 Claudia Veira1,2†Belén Dávila1,2†Elva Serna3†
Claudia Veira1,2†Belén Dávila1,2†Elva Serna3† Nilda Portillo3†
Nilda Portillo3† Gloria Yaluff3†
Gloria Yaluff3† Guzmán Álvarez2,4†
Guzmán Álvarez2,4† Hugo Cerecetto1,2,5†
Hugo Cerecetto1,2,5† Elena Aguilera1,2*†
Elena Aguilera1,2*†- 1Grupo de Química Orgánica Medicinal, Instituto de Química Biológica, Facultad de Ciencias, Universidad de la República, Montevideo, Uruguay
- 2Instituto de Investigación Una Salud, Universidad de la República, Montevideo, Uruguay
- 3Departamento de Medicina Tropical, Instituto de Investigaciones en Ciencias de la Salud, Universidad Nacional de Asunción, San Lorenzo, Paraguay
- 4Laboratorio de Moléculas Bioactivas, Centro Universitario Litoral Norte, Universidad de la República, Paysandú, Uruguay
- 5Área de Radiofarmacia, Centro de Investigaciones Nucleares, Facultad de Ciencias, Universidad de la República, Montevideo, Uruguay
Building upon our previous work on drug repurposing and the use of binary combinations as anti-Chagas agents, we detail our efforts to advance the most promising repositioned compounds and binary mixtures for in vitro usage beyond the hit-to-preclinical candidate stage. Accordingly, five repositioned active principles and four binary combinations of these agents that were selected for their in vitro isobolographic behaviors were evaluated in a murine model of acute Chagas disease. In the study, these nine systems were administered orally along with the negative and positive controls (benznidazole) for 15 d. The changes in the parasitemia levels and animal survival over time were evaluated together with the anti-Trypanosoma cruzi antibody levels at the end of the trials. Gabapentin was found to be the most notable repositioned drug as it decreased parasitemia and anti-T. cruzi antibody levels significantly while improving animal survival modestly. The best animal survival profile was achieved when gabapentin was combined with naftazone. Furthermore, the combination of naftazone and red clover extract, which is trademarked in Uruguay as Climodin, was another noteworthy binary mixture; this combination exhibited a consistent parasitemia reduction profile along with the best antibody reduction; however, it did not yield the best animal survival profile, which is in line with our previous in vitro findings. To advance preclinical studies, we further studied the mutagenicity of both binary combinations with and without metabolic activation, which showed that there were no observable mutagenic effects.
1 Introduction
Chagas disease (CD) is a neglected tropical disease that is endemic to Latin American countries with tropical and subtropical weather conditions (Pan American Health Organization, 2025); however, migratory movements to non-endemic areas have led to its global spread (Navarro et al., 2022). CD is estimated to have affected approximately 7.5 million people globally, with more than 100 million people at risk of infection, 30% of patients prone to suffering from Chagasic heart disease, and up to 10% of patients being affected by neurological, digestive, or mixed disorders (Pan American Health Organization, 2025). This illness is transmitted through various mechanisms, including the feces/urine of infected triatomine bugs, blood-sucking insects that feed on humans and animals, via transplacental processes, blood transfusion or organ transplant, and contaminated food and drinks (Pan American Health Organization, 2025). CD is caused by the protist parasite Trypanosoma cruzi (Chagas, 1909) (Kinetoplastida, Trypanosomatidae), whose lifecycle includes an insect vector and a vertebrate host at different stages, namely, the proliferative and presumably non-infective epimastigotes in the insect as well as the infective metacyclic trypomastigotes discharged in the feces/urine during the bloodmeal of the vector invading the host macrophages, fibroblasts, and muscle cells, where they transform into proliferative amastigotes (Pan American Health Organization, 2025). The available drugs against CD are two old nitroaromatics, namely, benznidazole (Bnz) and nifurtimox (Table 1), which have low efficacies during the chronic phase and severe side effects, including mutagenicity. Despite significant efforts over the years to develop new drugs for Chagas therapy since the introduction of Bnz and nifurtimox in the 1960s and 1970s, there have been no new therapeutic licensed medicines for CD or drug developments in phase III clinical trials (Cerecetto and González, 2010; Chatelain and Konar, 2015; Kratz, 2019). Consequently, research efforts must be intensified to discover new therapeutic strategies against CD.
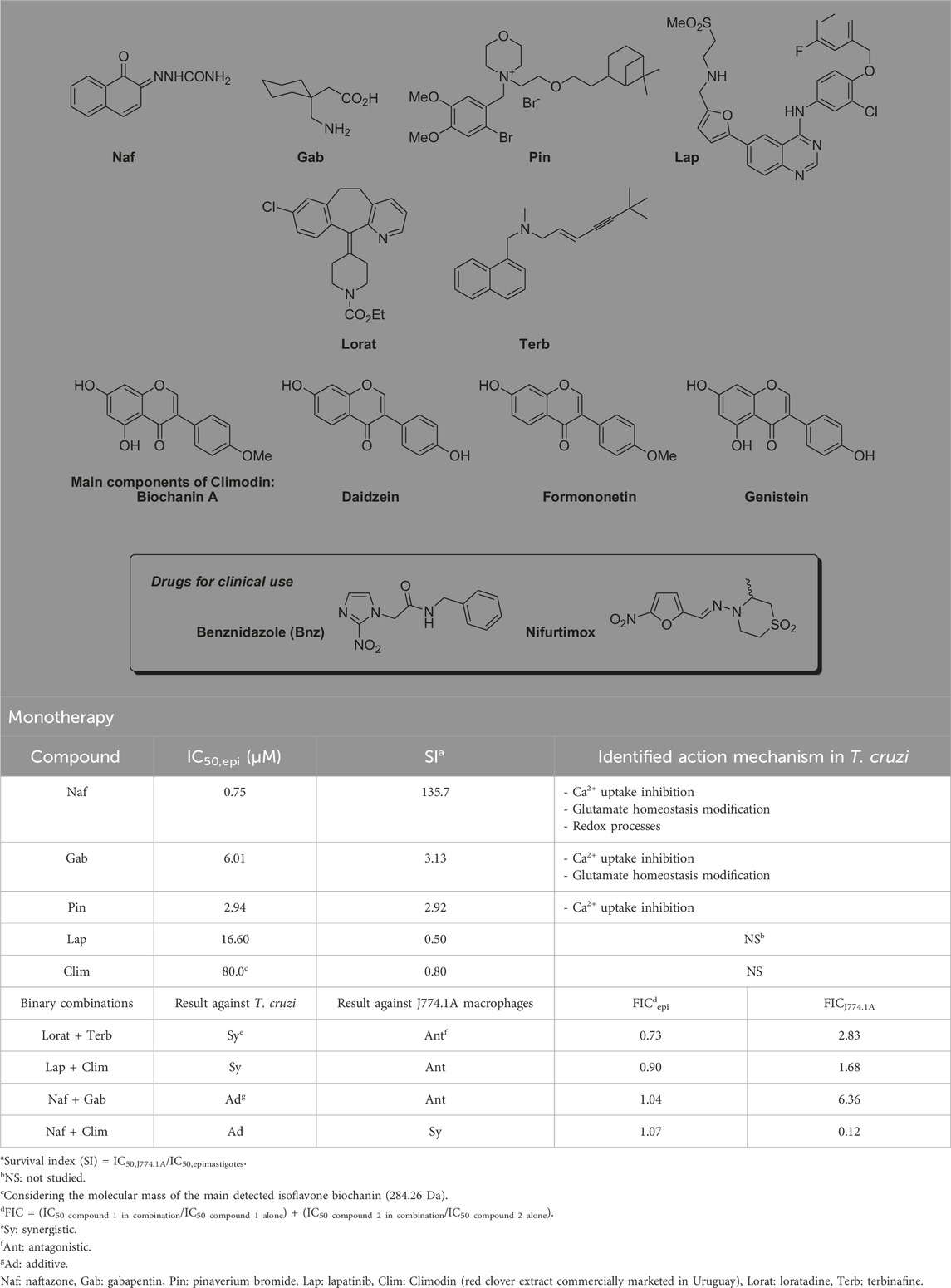
Table 1. Previously reported in vitro results of repurposed drugs and their binary combinations as therapeutic tools against Chagas disease (CD) (Veira et al., 2025).
