 Andrés F. Quintero-Mercado1,2*
Andrés F. Quintero-Mercado1,2* Sebastián Rojas3
Sebastián Rojas3 Yordan J. Romero-Contreras4
Yordan J. Romero-Contreras4 Luis F. Lozano4
Luis F. Lozano4 Mario Serrano4
Mario Serrano4 Celsa García1
Celsa García1- 1Facultad de Ciencias Agrarias, Departamento de Agronomía, Universidad Nacional de Colombia, Bogotá, Colombia
- 2Facultad de Ingeniería, Programa de Ingeniería Agronómica, Universidad del Magdalena, Santa Marta, Colombia
- 3Facultad de Ciencias, Departamento de Biología, Universidad Nacional de Colombia, Bogotá, Colombia
- 4Centro de Ciencias Genómicas, Universidad Nacional Autónoma de México, Cuernavaca, Morelos, Mexico
Colletotrichum tropicale is an endophyte that has been reported as a pathogen in ripe mango fruits (Mangifera indica L. cv. Azúcar) in Magdalena, Colombia, causing anthracnose. However, gene expression in the host that promotes its lifestyle transition remains unknown. This study aimed to analyze gene expression during the interaction between ripe mango fruit cv. Azúcar and C. tropicale to identify differentially expressed host genes that facilitate the pathogen’s infection process. RNA sequencing (RNA-seq) analysis was conducted at 0 and 12 h post inoculation (hpi), including de novo assembly and bioinformatic functional annotation using Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG). A total of 5,435 differentially expressed genes (DEGs) were identified in the interaction, of which only 421 DEGs were detected in ripe mango fruits. Among these, 379 were upregulated and 42 were downregulated (T0 vs. T12 hpi). GO functional annotation of downregulated fruit genes revealed that the molecular functions affected at 12 hpi were related to the plant’s defensive oxidative burst mediated by reactive oxygen species (ROS)—including NADPH oxidase activity, hydrogen peroxide formation, and the action of peroxidases and oxidoreductase enzymes—whereas upregulated genes were associated with stress response, defense, transferase activity, and kinase activity. KEGG analysis identified pathways related to mitogen-activated protein kinase (MAPK) signaling, pathogen-associated molecular pattern (PAMP)-triggered immunity, and phenylalanine metabolism. In conclusion, ripe mango fruit cv. Azúcar activates a defense response against C. tropicale at 12 hpi that does not overcome the pathogen’s initial quiescent phase but instead facilitates conditions for its establishment by suppressing oxidative burst pathways, which may later contribute to oxidative stress during the necrotrophic phase.
1 Introduction
Over the past three decades, mango (Mangifera indica L.) cultivation has expanded in production area and increased its commercial supply to new international markets. This fruit is of great importance due to its high nutritional value, as it contains macronutrients such as carbohydrates, lipids, fatty acids, and amino acids, as well as micronutrients including vitamins and minerals (Ledesma and Campbell, 2019; Maldonado-Celis et al., 2019). According to Arcila Cardona et al. (2022), the mango cv. Azúcar has been naturalized in Colombia, where it is grown in warm and dry areas ranging from 11 to 556 m above sea level (m.a.s.l.), mainly in regions such as Cesar, Córdoba, Cundinamarca, Huila, Magdalena, and Tolima (García Lozano et al., 2010). Among these, Magdalena contributed 20% to the national mango production according to the Ministerio de Agricultura y Desarrollo Rural, Colombia (2025).
The production system of mango cv. Azúcar is the fourth most economically important crop in the region of Magdalena (Colombia), after banana, African oil palm, and coffee (Salcedo, 2018). It is affected by anthracnose, a disease caused by the phytopathogenic fungus Colletotrichum, which is considered the most limiting factor in mango production across tropical and subtropical regions (Arauz, 2000; de Souza et al., 2013; Kamle, 2013; Ploetz, 2007). In this region, the disease can cause yield losses ranging from 20% to 50% both in the field and postharvest (Páez, 2003), as well as severe damage that reduces the productivity and competitiveness of various agroecosystems and plants in general, with losses reaching up to 100% (Crouch et al., 2014; da Silva et al., 2020; De Silva et al., 2017; Vieira et al., 2014). Regarding the endophytism of Colletotrichum, few reports confirm that this fungus can undergo a phase shift to a pathogenic lifestyle in mango. This has been evidenced in studies by Quintero-Mercado et al. (2019) and Páez-Redondo et al. (2024), in which C. tropicale was obtained as an endophyte from asymptomatic leaves and fruits of the cv. Azúcar, respectively, and when inoculated onto detached fruits, necrotic lesions characteristic of anthracnose symptoms developed. Similarly, Vieira et al. (2014) reported that endophytic isolates from mango cv. Tommy Atkins, including C. tropicale, C. asianum, C. endomangiferae, C. cliviae, C. dianesei, C. fructicola, and C. karstii, exhibited significant virulence in pathogenicity assays on detached fruits. For the breakdown of a mutualistic interaction between the fungus and the plant to occur, predisposing conditions must be present—provided by the host (e.g., biochemical changes), by the pathogen’s virulence, or by favorable environmental conditions in the ecosystem (De Silva et al., 2017; Delaye et al., 2013). The ability of this fungal microorganism to remain asymptomatic within plant tissue poses a phytosanitary risk due to the hidden production of infective propagules or initial inoculum (Freeman et al., 2001).
According to Kogel et al. (2006), endophytes must overcome the plant’s nonspecific defense responses, achieving successful penetration of the cell wall through the reprogramming of invaded cells, which leads to adaptation of the infective structures formed. This process preserves host integrity and ensures a long-lasting interaction, or a “friendly” infection, which in some cases is mediated by recognition through plant transmembrane kinase receptors. In this regard, the host must recognize conserved molecules of certain microorganisms, known as microbe-associated molecular patterns (MAMPs) or pathogen-associated molecular patterns (PAMPs) (Khare et al., 2018). According to Sánchez-Vallet et al. (2015), endophytes also develop defense mechanisms against the plant’s immune response.
Pathogenic symbioses are characterized by the production of enzymes that degrade the plant cell wall, in which damage-associated molecular pattern (DAMP) receptors perceive cellular debris, adenosine triphosphate (ATP), and carbohydrates to initiate an immune signaling response by the host. In contrast, mutualistic symbioses do not cause apparent damage to the integrity of the plant cell, and DAMP receptors are possibly silenced, repressing the plant’s immune signaling pathways (Plett and Martin, 2018).
In the case of a lifestyle shift from mutualistic symbiosis to pathogenesis, it can be stated that the microorganism is capable of activating mitogen-activated protein kinase (MAPK) pathways and the enzymatic complex NADPH oxidase that produces reactive oxygen species (ROS), possibly through the secretion of apoplastic as well as cytoplasmic effectors, with the purpose of damaging the cell wall (Eaton et al., 2011).
The use of omics tools, such as transcriptomics through RNA sequencing (RNA-seq), has not been explored to understand the endophyte–pathogen relationship of Colletotrichum in mango, particularly in cv. Azúcar from the region of Magdalena, Colombia. Such studies would allow the establishment of transcriptional profiles to help elucidate the nature of mutualism/endophytism and the host immune response (Dinkins et al., 2017; Eaton et al., 2011; Kaul et al., 2016), generating new knowledge of the genetic factors influencing the variation in lifestyles of Colletotrichum in organs such as ripe fruits. This would lead to the development of more effective control strategies against anthracnose and strengthen the agro-industrial competitiveness of mango cv. Azúcar. In this study, the differential gene expression of ripe mango fruit cv. Azúcar that favors the pathogenicity of the endophyte C. tropicale was analyzed.
2 Materials and methods
2.1 Disinfection of ripe mango fruits cv. Azúcar for inoculation tests
According to the BBCH scale proposed by Rajan et al. (2011), ripe fruits in phenological stage 801, detached and completely healthy, were used. Disinfection was carried out following the methodology proposed by Quintero-Mercado et al. (2019). Under laminar flow chamber conditions, fruits were immersed, in the following order, in sterile distilled water for 2 min, in 1% sodium hypochlorite for 1 min, and again in sterile distilled water for 2 min. Subsequently, they were sprayed with 70% ethanol for 1 min, and the excess was removed using sterile distilled water. Once disinfected, the fruits were left to dry completely on sterile Kraft paper for 45 min.
2.2 Preparation of the inoculum and inoculation of C. tropicale
The fungal inoculum was prepared from pure cultures with 20 days of growth on potato dextrose agar (PDA) medium supplemented with streptomycin sulfate (200 mg/L), under conditions of 26°C, a 12 h light/12 h dark photoperiod, and 80% relative humidity. The endophytic isolate was obtained from asymptomatic leaves of mango cv. Azúcar. Its pathogenicity was confirmed, and it was identified as Colletotrichum tropicale (GenBank® OR563797.1). In a laminar flow chamber, 20 mL of sterile distilled water were added to each culture, and the surface was scraped with a sterile spatula to collect the entire fungal content while avoiding contamination with the culture medium. The suspension was gently stirred and transferred to a sterile glass beaker, then macerated with a spatula and vortexed for 3 min. It was filtered through sterile medical gauze and collected in another aseptic glass beaker. The fungal suspension was adjusted to a concentration of 1 × 106 conidia/mL.
Using the drop-deposition inoculation technique, the inoculum was applied to six equidistant points on the epicarp of each fruit, with a volume of 10 μL per point and a second application at each site for a total of 20 μL of suspension. Six fruits, considered one biological replicate, were used for each evaluation time point (0 and 12 h post inoculation [hpi]), following the histological study by Quintero-Mercado (2025), in which it was identified that at 12 hpi the fungus employs quiescence as a pathogenic strategy in ripe mango cv. Azúcar, behaving as a hemibiotroph. At 0 hpi, inoculation was performed with sterile distilled water, and one additional fruit was inoculated with the fungal suspension to monitor symptoms and characteristic signs of the disease. Once inoculated, the fruits were kept under high-humidity conditions inside hermetically sealed plastic containers at a temperature between 22 °C and 25°C and 80% relative humidity in an environmental test chamber (Sanyo, now PHCbi). For each evaluation time point (0 and 12 hpi), three biological replicates were used. In each tube, 2 mL of NucleoProtect RNA reagent (Macherey-Nagel) were added to preserve RNA integrity.
2.3 Transcriptomic analysis
The samples preserved in NucleoProtect RNA (Macherey-Nagel) were sent to the Beijing Genomics Institute (BGI, Hong Kong) for total RNA extraction, library construction, and sequencing using DNBSeq™ technology with a PE150 read length.
To assess sequence quality, FastQC (version 0.11.8) (https://www.bioinformatics.babraham.ac.uk/projects/fastqc/) was used, followed by trimming with Trim-Galore (version 0.6.10) (https://www.bioinformatics.babraham.ac.uk/projects/trim_galore/). De novo assembly was performed using Trinity (version 2.15.1) (Grabherr et al., 2011), pooling all biological replicates for each time point or condition (T0 hpi = 3; T12 hpi = 3). The resulting transcripts were translated into proteins using TRANSDECODER (version 5.7.1) (https://github.com/TransDecoder/TransDecoder). The quality of the de novo assembly was assessed by performing BLAST (version 1.15.0) (Altschul et al., 1990) against UniProt release 2024_02 (https://www.uniprot.org/), and by aligning the reads of each biological replicate to the assembled transcripts using BOWTIE2 (version 2.5.2) (Langmead and Salzberg, 2012). Expectation-maximization (RSEM) (version 1.3.3) (Li and Dewey, 2011) was used to estimate gene abundance in each replicate, and DESeq2 (version 3.19) (Love et al., 2014) was applied for differential expression analysis. A log2 fold change (FC) ≥ 2 or ≤ −2 was used as the threshold.
2.4 Functional gene annotation through Gene Ontology (GO) in ripe mango fruit cv. Azúcar
For functional annotation, a BLAST search (version 1.15.0) (Altschul et al., 1990) was performed using three databases: the complete and annotated genome of Mangifera indica, compared against the plant-specific Nr database. Proteins with at least 35% identity and 70% coverage were selected.
After obtaining statistically significant genes (p-value, FDR < 0.05), GO analysis was conducted using the Blast2GO® program (https://www.blast2go.com) (Götz et al., 2008), included in the OmicsBox tool, for the upregulated and downregulated genes of ripe mango fruit to identify molecular functions, biological processes, and cellular components.
