 Yuanyuan Luo1†
Yuanyuan Luo1† Feiming Huang
Feiming Huang Shiheng Lu
Shiheng Lu Yan Yan
Yan Yan- 1Department of Ophthalmology, Renji Hospital, School of Medicine, Shanghai Jiao Tong University, Shanghai, China
- 2School of Life Sciences, Shanghai University, Shanghai, China
- 3Department of Ophthalmology, Shanghai Eye Diseases Prevention and Treatment Center/Shanghai Eye Hospital, School of Medicine, Tongji University, Shanghai, China
- 4Department of Ophthalmology, Eye and ENT Hospital, Fudan University, Shanghai, China
Introduction: Pathological myopia, a severe form of myopia, is characterized by an extreme elongation of the eyeball, leading to various vision-threatening complications. It is broadly classified into two primary types: high myopia, which primarily involves an excessive axial length of the eye with potential for reversible vision loss, and degenerative myopia, associated with progressive and irreversible retinal damage.
Methods: Leveraging data from DisGeNET, reporting 184 genes linked to high myopia and 39 genes associated with degenerative myopia, we employed the GenePlexus methodology in conjunction with screening tests to further explore the genetic landscape of pathological myopia.
Results and discussion: Our comprehensive analysis resulted in the discovery of 21 new genes associated with degenerative myopia and 133 genes linked to high myopia with significant confidence. Among these findings, genes such as ADCY4, a regulator of the cAMP pathway, were functionally linked to high myopia, while THBS1, involved in collagen degradation, was closely associated with the pathophysiology of degenerative myopia. These previously unreported genes play crucial roles in the underlying mechanisms of pathological myopia, thereby emphasizing the complexity and multifactorial nature of this condition. The importance of our study resides in the uncovering of new genetic associations with pathological myopia, the provision of potential biomarkers for early screening, and the identification of therapeutic targets.
1 Introduction
Myopia is a pathological condition that affects distance vision (Ohno-Matsui et al., 2016; Foster and Jiang, 2014). As a common disease, the eye focus disorder can be corrected with eyeglasses, contact lenses or surgery. According to International Myopia Institute (IMI) white papers (Resnikoff et al., 2019; Tahhan et al., 2023), more than 30% of the world population is suffering from myopia, amounting to more than 2 billion people. Specifically, a higher prevalence have been observed in some regions of the world like U.S. (42%), China mainland (47%), and Hong Kong (62%) (Pan et al., 2012; Holden et al., 2016), indicating myopia as an ongoing global health issues. According to predictions, by 2050, more than 40% of people all over the world will suffer from myopia (Holden et al., 2016). Therefore, studying the pathogenesis of myopia constitutes one of the most important parts of current biomedical research.
Although myopia is a common condition characterized by symptoms such as difficulty seeing distant objects, squinting, and eye strain, heterogeneity in myopia pathogenesis persists (Naiglin et al., 1999; Jones and Luensmann, 2012). High myopia and degenerative myopia represent two major classes of myopia, exhibiting similar nearsighted symptoms but differing in their pathological mechanisms (Russo et al., 2014). High myopia is characterized by a group of myopia primarily caused by inherited genetic conditions, resulting in excessive eyeball elongation (Grzybowski and Kanclerz, 2019; Verkicharla et al., 2015), whereas degenerative myopia presents slightly different characteristics. The pathogenesis of degenerative myopia generally involves the damage to the retina (Park et al., 2013; Grossniklaus and Green, 1992; Lam et al., 2005), crucial for capturing vision and transmitting it to the brain. Despite shared clinical features (e.g., blurred vision, eye strain), their divergent pathological trajectories—mechanical stretching in high myopia versus retinal cell death in degenerative myopia—demand subtype-specific diagnostic and therapeutic strategies. Critically, early differentiation remains challenging due to overlapping symptoms, highlighting the need for molecular biomarkers to guide intervention.
Previously, studies on myopia have identified a series of biomarkers for high myopia and degenerative myopia, respectively. For instance, in 2001, Young and King identified several regions within the MYP2 gene on chromosome 18p11.31 associated with autosomal dominant high myopia (Young et al., 2001). In 2004, two genes, MYP7 and PAX6, were identified through a genome-wide scan as being associated with high myopia (Hammond et al., 2004), highlighting the genetic influence on high myopia pathogenesis. Regarding degenerative myopia, inherited retinal degenerations are prevalent in older populations, representing one of the most common causes of blindness in Europe and North America (Francis, 2006; Duncan et al., 2018). Genes like MYP5 (Paluru et al., 2003) and COLA1A (Madhuri et al., 2021) have been identified as pathogenic in retinal degeneration, playing a significant role in the initiation and progression of myopia. However, existing gene catalogs remain incomplete: DisGeNET curates 184 high myopia-associated genes versus only 39 for degenerative myopia, reflecting a critical knowledge gap in the latter’s molecular drivers. Furthermore, conventional genome-wide association studies (GWAS) and linkage analyses face limitations in resolving polygenic interactions and tissue-specific networks, necessitating advanced computational approaches.
Recent efforts to bridge this gap have employed network-based algorithms. For instance, Zhang et al. (2024) utilized the random walk with restart (RWR) method to prioritize candidate genes for both subtypes. While RWR leverages protein-protein interaction (PPI) networks, its reliance on predefined seed genes and static network topology may overlook context-dependent gene functions. Additionally, the lack of integration with functional validation data limits its predictive specificity. These shortcomings emphasize the need for methodologies that synergize multi-omic data, dynamic network modeling, and machine learning to improve biomarker discovery.
To address these challenges, we applied GenePlexus, a novel computational framework that integrates network propagation, functional enrichment, and cross-validation, in conjunction with our screening tests to predict subtype-specific biomarkers of myopia. By analyzing DisGeNET-curated genes (184 for high myopia, 39 for degenerative myopia), our study achieves two advances: (Ohno-Matsui et al., 2016): identifying 21 novel degenerative myopia-associated genes (e.g., THBS1, implicated in collagen degradation and retinal integrity) and 133 high-confidence high myopia-associated genes (e.g., ADCY4, modulating cAMP signaling in axial growth); (Foster and Jiang, 2014); validating the functional relevance of predicted genes through pathway analysis. Our findings not only expand the genetic landscape of pathological myopia but also establish a paradigm for computationally deconvoluting complex ophthalmic disorders.
2 Methods
2.1 Pathological myopia associated genes
Pathological myopia, a form of myopia characterized by severe nearsightedness and structural changes in the eye, can lead to additional visual impairment. We employed DisGeNET (v7.0, https://disgenet.com/) (Piñero et al., 2019), a comprehensive database aggregating gene-disease associations from diverse sources such as expert-curated databases, scientific literature, and electronic health records, thereby facilitating in-depth genetic research, to collect genes related to pathological myopia. Using the keywords “high myopia” and “degenerative myopia,” we identified genes on two distinct levels. Utilizing DisGeNET, we identified 184 genes associated with high myopia and 39 genes linked to degenerative myopia. The identified 184 high myopia and 39 degenerative myopia genes were subsequently matched to the protein-protein interaction (PPI) network, yielding 170 proteins for high myopia and 34 proteins for degenerative myopia, detailed in Supplementary Table S1.
2.2 PPI network
In this research, we focused on identifying proteins associated with high myopia and degenerative myopia, employing GenePlexus within the framework of the PPI network. Serving as the foundation for our exploration, the PPI network, a critical asset for pinpointing and forecasting essential genes, allowed us to utilize its comprehensive structure effectively. In the PPI network, the associations between proteins are clearly displayed. Each protein can be considered by taking all other proteins as background. The method based on the PPI network can systematically overview all proteins and investigate individual protein at a system level. This is the special merit compared with the traditional methods that can only investigate proteins individually. Here, we revealed new candidate genes associated with high myopia and degenerative myopia, thereby broadening the scope of knowledge beyond existing genetic findings. This PPI network has wide applications in tackling various protein-related problems (Pan et al., 2022; Zhang and Chen, 2020; Zhao et al., 2020; Zhao et al., 2019; Huang et al., 2023; Li et al., 2022a; Huang et al., 2024).
