 Amalia Grau1,2
Amalia Grau1,2 Antonio Villalba3,4,5*
Antonio Villalba3,4,5* José I. Navas6Beatriz Hansjosten1
José I. Navas6Beatriz Hansjosten1 José M. Valencia1,2
José M. Valencia1,2 José R. García-March7
José R. García-March7 Patricia Prado7,8
Patricia Prado7,8 Guillermo Follana-Berná1,2Titouan Morage9
Guillermo Follana-Berná1,2Titouan Morage9 Maite Vázquez-Luis10Elvira Álvarez10
Maite Vázquez-Luis10Elvira Álvarez10 Pantelis Katharios11
Pantelis Katharios11 Christina Pavloudi11Elisabet Nebot-Colomer10
Christina Pavloudi11Elisabet Nebot-Colomer10 José Tena-Medialdea7
José Tena-Medialdea7 Monserrat Lopez-Sanmartín6
Monserrat Lopez-Sanmartín6 Claire Peyran9Hrvoje Čižmek12
Claire Peyran9Hrvoje Čižmek12 Georgia Sarafidou11
Georgia Sarafidou11 Yiannis Issaris13
Yiannis Issaris13 Inci Tüney-Kizilkaya14
Inci Tüney-Kizilkaya14 Salud Deudero10
Salud Deudero10 Serge Planes9
Serge Planes9 Gaetano Catanese1,2*
Gaetano Catanese1,2*- 1LIMIA - Laboratorio de Investigaciones Marinas y Acuicultura – Govern de les Illes Balears, Balearic Islands, Spain
- 2INAGEA (INIA-CAIB-UIB), Balearic Islands, Spain
- 3Centro de Investigacións Mariñas, Consellería do Mar, Xunta de Galicia, Vilanova de Arousa, Spain
- 4Departamento de Ciencias de la Vida, Universidad de Alcalá, Alcalá de Henares, Spain
- 5Research Centre for Experimental Marine Biology and Biotechnology, University of the Basque Country, Plentzia, Spain
- 6IFAPA Centro Agua del Pino, Junta de Andalucía, Cartaya, Spain
- 7Instituto de Investigación en Medio Ambiente y Ciencia Marina, Universidad Católica de Valencia, Calpe, Spain
- 8IRTA-Sant Carles de la Ràpita, Sant Carles de la Ràpita, Spain
- 9PSL Research University, EPHE - UPVD - CNRS, USR 3278 CRIOBE, Perpignan, France
- 10Instituto Español de Oceanografía (IEO, CSIC), Centro Oceanográfico de Baleares, Palma de Mallorca, Spain
- 11Institute of Marine Biology, Biotechnology and Aquaculture, Hellenic Centre for Marine Research, Heraklion, Greece
- 12Marine Explorers Society – 20000 Leagues, Zadar, Croatia
- 13Institute of Oceanography, Hellenic Centre for Marine Research, Attiki, Greece
- 14Department of Biology, Faculty of Science, Ege University, Bornova, Turkey
A mass mortality event (MME) affecting the fan mussel Pinna nobilis was first detected in Spain in autumn 2016 and spread north- and eastward through the Mediterranean Sea. Various pathogens have been blamed for contributing to the MME, with emphasis in Haplosporidium pinnae, Mycobacterium sp. and Vibrio spp. In this study, samples from 762 fan mussels (necropsies from 263 individuals, mantle biopsies from 499) of various health conditions, with wide geographic and age range, taken before and during the MME spread from various environments along Mediterranean Sea, were used to assess the role of pathogens in the MME. The number of samples processed by both histological and molecular methods was 83. The most important factor playing a main role on the onset of the mass mortality of P. nobilis throughout the Mediterranean Sea was the infection by H. pinnae. It was the only non-detected pathogen before the MME while, during MME spreading, its prevalence was higher in sick and dead individuals than in asymptomatic ones, in MME-affected areas than in non-affected sites, and it was not associated with host size, infecting both juveniles and adults. Conversely, infection with mycobacteria was independent from the period (before or during MME), from the affection of the area by MME and from the host health condition, and it was associated with host size. Gram (-) bacteria neither appeared associated with MME.
Introduction
The fan mussel Pinna nobilis is a Mediterranean endemic, marine bivalve mollusc now critically endangered (IUCN, 2019). During the last decades, P. nobilis populations have progressively decreased due to anthropogenic impacts, mainly related to the destruction of their habitats, including illegal fishing, ornamental harvesting, and accidental killing by anchoring, bottom nets and trawlers (Basso et al., 2015; Vázquez-Luis et al., 2017). However, an unusual mass mortality event (MME) of this species was first detected in the SE Spanish coast in autumn 2016 (Vázquez-Luis et al., 2017) and fast spread north- and eastward, reaching France, Italy, Morocco, Tunisia, Malta, Greece, Croatia, Bosnia and Herzegovina, Cyprus and Turkey in 3 years (IUCN, 2018, 2020; Carella et al., 2019; Katsanevakis et al., 2019, 2021; Panarese et al., 2019; Čelebičić et al., 2020; Čižmek et al., 2020; Lattos et al., 2020a; Öndes et al., 2020; Özalp and Kersting, 2020; Scarpa et al., 2020; Zotou et al., 2020; Acarli et al., 2021; Çinar et al., 2021).
Knowing the cause of this MME is crucial to implement measures to avoid its extinction and to find recovery ways. However, its primary cause (or interrelated causes) remains an enigma until now. Two microorganisms have been proposed as the most plausible aetiological agents. From a chronological point of view, a member of Haplosporida, a protistan group enclosing parasites highly pathogenic for bivalves (Hartikainen et al., 2014; Arzul and Carnegie, 2015), was first detected associated to high fan mussel mortality in Calpe (Spain) (Darriba, 2017). Afterward, Fanelli et al. (2018) pointed out a Haplosporidium sp. as the aetiological agent in different Italian localities and Catanese et al. (2018) described that haplosporidan as the new species Haplosporidium pinnae, proposing it as the most likely cause of the MME. Later, Carella et al. (2019) reported Mycobacterium sp., close to a terrestrial-like non-tuberculous mycobacterium (NTM), with stronger association with the P. nobilis MME along the Tyrrhenian coastline (Italy) than H. pinnae, thus introducing a new possible aetiological agent into the aetiological scientific debate. At the same time, two publications again identified H. pinnae in the massive die-offs observed in the Aegean (Katsanevakis et al., 2019) and Ionian Seas (Panarese et al., 2019) but without making a differential diagnosis with Mycobacterium sp., the newly detected possible aetiological agent, as is also the case with the detection of H. pinnae in the Ligurian Sea (Betti et al., 2021). Since then, mixed infections of these two pathogens affecting fan mussels have been reported in Adriatic Sea (Čižmek et al., 2020; Šarić et al., 2020), in Aegean Sea (Lattos et al., 2020a) and, with further co-infection with other pathogens, in the Tyrrhenian Sea, Ebro Delta (NE Spain) and Aegean Sea (Carella et al., 2020; Lattos et al., 2020b,2021; Scarpa et al., 2020). Additionally, the Gram (−) bacterium Vibrio mediterranei, previously described as an opportunistic emerging pathogen associated with mortalities in the wild (Rodrıguez et al., 2017), has again been detected in natural and stabled fan mussels (Prado et al., 2020a; Andree et al., 2021; Lattos et al., 2021). Nowadays, co-infection with H. pinnae and bacteria of various Vibrio spp., has been proposed as the cause of the MME in the Çanakkale Strait (Turkey) (Künili et al., 2021). The more recent findings could suggest a more complex scenario than that originally proposed in the first descriptions of the P. nobilis MMEs. Diseases with complex aetiology, influenced by various interacting drivers, frequently remain difficult to characterise due to reductionist approaches (De Lorgeril et al., 2018).
The International Union for the Conservation of Nature (IUCN, 2019) proposed a list of conservation recommendations with the objective of avoiding the plausible total extinction of the species. The main was “to investigate on the pathogens and vectors to conduct a deep study of the origin, epizootic dynamics and the complete life cycle of H. pinnae, as well as to clarify the pathogenicity and potential virulence of the reported mycobacteriosis.” However, the critically endangered status of this species involves many restrictions for experimentation, which complicates that research. To overcome these difficulties, contributions of multiple teams in various countries were assembled to configure a wide geographic, long term approach robust enough to support solid conclusions. This article reports the results of such a broad survey, based on histological and molecular diagnoses, including alive fan mussels collected before MME (2011–2014, from populations in a good state of conservation, but today extinct) and fan mussels in three classes of health condition (asymptomatic, sick and recently dead individuals) collected during the MME (2016–2020) in different areas, in order to: (1) assess the occurrence of potential pathogens (particularly H. pinnae, mycobacteria and Gram (−) bacteria) in fan mussels before the MME; (2) determine the occurrence of infectious agents in recently dead, sick and asymptomatic fan mussels during the different outbreaks of the ongoing MME; and (3) compare these two different scenarios, before and during MME, to unravel the relationships between the observed pathogenic microorganisms regarding the onset of the fan mussel MME.
Materials and Methods
Sampling
Samples involving 762 fan mussels (whole individuals or biopsies) from 30 areas scattered through eight countries (Spain, France, Monaco, Italy, Croatia, Greece, Cyprus and Turkey) that had been collected by scuba diving were gathered (Table 1 and Figure 1). Due to the urgency of the situation, many researchers have contributed to the sampling collection, forming an interdisciplinary panel of researchers (all of them experts in the biology of P. nobilis). In the case of samples obtained during the various outbreaks of massive mortality detected, the fixed samples were immediately sent to the diagnostic laboratories, where they were processed for further study. All histology samples were processed at LIMIA (with the sole exception of some samples from Croatia-Zagreb University and Greece-HCMR Lab). Likewise, most of the samples for molecular analysis were also processed at LIMIA (with the exception of few samples from Greece). The processing protocols were the same in all laboratories. The processing time was always less than 1 month after sampling. All the samples before MME were analysed at LIMIA. The diagnosis of all the histological samples was always carried out in LIMIA, and was also the molecular diagnosis for the most part, with the exception of Greece. In LIMIA, the molecular results obtained in Greece were subsequently verified.
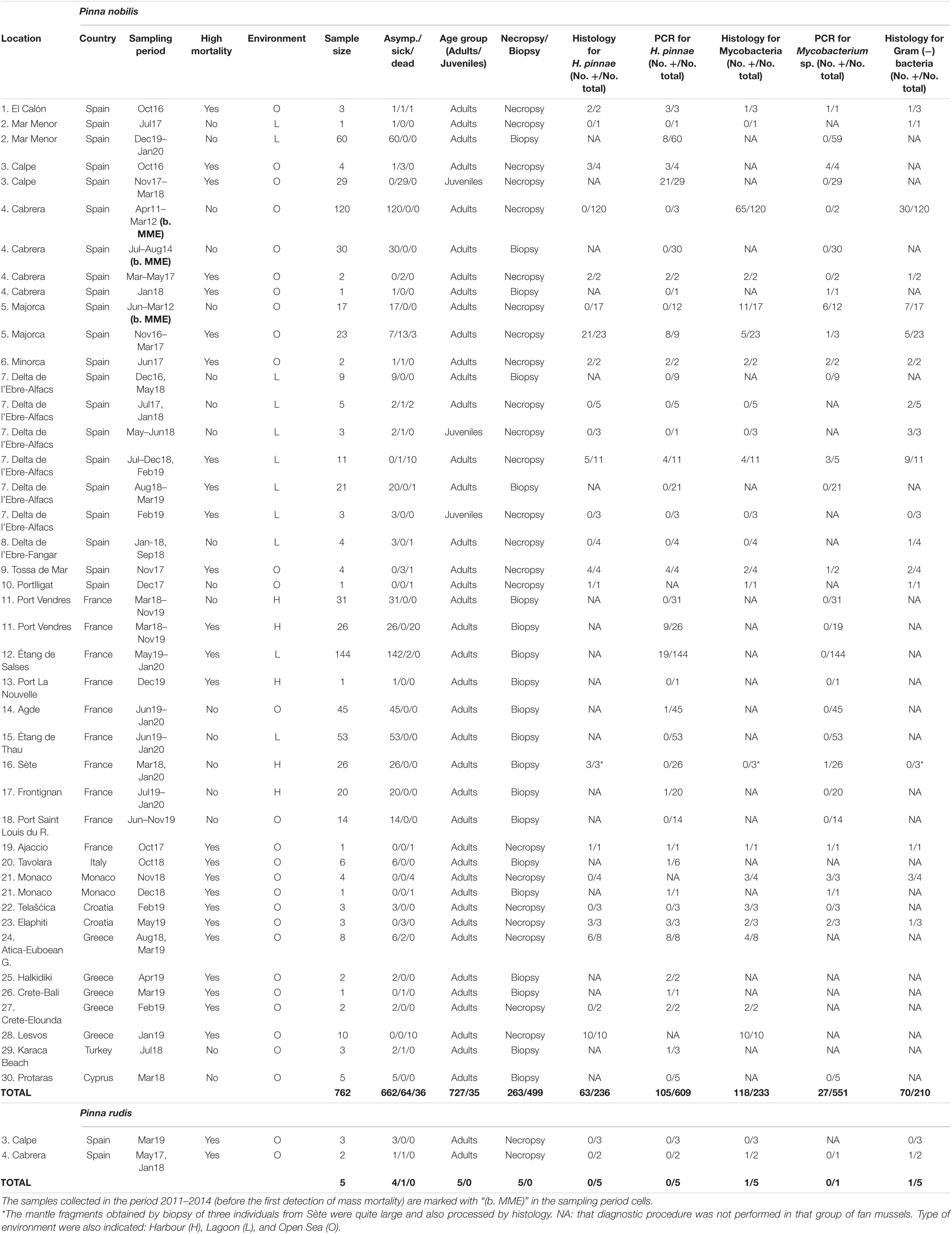
Table 1. Summary of sampling and diagnosis information, including sampling locations; sampling period; whether high mortality had occurred in the sampling location or not; sample size; number of fan mussels asymptomatic, sick and recently dead; age group (adults or juveniles); procedure of tissue removal (necropsy or biopsy); and the diagnostic results for Haplosporidium pinnae, mycobacteria and Gram (−) bacteria [No. positive (+) cases/No. diagnosed individuals].