Recently, our research group has been working on drug repurposing and binary combinations of medications as therapeutic tools in CD (Veira et al., 2025). In our past work, we identified new candidates among the already approved drugs for repurposing as well as other drugs described in the context of different models and T. cruzi strains that have in vitro anti-T. cruzi activities against all three forms of the parasite. As reported in our previous work (Veira et al., 2025), we confirmed that some of the drugs like naftazone (Naf), gabapentin (Gab), and pinaverium bromide (Pin) had similar action mechanisms in the protozoan as those described in mammals (Table 1). Furthermore, certain binary combinations of these drugs were found to have additive or synergistic qualities in their in vitro cytotoxic anti-T. cruzi activities, along with specific toxicity profiles against macrophages (as a mammalian model system). In terms of monotherapy, the drugs Naf, Gab, Pin, and lapatinib (Lap) as well as a red clover extract (Trifolium pratense extract) trademarked in Uruguay as Climodin (Clim; comprising the isoflavones biochanin A, formononetin, genistein, and daidzein) have been reported to display desirable biological behaviors (Table 1). For Naf, Gab, and Pin, the action mechanisms in mammals are also applicable to epimastigotes of T. cruzi (Table 1). Among the binary combinations studied, the best results in terms of synergistic effects against T. cruzi and antagonistic effects against macrophages were observed for loratadine (Lorat) combined with terbinafine (Terb) and for Lap with Clim (Lorat + Terb and Lap + Clim, respectively; Table 1). Additionally, the combination of Naf and Gab (Naf + Gab) exhibited additive effects on T. cruzi cytotoxicity along with the desired antagonism against macrophages. Another interesting binary mixture against T. cruzi was Naf and Clim (Naf + Clim) that was observed to promote additive effects; however, this mixture showed synergistic effects against J774.1A macrophages (Table 1) (Veira et al., 2025).
In the present study, we investigated the nine systems (five monotherapy and four combinations) shown in Table 1 in a murine model of acute CD; here, we analyzed the parasitemia and animal survival during treatment as well as the levels of anti-T. cruzi antibodies at the end of the assays. Additionally, we report the mutagenicities of the two best binary combinations with and without metabolic activation as a means of advancing preclinical studies.
2 Materials and methods
2.1 Animals
The animal protocols involved in this study were approved by the Paraguayan Ethics Committee for Animal Experimentation (no. IORG0010088). Accordingly, 3-month-old male Balb/c mice were infected with 1.0 × 104 T. cruzi CL Brener clone trypomastigotes on day 0 via intraperitoneal injection (DTU: TcVI, discrete typification unit prevailing in the Southern Cone countries; Romer et al. (2023)); these were obtained from donor mice at the onset of peak parasitemia (exceeding 1.0 × 106 parasites/mL). The parasitemia was monitored daily using optical microscopy from day 4 post-infection until all the mice in the group had tested positive for the infection. The treatment was initiated once parasitemia was confirmed in all animals.
2.2 Treatment in the murine model of CD
In brief, the treatment compounds were administered orally to unanesthetized animals via intragastric cannula once daily for 15 consecutive days. The parasitemia was then monitored weekly using a micromethod (Section 2.2.2). At the end of the treatment, approximately 200 µL of blood was collected from the tail vein of each unanesthetized mouse for serological analysis using ELISA to detect the IgG anti-T. cruzi antibodies (Test IICS Método ELISA abbreviated as IICS, Instituto de Investigación en Ciencias de la Salud, Asuncion, Paraguay), which uses antigens from the epimastigote lysate of the T. cruzi Y strain (DTU: TcII)) (Aria et al., 2016). On day 64 that marked the conclusion of the experiment, the animals were humanely euthanized via cervical dislocation performed by trained personnel in accordance with the ethical guidelines and approved protocols.
2.2.1 In vivo evaluation
The therapeutic efficacy of each treatment drug or combination against T. cruzi infection was evaluated using a group of six male Balb/c mice at doses expressed in terms of per kilogram of bodyweight. The selected doses for these in vivo evaluations were established by extrapolation of the effective concentrations observed in in vitro assays against T. cruzi to the murine model; this process also accounted for the pharmacological relevance as well as practical constraints of compound availability and formulation under academic laboratory conditions. As a reference, we considered the standard therapeutic dose of Bnz, which is administered at 50 mg/kg of bodyweight per animal per day corresponding to 192 μmol/kg (Aguilera et al., 2018). The different treatments were administered orally once daily starting at the acute phase of infection (Table 2). Wherever feasible, the test compounds were adjusted to match the standard molar equivalent dosage (192 μmol/kg) to facilitate comparison of trypanocidal efficacy. In cases where the compound solubility, formulation limitations, or toxicity profiles precluded exact equivalence, the closest practical dose was selected while maintaining biological relevance. For binary combinations, each drug component was dosed at either the molar equivalent (192 μmol/kg) or the highest achievable concentration. The trademarked drug Clim lacks direct in vivo data in this context and was thus administered at 50 mg/kg (175.9 μmol/kg) upon considering the molecular mass of the main detected isoflavone biochanin (284.26 Da).