2.5 Kyoto Encyclopedia of Genes and Genomes analysis for differentially expressed genes in ripe mango fruit cv. Azúcar
An enrichment analysis of specific molecular pathways was performed using KAAS–KEGG (version 2.1) (Kanehisa, 2004) (https://www.genome.jp/tools/kaas/) for the differentially expressed genes of ripe mango fruit cv. Azúcar.
2.6 Validation of gene expression data in ripe mango fruit cv. Azúcar by qRT-PCR
Total RNA was extracted using the InviTrap® Spin Plant RNA Mini Kit from 100 mg of inoculated tissue at different evaluation times (0, 12, and 24 h post inoculation [hpi]). RNA integrity was verified by electrophoresis on a 1.5% agarose gel. Samples were treated with DNase I (DNase I, Amplification Grade, Invitrogen™). For cDNA synthesis, the M-MLV Reverse Transcriptase Kit (Invitrogen™) was used.
Specific primers for each gene were designed with Primer3 (version 4.1.0) (Untergasser et al., 2012) and synthesized by Macrogen Inc. (Seoul, South Korea) (Supplementary Table 1). qRT-PCR was performed using the PowerTrack™ SYBR Green Master Mix Kit (Applied Biosystems™) and the QuantStudio™ 5 Real-Time PCR System (Applied Biosystems™, Thermo Fisher Scientific).
Amplification conditions were as follows: 50°C for 2 min and 95°C for 2 min, followed by 40 cycles of 95°C for 15 s and the optimal annealing temperature for each primer set for 60 s. Three biological replicates were used for each evaluation time and analyzed in technical triplicates. Results were normalized using the Pfaffl method and subsequently transformed to log2. Outliers were identified and removed using the IQR method.
Normality and homogeneity of variance were assessed for each gene and treatment. When assumptions were met, analysis of variance (ANOVA) followed by Tukey’s post hoc test was applied; otherwise, the Kruskal–Wallis test followed by Dunn’s test with Bonferroni correction for multiple comparisons was used. Actin was employed as the endogenous reference gene for expression normalization.
3 Results
3.1 Sequencing quality
The cleaning statistics of the reads, averaged across the three replicates for each evaluation time (0 and 12 hpi), yielded approximately 24 million reads for both cond_T0A and cond_T12A samples, with total read sizes of 7.1 Gb for T0A and 7.2 Gb for T12A. Quality parameters were Q20 > 96% and Q30 > 90% for all six samples, with a GC content of 44% (Supplementary Table 2).
3.2 Differential gene expression analysis of ripe mango cv. Azúcar fruit
Differential expression analysis of ripe mango cv. Azúcar fruit inoculated with the endophyte C. tropicale was performed using principal component analysis (PCA), where PC1 accounted for 58.20% and PC2 for 12.95% of the total data variance (Figure 1A). The correlation heatmap (Figure 1B) showed consistent statistical behavior among the three biological replicates for each evaluation time (0 and 12 hpi).
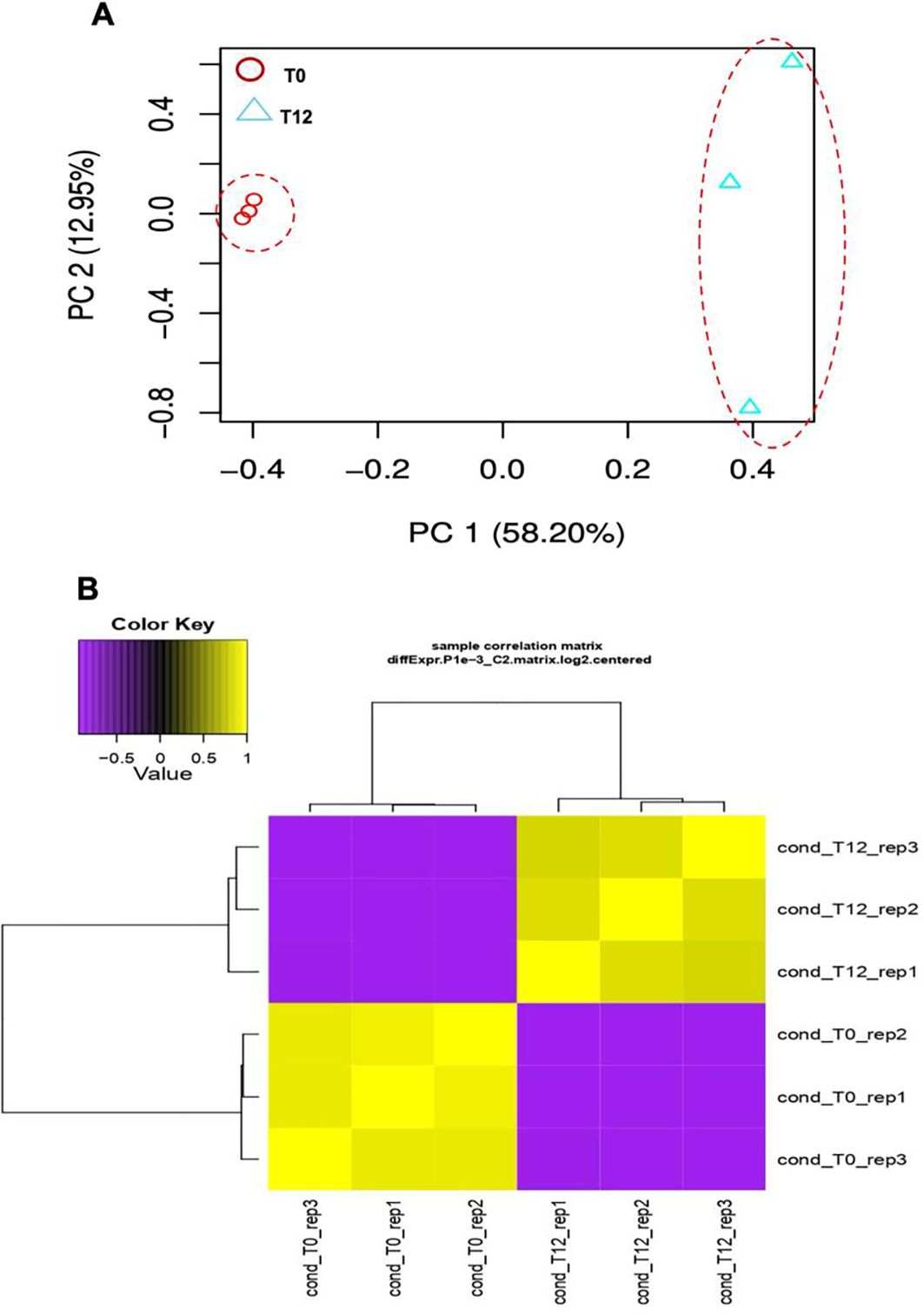
Figure 1. Transcriptomic analysis of the interaction between ripe mango fruit cv. Azúcar and C. tropicale under two evaluation time points (T0 vs. T12 h post inoculation). (A) Principal component analysis (PCA). Red dashed lines indicate separate clustering of each evaluation time point (T0 and T12 hpi). (B) Correlation heatmap of each evaluation time point (T0 and T12 hpi) across biological replicates (rep1, rep2, and rep3) (p < 0.05).
From the clean reads, de novo assembly yielded 152,318 transcripts corresponding to 89,357 genes. A total of 5,435 differentially expressed genes (DEGs) were identified (T0 vs. T12 hpi), visualized in a heatmap with log2 fold change (FC) ≥ 2 or ≤ −2 and p < 0.05 (Figure 2).
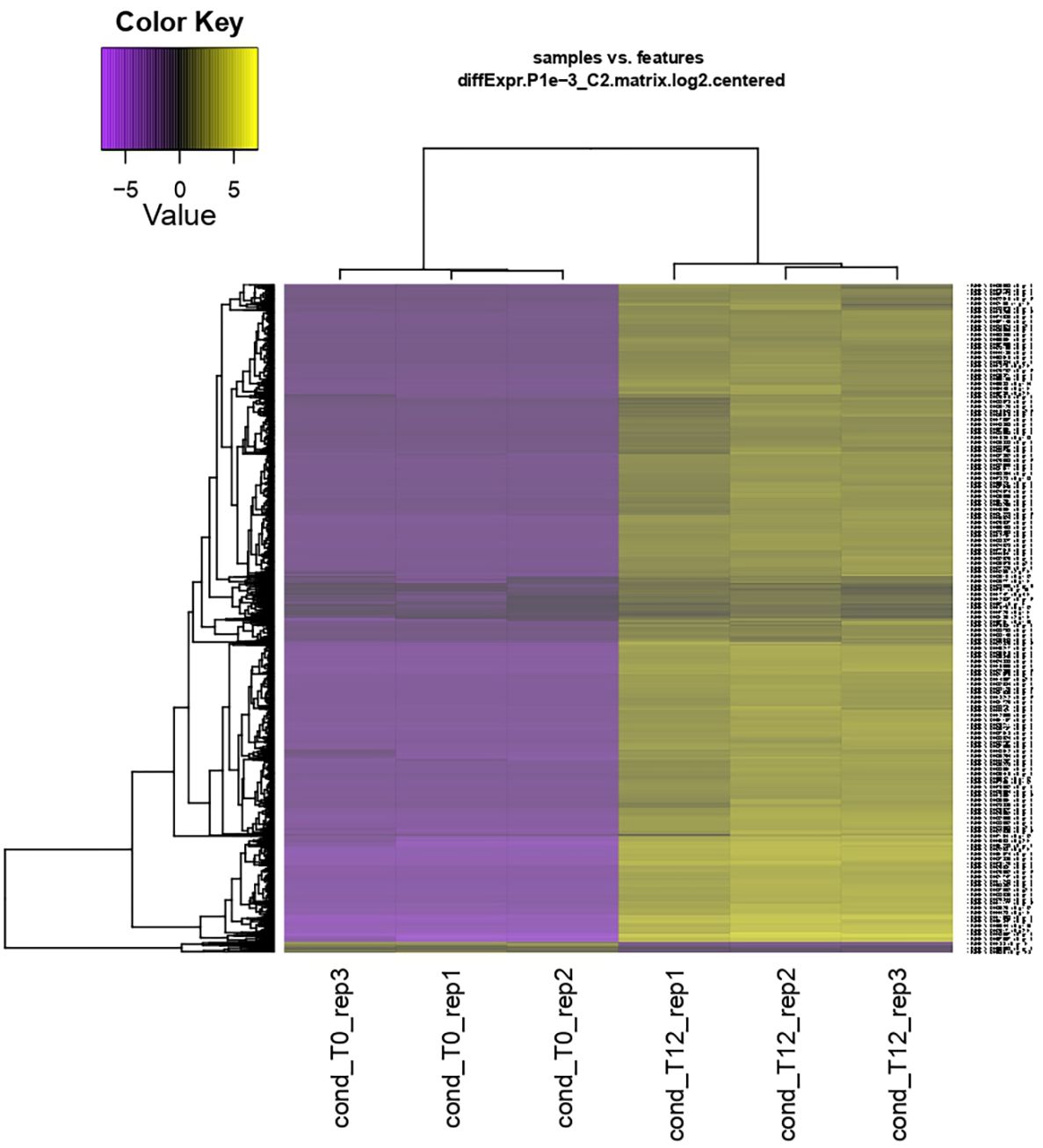
Figure 2. Hierarchical clustering and heatmap of differential expression analysis of the interaction between ripe mango fruit cv. Azúcar and C. tropicale at each evaluation time point (T0 and T12 hpi) across biological replicates (rep1, rep2, and rep3). Overexpressed genes are shown in yellow and repressed genes in purple. Log2 fold change (FC) ≥ 2 or ≤ −2 and p < 0.05.
3.3 Differentially expressed genes in ripe mango cv. Azúcar fruit
Of the 5,435 differentially expressed genes, only a total of 421 genes were identified in ripe mango fruit, with high significance against the Nr database for Mangifera indica, where 379 corresponded to upregulated genes and 42 to downregulated ones (T0 vs. T12 hpi) (Supplementary Data Sheet 1). Functional annotation by GO revealed that, for the upregulated genes, the largest number of sequences corresponding to gene subgroups were distributed in each category as follows: Biological Processes (response to stimulus: 151; response to stress: 113; defense response: 77), Cellular Components (membrane: 171; cell periphery: 103; plasma membrane: 100), and Molecular Functions (transferase activity: 69; phosphotransferase activity: 68; kinase activity: 68) (Figure 3). Regarding the downregulated genes, the highest number of sequences were grouped into the following subcategories: Biological Processes (biological process: 27; response to stimulus: 13; response to stress: 11), Cellular Components (cellular anatomical entity: 25; cellular component: 25; intracellular anatomical structure: 21), and Molecular Functions (cation binding: 14; metal ion binding: 13; calcium ion binding: 7) (Figure 4) (Supplementary Data Sheet 2).