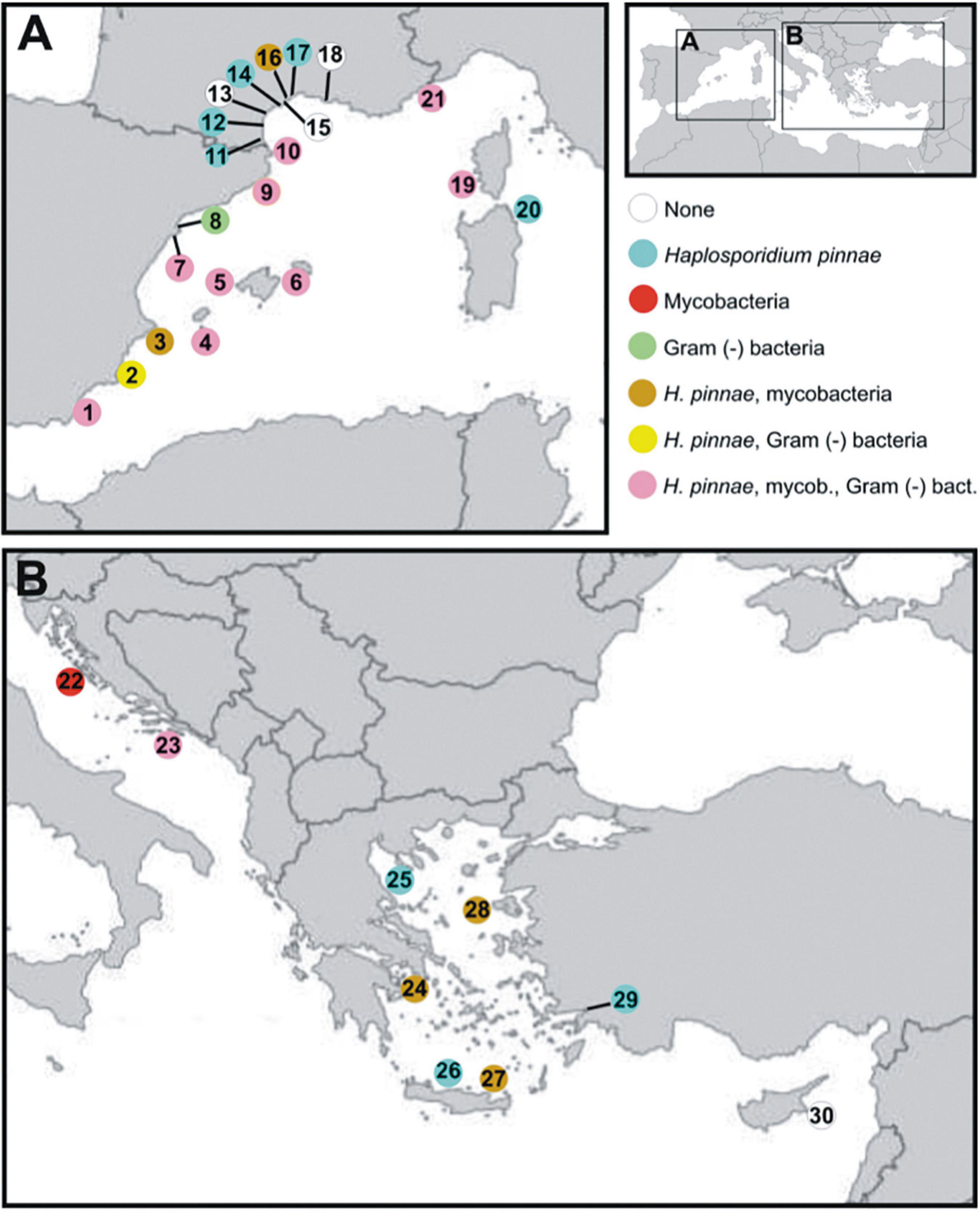
Figure 1. Map of the Mediterranean Sea showing sampling locations. Next is the list of sampling locations with indication of the ecosystem type (H: harbour; L: coastal lagoon; S: open sea). (A) Spain: El Calón, S (1); Mar Menor, including Isla Barón, L, Isla Perdiguera, L, and Pinna Point, L (2); Calpe, including Calpe, S, and Villajoyosa, H (3); Cabrera, including Castell de Cabrera, S, and Santa María, S (4); Majorca, including Andratx, S and Magaluf, S (5); Minorca, including Son Saura, S, Boya Rey, Cala Gandulf, S, and Rates, S (6); Delta de l’Ebre-Alfacs, L (7); Delta de l’Ebre-Fangar, L (8); Tossa de Mar, S (9); Portlligat, S (10). France: Agde, including Agde, S, and Port Ambonne, H (11); Étang de Salses, including Port Leucate, H, Leucate Nord, L, Leucate Centre, L, Leucate Sud, L, and Lido Port-Leucate, H, (12); Étang de Thau, including Grau des Quilles, L, Marseillan Plage, L, and Meze, L (13), Port de Frontignan, H (14); Port La Nouvelle, H (15); Port Saint Louis du Rhone, L (16); Port Vendres, including Plage de Peyrefite, S, and Port Vendres, H (17); Port-Sète, H (18); Ajaccio, S (19). Italy: Tavolara, S (20). Monaco: Monaco, S (21). (B) Croatia: Telašćica, S (22); Elaphiti, S (23). Greece: Atica-Euboean Gulf, including Lavrio, S, Avilda, S, and Oropos, S (24); Halkidiki, S (25); Crete-Bali, S (26); Crete-Elounda, S (27); Lesvos, S (28). Turkey: Karaka Beach, S (29). Cyprus: Protaras, S (30). Colours of the circles correspond to the pathogens [Haplosporidium pinnae, mycobacteria or Gram (−) bacteria] detected in each location.
The sample set included fan mussels in three classes of health condition (asymptomatic, sick and recently dead individuals), three environments (coastal lagoons, harbours, and open sea) and wide size/age range, with both juvenile and adults. Asymptomatic specimens had typical healthy responses and normal mantle appearance when sampling. Sick fan mussels showed no response to stimuli, with pale and retracted mantle, and slow valve closure. Recently dead individuals were those in which decomposition process had started but the presence of tissues that were still preserved and consistent allowed sampling. In the natural environment it is difficult to determine the time of death of these individuals. From observations of fan mussels maintained in captivity, recently dead individuals presumably corresponding to specimens that have died less than 36 h before (García-March personal communication).
A total of 126 whole specimens and 469 mantle biopsies were collected during MME spreading (2016–2020); additionally, 137 alive whole specimens and 30 mantle biopsies corresponded to the period 2011–2014, before the MME was first noticed. Samples in the period 2016–2020 were collected in the context of population surveillance or even awareness of first signs of abnormally high mortality in some of those populations by scientists, citizens and services for marine protected areas management, after the first detection of the MME in autumn 2016. Thus, high fan mussel mortality had affected some locations when sampling, while it had not occurred yet in others, which were sampled in the context of the alarming news from neighbouring areas. These samplings included collection of recently dead, sick and asymptomatic whole fan mussels, for a total of 126 specimens (more details in Table 1). The sacrifice of some asymptomatic and sick individuals was considered necessary to search for the causes of each mortality outbreak by analysing organs that cannot be sampled without killing the fan mussels. Most of those collected fan mussels were adults but 29 juvenile specimens (ca. 40 mm in dorso-ventral length) that had been settled on the anchorage ropes of finfish culture cages near Calpe and 6 more juveniles (ca. 180 mm in dorso-ventral length) collected from the “Banya” sandspit of Delta de l’Ebre-Alfacs were also included. Mantle biopsies were taken from other 469 adult specimens, without removing them from their habitat, during that period, using a specific procedure for P. nobilis preventing killing (Wesselmann et al., 2018). Additionally, samples that had been collected from 2011 to 2014 for other scientific purposes (characterisation of the gonad cycle and other physiological processes as well as search for potential pathogens) from Balearic Islands populations, extinct today, were included in this study to assess the potential pathogenic load of fan mussels before the MMEs, thus allowing comparison of the pathogenic load before and during MMEs. Namely, 120 live individuals were collected by scuba diving in the Cabrera National Park Marine Protected Area (CNP-MPA) and 17 live individuals from other fishing exploitation localities in Majorca Island in 2011–2012 (Deudero et al., 2017; Box et al., 2020); additionally, 30 mantle biopsies were taken from alive individuals located in CNP-MPA in 2014 (Table 1). A summary of sampling and diagnosis information is provided in Table 1. The number of samples processed by both histological and molecular methods was 83.
Furthermore, five spiny fan mussels Pinna rudis, a species not affected by high mortality (Vázquez-Luis et al., 2017; Catanese et al., 2018), were collected from two areas affected by the P. nobilis MME for comparative analysis.
Due to P. nobilis being a strictly protected species (Annex II of Barcelona Convention and Annex IV of the European Council Directive 92/43/EEC), all samplings were carried out under permission of regional and national authorities (listed below in Ethics Declarations: Permissions). For this reason, during the MME, a limited number of recently dead, sick and asymptomatic individuals were collected from each location. However, in the authorised survey conducted before the MME (2011–2012), approximately 9–12 individuals were collected every month. This number represented a very low percentage of the fan mussel population in the CNP-MPA at that time and the natural population showed no signs of disease (Vázquez-Luis et al., 2014; Deudero et al., 2015).
Specimens collected before MME were measured (dorso-ventral and antero-posterior shell lengths) and weighed (total and fresh weight) in the laboratory. Other biometric parameters were obtained from stored tissues. During MME, measures were taken, when possible, while performing visual census surveys, although sometimes it was not done due to the inherent difficulties of field sampling by scuba diving. In the case of mantle biopsies from live individuals, only shell width was measured when possible. Individuals collected for diagnosis embraced a wide length range, from juveniles to large adults (from 2.4 to 71 cm in shell length). Additional information, such as depth, temperature and environment, was also recorded when possible. The sampling effort covered all the depths that scuba divers can easily reach (0.3–35 m) and the different types of bottoms this long-living bivalve inhabits. Environments were directly recorded by direct census or compiled from data reported in scientific articles (Darriba, 2017; Katsanevakis et al., 2019; Čižmek et al., 2020), also indicating if sampling was made in coastal open sea, coastal lagoons or harbours. Although it is not specifically a lagoon, Delta de l’Ebre estuarine (Alfacs and Fangar bays) was considered as such, due to its semi-enclosed water bodies with freshwater inputs, its geomorphology, environmental conditions and paralic sedimentation that make it different in relation to the open sea (Prado et al., 2021).
Sample Processing and Histological Analysis
The biopsies consisted of mantle pieces taken in situ, which were preserved in absolute ethanol for molecular diagnosis. Exceptionally, the mantle biopsies from three individuals located in Sète were large enough to be also processed by histology.
The collected whole fan mussels (necropsies), either asymptomatic, sick or recently dead, were dissected and soft tissues preserved in 10% phosphate-buffered formalin or in Davidson’s fixative for histological analysis. For molecular diagnosis, before MME, samples of soft tissues were stored in absolute ethanol or at −80°C while during MME the samples were preserved only in absolute ethanol (Table 1). However, the whole soft tissues of the 29 juvenile fan mussels collected from Calpe were preserved in absolute ethanol and processed for molecular diagnosis. Additionally, the Ray’s fluid thioglycolate medium (RFTM) assay (Ray, 1963) was performed with gill pieces from individuals collected in 2011–2014 to assess infection with Perkinsus spp. For histology, a cross section of the visceral mass at the level of the digestive gland was taken from the fixed soft tissues in the case of adults, while the whole fixed soft tissues were used in the case of juveniles. Digestive gland was chosen for diagnosis because it is the target organ for H. pinnae sporulation (Catanese et al., 2018) as well as one of the organs infected with Mycobacterium sp. when it occurs; due to anatomical continuity, organs in contact with the digestive gland, namely the gonad, were partially included in the tissue pieces processed by histology. Most samples were dehydrated in an increasing ethanol gradient and cleared with X-free, embedded in paraffin wax and sectioned at 3–4 μm thickness; other samples were included in Technovit 7100 (Kulzer, Germany) and sectioned at 1–2 μm. In the case of the histological analyses of the samples prior to the MME (samples from 2011 to 2012) they were performed on new sections of the stored paraffin blocks. Sections from each block were stained with Mayer’s haematoxylin and eosin (MHE). Some additional sections were stained with Mayer’s haematoxylin-VOF (Gutiérrez, 1967), with which H. pinnae spores become highlighted (Catanese et al., 2018). Gram (Scharlab histo kit) and Ziehl-Neelsen (ZN; Luna, 1968) staining procedures were also performed to facilitate detection of bacteria and acid-fast bacteria (Mycobacterium sp.), respectively in most of the samples.
Molecular Analysis
Small pieces of the collected tissues were chopped with sterile scissors and used for DNA extraction; triplicate extractions were performed for each tissue, in order to minimise biases. Total DNA was extracted from mantle and/or digestive gland pieces (adults) or homogenised tissues (juveniles), using the NucleoSpin® Tissue DNA Isolation Kit (Macherey-Nagel), according to the manufacturer’s instructions.
For the Greek samples, DNA was extracted by precipitation with isopropanol and ammonium acetate (5 M) following the manual of Sambrook et al. (1989). Each sample was washed with 800 ul of sterile distilled water for 15 min, following centrifugation at 13,000 × g for 2 min. The supernatant was removed and the wash was repeated. Afterward, each sample was washed with 600 ul of lysis buffer (0.5 M Tris, 0.1 M EDTA, 2% SDS, pH 8.8) for 15 min, following centrifugation at 13,000 × g for 2 min and removal of the supernatant. The washes with the lysis buffer were repeated twice. The pellet was mixed with 600 μl of lysis buffer and 6 ul of proteinase K (20 mg/ml) and incubated at 55°C overnight.
In the final step of the DNA extraction protocol, i.e., the elution of the DNA pellet, replicate samples were pooled and their concentration was measured in a NanoDrop 1000 spectrophotometer.
The nucleic acids were subjected to PCR amplification employing the primer pairs HPNF3/HPNR3 (Catanese et al., 2018; López-Sanmartín et al., 2019) for H. pinnae, and mycgen-f/mycgen-r for Mycobacterium spp. (Böddinghaus et al., 1990; Carella et al., 2019), using the same PCR conditions applied by the authors who described them. All the obtained PCR products were purified using a mi-Gel Extraction Kit (Metabion, Germany) and bi-directionally sequenced using an ABI 3130 Genetic Analyser (Applied Biosystems, Carlsbad, CA, United States); the sequences were analysed using the BLAST algorithm at the NCBI server1 for species identification. PCR products obtained from samples collected before the onset of MME were sequenced to confirm and/or to identify the species detected by histology. However, the fact of obtaining PCR amplifications after years of sampling indicates good quality and acceptable storage of the samples.
Ray’s Fluid Thioglycolate Medium Assays
A piece of one gill lamella of each fan mussel collected in 2011–2012 (see Deudero et al., 2017) from CNP-MPA and Majorca, was incubated in Ray’s fluid thioglycolate medium (RFTM, Ray, 1963), supplemented with chloramphenicol and nystatin, for 1 week at room temperature in the dark, to detect hypnospores of Perkinsus spp. Afterward, tissues were crushed on a slide, stained with Lugol’s iodine solution, covered and observed under light microscopy to estimate the severity of Perkinsus infection using the modified Ray’s scale (Andrews and Hewatt, 1957) with six severity degrees, from 0 (no parasite detected) to 5 (very heavy infection).
Statistical Analysis
Due to the variable nature of the available samples (biopsies or necropsies), some could only be analysed by PCR, others by histology and others by both. In the absence of a gold standard to evaluate the sensitivity and specificity of each diagnostic method, the agreement between the histological and molecular diagnostic methods for both H. pinnae and Mycobacterium sp. was evaluated using Cohen’s kappa coefficient (κ) (Cohen, 1960) and interpreted according to Landis and Koch (1977). The following associations were tested using contingency tables and the chi square (X2) test: (1) association between the period regarding the MME (either before the first detection or during the MME spreading) and the occurrence of H. pinnae, mycobacteria or Gram (−) bacteria, respectively, in the fan mussels; (2) between the health condition of the fan mussels (either asymptomatic, sick or recently dead individuals) and the occurrence of the mentioned pathogens; (3) between the affection of the sampling area by MME (either yes or not) and the occurrence of the mentioned pathogens; (4) between the occurrence of the mentioned pathogens and host size (10 cm length) classes; (5) between the occurrence of H. pinnae and the environment type (either coastal lagoon, harbour or open sea) and (6) paired associations between the mentioned pathogens. Significance was considered at P ≤ 0.05.
Results
Histological and Molecular Diagnosis
Histological examination showed infection with H. pinnae in 63 of the 236 analysed fan mussels; no detection corresponded to individuals collected before the MME. Stages of the parasite sporulation process were detected in eight asymptomatic and eight sick individuals (Table 1 and Figure 2). Considering the 60 cases of detection of H. pinnae in histological sections that included digestive gland (the only organ where sporulation occurs), sporulation stages were detected in 16/60 samples (27%) of the infected individuals. Regarding diagnosis by PCR, H. pinnae was detected in 105/609 (17%) of the analysed fan mussels; again, no positive case was detected in individuals collected before the MME (Table 1).