The data for our study were sourced from the STRING database (version 10, https://www.string-db.org/) (Szklarczyk et al., 2023), comprising 4,274,001 interactions among 19,247 human proteins, as detailed in the file “9606. protein.links.v10. txt.gz.” These interactions are depicted as links between pairs of proteins, each identified by their Ensembl IDs. Furthermore, the PPIs are assigned a confidence score ranging from 1 to 999. This scoring system derives from the amalgamation of various sub-scores, including measures of neighborhood, fusion, co-occurrence, co-expression, experimental evidence, database integration, and text mining analysis. These metrics provide a multifaceted evaluation of the proteins’ interactions, accounting for factors such as genomic proximity, gene fusion events, occurrences across species, co-expression patterns, and corroboration from scientific literature.
For the construction of the PPI network, 19,247 human proteins were utilized as nodes. Connections (or edges) between these nodes were established based on the existence of a PPI between the corresponding proteins, given that they had a confidence score above zero. The strength of these interactions is quantitatively represented in the edge weights, with higher scores indicating stronger associations. This methodological approach enabled a detailed and nuanced assessment of the protein interactions, facilitating the identification of potential genetic markers for high and degenerative myopia with greater precision.
2.3 GenePlexus
Addressing the challenge of identifying genes within a molecular network
2.4 Screening tests
Our results suggest that certain candidate genes, as identified by GenePlexus, exhibit a strong association with the structure of the PPI network and may display distinct behaviors. For instance, certain nodes might have a higher likelihood of being identified as key nodes, irrespective of the chosen seed nodes. Conversely, candidate genes exhibiting more substantial connections with previously validated genes are more likely to be identified as novel genes associated with high myopia and degenerative myopia. To further refine the identification process for these critical candidate genes, we designed three additional screening tests.
2.4.1 Permutation test
The architecture of the PPI network plays a pivotal role in determining the results generated by GenePlexus, with specific genes within the network being inherently more likely to be flagged due to their positions. However, it's important to note that not all these prominently flagged genes are directly linked to high myopia and degenerative myopia. To effectively sift through these genes, a permutation test was implemented. This began with the creation of one thousand random gene sets, each tagged from
where
2.4.2 Interaction test
This examination focuses on assessing the potential connection between selected genes and severe vision conditions, namely high myopia and its degenerative counterpart. The analysis entails determining how each gene under consideration relates to others that have already been associated with these myopic conditions (Cai et al., 2017; Liang et al., 2021). The intensity of protein interactions, a crucial factor in this assessment, is quantified using confidence scores available in the STRING database. For any given gene, referred to as
Candidate genes that exhibit high MIS values are considered more likely to be involved in high myopia and degenerative myopia due to the strength of their protein-protein interactions. To narrow down the most promising candidates, we established a minimum MIS value of 0.9, a benchmark reflecting the highest level of confidence according to the STRING database’s scoring system. This stringent criterion ensures that only genes with the most substantial evidence of a link to myopia are selected for further investigation.
2.4.3 Enrichment test
The purpose of this concluding analysis is to enhance the process of selecting candidate genes by examining how closely their functional terms align with those associated with severe myopic conditions, both high and degenerative. A gene’s candidacy is strengthened if its functional descriptors closely resemble those of genes already confirmed to be involved with these types of myopia (Chen et al., 2019; Chen et al., 2016). This resemblance is quantitatively assessed through the calculation of an enrichment score (ES), which is derived using Equation 3:
Here,
Additionally, for every candidate gene
To single out the most promising candidate genes for a link to high and degenerative myopia, an MES threshold of 0.95 has been established. This threshold serves as a benchmark to identify genes whose functional profiles not only share a high degree of similarity with those known to affect myopia but also suggests a substantial likelihood of their involvement in these conditions.
2.5 Outline of the method
In this study, a computational method was designed to identify latent genes related to high and degenerative myopia. The entire procedures are illustrated in Figure 1. The method started with validated genes of high and degenerative myopia retrieved from DisGeNET. Then, they were fed into GenePlexus to identify possible genes in a PPI network. Finally, these possible genes were filtered by three screening tests (permutation, interaction, and enrichment tests) with specific measurements and thresholds. The identified genes were confirmed by biological analysis.
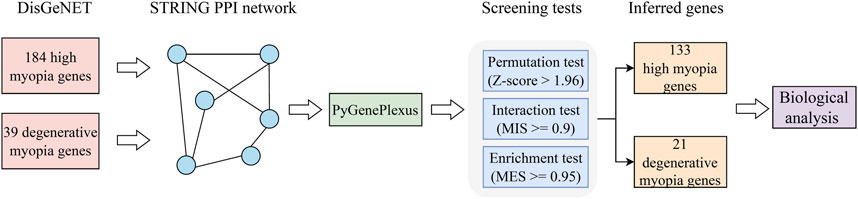
Figure 1. Flowchart depicting the procedure of extracting genes linked to pathological myopia. Initially, we gathered validated genes associated with high myopia and degenerative myopia from the DisGeNET database. Following this, we constructed a PPI network using data from the STRING database, a crucial step in visualizing the complex interplay between proteins. The GenePlexus tool was then utilized on this gene network to highlight candidates with a high likelihood of relevance. Subsequent refinement of these potential candidates was conducted through a series of three distinct screening tests, aiming to distill the list to a final set of genes deemed most pertinent. The culmination of our process involved a detailed evaluation of the relationship between these inferred genes and pathological myopia, employing a literature-based analysis to substantiate their relevance to the disease.
3 Results
3.1 Results of the GenePlexus
In this study, we explored the PPI network with the help of GenePlexus, focusing on proteins associated with pathological myopia. These proteins were designated as initial points of interest, or “seed nodes”. For each node within the network, we determined a specific likelihood of association with these seed nodes. Excluding the seed nodes themselves, the likelihoods assigned to all other nodes have been compiled in Supplementary Tables S2, S3. Subsequently, we identified the proteins corresponding to these designated nodes.
To assess the significance of each protein, we performed a permutation test to calculate p-values, with results detailed in Supplementary Tables S2, S3. Initially, proteins with a p-value below 0.05 were considered significant, leading to the identification of 2,600 proteins associated with high myopia and 1,427 proteins related to degenerative myopia. Further analysis was conducted through an interaction test, which assigned a MIS to each protein, as outlined in Supplementary Tables S2, S3. Proteins with an MIS of 0.9 or higher were selected, totaling 855 proteins for high myopia and 144 for degenerative myopia. To further evaluate their significance, we conducted an enrichment test on these filtered proteins. This test calculated the MES for each protein, with a threshold set at 0.95. Proteins below this cutoff were excluded, resulting in a final list of 133 proteins for high myopia and 21 for degenerative myopia, detailed in Supplementary Tables S2, S3. These proteins, termed “inferred proteins” in subsequent analyses, are considered to substantially correlate with the conditions of high myopia and degenerative myopia.
3.2 Associations between inferred proteins and validated proteins
In our research, we undertook multiple analyses to verify the reliability of proteins forecasted to be relevant to high and degenerative myopia. Specifically, we measured the number of interactions each predicted protein had within the PPI network with genes linked to these myopic conditions, setting a threshold for confidence scores at 0.9. The outcomes of these measurements are visually summarized in a bar plot illustrated in Figure 2.
