 Rénald Belley
Rénald Belley Paul V. R. Snelgrove
Paul V. R. Snelgrove- Departments of Ocean Sciences and Biology, Memorial University, St. John's, Newfoundland and Labrador, Canada
Current concern about biodiversity change associated with human impacts has raised scientific interest in the role of biodiversity in ecosystem functioning. However, studies on this topic face the challenge of evaluating and separating the relative contributions of biodiversity and environment to ecosystem functioning in natural environments. To investigate this problem, we collected sediment cores at different seafloor locations in Saanich Inlet and the Strait of Georgia, British Columbia, Canada, and measured benthic fluxes of oxygen and five nutrients (ammonium, nitrate, nitrite, phosphate, and silicate). We also measured 18 environmental variables at each location, identified macrofauna, and calculated a suite of species and functional diversity indices. Our results indicate that, examined separately, macrobenthic functional richness (FRic) predicted benthic flux better than species richness, explaining ~ 20% of the benthic flux variation at our sites. Environmental variables and functional diversity indices collectively explained 62.9% of benthic flux variation, with similar explanatory contributions from environmental variables (21.4%) and functional diversity indices (18.5%). The 22.9% shared variation between environmental variables and functional diversity indices demonstrate close linkages between species and environment. Finally, we also identified funnel feeding as a key functional group represented by a small number of species and individuals of maldanid and pectinariid polychaetes, which disproportionately affected benthic flux rates relative to their abundance. Our results indicate the primary importance of environment and functional diversity in controlling ecosystem functioning. Furthermore, these results illustrate the consequences of anthropogenic impacts, such as biodiversity loss and environmental changes, for ecosystem functioning.
Introduction
The loss of biodiversity and its impact on humanity (Cardinale et al., 2012) have raised considerable interest on potential links between biodiversity and ecosystem functioning in a wide range of ecosystems (Loreau et al., 2001, 2002; Solan et al., 2004; Loreau, 2010). This work points to a strong role for functional groups in controlling ecosystem functions (Hooper et al., 2005; Cardinale et al., 2006; Danovaro et al., 2008) but also a potential role for environment (Yachi and Loreau, 1999; Godbold and Solan, 2009; Belley et al., 2016). Although most of these studies focus on biodiversity loss by manipulating species in experiments (Cardinale et al., 2012; Naeem et al., 2012), natural gradients in environments offer an alternative “in situ” approach to linking function, biodiversity, and environment (Snelgrove et al., 2014).
In the world's ocean, seafloor habitats and the organisms that reside in and on marine sediments provide important ecosystem functions. These include recycling of organic matter that drives benthic-pelagic coupling and fuels surface waters with nutrients essential for primary production (Snelgrove et al., 2014). Despite a general consensus that biodiversity and environmental factors may both play a role in benthic ecosystem functioning, relatively few studies have attempted to separate abiotic and biotic contributions to ecosystem functioning. In fact, manipulative laboratory studies (i.e., changing species composition but not environmental conditions) typically underestimate the contribution of biodiversity to ecosystem functions (Duffy, 2009; Godbold, 2012). Nonetheless, studies that have tried to separate abiotic and biotic contributions generally found that both played an important role (Hiddink et al., 2009; Queirós et al., 2011; Godbold, 2012; Braeckman et al., 2014; Strong et al., 2015).
Measurements of benthic fluxes at the sediment-water interface offer one means of quantifying organic matter remineralization, an important ecosystem function in seafloor habitats (Giller et al., 2004). Multiple biological and environmental factors influence benthic fluxes. Previous studies point to the importance of environmental variables such as temperature (Hargrave, 1969; Cowan et al., 1996; Alonso-Pérez and Castro, 2014), and the quality and quantity of organic matter sinking to the seafloor (Berelson et al., 1996; Jahnke, 1996). Previous studies also report a strong positive influence of biological factors such as the presence of bio-irrigators and bioturbators on benthic fluxes and organic matter remineralization (Aller, 1982, 2014; Aller and Aller, 1998), and that focus has expanded to consider the importance of functional diversity on ecosystem functioning (Snelgrove et al., 1997, 2014; Raffaelli et al., 2003; Solan et al., 2004). Indeed, some studies report that functional diversity, defined as “the value and range of those species and organismal traits that influence ecosystem functioning” (Tilman, 2001), promotes organic matter remineralization and consequently, increases benthic fluxes (Braeckman et al., 2014).
Our study focuses on four different sites in the Salish Sea, a semi-enclosed sea between Vancouver Island and British Columbia, Canada (Figure 1). The large variation in species diversity (Macdonald et al., 2012) and environmental variables (Johannessen et al., 2005; Masson and Cummins, 2007) within the Salish Sea over a relatively small spatial scale provides an ideal location for a study that uses natural gradients to identify the influences of biodiversity and environment on ecosystem functioning. Saanich Inlet, a seasonally hypoxic fjord, supports a relatively low diversity benthic community that specializes on low-oxygen environments (Tunnicliffe, 1981; Matabos et al., 2012; Chu and Tunnicliffe, 2015). Strong seasonal variation in dissolved oxygen concentrations and temperature also characterizes the Strait of Georgia (Masson and Cummins, 2007; Johannessen et al., 2014). Finally, the Delta Dynamic Laboratory site within the Strait of Georgia offers a highly dynamic environment characterized by high organic and inorganic loading resulting from its proximity to the Fraser River outflow (Burd et al., 2008; Macdonald et al., 2012).
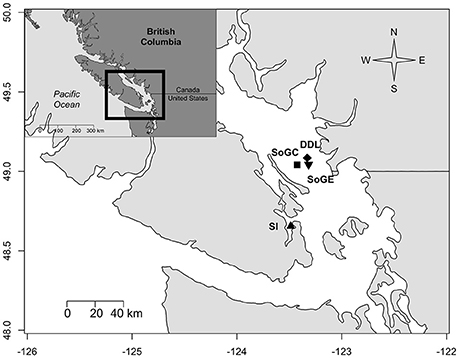
Figure 1. Map of stations sampled in Saanich Inlet and in the Strait of Georgia, British Columbia, Canada. Delta Dynamic Laboratory (DDL) and Strait of Georgia Central (SoGC) were sampled in July 2011. Saanich Inlet (SI) sampling occurred in July 2011 and September 2013, and Strait of Georgia East in May 2011 and September 2013.
The primary objective of this comparative field study was to evaluate the contributions of species and functional diversities, and environmental variables to benthic fluxes of oxygen and nutrients (ammonium, nitrate, nitrite, phosphate, and silicate) at contrasting sites. We addressed our objective by exploring the following questions at our study sites: (i) do benthic fluxes vary spatially, (ii) does benthic community composition vary spatially, (iii) which environmental variables explain benthic flux variation and remineralization, (iv) which species and functional diversity indices, if any, explain benthic flux variation and remineralization, and (v) how much benthic flux variation do biodiversity and environmental variables explain, respectively? This study builds from Belley et al. (2016), which investigated the effects of environmental variables on benthic flux variation at these and other deeper sites in the Northeast Pacific ocean.
Methods
Field Sampling
Samples were collected near the VENUS Observatory nodes in Saanich inlet and the Strait of Georgia, British Columbia, Canada (Figure 1). We collected push core sediments using the Remotely Operated Vehicle (ROV) ROPOS (www.ropos.com) on board the Canadian Coast Guard Ship John P. Tully (May 7–14, 2011), and the Research Vessels Thomas G. Thompson (June 30-July 3, 2011) and Falkor (September 6–18, 2013). Sampling occurred at the VENUS Delta Dynamic Laboratory (DDL) and the Strait of Georgia Central (SoGC) sites in July 2011, Saanich Inlet (SI) in July 2011 and September 2013, and the Strait of Georgia East (SoGE) in May 2011 and September 2013 (Table 1). The ROV collected 4–5 push-cores at each site (i.d. = 6.7 cm, L = 35.6 cm) at random locations within a bottom area that spanned ~ 25 × 25 m near the observatory nodes. One core per site served to determine prokaryotic cell abundance and biomass as well as sediment properties (summarized in Table 1), and the remaining cores were used for incubations to measure fluxes. A SBE 19plus V2 CTD mounted on the ROV recorded near-bottom dissolved oxygen (DO), temperature, and salinity. No specific permissions were required for these locations/activities and field studies did not involve endangered or protected species. Below we provide a brief overview of methodologies, but a more detailed description can be found in Belley et al. (2016).
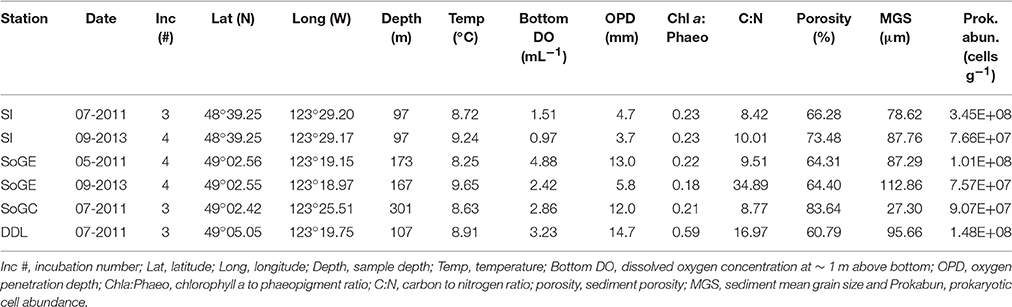
Table 1. Station names, sampling dates, number of incubations performed, locations, and environmental variables measured.
Incubations
At each sampling site, we acclimated 3–4 sediment cores (0.68 L ± 0.10 and 0.42 L ± 0.10, mean volume ± SD of sediment and water, respectively) for 4–24 h, allowing sufficient time for any sediment particles in suspension to settle back to the sediment surface. We used different acclimation times because of the high sampling intensity, tight dive schedule, and limited number of incubations that could be run simultaneously, but all acclimation times fall within the range of acclimation times typically reported in the literature (Valdemarsen et al., 2012; Link et al., 2013a; Nunnally et al., 2013). Moreover, comparison of results in relation to acclimation times showed no consistent pattern. We aerated the overlying water in each core for a minimum of 1 h using aquarium air pumps to avoid suboxic conditions during incubations. A previous study in this region revealed no significant effect of the overlying water aeration on benthic flux rate measurements (Belley et al., 2016). Sediment cores were then sealed with caps equipped with magnetic stirrers and gas-tight sampling ports, prior to incubating in the dark at in situ temperatures (8–9°C) for 12–24 h until 15–30% of available oxygen was consumed. Hall et al. (1989) showed a linear decrease in oxygen during the initial stage of incubations, which therefore provided reliable estimates of oxygen uptake rates.