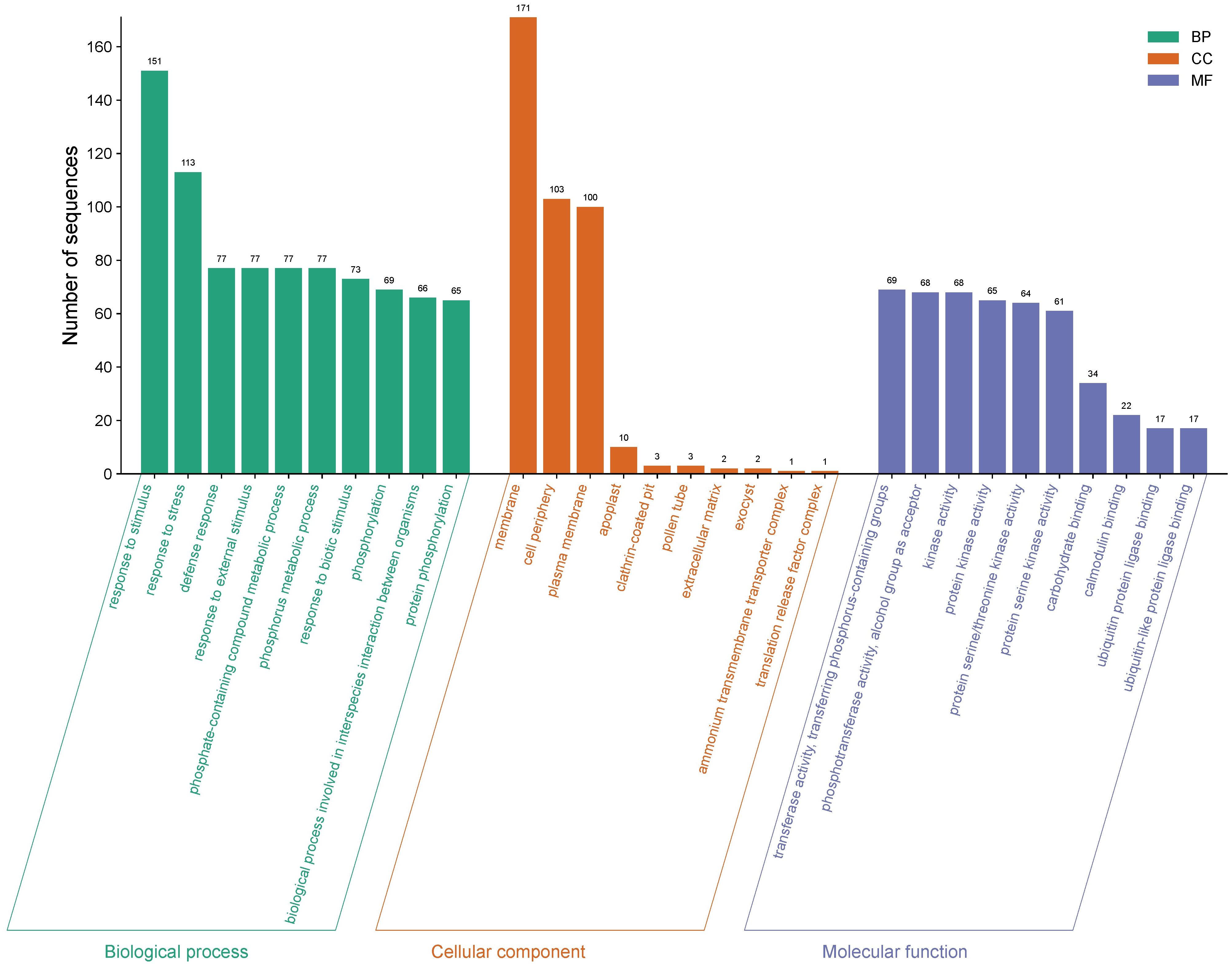
Figure 3. Gene Ontology (GO) terms for upregulated gene sequences in ripe mango fruit cv. Azúcar interacting with the endophyte C. tropicale at 12 hpi. Figure generated with SRPlot (Tang et al., 2023).
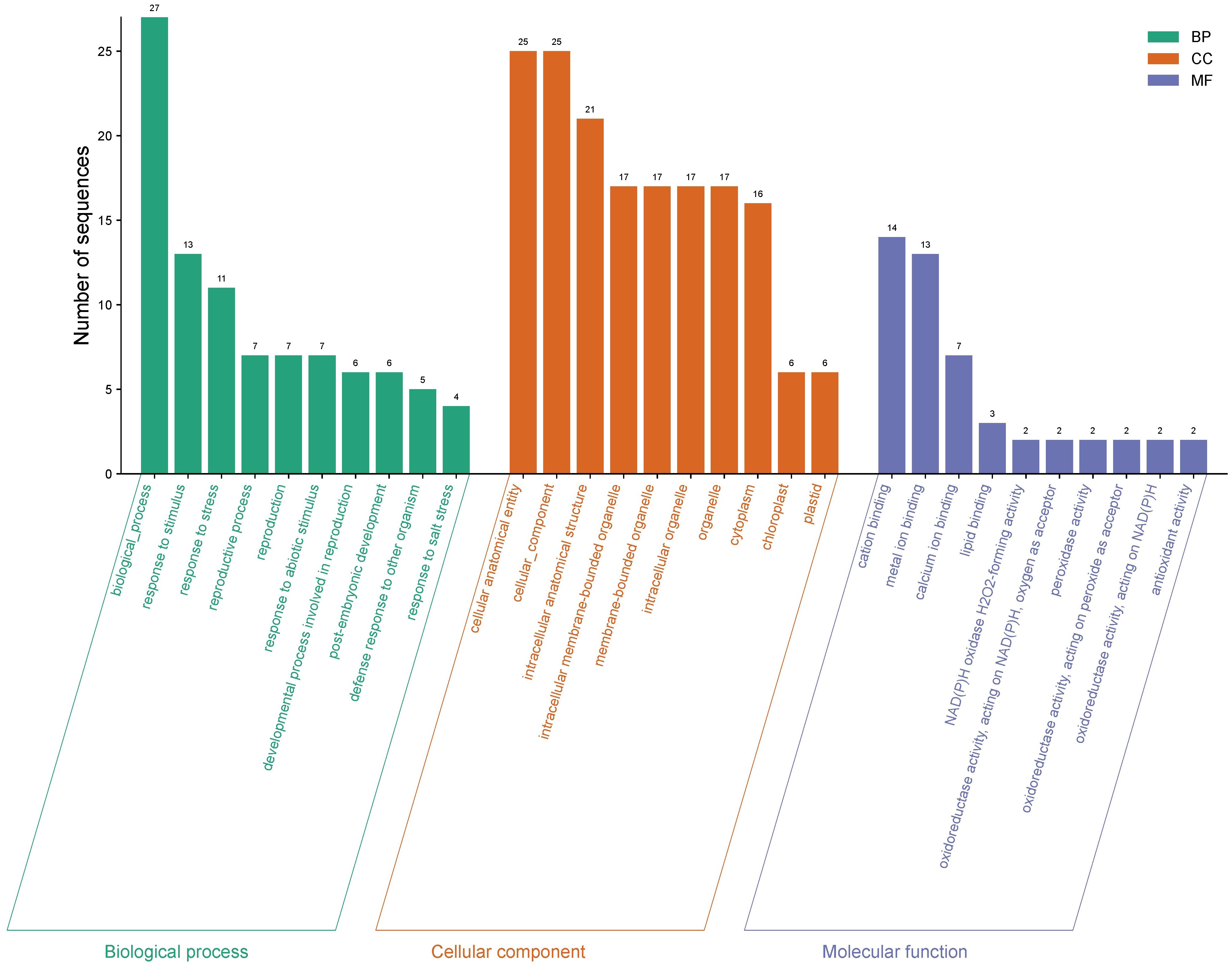
Figure 4. Gene Ontology (GO) terms for downregulated gene sequences in ripe mango fruit cv. Azúcar interacting with the endophyte C. tropicale at 12 hpi. Figure generated with SRPlot (Tang et al., 2023).
3.4 GO enrichment analysis of differentially expressed genes
A GO enrichment analysis was performed, and the enrichment score was calculated by selecting the top three most enriched terms per GO category. For the upregulated genes, the most significantly enriched terms were: Biological Processes – cell recognition (enrichment score: 10.52; p = 3.60 × 10-12; 16 sequences), response to oomycetes (8.69; p = 2.70 × 10-7; 10 sequences), and defense response to oomycetes (10.00; p = 3.10 × 10-7; 9 sequences); Cellular Components – ammonium transmembrane transporter complex (100.0; p = 0.0115; 1 sequence), translation release factor complex (20.0; p = 0.0454; 1 sequence), and cytochrome b6f complex (16.6; p = 0.0564; 1 sequence); Molecular Functions – naringenin-chalcone synthase activity (23.08; p = 1.70 × 10-6; 5 sequences), isoprenoid binding (17.94; p = 9.80 × 10-8; 7 sequences), and protein phosphatase inhibitor activity (17.5; p = 1.20 × 10-7; 7 sequences) (Figure 5).
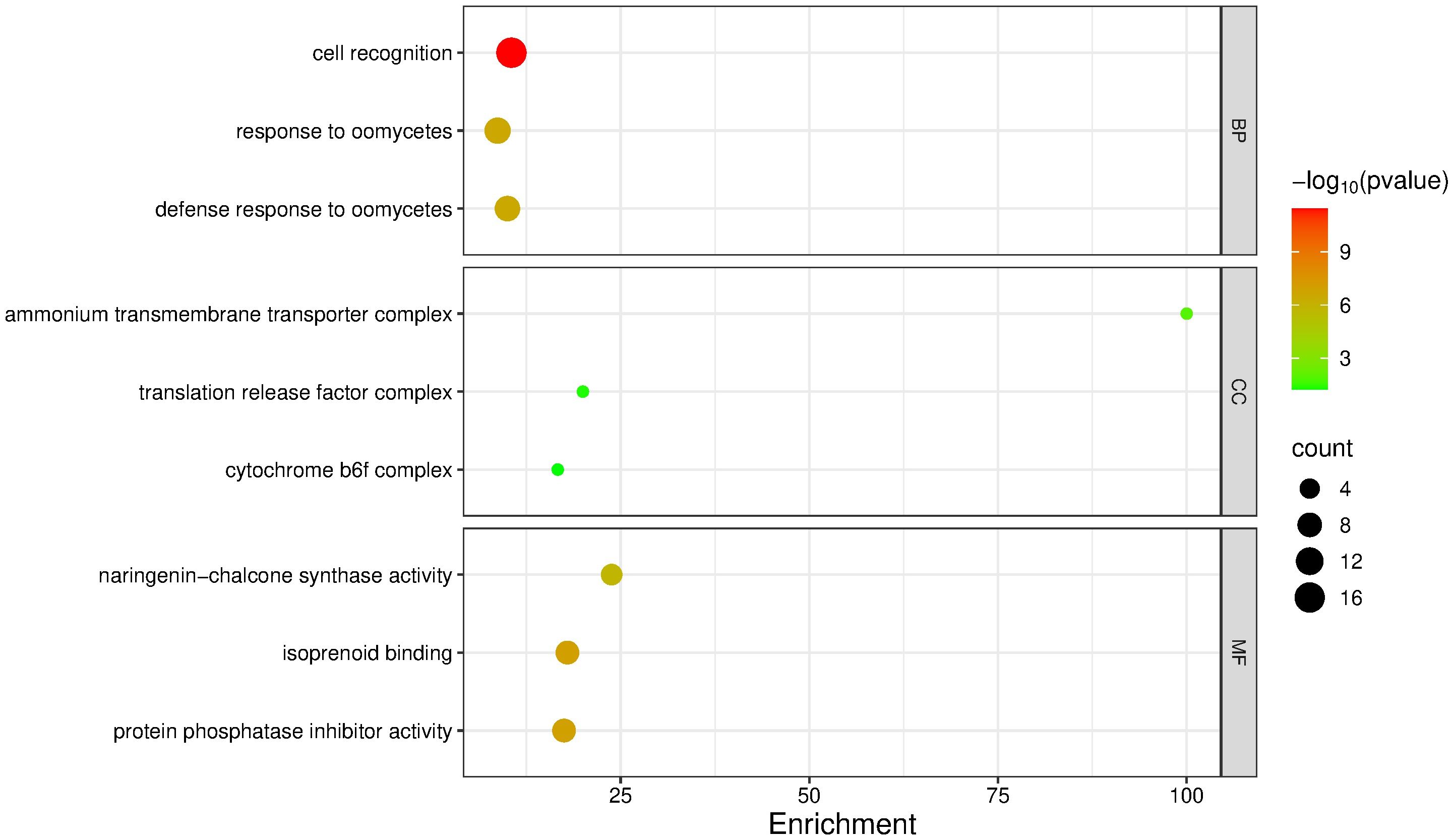
Figure 5. GO term enrichment analysis of upregulated genes in mango cv. Azúcar interacting with the endophyte C. tropicale at 12 hpi. Enrichment score (p < 0.05). Figure generated with SRPlot (Tang et al., 2023).
