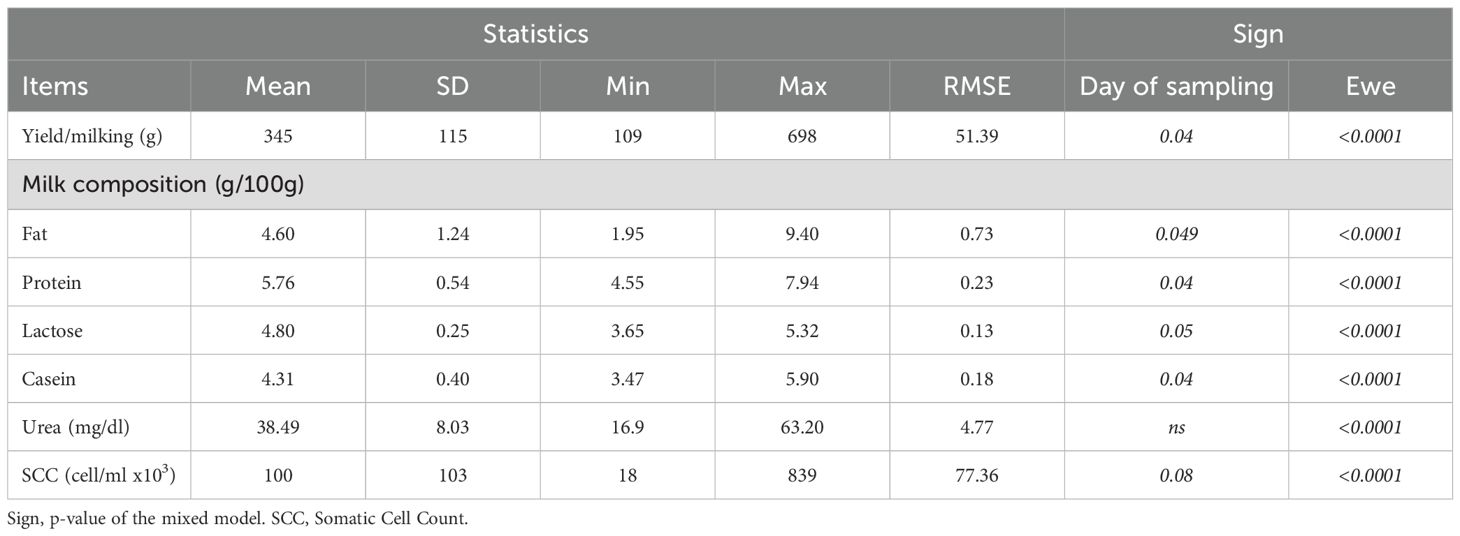
Table 2. Average milk yield and chemical composition (N = 170).
Fatty acids (FAs) composition of milk is presented in Table 3. The average FAs profile was represented by 61.9% saturated FAs (SFAs), 26.6% monounsaturated FAs (MUFAs), 6.0% polyunsaturated FAs (PUFAs), 2.9% branched-chain FAs (BCFAs), and 2.4% odd-chain FAs (OCFAs). Moreover, the PUFAs fraction showed an average concentration of c9,t11-18:2 (CLA) equal to 1.24 and an average n-6/n-3 ratio of 1.43. Finally, the atherogenic index (AI) and thrombogenic index (TI) averaged 2.19 and 2.29, respectively.
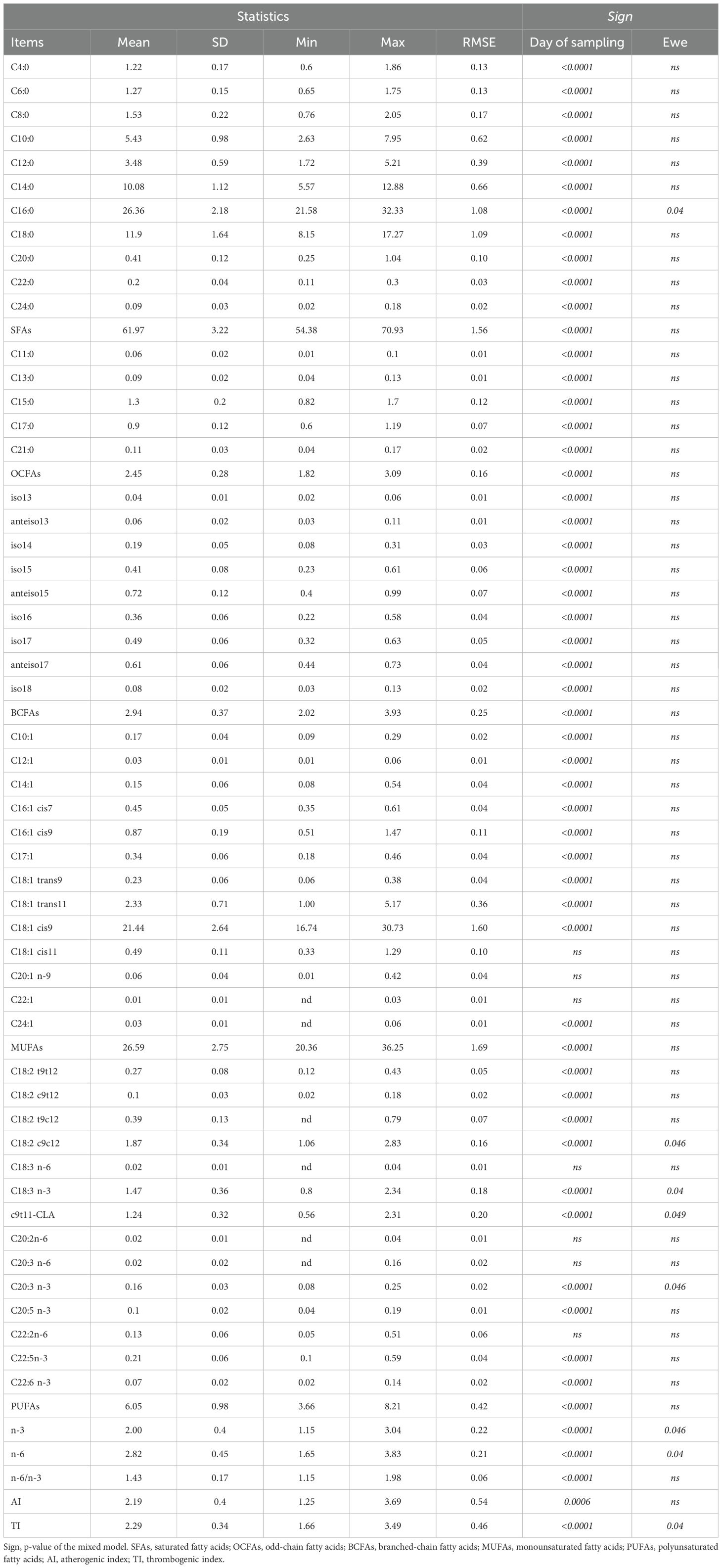
Table 3. Fatty acids composition (g/100g of total fatty acids) in Ciuta sheep milk samples (N = 110).
Descriptive statistics of average pH and colour parameters of milk are shown in Table 4. Milk showed pH values ranging from 6.31 to 6.97, with an overall mean of 6.63 and exhibited a generally white appearance, as indicated by the high average lightness value (L* = 86.52). Its colour profile was characterized by a moderate yellow saturation (b* = 12.40) and negative a* values which indicate green hues (a* = –8.87). The chromatic coordinates (Chroma = 12.44; Hue angle = –0.95) further confirmed the visual impression of a white milk with a perceptible greenish-yellow tint. The average white index (WI) and yellow index (YI) were equal to 79.60 and 20.47, respectively.