Oxygen Uptake
We measured oxygen consumption periodically (4–8 h intervals) using a 500-μm diameter oxygen microsensor (Unisense, Aarhus, Denmark) inserted through a small resealable hole on the top of the cap in May and July 2011, and with a non-invasive optical oxygen meter used in conjunction with oxygen optode patches (Fibox 4, PreSens, Regensburg, Germany) in September 2013. PreSens provided calibration details for each oxygen optode patch and we used the two-point calibration method for the oxygen microsensors recommended by Unisense, aerating water collected in situ for a minimum of 5 min and taking readings only after the signal stabilized for the fully saturated reading. The zero reading was obtained using a solution of sodium ascorbate and NaOH, both at final concentrations of 0.1 M. We determined oxygen uptake from the slope of the linear regression of oxygen concentration vs. time of incubations after correction for oxygen concentration in the replacement water (see example in Appendix 1 of Supplementary Material).
Nutrient Fluxes
At the beginning, midpoint, and end of the incubations we collected water samples with 60-mL, acid-rinsed plastic syringes, except in the SI, SoGC, and DDL incubations in July 2011, where high oxygen consumption shortened the incubation period to 12 h and we limited water sampling to the beginning and end of the incubations. We immediately replaced withdrawn water with an equivalent volume of bottom water of known oxygen and nutrient concentrations. Syringes and sample containers were initially rinsed with ~5 mL of sample water. At each sampling time we collected and stored two 25-mL water samples in acid-rinsed twist-cap 30-mL HDPE bottles. Upon collection, water samples were immediately placed in an upright position at −20°C until analyzed. We determined the concentrations of nutrients (, , , Si(OH)4, ) in the water samples using a Technicon Segmented Flow AutoAnalyzer II, following the method recommended by Technicon Industrial Systems (1973, 1977, 1979) with the exception of ammonia (hereafter referred as ammonium) analysis, which followed Kerouel and Aminot (1997). Nutrient fluxes were determined from the slope of the linear regression of nutrient concentrations vs. time of incubations after correction for the solute concentration in replacement water (see example in Appendix 1 of Supplementary Material).
Macrofaunal Identification and Taxonomic Diversity
After incubations, sediment cores were sectioned onto 0–2, 2–5, and 5–10 cm layers and processed over a 300 μm sieve prior to preservation in a 4% seawater-formaldehyde solution and subsequent transfer to 70% ethanol for identification. Specimens were sorted under a dissection microscope in the laboratory and identified to the lowest possible taxonomic level, usually to species. We determined abundance (N) for each taxon and taxonomic richness (S) as the number of taxa present in each sediment core. We also determined diversity indices including Simpson's index (Simp or 1 - D), Pielou's evenness (J'), Rarefaction (es25), and the Shannon-Wiener index (H') for each sediment core. All analyses presented in this study were performed on data from whole cores (0–10 cm), not in separate layers. Diversity indices were computed in R (R Core Team, 2016) using the package “vegan” (Oksanen et al., 2013).
Biological Traits and Functional Diversity
We selected five biological traits and 24 modalities based on their presumed influence on benthic fluxes and availability for all taxa (Table 2). These reflected behavior (bioturbation mode, feeding type, habitat, and mobility) and morphology (size). Biological traits were collected for each taxon from published sources (MarLIN, 2006; Macdonald et al., 2010; Link et al., 2013b; Queirós et al., 2013; Jumars et al., 2015; WoRMS Editorial Board, 2015). When biological traits information was unavailable for a specific taxon, we obtained information from one taxonomic rank higher. For example, the absence of species-specific information on the feeding type of the crustacean Diastylis abboti required us to use genus-level information. We used fuzzy coding that allowed more than one functional trait for a given taxon for each category, and scored from 0 to 1 based on the extent to which they displayed each trait. For example, the polychaete Paraprionospio pinnata can alternate between filter and surface deposit feeding depending on environmental conditions, so these two traits each scored 0.5 for the feeding type category. Trait category scores for each taxon and taxa abundance matrices were used to obtain functional diversity (FD) indices using the “FD” package (Laliberté and Legendre, 2010) in R (R Core Team, 2016). We then computed the following multidimensional FD indices for use in our analyses: functional richness (FRic), functional evenness (FEve), functional divergence (FDiv) (Villéger et al., 2008), functional dispersion (FDis) (Laliberté and Legendre, 2010), Rao's quadratic entropy (RaoQ) (Botta-Dukat, 2005), and an index of functional composition, the community-level weighted means of trait values (CWM) (Lavorel et al., 2008). In the statistical analyses described below, we included CWM as diversity variables because they provide measures of the range and distribution of functional traits value in sediment cores, and therefore represent good indicators of functional diversity (Lavorel et al., 2008).

Table 2. Biological traits used in the functional diversity analysis.
Oxygen Penetration Depth (OPD)
Immediately after recovery of the ROV we profiled oxygen concentrations as a function of depth in the sediment for one sediment core from each site. In each core, we performed three replicate profiles with Unisense oxygen microsensors (500 and 250 μm tip sizes in 2011 and 2013, respectively) in vertical increments of 1000 and 500 μm in 2011 and 2013, respectively. We defined the oxygen penetration depth (OPD) in the sediment as the mean depth at which oxygen concentration decreased below the suboxic level of 5 μmol L−1 (Thibodeau et al., 2010).
Prokaryotic Cells
We subcored the sediment cores with a cut off 10-mL sterile plastic syringe at depths of 0–2, 2–5, and 5–10 cm to sample sediment prokaryote abundances (hereafter abbreviated as prokabun). We placed 1 mL of sediment from each depth in a 20-mL scintillation vial containing 4 mL of a filtered-sterilized 2% seawater-formalin solution. Samples were frozen at −20°C until analysis. Sediment prokaryote abundance and biomass were determined following Danovaro (2010). In the statistical analyses described below, we included prokaryotic abundance as an environmental variable because we obtained site averages from one sample at each site and sampling time (i.e., core) as with other environmental variables; furthermore, we considered prokaryotic abundance a critical component of the biological environment for macrofauna.
Sediment Properties
We sectioned the upper 2-cm layer of sediment from one sediment core using inert plastic spatulas to characterize sediment properties. Each sediment layer was carefully placed in a Whirl-Pak bag and stored at −20°C until analyzed. We determined total organic matter (TOM) by ignition loss, and water content as the difference between the wet and dry sediment weights divided by the sediment wet weight (Danovaro, 2010). Sediment porosity and dry bulk density were calculated using formulas from Avnimelech et al. (2001) with a particle density of 2.65 g cm−3. We determined granulometric properties (sediment mean grain size; MGS) with a HORIBA Partica LA-950 laser diffraction particle size analyzer (Horiba Ltd. Kyoto. Japan). No sieving was performed prior to analysis because no large particles were present in our sediment samples. Samples were prepared for analyses of total organic carbon (TOC) and total nitrogen (TN) by drying for 24 h at 80°C, fuming with 1 M HCl for 24 h, and drying again for a minimum of 24 h. Finally, approximately 2 mg of sediment samples were weighed into a tin capsule and stored at 80°C until analyzed in a Perkin-Elmer 2400 Series II CHN analyzer. We used the carbon to nitrogen (C:N) mass ratio as a measure of organic matter nutritional quality on a long time scale (Le Guitton et al., 2015), where lower ratios indicate fresher and higher quality organic matter (Vidal et al., 1997; Godbold and Solan, 2009).
Chlorophyll-a and Phaeopigments
Concentrations of chlorophyll-a (chl a) and phaeopigments (phaeo) were quantified fluorimetrically following a modified version of Riaux-Gobin and Klein (1993). We placed 1–2 g of wet sediment in 90% acetone (v/v) at 4°C for 24 h and then analyzed the supernatant prior to and following acidification using a Turner Designs 10-AU-005-CE fluorometer (Turner Designs. Sunnyvale. USA). The remaining sediment was dried at 60°C for 24 h and weighed in order to standardize pigment concentrations per gram of sediment. The chl a:phaeo ratio provides a measure of organic matter quality on a short time scale (Le Guitton et al., 2015), where higher ratios indicate more recently settled phytoplankton particles and therefore fresher organic matter (Morata et al., 2011; Suykens et al., 2011).
Statistical Analyses
We examined spatial variation in benthic fluxes and taxonomic community composition using a permutational multivariate analysis of variance (PERMANOVA) performed with 9999 random permutations of appropriate units (Anderson, 2001; McArdle and Anderson, 2001). Previous benthic flux analyses (Belley et al., 2016) and preliminary analyses of benthic communities indicated no significant temporal variation at SI (July 2011 and September 2013) and SoGE (May 2011 and September 2013). We therefore grouped data from a single site collected on the two different occasions (i.e., SI and SoGE). Two separate analyses addressed two research questions: (1) a one-way PERMANOVA design using all benthic flux data with the factor “Site” (four levels: DDL, SI, SoGC and SoGE) tested for benthic flux spatial variation among sites, and (2) a one-way PERMANOVA design using all macrofaunal taxonomic data with the factor “Site” (four levels: DDL, SI, SoGC, and SoGE) tested for spatial variation in macrofaunal community composition among sites. Taxa that appeared only once were removed from the latter analysis (Clarke and Warwick, 1994), although this removal had little effect on overall patterns. We calculated the resemblance matrices from Euclidean distances of standardized benthic flux and from Bray-Curtis distances of fourth root transformed benthic community data. This transformation was applied to bring all taxa to a similar relative scale of abundance and therefore, increase the contribution of rare species (Anderson, 2001; Anderson et al., 2008). We verified homogeneity of multivariate dispersion using the PERMDISP routine (Anderson et al., 2008). When there were too few possible permutations for a meaningful test, we calculated a p-value based on 9999 Monte Carlo draws from the asymptotic permutation distribution (Terlizzi et al., 2005). We further analyzed significant terms within the full models using appropriate pair-wise comparisons. Multivariate patterns were visualized using non-metric multidimensional scaling (nMDS) ordinations of similarity matrices, and similarity percentage analyses (SIMPER) determined which taxa contributed most to dissimilarity between sites (Clarke, 1993). We completed nMDS, SIMPER, PERMANOVA, and PERMDISP analyses in PRIMER 6 (Clarke and Gorley, 2006) with the PERMANOVA+ add-on (Anderson et al., 2008).