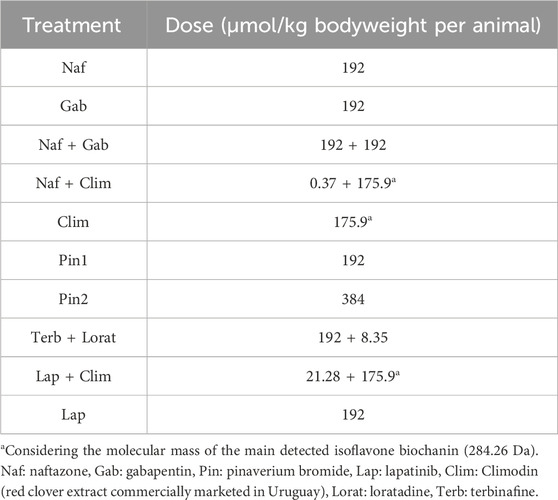
Table 2. Treatment groups and doses of the compounds administered to the acute murine model of Chagas disease.
2.2.2 Hemoconcentration and parasite quantification (micromethod)
Blood samples were collected from all unanesthetized animals by tail puncture and placed in calibrated capillary tubes to obtain a filling of 8–18 mm of blood per mouse. The volumes were then converted to microliters using a previously established laboratory table. The capillary tubes with the samples were centrifuged at 3,000×g for 40 s. Next, parasitemia was assessed under optical microscopy by examining the capillary tube contents, where the red blood cells (RBCs) settled at one end, serum remained at the opposite end, and trypomastigotes accumulated at the interface. The capillary tubes were then cut a few millimeters below the interface (toward the RBC portion), and their contents were spread over microscope slides and covered with a coverslip. The trypomastigotes were counted in 50 fields at ×40 magnification, and the total count was divided by 50 to calculate the average number of parasites per field, which represents the average number of trypanosomes per 5 mm3. These values were compared with those of untreated control animals. The mean and standard deviation values were calculated using OriginPro 9 and GraphPad Prism 5. Finally, the parasite suppression was statistically analyzed using one-way ANOVA (non-parametric).
2.3 Mutagenicity assay (Ames test)
The mutagenic potential of each compound was assessed using the Ames test. Cultures of Salmonella typhimurium strain TA98 (hisD3052, histidine-dependent) were grown to the exponential phase in Oxoid No. 2 medium up to a concentration of approximately 2 × 109 CFU/mL (Organisation for Economic Co-operation and Development, 2002). All procedures, including preparation of the bacterial suspensions, handling of the test compounds, S9 mixtures, and initial assay steps (up to incubation), were performed under sterile conditions using a laminar flow hood and sterilized materials (autoclaved at 121 °C for 20 min or used directly out of sterile manufacturer packaging). The addition of the histidine/biotin solution and plate seeding were performed under an upward sterile air current generated by a Bunsen burner flame. Five serial dilutions of dimethyl sulfoxide (DMSO) were then prepared, starting from the highest non-toxic concentration determined previously in a preliminary toxicity assay up to the third dilution. The positive control was established using 4-nitro-o-phenylenediamine (20 µg/plate) in the absence of metabolic activation; the negative controls included DMSO (50 µL/plate) and a phenotype control for assessing resistances to ampicillin, tetracycline, and crystal violet as well as sensitivity to ultraviolet light. Each binary combination was incubated for 1 h at 37 °C in 2 mL of agar containing 200 µL of a histidine/biotin (0.5 mM/0.5 mM) solution. After incubation, each mixture was plated on glucose-minimal agar and incubated at 37 °C for 48 h. To evaluate the effects of metabolic activation, the assays were repeated in the presence of the S9 fraction (Gibco™, Thermo Fisher Scientific, Grand Island, NY, United States). Here, the positive control was established using 2-aminofluorene (10 µg/plate) in the presence of metabolic activation. The colony counts were performed, and results were reported as mean ± standard deviation from duplicate plate readings. A compound was considered to be mutagenic when the number of revertant colonies was at least twice that of the negative control in two or more consecutive concentrations (Dávila et al., 2019).
3 Results
3.1 Monotherapy studies: in vivo results
Although molecular methods are often prioritized for determining parasitemia (Aguilera et al., 2018; 2022), microscopy has become a robust and sensitive method that provides very good final results. The oral treatment with Naf resulted in a clear decrease in parasitemia compared to phosphate-buffered saline (PBS) treatment during the assay period (Figure 1), leading to a slight shift in the timing of the first peak of parasitemia by 1 week. Similarly, Gab shifted the first peak of parasitemia by 1 week; however, in agreement with the in vitro IC50 value, there was a lower decrease in the parasite load at this time. Additionally, Gab shifted the second peak of parasitemia by 1 week. Contrary to the in vitro results, Clim almost completely abolished both peaks of parasitemia, and the parasite load values were not significantly different from those of Naf. The survival of mice for these three compounds wholly followed the survival index (SI) values determined in vitro (Table 1), with Naf promoting the greatest survival rate (first death at fourth week and 80% animal survival at the end of treatment), followed by Gab (first death at third week and 80% animal survival at the end of treatment) and finally Clim (first death at third week and 60% animal survival at the end of treatment). Regarding the levels of IgG anti-T. cruzi antibodies at the end of the assays (Table 3), Gab demonstrated the best behavior with values significantly lower than those of the PBS-treated animals. Furthermore, Naf displayed the worst antibody levels, in agreement with the finding that this group had the most significant level of parasitemia along with the PBS treatment group among all the treatment groups (Figure 1A).