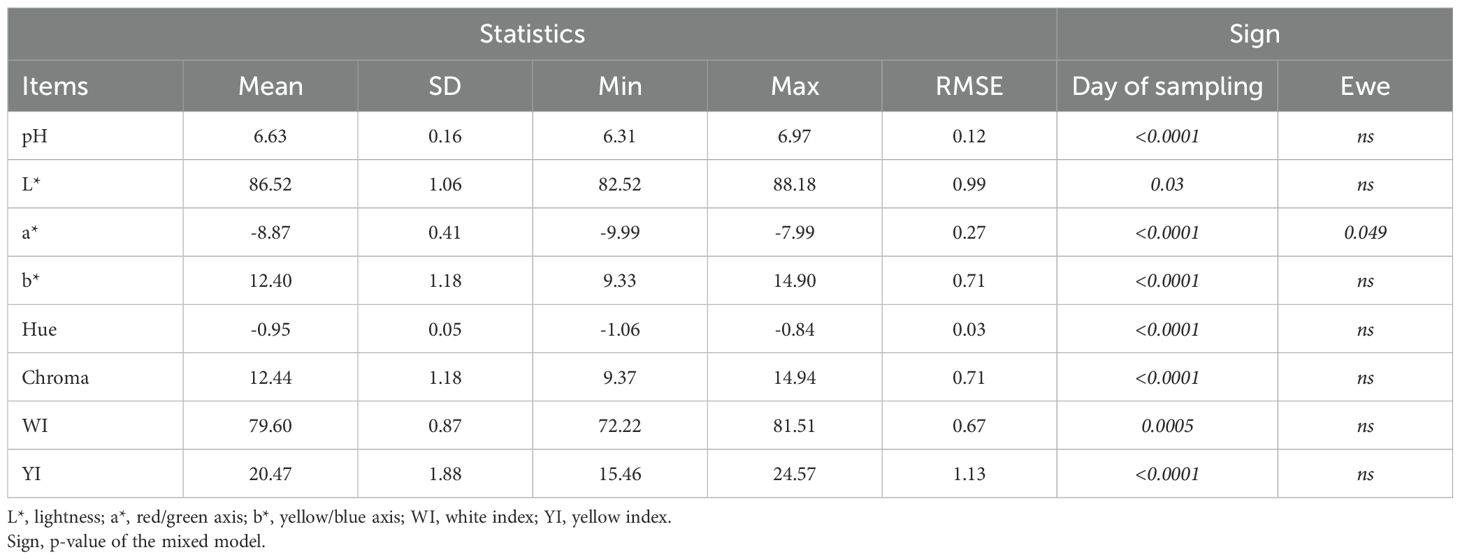
Table 4. pH and colour parameters in Ciuta sheep milk samples (N = 110).
The data presented in Table 5 provided a quantitative overview of the udder morphology in Ciuta ewes. Udder showed an average degree of suspension (US) of 6.46, with and udder depth (UD) of 6.54, a teat placement (TP) of 5.71 and a degree of separation (DS) of 2.99, on average.
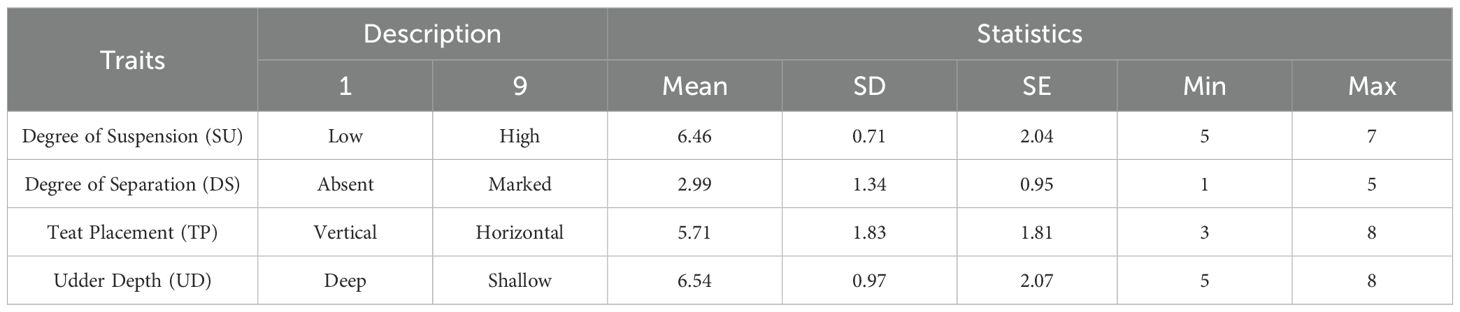
Table 5. Udder traits scores in Ciuta ewes (N = 10).
Descriptive statistics and results of statistical analysis for milking duration and for the frequency of discomfort behaviours are reported in Table 6. Overall, discomfort behaviours were mainly represented by moving (51% of the events) and kicking (32%). The other behaviours were observed at lower frequencies (turning body 10%; turning head 3% tail moving 3%) and therefore were not analysed in further detail, even though they were included in the calculation of the overall discomfort score (ODS). A statistically significant decrease of the duration of milking was observed (p=0.0131; Figure 2A), with a maximum average duration recorded on d08 (10.9 ± 0.63 min) and a minimum average duration on d50 (6.8 ± 0.83 min). No statistical difference among ewes was observed for the duration of milking. In line with the decrease of milking duration, the frequency of ODS also showed a highly significant decreasing trend (p<0.0001; Figure 2B), lowering from a maximum of 5.8 ± 1.7 events/min recorded on d15 to a minimum of 1.4 ± 0.40 on the last day of the trial (d64). Individual variation among ewes was statistically significant for the ODS (p=0.0404). The identical trend was observed for moving and kicking (Table 6). In particular, kicking was never recorded during the last observation day.
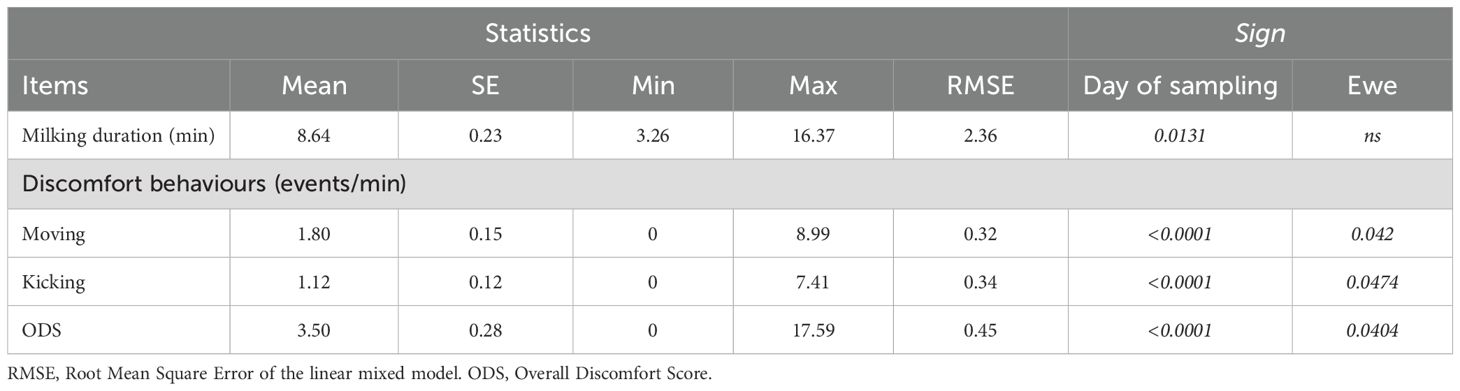
Table 6. Descriptive statistics and results from the mixed model on the milking duration (min) and the main discomfort behaviours (events/minute) observed in Ciuta sheep (N = 140).