We used two separate redundancy analyses (RDA) to identify the environmental variables (RDA #1) and functional diversity indices (RDA #2) that best explained benthic flux variation. RDA, a multivariate (i.e., multi-response) analysis, combines regression, and principal component analysis (PCA) and offers a key advantage over regression alone in that it determines which predictor variables explain the most variation in multiple response variables (Legendre and Legendre, 2012). First, RDA performs a multivariate multiple linear regression followed by a PCA of the fitted values. Therefore, it allows identification of linear combinations of variables that best explain the response matrix variation. Finally, RDA tests the significance of the explained variation using a permutation procedure (Legendre and Legendre, 2012). We used stepwise selection with a significance level of 0.05 and 9999 random permutations to obtain the model with the most parsimonious set of variables. This procedure can deal with a large number of explanatory variables by allowing only the selection of explanatory variables with the strongest contributions (Legendre and Legendre, 2012). Predictor variables containing outliers were transformed, and highly correlated (r > 0.85) predictor variables were excluded from the analyses (RDA #1 = chlorophyll-a, phaeopigments, total organic matter, nitrogen, water content, bulk density, and prokaryotic biomass; RDA #2 = Shannon-Wiener index, taxonomic richness, Rao's quadratic entropy, community-level weighted mean of epifauna). The optimal environmental model selection included 11 predictor variables: bottom water temperature, salinity, and dissolved O2 concentration, seafloor depth, sediment O2 penetration depth, chl a:phaeo ratio, carbon content, carbon:nitrogen ratio, porosity, mean grain size, and prokaryotic abundance. To correct for data skewness, we applied a natural logarithmic (Ln) transformation to three predictor variables (chl a:phaeo, carbon:nitrogen and prokaryotic abundance). The optimal diversity model selection included 25 predictor variables: abundance, Simpson's diversity, Pielou's evenness, Expected Species, functional richness, functional evenness, functional divergence, functional dispersion, community weighted means of carnivores, omnivores, scavengers, grazers, detritivores, filter feeders, surface, and sub-surface deposit feeders, funnel feeders, and predators, small, medium, and large-sized organisms, surficial modifiers, organisms with limited and slow movement through sediment, and infauna. We further analyzed multi-collinearity of the predictor variables from the full models with a variance inflation factor (VIF) test using the “vif” function from the “car” package (Fox and Weisberg, 2011), removing predictor variables with the highest VIF so that the best model selected contained only predictor variables with VIF < 5 and therefore, removed the negative effect of multi-collinearity on our results (Zuur et al., 2009). We verified the homogeneity of multivariate dispersion assumption using the PERMDISP routine (Anderson et al., 2008). Contributions of each predictor variable to benthic fluxes reported here are based on R2 and not on Adj. R2 calculations.
Finally, we performed variation partitioning (Legendre and Legendre, 2012) to determine relative contributions of environmental variables and functional diversity indices to benthic flux variation. Variation partitioning analysis allowed quantification of the portion of benthic flux variation explained by the two subsets of explanatory variables (diversity and environmental subsets) when controlling for the effect of the other subset. This is done by: (1) performing a RDA of the flux by diversity data (same steps as described for RDA#2 above), (2) performing a RDA of the flux by environmental data (same steps as described for RDA#1 above), (3) performing a RDA of the flux by diversity and environmental data, (4) computing the adjusted R2 of the three RDAs, and finally (5) computing fractions of adjusted variation by subtraction (Legendre and Legendre, 2012). Variation partitioning analysis is most often used when variables included in each RDA models differ at different scales (see examples in Legendre and Legendre, 2012).
We completed RDA and variation partitioning analyses in R (R Core Team, 2016) using the package “vegan” (Oksanen et al., 2013) and calculated the contribution of each predictor variable to benthic flux variation in PRIMER 6 (Clarke and Gorley, 2006) with the PERMANOVA+ add-on (Anderson et al., 2008).
Results
A total of 21 incubations spanned four different sites and three different time periods (Appendix 2 in Supplementary Material). In total, we identified 1942 specimens representing 119 different taxa (Appendix 3 in Supplementary Material). The most diverse and abundant animal Class was Polychaeta; the most abundant species, Mediomastus cf. californiensis (Capitellidae), occurred in highest densities in SoGE cores. The spionid Prionospio lighti also occurred in high densities, but mostly in the Strait of Georgia sites (i.e., DDL, SoGC, and SoGE). Malacostraca was the second most diverse and abundant animal Class; Cumella sp. (Cumacea), the most abundant taxon, occurred only at SoGE (Appendix 3 in Supplementary Material). SIMPER analysis revealed greatest dissimilarity between the SI benthic community and other sites (average dissimilarity of 73.8, 73.9, and 74.4% for SoGE, SoGC, and DDL, respectively). The taxa that contributed most to these dissimilarities were Cumella sp. (5.7% contribution), Levinsenia gracilis (5.9% contribution), and Prionospio lighti (6.4% contribution), which were more abundant at SoGE, SoGC, and DDL, respectively.
PERMANOVA indicated significant differences in benthic community assemblages among the four sampling sites and thus significantly greater variability in assemblages among sites than within sites [P (perm) < 0.01, Table 3]. Pair-wise comparisons showed significantly different benthic communities at each of our sampling sites [SI and SoGE, P (perm) < 0.001; SI and SoGC, P (perm) = 0.010; SI and DDL, P (perm) = 0.008; SoGE and SoGC, P (perm) = 0.006; SoGE and DDL, P (perm) = 0.005; SoGC and DDL, P (MC) = 0.039] (Figure 2A).
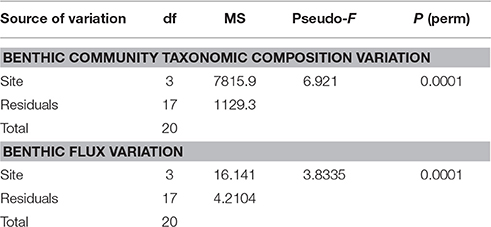
Table 3. Permutational analysis of variance (PERMANOVA) results testing the effect of site on benthic communities based on Bray-Curtis similarity matrices performed on fourth root transformed data, and on benthic fluxes based on Euclidean similarity matrices performed on normalized data.
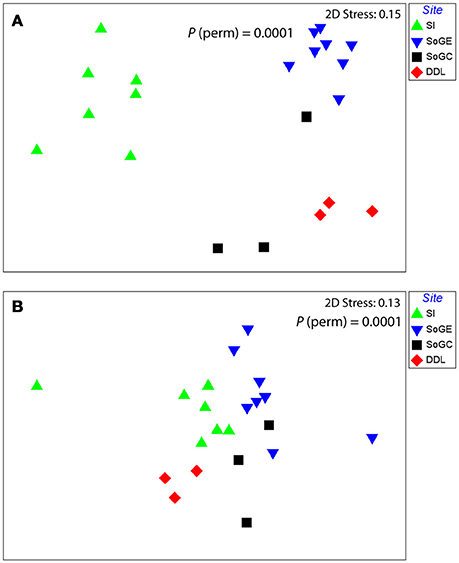
Figure 2. Non-metric multi-dimensional scaling (nMDS) plot of (A) benthic community taxonomic assemblages at each study site based on Bray-Curtis similarity matrices performed on fourth root transformed data, and (B) benthic fluxes at each study site based on Euclidean similarity matrices performed on normalized data.
PERMANOVA indicated significant differences in benthic fluxes among the sampling sites and also significantly greater variability in flux among sites than within sites [P (perm) < 0.01, Table 3]. Pair-wise comparisons showed that benthic fluxes at SoGE differed significantly from fluxes measured at all the other sites [DDL, P (perm) = 0.0073; SI, P (perm) = 0.0002; SoGC, P (perm) = 0.0345]. Pair-wise comparisons also showed that benthic fluxes at DDL differed significantly from fluxes measured at SoGC [P (MC) = 0.0339] (Figure 2B). Moreover, the nMDS plot clearly showed greater similarity in benthic fluxes within than across sites (Figure 2B).
Environmental Variables Explaining Multivariate Benthic Flux Variation
The best model that emerged from our redundancy analysis between benthic fluxes and environmental variables explained 58.3% (R2 = 0.583, Adj. R2 = 0.444) of the total multivariate benthic flux variation and included five environmental variables (Appendix 4 in Supplementary Material). Chl a:phaeo ratio contributed most to the variation (18.8%), followed by prokaryotic abundance (14.5%), depth (8.8%), temperature (8.8%), and porosity (7.4%) (Appendix 4 in Supplementary Material).
The first and second axes of the redundancy model accounted for 27.3 and 14.6% of total flux variation respectively (Appendix 5 in Supplementary Material). The first axis mostly separated SoGE from SoGC, DDL, and SI fluxes (Figure 3A). Chl a:phaeo ratio, prokaryotic abundance and depth contributed primarily to the first axis and explained 46.9% of fitted flux variation (Figure 3A, Appendix 5 in Supplementary Material). In explaining 25.0% of the fitted variation in fluxes, the second axis mostly separated DDL from SoGE, SoGC, and SI fluxes (Figure 3A) and correlated most strongly with prokaryotic abundance, and to a lesser extend with the chl a:phaeo ratio and temperature (Figure 3A, Appendix 5 in Supplementary Material).