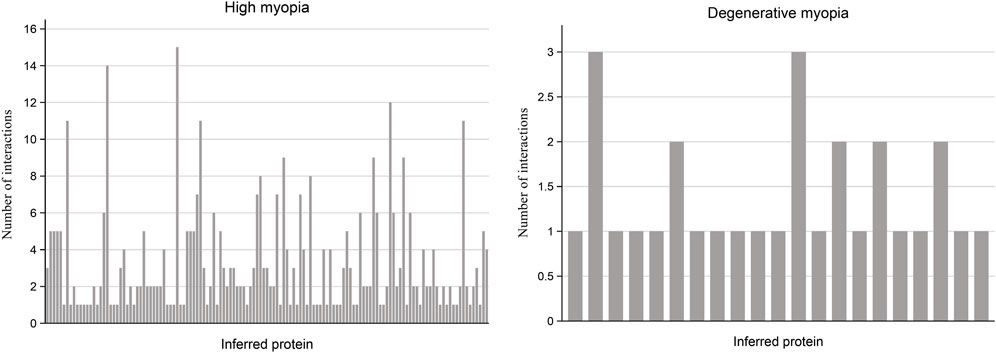
Figure 2. Bar plot illustrating the frequency of interactions between predicted proteins and known proteins linked to high myopia and degenerative myopia with high confidence scores. Among the recently discovered genes associated with high myopia and degenerative myopia, almost all have at least one interaction with known disease-related genes in the PPI network, indicating a strong connection between these inferred proteins and pathological myopia.
For the proteins associated with high myopia, our analysis revealed that 133 of them are connected to at least one gene within the PPI network known to be associated with high myopia. Notably, the protein HN1, identified by the code ENSP00000346839, was distinguished by its linkage to 15 genes related to high myopia, suggesting a potentially significant relationship between HN1 and this condition. Conversely, regarding degenerative myopia, 21 predicted proteins were identified as having at least one connection to a gene associated with degenerative myopia within the PPI network. By examining and comparing the connections between genes inferred from our study concerning high and degenerative myopia, and their specific disease-related genes, we ascertained that the genes identified through our computational analysis are strongly connected to both conditions. This linkage indicates a probable role of these inferred genes in the development and progression of high and degenerative myopia, thereby underscoring the accuracy of our research methodology.
4 Discussion
In this study, we introduced GenePlexus to predict novel genes associated with high myopia and degenerative myopia, building upon previously reported biomarkers for these two subtypes of myopia. We identified a series of novel biomarkers and compared our findings with the previous study using RWR algorithm (Zhang et al., 2024). This comparison revealed some common genes between the two methods, as illustrated in Figure 3. The detailed genes in three parts of each Venn diagram are listed in Supplementary Table S4. A literature review confirmed that some predicted myopia biomarkers identified in our study are associated with related pathological mechanisms, thereby validating the efficacy and accuracy of our predictions. The genes exclusively identified in this study are listed in Table 1.

Figure 3. Venn plot of pathological myopia-related genes identified based on the RWR method and those identified based on GenePlexus.

Table 1. Latent biomarkers identified in this study.
4.1 Validated biomarkers for degenerative myopia
Comparing our findings with previous predictions based on RWR method, we identified 10 shared genes associated with degenerative myopia predicted by both methods. Notably, SMAD5 (ENSP00000441954) emerged as the first in the prediction list. SMAD5 is a crucial component of the transforming growth factor beta (TGF-beta) signaling pathway (Gerjevic et al., 2012; Liu et al., 2013). According to recent publications, in 2017, Fisichella has summarized the specific role of TGF-beta signaling pathway for degeneration associated eye diseases including myopia (Fisichella, 2017), validating our prediction. Among the genes we predicted, TGFBR1 (ENSP00000364133) is notable for encoding a protein that interacts with type II TGF-beta receptors (Vander et al., 2018). Similarly, considering the specific role for TGF-beta signaling during retinal degeneration (Wang et al., 2019; Saika et al., 2009; Ma et al., 2019), it is reasonable for us to also predict such gene as degenerative myopia biomarkers, validating the efficacy of RWR and GenePlexus methods for degenerative myopia genes prediction. Additionally, WNT7A (ENSP00000285018), identified by both methods, is recognized for its involvement in developmental and homeostasis pathways (Lan et al., 2019). During the pathogenesis of degenerative myopia, WNT7A has been shown to regulate the plasticity of human retinal pigment epithelial cells, functionally associated with degenerative myopia (Kuznetsova et al., 2016). WNT7A-PAX6 signaling pathway has also been validated to be essential for retinal degeneration, associated with disease animal model design for degenerative myopia (Liu et al., 2017), implying the specific role of such gene during myopia pathogenesis. CDH1 (ENSP00000261769) is also one of the most important gene associated with degenerative myopia and has been selected by both methods. According to recent publications, retinal cadherins, encoded by CDH1 in situ, has been shown to be functionally associated with degenerative change in eye retina tissues (Yusuf et al., 2022). Cadherin associated pathways have also been shown to be pathological during the initiation and progression of myopia (Yang et al., 2022), consistent with our prediction on the specific role of CDH1 using both methods.
4.2 Biomarkers for degenerative myopia only predicted by GenePlexus
In addition to the gene identified by both methodologies as previously discussed, the GenePlexus approach has also pinpointed novel biomarkers linked to myopia. Specifically, it recognized two genes within the TGF-Beta signaling pathway—TGFB3 (ENSP00000238682) and TGFB2 (ENSP00000355896)—as being associated with degenerative myopia, a connection not made by the RWR method. Given the crucial role of the TGF-beta signaling in the pathogenesis of degenerative myopia, identifying these genes as potential biomarkers is well-founded (Fisichella, 2017). As two components of the signaling pathway, it is reasonable to predict such two genes as potential biomarkers. THBS1 (ENSP00000260356) is a subunit of a disulfide-linked homotrimer protein, functionally associated with cell-cell/cell-matrix interactions (Liu et al., 2020b; Duan et al., 2021). During the pathogenesis of degenerative myopia, increased collagen degradation associated matrix remodeling in retina has been shown to induce degenerative myopia (Ouyang et al., 2019; Pugazhendhi et al., 2020). Considering the specific role of THBS1 for collagen degradation (Wen et al., 2023; Inoue et al., 2013), its specific contribution on the pathogenesis of myopia can be validated. ACTA1 (ENSP00000355645) is another specific gene predicted to be associated with degenerative myopia. ACTA1 has been reported to participate in retinal development in guinea pig models (Srinivasalu et al., 2020). Degenerative change of extracellular matrix has been shown to be functionally associated with abnormal expression level of ACTA during myopia pathogenesis (Srinivasalu et al., 2020), validating the predictive efficacy of our newly presented GenePlexus based model. Novel predicted genes have also been shown to be functionally supported by recent publications to be associated with degenerative myopia, implying that GenePlexus is also an effective tool to identify disease subtyping associated biomarkers.
4.3 Validated biomarkers for high myopia
Using GenePlexus, we validated a set of biomarkers for high myopia previously predicted by the RWR method. For example, COL1A2 (ENSP00000297268), a component of type I collagen, has been identified as playing a significant role in extracellular matrix remodeling (Lamandé and Bateman, 2020). A 2021 study by researchers at Shanghai Ninth People’s Hospital, China, confirmed the link between type I collagen and high myopia mediated by microRNA-29a, supporting our findings (Zhu et al., 2022). Another gene associated with high myopia is SOX9 (ENSP00000245479), crucial for chondrocyte differentiation and skeletal growth (De Crombrugghe et al., 2000; Akiyama et al., 2002). Its interaction with BMP-2 and its role in scleral remodeling have been linked to high myopia (Li et al., 2015), validating our prediction. In 2012, researchers from Johnson and Johnson Vision Care also recognized specific role of SOX9 for extreme myopia (Ritchey et al., 2012). Additionally, GUCY2D (ENSP00000254854), which regulates cyclic GMP synthesis (Wimberg et al., 2018; Xue et al., 2019), has variants associated with high myopia, as demonstrated through genetic analyses (Haarman et al., 2022). Different direct protein functional associations, associations between GUCY2D and high myopia have been widely reported with a series of genetic association analyses (Haarman et al., 2022; Lazar et al., 2015; Stunkel et al., 2018). Local genetic variants around GUCY2D contributes to the pathogenesis of high myopia (Haarman et al., 2022; Koenekoop et al., 2007), which further indicates potential relationships between our predicted gene and such disease. SPP1 (ENSP00000378517) encodes a non-collagenous bone protein participating in cell-matrix interaction (Kramerova et al., 2019; Erikson et al., 2010). During the pathogenesis of high myopia, SPP1 has been shown to participate in the abnormal regulation of retinal pigment epithelium metabolism and further contributes to the pathogenesis of high myopia (Zhang, 2013). Familial genomic studies have pointed to SPP1 as a candidate gene for high myopia (Sánchez-Cazorla et al., 2023). These findings, validated by recent research, confirm the effectiveness and accuracy of the GenePlexus and RWR approaches in identifying relevant genes for high myopia.