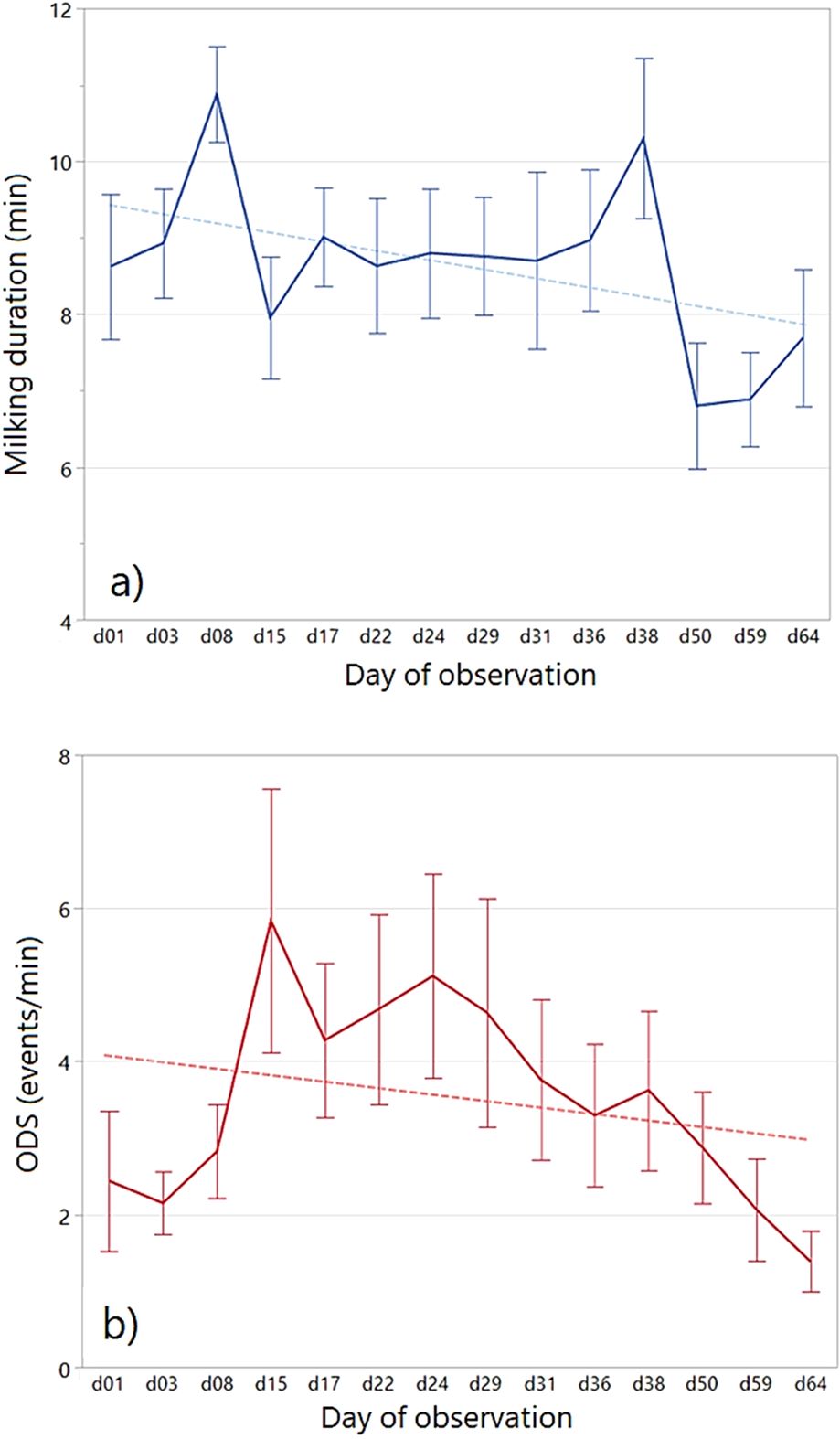
Figure 2. Temporal trend of (A) the duration of the milking phase, and (B) the overall discomfort score (ODS) (average individual values ± SE).
4 Discussion
Evidence from the literature suggests that the average milk yield recorded in the Ciuta sheep is markedly lower if compared to other indigenous multi-purpose sheep breeds. For example, Benedetti del Rio et al. (2024) reported for the Alpagota and Brogna breeds, an average daily yield between 0.8 and 1.2 litres recorded in 90 days of lactation. Signorelli et al. (2008) analysed milk yield and fatty acid composition in three Italian sheep breeds raised under similar management conditions. Over 140 days of lactation, average milk production was 1.07 litres in Altamurana, 0.65 in Gentile di Puglia, and 1.73 in the Sarda breed. A similar trend was also observed in native breeds selected for dairy production, such as the Comisana, whose daily yield ranges between 0.62 and 1.38 litres per lactation (Selvaggi et al., 2017; Stocco et al., 2025; Tufarelli et al., 2009), and the Valle del Belice breed, for which an average daily production of approximately 1.35 litres has been estimated (Cappio-Borlino et al., 1997).
(Simitzis et al., 2021; Ploumi et al., 1998; Dove, 1988)
The lower milk yield observed in Ciuta sheep was not unexpected and can be attributed to several factors. First, this was the first time that Ciuta ewes had been subjected to milking, as the breed is traditionally managed almost exclusively for lamb and meat production rather than for dairy purposes. Consequently, the animals were not accustomed to a milking routine, which likely limited their responsiveness. Moreover, the breed has never undergone genetic selection for productive traits such as milk, meat, or wool, which further constrains its potential for high yields. Management conditions also played a key role: it is well established that ewes milked twice daily produce significantly more milk than those milked once (Simitzis et al., 2021). Furthermore, in this study ewes were at different stages of lactation at the beginning of the trial (35 to 71 days post-partum), a factor known to affect yield, which generally decreases as lactation progresses (Ploumi et al., 1998). Finally, lambs were allowed to remain with their dams for part of the day, which not only reduced the amount of milk available for collection but may also have interfered with the milk ejection reflex (Dove, 1988).
The average fat content in milk resulted lower than that observed in most dairy sheep breeds, which typically ranges from 6.6% to 7.5% (Pulina et al., 2005). However, we recorded a maximum value of 9.4%, that is more consistent with literature data reported for Altamurana and Gentile di Puglia sheep milk fat content (Signorelli et al., 2008). Detailed sheep milk composition is difficult to describe whereas the great variability in milk quantity and composition is caused by genetic and physiological factors such as breed, individual characteristics, lactation stage, etc. Further elements of variability are linked to flock management, climate, altitude, the nutritional value of the diet, feed integration (Scintu and Piredda, 2007) (Salfer et al., 2018). Interestingly, from a physiological point of view, Labussiere (1988) classified dairy sheep into two categories: i) those that are easy to milk, which first release the cisternal milk, and then, following the neuroendocrine reflex, the alveolar milk (“double peaks” ewes); ii) and those that are difficult to milk, which release only the cisternal milk fraction (“single peak” ewes), thereby retaining a significant portion of the milk and fat in the alveolar part of the udder (Sagi and Morag, 1974). The same author has shown that ewes lacking the milk ejection reflex can retain up to 75% of the fat in the alveolar milk (Labussière et al., 1969), and this could partially explain the lower fat content of Ciuta milk, compared to other dairy breeds. Other authors have demonstrated that fat concentration of milk of sheep keep in a partial suckling regime was low, due to inhibition of reflex ejection and consequent failure to collect the alveolar milk, which is higher in fat (McKusick et al., 2001). In addition, Rassu et al. (2015) reported that 22 Sarda ewes subjected to a 28-day partial suckling regime with their lambs, yielded an average of 645 g of milk per day, with a fat concentration of 4.57%. This phenomenon is consistent with findings from other studies where milk yield was lower when lambs were allowed to suckle (Margetín et al., 2020).
The protein content in Ciuta milk was lower than the values reported by Benedetti del Rio et al. (2024) for other alpine sheep breeds, with Alpagota and Brogna milk containing 7.11% and 6.65%, respectively. However, similar values to Ciuta’s milk were found in Lamon and Foza breeds, in which protein content was 5.51% and 5.71%, respectively (Bittante et al., 2014). Similarly, 5.45% of protein was reported in milk of Sarda breed (Signorelli et al., 2008). The variation in protein and fat content observed among the Alpine sheep breeds can be attributed to both genetic and nutritional factors. As reported by Pulina et al. (2006), differences in feed quality and composition significantly affect milk chemical composition.
The feeding regimen of Ciuta ewes followed the farm’s standard management practices (hay and concentrate with part-time access to herbaceous pastures) and was not modified for the purposes of this study (Koidou et al., 2019; Wang et al., 2024)An estimate of dry matter intake (DMI) was made to assess whether the diet was adequate to sustain the productive performance of the ewes. On the basis of an average ewes body weight of 36 kg, and considering access to grassland for 7 h/day, the hay offered ad libitum, and 200 g/day of concentrate (see Table 1 for composition), the expected total DMI was calculated. Under full-time grazing sheep typically consume around 2.5-3.0% of body weight in DM per day (NRC, 2007). With restricted grazing time (7 h/day), intake is usually reduced to 60-70% of this potential (Molle et al., 2022), corresponding in this case to approximately 0.55-0.65 kg DM from grass. Additional contributions were provided by hay (0.30-0.50 kg DM/day) and concentrate (0.176 kg DM/day), resulting in a total DMI of about 1.0-1.3 kg DM/day per ewe. Utilizing the empirical equation proposed by Avondo and Lutri (2002), the predicted intake corresponded to 80 g DM/kg metabolic weight. For ewes averaging 36 kg live weight, this corresponds to a total DMI of approximately 1.2 kg DM/day. Nutritionally, this level of intake corresponded to 10–12 MJ of metabolizable energy (ME) and 160–220 g of crude protein. When compared with established requirements, these amounts were sufficient to meet maintenance needs (around 8 MJ ME and 60 g CP/day) and to sustain modest lactation levels of roughly 1.0 kg of milk/day (NRC, 2007). However, for higher milk yields the available energy would likely be limiting, indicating that either an increased level of concentrate supplementation or higher quality forage would be necessary.