For the downregulated genes, the most enriched terms were: Biological Processes – response to stress (3.37; p = 2.70 × 10-4; 11 sequences), defense response to another organism (5.49; p = 0.0021; 5 sequences), and lipid transport (20.0; p = 4.30 × 10-4; 3 sequences); Cellular Components – cellular anatomical entity (1.54; p = 0.0056; 25 sequences), cellular component (1.54; p = 0.0059; 25 sequences), and intracellular anatomical structure (1.55; p = 0.0132; 21 sequences); Molecular Functions – NAD(P)H oxidase (H2O2-forming) activity (200.0; p = 0.0001; 2 sequences), oxidoreductase activity acting on NAD(P)H with oxygen as acceptor (66.6; p = 0.00037; 2 sequences), and peroxidase activity (18.1; p = 0.00567; 2 sequences) (Figure 6).
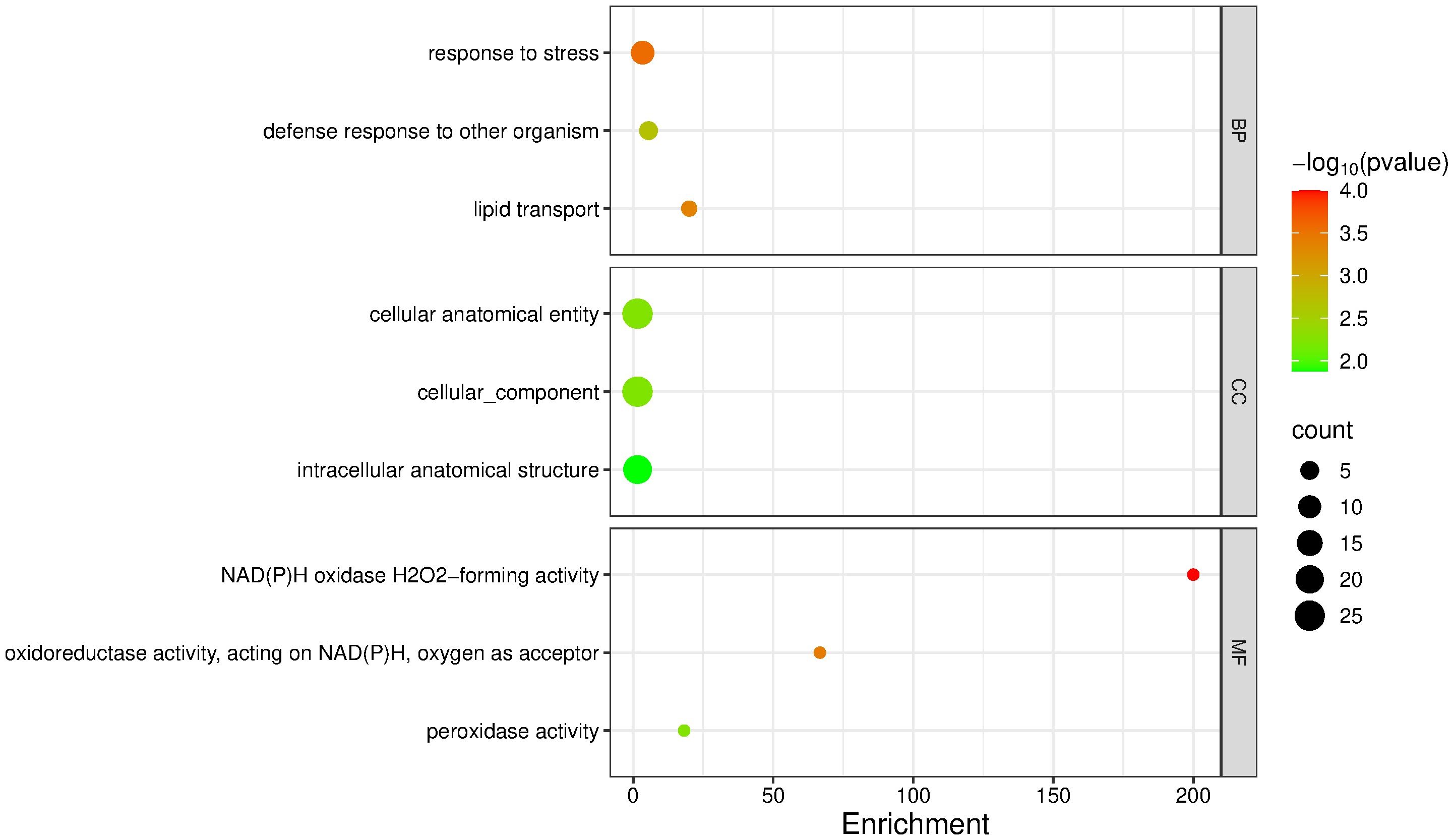
Figure 6. GO term enrichment analysis of downregulated genes in mango cv. Azúcar interacting with the endophyte C. tropicale at 12 hpi. Enrichment score (p < 0.05). Figure generated with SRPlot (Tang et al., 2023).
3.5 KEGG functional analysis of differentially expressed genes
The KEGG functional analysis revealed that upregulated genes at 12 hpi were mainly associated with plant defense and secondary metabolism pathways. The most represented pathways included biosynthesis of secondary metabolites (36 genes), phenylpropanoid biosynthesis (10), plant–pathogen interaction (9), MAPK signaling pathway in plants (8), and plant hormone signal transduction (6). Conversely, downregulated genes were associated with general metabolic pathways (3), biosynthesis of secondary metabolites (2), plant–pathogen interaction (2), phenylpropanoid biosynthesis (1), and MAPK signaling pathway in plants (1) (Figure 7).
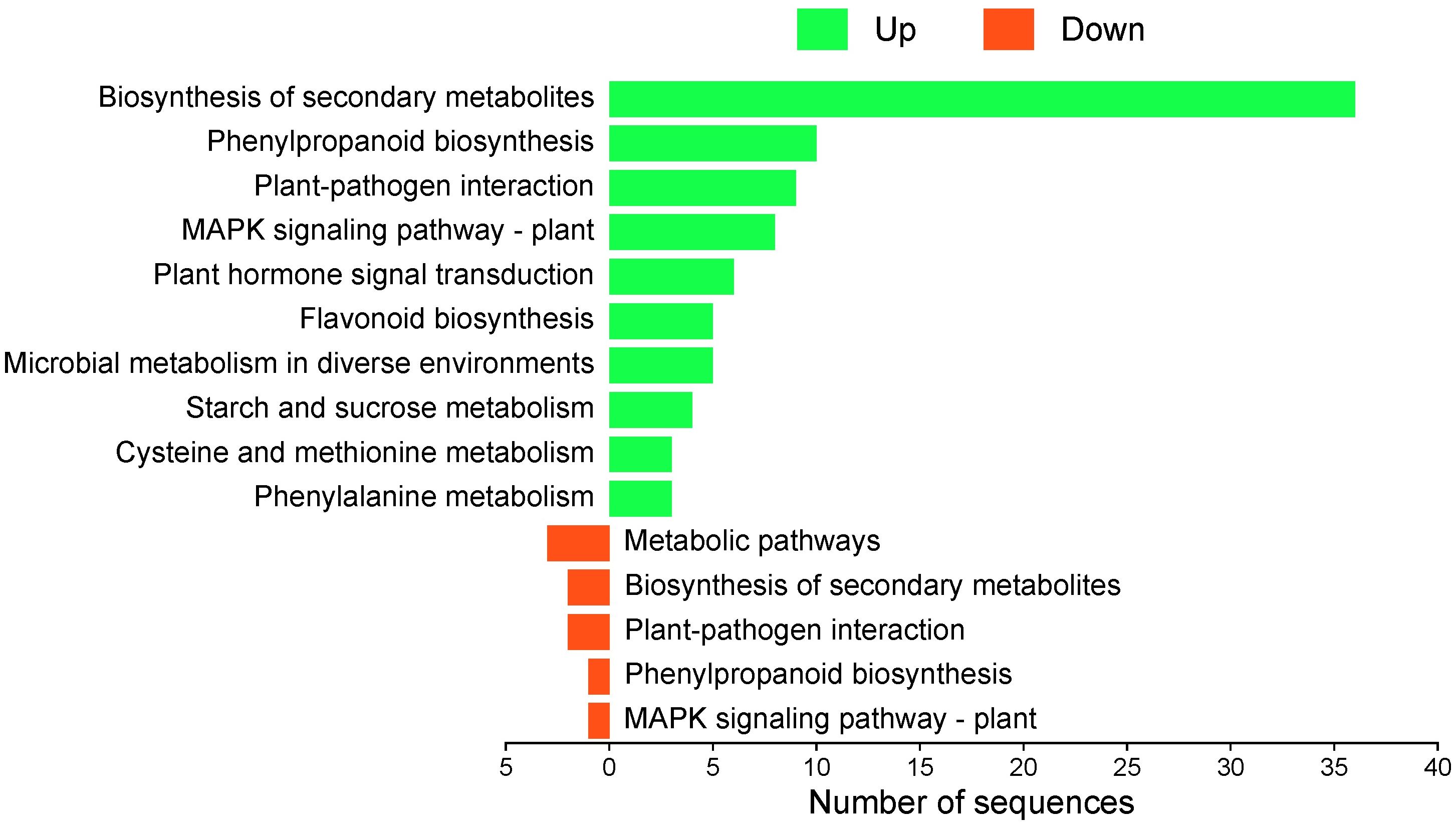
Figure 7. KEGG pathway enrichment analysis of differentially expressed genes in ripe mango fruit cv. Azúcar. Number of downregulated genes (red) and upregulated genes (green) at 12 hpi. Figure generated with SRPlot (Tang et al., 2023).
Significant KEGG pathways were mapped according to highly enriched genes at 12 hpi. Among the downregulated genes, most were involved in PAMP-triggered immunity (PTI) within the KEGG map ko04626 (plant–pathogen interaction). The most relevant were RbohC (respiratory burst oxidase homolog protein C), a key component in reactive oxygen species (ROS) production, and CaM/CML (calmodulin/calmodulin-like proteins), fundamental in calcium-dependent signaling and nitric oxide (NO) synthesis (Supplementary Figure 1). Repression of these pathways compromises the hypersensitive response (HR) (Figure 8, Supplementary Data 3).