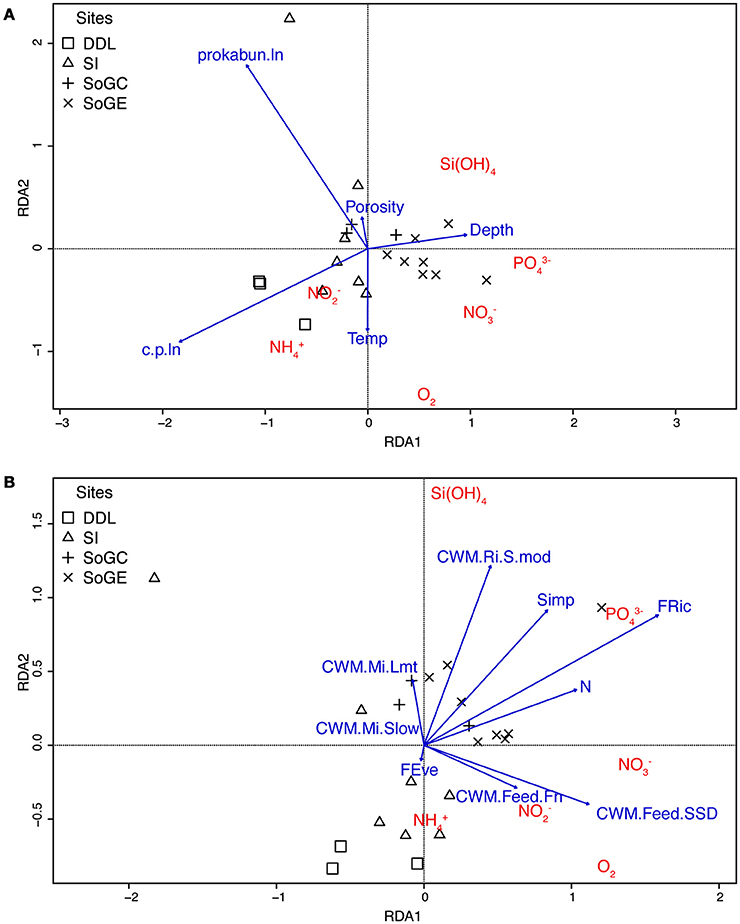
Figure 3. Plot of the redundancy analysis (RDA) models of (A) environmental variables, and (B) functional diversity indices best explaining variation in Salish Sea benthic fluxes measured in May/July 2011, and September 2013.
Functional Diversity Indices and Multivariate Benthic Flux Variation
The best model that emerged from our redundancy analysis between benthic fluxes and functional diversity indices explained 67.8% (R2 = 0.678, Adj. R2 = 0.414) of total multivariate benthic flux variation and included nine functional diversity indices (Appendix 6 in Supplementary Material). Functional richness (FRic) contributed most to the variation (19.7%), while the eight other functional diversity indices contributed to a lesser extent, with contributions ranging between 4.5 and 8.3% (Appendix 6 in Supplementary Material).
The first and second axes of the redundancy model accounted for 30.2 and 19.6% of total flux variation respectively (Appendix 7A in Supplementary Material). Again, the first axis mostly separated SoGE from SoGC, DDL, and SI fluxes (Figure 3B). Functional richness (FRic), community weighted means of sub-surface deposit feeders (CWM.Feed.SSD), abundance (N) and Simpson's diversity (Simp) contributed primarily to the first axis and explained 44.6% of the fitted flux variation (Figure 3B, Appendices 7A,B in Supplementary Material). As with the first axis, the second axis mostly separated SoGE and SoGC from DDL and SI fluxes (Figure 3B), explaining 28.9% of the fitted variation in fluxes and correlating most strongly with community weighted means of surficial modifiers (CWM.Ri.S.mod), Simpson's diversity (Simp) and functional richness (FRic) (Figure 3B, Appendices 7A,B in Supplementary Material).
Benthic Flux Variation Partitioning
Variation partitioning analysis of benthic fluxes between environmental variables and functional diversity indices identified by RDA indicated that environmental variables and functional diversity indices together explained 62.9% of benthic flux variation (R2 = 0.889, Adj. R2 = 0.629) (Figure 4, Appendix 8 in Supplementary Material). Environmental variables alone explained 21.4% of benthic flux variation, whereas functional diversity indices alone explained 18.5%; environmental variables and functional diversity indices shared 22.9% of the variation (Figure 4, Appendix 8 in Supplementary Material).
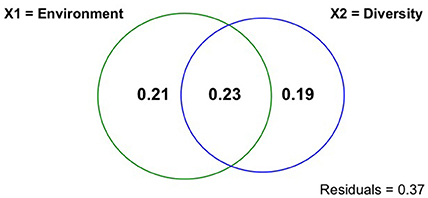
Figure 4. Venn diagram illustrating results of variation partitioning of benthic fluxes explained by environmental variables and functional diversity (FD) indices. X1 = environmental variables and X2 = functional diversity indices. Numbers correspond to variation explained by different fractions: environmental variable only = 0.21, FD indices only = 0.19, and intersection of environmental variables and functional diversity indices = 0.23.
Discussion
In this study we determined the environmental variables and functional diversity indices influencing benthic flux rates using redundancy analyses, and further evaluated their contributions using variation partitioning analysis. Our study is the first to use a powerful tool—variation partitioning analysis—to examine the contribution of environmental variables and functional diversity indices to multivariate flux rates and organic matter remineralization, in our case for soft sedimentary habitats. Our results show that environmental variables and functional diversity indices collectively explain the majority of the flux variation in our system and that they play a similar role in the control of flux rates. Furthermore, our results also indicate that environmental variables and functional diversity indices share a large proportion of the flux variation, which demonstrates the close links between the environment and resident species in delivery of key ecosystem functions.
Benthic Fluxes and Benthic Community Spatial Variation
Most studies have investigated the effects of abiotic and biotic factors influencing ecosystem processes and functions separately, but relatively few have attempted to separate the contribution of abiotic and biotic factors in the field (Hiddink et al., 2009; Queirós et al., 2011; Godbold, 2012; Braeckman et al., 2014; Strong et al., 2015). Our analyses demonstrate that despite significantly different macrofaunal communities at each of our sampling sites, differences in benthic fluxes were less consistent. On the one hand, SoGE fluxes differed significantly from the three other sites, and DDL fluxes also differed significantly from SoGC. On the other hand, SI fluxes were similar to those at DDL and SoGC. A previous study reported no consistent changes in ecosystem function with changes in functional diversity (Frid and Caswell, 2015) and we also found consistent differences in benthic communities at our study sites but not in benthic flux rates. Therefore, the specific attributes of our study system provide an opportunity to evaluate the contribution of environmental variables and functional diversity to benthic flux variation. Because communities consistently varied among all sites whereas functions did not, this might suggest that between-site differences in environmental variables and biodiversity have their own influence on ecosystem functions as reported by Strong et al. (2015).
Functional Diversity Effects on Multivariate Benthic Flux Variation
Based on the functional traits and modalities selected, functional richness (FRic), defined as “the amount of functional space filled by the community” (Villéger et al., 2008), influenced multivariate benthic flux variation more than any other functional diversity index, alone explaining 19.7% of the variation. This result indicates the primary importance of functional trait richness for benthic fluxes as suggested by Braeckman et al. (2014) for fine sandy sediments in the Southern North Sea. Our redundancy analysis indicated that, with the exception of ammonium, high fluxes of O2 and nutrients characterized sediment cores with the highest functional richness (FRic) (e.g., SoGE, Figure 3B). Similarly, we found positive relationships between functional richness and nutrient effluxes, especially phosphate and silicate, where efflux rates increased with increasing functional richness (Appendix 9 in Supplementary Material).
The larger influence of functional richness (FRic) on benthic flux variation than measures of species diversity (Simp, 5.0%) and abundance (N, 4.5%), suggests that a community composed of a few species in relatively low abundance could match or enhance benthic flux rates relative to another community comprised of more species in higher abundance if their functional trait diversities are similar (similar FRic). In our study, lower abundance (Mean ± SE = 28 ± 9, Appendix 10 in Supplementary Material) and Simpson's diversity (0.88 ± 0.3, Appendix 10 in Supplementary Material) at SI compared to DDL (N = 116 ± 44 and Simp = 0.93 ± 0.01, Appendix 10 in Supplementary Material) but similar functional richness (SI = 21.57 ± 25.50 and DDL = 19.13 ± 6.52, Appendix 10 in Supplementary Material) corresponded to similar benthic fluxes, as identified by our PERMANOVA. This result could have important implications for future studies and conservation efforts because it suggests greater importance of richness of functional traits (i.e., FRic) than species diversity (i.e., Simpson's diversity index) and species abundance in maintaining benthic ecosystem functioning (i.e., benthic fluxes). Similarly, a recent review of the biodiversity-ecosystem functioning (BEF) literature (Strong et al., 2015) also concluded that measures of functional diversity produced better BEF relationships compared to other measures of biodiversity such as species richness. Finally, a recent study using coastal marine benthic macrofaunal data from the Skagerrak-Baltic Sea region showed that although functional diversity usually decreases with decreasing taxonomic richness, in some cases functional diversity may remain high even at low taxonomic richness, suggesting that ecosystem processes and functions could potentially be maintained at lower taxonomic richness but similar functional diversity (Törnroos et al., 2015). This finding led them to suggest the primary importance of functional characteristics of species in maintaining ecosystem functions.
Our redundancy analysis indicated the importance of other functional diversity indices which demonstrate the important contribution of bioturbation and bio-irrigation of the sediment matrix to benthic flux variation. Functional diversity indices related to reworking of the sediment matrix (i.e., bioturbation), namely the community weighted means of taxa with limited (CWM.Mi.Lmt) and slow (CWM.Mi.Slow) movement through the sediment matrix, and of surficial modifiers (CWM.Ri.S.mod) explained 8.3%, 6.0% and 4.5% of benthic flux variation, respectively. Particle reworking and solute transport caused by infaunal movement through surface sediments are also known to increase microbial activities, organic matter degradation rates, and nutrient recycling (Aller et al., 2001). In their study, Lohrer et al. (2004) also showed a large positive effect of bioturbation activities of spatangoid urchins on benthic-pelagic fluxes. Moreover, the sediment resuspension created by groundfish activities (primarily the flatfish Lyopsetta exilis) plays a major role in ammonium, phosphate, and silica cycles in Saanich Inlet, with a lesser role for infauna (Yahel et al., 2008; Katz et al., 2009). In our study, many taxa contribute to bioturbation activities and increased benthic fluxes cannot be attributed to a single species. Nonetheless, a small subset of traits related to bioturbation activities clearly exhibited an important positive influence on benthic flux rates. In particular, these traits correspond to the taxa contributing the most to dissimilarity between SI and the other sites (SoGE, SoGC, and DDL) identified in our SIMPER analysis. For example, Cumella sp. (slow movement through the sediment and surficial modifier) played a particularly important role in differentiating SoGE, whereas L. gracilis (slow movement through the sediment and surficial modifier) differentiated SoGC, and P. lighti (limited movement through the sediment) differentiated DDL. Therefore, our results suggest that, among others, these taxa contributed not only to between-site differences in fauna but also to differences in benthic flux variation documented by our redundancy analysis. When combining the reworking and mobility categories, which both contribute to bioturbation, our redundancy analysis identified surface modifiers as our bioturbation trait least affecting benthic flux variation, a finding consistent with previous studies suggesting that surficial modifiers have the lowest impact on bioturbation among functional groups other than sediment stabilizers (Queirós et al., 2013). Interestingly, surficial modifiers comprised 69 of the 135 taxa (51.1%) identified in our study, with particularly high abundances at SoGE and SoGC (Appendix 10 in Supplementary Material) where we recorded the highest effluxes of phosphate and silicate. Our results suggest that despite their modest effect on bioturbation, the high abundances of surficial modifiers positively affected (though explaining only 4.5% of benthic flux variation) phosphate and silicate effluxes at SoGE and SoGC. Thus, weak bioturbators may contribute significantly when present in sufficiently high abundances, in this case at SoGE and SoGC where high functional richness in particular contributed to elevated effluxes of phosphate and silicate. More generally, Godbold and Solan (2009) showed that increased biodiversity (i.e., species richness) increased bioturbation activity (i.e., sediment mixing depth) in sediments near a fish farm in Scotland. Similarly, our results indicate that increased presence of bioturbating taxa led to increases in ecosystem processes measured (i.e., benthic flux rates; Figure 3B).
