 Peng Zhong
Peng Zhong Chumeng Zhang2
Chumeng Zhang2 Qinfeng Wu
Qinfeng Wu- 1Department of Cardiology, Jining No.1 People’s Hospital, Jining, Shandong, China
- 2The Second School of Clinical Medicine of Shandong First Medical University, Tai’an, Shandong, China
- 3College of Medical Information and Artificial Intelligence, Shandong First Medical University, Jinan, Shandong, China
- 4The Department of Otorhinolaryngology Head and Neck Surgery, Yantai Yuhuangding Hospital, Qingdao University, Qingdao, Shandong, China
Introduction: Cardiovascular diseases (CVDs), including myocardial infarction (MI), heart failure (HF), atrial fibrillation (AF), and arrhythmia, are major contributors to global mortality and often share overlapping risk factors and pathophysiological mechanisms. While genome-wide association studies (GWAS) have identified many loci for individual CVDs, the shared genetic architecture across related traits—particularly in East Asian populations—remains underexplored.
Materials and methods: We integrated large-scale GWAS summary statistics from East Asian populations to perform genome-wide and local genetic correlation analyses across four CVD phenotypes and five cardiometabolic traits (blood pressure and lipid levels). Using stratified LD score regression, we assessed tissue-specific heritability enrichment. Multi-trait analysis of GWAS (MTAG) was then employed to identify pleiotropic loci associated with multiple traits, with functional annotation and expression quantitative trait loci (eQTL) data used to explore biological relevance.
Results: We observed extensive genetic correlations among CVDs and between CVDs and cardiometabolic traits, with HF showing the strongest connections to both MI and arrhythmia. Notable genome-wide correlations were found between MI and SBP (rg = 0.35, P = 1.59 × 10−14) and between HF and DBP (rg = 0.54, P = 9.84 × 10−9). Stratified heritability analyses revealed significant enrichment in heart and arterial tissues, highlighting the relevance of cardiovascular-specific regulatory elements. MTAG identified several pleiotropic loci, including established genes such as APOB and MC4R, and novel East Asian-enriched signals such as QSOX2 and GUCY1A1/GUCY1B1. Functional data indicated that QSOX2 variants regulate gene expression in arterial and cardiac tissues, implicating redox regulation in HF and hypertension pathogenesis.
Conclusion: Our findings provide comprehensive insight into the shared genetic determinants of cardiovascular and metabolic diseases in East Asian populations. The identification of pleiotropic and ancestry-specific loci, along with tissue-specific regulatory patterns, underscores the need for integrative multi-trait and population-informed approaches in cardiovascular genetics and risk prediction.
1 Introduction
Cardiovascular diseases (CVDs), including myocardial infarction (MI), heart failure (HF), atrial fibrillation (AF), and arrhythmia, represent leading causes of morbidity and mortality worldwide (Kim, 2021). These conditions share overlapping pathophysiological mechanisms, such as myocardial ischemia, structural remodeling, and electrophysiological instability, which are increasingly understood to have genetic underpinnings. Genome-wide association studies (GWAS) have successfully identified susceptibility loci for individual conditions, especially in populations of European ancestry (Hartiala et al., 2021). However, the transferability of these findings to other populations, particularly East Asians, remains limited.
Given the substantial global burden and frequent co-occurrence of MI, HF, AF, and other arrhythmias, increasing attention has been directed toward their shared genetic architecture. Epidemiological data demonstrate considerable comorbidity across these conditions. For instance, up to 30% of patients with HF also develop AF, and prior MI significantly increases the risk of both arrhythmia and progressive HF (Packer, 2020; Wang et al., 2023). Furthermore, AF is now recognized as both a consequence and a driver of structural heart disease, with overlapping risk factors such as hypertension, diabetes, and aging (Li et al., 2022). These observations suggest that these CVDs may not be entirely distinct but instead represent manifestations of interconnected biological processes with common genetic underpinnings.
To systematically investigate these connections, multi-trait analytical frameworks offer a powerful approach to uncover shared and distinct loci contributing to this spectrum of disorders. In particular, Multi-Trait Analysis of GWAS (MTAG) enables the joint interrogation of genetically correlated phenotypes, thereby enhancing locus discovery and improving interpretability of pleiotropic associations (Turley et al., 2018).
In this study, we performed a multi-trait GWAS focusing on four major cardiovascular conditions—MI, HF, AF, and arrhythmia—in East Asian populations. By integrating functional annotations and tissue-specific enrichment analyses, we aimed to elucidate the shared and unique genetic determinants of these disorders and provide ancestry-informed insights into their biological mechanisms.
2 Materials and methods
2.1 GWAS data
GWAS summary statistics for the traits analyzed in this study were obtained from the GWAS Catalog (Buniello et al., 2019), with detailed information provided in Supplementary Table S1. Summary data for cardiometabolic traits—including systolic blood pressure (SBP), diastolic blood pressure (DBP), high-density lipoprotein (HDL), low-density lipoprotein (LDL), and triglycerides (TG)—were derived from a meta-analysis of East Asian cohorts (Chen et al., 2023), incorporating 92,615 participants from the Taiwan Biobank, Biobank Japan, and the East Asian subset of UK Biobank.