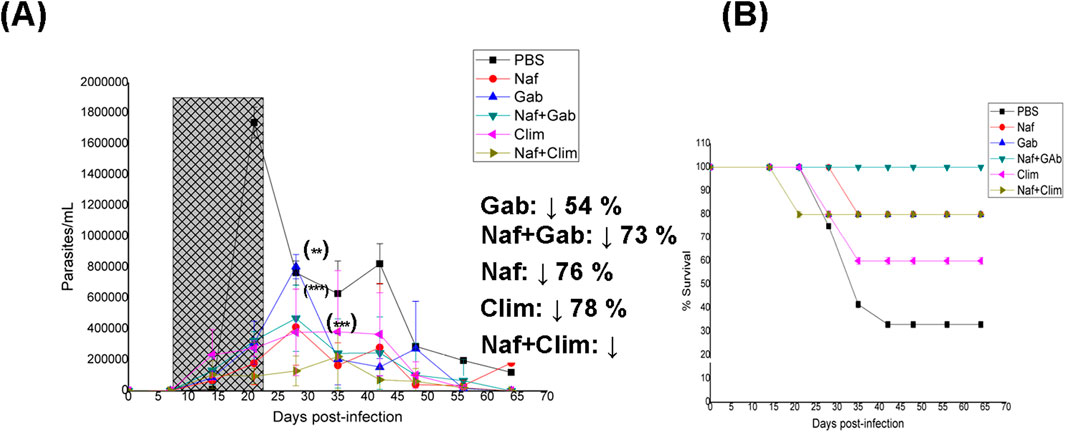
Figure 1. In vivo results for naftazone (Naf), gabapentin (Gab), and Climodin (Clim) as monotherapy and for the binary mixtures Naf + Gab and Naf + Clim. (A) Parasitemia vs. number of days post-infection; the period of oral treatment is marked in gray. Inset: Percentage decrease in parasitic load with respect to phosphate-buffered saline (PBS) treatment at the maximum parasitemia peak. (B) Animal survival vs. week of the assay. (**) p < 0.01; (***) p < 0.001 (with respect to the maximum parasitemia peak for PBS treatment).
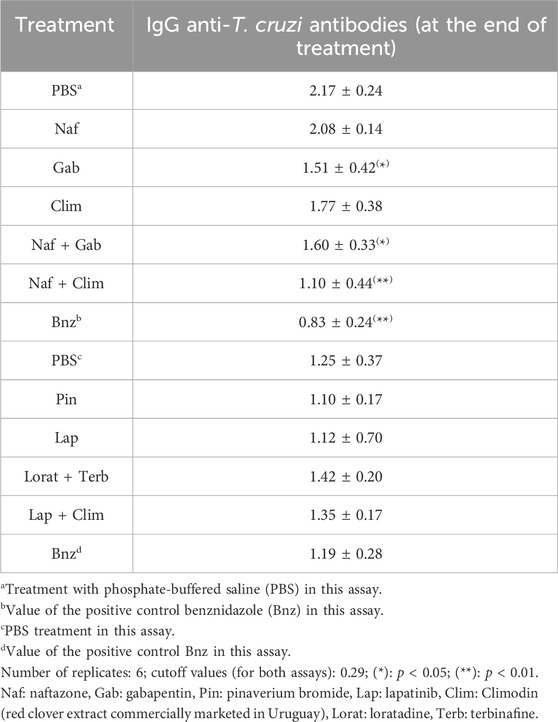
Table 3. Anti-T. cruzi antibody levels at the end of different treatments.
In the case of Pin, two different dosages were assayed owing to its better results compared to the PBS-treated animal counterparts, particularly in terms of the survival rate at the low dose of 192 μmol/kg bodyweight (Pin1, Figure 2A,B). At the high dose of 384 μmol/kg bodyweight (Pin2, Figure 2C), the parasitemia diminished significantly, abolishing almost both peaks of parasitemia along with parasite load values that were significantly lower than those of the PBS-treated animals. However, contrary to the in vitro results (Table 1) (Veira et al., 2025), the toxicity increased compared to the PBS-treated animals, resulting in a 17% decrease in animal survival (Figure 2D). Additionally, at the high dose, Pin did not significantly diminish the levels of IgG anti-T. cruzi antibodies at the end of the assay (Table 3). Unfortunately, none of the binary combinations with Pin yielded in vitro additivity or synergism against T. cruzi in our previous studies (Veira et al., 2025). Another repurposed drug that was studied in vivo as a monotherapy was Lap (Patel et al., 2013); the parasitemia and survival behaviors of Lap were very good (Figures 3A,B), almost abolishing both the maximum parasite peaks of the PBS-treated animals and reducing the number of deaths. On the other hand, the IgG anti-T. cruzi antibody levels were not improved by Lap at the end of treatment (Table 3).
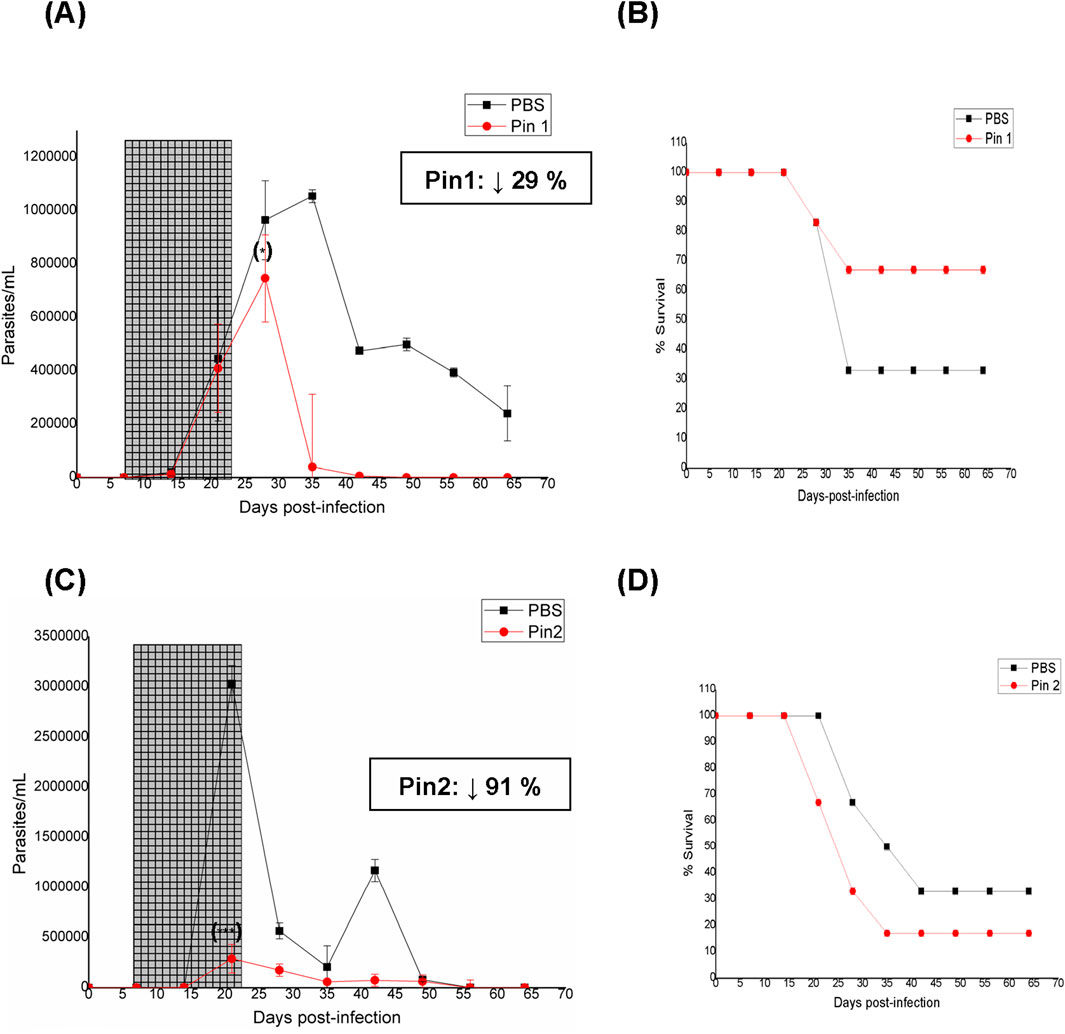
Figure 2. In vivo results for pinaverium bromide (Pin) as monotherapy. Dose of treatment: 192 μmol/kg animal bodyweight (Pin1). (A) Parasitemia vs. week of the assay. The period of oral treatment is marked in gray. Inset: Percentage decrease in parasitic load with respect to PBS treatment at the maximum parasitemia peak. (B) Animal survival vs. week of the assay. Dose of treatment: 384 μmol/kg animal bodyweight (Pin2). (C) Parasitemia vs. week of the assay. The period of oral treatment is marked in gray. Inset: Percentage decrease in parasitic load with respect to PBS treatment at the maximum parasitemia peak. (D) Animal survival vs. week of the assay. (*) p < 0.05; (***) p < 0.001 (with respect to the maximum parasitemia peak for PBS treatment).