Functional diversity indices related to biological irrigation of sediment (i.e., bio-irrigation), namely the community weighted mean of funnel feeders (CWM.Feed.Fn) and sub-surface deposit feeders (CWM.Feed.SSD) explained 8.1 and 6.0% of benthic flux variation, respectively. Many taxa were classified as sub-surface deposit feeders, including Cumella sp. and L. gracilis identified as particularly important in differentiating SoGE and SoGC, respectively. However, the presence of key taxa and their specific functions apparently disproportionately (relatively to their abundance) impacted flux rates (Appendices 6,7 in Supplementary Material). For instance, funnel feeders, a sub-group of deposit feeding animals that feed on surficial sediments but from below the sediment surface (Jumars et al., 2015), comprised only six polychaete taxa spanning two families (Maldanidae: Maldanidae spp., Maldane sp., Maldane sarsi, Praxillella sp., and Praxillella gracilis, and Pectinariidae: Pectinaria californiensis) represented by a total of only 12 specimens in our sediment cores. Yet, these taxa occurred mainly at SoGE (Appendix 10 in Supplementary Material), where we recorded particularly large silicate and phosphate releases, as well as nitrite intakes. Tube-building maldanids (i.e., Praxillella sp.) in particular can rapidly subduct freshly deposited organic matter that becomes available for deep-dwelling microbes and other infauna, and consequently enhance organic matter remineralization (Levin et al., 1997). Maldanids were therefore proposed as geochemical keystone species because of their feeding (Levin et al., 1997) and irrigation (Waldbusser et al., 2004) activities. The analysis of the three-dimensional organization of M. sarsi tubes also revealed increased concentrations of Fe, Mn, organic carbon, and bacteria, potentially resulting from tube irrigation, mucous secretion, and feeding activities (Dufour et al., 2008). Our results and previous studies point to the primary importance of functional traits related to bio-irrigation of sediments for the biogeochemical cycling of nutrients in sedimentary habitats. Moreover, these results point to the disproportionate importance (relative to their abundance) of some taxa and associated traits in sustaining ecosystem functions. Although our results show that overall, functional diversity influences benthic flux variation most strongly, they also suggest a strong identity effect, where a small number of taxa (i.e., six funnel feeder taxa) substantially impact ecosystem functions (Loreau et al., 2001; Strong et al., 2015).
Mobile bioturbators are known to increase bio-irrigation (Woodin et al., 2010) and organisms sediment reworking activities can also affect fluid transport within the sediment (Meysman et al., 2006). In our study, some taxa clearly influenced both bioturbation (i.e., particle mixing) and bio-irrigation (i.e., fluid exchange). For example, Cumella sp. and L. gracilis, which differentiated SoGE and SoGC, respectively, are sub-surface deposit feeders (a bio-irrigation trait) that move slowly through the sediment and act as surficial modifiers (bioturbation traits). This result suggests that some traits related to bioturbation and bio-irrigation can covary (e.g., increases in L. gracilis mean an increase in the following traits: sub-surface deposit feeders, slow movement through the sediment, and surficial modifiers). Taken together, biological sediment reworking and irrigation activities explained 32.9% of variation in benthic flux. These results mirror previous studies indicating that biological mixing of sediments and solute transport during feeding and irrigation stimulates microbial activity and organic matter remineralization (Aller and Aller, 1998; Aller, 2014).
Environmental Variables Explaining Multivariate Benthic Flux Variation
Of the environmental variables we examined, the chl a:phaeo ratio most strongly influenced benthic fluxes (i.e., 18.8%); this ratio reflects organic matter quality on a short time scale of days to weeks (Veuger and van Oevelen, 2011; Le Guitton et al., 2015). Prokaryotic cell abundance was the second most important environmental variable, explaining 14.5% of the variation in flux. Redundancy analysis indicated that high fluxes of ammonium (e.g., DDL) and nitrite (e.g., SI) characterized sites with the highest chl a:phaeo ratio and abundance of prokaryotic cells (Figure 3A). Similarly, most environmental variables explaining benthic flux variation identified in our study (i.e., chl a:phaeo ratio, temperature, and porosity) were previously reported as strong predictors for this region (see Belley et al., 2016). However, the redundancy analysis performed in our study identified prokaryotic abundance as an important variable explaining benthic flux variation not identified in our previous study (Belley et al., 2016). The fact that prokaryotic abundance remained an important variable explaining single flux variation (from multiple linear regression results) for all but silicate fluxes over a broad geographic area in our previous study (Belley et al., 2016) (i.e., Salish Sea but also sites in the open waters of the Northeast Pacific) can explain this discrepancy. We included water depth in our analysis because it often correlates well with other environmental variable known to influence benthic flux rates, such as organic flux to the seafloor (Jahnke, 1990; Berelson et al., 1996) and temperature (Hargrave, 1969; Cowan et al., 1996; Alonso-Pérez and Castro, 2014). Our model specifically accounted for differences in water depth, which explained 8.8% of the variation in benthic flux, and high fluxes of O2, nitrate, phosphate, and silicate (e.g., SoGE) generally characterized deeper sites. Overall, our results align with previous studies that reported increased fluxes of seafloor oxygen and nutrients with increased flux of fresh organic matter following phytoplankton blooms (Whitledge et al., 1986) and increased microbial abundance (Pfannkuche, 1993; Gooday, 2002).
Benthic Flux Variation Partitioning
Environmental variables and functional diversity indices collectively explained 62.9% of variation in benthic flux at our study sites. Environmental variables alone explained 21.4% of this variation, functional diversity indices alone explained 18.5%, whereas the two variable groups shared 22.9% of the variance. These results indicate that the abiotic and biotic variables measured in our study explained the majority of the variation in benthic flux, and that environment and macrofaunal functional diversity weigh almost equally in contribution. The meta-analysis conducted by Godbold (2012), who reported positive and similar abiotic and biotic (i.e., species identity and species richness) effects on ecosystem functions measured in the majority of experiments included in their analysis, support our results. However, the many field experiments that manipulated diversity and used species that comprised only a fraction of the natural community usually report higher influence of environmental variables on ecosystem functions. Godbold and Solan (2009) and Duffy (2009) propose that including a low number of species in manipulative experiments reduces the observed effect of biodiversity on ecosystem functions relative to environmental variables. Based on our results, and those from the few similar observational studies, we also advocate the use of natural communities in future studies to fully appreciate the full effect of biodiversity on ecosystem functioning. Although we acknowledge that correlative and regression analysis do not fully demonstrate causality, which requires manipulative experiments, we believe that mensurative data such as those we present here should inform manipulative experiments (which bring other limitations), in order to focus promising experimental directions.
We also measured many abiotic variables at the scale of sites (<25 m) and biotic variables at the scale of cores (10 s of cm). Although this approach may have underestimed small-scale (within site) variation in environmental variables relative to biotic variables, we argue that most abiotic variables measured in our study vary little in magnitude in any consistent way, noting the homogenizing effect of tidal exchange over the small (25 × 25 m) areas sampled within our study sites (e.g., bottom water temperature, dissolved O2 concentration, bottom depth). Admittedly, our RDA models identified five environmental and nine diversity variables best explaining benthic flux variation, and the inclusion of a larger number of diversity variables arguably may have increased the contribution of diversity variables to explaining the response variation. However, the stepwise selection of explanatory variables in both RDAs was based on their significance in explaining benthic flux variation. Therefore, both RDA models retained only significant predictors of benthic flux variation. Moreover, because we measured the environmental and diversity variables recognized by a wide range of studies as those most important for benthic fluxes, we believe our results provide an accurate estimate of the contributions of environmental and diversity variables to benthic flux variation at our study sites. Finally, examination of the stepwise analysis indicates that the top five diversity variables alone explained 44.2% of the variation in fluxes (based on R2 values from RDA #1), which is roughly comparable to the 58.3% explained by the environmental variables (based on R2 values from RDA #2).
The large proportion (22.9%) of the explained variation shared between environmental variables and functional diversity indices demonstrates the close interactions between resident species and their environment. Environmental variables greatly impact benthic community composition, however, the community also plays an important role in controlling ecosystem functioning. For example, the rate of particulate organic matter export to the seafloor strongly impacts benthic community composition (Wei et al., 2010). Decreases in dissolved oxygen concentration can also modify benthic community composition and can lead to lower sediment bioturbation and bio-irrigation rates (Levin et al., 2009; Belley et al., 2010; Rabalais et al., 2010) which, in turn, can decrease benthic flux rates (Aller, 1982; Link et al., 2013b; Aller, 2014). With increasing anthropogenic pressures on marine ecosystems (Halpern et al., 2008, 2015) and resulting decreases in marine biodiversity (Worm et al., 2006), the important proportion of the explained variation shared between the environment and the species inhabiting the sediments point to the need to limit anthropogenic impacts that might change the marine environment and potentially lead to loss of biodiversity and associated ecosystem functions in marine sedimentary habitats.