2.2 Global genetic correlation analysis
We assessed the genetic correlation (rg) between CVDS and cardiometabolic traits using linkage disequilibrium score regression (LDSC). LDSC estimates rg by analyzing the relationship between GWAS test statistics and LD scores, which represent the cumulative LD between a given SNP and its neighboring variants. This method operates on summary-level GWAS data, making it well-suited for large-scale meta-analyses and robust to confounding from sample overlap. In contrast to approaches requiring individual-level genotypes, LDSC is less susceptible to biases from population stratification or cryptic relatedness. As such, it offers a reliable framework for characterizing the genetic overlap between traits and for informing the selection of phenotypes in subsequent causal inference analyses (Bulik-Sullivan et al., 2015). The formula used in LDSC is as follows:
where
2.3 Cell-type-specific enrichment of SNP heritability
Stratified LD Score Regression (s-LDSC) extends the LDSC framework by quantifying the contribution of specific genomic annotations to the heritability of complex traits (Trynka et al., 2013). By modeling heritability across predefined functional categories while adjusting for linkage disequilibrium, s-LDSC enables the identification of biologically relevant regions contributing to disease risk. Unlike conventional approaches that rely on genome-wide significant variants, s-LDSC utilizes all SNPs to provide a more comprehensive view of the polygenic architecture. This method is computationally scalable and suitable for large-scale GWAS datasets. Importantly, s-LDSC can pinpoint enrichment in cell type–specific regulatory elements and functional annotations, aiding in the prioritization of genomic features for follow-up studies. Its integration into cross-trait analyses also allows for more nuanced interpretation of genetic correlations by revealing functional categories that drive shared heritability (Finucane et al., 2015).
To dissect tissue- and cell type-specific contributions to trait heritability, we employed s-LDSC using functional annotations derived from six chromatin marks (DHS, H3K27ac, H3K36me3, H3K4me1, H3K4me3, and H3K9ac) across 88 tissues and cell types from the Roadmap Epigenomics Consortium (Finucane et al., 2015). For each histone mark, annotations were grouped into nine biological categories: adipose, cardiovascular, central nervous system, digestive, immune/blood, liver, pancreas, musculoskeletal/connective tissue, and others. Trait-specific heritability enrichment was calculated for each annotation, and the results were visualized through hierarchical clustering based on normalized enrichment scores. This approach enabled the discovery of distinct enrichment profiles across traits and tissues, revealing shared biological underpinnings among genetically correlated phenotypes. Chromatin mark-specific signals provided further granularity, highlighting regulatory elements that may play central roles in tissue-relevant pathways. Together, these analyses support the functional interpretation of GWAS findings and inform the prioritization of candidate tissues and regulatory mechanisms involved in complex trait etiology.
2.4 Local genetic correlation analysis
To complement the genome-wide genetic correlation estimates obtained from LDSC, we applied ρ-HESS to assess local genetic correlations between trait pairs (Shi et al., 2017). While LDSC provides a genome-wide average estimate, ρ-HESS partitions the genetic covariance across 1,703 approximately independent genomic regions, enabling locus-level interrogation of shared genetic architecture. This method uses GWAS summary statistics and accounts for linkage disequilibrium patterns and potential sample overlap, without assuming specific distributions of effect sizes. For each region, ρ-HESS estimates local SNP heritability and covariance by projecting GWAS effect size vectors onto LD-derived eigenvectors. We applied this approach to all trait pairs with significant global genetic correlations from LDSC, using a Bonferroni-corrected threshold (P < 0.05/1,703) to determine statistical significance. This analysis allowed us to identify specific genomic intervals that disproportionately contribute to the observed genome-wide correlations, revealing heterogeneity in shared genetic architecture across loci.
2.5 Multi-trait analysis of GWAS
To enhance locus discovery and statistical power, we applied Multi-Trait Analysis of GWAS (MTAG), a method that leverages shared genetic architecture across genetically correlated traits to identify trait-specific SNP associations (Turley et al., 2018). MTAG uses GWAS summary statistics and estimates pairwise genetic correlations via LDSC (Bulik-Sullivan et al., 2015), accounting for sample overlap and trait correlation to generate unbiased SNP-level effect estimates. In our analysis, the included GWAS datasets were largely derived from non-overlapping cohorts; nonetheless, MTAG’s internal framework corrects for any residual overlap using covariance estimates from LDSC, thereby reducing inflation in test statistics. This framework is particularly suited for complex, polygenic traits with overlapping etiology. After stringent variant filtering, we retained SNPs that achieved genome-wide significance (P < 5 × 10−8) in MTAG and showed suggestive associations (P < 0.01) in the original single-trait GWAS, ensuring robustness and biological relevance. This approach enabled the identification of pleiotropic loci contributing to multiple cardiovascular phenotypes.