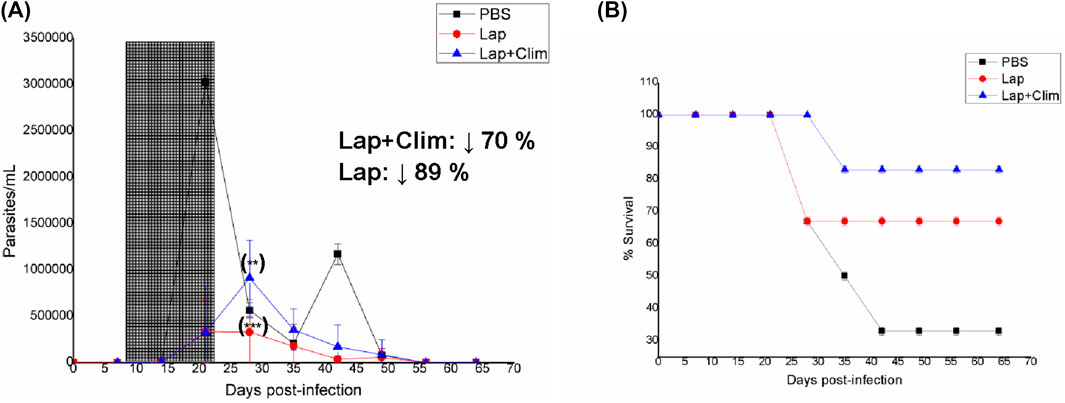
Figure 3. In vivo results for lapatinib (Lap) as monotherapy and for the binary mixture Lap + Clim. (A) Parasitemia vs. week of the assay; the period of oral treatment is marked in gray. Inset: Percentage decrease in parasitic load with respect to PBS treatment at the maximum parasitemia peak. (B) Animal survival vs. week of the assay. (**) p < 0.01; (***) p < 0.001 (with respect to the maximum parasitemia peak for PBS treatment).
3.2 Binary mixture studies: in vivo results
For the in vivo studies of the binary mixtures, we selected the three best combinations based on the in vitro results (Veira et al., 2025), namely, Lorat + Terb, Lap + Clim, and Naf + Gab, all of which are antagonists against the macrophage model (Table 1). Additionally, we included the Naf + Clim combination, which exhibited an undesirable profile against J774.1A cells and synergism in toxicity (Table 1).
The Lorat + Terb combination had a good FICJ774.1A in vitro and was studied orally, as indicated previously (Figure 4). The in vitro and in vivo studies of these two compounds, both as monotherapy and in combination with other entities, have been reported extensively by authors (Maldonado et al., 1993; Planer et al., 2014; Sayé et al., 2020); however, there are no reports on the combination of both drugs. The combination promoted a slight time shift of the first peak of parasitemia by 1 week, along with excellent decrease in the parasite load (75%) and abolished second peak. The animal survival rate also improved compared to the PBS-treated animals, as expected from the in vitro FICJ774.1A (Table 1) (first death at fifth week and 83% animal survival at the end of treatment). Unfortunately, the IgG anti-T. cruzi antibody levels were not good, and the Lorat + Terb treatment failed to reduce these values compared to PBS treatment (Table 3).
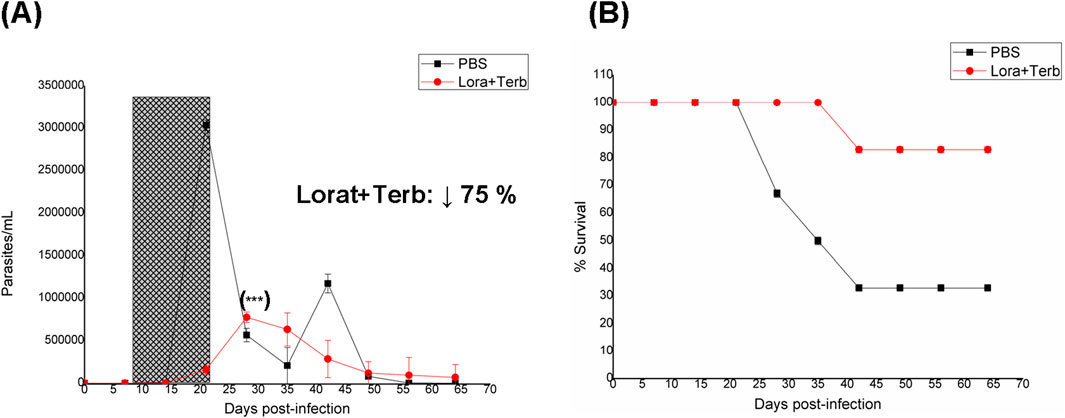
Figure 4. In vivo results for the binary mixture of loratadine and terbinafine (Lorat + Terb). (A) Parasitemia vs. week of the assay; the period of oral treatment is marked in gray. Inset: Percentage decrease in parasitic load with respect to PBS treatment at the maximum parasitemia peak. (B) Animal survival vs. week of the assay. (***) p < 0.001 (with respect to the maximum parasitemia peak for PBS treatment).