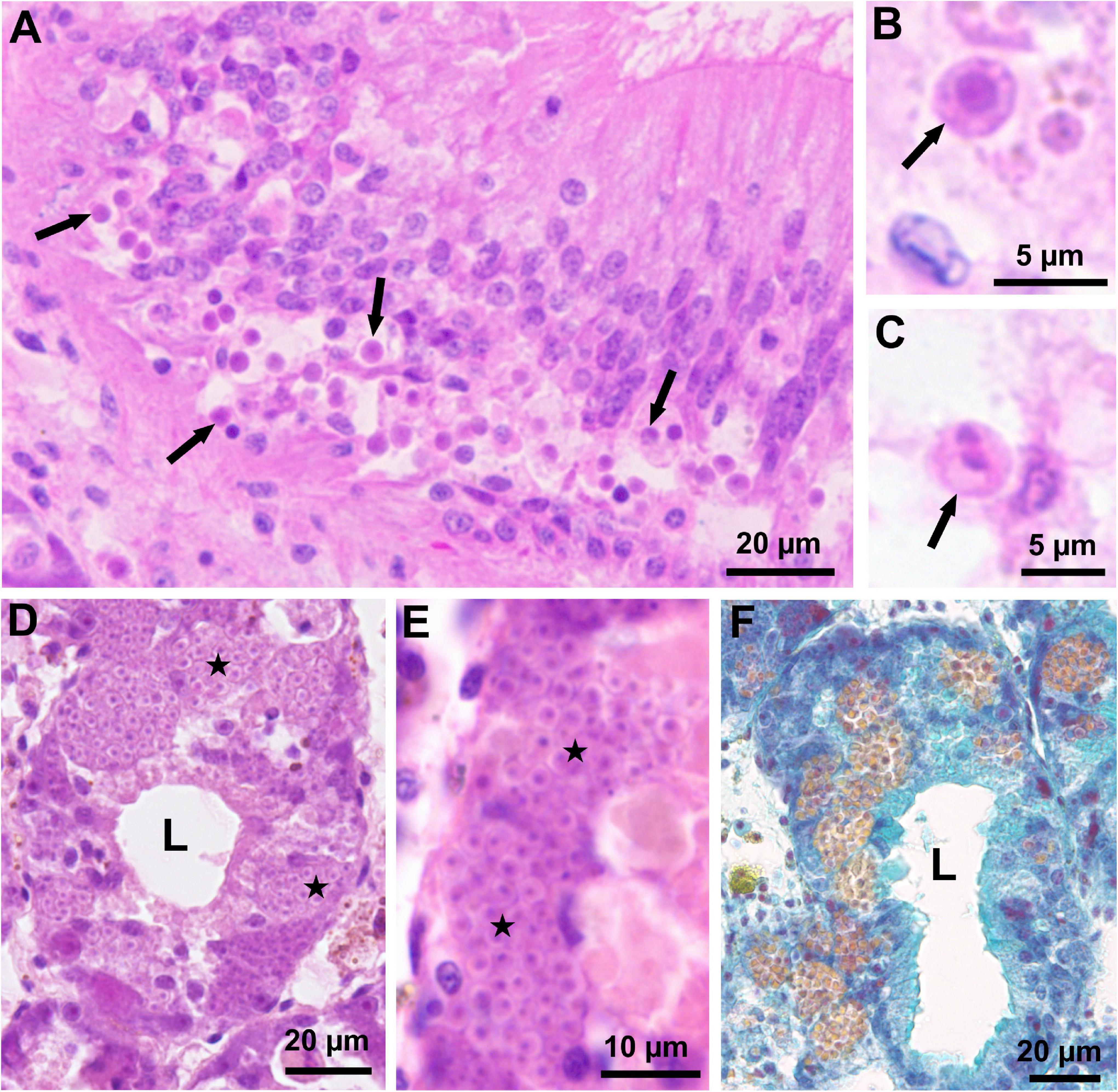
Figure 2. Light micrographs of histological sections through the visceral mass of fan mussels Pinna nobilis infected with Haplosporidium pinnae. (A) Section through the intestine showing numerous uninucleate cells of H. pinnae (arrows) associated with vacuolisation of the host intestinal epithelium (MHE; Lesvos, January 2019). (B) High magnification showing a uninucleate cell of H. pinnae (arrow) in the host connective tissue (MHE; Lesvos, January 2019). (C) High magnification showing a binucleate cell of H. pinnae (arrow) in the host connective tissue (MHE; Lesvos, January 2019). (D) Section through the host digestive gland showing masses of H. pinnae spores (stars) in the epithelium of a digestive tubule (MHE; Majorca, November 2016). (E) High magnification showing masses of H. pinnae spores (stars) in the epithelium of a digestive tubule (MHE; Majorca, November 2016). (F) Section through the host digestive gland, stained with VOF, showing sporulation stages of H. pinnae in the epithelium of digestive tubules; mature spores are easily distinguished due to their yellow colour (Majorca, November 2016). L, lumen of digestive tubules.
Regarding bacterial infections in the samples taken before MME, ZN staining revealed mycobacteria in the connective tissue of the digestive gland and surrounding area (Figures 3A,B) in 76/137 (55%) of the analysed individuals, while Gram staining showed infection with Gram (−) bacteria in 37/137 (27%) (Table 2); coinfection with mycobacteria and Gram (−) bacteria was detected in 24% of the individuals. Positive detection of Mycobacterium sp. occurred in 6/44 (14%) of the PCR-analysed individuals before MME (Table 2). During MME, 42/96 (44%) of the analysed individuals were positive to mycobacteria with the ZN staining and 33/73 (45%) to Gram (−) bacteria (Table 2); mixed infections were also detected. Total positive detection of Mycobacterium sp. occurred in 21/507 (4%) of the PCR-analysed individuals (Table 2).
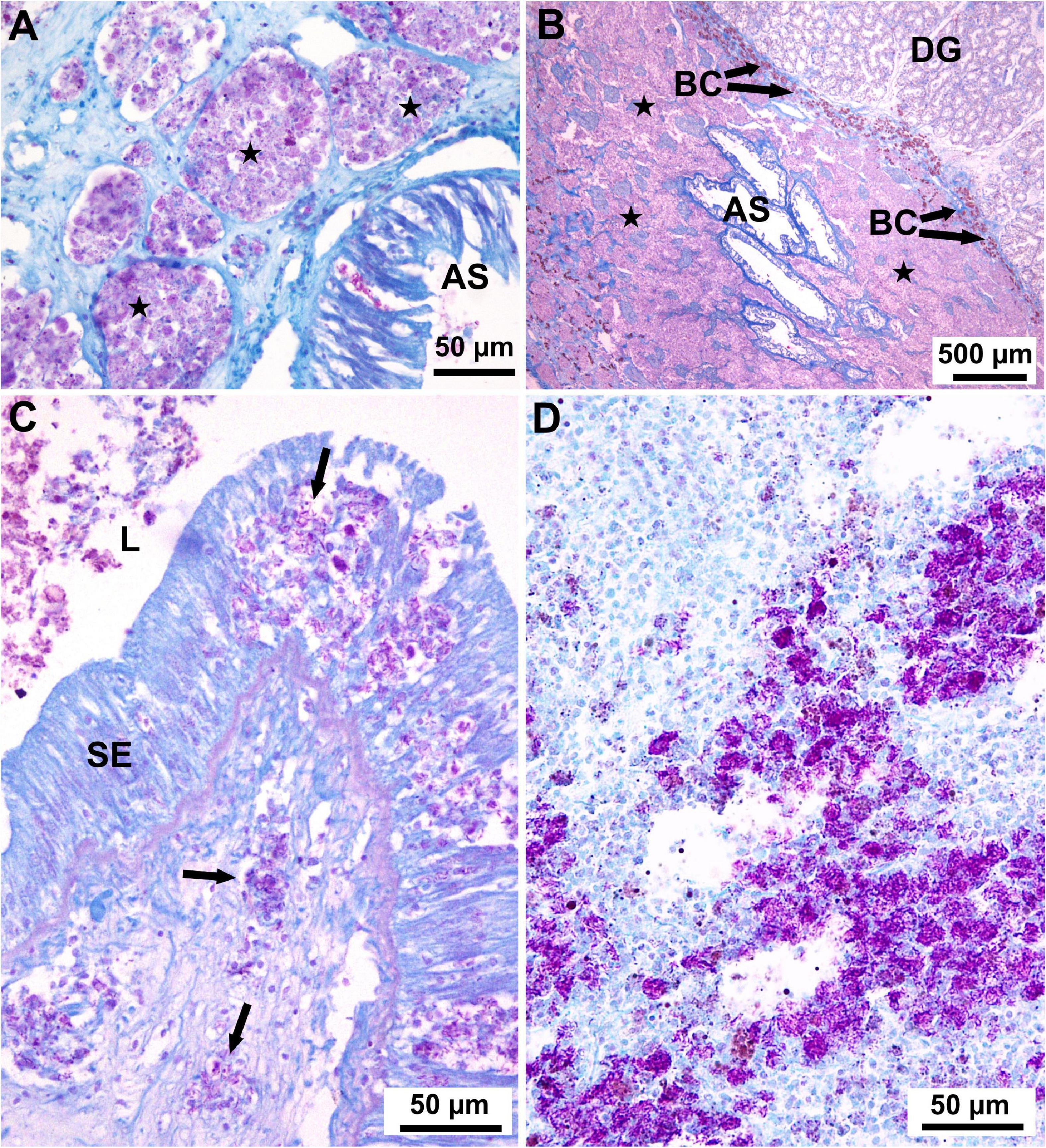
Figure 3. Light micrographs of histological sections through the visceral mass of fan mussels Pinna nobilis stained with Ziehl-Neelsen to show infection with mycobacteria. (A) Aggregations of haemocytes filled with mycobacteria (purple deposits) forming nodules (stars) in the connective tissue surrounding the anterior stomach (AS; Cabrera, January 2012). (B) Low magnification showing massive aggregation (stars) of haemocytes filled with mycobacteria throughout the connective tissue surrounding the anterior stomach (AS); BC: brown cells; DG: digestive gland (Cabrera, November 2011). (C) Section through the stomach showing free haemocytes (not enclosed in nodules, arrows) filled with mycobacteria (purple deposits), both in the stomach epithelium (SE) and in the subjacent connective tissue; L, lumen of the stomach (Majorca, March 2017). (D) Section through the connective tissue showing masses of mycobacteria (purple deposits) spreading outside haemocytes (Minorca, June 2017).
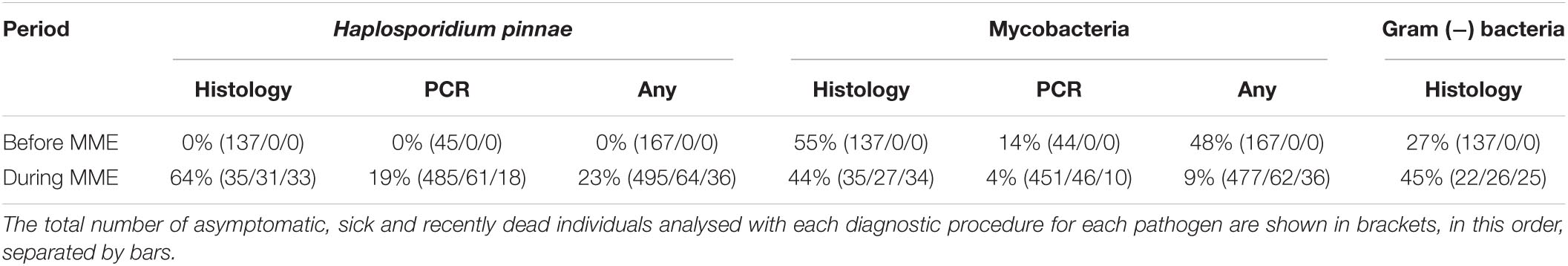
Table 2. Percentages of cases of infection with Haplosporidium pinnae, mycobacteria and Gram-negative bacteria detected before (2011–2014) and during (2016–2020) the MME, using histology, PCR assays or any of these procedures.
The tested DNA sequences through BLAST search showed 100% similarity with that of H. pinnae (accession number: LC338065; LR131124) and those of Mycobacterium sp. (accession numbers: X88906, MH569645, MH569646, MT642064, LR135613), respectively.
A total of 83 fan mussels were diagnosed both with histological and molecular procedures for H. pinnae, with agreement between them in 72/83 (87%) individuals (Table 3); that was “substantial” agreement (kappa = 0.734). Regarding diagnosis of mycobacteria, 42 fan mussels were diagnosed both with histology and PCR, with agreement only in 28/42 (67%) individuals (Table 3); that was “fair” agreement (kappa = 0.211). Remarkably, 20/34 (59%) of the positive cases for mycobacteria by histology were negative by PCR while every positive case for Mycobacterium sp. by PCR was also positive by histology. The high percentage of putative PCR-false negatives for Mycobacterium showed that the molecular procedure implemented in this study was not always appropriate to diagnose these bacteria, likely due to the fact that the mycobacteria infections were frequently restricted to digestive gland and surrounding connective tissue, while it occurred less frequently in the organs involved in the biopsy PCR assays (mantle).
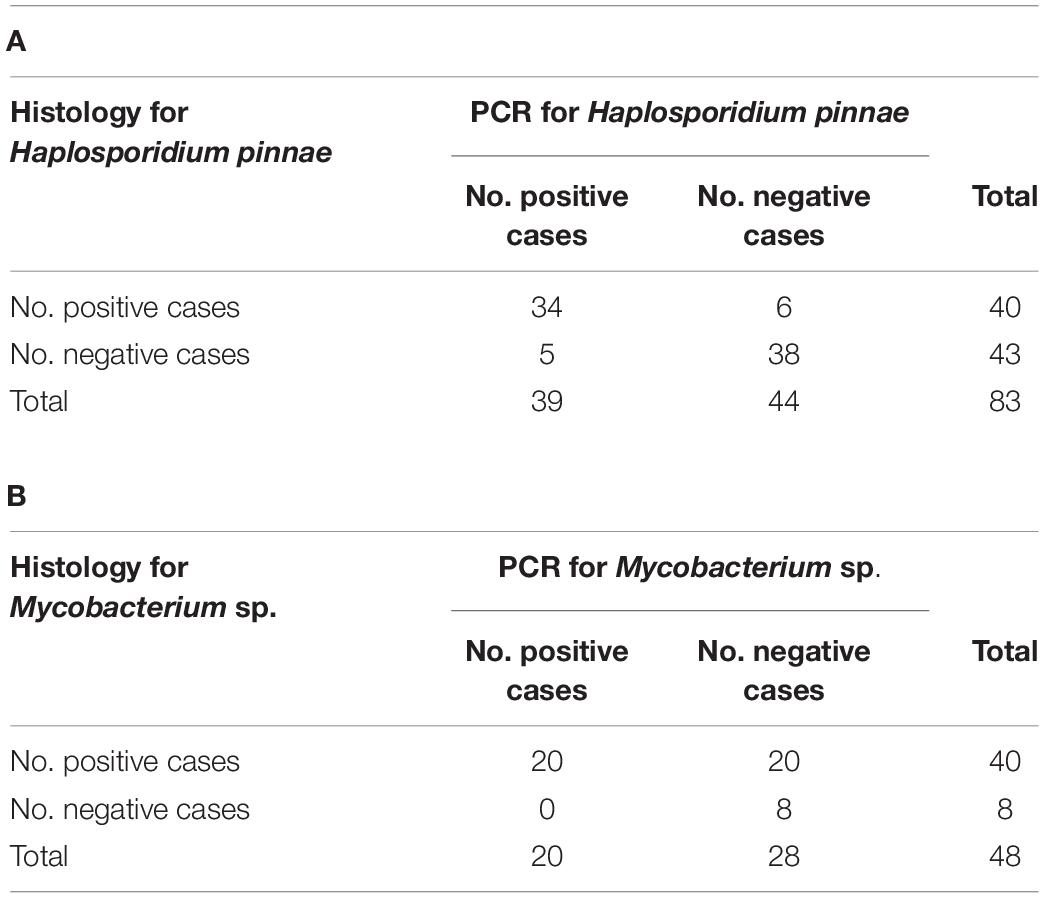
Table 3. Contingency tables for comparison between diagnostic procedures, histology and PCR assays, for Haplosporidium pinnae (A) and mycobacteria (B).
Infection with Perkinsus sp. was not detected in histological sections either before or during MME. The RFTM screening of perkinsosis performed before the MME revealed a moderate prevalence (47/128, 38%) and low severity degree (mean: 0.55) of Perkinsus sp. (Table 4).
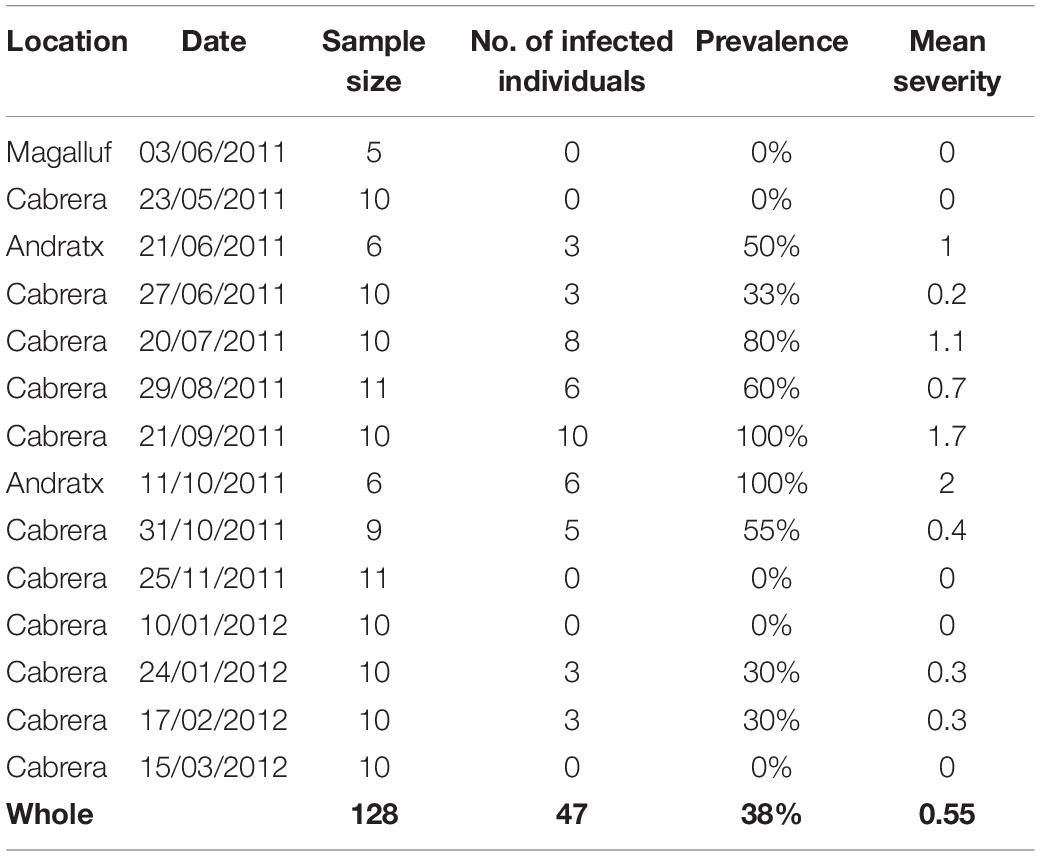
Table 4. Results of RTFM assays for perkinsosis.