3 Results
3.1 Genetic correlations between cardiovascular diseases and cardiometabolic traits
We performed genome-wide genetic correlation analysis using LDSC to evaluate shared genetic architecture among four major CVDs and five cardiometabolic traits (Figure 1; Supplementary Table S2). MI exhibited strong and significant positive genetic correlations with SBP (rg = 0.35, SE = 0.05, P = 1.59 × 10−14), DBP (rg = 0.30, SE = 0.05, P = 7.02 × 10−9), LDL (rg = 0.27, SE = 0.05, P = 1.15 × 10−7), and TG (rg = 0.21, SE = 0.03, P = 1.92 × 10−10). A significant inverse correlation was observed with HDL (rg = −0.20, SE = 0.05, P = 8.50 × 10−5). These results confirm the known involvement of blood pressure and lipid metabolism pathways in MI pathophysiology. HF also showed robust genetic correlations with both SBP (rg = 0.52, SE = 0.0891, P = 4.81 × 10−9) and DBP (rg = 0.54, SE = 0.0937, P = 9.84 × 10−9), highlighting a shared genetic basis linked to hemodynamic stress. Additionally, HF was modestly associated with TG (rg = 0.22, SE = 0.0699, P = 0.0021), and negatively correlated with HDL (rg = −0.19, SE = 0.0757, P = 0.0142), further suggesting convergence on metabolic dysregulation. In contrast, AF demonstrated weaker genetic correlations with cardiometabolic traits, with no significant associations observed for SBP or DBP. Modest but nominal associations were observed with LDL (rg = −0.22, SE = 0.10, P = 0.0296) and TG (rg = −0.18, SE = 0.07, P = 0.0109), suggesting a partially distinct genetic basis. Arrhythmia showed moderate genetic correlations with SBP (rg = 0.22, SE = 0.0486, P = 8.14 × 10−6) and DBP (rg = 0.18, SE = 0.0498, P = 0.0004), but weaker or no associations with lipid traits, including non-significant correlations with HDL and LDL. A small negative correlation with TG was noted (rg = −0.10, SE = 0.0442, P = 0.0185).
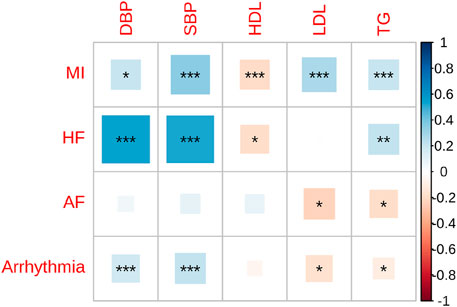
Figure 1. Genetic correlations between cardiovascular diseases and cardiometabolic traits in East Asians. This heatmap illustrates the pairwise genetic correlation between four cardiovascular disease phenotypes—myocardial infarction (MI), heart failure (HF), atrial fibrillation (AF), and arrhythmia—and six cardiometabolic traits—diastolic blood pressure (DBP), systolic blood pressure (SBP), high-density lipoprotein cholesterol (HDL), low-density lipoprotein cholesterol (LDL), triglycerides (TG), and HDL. The strength and direction of correlations are color-coded, with positive correlations shown in blue and negative correlations in red. The size of the squares represents the magnitude of the genetic correlation. Asterisks indicate levels of statistical significance: P < 0.05 (*), P < 0.01 (**), P < 0.001 (***).
3.2 Genetic correlations among cardiovascular diseases
To further examine shared genetic architecture across cardiovascular phenotypes, we estimated pairwise genetic correlations among MI, HF, AF, and arrhythmia (Table 1). MI showed a strong genetic correlation with HF (rg = 0.43, SE = 0.10, P = 1.76 × 10−5), and a moderate correlation with AF (rg = 0.27, SE = 0.11, P = 0.0165). However, the correlation between MI and arrhythmia was not significant (rg = 0.07, SE = 0.07, P = 0.2953). In contrast, HF exhibited significant genetic overlap with both AF (rg = 0.48, SE = 0.14, P = 0.0005) and arrhythmia (rg = 0.55, SE = 0.087, P = 3.78 × 10−10), indicating common underlying etiological pathways likely related to electrical and structural remodeling. As expected, AF and arrhythmia were highly correlated (rg = 1.0, SE = 0.08, P = 1.73 × 10−43), reflecting substantial phenotypic and mechanistic overlap.
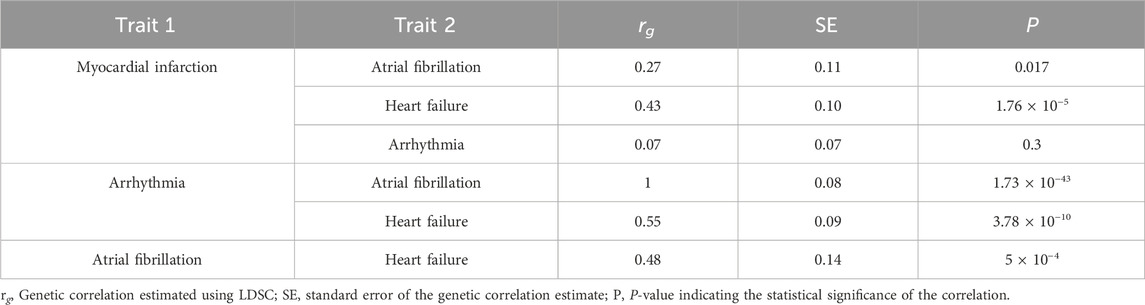
Table 1. Genetic correlations among cardiovascular diseases.