The in vivo Lap + Clim combination was observed to promote a slight time shift of the first peak of parasitemia by 1 week, along with an excellent decrease in parasite load (70%) and abolished second peak (Figure 3). The survival rates of the animals in this group also improved compared to the PBS-treated animals (first death at fourth week and 83% animal survival at the end of treatment). The Lap + Clim combination showed agreement with the in vitro FICJ774.1A value (Table 1), improving animal survival with respect to monotherapy with Clim (first death at third week and 60% animal survival at the end of treatment) or Lap (first death at third week and 63% animal survival at the end of treatment) (Figures 1B, 3B). Similar to the Lorat + Terb combination, the IgG anti-T. cruzi antibody levels of Lap + Clim were not good, meaning that this combination failed to reduce the antibody levels compared to PBS treatment (Table 3).
The orally administered Naf + Gab combination showed adequate in vitro behavior and was able to shift and reduce the first parasitemia peak (73%), along with almost abolishing the second peak (Figure 1). Additionally, this combination was the best among the different treatments in terms of animal survival rate (reaching 100% animal survival by the end of treatment) and had the highest in vitro FICJ774.1A value (Table 1) that was better than the individual values of Naf and Gab. Lastly, the Naf + Gab combination significantly diminished the IgG anti-T. cruzi antibody levels by the end of the study (Table 3).
The final binary mixture of Naf + Clim was the best among the different combinations studied herein in terms of the shift (2 weeks) and reduction (87%) of the first parasitemia peak as well as abolishing of the second peak (Figure 1). In agreement with the in vitro results (Table 1), the animal survival rate was worse than observed with the individual drugs (first death at second week and 80% animal survival at the end of treatment for Naf + Clim vs. first death at fourth week and 80% animal survival at the end of treatment for Naf vs. first death at third week and 60% animal survival at the end of treatment for Clim). Additionally, the Naf + Clim combination produced the best results in the present study (similar to Bnz) and reduced the IgG anti-T. cruzi antibody levels by the end of the study (Table 3).
3.3 Mutagenicity studies of the two best binary combinations
To advance preclinical studies, we conducted assays of the mutagenicities of the two best binary combinations observed herein, namely, Naf + Gab and Naf + Clim, according to the guidelines of the Organisation for Economic Co-operation and Development (Organisation for Economic Co-operation and Development, 2002). The Ames test was performed for both combinations using the Salmonella typhimurium strain TA98 with (+S9 fraction) and without (−S9 fraction) metabolic activation. Both combinations were found to be non-mutagenic both with and without metabolic activation (Table 4) since there were no dose-dependent increases in the numbers of revertant colonies, and the revertant count of each combination remained below the threshold of twice the revertant–vehicle control values (Alamón et al., 2020).
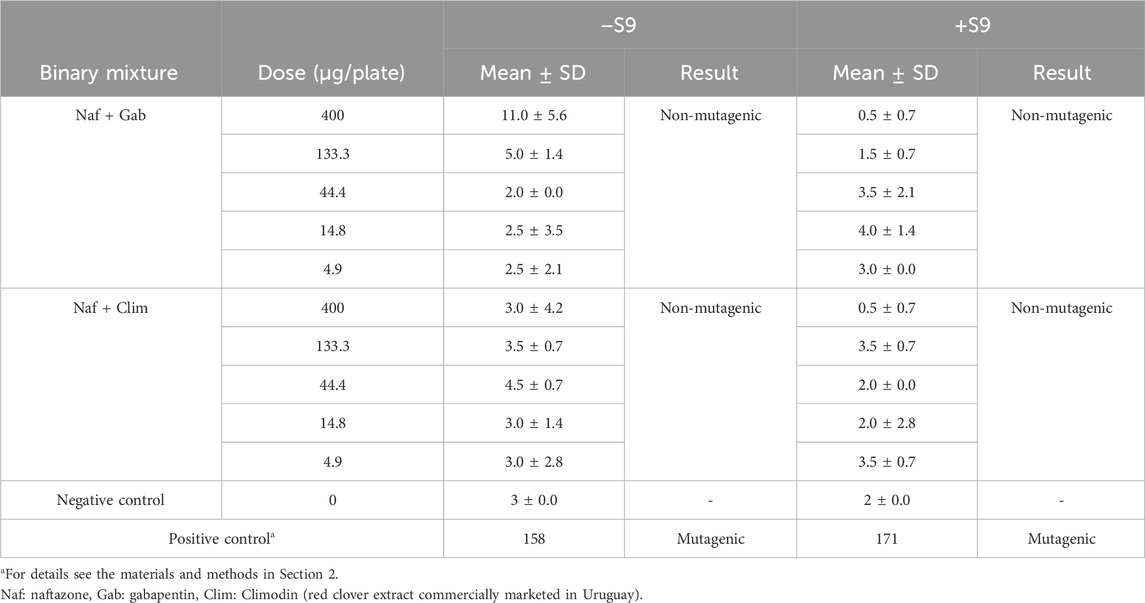
Table 4. Numbers of colonies observed in the Ames test strain TA98 without (−S9) and with (+S9) metabolic activation for binary mixtures Naf + Gab and Naf + Clim.
4 Discussion
In this work, we report the follow-up findings from our earlier study to overcome the hit-to-preclinical candidate stage for potential anti-Chagas agents based on drug repurposing processes developed by our group (Veira et al., 2025).
Our results confirm the good correlations between the in vitro results against epimastigotes and corresponding in vivo findings in a murine model of acute CD. These efforts allow validation of the in vitro procedures and results as a safe initial screening method without the need for animal experimentation.
In the case of monotherapy treatments, Naf and Gab were found to be appropriate drugs. For Naf, we observed good parasitemia diminishing levels with concomitant good animal survival profile; however, the IgG anti-T. cruzi antibody levels need be reduced, possibly through modification of the dosage schedule. For example, oral retreatment of the animals after a break could help reduce the presence of parasites, consequently lowering the antibody load. For Gab, we observed adequate reductions in parasitemia levels and animal death rates, with the best result for this monotherapy treatment being in terms of the IgG anti-T. cruzi antibody levels at the end of treatment; this result was in agreement with the parasitemia level at the end of treatment, where the animals treated with Gab showed near-zero parasites (Figure 1).