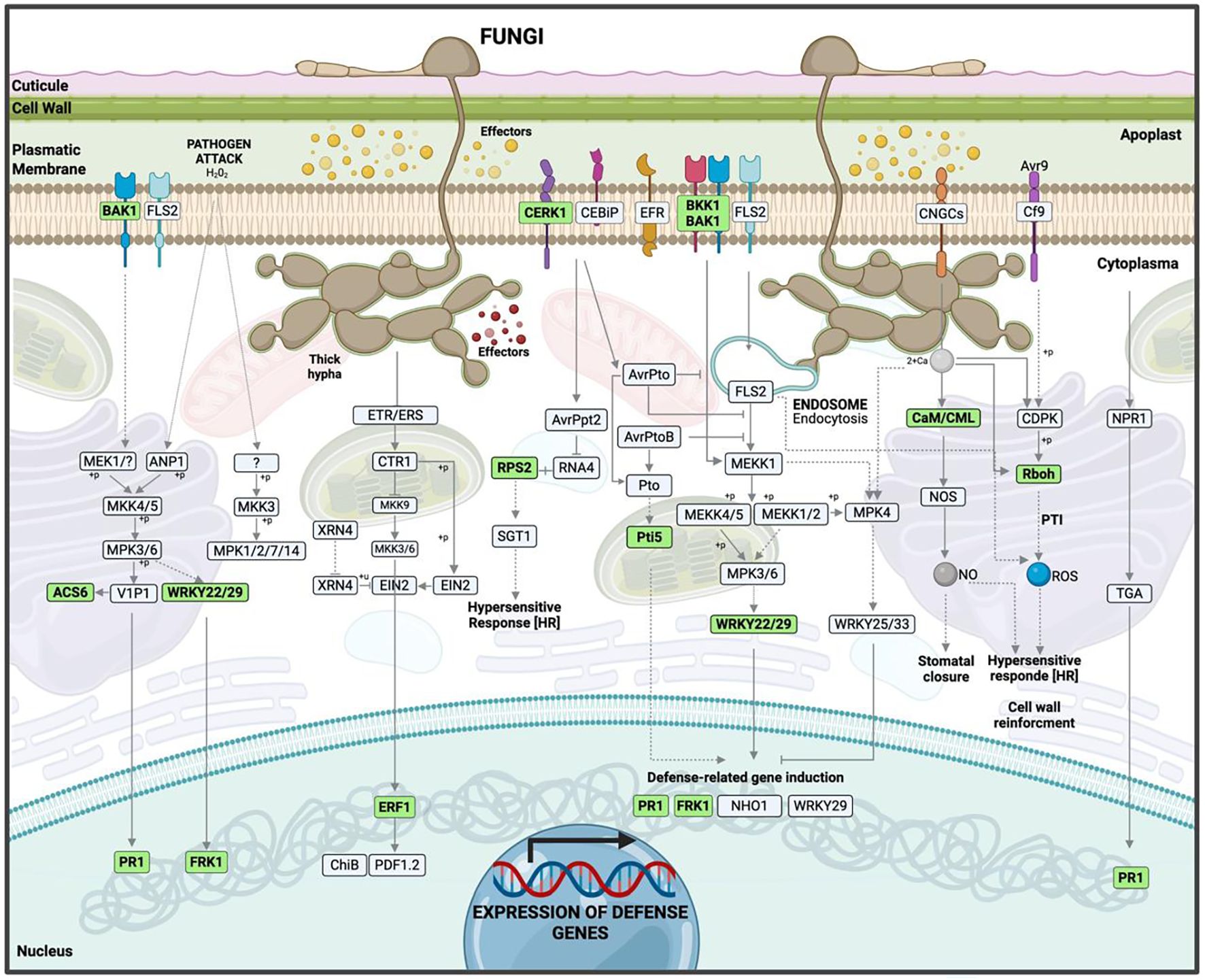
Figure 8. Hypothetical model of the interaction between ripe mango fruit cv. Azúcar and C. tropicale at 12 hpi, highlighting host upregulated pathways such as plant–pathogen interaction, plant hormone signal transduction, and MAPK signaling pathway – plant. Diagram created with BioRender.com.
For upregulated genes, the same KEGG map (ko04626) revealed activation of both PAMP-triggered immunity (PTI) and effector-triggered immunity (ETI). Receptor and co-receptor genes such as BAK1/BKK1 (brassinosteroid-insensitive 1-associated receptor kinase 1) and CERK1 (chitin elicitor receptor kinase 1) were identified, indicating fungal pattern recognition to initiate early defense signaling.
Calcium and redox signaling pathways were also observed, involving CaM/CML (calmodulin) and Rboh (respiratory burst oxidase). Transcription factor and defense-related genes were detected, including WRKY22/29, FRK1 (senescence-induced receptor-like serine/threonine-protein kinase), and PR1 (pathogenesis-related protein 1), which contribute to plant immunity by regulating defense gene transcription, phytoalexin accumulation, and microRNA production.
Specific resistance genes such as Pti5 (pathogenesis-related genes transcriptional activator PTI5) and RPS2 (disease resistance protein RPS2) were identified, associated with ETI (Supplementary Figure 2, Supplementary Data Sheet 3).
The phenylalanine metabolism pathway (ko04075: plant hormone signal transduction) was also identified among the upregulated genes, with PR1 activated, suggesting that PR1 expression is induced through salicylic acid (SA) biosynthesis to trigger a defense response (Supplementary Figure 3).
The MAPK signaling pathway (ko04016) revealed upregulated genes associated with pathogen perception and activation of the MAPK cascade (BAK1, BRI1-associated kinase 1); transcriptional responses through MAPK-activated factors (WRKY22/29); hormone biosynthesis such as ethylene (ACS6, 1-aminocyclopropane-1-carboxylate synthase 6; ERF1, ethylene response factor 1); expression of defense genes (FRK1, Flg22-induced receptor-like kinase 1; PR1, pathogenesis-related protein 1); and activation of ROS production (RbohD, respiratory burst oxidase homolog protein D) (Supplementary Figure 4, Figure 9).
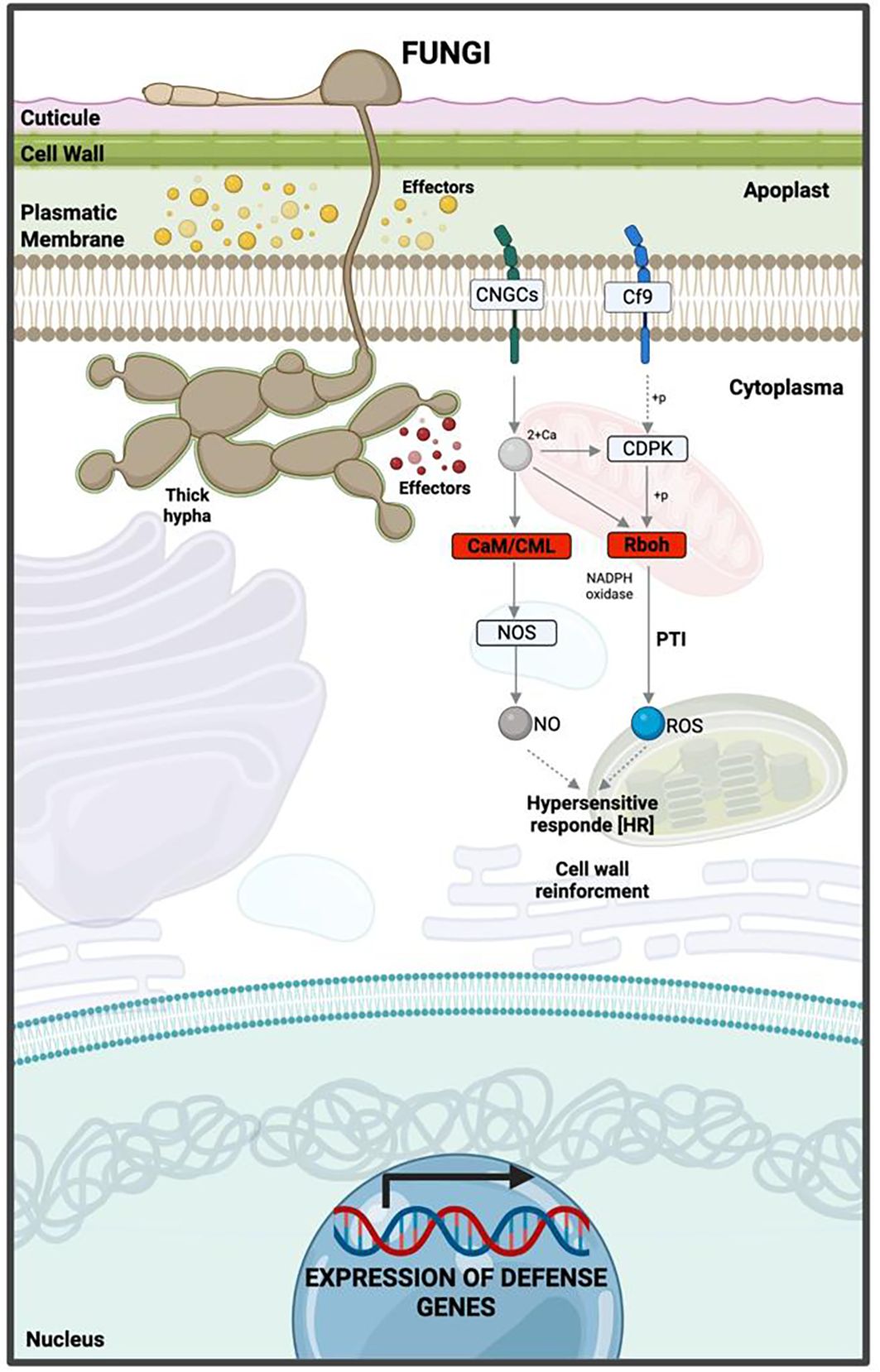
Figure 9. Hypothetical model of the interaction between ripe mango fruit cv. Azúcar and C. tropicale at 12 hpi, highlighting host downregulated pathways such as Rboh (respiratory burst oxidase homolog) and CaM/CML (calmodulin/calmodulin-like proteins). Diagram created with BioRender.com.
3.6 Validation of gene expression data in ripe mango cv. Azúcar by qRT-PCR
For qRT-PCR validation in ripe mango cv. Azúcar at 0, 12, and 24 hpi with C. tropicale, genes with the highest statistical significance (p < 0.05) were selected: one upregulated (LOX1: TRINITY_DN28581_c0_g1) and two downregulated (THI4: TRINITY_DN27837_c0_g1; MT3: TRINITY_DN25225_c1_g1).
LOX1 showed low expression at 0 hpi, which increased at 12 hpi (p < 0.05) and continued rising at 24 hpi (p < 0.001) (Figure 10). THI4 decreased slightly at 12 hpi and more strongly at 24 hpi (p < 0.001). MT3 displayed a progressive decrease beginning at 12 hpi and continued with marked significance (p < 0.001) at 24 hpi (Figure 10).
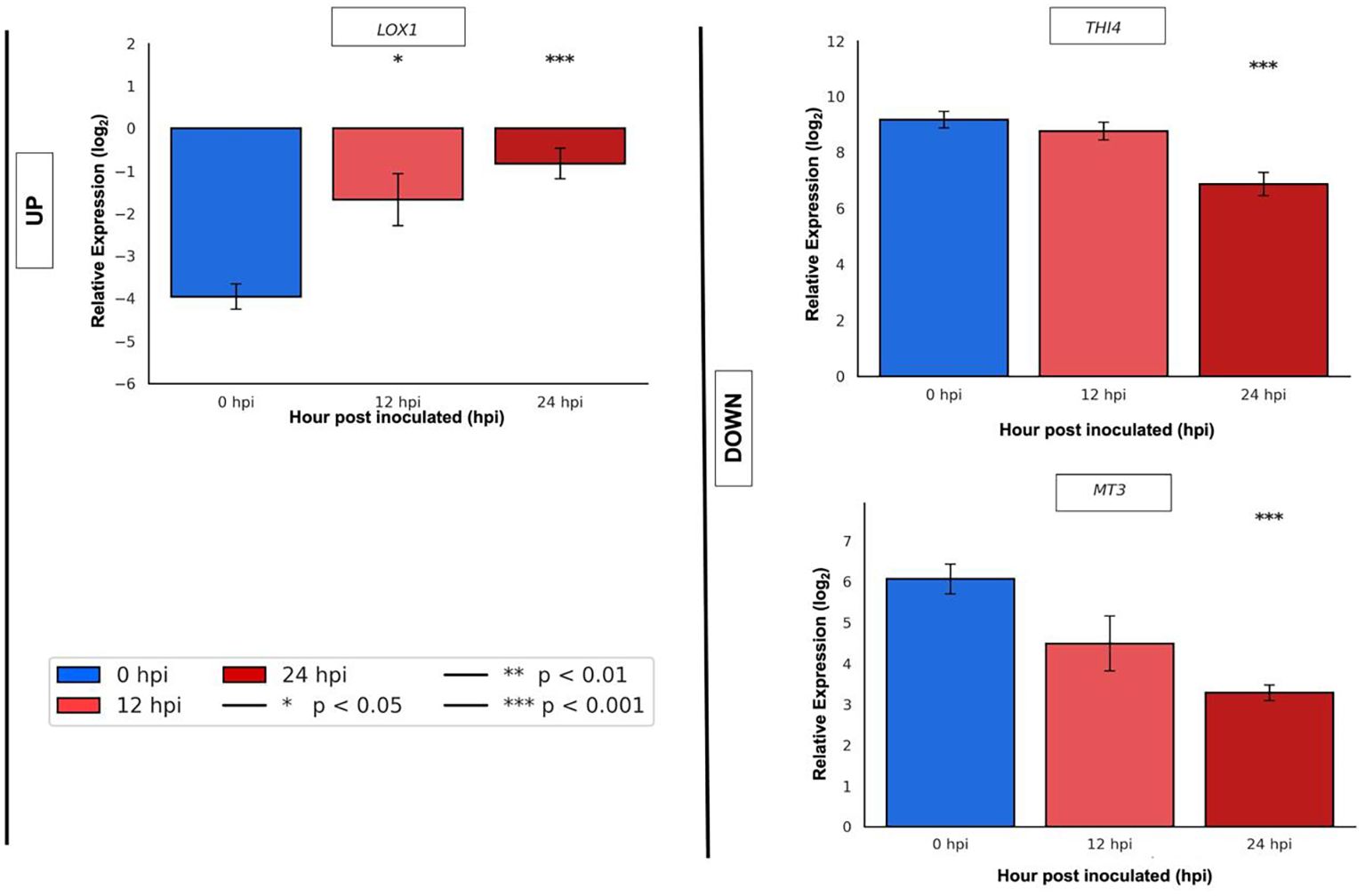
Figure 10. qRT-PCR validation of selected upregulated gene (LOX1: TRINITY_DN28581_c0_g1) and downregulated genes (THI4: TRINITY_DN27837_c0_g1; MT3: TRINITY_DN25225_c1_g1) in ripe mango fruit cv. Azúcar at 0, 12, and 24 hpi with the endophyte C. tropicale.