Effect of Other Variables on Benthic Flux Variation
The biological and environmental variables we measured could not explain approximately 37.1% of the benthic flux variation. Therefore, other factors not measured in our study presumably contribute to variability in benthic fluxes. Scavenging by trace metals (e.g., iron, manganese) (de la Rocha, 2003), and concentration gradients and molecular diffusion in sediment porewater and overlying water result in spatial and temporal variation in oxygen and nutrient fluxes at the seafloor (Schulz, 2000). For example, the dissolved oxygen contained in the bottom water penetrates the sediment following a diffusion gradient (i.e., from higher to lower concentration). Microorganisms such as bacteria and archaea (i.e., prokaryotes) utilize oxygen and other electron acceptors to degrade organic material within the sediment and therefore affect local concentrations. These changes in local concentrations generate nutrient fluxes directed either toward the water column or deeper in the sediment, depending on the specific chemical compound (Jorgensen, 2006). Moreover, macrobenthic organisms influence the distribution of chemical compounds and reaction rates by: (1) moving particles during feeding, burrowing, and tube construction, (2) disrupting the otherwise vertically stratified distribution of biogeochemical compounds during burrow and fecal pellet formation, (3) introducing new reactive organic substances during mucus secretion, (4) influencing bacterial communities that mediate chemical reactions during feeding and sediment mechanical disturbance, and (5) altering sediment during gut passage (Aller, 1982). Through their activities, macrofauna control pore water solute concentration profiles (Aller, 1982), increasing benthic fluxes by a factor of 3–4 in continental shelf sediments (Archer and Devol, 1992; Devol and Christensen, 1993). Although we measured bottom water dissolved oxygen concentrations at our study sites (environmental variable not retained by our RDA analysis), we did not measure in situ bottom water nutrient concentrations. Hence, differences in bottom water nutrient concentrations across our study sites could have influenced benthic flux rates and contributed to the 37.1% unexplained benthic flux variation at our study sites. However, we believe such a scenario unlikely for explaining within-site benthic flux variation because tidal currents would tend to minimize horizontal variation in bottom water properties within the 25 × 25 m sites that we sampled.
Summary
Our study indicates that environmental variables and functional diversity indices we measured explain the majority of flux variation in our Salish Sea sedimentary sites. Lability of organic matter, microbial abundance, benthic macrofaunal functional richness, and indices related to bioturbation and bio-irrigation were the most important variables in explaining benthic flux variation and organic matter remineralization at our seafloor study sites. Moreover, our results suggest that functional richness better predicts benthic flux rates than species diversity and abundance. We also identified funnel feeding as a key function provided by activities of a small number of species and individuals of maldanids and pectinariids polychaetes, which can affect benthic flux rates disproportionately relative to their abundance. Our results indicate that biodiversity and environment play a similar role in the control of organic matter remineralization. However, larger flux rates were recorded at sites with higher functional richness (e.g., SoGE) and funnel feeders, suggesting greater efficiency in organic matter processing with higher biodiversity. Given the increasing negative anthropogenic impacts on natural ecosystems and corresponding changes in biodiversity, our results point to the need to maintain functional richness in order to maintain ecosystem functioning. Results of this and other studies could help to predict the impact of non-random species loss associated with environmental changes (e.g., decrease of dissolved oxygen concentrations) on ecosystem functions such as nutrient flux rates and organic matter remineralization.
Author Contributions
Conceived and designed the experiments: RB, PS. Performed the experiments: RB. Analyzed the data: RB, PS. Contributed reagents/materials/analysis tools: PS. Wrote the paper: RB, PS.
Funding
Funding was provided by Natural Sciences and Engineering Research Council of Canada Discovery Grants (PS, grant #200372), the Natural Sciences and Engineering Research Council of Canada Canadian Healthy Oceans Network (PS, grant #206236), a Natural Sciences and Engineering Research Council of Canada Postgraduate Scholarships-Doctoral (RB), and the School of Graduate Studies at Memorial University (RB). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript. The Schmidt Ocean Institute (http://www.schmidtocean.org/) provided support (e.g., Research Vessel, technicians) for data collection.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors thank the CSSF ROPOS team as well as the officers and crews of the CCGS John P. Tully, R/V Thomas G. Thompson and R/V Falkor for their assistance with sample collection. Ocean Networks Canada kindly provided the opportunity to sample. We offer special thanks to C. Anstey and the Department of Fisheries and Oceans Canada who kindly provided nutrient analyses. We also thank Dr. S.K. Juniper and Dr. P. Archambault for their constructive thoughts on this study, Dr. V. Tunnicliffe, Dr. M. Matabos, Dr. R. Dewey, Dr. L. Pautet and J. Rose for help with sample collection, Dr. R. Stanley for advice on data analysis and help on figure production, Dr. C-L. Wei for advice on data analysis, Drs. H. Link and A. Piot for their constructive discussion on benthic fluxes, Dr. R Danovaro and E. Rastelli for help on prokaryotic cell counts, and Dr. R. Rivkin, Dr. S. Dufour, and L. Stolze for help with determination of sediment characteristics. We acknowledge the reviewers for their constructive comments which improved this manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmars.2016.00242/full#supplementary-material
References
Aller, R. C. (1982). “The effects of macrobenthos on chemical properties of marine sediment and overlying water,” in Animal-Sediment Relations: the Biogenic Alteration of Sediments, eds P. L. McCall and M. J. S. Tevesz (New York, NY: Plenum Press), 53–102.
Aller, R. C. (2014). “Sedimentary diagenesis, depositional environments, and benthic fluxes,” in Treatise on Geochemistry, 2nd Edn, eds H. D. Holland and K. K. Turekian. (Oxford: Elsevier), 293–334.
Aller, R. C., and Aller, J. Y. (1998). The effect of biogenic irrigation intensity and solute exchange on diagenetic reaction rates in marine sediments. J. Mar. Res. 56, 905–936. doi: 10.1357/002224098321667413
Aller, R. C., Aller, J. Y., and Kemp, P. F. (2001). “Effects of particle and solute transport on rates and extent of remineralization in bioturbated sediments,” in Organism-Sediment Interactions, eds J. Y. Aller, S. A. Woodin, and R. C. Aller. (Columbia: University of South Carolina Press), 315–333.
Alonso-Pérez, F., and Castro, C. G. (2014). Benthic oxygen and nutrient fluxes in a coastal upwelling system (Ria de Vigo, NW Iberian Peninsula): seasonal trends and regulating factors. Mar. Ecol. Prog. Ser. 511, 17–32. doi: 10.3354/meps10915
Anderson, M., Gorley, R., and Clarke, K. (2008). PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods. Plymouth, UK: PRIMER-E.
Anderson, M. J. (2001). A new method for non-parametric multivariate analysis of variance. Aust. Ecol. 26, 32–46. doi: 10.1111/j.1442-9993.2001.01070.pp.x
Archer, D., and Devol, A. (1992). Benthic oxygen fluxes on the Washington shelf and slope: a comparison of in situ microelectrode and chamber flux measurements. Limnol. Oceanogr. 37, 614–629. doi: 10.4319/lo.1992.37.3.0614
Avnimelech, Y., Ritvo, G., Meijer, L. E., and Kochba, M. (2001). Water content, organic carbon and dry bulk density in flooded sediments. Aquaculture Eng. 25, 25–33. doi: 10.1016/S0144-8609(01)00068-1
Belley, R., Archambault, P., Sundby, B., Gilbert, F., and Gagnon, J.-M. (2010). Effects of hypoxia on benthic macrofauna and bioturbation in the Estuary and Gulf of St. Lawrence Canada. Cont. Shelf Res. 30, 1302–1313. doi: 10.1016/j.csr.2010.04.010
Belley, R., Snelgrove, P. V. R., Archambault, P., and Juniper, S. K. (2016). Environmental drivers of benthic flux variation and ecosystem functioning in Salish Sea and Northeast Pacific sediments. PLoS ONE 11:e0151110. doi: 10.1371/journal.pone.0151110
Berelson, W. M., McManus, J., Coale, K. H., Johnson, K. S., Kilgore, T., Burdige, D., et al. (1996). Biogenic matter diagenesis on the sea floor: a comparison between two continental margin transects. J. Mar. Res. 54, 731–762. doi: 10.1357/0022240963213673
Botta-Dukat, Z. (2005). Rao's quadratic entropy as a measure of functional diversity based on multiple traits. J. Veg Sci. 16, 533–540. doi: 10.1111/j.1654-1103.2005.tb02393.x
Braeckman, U., Foshtomi, M. Y., Van Gansbeke, D., Meysman, F., Soetaert, K., Vincx, M., et al. (2014). Variable importance of macrofaunal functional biodiversity for biogeochemical cycling in temperate coastal sediments. Ecosystems 17, 720–737. doi: 10.1007/s10021-014-9755-7
Burd, B. J., Macdonald, R. W., Johannessen, S. C., and van Roodselaar, A. (2008). Responses of subtidal benthos of the Strait of Georgia, British Columbia, Canada to ambient sediment conditions and natural and anthropogenic depositions. Mar. Environ. Res. 66, S62–S79. doi: 10.1016/j.marenvres.2008.08.009
Cardinale, B. J., Duffy, J. E., Gonzalez, A., Hooper, D. U., Perrings, C., Venail, P., et al. (2012). Biodiversity loss and its impact on humanity. Nature 486, 59–67. doi: 10.1038/nature11148
Cardinale, B. J., Srivastava, D. S., Emmett Duffy, J., Wright, J. P., Downing, A. L., Sankaran, M., et al. (2006). Effects of biodiversity on the functioning of trophic groups and ecosystems. Nature 443, 989–992. doi: 10.1038/nature05202
Chu, J. W. F., and Tunnicliffe, V. (2015). Oxygen limitations on marine animal distributions and the collapse of epibenthic community structure during shoaling hypoxia. Glob. Change Biol. 21, 2989–3004. doi: 10.1111/gcb.12898
Clarke, K. R. (1993). Nonparametric multivariate analyses of changes in community structure. Aust. J. Ecol. 18, 117–143. doi: 10.1111/j.1442-9993.1993.tb00438.x
Clarke, K. R., and Warwick, R. M. (1994). Similarity-based testing for community pattern: the two-way layout with no replication. Mar. Biol. 118, 167–176. doi: 10.1007/BF00699231
Cowan, J. L. W., Pennock, J. R., and Boynton, W. R. (1996). Seasonal and interannual patterns of sediment-water nutrient and oxygen fluxes in Mobile Bay, Alabama (USA): regulating factors and ecological significance. Mar. Ecol. Prog. Ser. 141, 229–245. doi: 10.3354/meps141229
Danovaro, R. (2010). Methods for the Study of Deep-Sea Sediments, their Functioning and Biodiversity. Boca Raton, FL: CRC Press.