In the case of P. rudis, H. pinnae was not detected, while mycobacteria were observed in one asymptomatic individual (Table 1). Gram (−) bacteria were detected in the only sick individual of the five examined.
Histological Remarks of Haplosporidium pinnae Infection
Uninucleate and binucleate cells of the parasite and, less frequently plasmodia with up to six nuclei, occurred in the connective tissue throughout the visceral mass, mantle and gills, as well as in the stomach and intestine epithelia (Figures 2A–C). The infection was frequently associated with heavy haemocytic infiltration of the connective tissue and the affected epithelia. Additionally, stages of parasite sporulation process, including uninucleate and binucleated cells, multinucleate plasmodia, and sporocysts enclosing sporoblasts or more or less mature spores, were detected in the epithelium of digestive tubules (Figures 2D–F) of eight asymptomatic and eight sick individuals from El Calón (1 fan mussel), Calpe (3), Majorca (8), and Attica (4). Parasite sporulation caused severe damage of the digestive tubule epithelium.
Histopathological Remarks of Bacterial Infections
In the samples taken before MME, multiple nodular aggregates of haemocytes were often observed scattered in the connective tissue circumscribing the visceral mass and the anterior stomach, sometimes forming aggregate-rich regions encapsulated by a haemocytic and brown cells response (Figures 3A,B). In some instances, its larger size became a bacterial abscess. Acid-fast positive bacteria were seen within the affected haemocytes and brown cells in the core of the lesion. Less frequently, Gram negative bacteria were also observed. Both nodules and abscesses appeared to effectively sequester the bacteria, since haemocytes enclosing bacteria were not seen outside aggregates. However, mycobacterial infection showed some differences in the samples collected during MME. Mycobacteria-filled haemocytes located in nodular aggregates, were also observed. They were also detected in the connective tissue surrounding visceral mass and anterior stomach but also in the mantle and in the connective tissue surrounding gonadal follicles. In addition, mycobacteria-filled isolated haemocytes were frequently seen spreading throughout the connective tissue of the visceral mass and mantle as well as in the digestive epithelia (Figure 3C). Occasionally, free mycobacteria were seen (Figure 3D).
For comparison, micrographs showing the normal structure of the fan mussel digestive gland, corresponding to a healthy condition, are provided in Figure 4.
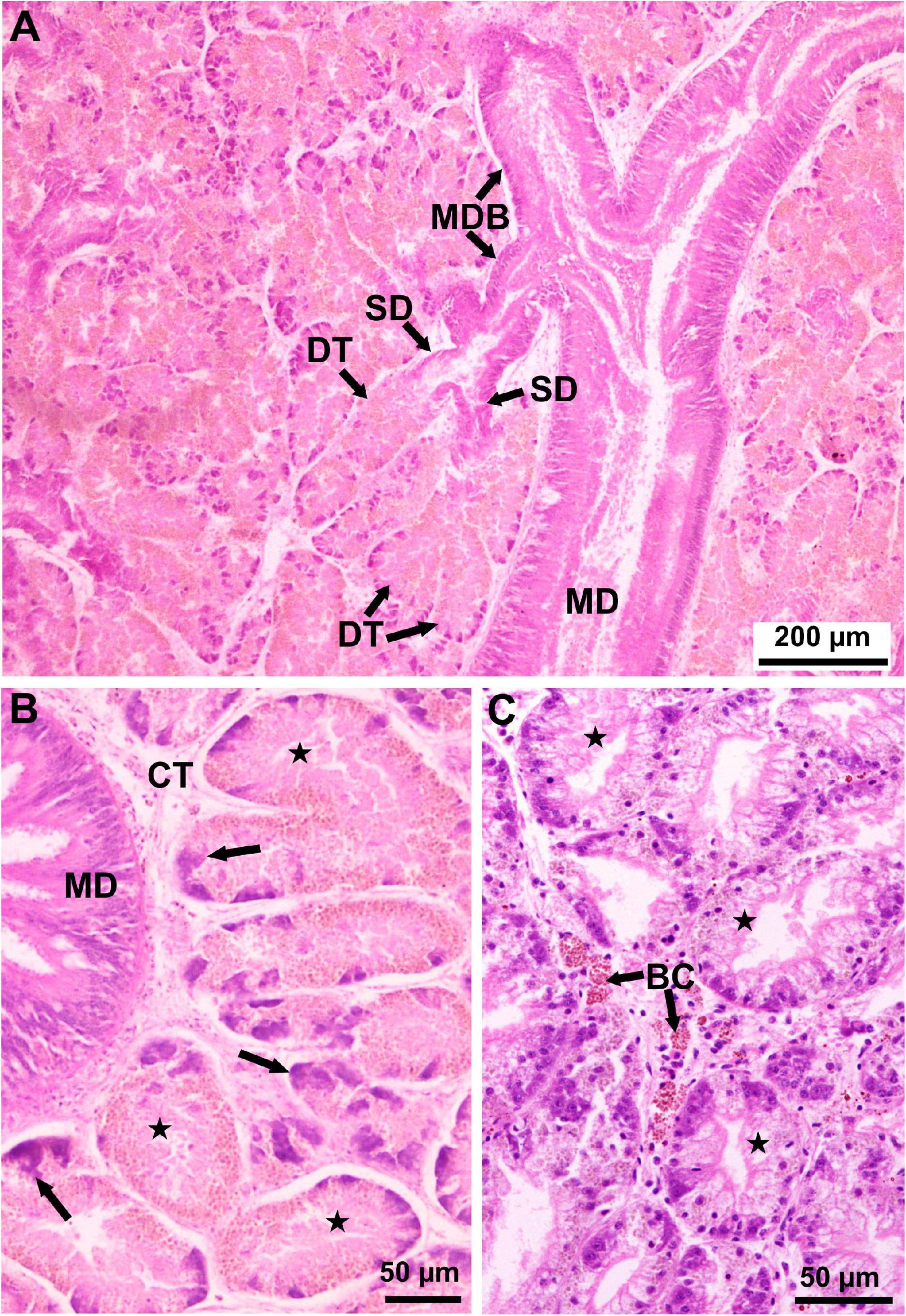
Figure 4. Light micrographs of histological sections stained with MHE showing the normal (healthy) structure of the Pinna nobilis digestive gland. (A) Lower magnification showing a main duct (MD) leading to main duct branches (MDB) from which primary ducts (PD) arise leading to secondary ducts (SD) ending in blind digestive tubules (DT) (Cabrera, May 2011). (B) Higher magnification showing digestive gland tubules (stars) in absorptive phase surrounding a stomach branch (SB); the digestive tubules show highly reduced lumen and tall epithelium, consisting of acidophilic absorptive cells and basophilic young, differentiating cells (arrows); CT, connective tissue (Cabrera, May 2011). (C) Higher magnification showing digestive gland tubules (stars) in holding phase, in which digestive tubules show larger lumen; some brown cells (BC) occur in the connective tissue (Cabrera, March 2012).
Pathogen Occurrence in Samples Collected Before and During Mass Mortality Event
Remarkably, H. pinnae was not detected in any of the 167 fan mussels analysed before the MME, while it occurred in 63/99 (64%) of fan mussels examined by histology collected during MME (Table 2 and Figure 5A). Conversely, mycobacteria were detected both before and during MME (Table 2 and Figure 5A). Considering the histological diagnoses, differences in distribution into negative and positive cases between samples taken before and during MME were significant for H. pinnae and Gram (−) bacteria but not for mycobacteria. The ratios of individuals in the health condition classes were different between diagnostic procedures (Table 2), which contributed to differences in the percentages of positive cases between procedures.
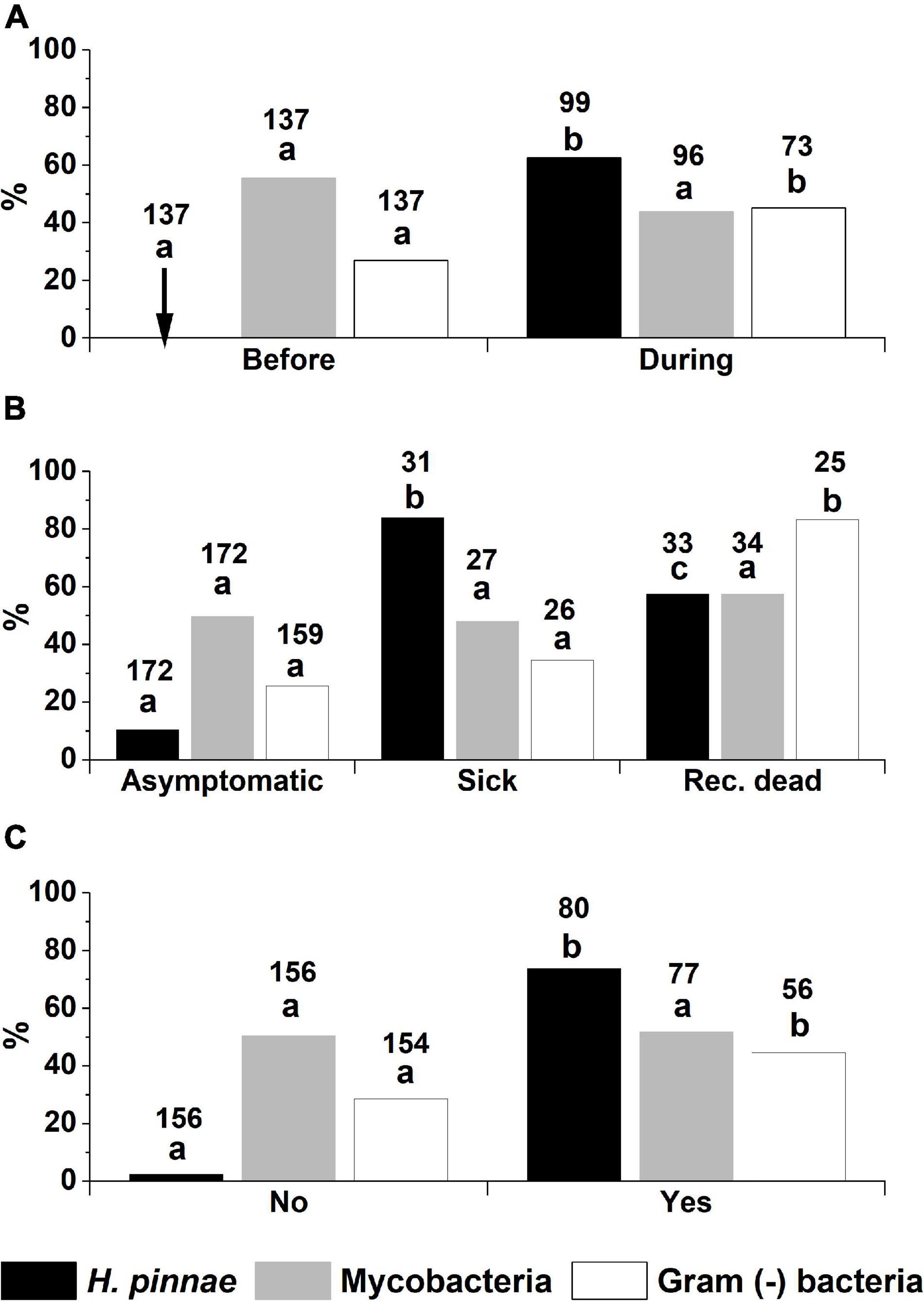
Figure 5. Percentages of infection with Haplosporidium pinnae, mycobacteria and Gram (−) bacteria, diagnosed with histology, in fan mussels classified according to (A) the period of sampling (either before or during the MME), (B) the fan mussel health condition (asymptomatic, sick or recently dead), and (C) the background of high mortality at sampling location (either yes or no). Different letters above bars denote significant differences between fan mussel classes for each pathogen. The arrow in the top graph points out 0% cases of infection with H. pinnae. The numbers above bars correspond to the number of fan mussels analysed in each case.
Association Between Pathogen Occurrence and Health Condition
The distribution into negative and positive histological detections of H. pinnae was significantly different between the three classes of health condition, with the lowest percentage of infection in the asymptomatic individuals and the highest in the sick ones (Figure 5B). No significant difference was found between histological mycobacteria detections of the three health conditions, while Gram (−) infections showed significant differences between the recently dead condition and the other ones (Figure 5B). Considering the conditions in which a putative aetiological agent of the MME should be more likely found, namely in sick individuals from high mortality affected areas, H. pinnae was detected in 26/31 (88%) of the sick fan mussels from MME-affected areas that were analysed with histology, mycobacteria in 13/27 (48%), and Gram (−) bacteria in 9/26 (35%) (Figure 5B). Regarding the 26 sick fan mussels from MME-affected areas that were found infected with H. pinnae by histology, 11/26 (42%) of them were simultaneously infected with mycobacteria and 5/26 (19%) of them were also infected with Gram (−) bacteria. During MME, Mycobacterium sp. but no Haplosporidium was detected in only four recently dead individuals.
Relationship Between Pathogen Occurrence and the Local Affection by Mass Mortality Event
As it has been explained before (see the section “Materials and Methods”), not all the locations were affected by a high mortality at the moment of sampling. The frequency of histological detection of H. pinnae was significantly higher in the locations that had been affected by high mortality than in the sites that had not been affected when sampling (Figure 5C). That was not the case for histological detection of mycobacteria, which showed similar distribution of positive and negative cases in the high mortality-affected and in the non-affected locations. Detection of Gram (−) bacteria was higher in high mortality-affected sites (Figure 5C).
Juvenile Condition and Occurrence of Pathogens
A total of 35 juvenile fan mussels were analysed, 29/35 of which were sick individuals taken from Calpe and 1/35 sick plus 5/35 asymptomatic specimens collected from Delta de l’Ebre-Alfacs. The 21/29 sick juveniles from Calpe were positive for H. pinnae by PCR (Table 1). However, every juvenile (29/29) from that location was PCR-negative for Mycobacterium sp. (Table 1), even though the use of the whole soft tissues would facilitate the detection of this bacterium by PCR. Regarding the six juveniles from Delta de l’Ebre-Alfacs, everyone was negative (6/6) for H. pinnae (histology and PCR) and for mycobacteria (histology) but 3/6 showed Gram (−) bacteria in histological sections.
Relationship of Pathogen Occurrence With Host Size
The percentage of fan mussels found infected with H. pinnae by histology was not associated with size (dorso-ventral length; Figure 6). However, a significant association was found between the percentage of fan mussels found infected with mycobacteria by histology and the host size; the same applied for infection with Gram (−) bacteria. The higher the size class, the higher the percentage of infection, both for mycobacteria and Gram (−) bacteria (Figure 6).