4.4 Biomarkers for high myopia only predicted by GenePlexus
Comparing with RWR predicted high myopia associated genes, we further predicted variousGenePlexus specific high myopia associated genes, which have also been validated by recent publications, implying the efficacy and accuracy of GenePlexus approach. The first predicted gene is FGF7 (ENSP00000267843), a functional fibroblast growth factor regulating cell growth, tissue repair and morphogenesis (Nunes et al., 2016; Farooq et al., 2021; Prudovsky, 2021). Associated with FGF10, FGF7 has been confirmed to regulate specific mesenchymal-epithelial transition pathologically associated with extreme high myopia (Prudovsky, 2021), validating our prediction. The next predicted gene is ADCY4 (ENSP00000312126). ADCY4 is also a regulator for the secondary messenger cyclic adenosine monophosphate (cAMP) (Fan et al., 2019). The level of secondary messenger cyclic adenosine monophosphate in retina has been shown to be associated with the pathogenesis of high myopia by interfering with collagen metabolism (Zhou et al., 2012; Chun et al., 2015; Zhao et al., 2021). Therefore, it’s reasonable to speculate that ADCY4, as cAMP regulator is functionally associated with high myopia. MTOR (ENSP00000354558) encodes a serine/threonine protein kinase regulating cell metabolism, growth, and survival (Bhalla et al., 2017; Wullschleger et al., 2006). Gene polymorphisms of MTOR has been shown to be highly connected with myopia severity (Li et al., 2022b). BMPR2 (ENSP00000363708) is also a member of serine/threonine kinases participating in adipogenesis (Motomura et al., 2019; Schleinitz et al., 2011). During the pathogenesis of high myopia, BMPR2 has been shown to be associated with bidirectional regulator effects on retinal pigment epithelium (Zhang et al., 2012). Therefore, it is reasonable to predict such gene as a potential biomarker for high myopia. SMAD4 (ENSP00000341551) is a member of Smad family functionally associated with TGF-beta signaling (Zhao et al., 2018; Liu et al., 2023). In 2021, researchers from Fudan University confirmed aberrant TGF-beta signaling and SMAD signaling activation during the pathogenesis of high myopia (Zhu et al., 2021), validating our prediction. As we have discussed above, GenePlexus also recognized a series of novel high myopia biomarkers validated by previous publications, implying the efficacy and accuracy of such computational methods comparing with previous RWR methods.
In conclusion, as highlighted previously, the biomarkers we predicted for distinguishing between high myopia and degenerative myopia have been corroborated by recent scholarly articles. A comparison with the RWR method revealed that approximately 50% of our predicted biomarkers were also identified by RWR, underscoring the effectiveness and precision of predictions made using GenePlexus. This demonstrates that the integration of machine learning models like GenePlexus into the research process can successfully unveil novel biomarkers for various myopia subtypes. Furthermore, employing a range of effective machine learning models enhances the overall accuracy and efficiency of predictive modeling and biomarker discovery, suggesting a robust approach for the identification of disease-specific markers.
4.5 Enrichment analysis
For the identified genes of high and degenerative myopia, we conducted the enrichment analysis to uncover the biological meanings behind these genes. The recently proposed tool g: Profiler was employed to do this analysis (Kolberg et al., 2023), which can identify significantly enriched biological pathways and gene ontology (GO) terms for a given gene set. The enriched GO terms and pathways are listed in Figures 4, 5.
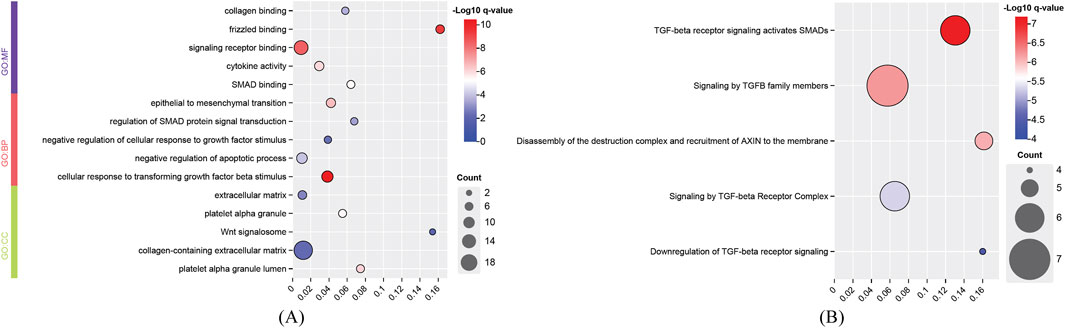
Figure 4. Enrichment analysis results of identified genes related to degenerative myopia. (A) Enriched gene ontology (GO) terms; (B) Enriched pathways. BP, biological process; CC, cellular component; MF, molecular function.
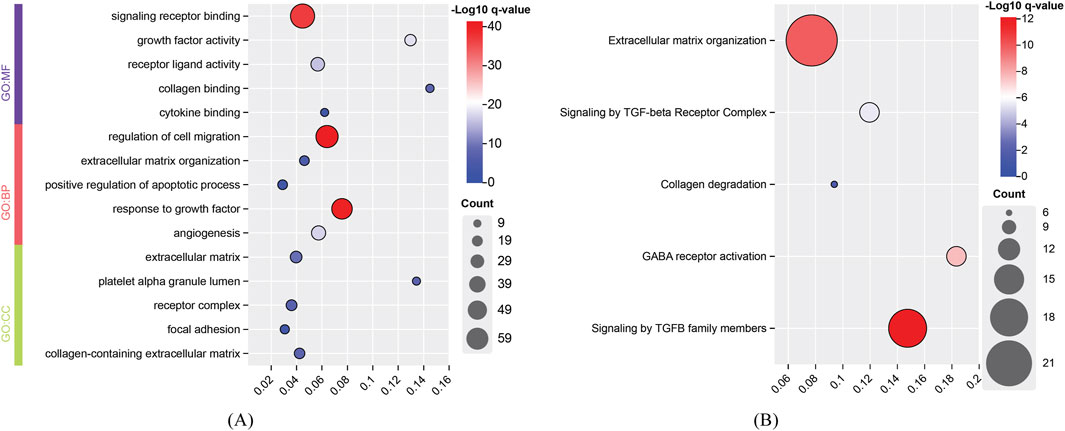
Figure 5. Enrichment analysis results of identified genes related to high myopia. (A) Enriched gene ontology (GO) terms; (B) Enriched pathways. BP, biological process; CC, cellular component; MF, molecular function.