4 Discussion
The analysis of overexpressed genes in ripe mango fruit cv. Azúcar at 12 hpi reveals a significant activation of GO terms related to kinase activity, serine/threonine residue phosphorylation, calmodulin binding, and ubiquitin ligase activity. In other words, during the interaction with the endophyte C. tropicale, signaling pathways associated with pathogen perception, defense signal transduction, and protein regulation through the ubiquitin–proteasome system were activated. Regarding the role of protein kinases and receptors with carbohydrate-binding capacity, it is specified that their activation depends on pathogen-associated molecular patterns (PAMPs), leading to an initial defensive response. Likewise, the overexpressed genes associated with calmodulin and ubiquitin indicate their involvement in calcium-dependent signaling pathways and protein remodeling. This suggests that the ripe fruit perceives the presence of the fungus, triggering a robust defense response that does not ultimately result in the complete elimination of the pathogen.
The transcriptomic study conducted by Alkan et al. (2015) on the interaction between green/ripe tomato fruit and C. gloeosporioides reported host genes overexpressed during the quiescent phase of the fungus, with functions related to kinase activity, calmodulin binding, and ubiquitin ligases. Moreover, the pathogen was shown to alkalinize plant tissue to stimulate signaling pathways dependent on calcium, reactive oxygen species (ROS), and serine/threonine kinases.
For the repressed genes in ripe mango fruit cv. Azúcar, the GO results suggest that at 12 hpi, corresponding to the biotrophic stage of C. tropicale, the fungus impacts molecular functions associated with redox homeostasis, including NADPH oxidase involved in hydrogen peroxide production, peroxidases, antioxidant activities, and oxidoreductase enzymes. These functions are critical for the generation of ROS, which are essential for the activation of the host immune response, such as the hypersensitive response (HR).
Reactive oxygen species (ROS), including singlet oxygen (¹O2), superoxide anion (O2-), hydroxyl radical (OH•), nitric oxide (•NO), and hydrogen peroxide (H2O2) (Groß et al., 2013; Shetty et al., 2008; Wang et al., 2010), are released into the apoplast to establish the oxidative burst, which constitutes a plant defensive reaction in response to pathogen attack (Kámán-Tóth et al., 2019; Marino et al., 2012). For ROS generation, plant cells employ enzymatic activities related to class III peroxidases, NADPH oxidases, amine and polyamine oxidases (AOs and PAOs), respiratory burst oxidase homologs (RBOHs), glycolate oxidase, oxalate oxidase, xanthine oxidase, photosynthesis, photorespiration, and respiration (Baxter et al., 2014; Heller and Tudzynski, 2011; Kärkönen and Kuchitsu, 2015; Marino et al., 2012). Oxidative stress occurs when there is an imbalance between ROS production and antioxidant activity, which can lead to severe cellular damage and ultimately programmed cell death. Therefore, enzymes such as peroxidases, among others, are required to maintain system homeostasis through detoxification (dos Santos and Franco, 2023; Hasanuzzaman et al., 2017).
Macho and Zipfel (2015) reported that cytoplasmic effectors can silence RBOH enzymes and, consequently, inhibit ROS production. Similarly, Singh et al. (2016) suggested that these effectors can suppress RBOH-mediated pathways, whose function is to generate robust ROS production. Jwa and Hwang (2017) indicated that apoplastic effectors can eliminate ROS from the extracellular space to reduce pathogen damage and promote intracellular infection, through the silencing of PAMP recognition receptors (PRRs), which in turn inactivate NADPH oxidase-dependent RBOH pathways. Segal and Wilson (2018) reported that pathogens can eliminate host ROS to suppress its defensive response. Through RNA-seq studies, Alkan et al. (2015) identified that Colletotrichum gloeosporioides, to accomplish its lifestyle transition from quiescent to necrotrophic in green tomato fruit, can modulate the host redox environment by inducing alkalinization and delaying ROS activation. Studies conducted by Zhang et al. (2021) described that the hemibiotroph C. orbiculare produces the specific effector SIB1, which suppresses the oxidative burst or ROS production in Nicotiana benthamiana, with its highest expression observed at 12 hpi (during appressorium formation or the early infection stage). The oxidative burst in plants, as a defense strategy induced by the release of ROS and mediated by H2O2 during the hemibiotrophic stage of fungi, can be reduced at this stage and subsequently increase during the transition to the necrotrophic lifestyle. Eloy et al. (2015), in the cowpea (Vigna unguiculata)–C. gloeosporioides pathosystem, demonstrated that changes in H2O2 levels modulate fungal lifestyle transitions, with the necrotrophic stage being favored by the increase in ROS. The effectors ChEC3, ChEC3a, ChEC5, ChEC6, and ChEC34 from C. higginsianum reported by Kleemann et al. (2012), as well as CgDN3 from C. orbiculare described by Yoshino et al. (2012), were shown to be overexpressed and to suppress cell death in N. benthamiana leaves during the biotrophic phase. Gao et al. (2024) identified the CfMsg5 gene of C. fructicola during its interaction with Camellia oleifera through RNA-seq analysis, which contributes to ROS detoxification, regulation of the MAPK cell wall integrity (CWI) pathway, and unfolded protein response (UPR), thereby promoting its pathogenicity.
In this context, the endophyte C. tropicale, which behaves as a hemibiotroph, can repress oxidative burst pathways associated with defense in ripe mango fruit cv. Azúcar, thereby facilitating its establishment during the quiescent or biotrophic phase. This repression could lead to oxidative stress and ROS overaccumulation, which may subsequently promote cell death and, consequently, the necrotrophic phase of C. tropicale, consistent with its hemibiotrophic lifestyle.
These results, concerning the overexpressed genes in mango cv. Azúcar, are consistent with the findings of Plett and Martin (2018), who reported that hemibiotrophs can activate plant defense responses through pathways related to PR proteins, ethylene, salicylic acid (SA), and ROS, leading to a hypersensitive response (Figure 10). In the case of ethylene, it has been described that Ethylene signaling increases during the quiescent phase, along with defense responses, and may even induce release from this same phase (Alkan et al., 2015). Hong et al. (2016), through RNA-seq studies on the mango fruit cv. Zill–C. gloeosporioides interaction, reported the role of overexpressed PR and ERF genes as positive regulators of host resistance against fungal infection. Our study reveals that ripe mango fruit cv. Azúcar is capable of overexpressing genes associated with ethylene and salicylic acid pathways.
Prusky et al. (2013) stated that ripe fruit reduces its antifungal defense against quiescent fungi, which induce high levels of ethylene and salicylic acid, thereby promoting programmed cell death. This is consistent with our study, which reveals that ripe mango fruit cv. Azúcar activates defense pathways mediated by ethylene and salicylic acid in response to infection by the endophyte C. tropicale.
LOX genes (linoleate:oxygen oxidoreductase; lipoxygenases) belong to the family of fatty acid or lipid substrate dioxygenases (Feussner and Wasternack, 2002). In plants, they are key enzymes that synthesize oxylipins, which act as signaling molecules during plant–pathogen interactions, leading to the production of C6 volatiles and jasmonates (Viswanath et al., 2020). Studies by Peng et al. (1994) demonstrated that LOX1 is induced early in rice as a response to infection by Magnaporthe grisea. Similarly, pepper leaves inoculated with Xanthomonas campestris and Colletotrichum coccodes showed high expression levels of the CaLOX gene, and in Arabidopsis, overexpression of the same gene conferred increased resistance to infection by P. syringae, Hyaloperonospora arabidopsidis, and Alternaria brassicicola (Hwang and Hwang, 2010). Hou et al. (2018) determined the role of the DkLOX3 gene in Arabidopsis, where its overexpression enhanced resistance to P. syringae and Botrytis cinerea, as evidenced by greater ROS accumulation followed by cell death. Rustérucci et al. (1999) proposed that the synthesis of fatty acid hydroperoxides and volatile products with signaling roles act as precursors for LOX genes to mount pathogen defense. Likewise, these genes have been shown to confer antimicrobial activity (Croft et al., 1993; Weber et al., 1999). LOX-derived products also participate in hypersensitive response (HR) processes that enable the plant to contain pathogen infection and thereby establish a defense mechanism (Viswanath et al., 2020).
Joshi et al. (2020) reported that plants can use THI4 (thiazole synthase), which leads to thiamine biosynthesis. Among the main functions of thiamine (vitamin B1) in plants under stress conditions, a prominent role is its ability to trigger ROS release and subsequently induce resistance (Li et al., 2016; Yin et al., 2022). Yang et al. (2024) demonstrated that the thiazole moiety of thiamine (THI2 = TaTHI2) plays an important role in plant immunity against Chinese wheat mosaic virus (CWMV), as it activates ROS production and enhances resistance in wheat.
Metallothioneins (MTs) are characterized as low molecular weight (6–7 kDa) metal-binding proteins with high cysteine content (~30%) (Li et al., 2023). In plants, MT genes such as MT3 are expressed in ripe fruits (Yang et al., 2015), and among their main functions is the regulation or neutralization of reactive oxygen species (ROS), which cause DNA damage (Akashi et al., 2004; Chubatsu and Meneghini, 1993; Miura et al., 1997; Wong et al., 2004). Dauch and Jabaji-Hare (2006) provided the first report of an MT gene (MT3) induced in Abutilon theophrasti plants in response to Colletotrichum coccodes infection, where the gene expression decreased 5 days after fungal inoculation, coinciding with the appearance of the first disease symptoms.
These results confirm that the endophyte C. tropicale, at early stages of its pathogenic process (hemibiotrophic with quiescence) at 12 hpi, is able to modulate defense response genes in ripe mango fruit cv. Azúcar—upregulating genes such as LOX1, which participates in the hypersensitive response (HR), and downregulating genes involved in primary metabolism and antioxidant homeostasis, such as THI4 and MT3. This modulation could lead to oxidative stress that, in turn, favors the necrotrophic phase of the endophyte, as observed at 24 hpi, where the increased expression of LOX1 may contribute to enhanced ROS release accompanied by cell death, while the decreased expression of MT3 would reduce its capacity to scavenge ROS and to provide cellular protection.
5 Conclusions
Based on the RNA-seq study of the interaction between ripe mango fruit cv. Azúcar and the endophyte C. tropicale, 5,435 differentially expressed genes were identified from both the host and the fungus at 12 hpi. The main findings include: (1) repression of genes in ripe mango fruit associated with molecular functions involved in the plant’s defensive oxidative burst mediated by ROS (NADPH oxidase with hydrogen peroxide production, peroxidases, antioxidant activities, and oxidoreductase enzymes), possibly as an effect of colonization by the endophyte C. tropicale to establish its initial infective stage (quiescence) as a hemibiotroph; (2) validation of repressed genes in ripe fruit, THI4 and MT3, with functions related to ROS release and ROS scavenging, respectively; (3) activation of signaling pathways in ripe fruit linked to pathogen perception, defense signal transduction, and protein regulation via the ubiquitin–proteasome system.