Danovaro, R., Gambi, C., Dell'Anno, A., Corinaidesi, C., Fraschetti, S., Vanreusel, A., et al. (2008). Exponential decline of deep-sea ecosystem functioning linked to benthic biodiversity loss. Curr. Biol. 18, 1–8. doi: 10.1016/j.cub.2007.11.056
de la Rocha, C. L. (2003). “The biological pump,” in The Oceans and Marine Giochemistry, eds H. D. Holland and K. K. Tukekian (Oxford: Elsevier-Pergamon), 83–111.
Devol, A. H., and Christensen, J. P. (1993). Benthic fluxes and nitrogen cycling in sediments of the continental margin of the eastern North Pacific. J. Mar. Res. 51, 345–372. doi: 10.1357/0022240933223765
Duffy, J. E. (2009). Why biodiversity is important to the functioning of real-world ecosystems. Front. Ecol. Environ. 7:70195. doi: 10.1890/070195
Dufour, S. C., White, C., Desrosiers, G., and Juniper, S. K. (2008). Structure and composition of the consolidated mud tube of Maldane sarsi (Polychaeta: Maldanidae). Estuar Coast Shelf Sci. 78, 360–368. doi: 10.1016/j.ecss.2007.12.013
Frid, C. L. J., and Caswell, B. A. (2015). Is long-term ecological functioning stable: the case of the marine benthos? J. Sea Res. 98, 15–23. doi: 10.1016/j.seares.2014.08.003
Giller, P. S., Hillebrand, H., Berninger, U. G., Gessner, M. O., Hawkins, S., Inchausti, P., et al. (2004). Biodiversity effects on ecosystem functioning: emerging issues and their experimental test in aquatic environments. Oikos 104, 423–436. doi: 10.1111/j.0030-1299.2004.13253.x
Godbold, J. A. (2012). “Effects of biodiversity-environment conditions on the interpretation of biodiversity-function relations,” in Marine Biodiversity and Ecosystem Functioning: Frameworks, Methodologies, and Integration, 1st edn, eds M. Solan, R. J. Aspden, and D. M. Paterson. (Oxford: Oxford University Press), 114.
Godbold, J. A., and Solan, M. (2009). Relative importance of biodiversity and the abiotic environment in mediating an ecosystem process. Mar. Ecol. Prog. Ser. 396, 273–282. doi: 10.3354/meps08401
Gooday, A. J. (2002). Biological responses to seasonally varying fluxes of organic matter to the ocean floor: a review. J. Oceanogr. 58, 305–332. doi: 10.1023/A:1015865826379
Hall, P. O. J., Anderson, L. G., van der Loeff, M. M. R., Sundby, B., and Westerlund, S. F. G. (1989). Oxygen uptake kinetics in the benthic boundary layer. Limnol. Oceanogr. 34, 734–746. doi: 10.4319/lo.1989.34.4.0734
Halpern, B. S., Frazier, M., Potapenko, J., Casey, K. S., Koenig, K., Longo, C., et al. (2015). Spatial and temporal changes in cumulative human impacts on the world's ocean. Nat. Commun. 6:7615. doi: 10.1038/ncomms8615
Halpern, B. S., Walbridge, S., Selkoe, K. A., Kappel, C. V., Micheli, F., D'Agrosa, C., et al. (2008). A global map of human impact on marine ecosystems. Science 319, 948–952. doi: 10.1126/science.1149345
Hargrave, B. T. (1969). Similarity of oxygen uptake by benthic communities. Limnol. Oceanogr. 14, 801–805. doi: 10.4319/lo.1969.14.5.0801
Hiddink, J. G., Davies, T. W., Perkins, M., Machairopoulou, M., and Neill, S. P. (2009). Context dependency of relationships between biodiversity and ecosystem functioning is different for multiple ecosystem functions. Oikos 118, 1892–1900. doi: 10.1111/j.1600-0706.2009.17556.x
Hooper, D. U., Chapin, F. S., Ewel, J. J., Hector, A., Inchausti, P., Lavorel, S., et al. (2005). Effects of biodiversity on ecosystem functioning: a consensus of current knowledge. Ecol. Monogr. 75, 3–35. doi: 10.1890/04-0922
Jahnke, R. A. (1990). Early diagenesis and recycling of biogenic debris at the seafloor, Santa Monica Basin, California. J. Mar. Res. 48, 413–436. doi: 10.1357/002224090784988773
Jahnke, R. A. (1996). The global ocean flux of particulate organic carbon: areal distribution and magnitude. Glob. Biogeochem. Cycles. 10, 71–88. doi: 10.1029/95GB03525
Johannessen, S. C., Masson, D., and Macdonald, R. W. (2014). Oxygen in the deep Strait of Georgia, 1951–2009: the roles of mixing, deep-water renewal, and remineralization of organic carbon. Limnol. Oceanogr. 59, 211–222. doi: 10.4319/lo.2014.59.1.0211
Johannessen, S. C., O'Brien, M. C., Denman, K. L., and Macdonald, R. W. (2005). Seasonal and spatial variations in the source and transport of sinking particles in the Strait of Georgia, British Columbia, Canada. Mar. Geol. 216, 59–77. doi: 10.1016/j.margeo.2005.01.004
Jorgensen, B. B. (2006). “Bacteria and marine biogeochemistry,” in Marine Geochemistry, eds H. D. Schulz and M. Zabel. 2nd edn (Berlin: Springer), 169–206.
Jumars, P. A., Dorgan, K. M., and Lindsay, S. M. (2015). Diet of worms emended: an update of polychaete feeding guilds. Annu. Rev. Mar. Sci. 7, 497–520. doi: 10.1146/annurev-marine-010814-020007
Katz, T., Yahel, G., Yahel, R., Tunnicliffe, V., Herut, B., Snelgrove, P., et al. (2009). Groundfish overfishing, diatom decline, and the marine silica cycle: lessons from Saanich Inlet, Canada, and the Baltic Sea cod crash. Glob. Biogeochem. Cycol 23, GB4032. doi: 10.1029/2008GB003416
Kerouel, R., and Aminot, A. (1997). Fluorometric determination of ammonia in sea and estuarine waters by direct segmented flow analysis. Mar. Chem. 57, 265–275. doi: 10.1016/S0304-4203(97)00040-6
Laliberté, E., and Legendre, P. (2010). A distance-based framework for measuring functional diversity from multiple traits. Ecology 91, 299–305. doi: 10.1890/08-2244.1
Lavorel, S., Grigulis, K., McIntyre, S., Williams, N. S. G., Garden, D., Dorrough, J., et al. (2008). Assessing functional diversity in the field – methodology matters! Funct. Ecol. 22, 134–147. doi: 10.1111/j.1365-2435.2007.01339.x
Le Guitton, M., Soetaert, K., Damsté, J. S. S., and Middelburg, J. J. (2015). Biogeochemical consequences of vertical and lateral transport of particulate organic matter in the southern North Sea: a multiproxy approach. Estuar Coast Shelf Sci. 165, 117–127. doi: 10.1016/j.ecss.2015.09.010
Levin, L. A., Ekau, W., Gooday, A. J., Jorissen, F., Middelburg, J. J., Naqvi, W., et al. (2009). Effects of natural and human-induced hypoxia on coastal benthos. Biogeosciences 6, 2063–2098. doi: 10.5194/bg-6-2063-2009
Levin, L., Blair, N., DeMaster, D., Plaia, G., Fornes, W., Martin, C., et al. (1997). Rapid subduction of organic matter by maldanid polychaetes on the North Carolina slope. J. Mar. Res. 55, 595–611. doi: 10.1357/0022240973224337
Link, H., Chaillou, G., Forest, A., Piepenburg, D., and Archambault, P. (2013a). Multivariate benthic ecosystem functioning in the Arctic – benthic fluxes explained by environmental parameters in the southeastern Beaufort Sea. Biogeosciences 10, 5911–5929. doi: 10.5194/bg-10-5911-2013
Link, H., Piepenburg, D., and Archambault, P. (2013b). Are hotspots always hotspots? The relationship between diversity, resource and ecosystem functions in the Arctic. PLoS ONE 8:e74077. doi: 10.1371/journal.pone.0074077
Lohrer, A. M., Thrush, S. F., and Gibbs, M. M. (2004). Bioturbators enhance ecosystem function through complex biogeochemical interactions. Nature 431, 1092–1095. doi: 10.1038/nature03042
Loreau, M. (2010). Linking biodiversity and ecosystems: towards a unifying ecological theory. Philos Trans. R. Soc B 365, 49–60. doi: 10.1098/rstb.2009.0155
Loreau, M., Naeem, S., and Inchausti, P. (2002). Biodiversity and Ecosystem Functioning: Synthesis and Perspectives. Toronto, ON: Oxford University Press.
Loreau, M., Naeem, S., Inchausti, P., Bengtsson, J., Grime, J. P., Hector, A., et al. (2001). Biodiversity and ecosystem functioning: current knowledge and future challenges. Science 294, 804–808. doi: 10.1126/science.1064088
Macdonald, T. A., Burd, B. J., Macdonald, V. I., and van Roodselaar, A. (2010). Taxonomic and Feeding Guild Classification for the Marine Benthic Macroinvertebrates of the Strait of Georgia, British Columbia. Canadian technical report of fisheries and aquatic sciences
Macdonald, T. A., Burd, B. J., and van Roodselaar, A. (2012). Size structure of marine soft-bottom macrobenthic communities across natural habitat gradients: implications for productivity and ecosystem function. PLoS ONE 7:e40071. doi: 10.1371/journal.pone.0040071
MarLIN. (2006). BIOTIC - Biological Traits Information Catalogue [Online]. Plymouth: Marine Biological Association of the United Kingdom. Available online: http://www.marlin.ac.uk/biotic (Accessed 2015-12-02)
Masson, D., and Cummins, P. F. (2007). Temperature trends and interannual variability in the Strait of Georgia, British Columbia. Cont. Shelf Res. 27, 634–649. doi: 10.1016/j.csr.2006.10.009
Matabos, M., Tunnicliffe, V., Juniper, S. K., and Dean, C. (2012). A year in hypoxia: epibenthic community responses to severe oxygen deficit at a subsea observatory in a coastal inlet. PLoS ONE 7:e45626. doi: 10.1371/journal.pone.0045626
McArdle, B. H., and Anderson, M. J. (2001). Fitting multivariate models to community data: a comment on distance-based redundancy analysis. Ecology 82, 290–297. doi: 10.1890/0012-9658(2001)082[0290:FMMTCD]2.0.CO;2
Meysman, F. J. R., Middelburg, J. J., and Heip, C. H. R. (2006). Bioturbation: a fresh look at Darwin's last idea. Trends Ecol. Evol. 21, 688–695. doi: 10.1016/j.tree.2006.08.002
Morata, N., Poulin, M., and Renaud, P. E. (2011). A multiple biomarker approach to tracking the fate of an ice algal bloom to the sea floor. Polar Biol. 34, 101–112. doi: 10.1007/s00300-010-0863-3
Naeem, S., Duffy, J. E., and Zavaleta, E. (2012). The functions of biological diversity in an age of extinction. Science 336, 1401–1406. doi: 10.1126/science.1215855
Nunnally, C. C., Rowe, G. T., Thornton, D. C. O., and Quigg, A. (2013). Sedimentary oxygen consumption and nutrient regeneration in the northern Gulf of Mexico hypoxic zone. J. Coast. Res. 63, 84–96. doi: 10.2112/SI63-008.1
Oksanen, J., Blanchet, F. G., Kindt, R., Legendre, P., Minchin, P. R., O'Hara, R. B., et al. (2013). “Vegan: Community Ecology Package”. R package version 2.0-10 ed.).