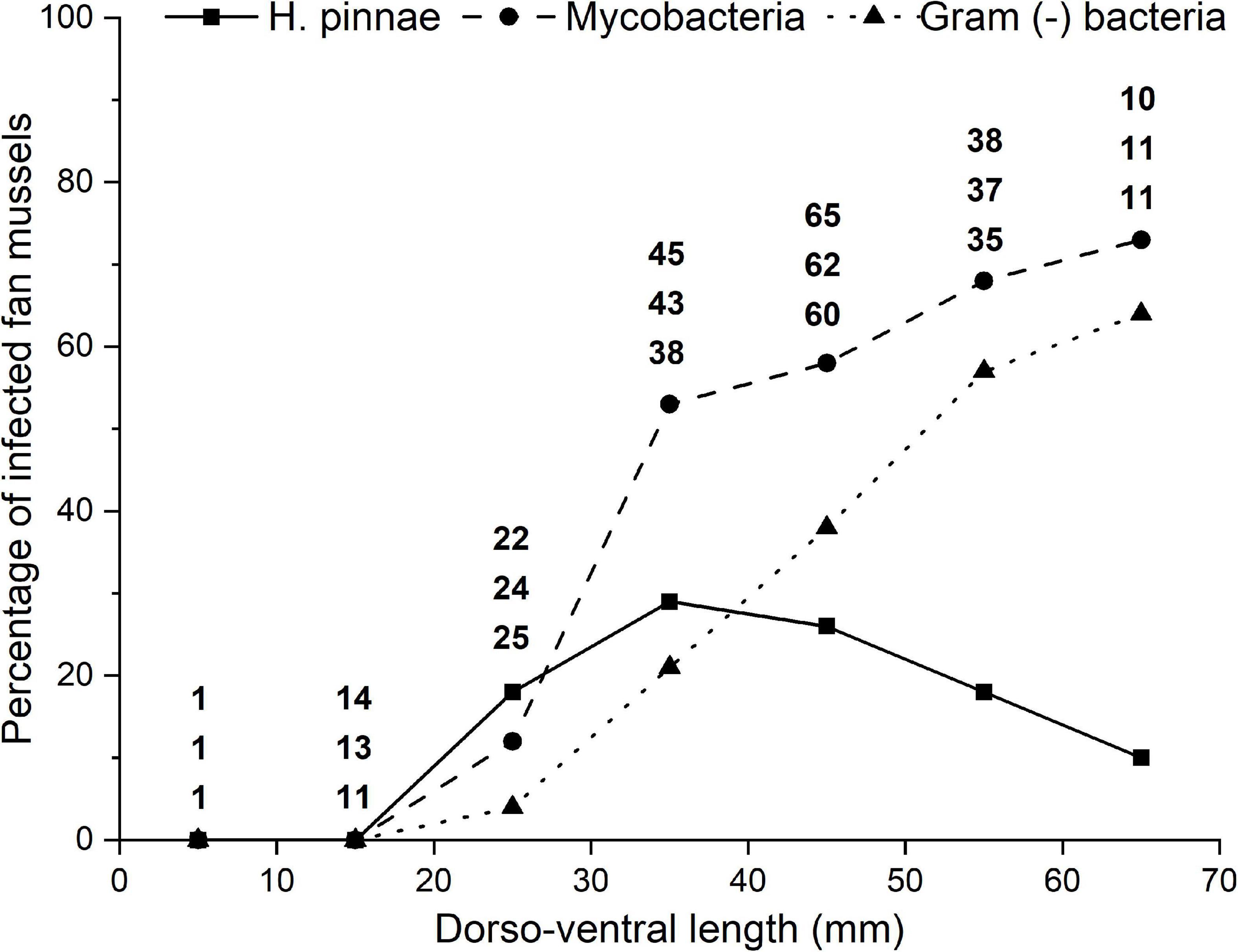
Figure 6. Variation of the percentage of fan mussels found infected by histology with size (dorso-ventral length) classes of the host. Infections with Haplosporidium pinnae, mycobacteria and Gram (−) bacteria are distinguished with different line patterns and symbols. The numbers above each size class correspond to the number of fan mussels of that class diagnosed for H. pinnae (top), mycobacteria (mid) and Gram (−) bacteria (bottom).
Association Between Pathogens
The paired tests to evaluate the association between pathogens, based on histological diagnosis, showed that the occurrence of H. pinnae was independent of that of mycobacteria and Gram (−) bacteria, while the occurrence of mycobacteria was associated with that of Gram (−) bacteria (Table 5).
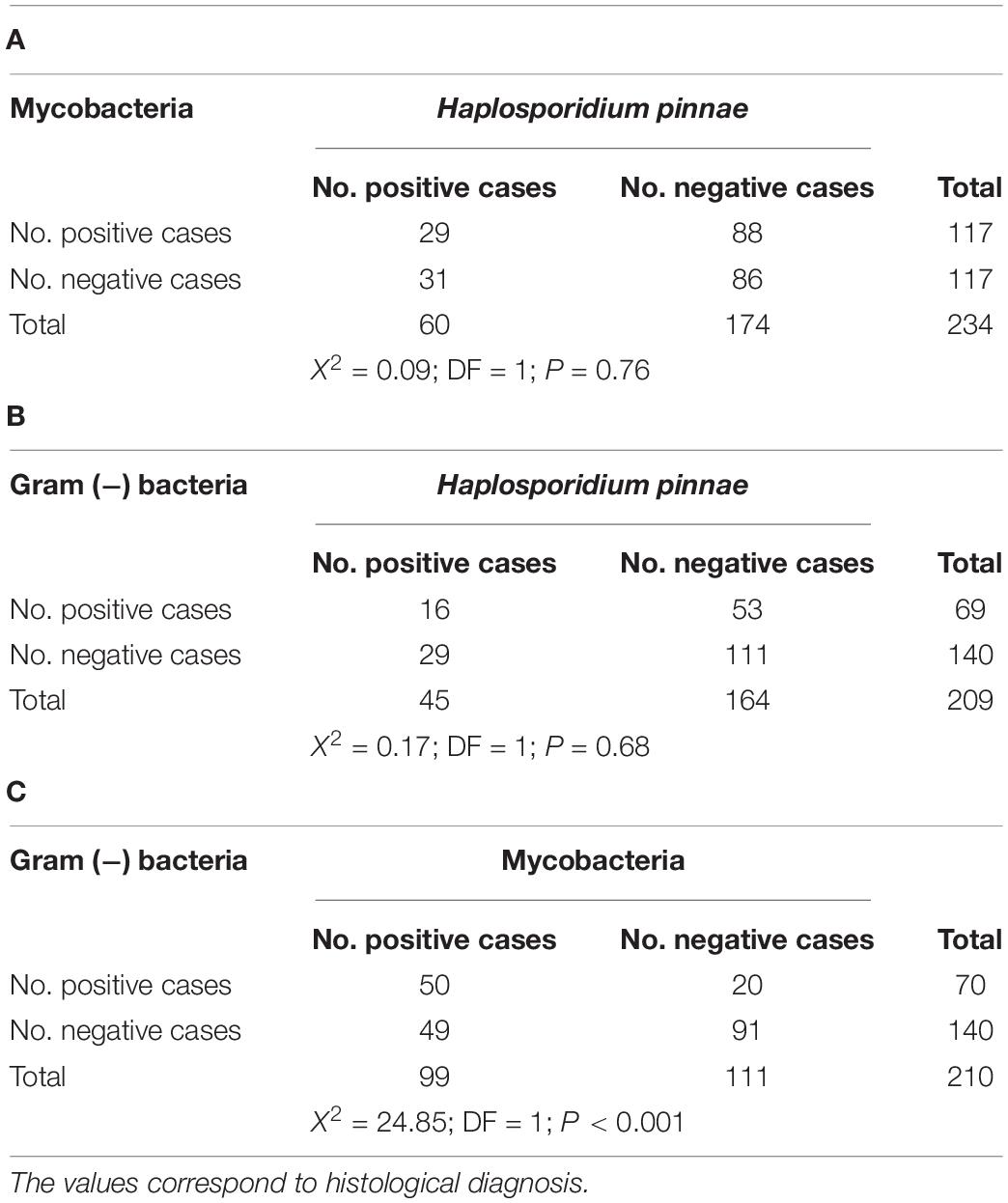
Table 5. Contingency tables and statistical test to evaluate the association between the occurrence of Haplosporidium pinnae and that of mycobacteria (A), H. pinnae and Gram (−) bacteria (B), and mycobacteria and Gram (−) bacteria (C).
Relationship Between the Occurrence of Haplosporidium pinnae and the Environment
The highest percentage of individuals infected with H. pinnae corresponded to those living in open sea, while the lowest percentage corresponded to those from coastal lagoons and those from harbours in intermediate position (Figure 7). The differences in the distribution of fan mussels into H. pinnae-infected and non-infected categories between environments were significant.
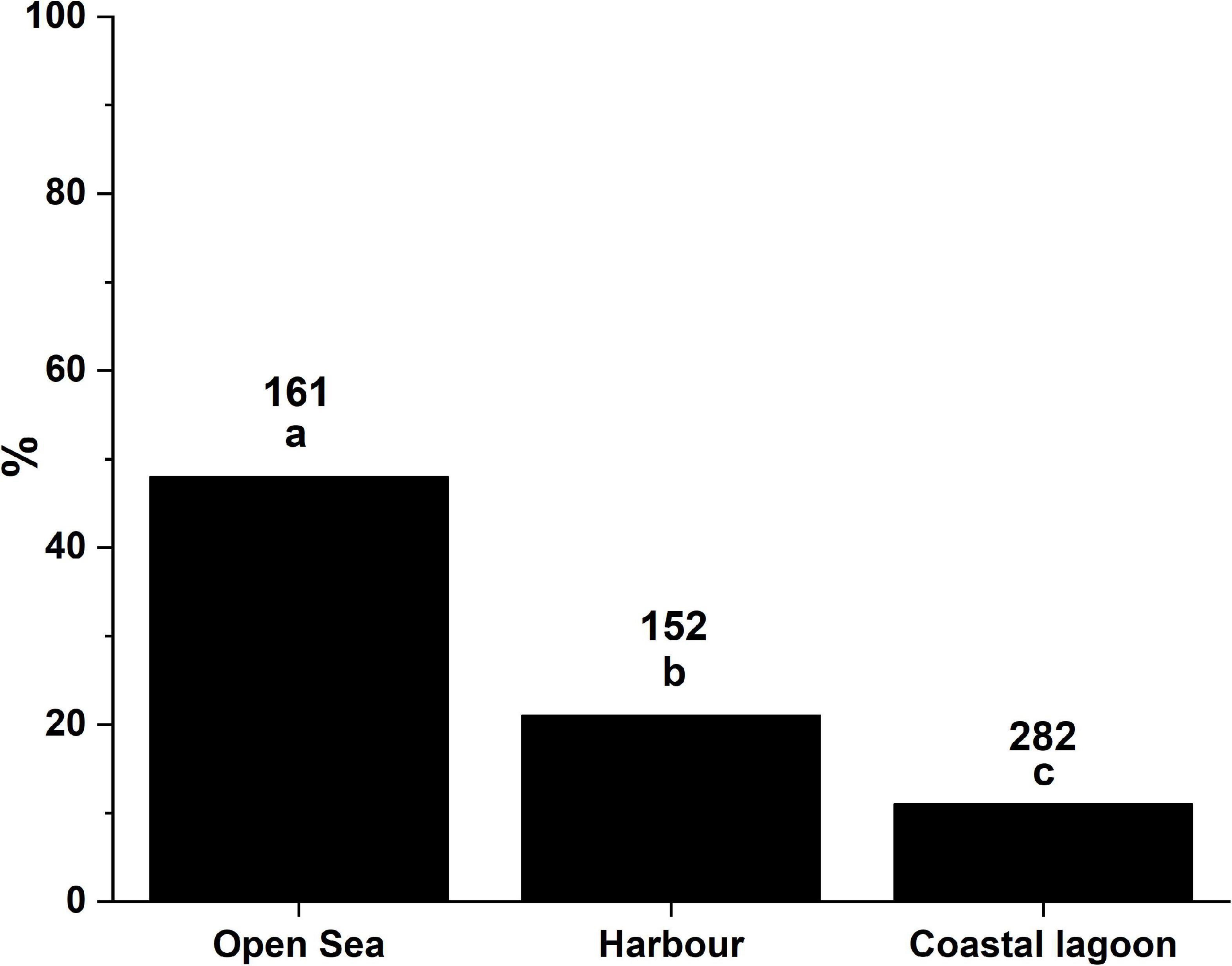
Figure 7. Percentages of infection with Haplosporidium pinnae corresponding to different ecosystems. Different letters above bars denote significant differences between environments. The numbers above bars correspond to the number of fan mussels diagnosed in each case.
Geographic Distribution of the Pathogens
Haplosporidium pinnae was detected in 24/30 locations while it was not in 6/30 sites (Table 1 and Figure 1). In 4/6 of the locations where H. pinnae was not detected, abnormally high mortality had not been recorded, while in the other 2/6, Port La Nouvelle and Telašćica, which had been affected by MME, the analysed fan mussels were asymptomatic. Regarding the 24 locations in which the parasite was detected, MME had affected 20/24 and the other 4/24 sites (Mar Menor, Agde, Sète, and Frontignan) that had not been affected by MME when sampling, were close to affected locations. The occurrence of mycobacteria was tested, either by histology or PCR, in 26 locations; positive cases were recorded in 16/26, 14 that had been affected by MME and 2 that had not (Table 1 and Figure 1). Regarding the 10 locations in which mycobacteria were not detected, diagnoses were based on mantle biopsies analysed with PCR (Table 1), a procedure likely yielding high proportion of false negatives, as mentioned above. Considering the 16 locations for which histological diagnosis for mycobacteria was performed, mycobacteria were detected in 14/16, all them except in Mar Menor (only one specimen), and Delta de l’Ebre Fangar (four individuals) (Table 1). Gram (−) bacteria were analysed in samples from 13 locations, 9/13 had been affected by MME and 4/13 had not; infections were found in 12/13 sites except in Sète (only three mantle biopsies) (Table 1).
Discussion
Samples of fan mussels taken through the Mediterranean coast by various teams of different countries were used to gather a wide-geographic, long term data set with which assess the role of previously blamed and suspicious pathogens in the panzootic MME. This approach allowed compiling highly diverse information, including samples taken before and during MME; asymptomatic, sick and recently dead animals; wide geographic and fan mussel size (from juvenile to old adults) ranges; and different environments (coastal lagoon, harbour, open sea). The involvement of various sampling/research teams and the different protection degree of P. nobilis, depending on country and sampling period (before and during mortality outbreaks), explained the heterogeneity of diagnostic techniques and sample types (either necropsy or biopsy), but that was compensated with the unprecedentedly high number of analysed fan mussels and highly diverse scenarios.
Concerning the health status of the analysed individuals, accurately describing what is considered as “recently dead” specimens in the wild is important in the context of the current controversy in the scientific community on the cause of the P. nobilis MME. In our study, the individuals considered “recently dead” were dead animals in which the decomposition, lytic post-mortem processes had begun although the presence of still consistent soft tissues allowed taking soft tissue pieces and their histological analysis. Previous reports on the mass mortality of P. nobilis did not explain the meaning of “recently dead” individuals. Remarkably, the detection of multiple pathogens only in individuals classified as “recently dead” may be due to the decomposition process, i.e., post-mortem proliferation of opportunistic or saprophagous organisms (particularly bacteria) in degraded specimens, which were in autolytic phase at sampling (Dent et al., 2004; Tsokos, 2005). If decomposition and tissue lysis is advanced, bacterial proliferation is obvious while the detection of other pathogens (such as H. pinnae) that occurred before the post-mortem lysis could be very difficult or impossible, causing interpretational problems (Tsokos, 2005). In this sense, in marine environments, as the putrefaction proceedes, Gram-negative, non-pigmented rods constitute a major component of the flora (Masami and Hiroshi, 1959). However, parasites do not survive in dead molluscs because tissue putrefaction probably leads to an acidic degradation of the cyst or oocyst wall (Gómez-Couso et al., 2003).
Regarding heterogeneity of diagnostic techniques and procedures, histology and PCR showed substantial agreement for detection of H. pinnae. However, the PCR procedure to detect Mycobacterium sp. seemed to yield a high number of false negatives, likely due to the solely use of mantle biopsies for PCR assays, because the mycobacteria infections were frequently restricted to the digestive gland and the surrounding connective tissue. According to this and considering that Gram (−) bacteria were only evaluated with histology (using Gram staining), all the analyses involving comparisons between different pathogens were based on histological diagnoses. Nevertheless, positive PCR results were useful at least to suggest (pending of confirmation) the occurrence of mycobacteria and to get the DNA sequence for its correct identification in various geographic locations for which PCR-based diagnoses were the only available diagnostic. The use of histological techniques in studies looking for aetiology is important to confirm true infections because just a positive PCR result does not involve true infection on its own (Burreson, 2008).
Histopathological features of the infections of fan mussels with H. pinnae and those with mycobacteria observed in this study are consistent with descriptions provided by Catanese et al. (2018) and Carella et al. (2019, 2020), respectively. The connective tissue of the visceral mass was the main location for mycobacteria; nevertheless, the detection of some positive PCR-results for Mycobacterium sp. from mantle biopsies as well as the reports of mycobacteria infections occurring in mantle (Carella et al., 2019, 2020) and gills (Lattos et al., 2020a) evince a broader histological range, especially in advanced infections in which mycobacteria occur free in the host tissues in addition to those sequestered in haemocytic nodules.