This feeding system might have resulted in low intake of readily fermentable carbohydrates and reduced energy density. This condition could also explain the atypical fat-to-protein ratio observed in Ciuta milk, in which protein levels exceed fat content. This is often associated to suboptimal ruminal fermentation or energy deficiency, which may reduce the availability of precursor for milk fat synthesis. Specifically, low fibre digestibility due to mature hay may lead to a decreased production of acetate and butyrate (Cannas, 2004). Moreover, in the absence of dietary lipid supplementation, no increase in propionate production is observed, resulting in lower efficiency in energy availability for milk synthesis (Angeles-Hernandez et al., 2020).
A significant negative relationship between sheep milk protein percentage and milk urea concentration and its related modulation by dietary energy level were discussed by Nudda et al. (2020). Giovanetti et al. (2019) found a marked negative linear relationship between dietary energy content and milk urea level in dairy ewes in mid and late lactation. Contrary to what happens with the dietary energy content, in dairy sheep milk MUC is positively correlated with dietary crude protein (CP) content and, to a lesser extent, with protein intake (Nudda et al., 2020). Milk urea content has been identified as the best predictor of dietary CP content because it allows to estimate how much CP is consumed and how well it is utilized by an animal (Cannas et al., 1998). However, at pasture, the botanical species and the presence of legumes in the herbage, make this prediction more difficult to evidence and this depends mainly on the fact that forage legumes contain tannic phenols, which play a role in modulating faecal N excretion and N use efficiency (Molle et al., 2009).
The percentage of lactose in Ciuta sheep milk was similar with values found in other Italian autochthonous breeds (Bittante et al., 2014; Stocco et al., 2025), with little differences which may be related to breed-specific characteristics (Selvaggi et al., 2017). As reported from Todaro et al. (2023), the low variability of this parameter can be related to the fact that lactose content does not change during the lactation of healthy animals, as it is the main osmotically active component of the milk. Even the casein content (74.8% of total protein) was in line with the casein-to-protein ratio reported for Alpagota and Brogna milk, where casein represented approximately 73.85% and 74.44% of total protein, respectively (Benedetti del Rio et al., 2024). These findings suggest a similar casein profile among these Alpine breeds, despite differences in total protein content.
Overall, Ciuta sheep milk showed a good hygienic-sanitary profile. Somatic cell count (SCC) is widely recognized in the dairy industry as a biomarker of udder’s health and milk hygiene. SCC in Ciuta sheep milk showed average values consistently below the suggested threshold of 250,000 cells/mL) (Albenzio et al., 2019) on almost all days of sampling. However, a maximum of 839,000 cells/mL, was registered in the last sampling. This increase suggests the occurrence of an acute and likely localized event in one or more animals, rather than a general deterioration of milking hygiene or stall conditions. Since all previous samplings showed low SCC and the hygiene milking protocol remained unchanged, this spike is likely attributable to individual animal events such as a transient infection or a physical trauma affecting udder integrity.
The FAs profile in Ciuta milk, mainly composed of C14:0, C16:0, C18:0, C18:1c9, is in line to what reported in literature for sheep milk (Cabiddu et al., 2005).
In a previous study, Pellattiero et al. (2015) reported that milk from Brogna, Foza, and Lamon sheep contained higher concentrations of SFAs, averaging approximately 73.5%, and lower proportions of MUFAs and PUFAs, at 21.9% and 4.8%, respectively, compared to Ciuta milk (61.9%, 26.6%, 6.0%). While such differences may partly reflect genetic variation among breeds (Moioli et al., 2007), they are more likely attributable to differences in diet composition and feeding strategies, particularly in terms of forage proportion and quality (Nudda et al., 2014). In Pellattiero et al. (2015) study, the animals were fed a total mixed ration (TMR), conditions known to increase SFAs and reduce the content of PUFAs in milk. By contrast, Ciuta sheep were managed under an extensive system with minimal concentrate supplementation which probably contributed to increase PUFAs content in milk.
In addition, Ciuta milk showed a lower C18:0 proportion, suggesting a reduced extent of ruminal biohydrogenation which can occur when the diet includes fresh grass rich in PUFAs and secondary plant metabolites, such as tannins and terpenes, which are known to partially inhibit the activity of rumen microbes responsible for the hydrogenation of unsaturated FAs (Poulopoulou et al., 2019; Toral et al., 2018; Vasta et al., 2019). As a result, higher amounts of dietary MUFAs and PUFAs escape microbial saturation in the rumen and are directly transferred into milk fat. Furthermore, the average concentration of c9,t11-18:2 (CLA) in Ciuta milk was consistent with milk produced from lactating ewes under forage and pasture-based feeding systems compared to those kept indoors on conserved forages and concentrates (Atti et al., 2006; Tsiplakou et al., 2006). CLAs are the most investigated FAs in ruminant milk due to their functional role in human health, which include potential anticarcinogenic, antiatherogenic, and immunomodulatory properties (Badawy et al., 2023). The values observed in Ciuta sheep are similar to those reported for other local Alpine breeds with forage-based and pasture-based farming systems (Benedetti del Rio et al., 2024). The increased CLAs content in milk from grazing ewes is related to ruminal biohydrogenation process occurring in the rumen, where C18:2 is partially hydrogenated by rumen bacteria. This process produces intermediates, such as t11-C18:1, which is subsequently desaturated in the mammary gland by the delta-9 desaturase enzyme, leading to the formation of c9,t11-CLA (Chilliard et al., 2007). Pasture-based diets, containing high levels of C18:3, influence this pathway by modulating rumen microbial activity and reducing complete hydrogenation of unsaturated FAs, thereby enhancing c9,t11-CLA synthesis and deposition in milk fat (Vasta et al., 2019).
The nutritional quality of milk fat is influenced by the proportion between n-6 and n-3 PUFAs, expressed as the n-6/n-3 ratio. In Ciuta sheep milk, this ratio showed an average value which can considered low and nutritionally favourable. Indeed, values below 4 are generally associated with anti-inflammatory activity and lower risk of cardiovascular and other chronic diseases (Simopoulos, 2002) (Benedetti del Rio et al., 2024).
The FAs profile of Ciuta sheep also exhibited a relatively high content of other functional FAs. In particular, the average concentration of BCFAs and OCFAs were higher than those typically reported in literature for sheep milk, which are approximately 2.2% and 2%, respectively (Nudda et al., 2021). According to Vlaeminck et al. (2006), higher forage-to-concentrate ratios and grazing-based systems tend to promote ruminal microbial populations that increase the synthesis and accumulation of BCFAs and OCFAs in milk fat. BCFAs are primary synthesized by rumen bacteria and are considered markers of microbial activity, particularly under high-forage diets. OCFAs, by contrast, are made both from microbial lipids activity in the rumen and de novo synthesis in the mammary gland. The feeding system adopted for Ciuta sheep likely promoted ruminal microbial activity and fermentation pathways favourable to the synthesis and accumulation into milk of these functional FAs.
The AI and TI showed average values consistent with those reported for semi-intensive farming systems (Kasapidou et al., 2021) and indicative of a favourable fatty acid profile. Lower AI and TI values are associated with higher levels of unsaturated FAs (MUFAs and PUFAs) and reduced concentrations of SFAs considered pro-atherogenic, such as C12:0, C14:0, and C16:0 (Ulbricht and Southgate, 1991). Similar results were observed by De Renobales et al. (2012), who reported lower AI in milk from grazing ewes compared to TMR-fed animals. In this context, it is worth noting that Ciuta milk contains a variety of bioactive compounds with potential health benefits, including medium- and short-chain FAs, BCFAs, OCFAs, and naturally occurring trans-FAs. In a recent review, these dairy components have been associated with cardiometabolic health and with favourable health outcomes (Mozaffarian and Wu, 2018).
(Mohapatra et al., 2019; Todaro et al., 2023; Pulina et al., 2021) Regarding the colour, the observed L* value was consistent with the typical optical properties of sheep milk (Milovanovic et al., 2021). The colour parameters were generally in line with previously reported characteristics for ovine milk, although with some minor differences. Interestingly, Ciuta sheep milk showed a more marked greenish component compared to what is typically described in literature (Jiménez Sobrino et al., 2018; Milovanovic et al., 2020). This difference can be partly attributed to the composition of pigments in the milk, which are influenced by both fat-soluble and water-soluble compounds. The main pigments of milk are riboflavin, a green compound present in the aqueous phase which is a strong photosensitizer, carotene and, to a lesser extent, lutein. It is well known that fresh grass is rich in natural pigments, such as β-carotene and riboflavin. Although sheep efficiently convert carotenoids into retinol, with limited transfer of β-carotene to milk, riboflavin remains the main pigment present in the aqueous phase of sheep milk (Nozière et al., 2006; Magan et al., 2020) Even the WI and YI observed in this study further highlighted the distinctive optical properties of ovine milk (Raynal-Ljutovac et al., 2008). The pigments contained in sheep milk, in synergy with the specific micellar architecture, contribute to its intense whiteness and subtle green-yellow reflections.