Altogether, these findings lead to the conclusion that ripe mango fruit cv. Azúcar initiates a defense response against the endophyte C. tropicale that does not fully overcome its initial pathogenic process at the cellular level through quiescence but rather creates favorable conditions for its establishment by suppressing oxidative burst pathways, which subsequently promote oxidative stress and facilitate the transition to its necrotrophic phase.
Thus, this study reveals the gene expression patterns that regulate the lifestyle switch of an endophyte such as C. tropicale in ripe mango fruit cv. Azúcar, based on its pathogenic strategy as a hemibiotroph with quiescence. The host transcriptional profile revealed in this study could be used to propose management strategies for anthracnose during the postharvest stage of ripe mango fruits cv. Azúcar, focusing on the external induction of defense responses and on the regulation of redox balance through antioxidants that counteract oxidative stress, thereby preventing the activation of the necrotrophic phase of the fungus.
Data availability statement
The RNA sequencing data have been deposited in the Sequence Read Archive (SRA) of the National Center for Biotechnology Information (NCBI), with accession number PRJNA1337713.
Author contributions
AQ: Writing – original draft, Writing – review & editing, Methodology, Visualization. SR: Validation, Writing – original draft, Investigation. YR: Writing – original draft, Investigation. LL: Methodology, Investigation, Writing – original draft. MS: Investigation, Writing – review & editing, Methodology, Writing – original draft, Supervision. CG: Methodology, Supervision, Writing – original draft, Writing – review & editing, Investigation.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This work was partially supported by Programa de Apoyo a Proyectos de Investigación e Innovación Tecnológica (DGAPA-PAPIIT) grant No. N203023 to MS and the Facultad de Ciencias Agrarias of the Universidad Nacional de Colombia sede Bogotá for its partial support in the publication of this article.
Acknowledgments
We thank Michael F. Dunn for the critical reading and comments on the manuscript. The authors would like to thank the laboratories at the Universidad Nacional de Colombia, Bogotá campus: Biotecnología Vegetal (Facultad de Ciencias Agrarias) and Biología Molecular (Departamento de Biología); as well as the TANDEM Max Planck group, Genómica Evolutiva del Metabolismo Especializado (GEME), for providing the facilities and infrastructure required for carrying out the experiments.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ffunb.2025.1699983/full#supplementary-material
Supplementary Data Sheet 1 | DEGs_Mango_cv_Azúcar_T0vsT12.
Supplementary Data Sheet 2 | GO_Mango_cv_Azúcar_UP_DOWN.
Supplementary Data Sheet 3 | KEGG_Mango_cv_Azúcar.
References
Akashi K., Nishimura N., Ishida Y., and Yokota A. (2004). Potent hydroxyl radical-scavenging activity of drought-induced type-2 metallothionein in wild watermelon. Biochem. Biophys. Res. Commun. 323, 72–78. doi: 10.1016/j.bbrc.2004.08.056
Alkan N., Friedlander G., Ment D., Prusky D., and Fluhr R. (2015). Simultaneous transcriptome analysis of Colletotrichum gloeosporioides and tomato fruit pathosystem reveals novel fungal pathogenicity and fruit defense strategies. New Phytol. 205, 801–815. doi: 10.1111/nph.13087
Altschul S. F., Gish W., Miller W., Myers E. W., and Lipman D. J. (1990). Basic local alignment search tool. J. Mol. Biol. 215, 403–410. doi: 10.1016/S0022-2836(05)80360-2
Arauz L. F. (2000). Mango anthracnose: economic impact and current options for integrated managaement. Plant Dis. 84, 600–611. doi: 10.1094/PDIS.2000.84.6.600
Arcila Cardona Á. M., Castillo Urquiza G. P., Pérez Artiles L., Abaunza González C. A., Yacomelo Hernández M. J., and León Pacheco R. I. (2022). Modelo productivo de mango de azúcar (Mangifera indica L.) para el departamento del Magdalena (Mosquera, Colombia: Corporación Colombiana de investigación agropecuaria - AGROSAVIA). doi: 10.21930/agrosavia.model.7405170
Baxter A., Mittler R., and Suzuki N. (2014). ROS as key players in plant stress signalling. J. Exp. Bot. 65, 1229–1240. doi: 10.1093/jxb/ert375
Chubatsu L. S. and Meneghini R. (1993). Metallothionein protects DNA from oxidative damage. Biochem. J. 291, 193–198. doi: 10.1042/bj2910193
Croft K. P. C., Juttner F., and Slusarenko A. J. (1993). Volatile Products of the Lipoxygenase Pathway Evolved from Phaseolus vulgaris (L.) Leaves Inoculated with Pseudomonas syringae pv phaseolicola. Plant Physiol. 101, 13–24. doi: 10.1104/pp.101.1.13
Crouch J., O’Connell R., Gan P., Buiate E., Torres M. F., Beirn L., et al. (2014). “The genomics of Colletotrichum,” in Genomics of Plant-Associated Fungi: Monocot Pathogens (Springer Berlin Heidelberg, Berlin, Heidelberg). doi: 10.1007/978-3-662-44053-7_3
da Silva L. L., Moreno H. L. A., Correia H. L. N., Santana M. F., and de Queiroz M. V. (2020). Colletotrichum: species complexes, lifestyle, and peculiarities of some sources of genetic variability. Appl. Microbiol. Biotechnol. 104, 1891–1904. doi: 10.1007/s00253-020-10363-y
Dauch A. L. and Jabaji-Hare S. H. (2006). Metallothionein and bZIP Transcription Factor Genes from Velvetleaf and Their Differential Expression Following Colletotrichum coccodes Infection. Phytopathology 96, 1116–1123. doi: 10.1094/PHYTO-96-1116
Delaye L., García-Guzmán G., and Heil M. (2013). Endophytes versus biotrophic and necrotrophic pathogens-are fungal lifestyles evolutionarily stable traits? Fungal Divers. 60, 125–135. doi: 10.1007/s13225-013-0240-y
De Silva D. D., Crous P. W., Ades P. K., Hyde K. D., and Taylor P. W. J. (2017). Life styles of Colletotrichum species and implications for plant biosecurity. Fungal Biol. Rev. 31. doi: 10.1016/j.fbr.2017.05.001
de Souza A., Delphino Carboni R. C., Wickert E., de Macedo Lemos E. G., and de Goes A. (2013). Lack of host specificity of Colletotrichum spp. isolates associated with anthracnose symptoms on mango in Brazil. Plant Pathol. 62, 1038–1047. doi: 10.1111/ppa.12021
Dinkins R. D., Nagabhyru P., Graham M. A., Boykin D., and Schardl C. L. (2017). Transcriptome response of Lolium arundinaceum to its fungal endophyte Epichloë coenophiala. New Phytol. 213, 324–337. doi: 10.1111/nph.14103
dos Santos C. and Franco O. L. (2023). Pathogenesis-related proteins (PRs) with enzyme activity activating plant defense responses. Plants 12, 2226. doi: 10.3390/plants12112226
Eaton C. J., Cox M. P., and Scott B. (2011). What triggers grass endophytes to switch from mutualism to pathogenism? Plant Sci. 180, 190–195. doi: 10.1016/j.plantsci.2010.10.002
Eloy Y. R. G., Vasconcelos I. M., Barreto A. L. H., Freire-Filho F. R., and Oliveira J. T. A. (2015). H2O2 plays an important role in the lifestyle of Colletotrichum gloeosporioides during interaction with cowpea [Vigna unguiculata (L.) Walp. Fungal Biol. 119, 747–757. doi: 10.1016/j.funbio.2015.05.001
Feussner I. and Wasternack C. (2002). The lipoxygenase pathway. Annu. Rev. Plant Biol. 53, 275–297. doi: 10.1146/annurev.arplant.53.100301.135248
Freeman S., Horowitz S., and Sharon A. (2001). Pathogenic and nonpathogenic lifestyles in Colletotrichum acutatum from strawberry and other plants. Phytopathology 91. doi: 10.1094/PHYTO.2001.91.10.986
Gao Y., Zhang S., Sheng S., and Li H. (2024). A Colletotrichum fructicola dual specificity phosphatase CfMsg5 is regulated by the CfAp1 transcription factor during oxidative stress and promotes virulence on Camellia oleifera. Virulence 15. doi: 10.1080/21505594.2024.2413851
García Lozano J., Gómez Barros G., de J., Forero Longas F., Sandoval A., Vásquez L., et al. (2010). Descripción de las variedades de mango criollo Colombiano: boletín técnico (Corporación Colombiana de Investigación Agropecuaria). Available online at: http://hdl.handle.net/20.500.12324/1239 (Accessed January 20, 2025).