In the case of binary combination treatments, the best results were obtained with Naf + Gab and Naf + Clim. Except for the animal survival rate for the Naf + Clim mixture, all analyzed parameters for both combinations were good to excellent. Among the binary combinations assessed herein, Naf + Clim was the most effective in reducing the parasite load, achieving the maximum peak parasitemia abolishment, and maintaining low antibody levels at the end of treatment. For the other two binary combinations studied, namely, Lorat + Terb and Lap + Clim, the parasitemia reductions and survival rates of the animals were very good; however, the IgG anti-T. cruzi antibody levels at the end of experiment were insufficiently reduced.
As approved drugs, Naf, Gab, and Clim have interesting clinical profiles that could be advantageous for people suffering from CD. Naf is commonly used to treat varicose veins, hemorrhoids, and venous diseases to stabilize capillary walls, achieve anti-inflammatory actions, improve blood rheology, protect endothelial cells from oxidative stress, and modulate enzymatic activities (Kitchens et al., 2020). The action mechanism of Naf is related to controlling the oxidation–reduction reactions at the lysosomal membranes (Agha and Gad, 1995) as well as inhibiting cellular Ca2+ uptake via the Naf glucuronide metabolite (Mattei et al., 1999), where Ca2+ plays a critical role in the glutamate-induced lysosomal membrane permeability (Yan et al., 2016). The clinical manifestations of CD-related cardiac conditions, such as arrhythmias, structural heart diseases, cardiomyopathies, and cardiac insufficiency (Frade et al., 2025), may benefit from Naf treatment owing to its modification of calcium homeostasis and anti-inflammatory effects on the heart. Similarly, Gab is commonly used to treat partial seizures and neuropathic pain, and its action mechanism is through inhibition of the Ca2+ channels (Varadi G., 2024) that could also offer benefits against chronic Chagasic cardiomyopathy. The natural product extract Clim marketed in Uruguay is mainly composed of the isoflavones biochanin A, daidzein, formononetin, and genistein; it affects the climacteric symptoms by acting positively on the type β estrogen receptor (ERβ). It has been reported that the activation of estrogen receptors like ERα, ERβ, and GPR30 could have cardioprotective effects (Montoya and Krysan, 2018); these effects could be advantageous in CD leading to cardiovascular issues, such as fibrosis opening doors to new therapeutic strategies, and particularly for heart diseases that often follow chronic Chagasic infections (Frade et al., 2025). In addition to the availability of such natural product extract drugs in countries like Uruguay, the clinical and therapeutic effects of the drugs make them good candidates for CD treatment in the future.
The limitations of existing treatments for CD, which still affects a large portion of the global population with limited executive power, and the limited interest shown by pharmaceutical companies toward suitable drug development, make studies like those reported by our research group relevant. The present work aims to promote the development of new dosage regimens with repurposed drugs like Naf, Gab, and Clim as well as their combinations Naf + Gab and Naf + Clim through modifications of the doses and administration times, among others.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The animal study was approved by the Paraguayan Ethics Committee for Animal Experimentation-IICS-UNA (protocol no. IORG0010088). The study was conducted in accordance with all local legislation and institutional requirements.
Author contributions
CV: Writing – review and editing, Data curation, Investigation, Formal Analysis. BD: Formal Analysis, Writing – review and editing, Data curation, Investigation. ES: Investigation, Writing – review and editing, Data curation, Formal Analysis. NP: Formal Analysis, Writing – review and editing, Investigation, Data curation. GY: Writing – review and editing, Formal Analysis, Data curation, Conceptualization, Investigation. GÁ: Writing – review and editing, Investigation, Conceptualization, Formal Analysis, Data curation. HC: Writing – review and editing, Software, Methodology, Investigation, Writing – original draft, Supervision, Funding acquisition, Resources, Visualization, Data curation, Conceptualization, Formal Analysis, Project administration, Validation. EA: Data curation, Software, Visualization, Investigation, Methodology, Conceptualization, Resources, Funding acquisition, Project administration, Validation, Writing – review and editing, Writing – original draft, Formal Analysis, Supervision.
Funding
The authors declare that financial support was received for the research and/or publication of this article. This work was supported by the Agencia Nacional de Investigación e Innovación (ANII, project FCE_3_2022_1_172684), Programa de Desarrollo de las Ciencias Básicas (PEDECIBA), and the Universidad de la República (UdelaR).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Agha, A. M., and Gad, M. Z. (1995). Lipid peroxidation and lysosomal integrity in different inflammatory models in rats: the effects of indomethacin and naftazone. Pharmacol. Res. 32, 279–285. doi:10.1016/s1043-6618(05)80015-5
Aguilera, E., Varela, J., Serna, E., Torres, S., Yaluff, G., Bilbao, N. V. d., et al. (2018). Looking for combination of benznidazole and Trypanosoma cruzi-triosephosphate isomerase inhibitors for chagas disease treatment. Mem. Inst. Oswaldo Cruz. 113, 153–160. doi:10.1590/0074-02760170267
Aguilera, E., Sánchez, C., Cruces, M. E., Dávila, B., Minini, L., Mosquillo, F., et al. (2022). Preclinical studies and drug combination of low-cost molecules for chagas disease. Pharmaceuticals (Basel) 16, 20. doi:10.3390/ph16010020
Alamón, C., Dávila, B., García, M. F., Sánchez, C., Kovacs, M., Trias, E., et al. (2020). Sunitinib-containing carborane pharmacophore with the ability to inhibit tyrosine kinases receptors FLT3, KIT and PDGFR-β, exhibits powerful in vivo anti-glioblastoma activity. Cancers 12, 3423. doi:10.3390/cancers12113423
Aria, L., Acosta, M. E., Guillen, Y., Rojas, A., Meza, T., and Infanzón, B. (2016). Desempeño del Kit ELISA Chagas IICS V.1 para el diagnóstico de la enfermedad de Chagas. Mem. Inst. Investig. Cienc. Salud. 14, 7–13. doi:10.18004/mem.iics/1812-9528/2016.014(03)07-013
Cerecetto, H., and González, M. (2010). Synthetic medicinal chemistry in Chagas’ disease: compounds at the final stage of “Hit-to-Lead” phase. Pharmaceuticals (Basel) 3, 810–838. doi:10.3390/ph3040810
Chatelain, E., and Konar, N. (2015). Translational challenges of animal models in Chagas disease drug development: a review. Drug Des. Dev. Ther. 9, 4807–4823. doi:10.2147/DDDT.S90208
Dávila, B., Sánchez, C., Fernández, M., Cerecetto, H., Lecot, N., Cabral, P., et al. (2019). Selective hypoxia-cytotoxin 7-fluoro-2-aminophenazine 5,10-dioxide: toward “candidate-to-drug” stage in the drug-development pipeline. ChemistrySelect 4, 9396–9402. doi:10.1002/slct.201902601
Frade, A. F., Guérin, H., Nunes, J. P. S., Silva, L. F. S. E., Roda, V. M. D. P., Madeira, R. P., et al. (2025). Cardiac and digestive forms of chagas disease: an update on pathogenesis, genetics, and therapeutic targets. Mediat. Inflamm. 2025, 8862004. doi:10.1155/mi/8862004
Kitchens, B. P., Snyder, R. J., and Cuffy, C. A. (2020). A literature review of pharmacological agents to improve venous leg ulcer healing. Wounds 32, 195–207.