For a satisfactory milkability of ewes and their overall suitability for dairy production it is widely recognized that udder morphology plays a key role. In this study, we investigated the udder conformation of Ciuta ewes in order to tentatively assess whether morphological and physiological traits were present that could support their potential use as dairy sheep. According to many authors (Pourlis, 2020 and references therein), the udder must be well attached, with vertical teats of medium size and high cistern, to allow good milk storage between milkings. The teats should be inserted vertically at the lower part of the cistern, as overly horizontal teats create difficulties during milking, especially when mechanical milking is used. Unfortunately, large cisterns are generally associated with unfavourable position of the teats (Labussiere, 1988).
Overall, the results indicated favourable udder conformation for traits related to suspension and depth, while showed greater variability in teats placement and degree of separation of halves.
The values observed for the degree of suspension suggested that most animals exhibit good udder support, which is desirable for milking and udder health. Similarly, udder depth indicated a generally well-positioned udder, far enough from the ground to reduce contamination risks.
Teat placement showed higher variability, ranging from ideal to less optimal configurations, which could affect milkability, especially in the context of mechanical milking, and would require selective breeding for improvement. The degree of separation had the lowest score and the highest variability, indicating poor strength of suspensor ligament, a trait that could potentially limit milking efficiency. This reflects also a more globular or asymmetrical udder conformation. While the Ciuta breed is not traditionally selected for dairy purposes, these preliminary observations on a limited number of individuals indicated the potential for favourable udder traits that may warrant further investigation. The low DS score may represent a limiting factor, but it should be weighed against the overall potential of this breed.
Finally, regarding the milking behaviour of Ciuta sheep involved in the study, we observed both the lower milking duration and the lower occurrence of discomfort behaviours with a progressive habituation of sheep to the milking routine. In fact, the reduction of these behaviours is likely due to sheep’s accustomedness to human contact and handling, fostered by repeated experiences (Gygax and Vögeli, 2016; Murray et al., 2009) and associated with positive events (Dodd et al., 2012), such as the provision of feed at the entrance to the milking stall (Murray et al., 2009). Although physiological stress markers were not measured, the selected behaviours have been previously associated with reactivity in sheep (Dodd et al., 2012). Moreover, studies conducted on cattle (Hemsworth et al., 1989) and goats (Andrioli et al., 2024) have shown that animals that frequently interact with humans show a significant reduction in distress behaviours, such as kicking. As already mentioned above, the inter-individual variability was statistically significant for all the discomfort behaviours considered, but not for milking duration (Table 5). This suggests that Ciuta sheep respond differently to handling and milking, depending on the individual’s aptitude, a phenomenon already described by Barnard et al. (2016).
Even if the temporal trend was not the same for all the ewes, animals that initially showed more frequent discomfort behaviours continued to be more nervous than others throughout the test period. However, even in these more agitated individuals, a decrease in discomfort behaviours was observed, indicating a certain degree of adaptation to milking (see an example in Figure 3 for ODS). The high level of individual variability suggests that there is room for selecting less reactive individuals, which can therefore adapt better and more quickly to handling and milking operations.
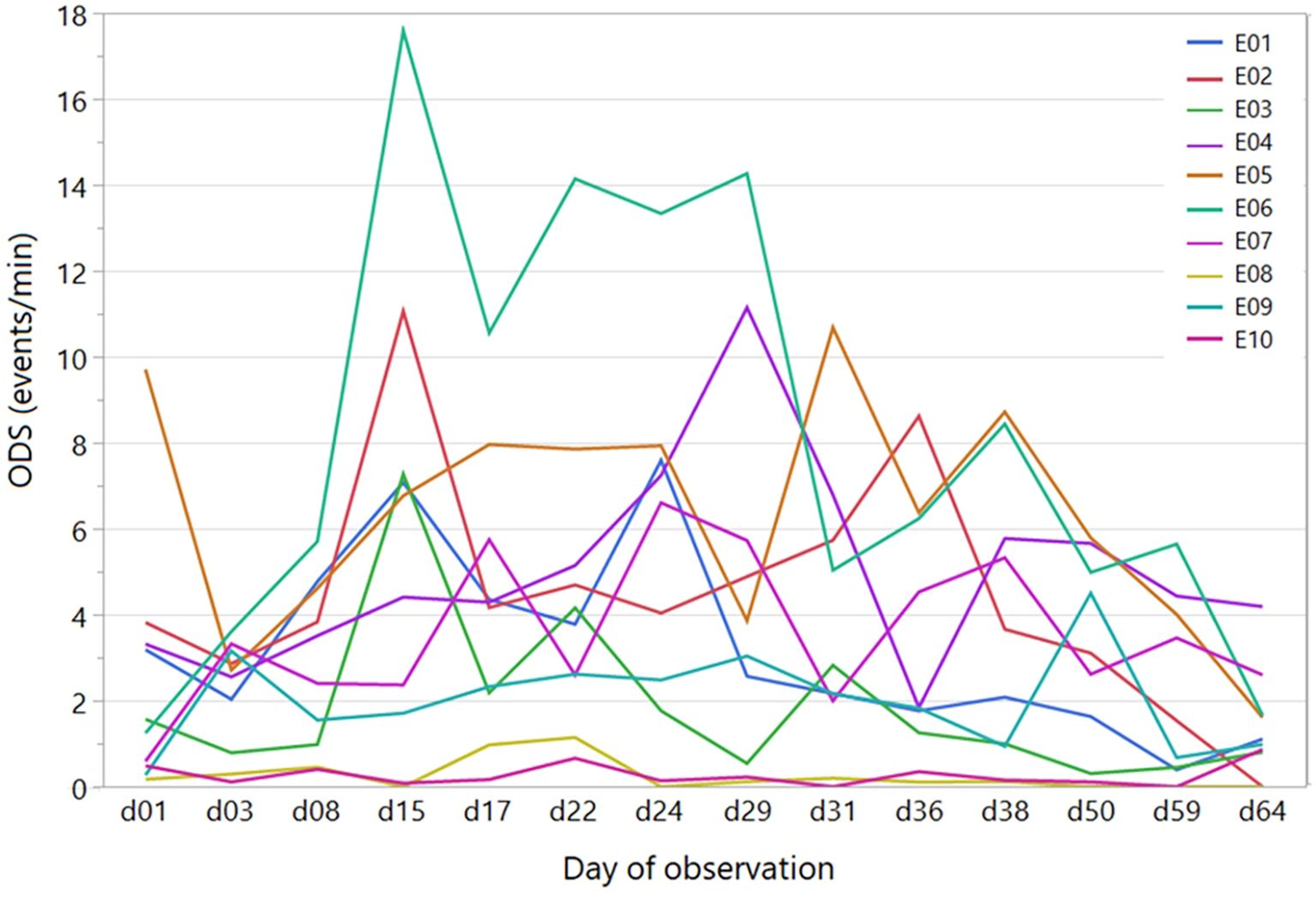
Figure 3. Individual temporal trends of the overall discomfort score (ODS) throughout the whole observation period. Lines of different colours represent different ewes.
5 Conclusions
Results from this study represent a complete novelty for the current conditions of Ciuta sheep farming. The average milk yield was 345 g per ewe and the chemical composition of milk was characterized by 4.6% fat, 5.76% protein, 4.80% lactose and 4.31% casein. Milk hygienic properties were satisfactory, with an average SCC of 100,000 cells/mL. The FAs profile was represented by 61.9% saturated FAs (SFAs), 26.6% monounsaturated FAs (MUFAs), 6.0% polyunsaturated FAs (PUFAs), 2.9% branched-chain FAs (BCFAs), and 2.4% odd-chain FAs (OCFAs). Moreover, the PUFAs fraction showed an average concentration of c9,t11-18:2 (CLA) equal to 1.24 and an average n-6/n-3 ratio of 1.43.
Ciuta milk yield was limited, likely due to the partial suckling of lambs during the trial in the experimental group, as well as the fact that these ewes were milked for the first time and had generally undergone little previous handling, which may have led to a weak ejection reflex. These aspects need further investigations, possibly through a milking trial conducted after lambs weaning, in order to better assess the true productive potential of this breed.
Furthermore, the study highlighted how the Ciuta sheep might be adaptable to milking. The behavioural and production variability observed could represent an opportunity for farmers, who could begin selecting animals based on temperament and production yields, thus improving the efficiency of milking operations and the quantity of milk produced.