The development of degenerative myopia involves complex interactions among various signaling pathways, including those mediated by frizzled receptors, signaling receptors, and cellular responses to transforming growth factor beta (TGF-β) stimuli. Frizzled receptors, such as FZD8, are integral to the Wnt signaling pathway, which influences ocular development and may contribute to myopia progression (Murillo-Garzón et al., 2018). The development of degenerative myopia involves complex interactions among various signaling pathways, including those mediated by frizzled receptors, signaling receptors, and cellular responses to transforming growth factor beta (TGF-β) stimuli. The cellular response to TGF-β stimuli involves the activation of Smad proteins, which translocate to the nucleus to regulate gene expression. Aberrant TGF-β1 signaling activation by MAF has been implicated in pathological lens growth, a key factor in the development of high myopia (Zhu et al., 2021).
The development of high myopia is influenced by signaling receptors, cell migration, and growth factor responses, with integrins playing a key role in cell attachment and migration, particularly in ocular tissue remodeling (Meguro et al., 2020). Cell migration, particularly in the sclera, is critical for myopia progression, with EGFR signaling playing a significant role in this process (Dong et al., 2020). Growth factors like IGF1 and FGF2 regulate ocular growth, contributing to myopia, and TGF-β family members are important for early eye development and myopia pathogenesis (Shu and Lovicu, 2021). In conclusion, high myopia arises from the interplay of signaling receptors, cell migration, and growth factor responses, leading to ocular remodeling and eye elongation.
4.6 Medical significance of this study
The medical significance of this study includes: (1) Early screening and precision diagnosis. The newly identified genes can serve as potential biomarkers for the development of early screening tools, facilitating the differentiation between high myopia and degenerative myopia and informing timely clinical interventions. (2) Development of therapeutic targets. The identified genes and pathways (e.g., TGF-β signaling, WNT7A-PAX6 axis) offer promising avenues for targeted therapeutic development. For example: Inhibiting THBS1 may reduce retinal collagen degradation, thereby delaying degenerative progression. Modulating ADCY4 or cAMP levels may help regulate excessive axial elongation. (3) Broader methodological applications. The computational framework is extensible to other complex diseases, such as glaucoma and age-related macular degeneration, advancing the application of computational biology in ophthalmology and beyond. (4) Potential for clinical translation. The study findings provide a theoretical foundation for gene-editing approaches (e.g., CRISPR) and small-molecule drug development (e.g., targeting TGF-β receptors), potentially driving the advancement of personalized treatment strategies.
4.7 Limitations of this study
This study designed a computational method to identify latent biomarkers of high and degenerative myopia. Some key genes were found out. However, some limitations exist. First, the proposed method was highly relied on the PPI network. The accuracy of PPI information determined the results of this method. At present, this information is far from complete, reducing the effectiveness of the method. Second, although some identified genes can be supported by some published literature, they still need to be validated by extensively wet experiments. In future, we will continue this work to infer more reliable biomarkers.
5 Conclusion
In our study, GenePlexus was utilized to explore the Protein-Protein Interaction (PPI) network, with a particular emphasis on genes implicated in both high and degenerative myopia, serving as initial seed nodes. This exploration entailed the application of three distinct screening techniques, resulting in the identification of 133 proteins related to high myopia and 21 proteins linked to degenerative myopia. Importantly, genes that emerged as highly relevant to pathological myopia were either directly associated with the condition or involved in recognized pathways pertinent to myopia, such as the regulation of gap junction trafficking and electrical signaling across these cellular junctions. The validation of genetic markers identified in our research, corroborated by existing literature, underscores GenePlexus’s precision and efficacy in elucidating the biological underpinnings of myopia. This methodology not only reinforces our comprehension of myopia but also proposes an innovative approach for biomarker discovery and the examination of underlying mechanisms in other complex conditions, providing a new avenue for the exploration of disease biomarkers and the deciphering of intricate disease pathways.
Data availability statement
Publicly available datasets were analyzed in this study. This data can be found here: https://disgenet.com/.
Author contributions
YL: Formal Analysis, Methodology, Writing–original draft. YW: Formal Analysis, Methodology, Writing–original draft. LL: Formal Analysis, Writing–review and editing. FH: Methodology, Writing–review and editing. SL: Conceptualization, Data curation, Validation, Writing–review and editing. YY: Conceptualization, Supervision, Validation, Writing–review and editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This work was supported by the National Natural Science Foundation of China (82171061).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fgene.2025.1533567/full#supplementary-material
SUPPLEMENTARY TABLE S1 | Listing of the 184 genes associated with high myopia and 39 genes associated with degenerative myopia, along with annotated information sourced from DisGeNET.
SUPPLEMENTARY TABLE S2 | Detailed results of screening tests, including each filtering condition, for genes associated with high myopia, presented in the corresponding sheet.
SUPPLEMENTARY TABLE S3 | Detailed results of screening tests, including each filtering condition, for genes associated with degenerative myopia, presented in the corresponding sheet.
SUPPLEMENTARY TABLE S4 | Intersection of genes identified by RWR and GenePlexus.
References
Akiyama, H., Chaboissier, M.-C., Martin, J. F., Schedl, A., and De Crombrugghe, B. (2002). The transcription factor Sox9 has essential roles in successive steps of the chondrocyte differentiation pathway and is required for expression of Sox5 and Sox6. Genes and Dev. 16 (21), 2813–2828. doi:10.1101/gad.1017802
Bhalla, M., Law, D., Dowd, G. C., and Ireton, K. (2017). Host serine/threonine kinases mTOR and protein kinase C-α promote InlB-mediated entry of Listeria monocytogenes. Infect. Immun. 85 (7). doi:10.1128/IAI.00087-17
Cai, Y.-D., Zhang, Q., Zhang, Y.-H., Chen, L., and Huang, T. (2017). Identification of genes associated with breast cancer metastasis to bone on a protein-protein interaction network with a shortest path algorithm. J. proteome Res. 16 (2), 1027–1038. doi:10.1021/acs.jproteome.6b00950
Chen, L., Pan, X., Zhang, Y. H., Liu, M., Huang, T., and Cai, Y. D. (2019). Classification of widely and rarely expressed genes with recurrent neural network. Comput. Struct. Biotechnol. J. 17, 49–60. doi:10.1016/j.csbj.2018.12.002
Chen, L., Zhang, Y. H., Zheng, M., Huang, T., and Cai, Y. D. (2016). Identification of compound-protein interactions through the analysis of gene ontology, KEGG enrichment for proteins and molecular fragments of compounds. Mol. Genet. genomics MGG. 291 (6), 2065–2079. doi:10.1007/s00438-016-1240-x
Chun, R. K. M., Shan, S. W., Lam, T. C., Wong, C. L., Li, K. K., Do, C. W., et al. (2015). Cyclic adenosine monophosphate activates retinal apolipoprotein A1 expression and inhibits myopic eye growth. Investigative Ophthalmol. and Vis. Sci. 56 (13), 8151–8157. doi:10.1167/iovs.14-14233
De Crombrugghe, B., Lefebvre, V., Behringer, R. R., Bi, W., Murakami, S., and Huang, W. (2000). Transcriptional mechanisms of chondrocyte differentiation. Matrix Biol. 19 (5), 389–394. doi:10.1016/s0945-053x(00)00094-9
Dong, L., Shi, X. H., Li, Y. F., Jiang, X., Wang, Y. X., Lan, Y. J., et al. (2020). Blockade of epidermal growth factor and its receptor and axial elongation in experimental myopia. Faseb J. 34 (10), 13654–13670. doi:10.1096/fj.202001095RR
Duan, F.-M., Fu, L.-J., Wang, Y.-H., Adu-Gyamfi, E. A., Ruan, L.-L., Xu, Z.-W., et al. (2021). THBS1 regulates trophoblast fusion through a CD36-dependent inhibition of cAMP, and its upregulation participates in preeclampsia. Genes and Dis. 8 (3), 353–363. doi:10.1016/j.gendis.2020.05.007
Duncan, J. L., Pierce, E. A., Laster, A. M., Daiger, S. P., Birch, D. G., Ash, J. D., et al. (2018). Inherited retinal degenerations: current landscape and knowledge gaps. Transl. Vis. Sci. Technol. 7 (4), 6. doi:10.1167/tvst.7.4.6
Erikson, D. W., Hsieh, Y.-H., Hayashi, K., Burghardt, R. C., Bayless, K. J., Chang, P.-L., et al. (2010). SPP1 (osteopontin) and SPARC (osteonectin) may interact developmentally during mouse pregnancy. Oxford University Press.