3.3 Partitioning heritability and chromatin marker enrichment in cardiovascular diseases and cardiometabolic traits
s-LDSC analysis demonstrated significant heritability enrichment for cardiovascular and cardiometabolic traits in cardiac and vascular tissues, particularly the left ventricle, right atrium, aorta, coronary artery, and tibial artery (Supplementary Figure S1). Among these, MI and SBP exhibited the highest enrichment in coronary and aortic tissues, respectively, consistent with their biological roles. Genomic regions bearing active chromatin marks—such as H3K27ac and H3K4me3—in these tissues were preferentially enriched, indicating that regulatory elements, including enhancers and promoters active in cardiomyocytes and endothelial cells, are likely to harbor causal variants. Notably, the enrichment profiles were broadly consistent across MI and related cardiometabolic traits, pointing to shared regulatory architecture and suggesting that pleiotropic effects may be mediated by tissue-specific regulatory programs. These results underscore the importance of cardiovascular tissues in shaping the genetic basis of MI and its metabolic risk factors, and emphasize the value of investigating the functional roles of these tissue-specific regulatory elements.
3.4 Local genetic correlations between cardiovascular diseases and cardiometabolic traits
To further explore the genetic interplay between cardiovascular diseases and cardiometabolic traits, we conducted a regional analysis of local genetic correlations. This analysis revealed several loci with statistically significant genetic overlap between CVD phenotypes and metabolic risk traits. For instance, a positive local genetic correlation was observed between MI and HDL cholesterol on chromosome 12 (Supplementary Table S3), a region enriched with genes implicated in lipid metabolism, including HNF1A (Ai-Ghalayini et al., 2020), which is known to affect HDL levels and coronary artery disease risk. In contrast, we identified negative local correlations between MI and LDL cholesterol at multiple loci. On chromosome 9 (Supplementary Table S3), the region includes ABO, a gene involved in coagulation and lipid regulation (Li et al., 2015), and SURF4, which facilitates the secretion of lipoproteins such as VLDL and LDL (Wang et al., 2021). Two additional regions on chromosome 19 were noteworthy: the first (positions 9,383,877–11,849,449; Supplementary Table S3) encompasses LDLR, a gene central to LDL clearance and cardiovascular risk (Franceschini et al., 2009); the second (positions 43,862,455–45,579,043; Supplementary Table S3) contains the APOE gene cluster (APOE, APOC1, APOC2, and APOC4), all of which play essential roles in lipid transport and homeostasis. Notably, genetic variants in APOE have been consistently associated with LDL levels and predisposition to cardiovascular disease (Chaudhary et al., 2012).
3.5 Multi-trait GWAS analysis identifies pleiotropic loci for cardiovascular diseases and cardiometabolic traits
To identify pleiotropic loci contributing to CVDs and related cardiometabolic traits, we conducted a multi-trait GWAS using MTAG across MI, HF, SBP, DBP, and other relevant traits. This analysis revealed several new genome-wide significant loci (P < 5 × 10−8) exhibiting consistent, albeit modest, associations across traits (Table 2). Additionally, we identified several previously unreported East Asian-specific loci characterized by substantially higher minor allele frequencies in East Asian populations compared to other populations (Supplementary Table S4). Notably, the locus tagged by rs13414987 (APOB) was jointly associated with HF and MI (P = 1.35 × 10−8), implicating a gene central to lipid metabolism and atherosclerosis (Supplementary Figure S2). Similarly, rs1942867 (MC4R) showed pleiotropic effects on HF and MI (P = 4.24 × 10−8), consistent with MC4R’s known role in energy homeostasis and obesity, both critical contributors to cardiac risk (Supplementary Figure S2). Of particular interest, rs3824359 in QSOX2 was significantly associated with HF and SBP (P = 7.06 × 10−10), a novel pleiotropic locus not previously reported in cardiometabolic GWAS (Supplementary Figure S2). To explore the functional relevance of rs3824359, we examined eQTL data from GTEx and found that this variant significantly influences QSOX2 expression across multiple tissues. The lead allele was associated with increased QSOX2 expression in cultured fibroblasts (P = 2.00 × 10−20), testis, tibial artery (P = 1.60 × 10−12), and subcutaneous adipose tissue, as well as in cardiovascular-relevant tissues including the heart (left ventricle and atrial appendage), skeletal muscle, and arterial vasculature (Supplementary Table S5). Notably, the variant also modulated gene expression in esophageal and brain tissues, suggesting a broad regulatory footprint. Given QSOX2’s role in oxidative protein folding and redox homeostasis, its upregulation in vascular and cardiac tissues could implicate reactive oxygen species modulation as a shared pathway in HF and blood pressure regulation. These findings nominate QSOX2 as a previously unrecognized contributor to cardiovascular pathophysiology and highlight the potential of pleiotropic loci to reveal convergent molecular mechanisms underlying complex traits.