Although some authors suggest the multifactorial causality of the P. nobilis mortality, the results of this work supported that the onset of the MME is strongly associated with the detection of H. pinnae, which exhibits a preeminent role with respect to the others pathological agents considered in the study. Although it is true that the causal relationship with the disease can only be unequivocally established with controlled laboratory experiments, the strong relationship between the presence of H. pinnae with the MME is unquestionable, as shown in our results. This strong relationship, however, is not exhibited by other putative pathogenic agents identified as possible aetiological agents. It was the only analysed pathogen that was not detected in the period before MME was noticed, while it was widely detected during MME. Infection with H. pinnae was significantly associated with health condition, with significantly higher probability of finding the parasite in sick and dead animals than in asymptomatic ones; the lower histological detection of H. pinnae in recently dead individuals than in sick ones could be due to the difficulty of detection of the parasite in post-mortem delayed tissues (Gómez-Couso et al., 2003; Alaeddini et al., 2010; Allen-Hall and McNevin, 2013). The frequency of histological detection of H. pinnae was significantly higher in areas that had been affected by high mortality. Assuming that the most likely conditions to detect a putative aetiological agent of the MME would be in sick fan mussels from affected areas, H. pinnae complied with that contingency and did it more frequently without mycobacteria or Gram (−) bacteria co-infection. Considering that the MME has affected fan mussels of all ages (Vázquez-Luis et al., 2017; Zotou et al., 2020), from juveniles to old adults, the detection of H. pinnae in many juveniles, more frequently in the sick ones, from Calpe, an area heavily affected by high mortality early in the MME, together with the lack of association between the occurrence of this parasite and the host size (age), contributes to support the strong association between H. pinnae and the MME. The lack of detection of this parasite in the six juvenile fan mussels from Delta de l’Ebre-Alfacs could be due to the fact that three of them were taken before abnormal high mortality got the area and that the other three individuals, taken once high mortality had been observed, were asymptomatic. Finally, this parasite was not detected in any specimen of P. rudis, which has not been affected by MME even in the areas where P. nobilis became extinct.
On the contrary, evidence supporting association of MME with mycobacteria was not found. Infection with mycobacteria was present with a high prevalence, in the healthy, and reproductively capable fan mussel population of the Balearic Islands, regularly monitored before the onset of the MMEs (Hendriks et al., 2012; Vázquez-Luis et al., 2014; Alomar et al., 2015; Deudero et al., 2015, 2017; Box et al., 2020). These mycobacteria were identified as Mycobacterium sp., very closely related (with 100% sequence similarity) to those detected by Carella et al. (2019) during MME (Box et al., 2020). The percentage of individuals infected with mycobacteria in the period before detection of MME was not significantly different from that during the MME spreading. The probability of finding infection with mycobacteria did not significantly vary depending on health condition. The percentage of infection with mycobacteria in the high mortality-affected areas was not significantly different from that of the non-affected sites. Mycobacteria were not detected in half of the sick individuals from high mortality-affected areas. The lack of detection of mycobacteria in juveniles and the significant association of the occurrence of mycobacteria with host size (age) would support a dominant chronic condition of the infection with mycobacteria rather than acute infection leading to host death. Histological observation revealed mycobacterial chronic nodular lesions, resembling those described by Carella et al. (2019). The digestive route of infection could be the most plausible, due to the localisation of lesions at the stomach, digestive gland and surrounding area. These observations support the hypothesis of the opportunistic character of mycobacteria, acting as a fan mussel coloniser, being sequestered in nodules, close to the “entrance door.” Moreover, mycobacteria were also detected in an asymptomatic, P. rudis, suggesting an opportunistic character in the Pinnidae family. Mycobacteria have been mainly found in marine bivalves when asymptomatic populations are studied focussing on their role as carriers involved in human disease (Aubry et al., 2002; Kotlowski et al., 2004; Hashish et al., 2018). Interestingly, mycobacteria were detected during a study of microbiome assessment of the predominant bacterial populations in asymptomatic Pinna nobilis individuals from a healthy population of the Eastern Adriatic Sea before MME arrival (Pavlinec et al., 2020). The detection of Mycobacteria in a few recently dead individuals, without detecting H. Pinnae, could be explained by their greater resistance capacity to environmental factors, withstanding the autolysis process and inducing their minor cellular fragmentation in the sample (Lindahl, 1993).
Gram (−) bacteria neither appeared associated with MME. They were detected both before the first detection of MME and during the period of MMEs. The higher probability of finding them in recently dead individuals than in sick and asymptomatic fan mussels suggests their post-mortem proliferation rather than being the cause of death. If so, the higher percentage of infection with Gram (−) bacteria in areas with high mortality background than in MME non-affected areas would be due to the fact that the proportion of recently dead individuals collected from the former areas was higher than that from the latter sites.
Gram (−) bacteria were only detected in one third of the sick individuals from MME-affected areas. As in the case of mycobacteria, the fact that the prevalence of infection with Gram (−) bacteria increased with host size (age) while MME equally affected fan mussels of all sizes (ages), debilitates a potential role of Gram (−) bacteria in fan mussel MME. These bacteria were also detected in a sick P. rudis. The opportunistic condition of a Gram (−) bacterium, V. mediterranei, was suggested to explain high mortality in stabled fan mussels; the bacteria would have taken advantage of the host stress due to captivity and other possible factors to proliferate through host tissues (Prado et al., 2020a).
The occurrence of H. pinnae in the tissues of fan mussels was independent of those of mycobacteria and Gram (−) bacteria, while these two types of bacteria were significantly associated, which could be consistent with an opportunistic character of both types of bacterial infections. Accordingly, the pathophysiological response in sick animals is much greater in co-infections with H. pinnae in comparison with co-infections without presence of haplosporidan parasite (Lattos et al., 2021).
No case of infection with protistans Perkinsus spp. was detected with histology. Cases of infection with these parasites were only detected with the RFTM procedure, which was used in before the first evidence of MME. Diagnosis of Perkinsus spp. with RFTM is more sensitive than with histology (Bushek et al., 1994; Rodríguez and Navas, 1995; Villalba et al., 2005). Therefore, cases of light perkinsosis could have been unnoticed with histology during the MME spreading but, if systemic (moderate and heavy) infections had occurred, they would have been detected even with histology. The occurrence of cases of perkinsosis before the MME, as light infections, and the lack of detection of infections with Perkinsus sp. by histology through the study showed that perkinsosis is not an emerging disease for P. nobilis and it did not reach moderate or heavy intensity in P. nobilis, which allowed discarding association of perkinsosis with MME. No other serious pathogens with potential to cause high mortality were detected in the study.
The aetiology of the fan mussel MME was addressed previously. Since H. pinnae was proposed as its most likely cause (Vázquez-Luis et al., 2017; Catanese et al., 2018), subsequent surveys performed in areas affected by high mortality of fan mussels emphasised the search for this parasite and, as it was found, it was blamed for the high mortality in the respective areas (Fanelli et al., 2018; Katsanevakis et al., 2019; Panarese et al., 2019). The detection of more cases of infection with Mycobacterium sp., associated with heavier histopathological damage, than with H. pinnae in sick fan mussels from Campania and Sicily (South Italy) led to Carella et al. (2019) to state that Mycobacterium sp. was associated with the fan mussel high mortality in those areas but H. pinnae was not. However, Carella et al. (2020), in a broader (involving more fan mussels with wider geographic range) study, proposed that Mycobacterium sp., Vibrio spp., H. pinnae and Perkinsus sp. cooperate to disease pathogenesis, among which Mycobacterium sp. and H. pinnae seemed to be the most relevant. Subsequent surveys in other Mediterranean regions highlighted the occurrence of both Mycobacterium sp. and H. pinnae in fan mussels from MME-affected areas (Lattos et al., 2020a; Scarpa et al., 2020; Čižmek et al., 2020; Šarić et al., 2020).
The lack of detection of Mycobacterium sp. and H. pinnae in some sick fan mussels from affected areas was highlighted to argue that other factor/s could be the cause of (or contribute to) the MME (Scarpa et al., 2020; Šarić et al., 2020). The critically endangered condition of P. nobilis implies exceptional difficulties to perform ad hoc experiments addressing the aetiology of the MME. Previous studies on the aetiology of the fan mussel MME were based on scarce number of specimens, from 4 (Darriba, 2017) to 46 (Scarpa et al., 2020). Our study has been the broadest attempt, in terms of time period, geographic range and sample size, addressing the aetiology of the MME; the results supported that H. pinnae was the pathological agent of those considered in the study with a preeminent role in the onset of the MMEs; even the argument used in other studies to weaken or discard the consideration of H. pinnae as the aetiological agent of the MME, namely its absence in sick fan mussels from MME-affected areas, would not apply to our results because 26 out of the 27 sick individuals collected from affected areas that were diagnosed with histology showed infection with H. pinnae. Nevertheless, assuming the strong association between H. pinnae and the MME found in this study, it cannot be considered as a conclusive evidence of a cause-effect relationship.
Scarpa et al. (2020) stated that the Mediterranean mussel Mytilus galloprovincialis and the carpet shell clam Ruditapes decussatus are also hosts of H. pinnae, and thus P. nobilis is not the exclusive bivalve host of this parasite, all it based on PCR-positive diagnosis for H. pinnae obtained from tissues of three clams and one mussel. However, only PCR amplification cannot be considered as a valid evidence of a true infection and, thus, of a parasite-host relationship; further validation is needed with an established technique, typically histology or in situ hybridisation, as thoroughly explained by Burreson (2008). The fact that the three clam samples that gave PCR-positive results for H. pinnae had been collected in 2014 would involve that the parasite occurred at that time in The Mediterranean Sea (at least in the eastern coast of Sardinia). If H. pinnae really occurred at that time, did it infect its currently well-known host P. nobilis long time before the first evidence of MME, in 2016? According to our results, H. pinnae was not detected in samples of 167 fan mussels collected in the period 2011–2014 from various locations in the Balearic Islands while it was widely detected since 2016 onward in the same locations, where the fan mussel is nowadays almost extinct. Assessing whether H. pinnae occurred or not in fan mussels before 2016 in other areas would be useful if recorded samples were available.
Considering that all the known proliferation patterns of haplosporidans have been found in H. pinnae (see information below), likely this species can be directly transmitted among fan mussels and endure free in the environment keeping infective potential, all that contributing to facilitate proliferation through fan mussel populations. According to our results, stages of the H. pinnae sporulation process were only detected in 18% of the infected fan mussels. The stages of the sporulation process (multinucleate plasmodia, sporocysts and spores) are more easily detected than the smaller uni- and binucleate stages during histological examination, thus special care is needed to detect the parasite when sporulation does not occur. The detection of uni- and binucleate stages is particularly difficult in decomposed tissues of dead fan mussels.
Results showed a wide geographic distribution of H. pinnae. The earliest H. pinnae detections corresponded to SE Spain (El Calón and Calpe) and Majorca Island (autumn 2016), being found later (2017) northward in Minorca island, NE Spain (Tossa de Mar and Portlligat) and eastward in Corsica, coinciding with the transmission models of a waterborne parasite dictated by surface currents (Cabanellas-Reboredo et al., 2019), which completely fits with the reduced SIR model validated with available data of Haplosporidium pinnae transmission in controlled water tanks (Giménez-Romero et al., 2021). The parasite was detected through 2018 and 2019 in more eastern Mediterranean locations as sampling effort coincided with MME spread. Both mycobacteria and Gram (−) bacteria were also widespread in P. nobilis populations but, very likely, the lack of their detection in some areas would be due to the lack of using histological diagnostic techniques with ZN and Gram staining, respectively.
Remarkably, differences in the prevalence of H. pinnae between fan mussels living in open sea, in harbours and in coastal lagoons were significant. Accurately determining whether particular environmental conditions make difficult or impede parasite proliferation could be relevant to recovery purposes, especially if some coastal lagoons remain as sanctuaries of P. nobilis. In fact, environmental factors (water temperature, salinity, presence of pollutants, trophic conditions) have significant effects on the pathogens themselves, on the host and on the host-pathogen relationship (Barbosa Solomieu et al., 2015). Notably, the dynamics of haplosporidan parasites is modulated by environmental parameters, especially temperature and salinity (Burreson and Ford, 2004; Arzul and Carnegie, 2015). Specific tolerance of H. pinnae to temperature and salinity has been suggested (temperatures above 13.5°C and salinity range between 36.5–39.7 psu; Cabanellas-Reboredo et al., 2019; García-March et al., 2020; Prado et al., 2021; Nebot-Colomer et al., 2021). Accordingly, salinity range outside the common Mediterranean salinity range (36.7–39.5 psu; Vidal-Vijande et al., 2011) is a shared feature of these coastal lagoons: on the one hand, the Mar Menor, exhibiting high salinity, up to 47 psu (Pérez-Ruzafa et al., 2005; Giménez-Casalduero et al., 2020); on the other hand, the other Spanish and French coastal lagoons, strongly affected by freshwater supplies (Prado et al., 2014, 2020b). Recent data demonstrated that until now the lagoons of the Gulf of Lion are still free from parasite (almost 90% of P. nobilis population; Peyran et al., 2021). Interestingly, mortality of fan mussels has been shown to be associated with salinity gradient in one of the coastal lagoons involved in the study, Delta de l’Ebre-Alfacs, influenced by freshwater discharges; the lower the salinity the lower the cumulative mortality (Prado et al., 2021). However, many coastal lagoons (as well as harbours) share higher anthropogenic pressures implying higher stress for fan mussels living there (García-March et al., 2020; Giménez-Casalduero et al., 2020; Prado et al., 2021), which could influence in their redox balance, cause cellular damage, and depress their immune system (Rivera-Ingraham et al., 2013; Capó et al., 2015; Espinosa and Rivera-Ingraham, 2016). In this sense, before the MME (deeply associated with H. pinnae according to our results), anthropogenic stressors rather than environmental or other variables explained most of the variability in the spatial distribution of the density of P. nobilis in the Balearic Islands (Deudero et al., 2015). Indeed, environmental adverse conditions such as pollutants or infections can induce oxidative stress in marine bivalves. However, when analysing the differential effects of H. pinnae and Mycobacterium sp. on P. nobilis antioxidant response, significant effects on biomarkers were only observed in the presence of H. pinnae (Box et al., 2020). Unfortunately, we do not have data in P. nobilis to relate the effect of anthropic pressure on biomarkers before the presence of the parasite, which prevents us from elucidating whether the anthropic pressure may also have an adverse effect on P. nobilis inhabiting coastal lagoons, the last relicts for the survival of the species.
Special attention deserves the known stages of the H. pinnae life cycle, which have implications in the potential for disease spreading and parasite permanence in the ecosystem. The life cycle of haplosporidan parasites is a complex issue. Binary fission is the only known proliferation process (red dotted frame in Figure 8) of some haplosporidan species infecting oysters, such as Bonamia ostreae and Bonamia exitiosa, in which uninucleate and binucleate cells can leave the host with potential to infect new hosts, thus making possible direct parasite transmission among individuals of the type host species (Sas et al., 2020); additionally other potential transmission vehicles or reservoirs have been identified, including oyster larvae (Arzul et al., 2011) and other invertebrate species (Lynch et al., 2007, 2010; Costello et al., 2020). In other haplosporidan species, such as Haplosporidium nelsoni, the only known stages are plasmodia and those of sporogonia (area enclosed with blue dashed outline in Figure 8); for these haplosporidan species, their transmission way is unknown; direct transmission has never been experimentally confirmed and the need of a host has been suggested (Burreson and Ford, 2004; Arzul and Carnegie, 2015); the spores are likely enduring stages whose infective potential last longer than that of naked uninucleate and binucleate stages. Among haplosporidans infecting molluscs, the species in which the occurrence of all the mentioned proliferation patterns have been demonstrated through the host are scarce; that is the case of Bonamia perspora (Carnegie et al., 2006) and H. pinnae (Catanese et al., 2018), whose sporulation processes take place in different tissues: connective tissue and epithelium of digestive gland tubules, respectively.