In conclusion, the Ciuta sheep represents not only a genetic heritage to be preserved, but also a potential economic resource for mountain communities that choose to focus on the quality and uniqueness of their products.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The animal studies were approved by Animal Welfare Organization of the University of Milan. The studies were conducted in accordance with the local legislation and institutional requirements. Written informed consent was obtained from the owners for the participation of their animals in this study.
Author contributions
MG: Writing – original draft, Data curation, Investigation, Writing – review & editing, Formal Analysis. AL: Conceptualization, Writing – original draft, Investigation, Writing – review & editing, Methodology, Formal Analysis, Data curation. VM: Writing – original draft, Validation, Resources, Data curation, Methodology, Supervision, Conceptualization, Writing – review & editing. SM: Data curation, Writing – original draft, Conceptualization, Resources, Methodology, Validation, Supervision, Writing – review & editing. BM: Formal Analysis, Writing – review & editing, Writing – original draft. SC: Writing – review & editing, Data curation, Writing – original draft. FB: Resources, Project administration, Validation, Conceptualization, Supervision, Writing – review & editing, Writing – original draft, Funding acquisition.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by the following funder: Regione Lombardia, PSR 2014-2020 (FEASR): bando Misura 16, Operazione 16.2.01 “Progetti pilota e sviluppo di innovazione” within the framework of the project “Val3Ciuta – Valorizzazione della triplice attitudine della Pecora Ciuta attraverso la caratterizzazione delle sue produzioni.”
Acknowledgments
The authors sincerely thank the PatriMont – Pro Patrimonio Montano association (https://patrimont.org/en/) and all the participating farmers (Agriturismo La Pedruscia, Agriturismo La Pecora Nera, Azienda Agricola Marco Paganoni), for their generous cooperation in providing data and samples, and for their valuable contribution to this study.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that Generative AI was used in the creation of this manuscript. During the preparation of this work the authors used ChatGpt to improve the English style. After using this tool, the authors reviewed and edited the content as needed and take full responsibility for the content of the published article.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Albenzio M., Figliola L., Caroprese M., Marino R., Sevi A., and Santillo A. (2019). Somatic cell count in sheep milk. Small Ruminant Res. 176, 24–30. doi: 10.1016/j.smallrumres.2019.05.013
Andrioli M., Grajales-Cedeño J. K., Negrão J. A., and Paranhos da Costa M. J. R. (2024). ffects of training of Saanen goats for the first milking on behavior, milk yield, and milk quality traits. Appl. Anim. Behav. Sci. 281, 106444. doi: 10.1016/j.applanim.2024.106444
Angeles-Hernandez J. C., Vieyra Alberto R., Kebreab E., Appuhamy J. A. D. R. N., Dougherty H. C., Castelan-Ortega O., et al. (2020). “Effect of forage to concentrate ratio and fat supplementation on milk composition in dairy sheep: A meta-analysis,” in Livest. Sci., 238, 104069. (Elsevier B.V). doi: 10.1016/j.livsci.2020.104069
Atti N., Rouissi H., and Othmane M. H. (2006). Milk production, milk fatty acid composition and conjugated linoleic acid (CLA) content in dairy ewes raised on feedlot or grazing pasture. Livestock Sci. 104, 121–127. doi: 10.1016/j.livsci.2006.03.014
Avondo M. and Lutri L. (2002). Feed Intake. Chapter 5 in Dairy Sheep Feeding and Nutrition. 105–122. Pulina G. Editor. Avenue media, Bologna (Italy).
Badawy S., Liu Y., Guo M., Liu Z., Xie C., Marawan M. A., et al. (2023). “Conjugated linoleic acid (CLA) as a functional food: Is it beneficial or not?,” in Food Res. Int, 172, 113158. (Elsevier Ltd). doi: 10.1016/j.foodres.2023.113158
Barnard S., Matthews L., Messori S., Podaliri-Vulpiani M., and Ferri N. (2016). Laterality as an indicator of emotional stress in ewes and lambs during a separation test. Anim. Cogn. 19, 207–214. doi: 10.1007/s10071-015-0928-3
Benedetti del Rio E., Berton M., Amalfitano N., Ramanzin M., and Sturaro E. (2024). Added value of products from endangered local sheep breeds in mountain areas. Animals 14, 2855. doi: 10.3390/ani14192855
Bigi D. and Zanon A. (2020). Atlante delle razze autoctone. Bovini, Equini, Ovicaprini, Suini allevati in Italia. 2nd Edition (Milano (Italy): Edagricole - Edizioni Agricole di New Business Media).
Bittante G., Pellattiero E., Malchiodi F., Cipolat-Gotet C., Pazzola M., Vacca G. M., et al. (2014). Quality traits and modeling of coagulation, curd firming, and syneresis of sheep milk of Alpine breeds fed diets supplemented with rumen-protected conjugated fatty acid. J. Dairy Sci. 97, 4018–4028. doi: 10.3168/jds.2013-7345
Bojovic M. and McGregor A. (2023). A review of megatrends in the global dairy sector: what are the socioecological implications? Agric. Hum. Values 40, 373–394. doi: 10.1007/s10460-022-10338-x
Cabiddu A., Decandia M., Addis M., Piredda G., Pirisi A., Molle G., et al. Managing Mediterranean pastures in order to enhance the level of beneficial fatty acids in sheep milk. Small Rumin. Res. 59, 169–180. doi: 10.1016/j.smallrumres.2005.05.005
Cannas A. (2004). “Energy and protein requirements,” in Dairy sheep nutrition. Ed. Pulina G. (Wallingford (UK): CAB International).
Cannas A., Pes A., Mancuso R., Vodret B., and Nudda A. (1998). Effect of dietary energy and protein concentration on the concentration of milk urea nitrogen in dairy ewes. J. Dairy Sci. 81, 499–508. doi: 10.3168/jds.S0022-0302(98)75602-4
Cappio-Borlino A., Portolano B., Todaro M., Macciotta N. P. P., Giaccone P., and Pulina G. (1997). Lactation curves of valle del belice dairy ewes for yields of milk, fat, and protein estimated with test day models. J. Dairy Sci. 80, 3023–3029. doi: 10.3168/jds.S0022-0302(97)76269-6
Casu S., Pernazza I., and Carta A. (2006). Feasibility of a linear scoring method of udder morphology for the selection scheme of Sardinian sheep. J. Dairy Sci. 89, 2200–2209. doi: 10.3168/jds.S0022-0302(06)72290-1
Chilliard Y., Glasser F., Ferlay A., Bernard L., Rouel J., and Doreau M. (2007). Diet, rumen biohydrogenation and nutritional quality of cow and goat milk fat. Eur. J. Lipid Sci. Technol. 109, 828–855). doi: 10.1002/ejlt.200700080
Christie W. W. (2003). “Preparation of derivatives of fatty acids,” in Lipid analysis: isolation, separation and structural analysis of lipids, Bridgewater (England): The Oily Press, 205–225.
Claps S., Mecca M., Di Trana A., and Sepe L. (2020). Local small ruminant grazing in the monti foy area (Italy): the relationship between grassland biodiversity maintenance and added-value dairy products. Front. Veterinary Sci. 7. doi: 10.3389/fvets.2020.546513
De Renobales M., Amores G., Arranz J., Virto M., Barrón L. J. R., Bustamante M. A., et al. (2012). Part-time grazing improves sheep milk production and its nutritional characteristics. Food Chem. 130, 90–96. doi: 10.1016/j.foodchem.2011.07.002
Dodd C. L., Pitchford W. S., Hocking Edwards J. E., and Hazel S. J. (2012). Measures of behavioural reactivity and their relationships with production traits in sheep: A review. Appl. Anim. Behav. Sci. 140, 1–15). doi: 10.1016/j.applanim.2012.03.018
Dove H. (1988). Estimation of the intake of milk by lambs, from the turnover of deuterium- or tritium-labelled water. Br. J. Nutr. 60, 375–387. doi: 10.1079/bjn19880107
EFSA AHAW Panel (2014). Scientific Opinion on the welfare risks related to the farming of sheep for wool, meat and milk production. EFSA J. 12, 3933. doi: 10.2903/j.efsa.2014.3933
FAO (2009). Livestock keepers : guardians of biodiversity (Roma (Italy): Animal Production and Health Paper No. 67). Food & Agriculture Organization of the United Nations).
FAO (2019). The state of the world’s biodiversity for food and agriculture. Available online at: http://www.fao.org/3/CA3129EN/CA3129EN.pdf.