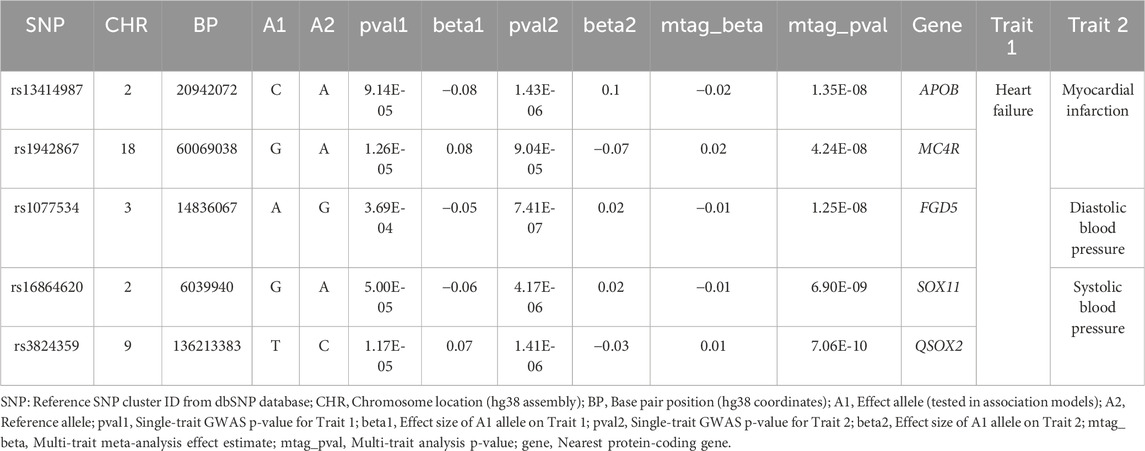
Table 2. New pleiotropic loci identified in multi-trait GWAS of heart failure.
Moreover, the region encompassing GUCY1A1 and GUCY1B1—encoding key subunits of soluble guanylate cyclase (sGC), the intracellular receptor for nitric oxide—emerged as a candidate with dual associations to both MI and blood pressure traits (Table 3). This pathway mediates cGMP synthesis, promoting vasodilation and vascular tone regulation. Regional association analyses revealed evidence of allelic heterogeneity, with at least two statistically independent association peaks, implying the presence of multiple functional variants or distinct regulatory elements within this locus (Figure 2). Prior studies have linked this region to blood pressure phenotypes, and the cumulative burden of risk alleles in this region has been associated with heightened susceptibility to stroke and coronary disease. Our eQTL investigation using GTEx data further supports the functional relevance of this locus, as risk alleles were associated with reduced expression of GUCY1A1 and GUCY1B1 in arterial tissues, potentially impairing NO-sGC signaling and contributing to vascular dysfunction (Supplementary Table S5). These findings underscore the mechanistic convergence between endothelial signaling, hemodynamic regulation, and ischemic cardiac events.
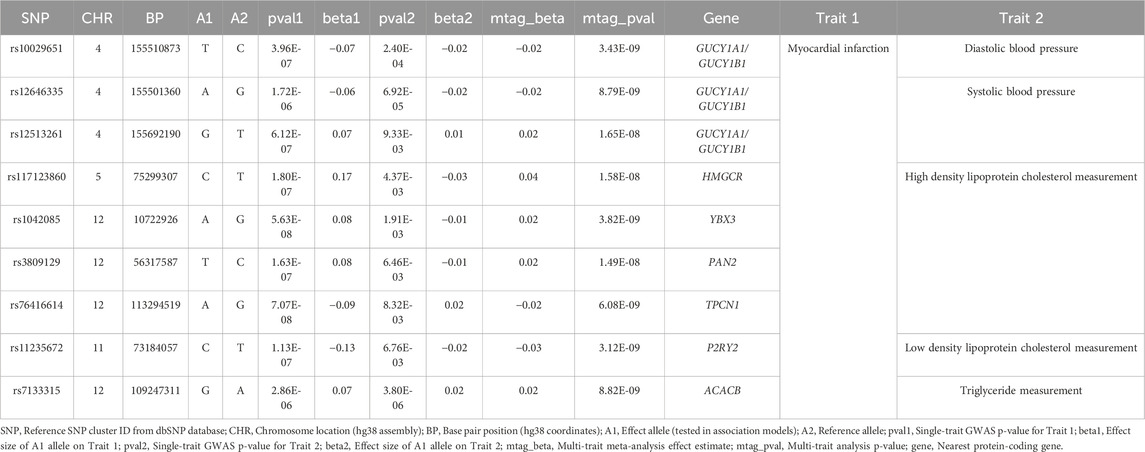
Table 3. New pleiotropic loci identified in multi-trait GWAS of myocardial infarction.