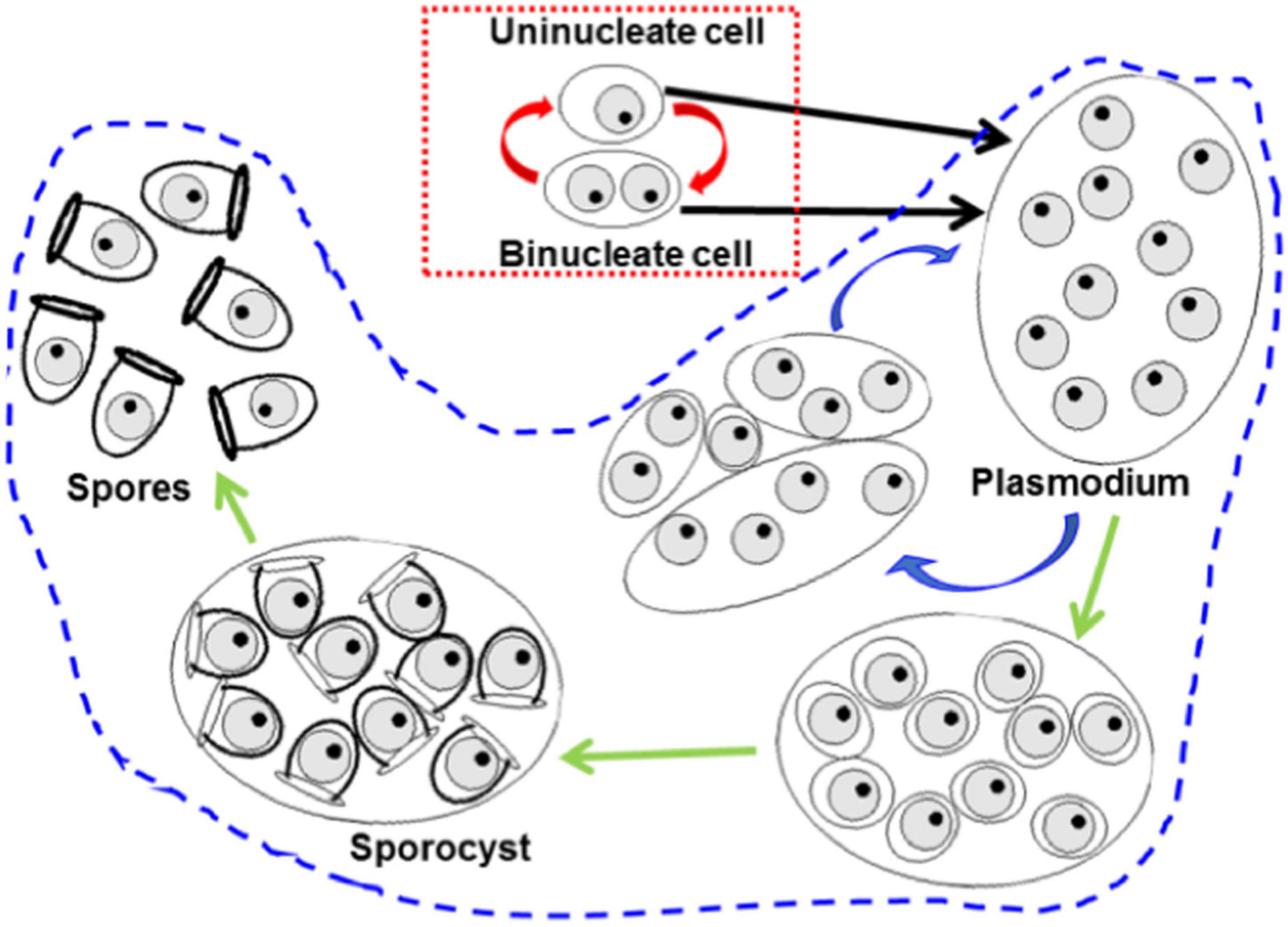
Figure 8. Scheme of the known proliferation patterns trough host tissues of haplosporidan species. The red dotted frame encloses binary fission proliferation, based on karyokinesis in uninucleate cells giving rise to binucleate cells, followed by cytokinesis (red arrows). The area enclosed with blue-dotted outline includes formation of multinucleate plasmodia through karyokinesis without cytokinesis; eventually the plasmodia fragment by plasmotomy and the daughter cells repeat the process again and again (blue arrows); under unknown stimuli, plasmodia enter in sporogony process (green arrows) yielding uninucleate spores.
Knowing the aetiology of the fan mussel MME is essential to avoid the complete extinction of this species and to design measures to recover their populations. The results supported a strong association of H. pinnae with the MME and allowed discarding an essential role of mycobacteria and Gram (−) bacteria.
Permissions
Samplings were performed with the necessary permits of the competent authorities (national and/or local). The corresponding permissions were detailed below: GOIB-ENB-AC-01/31/2011, SPE-CMAP-RE127 (7/11/2011), CE-5614-05-24-2013, CMAP-DGENB-40/2016, GC-DTS-SF/1088-2017, GC-DTS-02-11-2017, GC-DTS-17-11-2017, DIV/BDM/AUSPP/45/2017, DIV/BDM/AUSPP/47/2017, GMEE175828/2195 (11/14/2018), KLASA:UP/I-612-07/18-48/134, URBROJ: 517-05-1-1-18-4, prefectural decree 2018-s-24, prefectural decree 2018-s-28, SGPM/BDM/AUTSPP/57/2018, PTOP-2019-404, GC-8428-05-23-2019, MAF-FA67852565-140.03.03-E.2935960 (09/26/2019).
Data Availability Statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article.
Author Contributions
AG and GC conceived the study. AG, GC, BH, JV, PK, CP, and GS analysed the samples. AG, GC, AV, and JN analysed the data. AG, AV, and GC wrote the manuscript with input from all authors and revised the final manuscript. The rest of the authors participated in the field work. All the authors reviewed the manuscript improving it substantially. All documents were edited and approved by all authors.
Funding
This work was funded by: DG Pesca i Medi Mari (GOIB), EsMarEs (order IEO by MITECO, Spanish Government), Life UFE IP-PAF INTEMARES (LIFE15 IPE ES 012) “Gestión integrada, innovadora y participativa de la Red Natura 2000 en el medio marino español,” the research project “Estado de conservación del bivalvo amenazado Pinna nobilis en el PNAC” (OAPN 024/2010), the project RECONNECT (MIS 5017160) of the Programme Interreg V-B “Balkan-Mediterranean 2014–2020.” MTES (French Government), DREAL (Direction Régionale Environnement Aménagement Logement) and Région Occitanie (France) for funding research and monitoring of Pinna. GC and PP were contracted under the INIA-CCAA cooperative research programme for postdoctoral incorporation from the Spanish National Institute for Agricultural and Food Research and Technology (INIA) (DOC INIA 8/2013 and 15/2015). MV-L was supported by a Juan de la Cierva-Incorporación postdoctoral contract (ICJI-2016-29329, MICIU Programme). ML-S and EÁ were supported by a Personal Técnico de Apoyo contract MINECO programme (PTA2015-11709-I and PTA2015-10829-I, respectively). CP and GS were supported by the project RECONNECT (MIS 5017160) financed by the Transnational Cooperation Programme Interreg V-B “Balkan-Mediterranean 2014–2020” and co-funded by the European Union and national funds of the participating countries. CP was supported by Sorbonne University.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
The authors thank all the members of the ‘scuba diver teams’ for their help in sampling, namely F. Giménez, A. Izquierdo, F. Garfella, E. Massutí, A. Barrajón, D. Moreno, S. Jiménez, J.L. Crespo, M. Guimerans, and E. Cortés (Spain), C. Steiner (France), G. Ceccherelli, and M.F. Cinti (Italy), B. Colic (Croatia), L. Hadjioannou (Cyprus), M. Salomidi, V. Gerakaris, G. Chatzigeorgiou, P. Grigoriou, E. Chatzinikolaou, Hippocampus Bali dive center (Greece). J. Barja provides the samples from Almería. D. Karagiannis of NRL for Mollusc Diseases (Greek Government) provided the samples from Lesvos. M. Moreno de Pintos (Spanish Ministry of Environment) and M. Otero (IUCN) provided logistical support. A. Simal provided technical assistance in histological processing. R. Graèan carried out histological analysis of Telascica samples.
Footnotes
References
Acarli, S., Acarli, D., and Kale, S. (2021). Current Status of Critically Endangered Fan Mussel Pinna nobilis (Linnaeus 1758) Population in Çanakkale Strait, Turkey. Mar. Sci. Tech. Bull. 10, 62–70. doi: 10.33714/masteb.793885
Alaeddini, R., Walsh, S. J., and Abbas, A. (2010). Forensic implications of genetic analyses from degraded DNA—a review. Forensic Sci. Int. Genet. 4, 148–157. doi: 10.1016/j.fsigen.2009.09.007
Allen-Hall, A., and McNevin, D. (2013). Non-cryogenic forensic tissue preservation in the field: a review. Aust. J. Forensic Sci. 45, 450–460.
Alomar, C., Vázquez-Luis, M., Magraner, K., Lozano, L., and Deudero, S. (2015). Evaluating stable isotopic signals at bivalve Pinna nobilis under different human pressures. J. Exp. Mar. Biol. Ecol. 467, 77–86. doi: 10.1016/j.jembe.2015.03.006
Andree, K. B., Carrasco, N., Carella, F., Furones, D., and Prado, P. (2021). Vibrio mediterranei, a Potential Emerging Pathogen of Marine Fauna: Investigation of pathogenicity using a bacterial challenge in Pinna nobilis and development of a species-specific PCR. J. Appl. Microbiol. 2021, 1–15. doi: 10.1111/jam.14756
Andrews, J. D., and Hewatt, W. G. (1957). Oyster mortality studies in Virginia. II. The fungus disease caused by Dermocystidium marinum in oysters of Chesapeake Bay. Ecol. Monogr. 27, 1–26.
Arzul, I., and Carnegie, R. B. (2015). New perspective on the haplosporidian parasites of molluscs. J. Invertebr. Pathol. 131, 32–42. doi: 10.1016/j.jip.2015.07
Arzul, I., Langlade, A., Chollet, B., Robert, M., Ferrand, S., Omnes, E., et al. (2011). Can the protozoan parasite Bonamia ostreae infect larvae of flat oysters Ostrea edulis? Vet. Parasitol. 179, 69–76. doi: 10.1016/j.vetpar.2011.01.060
Aubry, A., Chosidow, O., Caumes, E., Robert, J., and Cambau, E. (2002). Sixty-three cases of Mycobacterium marinum infection: clinical features, treatment, and antibiotic susceptibility of causative isolates. Arch. Intern. Med. 162, 1746–1752. doi: 10.1001/archinte.162.15.1746
Barbosa Solomieu, V., Renault, T., and Travers, M. A. (2015). Mass mortality in bivalves and the intricate case of the Pacific oyster, Crassostrea gigas. J. Invertebr. Pathol. 131, 2–10. doi: 10.1016/j.jip.2015.07.011
Basso, L., Vázquez-Luis, M., García-March, J. R., Deudero, S., Alvarez, E., Vicente, N., et al. (2015). The Pen Shell, Pinna nobilis: A Review of Population Status and Recommended Research Priorities in the Mediterranean Sea. Adv. Mar. Biol. 71, 109–160. doi: 10.1016/bs.amb.2015.06.002
Betti, F., Venturini, S., Merotto, L., Cappanera, V., Ferrando, S., Aicardi, S., et al. (2021). Population trends of the fan mussel Pinna nobilis from Portofino MPA (Ligurian Sea, Western Mediterranean Sea) before and after a mass mortality event and a catastrophic storm. Eur. Zool. J. 88, 18–25. doi: 10.1080/24750263.2020.1850891
Böddinghaus, B., Rogall, T., Flohr, T., Blöcker, H., and Böttger, E. C. (1990). Detection and identification of mycobacteria by amplification of rRNA. J. Clin. Microbiol. 28, 1751–1759. doi: 10.1128/JCM.28.8.1751-1759.1990
Box, A., Capó, X., Tejada, S., Catanese, G., Grau, A., Deudero, S., et al. (2020). Reduced Antioxidant Response of the Fan Mussel Pinna nobilis Related to the Presence of Haplosporidium pinnae. Pathogens 9:932. doi: 10.3390/pathogens9110932
Burreson, E. M. (2008). Misuse of PCR assay for diagnosis of mollusc protistan infections. Dis. Aquat. Org. 80, 81–83. doi: 10.3354/dao01925
Burreson, E. M., and Ford, S. E. (2004). A review of recent information on the Haplosporidia, with special reference to Haplosporidium nelsoni (MSX disease). Aquat. Living. Resour. 17, 499–517. doi: 10.1051/alr:2004056
Bushek, D., Ford, S. E., and Allen, S. K. (1994). Evaluation of methods using Ray‘s fluid thioglycollate medium for diagnosis of Perkinsus marinus infection in the eastern oyster, Crassostrea virginica. Ann. Rev. Fish. Dis. 4, 201–217. doi: 10.1016/0959-8030(94)90029-9
Cabanellas-Reboredo, M., Vázquez-Luis, M., Mourre, B., Alvarez, E., Deudero, S., Amores, A., et al. (2019). Tracking the dispersion of a pathogen causing mass mortality in the pen shell Pinna nobilis: a collaborative effort of scientists and citizens. Sci. Rep. 9:13355. doi: 10.1038/s41598-019-49808-4
Capó, X., Tejada, S., Box, A., Deudero, S., and Sureda, A. (2015). Oxidative status assessment of the endemic bivalve Pinna nobilis affected by the oil spill from the sinking of the Don Pedro. Mar. Environ. Res. 110, 19–24. doi: 10.1016/j.marenvres.2015.07.013
Carella, F., Aceto, S., Pollaro, F., Miccio, A., Iaria, C., Carrasco, N., et al. (2019). A mycobacterial disease is associated with the silent mass mortality of the pen shell Pinna nobilis along the Tyrrhenian coastline of Italy. Sci. Rep. 9, 1–12. doi: 10.1038/s41598-018-37217-y
Carella, F., Antuofermo, E., Farina, S., Salati, F., Mandas, D., Prado, P., et al. (2020). In the wake of the ongoing Mass Mortality Events: co-occurrence of Mycobacterium, Haplosporidium and other pathogens in Pinna nobilis collected in Italy and Spain (Mediterranean Sea). Front. Mar. Sci. 7:48. doi: 10.3389/fmars.2020.00048
Carnegie, R. B., Burreson, E. M., Hine, P. M., Stokes, N. A., Audemard, C., Bishop, M. J., et al. (2006). Bonamia perspora n. sp. (Haplosporidia), a parasite of the oyster Ostreola equestris, is the first Bonamia species known to produce spores. J. Euk. Mic. 53, 232–245. doi: 10.1111/j.1550-7408.2006.00100.x
Catanese, G., Grau, A., Valencia, J. M., Garcia-March, J. R., Vázquez-Luis, M., Alvarez, E., et al. (2018). Haplosporidium pinnae sp. nov., a haplosporidan parasite associated with mass mortalities of the fan mussel. Pinna nobilis in the Western Mediterranean Sea. J. Invertebr. Pathol. 157, 9–24. doi: 10.1016/j.jip.2018.07.006
Čelebičić, M., Gradaščević, N., and Viteškić, V. (2020). Is Pinna nobilis l. critically endangered in Neum Bay, Bosnia and Herzegovina? Veterinaria 69, 221–227.
Çinar, M. E., Bilecenoğlu, M., Yokeş, M. B., and Güçlüsoy, H. (2021). The last fortress fell: mass mortality of Pinna nobilis in the Sea of Marmara. Medit. Mar. Sci. 22, 669–676. doi: 10.12681/mms.27137
Čižmek, H., Čolić, B., Gračan, R., Grau, A., and Catanese, G. (2020). Emergency situation for pen shells in the Mediterranean: the Adriatic Sea, one of the last Pinna nobilis shelter, is now affected by the Mass Mortality Event. J. Invertebr. Pathol. 173:107388. doi: 10.1016/j.jip.2020.107388
Cohen, J. (1960). A coefficient of agreement for nominal scales. Educ. Psychol. Meas. 20, 37–46. doi: 10.1177/001316446002000104
Costello, K. E., Lynch, S. A., McAllen, R., O’Riordan, R. M., and Culloty, S. C. (2020). The role of invasive tunicates as reservoirs of molluscan pathogens. Biol. Invasions. 23, 641–655. doi: 10.1007/s10530-020-02392-5
Darriba, S. (2017). First haplosporidan parasite reported infecting a member of the Superfamily Pinnoidea (Pinna nobilis) during a mortality event in Alicante (Spain, Western Mediterranean). J. Invertebr. Pathol. 148, 14–19. doi: 10.1016/j.jip.2017.05.006
De Lorgeril, J., Lucasson, A., Petton, B., Toulza, E., Montagnani, C., Clerissi, C., et al. (2018). Immune-suppression by OsHV-1 viral infection causes fatal bacteraemia in Pacific oysters. Nat. Commun. 9:4215. doi: 10.1038/s41467-018-06659-3
Dent, B. B., Forbes, S. L., and Stuart, B. H. (2004). Review of human decomposition processes in soil. Env. Geol. 45, 576–585. doi: 10.1007/s00254-003-0913-z
Deudero, S., Grau, A., Vázquez-Luis, M., Álvarez, E., Alomar, C., and Hendriks, I. (2017). Reproductive investment of the pen shell Pinna nobilis Linnaeus, 1758 in Cabrera National Park (Spain). Med. Mar. Sci. 18, 271–284. doi: 10.12681/mms.1645
Deudero, S., Vázquez-Luis, M., and Álvarez, E. (2015). Human Stressors Are Driving Coastal Benthic Long-Lived Sessile Fan Mussel Pinna nobilis Population Structure More than Environmental Stressors. PLoS One 10:e0134530. doi: 10.1371/journal.pone.0134530
Espinosa, F., and Rivera-Ingraham, G. A. (2016). Subcellular evidences of redox imbalance in well-established populations of an endangered limpet. Reasons for alarm? Mar. Poll. Bull. 109, 72–80. doi: 10.1016/j.marpolbul.2016.06.019
Fanelli, G., Rubino, F., Paoletti, B., Della Salda, L., Mosca, F., Morelli, S., et al. (2018). Haplosporidium sp. (Ascetosporea: Haplosporidiidae) come causa di mortalità di massa di Pinna nobilis (L.) a Taranto e in Toscana. In: VII Convegno Nazionale Società Italiana di Ricerca Applicata alla Molluschicoltura. Italy: Napoli.