Fan, Y., Mu, J., Huang, M., Imani, S., Wang, Y., Lin, S., et al. (2019). Epigenetic identification of ADCY4 as a biomarker for breast cancer: an integrated analysis of adenylate cyclases. Epigenomics-Uk 11 (14), 1561–1579. doi:10.2217/epi-2019-0207
Farooq, M., Khan, A. W., Kim, M. S., and Choi, S. (2021). The role of fibroblast growth factor (FGF) signaling in tissue repair and regeneration. Cells 10 (11), 3242. doi:10.3390/cells10113242
Foster, Pa, and Jiang, Y. (2014). Epidemiology of myopia. Eye 28 (2), 202–208. doi:10.1038/eye.2013.280
Francis, P. J. (2006). Genetics of inherited retinal disease. J. R. Soc. Med. 99 (4), 189–191. doi:10.1177/014107680609900417
Gerjevic, L. N., Liu, N., Lu, S., and Harrison-Findik, D. D. (2012). Alcohol activates TGF-beta but inhibits BMP receptor-mediated Smad signaling and Smad4 binding to hepcidin promoter in the liver. Int. J. hepatology 2012, 459278. doi:10.1155/2012/459278
Grossniklaus, H. E., and Green, W. R. (1992). Pathologic findings in pathologic myopia. Retina 12 (2), 127–133. doi:10.1097/00006982-199212020-00009
Grzybowski, A., and Kanclerz, P. (2019). The standardized definition of high myopia. Graefe's Archive Clin. Exp. Ophthalmol. 257 (8), 1805. doi:10.1007/s00417-019-04397-w
Haarman, A. E., Thiadens, A. A., van Tienhoven, M., Loudon, S. E., de Klein, J. A., Brosens, E., et al. (2022). Whole exome sequencing of known eye genes reveals genetic causes for high myopia. Hum. Mol. Genet. 31 (19), 3290–3298. doi:10.1093/hmg/ddac113
Hammond, C. J., Andrew, T., Mak, Y. T., and Spector, T. D. (2004). A susceptibility locus for myopia in the normal population is linked to the PAX6 gene region on chromosome 11: a genomewide scan of dizygotic twins. Am. J. Hum. Genet. 75 (2), 294–304. doi:10.1086/423148
Holden, B. A., Fricke, T. R., Wilson, D. A., Jong, M., Naidoo, K. S., Sankaridurg, P., et al. (2016). Global prevalence of myopia and high myopia and temporal trends from 2000 through 2050. Ophthalmology 123 (5), 1036–1042. doi:10.1016/j.ophtha.2016.01.006
Huang, F., Fu, M., Li, J., Chen, L., Feng, K., Huang, T., et al. (2023). Analysis and prediction of protein stability based on interaction network, gene ontology, and KEGG pathway enrichment scores. BBA - Proteins Proteomics. 1871 (3), 140889. doi:10.1016/j.bbapap.2023.140889
Huang, F., Guo, W., Chen, L., Feng, K., Huang, T., and Cai, Y.-D. (2024). Identifying autophagy-associated proteins and chemicals with a random walk-based method within heterogeneous interaction network. Front. Bioscience-Landmark 29 (1), 21. doi:10.31083/j.fbl2901021
Inoue, M., Jiang, Y., Barnes, R. H., Tokunaga, M., Martinez-Santibañez, G., Geletka, L., et al. (2013). Thrombospondin 1 mediates high-fat diet-induced muscle fibrosis and insulin resistance in male mice. Endocrinology 154 (12), 4548–4559. doi:10.1210/en.2013-1587
Jones, D., and Luensmann, D. (2012). The prevalence and impact of high myopia. Eye and contact lens 38 (3), 188–196. doi:10.1097/ICL.0b013e31824ccbc3
Koenekoop, R. K., Lopez, I., Den Hollander, A. I., Allikmets, R., and Cremers, F. P. (2007). Genetic testing for retinal dystrophies and dysfunctions: benefits, dilemmas and solutions. Clin. and Exp. Ophthalmol. 35 (5), 473–485. doi:10.1111/j.1442-9071.2007.01534.x
Kolberg, L., Raudvere, U., Kuzmin, I., Adler, P., Vilo, J., and Peterson, H. (2023). g:Profiler-interoperable web service for functional enrichment analysis and gene identifier mapping (2023 update). Nucleic acids Res. 51 (W1), W207–W212. doi:10.1093/nar/gkad347
Kramerova, I., Kumagai-Cresse, C., Ermolova, N., Mokhonova, E., Marinov, M., Capote, J., et al. (2019). Spp1 (osteopontin) promotes TGFβ processing in fibroblasts of dystrophin-deficient muscles through matrix metalloproteinases. Hum. Mol. Genet. 28 (20), 3431–3442. doi:10.1093/hmg/ddz181
Kuznetsova, A. V., Aleksandrova, M. A., Kurinov, A. M., Chentsova, E. V., and Makarov, P. V. (2016). Plasticity of adult human retinal pigment epithelial cells. Int. J. Clin. Exp. Med. 9 (11), 20892–20906.
Lam, D. S., Fan, D. S., Chan, W.-M., Tam, B. S., Kwok, A. K., Leung, A. T., et al. (2005). Prevalence and characteristics of peripheral retinal degeneration in Chinese adults with high myopia: a cross-sectional prevalence survey. Optometry Vis. Sci. 82 (4), 235–238. doi:10.1097/01.opx.0000159359.49457.b4
Lamandé, S. R., and Bateman, J. F. (2020). Genetic disorders of the extracellular matrix. anatomical Rec. 303 (6), 1527–1542. doi:10.1002/ar.24086
Lan, L., Wang, W., Huang, Y., Bu, X., and Zhao, C. (2019). Roles of Wnt7a in embryo development, tissue homeostasis, and human diseases. J. Cell. Biochem. 120 (11), 18588–18598. doi:10.1002/jcb.29217
Lazar, C. H., Mutsuddi, M., Kimchi, A., Zelinger, L., Mizrahi-Meissonnier, L., Marks-Ohana, D., et al. (2015). Whole exome sequencing reveals GUCY2D as a major gene associated with cone and cone–rod dystrophy in Israel. Investigative Ophthalmol. and Vis. Sci. 56 (1), 420–430. doi:10.1167/iovs.14-15647
Li, H., Cui, D., Zhao, F., Huo, L., Hu, J., and Zeng, J. (2015). BMP-2 is involved in scleral remodeling in myopia development. PloS one 10 (5), e0125219. doi:10.1371/journal.pone.0125219
Li, X., Long, J., Liu, Y., Cai, Q., Zhao, Y., Jin, L., et al. (2022b). Association of MTOR and PDGFRA gene polymorphisms with different degrees of myopia severity. Exp. Eye Res. 217, 108962. doi:10.1016/j.exer.2022.108962
Li, X., Lu, L., and Chen, L. (2022a). Identification of protein functions in mouse with a label space partition method. Math. Biosci. Eng. 19 (4), 3820–3842. doi:10.3934/mbe.2022176
Liang, F., Fu, X., Ding, S., and Li, L. (2021). Use of a network-based method to identify latent genes associated with hearing loss in children. Front. cell Dev. Biol. 9, 783500. doi:10.3389/fcell.2021.783500
Liu, A., Yu, C., Qiu, C., Wu, Q., Huang, C., Li, X., et al. (2023). PRMT5 methylating SMAD4 activates TGF-β signaling and promotes colorectal cancer metastasis. Oncogene 42 (19), 1572–1584. doi:10.1038/s41388-023-02674-x
Liu, C.-H., Wang, Z., Sun, Y., and Chen, J. (2017). Animal models of ocular angiogenesis: from development to pathologies. FASEB J. 31 (11), 4665–4681. doi:10.1096/fj.201700336R
Liu, R., Mancuso, C. A., Yannakopoulos, A., Johnson, K. A., and Krishnan, A. (2020a). Supervised learning is an accurate method for network-based gene classification. Bioinformatics 36 (11), 3457–3465. doi:10.1093/bioinformatics/btaa150
Liu, X., Xu, D., Liu, Z., Li, Y., Zhang, C., Gong, Y., et al. (2020b). THBS1 facilitates colorectal liver metastasis through enhancing epithelial–mesenchymal transition. Clin. Transl. Oncol. 22, 1730–1740. doi:10.1007/s12094-020-02308-8
Liu, Y., Zheng, W.-K., Gao, W.-S., Shen, Y., and Ding, W.-Y. (2013). Function of TGF-beta and p38 MAKP signaling pathway in osteoblast differentiation from rat adipose-derived stem cells. Eur. Rev. Med. and Pharmacol. Sci. 17 (12), 1611–1619.