Friard O. and Gamba M. (2016). BORIS: a free, versatile open-source event-logging software for video/audio coding and live observations. Methods Ecol. Evol. 7, 1325–1330. doi: 10.1111/2041-210X.12584
Giovanetti V., Boe F., Decandia M., Bomboi G. C., Atzori A. S., Cannas A., et al. (2019). Milk urea concentration in dairy sheep: Accounting for dietary energy concentration. Animals 9, 1118. doi: 10.3390/ani9121118
Gygax L. and Vögeli S. (2016). Reactions of sheep towards three sets of emotional stimuli: (In)Consistency in respect to stimulus valence and sheep identity. Appl. Anim. Behav. Sci. 174, 51–57. doi: 10.1016/j.applanim.2015.11.015
Hemsworth P. H., Barnett J. L., Tilbrook A. J., and Hansen C. (1989). The effects of handling by humans at calving and during milking on the behaviour and milk cortisol concentrations of primiparous dairy cows. Appl. Anim. Behavour Sci. 22, 313–326. doi: 10.1016/0168-1591(89)90026-9
Jiménez Sobrino L., Poveda Colado J. M., Garzón Sigler A. I., Martínez Marín A. L., Núñez Sánchez N., Asensio J. R., et al. (2018). Composition and colour indices of sheep’s bulk-tank milk are influenced by production practices. Ital. J. Anim. Sci. 17, 477–488. doi: 10.1080/1828051X.2017.1383860
Kasapidou E., Basdagianni Z., Papadopoulos V., Karaiskou C., Kesidis A., and Tsiotsias A. (2021). Effects of intensive and semi-intensive production on sheep milk chemical composition, physicochemical characteristics, fatty acid profile, and nutritional indices. Animals 11, 2578. doi: 10.3390/ani11092578
Kiełczewska K., Kowalik J., Dabrowska˛ A., Jankowska A., and Wachowska M. (2021). The effect of high-pressure treatment and skimming on caprine milk proteins. Appl. Sci. (Switzerland) 11, 5982. doi: 10.3390/app11135982
Koidou M., Mountousis I., Dotas V., Zagorakis K., and Yiakoulaki M. (2019). Temporal variations of herbage production and nutritive value of three grasslands at different elevation zones regarding grazing needs and welfare of ruminants. Arch. Anim. Breed. 62, 215–226. doi: 10.5194/aab-62-215-2019
Labussiere J. (1988). “Review of physiological and anatomical factors influencing the milking ability of ewes and the organization of milking,” in Livestock production science, vol. Vol. 18. .
Labussière J., Combaud J. F., Petrequin P., Tessonnière R., and Gouget R. (1969). Importance, composition et signification des differentes fractions de lait obtenues successivement au cours de la traite mecanique des brebis. Annales Zootechnie 18, 185–196. Lyon (France): HAL Open Science, Available online at: https://hal.science/hal-00886964v1.
Lauvie A., Alexandre G., Angeon V., Couix N., Fontaine O., Gaillard C., et al. (2023). Is the ecosystem services concept relevant to capture the multiple benefits from farming systems using livestock biodiversity? A framework proposal. Genet. Resour. 4, 15–28. doi: 10.46265/genresj.MRBT4299
Liu Z., Wang J., Li C., and Rochfort S. (2020). Development of one-step sample preparation methods for fatty acid profiling of milk fat. Food Chem. 315, 126281. doi: 10.1016/j.foodchem.2020.126281
Lopez A., Celozzi S., Mattiello S., Marcolli B., Greco M., Moretti V. M., et al. (2024). “Wool fiber quality of Pecora Ciuta, a local sheep breed from the Italian Alps,”. ABstract presented at 25 °Convegno Della Società Italiana Di Patologia e Allevamento Degli Ovini e Dei Caprini (SIPAOC).
Madududu P., Jourdain D., Tran D., Degieter M., Karuaihe S., Ntuli H., et al. (2024). Consumers’ willingness-to-pay for dairy and plant-based milk alternatives towards sustainable dairy: A scoping review. Sustain. Production Consumption 51, 261–277). doi: 10.1016/j.spc.2024.09.010
Magan J. B., O’Callaghan T. F., Zheng J., Zhang L., Mandal R., Hennessy D., et al. (2020). Effect of diet on the Vitamin B profile of bovine milk-based protein ingredients. Foods 9, 578. doi: 10.3390/foods9050578
Margetín M., Oravcová M., Margetínová J., Vavrišínová K., and Janíček M. (2020). The influence of lamb rearing system on ewe milk and lamb growth traits in dairy sheep. J. Anim. Feed Sci. 29, 27–34. doi: 10.22358/JAFS/118129/2020
McKusick B. C., Thomas D. L., and Berger Y. M. (2001). Effect of weaning system on commercial milk production and lamb growth of East Friesian dairy sheep. J. Dairy Sci. 84, 1660–1668. doi: 10.3168/jds.S0022-0302(01)74601-2
Milovanovic B., Djekic I., Miocinovic J., Djordjevic V., Lorenzo J. M., Barba F. J., et al. (2020). What is the color of milk and dairy products and how is it measured? Foods 9, 1629. doi: 10.3390/foods9111629
Milovanovic B., Tomovic V., Djekic I., Miocinovic J., Solowiej B. G., Lorenzo J. M., et al. (2021). Colour assessment of milk and milk products using computer vision system and colorimeter. Int. Dairy J. 120, 105084. doi: 10.1016/j.idairyj.2021.105084
Mohapatra A., Shinde A. K., and Singh R. (2019). Sheep milk: A pertinent functional food. Small Ruminant Res. 181, 6–11). doi: 10.1016/j.smallrumres.2019.10.002
Moioli B., D’Andrea M., and Pilla F. (2007). Candidate genes affecting sheep and goat milk quality. Small Ruminant Res. 68, 179–192. doi: 10.1016/j.smallrumres.2006.09.008
Molle G., Giovanetti V., Cabiddu A., Cuccureddu M., Sitzia M., and Decandia M. (2009). “The effect of different grass-legume mixtures on nitrogen excretion of grazing sheep,”. Eds. Papachristou T. G., Parissi Z. M., Salem H.B., and Morand-Fehr P. (Nutritional and foraging ecology of sheep and goats), 55–60.
Molle G., Cannas A., and Gregorini P. (2022). A review on the effects of part-time grazing herbaceous pastures on feeding behaviour and intake of cattle, sheep and horses. Livestock Science, 263, 104982.
Mozaffarian D. and Wu J. H. Y. (2018). Flavonoids, dairy foods, and cardiovascular and metabolic health: A review of emerging biologic pathways. Circ. Res. 122, 369–384). doi: 10.1161/CIRCRESAHA.117.309008
Murray T. L., Blache D. B., and Bencini R. (2009). The selection of dairy sheep on calm temperament before milking and its effect on management and milk production. Small Ruminant Res. 87, 45–49. doi: 10.1016/j.smallrumres.2009.10.005
National Research Council. (2007). Nutrient Requirements of Small Ruminants: sheep, goats, cervids and new world camelids. Washington DC (USA): The National Academies Press.
Nozière P., Graulet B., Lucas A., Martin B., Grolier P., and Doreau M. (2006). Carotenoids for ruminants: From forages to dairy products. Anim. Feed Sci. Technol. 131, 418–450). doi: 10.1016/j.anifeedsci.2006.06.018
Nudda A., Atzori A. S., Correddu F., Battacone G., Lunesu M. F., Cannas A., et al. (2020). Effects of nutrition on main components of sheep milk. Small Ruminant Res. 184, 106015. doi: 10.1016/j.smallrumres.2019.11.001
Nudda A., Battacone G., Neto O. B., Cannas A., Francesconi A. H. D., Atzori A. S., et al. (2014). Feeding strategies to design the fatty acid profile of sheep milk and cheese. In Rev. Bras. Zootecnia 43, 445–456). doi: 10.1590/S1516-35982014000800008
Nudda A., Correddu F., Cesarani A., Pulina G., and Battacone G. (2021). Functional odd- and branched-chain fatty acid in sheep and goat milk and cheeses. Dairy 2, 79–89. doi: 10.3390/dairy2010008
Pellattiero E., CecChinato A., Tagliapietra F., Schiavon S., and Bittante G. (2015). The use of 2-dimensional gas chromatography to investigate the effect of rumen-protected conjugated linoleic acid, breed, and lactation stage on the fatty acid profile of sheep milk. J. Dairy Sci. 98, 2088–2102. doi: 10.3168/jds.2014-8395
Ploumi K., Belibasaki S., and Triantaphyllidis G. (1998). “Some factors affecting daily milk yield and composition in a flock of Chios ewes,” in Small ruminant research, 28, 89–92.