Götz S., Garcia-Gomez J. M., Terol J., Williams T. D., Nagaraj S. H., Nueda M. J., et al. (2008). High-throughput functional annotation and data mining with the Blast2GO suite. Nucleic Acids Res. 36, 3420–3435. doi: 10.1093/nar/gkn176
Grabherr M. G., Haas B. J., Yassour M., Levin J. Z., Thompson D. A., Amit I., et al. (2011). Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 29, 644–652. doi: 10.1038/nbt.1883
Groß F., Durner J., and Gaupels F. (2013). Nitric oxide, antioxidants and prooxidants in plant defence responses. Front. Plant Sci. 4. doi: 10.3389/fpls.2013.00419
Hasanuzzaman M., Nahar K., Hossain Md., Mahmud J., Rahman A., Inafuku M., et al. (2017). Coordinated actions of glyoxalase and antioxidant defense systems in conferring abiotic stress tolerance in plants. Int. J. Mol. Sci. 18, 200. doi: 10.3390/ijms18010200
Heller J. and Tudzynski P. (2011). Reactive oxygen species in phytopathogenic fungi: signaling, development, and disease. Annu. Rev. Phytopathol. 49, 369–390. doi: 10.1146/annurev-phyto-072910-095355
Hong K., Gong D., Zhang L., Hu H., Jia Z., Gu H., et al. (2016). Transcriptome characterization and expression profiles of the related defense genes in postharvest mango fruit against Colletotrichum gloeosporioides. Gene 576, 275–283. doi: 10.1016/j.gene.2015.10.041
Hou Y., Ban Q., Meng K., He Y., Han S., Jin M., et al. (2018). Overexpression of persimmon 9-lipoxygenase DkLOX3 confers resistance to Pseudomonas syringae pv. tomato DC3000 and Botrytis cinerea in Arabidopsis. Plant Growth Regul. 84, 179–189. doi: 10.1007/s10725-017-0331-y
Hwang I. S. and Hwang B. K. (2010). The pepper 9-lipoxygenase gene caLOX1 functions in defense and cell death responses to microbial pathogens. Plant Physiol. 152, 948–967. doi: 10.1104/pp.109.147827
Joshi J., Beaudoin G. A. W., Patterson J. A., García-García J. D., Belisle C. E., Chang L.-Y., et al. (2020). Bioinformatic and experimental evidence for suicidal and catalytic plant THI4s. Biochem. J. 477, 2055–2069. doi: 10.1042/BCJ20200297
Jwa N.-S. and Hwang B. K. (2017). Convergent evolution of pathogen effectors toward reactive oxygen species signaling networks in plants. Front. Plant Sci. 8. doi: 10.3389/fpls.2017.01687
Kámán-Tóth E., Dankó T., Gullner G., Bozsó Z., Palkovics L., and Pogány M. (2019). Contribution of cell wall peroxidase- and NADPH oxidase-derived reactive oxygen species to Alternaria brassicicol -induced oxidative burst in Arabidopsis. Mol. Plant Pathol. 20, 485–499. doi: 10.1111/mpp.12769
Kamle M. (2013). A species-specific PCR based assay for rapid detection of mango anthracnose pathogen Colletotrichum gloeosporioides Penz. and Sacc. J. Plant Pathol. Microbiol. 04. doi: 10.4172/2157-7471.1000184
Kanehisa M. (2004). The KEGG resource for deciphering the genome. Nucleic Acids Res. 32, 277D–2280. doi: 10.1093/nar/gkh063
Kärkönen A. and Kuchitsu K. (2015). Reactive oxygen species in cell wall metabolism and development in plants. Phytochemistry 112, 22–32. doi: 10.1016/j.phytochem.2014.09.016
Kaul S., Sharma T., and Dhar M. K. (2016). Omics” tools for better understanding the plant–endophyte interactions. Front. Plant Sci. 7. doi: 10.3389/fpls.2016.00955
Khare E., Mishra J., and Arora N. K. (2018). Multifaceted interactions between endophytes and plant: Developments and Prospects. Front. Microbiol. 9. doi: 10.3389/fmicb.2018.02732
Kleemann J., Rincon-Rivera L. J., Takahara H., Neumann U., van Themaat E. V. L., van der Does H. C., et al. (2012). Sequential delivery of host-induced virulence effectors by appressoria and intracellular hyphae of the phytopathogen Colletotrichum higginsianum. PLoS Pathog. 8, e1002643. doi: 10.1371/journal.ppat.1002643
Kogel K. H., Franken P., and Hückelhoven R. (2006). Endophyte or parasite - what decides? Curr. Opin. Plant Biol. 9, 358–363. doi: 10.1016/j.pbi.2006.05.001
Langmead B. and Salzberg S. L. (2012). Fast gapped-read alignment with Bowtie 2. Nat. Methods 9, 357–359. doi: 10.1038/nmeth.1923
Ledesma N. and Campbell R. J. (2019). The status of mango cultivars, market perspectives and mango cultivar improvement for the future. Acta Hortic. 23–28. doi: 10.17660/ActaHortic.2019.1244.3
Li B. and Dewey C. N. (2011). RSEM: accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinf. 12, 323. doi: 10.1186/1471-2105-12-323
Li Y., Lee S. H., Piao M., Kim H. S., and Lee K. Y. (2023). Metallothionein 3 inhibits 3T3-L1 adipocyte differentiation via reduction of reactive oxygen species. Antioxidants 12, 640. doi: 10.3390/antiox12030640
Li C.-L., Wang M., Wu X.-M., Chen D.-H., Lv H.-J., Shen J.-L., et al. (2016). THI1, a thiamine thiazole synthase, interacts with Ca2+-dependent protein kinase CPK33 and modulates the S-type anion channels and stomatal closure in Arabidopsis. Plant Physiol. 170, 1090–1104. doi: 10.1104/pp.15.01649
Love M. I., Huber W., and Anders S. (2014). Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550. doi: 10.1186/s13059-014-0550-8
Macho A. P. and Zipfel C. (2015). Targeting of plant pattern recognition receptor-triggered immunity by bacterial type-III secretion system effectors. Curr. Opin. Microbiol. 23, 14–22. doi: 10.1016/j.mib.2014.10.009
Maldonado-Celis M. E., Yahia E. M., Bedoya R., Landázuri P., Loango N., Aguillón J., et al. (2019). Chemical Composition of Mango (Mangifera indica L.) Fruit: Nutritional and Phytochemical Compounds. Front Plant Sci 10. doi: 10.3389/fpls.2019.01073
Marino D., Dunand C., Puppo A., and Pauly N. (2012). A burst of plant NADPH oxidases. Trends Plant Sci. 17, 9–15. doi: 10.1016/j.tplants.2011.10.001
Ministerio de Agricultura y Desarrollo Rural (2025). Base agrícola eva 2020 (Biblioteca Digital Agronet). Available online at: https://www.agronet.gov.co/estadistica/paginas/home.aspx?cod=59 (Accessed April 13, 2025).
Miura T., Muraoka S., and Ogiso T. (1997). Antioxidant activity of metallothionein compared with reduced glutathione. Life Sci. 60, 301–309. doi: 10.1016/S0024-3205(97)00156-2
Páez A. (2003). Tecnologías sostenibles para el manejo de la Antracnosis en papaya y mango. [Boletín técnico n.o 8] (Corpoica: Corporación Colombiana de Investigación Agropecuaria), 1–18. Corpoica. Available online at: https://bibliotecadigital.agronet.gov.co/handle/11348/6459 (Accessed June 17, 2025).
Páez-Redondo A. R., García-Merchán V. H., Rincón J. D., Rodríguez H. A., Morales J. G., and Hoyos-Carvajal L. M. (2024). Colletotrichum species associated with anthracnose disease on mango (Mangifera indica L.) cv. Azúcar in Colombia. Can. J. Plant Pathol. 46, 616–631. doi: 10.1080/07060661.2024.2402719
Peng Y. L., Shirano Y., Ohta H., Hibino T., Tanaka K., and Shibata D. (1994). A novel lipoxygenase from rice. Primary structure and specific expression upon incompatible infection with rice blast fungus. J. Biol. Chem. 269, 3755–3761. doi: 10.1016/S0021-9258(17)41924-7
Plett J. M. and Martin F. M. (2018). Know your enemy, embrace your friend: using omics to understand how plants respond differently to pathogenic and mutualistic microorganisms. Plant J. 93, 729–746. doi: 10.1111/tpj.13802
Ploetz R. C. (2007). Diseases of tropical perennial crops: challenging problems in diverse environments. Plant Dis. 91, 644–663. doi: 10.1094/PDIS-91-6-0644
Prusky D., Alkan N., Mengiste T., and Fluhr R. (2013). Quiescent and necrotrophic lifestyle choice during postharvest disease development. Annu. Rev. Phytopathol. 51, 155–176. doi: 10.1146/annurev-phyto-082712-102349
Quintero-Mercado A. F. (2025). Identificación de la expresión génica en mango (Mangifera indica L. cv. Azúcar) que favorece la patogenicidad del endófito Colletotrichum tropicale en frutos. Universidad Nacional de Colombia, Bogotá D.C.
Quintero-Mercado A., Dangon-Bernier F., and Páez-Redondo A. (2019). Aislamientos endofíticos de Colletotrichum spp. a partir de hojas y ramas de mango (Mangifera indica L.) cultivar Azúcar en el municipio de Ciénaga, Magdalena, Colombia. Rev. Acad. Colomb Cienc Exactas Fis Nat. 43. doi: 10.18257/raccefyn.788
Rajan S., Tiwari D., Singh V. K., Saxena P., Reddy Y. T. N., Singh S., et al. (2011). Application of extended BBCH Scale for phenological studies in mango (Mangifera indica L.). J. Appl. Horticulture 13, 108–114. doi: 10.37855/jah.2011.v13i02.25
Rustérucci C., Montillet J.-L., Agnel J.-P., Battesti C., Alonso B., Knoll A., et al. (1999). Involvement of lipoxygenase-dependent production of fatty acid hydroperoxides in the development of the hypersensitive cell death induced by cryptogein on tobacco leaves. J. Biol. Chem. 274, 36446–36455. doi: 10.1074/jbc.274.51.36446
Salcedo R. (2018). Revista de la asociación hortifrutícola de Colombia, ASOHOFRUCOL- FNFH. No. 57. Magdalena fortalece su potencial exportador frutas y hortalizas 27.
Sánchez-Vallet A., Mesters J. R., and Thomma B. P. H. J. (2015). The battle for chitin recognition in plant-microbe interactions. FEMS Microbiol. Rev. 39, 171–183. doi: 10.1093/femsre/fuu003
Segal L. M. and Wilson R. A. (2018). Reactive oxygen species metabolism and plant-fungal interactions. Fungal Genet. Biol. 110, 1–9. doi: 10.1016/j.fgb.2017.12.003
Shetty N. P., Jørgensen H. J. L., Jensen J. D., Collinge D. B., and Shetty H. S. (2008). Roles of reactive oxygen species in interactions between plants and pathogens. Eur. J. Plant Pathol. 121, 267–280. doi: 10.1007/s10658-008-9302-5
Singh R., Dangol S., Chen Y., Choi J., Cho Y.-S., Lee J.-E., et al. (2016). Magnaporthe oryzae effector AVR-pii helps to establish compatibility by inhibition of the rice NADP-malic enzyme resulting in disruption of oxidative burst and host innate immunity. Mol. Cells 39, 426–438. doi: 10.14348/molcells.2016.0094
Tang D., Chen M., Huang X., Zhang G., Zeng L., Zhang G., et al. (2023). SRplot: A free online platform for data visualization and graphing. PLoS One 18, e0294236. doi: 10.1371/journal.pone.0294236
Untergasser A., Cutcutache I., Koressaar T., Ye J., Faircloth B. C., Remm M., et al. (2012). Primer3—new capabilities and interfaces. Nucleic Acids Res. 40, e115–e115. doi: 10.1093/nar/gks596
Vieira W. A. S., Michereff S. J., de Morais M. A., Hyde K. D., and Câmara M. P. S. (2014). Endophytic species of Colletotrichum associated with mango in northeastern Brazil. Fungal Divers. 67. doi: 10.1007/s13225-014-0293-6
Viswanath K. K., Varakumar P., Pamuru R. R., Basha S. J., Mehta S., and Rao A. D. (2020). Plant lipoxygenases and their role in plant physiology. J. Plant Biol. 63, 83–95. doi: 10.1007/s12374-020-09241-x
Wang C.-F., Huang L.-L., Zhang H.-C., Han Q.-M., Buchenauer H., and Kang Z.-S. (2010). Cytochemical localization of reactive oxygen species (O2– and H2O2) and peroxidase in the incompatible and compatible interaction of wheat – Puccinia striiformis f. sp. tritici. Physiol. Mol. Plant Pathol. 74, 221–229. doi: 10.1016/j.pmpp.2010.02.002
Weber H., Chételat A., Caldelari D., and Farmer E. E. (1999). Divinyl ether fatty acid synthesis in late blight–diseased potato leaves. Plant Cell 11, 485–493. doi: 10.1105/tpc.11.3.485
Wong H. L., Sakamoto T., Kawasaki T., Umemura K., and Shimamoto K. (2004). Down-regulation of metallothionein, a reactive oxygen scavenger, by the small GTPase osRac1 in rice. Plant Physiol. 135, 1447–1456. doi: 10.1104/pp.103.036384
Yang J., Chen L., Zhang J., Liu P., Chen M., Chen Z., et al. (2024). TaTHI2 interacts with Ca2+ -dependent protein kinase TaCPK5 to suppress virus infection by regulating ROS accumulation. Plant Biotechnol. J. 22, 1335–1351. doi: 10.1111/pbi.14270
Yang M., Zhang F., Wang F., Dong Z., Cao Q., and Chen M. (2015). Characterization of a type 1 metallothionein gene from the stresses-tolerant plant Ziziphus jujuba. Int. J. Mol. Sci. 16, 16750–16762. doi: 10.3390/ijms160816750
Yin H., Wang Z., Li H., Zhang Y., Yang M., Cui G., et al. (2022). MsTHI1 overexpression improves drought tolerance in transgenic alfalfa (Medicago sativa L.). Front. Plant Sci. 13. doi: 10.3389/fpls.2022.992024
Yoshino K., Irieda H., Sugimoto F., Yoshioka H., Okuno T., and Takano Y. (2012). Cell Death of Nicotiana benthamiana Is Induced by Secreted Protein NIS1 of Colletotrichum orbiculare and Is Suppressed by a Homologue of CgDN3. Mol. Plant-Microbe Interactions® 25, 625–636. doi: 10.1094/MPMI-12-11-0316
Keywords: Colletotrichum tropicale, genome-wide expression analysis, Mangifera indica L. cv. Azúcar, oxidative burst, oxidative stress, MAPK, quiescence
Citation: Quintero-Mercado AF, Rojas S, Romero-Contreras YJ, Lozano LF, Serrano M and García C (2025) Differential gene expression in ripe mango fruit (Mangifera indica L. cv. Azúcar) that favors the pathogenicity of the endophyte Colletotrichum tropicale. Front. Fungal Biol. 6:1699983. doi: 10.3389/ffunb.2025.1699983
Received: 05 September 2025; Accepted: 20 October 2025;
Published: 06 November 2025.
Edited by:
Subhadeep Das, Adamas University, IndiaReviewed by:
Darshan Dharajiya, Gujarat Biotechnology Research Centre (GBRC), IndiaMd Motiur Rahman, Purdue University, United States
Copyright © 2025 Quintero-Mercado, Rojas, Romero-Contreras, Lozano, Serrano and García. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Andrés F. Quintero-Mercado, YXF1aW50ZXJvbWVAdW5hbC5lZHUuY28=