Ma, W., Silverman, S. M., Zhao, L., Villasmil, R., Campos, M. M., Amaral, J., et al. (2019). Absence of TGFβ signaling in retinal microglia induces retinal degeneration and exacerbates choroidal neovascularization. Elife 8, e42049. doi:10.7554/eLife.42049
Madhuri, V., Selina, A., Loganathan, L., Kumar, A., Kumar, V., Raymond, R., et al. (2021). Osteogenesis imperfecta: novel genetic variants and clinical observations from a clinical exome study of 54 Indian patients. Ann. Hum. Genet. 85 (1), 37–46. doi:10.1111/ahg.12403
Mancuso, C. A., Liu, R., and Krishnan, A. (2023). PyGenePlexus: a Python package for gene discovery using network-based machine learning. Bioinformatics 39 (2), btad064. doi:10.1093/bioinformatics/btad064
Meguro, A., Yamane, T., Takeuchi, M., Miyake, M., Fan, Q., Zhao, W., et al. (2020). Genome-wide association study in asians identifies novel loci for high myopia and highlights a nervous system role in its pathogenesis. Ophthalmology 127 (12), 1612–1624. doi:10.1016/j.ophtha.2020.05.014
Motomura, M., Shimokawa, F., Kobayashi, T., Yamashita, Y., Mizoguchi, I., Sato, Y., et al. (2019). Relationships between expression levels of genes related to adipogenesis and adipocyte function in dogs. Mol. Biol. Rep. 46 (5), 4771–4777. doi:10.1007/s11033-019-04923-3
Murillo-Garzón, V., Gorroño-Etxebarria, I., Åkerfelt, M., Puustinen, M. C., Sistonen, L., Nees, M., et al. (2018). Frizzled-8 integrates Wnt-11 and transforming growth factor-β signaling in prostate cancer. Nat. Commun. 9 (1), 1747. doi:10.1038/s41467-018-04042-w
L. Naiglin, J. Clayton, C. Gazagne, F. Dallongeville, F. Malecaze, and P. Calvas (1999). Familial high myopia: evidence of an autosomal dominant mode of inheritance and genetic heterogeneity. Ann. Genet. 42 (3), 140–146.
Nunes, Q. M., Li, Y., Sun, C., Kinnunen, T. K., and Fernig, D. G. (2016). Fibroblast growth factors as tissue repair and regeneration therapeutics. PeerJ 4, e1535. doi:10.7717/peerj.1535
Ohno-Matsui, K., Lai, T. Y., Lai, C.-C., and Cheung, C. M. G. (2016). Updates of pathologic myopia. Prog. Retin. eye Res. 52, 156–187. doi:10.1016/j.preteyeres.2015.12.001
Ouyang, X., Han, Y., Xie, Y., Wu, Y., Guo, S., Cheng, M., et al. (2019). The collagen metabolism affects the scleral mechanical properties in the different processes of scleral remodeling. Biomed. and Pharmacother. 118, 109294. doi:10.1016/j.biopha.2019.109294
Paluru, P., Ronan, S. M., Heon, E., Devoto, M., Wildenberg, S. C., Scavello, G., et al. (2003). New locus for autosomal dominant high myopia maps to the long arm of chromosome 17. Investigative Ophthalmol. and Vis. Sci. 44 (5), 1830–1836. doi:10.1167/iovs.02-0697
Pan, C. W., Ramamurthy, D., and Saw, S. M. (2012). Worldwide prevalence and risk factors for myopia. Ophthalmic Physiological Opt. 32 (1), 3–16. doi:10.1111/j.1475-1313.2011.00884.x
Pan, X., Chen, L., Liu, I., Niu, Z., Huang, T., and Cai, Y. D. (2022). Identifying protein subcellular locations with embeddings-based node2loc. IEEE/ACM Trans. Comput. Biol. Bioinform 19 (2), 666–675. doi:10.1109/TCBB.2021.3080386
Park, H., Tan, C. C., Faulkner, A., Jabbar, S. B., Schmid, G., Abey, J., et al. (2013). Retinal degeneration increases susceptibility to myopia in mice. Mol. Vis. 19, 2068–2079.
Piñero, J., Ramírez-Anguita, J. M., Saüch-Pitarch, J., Ronzano, F., Centeno, E., Sanz, F., et al. (2019). The DisGeNET knowledge platform for disease genomics: 2019 update. Nucleic acids Res. 48 (D1), D845–D855. doi:10.1093/nar/gkz1021
Prudovsky, I. (2021). Cellular mechanisms of FGF-stimulated tissue repair. Cells 10 (7), 1830. doi:10.3390/cells10071830
Pugazhendhi, S., Ambati, B., and Hunter, A. A. (2020). Pathogenesis and prevention of worsening axial elongation in pathological myopia. Clin. Ophthalmol. 14, 853–873. doi:10.2147/OPTH.S241435
Resnikoff, S., Jonas, J. B., Friedman, D., He, M., Jong, M., Nichols, J. J., et al. (2019). Myopia–a 21st century public health issue. Investigative Ophthalmol. Vis. Sci. 60 (3). Mi-Mii. doi:10.1167/iovs.18-25983
Ritchey, E. R., Zelinka, C. P., Tang, J., Liu, J., and Fischer, A. J. (2012). The combination of IGF1 and FGF2 and the induction of excessive ocular growth and extreme myopia. Exp. Eye Res. 99, 1–16. doi:10.1016/j.exer.2012.03.019
Russo, A., Semeraro, F., Romano, M. R., Mastropasqua, R., Dell’Omo, R., and Costagliola, C. (2014). Myopia onset and progression: can it be prevented? Int. Ophthalmol. 34, 693–705. doi:10.1007/s10792-013-9844-1
Saika, S., Yamanaka, O., Okada, Y., Tanaka, S.-I., Miyamoto, T., Sumioka, T., et al. (2009). TGF beta in fibroproliferative diseases in the eye. Front. Bioscience-Scholar 1 (1), 376–390. doi:10.2741/S32
Sánchez-Cazorla, E., González-Atienza, C., López-Vázquez, A., Arruti, N., Nieves-Moreno, M., Noval, S., et al. (2023). Whole-Exome sequencing of 21 families: candidate genes for early-onset high myopia. Int. J. Mol. Sci. 24 (21), 15676. doi:10.3390/ijms242115676
Schleinitz, D., Klöting, N., Böttcher, Y., Wolf, S., Dietrich, K., Tönjes, A., et al. (2011). Genetic and evolutionary analyses of the human bone morphogenetic protein receptor 2 (BMPR2) in the pathophysiology of obesity. PloS one 6 (2), e16155. doi:10.1371/journal.pone.0016155
Shu, D. Y., and Lovicu, F. J. (2021). Insights into bone morphogenetic protein-(BMP-) signaling in ocular lens biology and pathology. Cells 10 (10), 2604. doi:10.3390/cells10102604
Srinivasalu, N., Zhang, S., Xu, R., Reinach, P. S., Su, Y., Zhu, Y., et al. (2020). Crosstalk between EP2 and PPARα modulates hypoxic signaling and myopia development in Guinea pigs. Investigative Ophthalmol. and Vis. Sci. 61 (8), 44. doi:10.1167/iovs.61.8.44
Stunkel, M. L., Brodie, S. E., Cideciyan, A. V., Pfeifer, W. L., Kennedy, E. L., Stone, E. M., et al. (2018). Expanded retinal disease spectrum associated with autosomal recessive mutations in GUCY2D. Am. J. Ophthalmol. 190, 58–68. doi:10.1016/j.ajo.2018.03.021