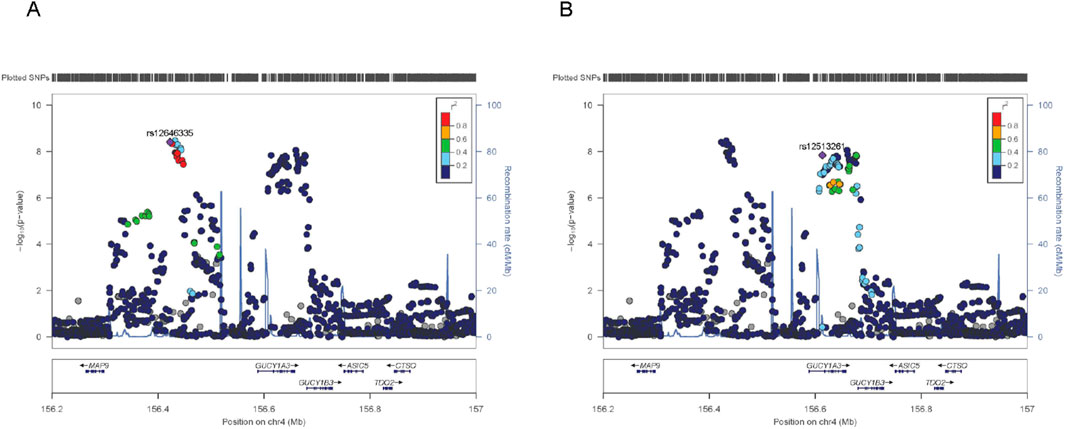
Figure 2. Regional association plots at the GUCY1A1/GUCY1B1 locus. (A) Association plot centered on signal 1, using the lead SNP rs12646335 as the index variant. (B) Association plot centered on signal 2, using the lead SNP rs12513261as the index variant. Each plot displays SNPs within the GUCY1A1/GUCY1B1 locus on chromosome 4, colored by their linkage disequilibrium (LD, measured as r2) with the respective lead SNP, based on the East Asian reference panel from the 1000 Genomes Project. SNPs in strong LD (r2 ≥ 0.8) are shown in red, moderate LD (0.6 ≤ r2 < 0.8) in orange, weak LD (0.4 ≤ r2 < 0.6) in green, and very low LD (r2 < 0.4) in light blue. Variants lacking LD information are shown in grey. Gene annotations and local recombination rates (blue curves) are provided for genomic context. Although both signals map to the same genomic region, their distinct LD structures suggest the presence of multiple independent regulatory elements at the GUCY1A1/GUCY1B1 locus.
4 Discussion
In this study, we systematically dissected the shared genetic architecture of CVDs and cardiometabolic traits in East Asian populations through a multi-layered analytical framework. By integrating genome-wide genetic correlation analyses, local heritability mapping, tissue-specific enrichment profiling, and multi-trait genome-wide association studies, we identified several pleiotropic loci that illuminate converging biological mechanisms underlying cardiovascular and metabolic risk.
Our genetic correlation results affirm extensive shared heritability between key cardiovascular conditions. The strong correlations between HF and both systolic and diastolic blood pressure, as well as the consistent links between MI and LDL, TG, and HDL, support the notion that vascular dysfunction and lipid dysregulation are foundational to CVD pathogenesis. In contrast, the weaker correlations between arrhythmia and lipid traits suggest partially distinct genetic pathways for electrophysiological disorders. Our analysis also revealed extensive genetic connectivity among four major CVD phenotypes suggesting that these conditions, though clinically distinct, are influenced by overlapping genetic factors in East Asians. Notably, HF exhibited the strongest and most consistent genetic correlations with MI, AF and arrhythmia, underscoring its central role at the intersection of structural, electrical, and ischemic heart disease (Winata et al., 2025). These findings reflect clinical observations that HF often co-occurs with rhythm disturbances and evolves downstream of MI, serving as both a consequence and an amplifier of other cardiovascular pathologies. While MI showed moderate correlation with AF and strong correlation with HF, arrhythmia and AF were nearly genetically indistinguishable (rg = 1.0), consistent with their shared electrophysiological basis. Taken together, these results support a model in which HF acts as a genetically and pathophysiologically integrative phenotype, bridging hemodynamic overload, myocardial injury, and conduction abnormalities.
Critically, our MTAG analysis yielded several genome-wide significant pleiotropic loci, including established cardiovascular genes and novel signals with potential functional relevance. APOB encodes apolipoprotein B, the primary structural protein of LDL, which plays a central role in lipid transport and the formation of atherosclerotic plaques. Variants in APOB have been repeatedly linked to altered LDL-C levels and coronary artery disease risk across ancestries. In our analysis, rs13414987 at the APOB locus was jointly associated with both HF and MI, highlighting the gene’s pleiotropic effect on vascular integrity and downstream cardiac remodeling. This finding reinforces the contribution of lipid dysregulation to both atherosclerotic burden and cardiac decompensation, and suggests that APOB-mediated pathways may serve as early upstream determinants of adverse cardiovascular outcomes (Giannakopoulou et al., 2025; Vorster et al., 2025). Similarly, MC4R (melanocortin 4 receptor), a G protein–coupled receptor expressed predominantly in the hypothalamus, is known for its key role in regulating appetite and energy homeostasis. Common and rare variants in MC4R have been implicated in obesity, metabolic syndrome, and type 2 diabetes. In our study, the MC4R locus was significantly associated with both HF and MI, suggesting a broader influence of central energy regulation on cardiovascular risk. This observation supports the concept that neuroendocrine control of body weight and energy balance exerts long-term effects on cardiac structure and function, potentially through mechanisms involving insulin resistance, adiposity-driven inflammation, and neurohumoral activation (Guo et al., 2025). Notably, MC4R has also been linked to blood pressure regulation, further implicating this locus as a point of intersection between metabolic and hemodynamic stressors.
Beyond these well-characterized genes, we also identified novel pleiotropic signals with functional potential. Among them, QSOX2 emerged as a previously unrecognized locus significantly associated with both HF and SBP. eQTL data revealed that rs3824359 modulates QSOX2 expression in multiple tissues including arteries and the heart, suggesting a role for oxidative protein folding and redox balance in vascular homeostasis and cardiac stress response (Ning et al., 2024). Similarly, we identified distinct association peaks at the GUCY1A1/GUCY1B1 locus, implicating reduced expression of nitric oxide receptor components in arterial tissues as a potential shared mechanism linking MI and elevated blood pressure (Vishnolia et al., 2021). These findings extend prior evidence on NO–sGC–cGMP signaling and offer mechanistic insights into the vascular origins of ischemic and hypertensive disease (Russo et al., 2024; Gawrys et al., 2025).