Kratz, J. M. (2019). “Drug discovery for chagas disease: a viewpoint,” in Acta tropica (Amsterdam, Netherlands: Elsevier B.V). doi:10.1016/j.actatropica.2019.105107
Maldonado, R. A., Molina, J., Payares, G., and Urbina, J. A. (1993). Experimental chemotherapy with combinations of ergosterol biosynthesis inhibitors in murine models of Chagas disease. Antimicrob. Agents Chemother. 37, 1353–1359. doi:10.1128/AAC.37.6.1353
Mattei, C., Molgó, J., Joseph, X., Israel, M., and Bloy, C. (1999). Naftazone reduces glutamate cerebrospinal fluid levels in rats and glutamate release from mouse cerebellum synaptosomes. Neurosci. Lett. 271, 183–186. doi:10.1016/s0304-3940(99)00552-2
Montoya, M. C., and Krysan, D. J. (2018). Repurposing estrogen receptor antagonists for the treatment of infectious disease. mBio 9, e02272-18–18. doi:10.1128/mBio.02272-18
Navarro, M., Reguero, L., Subirà, C., Blázquez-Pérez, A., and Requena- Méndez, A. (2022). Estimating Chagas disease prevalence and number of underdiagnosed, and undertreated individuals in Spain. Travel Med. Infect. Dis. 47, 102284. doi:10.1016/j.tmaid.2022.102284
OECD – Organisation for Economic Co-operation and Development (2002). Test no. 423: acute oral toxicity - acute toxic class Method, OECD Guidelines for the testing of Chemicals, section 4. Paris: OECD Publishing. doi:10.1787/9789264071001-en
PAHO - Pan American Health Organization (2025). World Chagas disease day 2025. Available online at: https://www.paho.org/en/campaigns/world-chagas-disease-day-2025 (accessed on June 30, 2025).
Patel, G., Karver, C. E., Behera, R., Guyett, P. J., Sullenberger, C., Edwards, P., et al. (2013). Kinase scaffold repurposing for neglected disease drug discovery: discovery of an efficacious, lapatanib-derived lead compound for trypanosomiasis. J. Med. Chem. 56, 3820–3832. doi:10.1021/jm400349k
Planer, J. D., Hulverson, M. A., Arif, J. A., Ranade, R. M., Don, R., and Buckner, F. S. (2014). Synergy testing of FDA-approved drugs identifies potent drug combinations against Trypanosoma cruzi. PLoS Negl. Trop. Dis. 8, e2977. doi:10.1371/journal.pntd.0002977
Romer, G., Bracco, L. A., Ricci, A. D., Balouz, V., Berná, L., Villar, J. C., et al. (2023). Deep serological profiling of the Trypanosoma cruzi TSSA antigen reveals different epitopes and modes of recognition by chagas disease patients. PLoS Negl. Trop. Dis. 17, e0011542. doi:10.1371/journal.pntd.0011542
Sayé, M., Gauna, L., Valera-Vera, E., Reigada, C., Miranda, M. R., and Pereira, C. A. (2020). Crystal violet structural analogues identified by in silico drug repositioning present anti-Trypanosoma cruzi activity through inhibition of proline transporter TcAAAP069. PLoS Negl. Trop. Dis. 14, e0007481. doi:10.1371/journal.pntd.0007481
Varadi, G. (2024). Mechanism of analgesia by gabapentinoid drugs: involvement of modulation of synaptogenesis and trafficking of glutamate-gated ion channels. J. Pharmacol. Exp. Ther. 388, 121–133. doi:10.1124/jpet.123.001669
Veira, C., Benítez, D., Pérez-Díaz, L., Álvarez, G., Cerecetto, H., and Aguilera, E. (2025). Looking for approved-medicines to be repositioned as anti-Trypanosoma cruzi agents. Identification of new chemotypes with good individual- or in combination-biological behaviours. Mem. Inst. Oswaldo Cruz. 120, e240183. doi:10.1590/0074-02760240183
Keywords: Chagas disease, Trypanosoma cruzi, drug repurposing, gabapentin, naftazone, red clover extract, pinaverium bromide
Citation: Veira C, Dávila B, Serna E, Portillo N, Yaluff G, Álvarez G, Cerecetto H and Aguilera E (2025) Drug repurposing for Chagas disease: overcoming the hit-to-preclinical candidate stage. Front. Chem. Biol. 4:1662907. doi: 10.3389/fchbi.2025.1662907
Received: 09 July 2025; Accepted: 30 October 2025;
Published: 26 November 2025.
Edited by:
Kaio Cesar Chaboli Alevi, Sao Paulo State University, BrazilReviewed by:
Carlos Eliel Maya-Ramírez, National Polytechnic Institute, MexicoEsteban Rodriguez-Arce, Universidad de Chile, Chile
Copyright © 2025 Veira, Dávila, Serna, Portillo, Yaluff, Álvarez, Cerecetto and Aguilera. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Elena Aguilera, ZWFndWlsZXJhQGZjaWVuLmVkdS51eQ==
†ORCID: Claudia Veira, orcid.org/0009-0001-1946-5938; Belén Dávila, orcid.org/0009-0006-3283-8192; Elva Serna, orcid.org/0000-0002-3481-012X; Nilda Portillo, orcid.org/0000-0001-6216-3516; Gloria Yaluff, orcid.org/0000-0002-7568-4756; Guzmán Álvarez, orcid.org/0000-0001-8385-6100; Hugo Cerecetto, orcid.org/0000-0003-1256-3786; Elena Aguilera, orcid.org/0000-0002-1052-687X