Szklarczyk, D., Kirsch, R., Koutrouli, M., Nastou, K., Mehryary, F., Hachilif, R., et al. (2023). The STRING database in 2023: protein-protein association networks and functional enrichment analyses for any sequenced genome of interest. Nucleic acids Res. 51 (D1), D638–D646. doi:10.1093/nar/gkac1000
Tahhan, N., Wolffsohn, J. S., Sankaridurg, P., Jonas, J. B., Bullimore, M. A., Flitcroft, I., et al. (2023). Editorial: international myopia Institute white paper series 2023. Investigative Ophthalmol. and Vis. Sci. 64 (6), 1. doi:10.1167/iovs.64.6.1
Vander, A. A., Cao, J., and Li, X. (2018). TGF-β receptors: in and beyond TGF-β signaling. Cell. Signal. 52, 112–120. doi:10.1016/j.cellsig.2018.09.002
Verkicharla, P. K., Ohno-Matsui, K., and Saw, S. M. (2015). Current and predicted demographics of high myopia and an update of its associated pathological changes. Ophthalmic Physiological Opt. 35 (5), 465–475. doi:10.1111/opo.12238
Wang, K., Li, H., Sun, R., Liu, C., Luo, Y., Fu, S., et al. (2019). Emerging roles of transforming growth factor β signaling in wet age-related macular degeneration. Acta Biochimica Biophysica Sinica 51 (1), 1–8. doi:10.1093/abbs/gmy145
Wen, Z., Zhang, Y., Wang, X., Wu, Y., Mao, J., Li, Q., et al. (2023). THBS1-Mediated degradation of collagen via the PI3K/AKT pathway facilitates the metastasis and poor prognosis of OSCC. Int. J. Mol. Sci. 24 (17), 13312. doi:10.3390/ijms241713312
Wimberg, H., Lev, D., Yosovich, K., Namburi, P., Banin, E., Sharon, D., et al. (2018). Photoreceptor guanylate cyclase (GUCY2D) mutations cause retinal dystrophies by severe malfunction of Ca2+-dependent cyclic GMP synthesis. Front. Mol. Neurosci. 11, 348. doi:10.3389/fnmol.2018.00348
Wullschleger, S., Loewith, R., and Hall, M. N. (2006). TOR signaling in growth and metabolism. Cell 124 (3), 471–484. doi:10.1016/j.cell.2006.01.016
Xue, F., Wei, T., Sun, J., Luo, Y., Huo, Y., Yu, P., et al. (2019). GUCY2D gene loss-of-function mutations responsible for leber congenital amaurosis 1. Res. Square. doi:10.21203/rs.2.11649/v2
Yang, J., Ouyang, X., Fu, H., Hou, X., Liu, Y., Xie, Y., et al. (2022). Advances in biomedical study of the myopia-related signaling pathways and mechanisms. Biomed. and Pharmacother. 145, 112472. doi:10.1016/j.biopha.2021.112472
Young, T. L., Atwood, L. D., Ronan, S. M., Dewan, A. T., Alvear, A. B., Peterson, J., et al. (2001). Further refinement of the MYP2 locus for autosomal dominant high myopia by linkage disequilibrium analysis. Ophthalmic Genet. 22 (2), 69–75. doi:10.1076/opge.22.2.69.2233
Yusuf, I. H., Garrett, A. M., MacLaren, R. E., and Issa, P. C. (2022). Retinal cadherins and the retinal cadherinopathies: current concepts and future directions. Prog. Retin. eye Res. 90, 101038. doi:10.1016/j.preteyeres.2021.101038
Zhang, J., Huang, T., Sun, Q., and Zhang, J. (2024). Identifying pathological myopia associated genes with A random walk-based method in protein-protein interaction network. Curr. Bioinforma. 19 (4), 375–384. doi:10.2174/0115748936268218231114070754
Zhang, X. L., and Chen, L. (2020). Prediction of membrane protein types by fusing protein-protein interaction and protein sequence information. Bba-Proteins Proteom 1868 (12), 140524. doi:10.1016/j.bbapap.2020.140524
Zhang, Y. (2013). Role of retinal pigment epithelium in myopia development and control. Berkeley: University of California.
Zhang, Y., Liu, Y., and Wildsoet, C. F. (2012). Bidirectional, optical sign-dependent regulation of BMP2 gene expression in chick retinal pigment epithelium. Investigative Ophthalmol. and Vis. Sci. 53 (10), 6072–6080. doi:10.1167/iovs.12-9917
Zhao, F., Zhou, H., Chen, W., Zhao, C., Zheng, Y., Tao, Y., et al. (2021). Declines in PDE4B activity promote myopia progression through downregulation of scleral collagen expression. Exp. Eye Res. 212, 108758. doi:10.1016/j.exer.2021.108758
Zhao, M., Mishra, L., and Deng, C. X. (2018). The role of TGF-β/SMAD4 signaling in cancer. Int. J. Biol. Sci. 14 (2), 111–123. doi:10.7150/ijbs.23230
Zhao, R., Chen, L., Zhou, B., Guo, Z.-H., Wang, S., and Aorigele, (2019). Recognizing novel tumor suppressor genes using a network machine learning strategy. IEEE Access 7, 155002–155013. doi:10.1109/access.2019.2949415
Zhao, R., Hu, B., Chen, L., and Zhou, B. (2020). Identification of latent oncogenes with a network embedding method and random forest. BioMed Res. Int. 2020, 5160396. doi:10.1155/2020/5160396
Zhou, X., Tao, Y., Pan, M., Lu, R., Chen, J.-F., and Qu, J. (2012). cAMP regulates visual development and myopia occurrence in Guinea pigs probably via reducing scleral collagen synthesis. Investigative Ophthalmol. and Vis. Sci. 53 (14), 4661.
Zhu, X., Du, Y., Li, D., Xu, J., Wu, Q., He, W., et al. (2021). Aberrant TGF-β1 signaling activation by MAF underlies pathological lens growth in high myopia. Nat. Commun. 12 (1), 2102. doi:10.1038/s41467-021-22041-2
Keywords: pathological myopia, high myopia, degenerative myopia, disease gene, GenePlexus, DisGeNET
Citation: Luo Y, Wang Y, Liu L, Huang F, Lu S and Yan Y (2025) Identifying pathological myopia associated genes with GenePlexus in protein-protein interaction network. Front. Genet. 16:1533567. doi: 10.3389/fgene.2025.1533567
Received: 27 November 2024; Accepted: 18 February 2025;
Published: 05 March 2025.
Edited by:
Lin Lu, Columbia University, United StatesReviewed by:
Guohua Huang, Shaoyang University, ChinaShuaiqun Wang, Shanghai Maritime University, China
Copyright © 2025 Luo, Wang, Liu, Huang, Lu and Yan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Shiheng Lu, bHVkaWNlQDE2My5jb20=; Yan Yan, aHoyMDA0eWFuQDE2My5jb20=
†These authors have contributed equally to this work