Pfannkuche, O. (1993). Benthic response to the sedimentation of particulate organic matter at the BIOTRANS station, 47°N, 20°W. Deep Sea Res. II 40, 135–149. doi: 10.1016/0967-0645(93)90010-K
Queirós, A. M., Birchenough, S. N. R., Bremner, J., Godbold, J. A., Parker, R. E., Romero-Ramirez, A., et al. (2013). A bioturbation classification of European marine infaunal invertebrates. Ecol. Evol. 3, 3958–3985. doi: 10.1002/ece3.769
Queirós, A. M., Hiddink, J. G., Johnson, G., Cabral, H. N., and Kaiser, M. J. (2011). Context dependence of marine ecosystem engineer invasion impacts on benthic ecosystem functioning. Biol. Invasions 13, 1059–1075. doi: 10.1007/s10530-011-9948-3
R Core Team (2016). “R: A language and Environment for Statistical Computing”. 3.2.4 edn. Vienna: R Foundation for Statistical Computing.
Rabalais, N. N., Díaz, R. J., Levin, L. A., Turner, R. E., Gilbert, D., and Zhang, J. (2010). Dynamics and distribution of natural and human-caused hypoxia. Biogeosciences 7, 585–619. doi: 10.5194/bg-7-585-2010
Raffaelli, D., Emmerson, M., Solan, M., Biles, C., and Paterson, D. (2003). Biodiversity and ecosystem processes in shallow coastal waters: an experimental approach. J. Sea Res. 49, 133–141. doi: 10.1016/S1385-1101(02)00200-9
Riaux-Gobin, C., and Klein, B. (1993). “Microphytobenthic biomass measurement using HPLC and conventional pigment analysis,” in Handbook of Methods in Aquatic Microbial Ecology, eds P. Kemp, B. Sherr, E. Sherr, and J. Cole (Boca Raton, FL: Lewis Publishers), 369–376.
Schulz, H. D. (2000). “Quantification of early diagenesis: dissolved constituents in marine pore water,” in Marine Geochemistry, eds H. D. Schulz and M. Zabel (New York, NY: Springer-Verlag Heidelberg), 87–128.
Snelgrove, P., Blackburn, T. H., Hutchings, P. A., Alongi, D. M., Grassle, J. F., Hummel, H., et al. (1997). The importance of marine sediment biodiversity in ecosystem processes. Ambio 26, 578–583.
Snelgrove, P. V. R., Thrush, S. F., Wall, D. H., and Norkko, A. (2014). Real world biodiversity-ecosystem functioning: a seafloor perspective. Trends Ecol. Evol. 29, 398–405. doi: 10.1016/j.tree.2014.05.002
Solan, M., Cardinale, B. J., Downing, A. L., Engelhardt, K. A. M., Ruesink, J. L., and Srivastava, D. S. (2004). Extinction and ecosystem function in the marine benthos. Science 306, 1177–1180. doi: 10.1126/science.1103960
Strong, J. A., Andonegi, E., Bizsel, K. C., Danovaro, R., Elliott, M., Franco, A., et al. (2015). Marine biodiversity and ecosystem function relationships: the potential for practical monitoring applications. Estuar Coast Shelf Sci. 161, 46–64. doi: 10.1016/j.ecss.2015.04.008
Suykens, K., Schmidt, S., Delille, B., Harlay, J., Chou, L., De Bodt, C., et al. (2011). Benthic remineralization in the northwest European continental margin (northern Bay of Biscay). Cont. Shelf Res. 31, 644–658. doi: 10.1016/j.csr.2010.12.017
Technicon Industrial Systems (1973). Orthophosphate in Water and Seawater. Autoanalyzer II Industrial Method No. 155-71 W. Tarrytown, NY: Technicon Industrial Systems.
Technicon Industrial Systems (1977). Silicates in Water and Seawater. Autoanalyzer II Industrial Method No. 186-72 W. Tarrytown, NY: Technicon Industrial Systems.
Technicon Industrial Systems (1979). Nitrate and Nitrite in Water and Seawater. Autoanalyzer II Industrial Method No. 158-71 W. Tarrytown, NY: Technicon Industrial Systems.
Terlizzi, A., Benedetti-Cecchi, L., Bevilacqua, S., Fraschetti, S., Guidetti, P., and Anderson, M. J. (2005). Multivariate and univariate asymmetrical analyses in environmental impact assessment: a case study of Mediterranean subtidal sessile assemblages. Mar. Ecol. Prog. Ser. 289, 27–42. doi: 10.3354/meps289027
Thibodeau, B., Lehmann, M. F., Kowarzyk, J., Mucci, A., Gelinas, Y., Gilbert, D., et al. (2010). Benthic nutrient fluxes along the Laurentian Channel: impacts on the N budget of the St. Lawrence marine system. Estuar. Coast. Shelf Sci. 90, 195–205. doi: 10.1016/j.ecss.2010.08.015
Tilman, D. (2001). “Functional diversity,” in Encyclopedia of Biodiversity, ed S. A. Levin (San Diego, CA: Academic Press), 109–120.
Törnroos, A., Bonsdorff, E., Bremner, J., Blomqvist, M., Josefson, A. B., Garcia, C., et al. (2015). Marine benthic ecological functioning over decreasing taxonomic richness. J. Sea Res. 98, 49–56. doi: 10.1016/j.seares.2014.04.010
Tunnicliffe, V. (1981). High species diversity and abundance of the epibenthic community in an oxygen-deficient basin. Nature 294, 354–356. doi: 10.1038/294354a0
Valdemarsen, T., Bannister, R. J., Hansen, P. K., Holmer, M., and Ervik, A. (2012). Biogeochemical malfunctioning in sediments beneath a deep-water fish farm. Environ. Pollut. 170, 15–25. doi: 10.1016/j.envpol.2012.06.007
Veuger, B., and van Oevelen, D. (2011). Long-term pigment dynamics and diatom survival in dark sediment. Limnol. Oceanogr. 56, 1065–1074. doi: 10.4319/lo.2011.56.3.1065
Vidal, M., Morgui, J. A., Latasa, M., Romero, J., and Camp, J. (1997). Factors controlling seasonal variability of benthic ammonium release and oxygen uptake in Alfacs bay (Ebro delta, NW Mediterranean). Hydrobiologia 350, 169–178. doi: 10.1023/A:1003096117678
Villéger, S., Mason, N. W. H., and Mouillot, D. (2008). New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology 89, 2290–2301. doi: 10.1890/07-1206.1
Waldbusser, G. G., Marinelli, R. L., Whitlatch, R. B., and Visscher, P. T. (2004). The effects of infaunal biodiversity on biogeochemistry of coastal marine sediments. Limnol. Oceanogr. 49, 1482–1492. doi: 10.4319/lo.2004.49.5.1482
Wei, C. L., Rowe, G. T., Escobar-Briones, E., Boetius, A., Soltwedel, T., Caley, M. J., et al. (2010). Global patterns and predictions of seafloor biomass using Random Forests. PLoS ONE 5:e15323. doi: 10.1371/journal.pone.0015323
Whitledge, T. E., Reeburgh, W. S., and Walsh, J. J. (1986). Seasonal inorganic nitrogen distributions and dynamics in the southeastern Bering Sea. Cont. Shelf Res. 5, 109–132. doi: 10.1016/0278-4343(86)90012-9
Woodin, S. A., Wethey, D. S., and Volkenborn, N. (2010). Infaunal hydraulic ecosystem engineers: cast of characters and impacts. Integr. Comp. Biol. 50, 176–187. doi: 10.1093/icb/icq031
Worm, B., Barbier, E. B., Beaumont, N., Duffy, J. E., Folke, C., Halpern, B. S., et al. (2006). Impacts of biodiversity loss on ocean ecosystem services. Science 314, 787–790. doi: 10.1126/science.1132294
WoRMS Editorial Board (2015). World Register of Marine Species [Online]. Available online at: http://www.marinespecies.org (Accessed 2015-12-02)
Yachi, S., and Loreau, M. (1999). Biodiversity and ecosystem productivity in a fluctuating environment: the insurance hypothesis. Proc. Natl. Acad. Sci. U.S.A. 96, 1463–1468. doi: 10.1073/pnas.96.4.1463
Keywords: biodiversity, ecosystem functioning, functional diversity, environmental variables, benthic fluxes, organic matter remineralization, Salish Sea
Citation: Belley R and Snelgrove PVR (2016) Relative Contributions of Biodiversity and Environment to Benthic Ecosystem Functioning. Front. Mar. Sci. 3:242. doi: 10.3389/fmars.2016.00242
Received: 08 September 2016; Accepted: 04 November 2016;
Published: 18 November 2016.
Edited by:
Thomas Good, NOAA Fisheries, USAReviewed by:
Ana M. Queiros, Plymouth Marine Laboratory, UKPaul E Renaud, Norwegian Institute for Water Research, Norway
Copyright © 2016 Belley and Snelgrove. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rénald Belley, cmVuYWxkLmJlbGxleUBtdW4uY2E=