García-March, J. R., Tena-Medialdea, J., Henandis-Caballero, S., Vázquez-Luis, M., López, D., Téllez, C., et al. (2020). Can we save a marine species affected by a highly-infective-highly-lethal waterborne disease from extinction? Biol. Conserv. 243:108498. doi: 10.1016/j.biocon.2020.108498
Giménez-Casalduero, F., Gomariz-Castillo, F., Alonso-Sarría, F., Cortés, E., Izquierdo-Muñoz, A., and Ramos-Esplá, A. A. (2020). Pinna nobilis in the Mar Menor coastal lagoon: a story of colonization and uncertainty. Mar. Ecol. Prog. Ser. 652, 77–94. doi: 10.3354/meps13468
Giménez-Romero, A., Grau, A., Hendriks, I. E., and Matias, M. A. (2021). Modelling parasite-produced marine diseases: The case of the mass mortality event of Pinna nobilis. Ecol. Model. 459:109705. doi: 10.1016/j.ecolmodel.2021.109705
Gómez-Couso, H., Freire-Santos, F., Ortega-Iñarrea, M., Castro-Hermida, J., and Ares-Mazas, M. (2003). Environmental dispersal of Cryptosporidium parvum oocysts and cross transmission in cultured bivalve molluscs. Parasitol. Res. 90, 140–142. doi: 10.1007/s00436-002-0748-7
Gutiérrez, M. (1967). Coloración histoloógica para ovarios de peces, crustáceos y moluscos. Inv. Pesq. 31, 265–271.
Hartikainen, H., Ashford, O. S., Berney, C., Okamura, B., Feist, S. W., Baker-Austin, C., et al. (2014). Lineage-specific molecular probing reveals novel diversity and ecological partitioning of haplosporidians. ISME J. 8, 177–186. doi: 10.1038/ismej.2013.136
Hashish, E., Merwad, A., Elgaml, S., Amer, A., Kamal, H., Elsadek, A., et al. (2018). Mycobacterium marinum infection in fish and man: epidemiology, pathophysiology and management; a review. Vet. Q. 38, 35–46. doi: 10.1080/01652176.2018.1447171
Hendriks, I. E., Basso, L., Deudero, S., Cabanellas-Reboredo, M., and Alvarez, E. (2012). Relative growth rates of the noble pen shell Pinna nobilis throughout ontogeny around the Balearic Islands (Western Mediterranean, Spain). J. Shellfish. Res. 31, 749–756. doi: 10.2983/035.031.0319
IUCN (2018). Pinna nobilis mass mortality outbreak in the Mediterranean – call to action for central and western Mediterranean countries. 2018. Gland: IUCN.
Katsanevakis, S., Carella, F., Cinar, M., Čižmek, H., Jimenez, C., Kersting, D. K., et al. (2021). The Fan mussel Pinna nobilis on the brink of extinction in the Mediterranean. Ref. Mod. Earth Syst. Env. Sci. 2021:2. doi: 10.1016/B978-0-12-821139-7.00070-2
Katsanevakis, S., Tsirintanis, K., Tsaparis, D., Doukas, D., Sini, M., Athanassopoulou, F., et al. (2019). The cryptogenic parasite Haplosporidium pinnae invades the Aegean Sea and causes the collapse of Pinna nobilis populations. Aquat. Invas. 14, 150–164. doi: 10.3391/ai.2019.14.2.01
Kotlowski, R., Martin, A., Ablordey, A., Chemlal, K., Fonteyne, P. A., and Portaels, F. (2004). One-tube cell lysis and DNA extraction procedure for PCR-based detection of Mycobacterium ulcerans in aquatic insects, mollusks and fish. J. Med. Microbiol. 53, 927–933. doi: 10.1099/jmm.0.45593-0
Künili, I. E., Gürkan, S. E., Aksu, A., Turgay, E., Çakir, F., Gürkan, M., et al. (2021). Mass mortality in endangered fan mussels Pinna nobilis (Linnaeus 1758) caused by co-infection of Haplosporidium pinnae and multiple Vibrio infection in Çanakkale Strait, Turkey. Biomarkers 2021:1910344. doi: 10.1080/1354750X.2021.1910344
Landis, J., and Koch, G. (1977). The Measurement of Observer Agreement for Categorical Data. Biometrics 33, 159–174. doi: 10.2307/2529310
Lattos, A., Bitchava, K., Giantsis, I. A., Theodorou, J. A., Batargias, C., and Michaelidis, B. (2021). The Implication of Vibrio Bacteria in the Winter Mortalities of the Critically Endangered Pinna nobilis. Microorganisms 9:922. doi: 10.3390/microorganisms9050922
Lattos, A., Giantsis, I. A., Karagiannis, D., and Michaelidis, B. (2020a). First detection of the invasive Haplosporidian and Mycobacteria parasites hosting the endangered bivalve Pinna nobilis in Thermaikos Gulf, North Greece. Mar. Env. Res. 155:104889. doi: 10.1016/j.marenvres.2020.104889
Lattos, A., Giantsis, I. A., Karagiannis, D., Theodorou, J. A., and Michaelidis, B. (2020b). Gut Symbiotic Microbial Communities in the IUCN Critically Endangered Pinna nobilis suffering from Mass Mortalities, Revealed by 16S rRNA Amplicon NGS). Pathogens 9:1002. doi: 10.3390/pathogens9121002
López-Sanmartín, M., Catanese, G., Grau, A., Valencia, J. M., Garcıía-March, J. R., and Navas, J. I (2019). Real-Time PCR based test for the early diagnosis of Haplosporidium pinnae affecting fan mussel Pinna nobilis. PLoS One 14:e0212028. doi: 10.1371/journal.pone.0212028
Luna, L. G. (1968). Manual of histologic staining methods of the Armed Forces Institute of Pathology, Chap. New York, NY. McGrawHill Book Company, 258.
Lynch, S. A., Abollo, E., Ramilo, A., Cao, A., Culloty, S. C., and Villalba, A. (2010). Observations raise the question if the Pacific oyster, Crassostrea gigas, can act as either a carrier or a reservoir for Bonamia ostreae or Bonamia exitiosa. Parasitology 137, 1515–1526. doi: 10.1017/S0031182010000326
Lynch, S. A., Armitage, D. V., Coughlan, J., Mulcahy, M. F., and Culloty, S. C. (2007). Investigating the possible role of benthic macroinvertebrates and zooplankton in the life cycle of the haplosporidian Bonamia ostreae. Exp. Parasitol. 115, 359–368. doi: 10.1016/j.exppara.2006.09.021
Masami, K., and Hiroshi, K. (1959). Studies on the Food Putrefaction (III): On amines production of putrefactive bacteria in fish. Bull. Univ. Osaka Prefect., Ser. B Agric. biol. 9, 117–127.
Nebot-Colomer, E., Álvarez, E., Belando, M. D., Deudero, S., Catanese, G., Bernardeau Esteller, J., et al. (2021). Living under threat: Will one of the last Pinna nobilis populations be able to survive? Aquat. Conserv. Mar. Freshw. Ecosyst 2021, 1–13. doi: 10.1002/aqc.3738
Öndes, F., Alan, V., Açkali, B., and Güçlüsoy, H. (2020). Mass mortality of the fan mussel, Pinna nobilis in Turkey (eastern Mediterranean). Mar. Ecol. 00:e12607. doi: 10.1111/maec.12607
Özalp, H. B., and Kersting, D. K. (2020). A pan-Mediterranean extinction? Pinna nobilis mass mortality has reached the Turkish straits system. Mar. Biodivers. 50:81. doi: 10.1007/s12526-020-01110-7
Panarese, R., Tedesco, P., Chimienti, G., Latrofa, M. S., Quaglio, F., Passantino, G., et al. (2019). Haplosporidium pinnae associated with mass mortality in endangered Pinna nobilis (Linnaeus 1758) fan mussels. J. Invertebr. Pathol. 164, 32–37. doi: 10.1016/j.jip.2019.04.005
Pavlinec, Z., Zupicic, I. G., Oraic, D., Petani, B., Mustac, B., Mihaljevic, Z., et al. (2020). Assessment of predominant bacteria in noble pen shell (Pinna nobilis) collected in the Eastern Adriatic Sea. Environ. Monit. Assess 192:581. doi: 10.1007/s10661-020-08541-6
Pérez-Ruzafa, A., Marcos, C., and Gilabert, J. (2005). “The ecology of the Mar Menor Coastal Lagoon: a fast-changing ecosystem under human pressure,” in Coastal lagoons. Ecosystem processes and modeling for sustainable use and development, eds I. E. Gonenc and J. P. Wolflin (Boca Raton: CRC Press), 392–422.
Peyran, C., Boissin, E., Morage, T. Nebot-Colomer, E., Iwankow, G., and Planes, S. (2021). Genetic homogeneity of the critically endangered fan mussel, Pinna nobilis, throughout lagoons of the Gulf of Lion (North-Western Mediterranean Sea). Sci. Rep. 11:7805. doi: 10.1038/s41598-021-87493-4
Prado, P., Caiola, N., and Ibáñez, C. (2014). Habitat use by a large population of Pinna nobilis in shallow waters. Sci. Mar. 78, 555–565. doi: 10.3989/scimar.04087.03A
Prado, P., Carrasco, N., Catanese, G., Grau, A., Cabanes, P., Carella, F., et al. (2020a). Presence of Vibrio mediterranei associated to major mortality in stabled individuals of Pinna nobilis L. Aquaculture. 519:734899. doi: 10.1016/j.aquaculture.2019.734899
Prado, P., Andree, K. A., Trigos, S., Carrasco, N., Caiola, N., García-March, J. R., et al. (2020b). Breeding, planktonic and settlement factors shape recruitment patterns of one of the last remaining major population of Pinna nobilis within Spanish waters. Hydrobiologia. 847, 771–786. doi: 10.1007/s10750-019-04137-5
Prado, P., Grau, A., Catanese, G., Cabanes, P., Carella, F., Fernández-Tejedor, M., et al. (2021). Pinna nobilis in suboptimal environments are more resilient to disease but more vulnerable to catastrophic events. Mar. Environ. Res. 163:105220. doi: 10.1016/j.marenvres.2020.105220
Ray, S. M. (1963). A review of the culture method of detecting Dermocystidium marinum with suggested modifications and precautions. Proc. Natl. Shellfish Assoc. 54, 55–69.
Rivera-Ingraham, G. A., Malanga, G., Puntarulo, S., Pérez, A. F., Ruiz-Tabares, A., Maestre, M., et al. (2013). Antioxidant defenses and trace metal bioaccumulation capacity of Cymbula nigra (Gastropoda: Patellidae). Water Air. Soil. Pollut. 224, 1458–1471. doi: 10.1007/s11270-013-1458-8
Rodríguez, F., and Navas, J. I. (1995). A comparison of gill and hemolymph assays for the thioglycollate diagnosis of Perkinsus atlanticus (Apicomplexa, Perkinsea) in clams, Ruditapes decussatus (L.) and Ruditapes philippinarum (Adams et Reeve). Aquaculture. 132, 145–152. doi: 10.1016/0044-8486(94)00393-3
Rodrıguez, S., Balboa, S., Olveira, G., Montes, J., Moreno, D., Barrajon, A., et al. (2017). “First report of mass mortalities in natural population of Pinna nobilis. A microbial perspective. FEMS7- 3233,” in The 7th Congress of European Microbiologists (Valencia: FEMS).
Sambrook, J., Fritsch, F. J., and Maniatis, T. (1989). Molecular Cloning, a Laboratory Manual, 2nd Edn. Plainview, N.Y: Cold Spring Harbor Laboratory Press, 545.
Šarić, T., Župan, I., Aceto, S., Villari, G., Palić, D., De Vico, G., et al. (2020). Epidemiology of Noble Pen Shell (Pinna nobilis L. 1758). Mass Mortality Events in Adriatic Sea Is Characterised with Rapid Spreading and Acute Disease Progression. Pathogens. 9:776. doi: 10.3390/pathogens9100776
Sas, H., Deden, B., Kamermans, P., Ermgassen, P. S. E., Pogoda, B., Preston, J., et al. (2020). Bonamia infection in native oysters (Ostrea edulis) in relation to European restoration projects. Aquatic Conserv: Mar. Freshw. Ecosyst. 30, 2150–2162. doi: 10.1002/aqc.3430
Scarpa, F., Sanna, D., Azzena, I., Mugetti, D., Cerruti, F., Hosseini, S., et al. (2020). Multiple Non-Species-Specific Pathogens Possibly Triggered the Mass Mortality in Pinna nobilis. Life. 10:238. doi: 10.3390/life10100238
Tsokos, M. (2005). “Postmortem Changes and Artifacts Occurring During the Early Postmortem Interval,” in Forensic Pathology Reviews, Vol. 3, ed. M. Tsokos (Totowa: Humana Press), 183–238. doi: 10.1007/978-1-59259-910-3_5
Vázquez-Luis, M., Álvarez, E., Barrajón, A., García-March, J. R., Grau, A., Hendriks, I. E., et al. (2017). S.O.S. Pinna nobilis: A Mass Mortality Event in Western Mediterranean Sea. Front. Mar. Sci. 4:220. doi: 10.3389/fmars.2017.00220
Vázquez-Luis, M., March, D., Alvarez, E., Alvarez-Berastegui, D., and Deudero, S. (2014). Spatial distribution modelling of the endangered bivalve Pinna nobilis in a Marine Protected Area. Mediterr. Mar. Sci. 15, 626–634. doi: 10.12681/mms.796
Vidal-Vijande, E., Pascual, A., Barnier, B., Molines, J. M., and Tintore, J. (2011). Analysis of a 44-year hindcast for the Mediterranean Sea: comparison with altimetry and in situ observations. Sci. Mar. 75, 71–86. doi: 10.3989/scimar.2011.75n1071
Villalba, A., Casas, S. M., López, C., and Carballal, M. J. (2005). Study of perkinsosis in the carpet shell clam Tapes decussatus in Galicia (NW Spain). II. Temporal pattern of disease dynamics and association with clam mortality. Dis. Aquat. Org. 65, 257–267. doi: 10.3354/dao065257
Wesselmann, M., González-Wanguemert, M., Serrao, E. A., Engelen, A. H., Renault, L., Garcia-March, J. R., et al. (2018). Genetic and oceanographic tools reveal high population connectivity and diversity in the endangered pen shell Pinna nobilis. Sci. Rep. 8, 1–11. doi: 10.1038/s41598-018-23004-2
Keywords: Pinna nobilis, Haplosporidium pinnae, Mycobacterium sp., Gram negative bacteria, molecular biology, histology
Citation: Grau A, Villalba A, Navas JI, Hansjosten B, Valencia JM, García-March JR, Prado P, Follana-Berná G, Morage T, Vázquez-Luis M, Álvarez E, Katharios P, Pavloudi C, Nebot-Colomer E, Tena-Medialdea J, Lopez-Sanmartín M, Peyran C, Čižmek H, Sarafidou G, Issaris Y, Tüney-Kizilkaya I, Deudero S, Planes S and Catanese G (2022) Wide-Geographic and Long-Term Analysis of the Role of Pathogens in the Decline of Pinna nobilis to Critically Endangered Species. Front. Mar. Sci. 9:666640. doi: 10.3389/fmars.2022.666640
Received: 10 February 2021; Accepted: 15 February 2022;
Published: 17 March 2022.
Edited by:
Cosimo Solidoro, Istituto Nazionale di Oceanografia e di Geofisica Sperimentale, ItalyReviewed by:
Free Espinosa Torre, Seville University, SpainIoannis A. Giantsis, University of Western Macedonia, Greece
Copyright © 2022 Grau, Villalba, Navas, Hansjosten, Valencia, García-March, Prado, Follana-Berná, Morage, Vázquez-Luis, Álvarez, Katharios, Pavloudi, Nebot-Colomer, Tena-Medialdea, Lopez-Sanmartín, Peyran, Čižmek, Sarafidou, Issaris, Tüney-Kizilkaya, Deudero, Planes and Catanese. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Antonio Villalba, YW50b25pby52aWxsYWxiYS5nYXJjaWFAeHVudGEuZ2Fs; Gaetano Catanese, Z2NhdGFuZXNlQGRncGVzY2EuY2FpYi5lcw==