Poulopoulou I., Evangelos Z., Styliani A., Theofilaktos M., and Ioannis H. (2019). Effects of terpenes administration on fatty acid profile and coagulation properties of ewes’ milk. Emirates J. Food Agric. 31, 980–985. doi: 10.9755/ejfa.2019.v31.i12.2048
Pourlis A. (2020). Ovine mammary morphology and associations with milk production, milkability and animal selection. Small Ruminant Res. 184, 106009. doi: 10.1016/j.smallrumres.2019.10.010
Pulina G., Atzori A. S., Dimauro C., Ibba I., Gaias G. F., Correddu F., et al. (2021). The milk fingerprint of Sardinian dairy sheep: quality and yield of milk used for Pecorino Romano P.D.O. cheese production on population-based 5-year survey. Ital. J. Anim. Sci. 20, 171–180. doi: 10.1080/1828051X.2021.1875896
Pulina G., Macciotta N., and Nudda A. (2005). Milk composition and feeding in the Italian dairy sheep. Ital. J. Anim. Sci. 4, 5–14). doi: 10.4081/ijas.2005.1s.5
Pulina G., Nudda A., Battacone G., and Cannas A. (2006). Effects of nutrition on the contents of fat, protein, somatic cells, aromatic compounds, and undesirable substances in sheep milk. Anim. Feed Sci. Technol. 131, 255–291. doi: 10.1016/j.anifeedsci.2006.05.023
Rassu S. P. G., Nudda A., Carzedda C., Battacone G., Bencini R., and Pulina G. (2015). A partial suckling regime increases milk production in Sarda dairy sheep without affecting meat quality of lambs. Small Ruminant Res. 125, 15–20. doi: 10.1016/j.smallrumres.2015.02.015
Raynal-Ljutovac K., Lagriffoul G., Paccard P., Guillet I., and Chilliard Y. (2008). Composition of goat and sheep milk products: An update. Small Ruminant Res. 79, 57–72. doi: 10.1016/j.smallrumres.2008.07.009
Reefmann N., Bütikofer Kaszàs F., Wechsler B., and Gygax L. (2009). Ear and tail postures as indicators of emotional valence in sheep. Appl. Anim. Behav. Sci. 118, 199–207. doi: 10.1016/j.applanim.2009.02.013
Rico D. E., Marshall E. R., Choi J., Kaylegian K. E., Dechow C. D., Harvatine K. J., et al (2014). Within-milking variation in milk composition and fatty acid profile of Holstein dairy cows. J. Dairy Sci. 97, 4259–4268. doi: 10.3168/jds.2013-7731
Sagi R. and Morag M. (1974). Udder conformation, milk yield and milk fractionation in the dairy ewe. Annales Zootechnie 23, 185–192. doi: 10.1051/animres:19740207
Salfer I. J., Morelli M. C., Ying Y., Allen M. S., and Harvatine K. J. (2018). The effects of source and concentration of dietary fiber, starch, and fatty acids on the daily patterns of feed intake, rumination, and rumen pH in dairy cows. J. Dairy Sci. 101, 10911–10921. doi: 10.3168/jds.2018-15071
Santillo A., della Malva A., and Albenzio M. (2025). Preserving biodiversity of sheep and goat farming in the apulia region,” in Animals, 15, 1610. (Multidisciplinary Digital Publishing Institute (MDPI). doi: 10.3390/ani15111610
Scintu M. F. and Piredda G. (2007). Typicity and biodiversity of goat and sheep milk products. Small Ruminant Res. 68, 221–231. doi: 10.1016/j.smallrumres.2006.09.005
Selvaggi M., D’Alessandro A. G., and Dario C. (2017). Environmental and genetic factors affecting milk yield and quality in three Italian sheep breeds. J. Dairy Res. 84, 27–31. doi: 10.1017/S0022029916000765
Signorelli F., Contarini G., Annicchiarico G., Napolitano F., Orrù L., Catillo G., et al. (2008). Breed differences in sheep milk fatty acid profiles: Opportunities for sustainable use of animal genetic resources. Small Ruminant Res. 78, 24–31. doi: 10.1016/j.smallrumres.2008.04.003
Simitzis P., Seferlis I., Goliomytis M., Bizelis I., and Politis I. (2021). Handling and milking frequency affect milk yield and behavior in dairy ewes. Small Ruminant Res. 198, N106351. doi: 10.1016/j.smallrumres.2021.106351
Simopoulos A. P. (2002). The importance of the ratio of omega-6/omega-3 essential fatty acids. BioMed. Pharmacother. 56, 365–379. Available online at: www.elsevier.com/locate/biopha.
Skordos D., Ragkos A., Karanikolas P., and Vlahos G. (2024). How valuable are the ecosystem services of local animal breeds? The case of Kalaritiko sheep in Greece. Livestock Sci. 279, 105391. doi: 10.1016/j.livsci.2023.105391
Stocco G., Cipolat-Gotet C., Summer A., Tiezzi F., Blotta S., Negro A., et al. (2025). Modeling the relationships among technological properties of sheep individual animal factors, milk composition, and minerals using generalized additive mixed models. J. Dairy Sci. 108, 3334–3353. doi: 10.3168/jds.2024-25846
Sturaro E., Marchiori E., Cocca G., Penasa M., Ramanzin M., and Bittante G. (2013). Dairy systems in mountainous areas: Farm animal biodiversity, milk production and destination, and land use. Livestock Sci. 158, 157–168. doi: 10.1016/j.livsci.2013.09.011
Todaro M., Gannuscio R., Mancuso I., Ducato B., and Scatassa M. L. (2023). Relationships between chemical and physical parameters of bulk milk from Valle del Belice sheep. Ital. J. Anim. Sci. 22, 953–958. doi: 10.1080/1828051X.2023.2252846
Toral P. G., Monahan F. J., Hervas G., Frutos P., and Moloney A. P. (2018). “Review: Modulating ruminal lipid metabolism to improve the fatty acid composition of meat and milk. challenges and opportunities,” in Animal, vol. 12. (Cambridge University Press), 12, s272–s281. doi: 10.1017/S1751731118001994
Tsiplakou E., Mountzouris K. C., and Zervas G. (2006). Concentration of conjugated linoleic acid in grazing sheep and goat milk fat. Livestock Sci. 103, 74–84. doi: 10.1016/j.livsci.2006.01.010
Tufarelli V., Dario M., and Laudadio V. (2009). Milk yield and composition of lactating Comisana ewes fed total mixed rations containing nitrogen sources with different ruminal degradability. Livestock Sci. 122, 349–353. doi: 10.1016/j.livsci.2008.08.012
Turri F. (2025). Atlante delle risorse genetiche animali lombardi (Milano (Italy): RTI EY-Carmi e Ubertis).
Ulbricht T. L. V. and Southgate D. A. T. (1991). Coronary heart disease: seven dietary factors. The Lancet. 338, 985–992. doi: 10.1016/0140-6736(91)91846-M
Vasta V., Daghio M., Cappucci A., Buccioni A., Serra A., Viti C., et al. (2019). Invited review: Plant polyphenols and rumen microbiota responsible for fatty acid biohydrogenation, fiber digestion, and methane emission: Experimental evidence and methodological approaches. J. Dairy Sci. 102, 3781–3804. doi: 10.3168/jds.2018-14985
Vlaeminck B., Fievez V., Cabrita A. R. J., Fonseca A. J. M., and Dewhurst R. J. (2006). Factors affecting odd- and branched-chain fatty acids in milk: A review. Anim. Feed Sci. Technol. 131, 389–417). doi: 10.1016/j.anifeedsci.2006.06.017
Keywords: Ciuta sheep, alpine farming, local breeds, milk quality, behaviour
Citation: Greco M, Lopez A, Moretti VM, Mattiello S, Marcolli B, Celozzi S and Bellagamba F (2025) Preliminary study on the milk producing ability of Ciuta sheep. Front. Anim. Sci. 6:1698620. doi: 10.3389/fanim.2025.1698620
Received: 03 September 2025; Accepted: 10 October 2025;
Published: 24 October 2025.
Edited by:
Bianca Castiglioni, National Research Council (CNR), ItalyReviewed by:
Paola Cremonesi, Research Council (CNR), ItalyFiorenzo Piccioli-Cappelli, Catholic University of the Sacred Heart, Italy
Copyright © 2025 Greco, Lopez, Moretti, Mattiello, Marcolli, Celozzi and Bellagamba. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Annalaura Lopez, YW5uYWxhdXJhLmxvcGV6QHVuaW1pLml0