Our study has several limitations that should be acknowledged. First, although we identified multiple novel pleiotropic loci in East Asian populations, we were unable to perform replication in independent cohorts due to the limited availability of comparable datasets with matched phenotypes. Second, the GWAS summary statistics used in our analyses were derived from different studies, which may vary in terms of case definitions, diagnostic criteria, and phenotype ascertainment. Such heterogeneity in phenotype definitions could introduce noise and reduce the precision of association signals. Third, while some of the identified loci have been reported in European or other populations, many appear to be specific or stronger in East Asians. These differences may reflect ancestry-specific allele frequencies or linkage disequilibrium patterns, limiting the generalizability of our findings. Future studies involving harmonized phenotyping and multi-ancestry analyses will be critical to validate these associations and assess their relevance across diverse populations.
In conclusion, our study provides a comprehensive analysis of the genetic relationships among major CVDs and their cardiometabolic risk factors in East Asian populations. The identification of shared loci and enriched biological pathways emphasizes the interconnected nature of cardiovascular and metabolic disorders and highlights the need for integrative risk prediction and therapeutic strategies. Future efforts incorporating multi-omic data and trans-ancestry replication will be crucial to further refine these insights and translate them into precision medicine applications.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding authors.
Ethics statement
Written informed consent was obtained from the individual(s) for the publication of any potentially identifiable images or data included in this article.
Author contributions
PZ: Conceptualization, Funding acquisition, Writing – original draft, Formal Analysis, Data curation. CZ: Resources, Formal Analysis, Data curation, Writing – review and editing. QW: Software, Visualization, Writing – review and editing, Data curation. XC: Supervision, Writing – original draft, Project administration, Investigation, Writing – review and editing, Funding acquisition.
Funding
The author(s) declare that no financial support was received for the research and/or publication of this article.
Acknowledgments
We thank the GWAS Catalog project for providing valuable resources and data that contributed to this study.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fgene.2025.1635378/full#supplementary-material
References
Ai-Ghalayini, K. W., Salama, M. A., Al Mahdi, H. B., Al-Harthi, S., Alhejily, W. A., Alasnag, M. A., et al. (2020). Identification of genetic variants associated with myocardial infarction in Saudi Arabia. Heart Surg. forum 23, E517–E523. doi:10.1532/hsf.2955
Bulik-Sullivan, B., Finucane, H. K., Anttila, V., Gusev, A., Day, F. R., Loh, P. R., et al. (2015). An atlas of genetic correlations across human diseases and traits. Nat. Genet. 47, 1236–1241. doi:10.1038/ng.3406
Buniello, A., MacArthur, J. A. L., Cerezo, M., Harris, L. W., Hayhurst, J., Malangone, C., et al. (2019). The NHGRI-EBI GWAS catalog of published genome-wide association studies, targeted arrays and summary statistics 2019. Nucleic Acids Res. 47, D1005–D1012. doi:10.1093/nar/gky1120
Chaudhary, R., Likidlilid, A., Peerapatdit, T., Tresukosol, D., Srisuma, S., Ratanamaneechat, S., et al. (2012). Apolipoprotein E gene polymorphism: effects on plasma lipids and risk of type 2 diabetes and coronary artery disease. Cardiovasc. Diabetol. 11, 36. doi:10.1186/1475-2840-11-36
Chen, C. Y., Chen, T. T., Feng, Y. C. A., Yu, M., Lin, S. C., Longchamps, R. J., et al. (2023). Analysis across Taiwan Biobank, Biobank Japan, and UK Biobank identifies hundreds of novel loci for 36 quantitative traits. Cell genomics 3, 100436. doi:10.1016/j.xgen.2023.100436
Finucane, H. K., Bulik-Sullivan, B., Gusev, A., Trynka, G., Reshef, Y., Loh, P. R., et al. (2015). Partitioning heritability by functional annotation using genome-wide association summary statistics. Nat. Genet. 47, 1228–1235. doi:10.1038/ng.3404
Franceschini, N., Muallem, H., Rose, K. M., Boerwinkle, E., and Maeda, N. (2009). Low density lipoprotein receptor polymorphisms and the risk of coronary heart disease: the atherosclerosis risk in communities study. J. thrombosis haemostasis JTH 7, 496–498. doi:10.1111/j.1538-7836.2008.03262.x
Gawrys, O., Kala, P., Sadowski, J., Melenovský, V., Sandner, P., and Červenka, L. (2025). Soluble guanylyl cyclase stimulators and activators: promising drugs for the treatment of hypertension? Eur. J. Pharmacol. 987, 177175. doi:10.1016/j.ejphar.2024.177175
Giannakopoulou, S. P., Antonopoulou, S., Barkas, F., Liberopoulos, E., Chrysohoou, C., Sfikakis, P. P., et al. (2025). Concordance-discordance between apolipoprotein B and lipid biomarkers in predicting 20-year atherosclerotic cardiovascular disease risk: the ATTICA study (2002-2022). Eur. J. Clin. investigation, e70077. doi:10.1111/eci.70077
Guo, D. F., Williams, P. A., Olson, A., Morgan, D. A., Herz, H., Resch, J., et al. (2025). Loss of MRAP2 in MC4R neurons protect from obesity-associated autonomic and cardiovascular dysfunctions. Cardiovasc. Res., cvaf067. doi:10.1093/cvr/cvaf067
Hartiala, J. A., Han, Y., Jia, Q., Hilser, J. R., Huang, P., Gukasyan, J., et al. (2021). Genome-wide analysis identifies novel susceptibility loci for myocardial infarction. Eur. Heart J. 42, 919–933. doi:10.1093/eurheartj/ehaa1040
Kim, S. J. (2021). Global awareness of myocardial infarction symptoms in general population. Korean circulation J. 51, 997–1000. doi:10.4070/kcj.2021.0320
Li, H., Song, X., Liang, Y., Bai, X., Liu-Huo, W. S., Tang, C., et al. (2022). Global, regional, and national burden of disease study of atrial fibrillation/flutter, 1990-2019: results from a global burden of disease study, 2019. BMC public health 22, 2015. doi:10.1186/s12889-022-14403-2
Li, S., Xu, R. X., Guo, Y. L., Zhang, Y., Zhu, C. G., Sun, J., et al. (2015). ABO blood group in relation to plasma lipids and proprotein convertase subtilisin/kexin type 9. Nutr. metabolism, Cardiovasc. Dis. NMCD 25, 411–417. doi:10.1016/j.numecd.2014.10.015
Ning, Z., Huang, Y., Lu, H., Zhou, Y., Tu, T., Ouyang, F., et al. (2024). Novel drug targets for atrial fibrillation identified through Mendelian randomization analysis of the blood proteome. Cardiovasc. drugs Ther. 38, 1215–1222. doi:10.1007/s10557-023-07467-8
Packer, M. (2020). What causes sudden death in patients with chronic heart failure and a reduced ejection fraction? Eur. heart J. 41, 1757–1763. doi:10.1093/eurheartj/ehz553
Russo, P., Vitiello, L., Milani, F., Volterrani, M., Rosano, G. M. C., Tomino, C., et al. (2024). New therapeutics for heart failure worsening: focus on vericiguat. J. Clin. Med. 13, 4209. doi:10.3390/jcm13144209
Shi, H., Mancuso, N., Spendlove, S., and Pasaniuc, B. (2017). Local genetic correlation gives insights into the shared genetic architecture of complex traits. Am. J. Hum. Genet. 101, 737–751. doi:10.1016/j.ajhg.2017.09.022
Trynka, G., Sandor, C., Han, B., Xu, H., Stranger, B. E., Liu, X. S., et al. (2013). Chromatin marks identify critical cell types for fine mapping complex trait variants. Nat. Genet. 45, 124–130. doi:10.1038/ng.2504
Turley, P., Walters, R. K., Maghzian, O., Okbay, A., Lee, J. J., Fontana, M. A., et al. (2018). Multi-trait analysis of genome-wide association summary statistics using MTAG. Nat. Genet. 50, 229–237. doi:10.1038/s41588-017-0009-4
Vishnolia, K. K., Rakovic, A., Hoene, C., Tarhbalouti, K., Aherrahrou, Z., and Erdmann, J. (2021). sGC activity and regulation of blood flow in a zebrafish model system. Front. physiology 12, 633171. doi:10.3389/fphys.2021.633171
Vorster, A., Kruger, R., Mels, C. M., and Breet, Y. (2025). Cardiovascular disease risk factors are associated with conventional lipids and apolipoproteins in South African adults of African ancestry. Lipids health Dis. 24, 177. doi:10.1186/s12944-025-02591-w
Winata, I. G., Immanuel, S. S., Leonardo, L., Rinaldi, F. X., Tandecxi, G., and Wijaya, R. (2025). Examining the interplay between endometriosis and later-life cerebro-cardiovascular diseases: a systematic review, meta-analysis, and trial sequential analysis. Narra J. 5, e1935. doi:10.52225/narra.v5i1.1935
Wang, B., Shen, Y., Zhai, L., Xia, X., Gu, H. M., Wang, M., et al. (2021). Atherosclerosis-associated hepatic secretion of VLDL but not PCSK9 is dependent on cargo receptor protein Surf4. J. lipid Res. 62, 100091. doi:10.1016/j.jlr.2021.100091
Keywords: heart failure, cardiovascular diseases, multi-trait GWAS, cardiometabolic traits, genetic correlation
Citation: Zhong P, Zhang C, Wu Q and Chang X (2025) Shared genetic loci connect cardiovascular disease with blood pressure and lipid traits in East Asian populations. Front. Genet. 16:1635378. doi: 10.3389/fgene.2025.1635378
Received: 26 May 2025; Accepted: 13 June 2025;
Published: 24 June 2025.
Edited by:
Hui-Qi Qu, Children’s Hospital of Philadelphia, United StatesReviewed by:
Baohong Liu, Chinese Academy of Agricultural Sciences, ChinaXuming Zhu, Icahn School of Medicine at Mount Sinai, United States
Copyright © 2025 Zhong, Zhang, Wu and Chang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Qinfeng Wu, MjU0MzgxNjcxNkBxcS5jb20=; Xiao Chang, Y2hhbmd4aWFvQHNkZm11LmVkdS5jbg==