 Kristin H. Westdal
Kristin H. Westdal Pierre R. Richard
Pierre R. Richard Jessica Sportelli4
Jessica Sportelli4 Steve H. Ferguson
Steve H. Ferguson- 1Department of Biological Sciences, University of Manitoba, Winnipeg, MB, Canada
- 2Oceans North, Ottawa, ON, Canada
- 3Independent Researcher, Winnipeg, MB, Canada
- 4Scripps Institution of Oceanography, University of California San Diego, La Jolla, CA, United States
- 5Independent Researcher, Churchill, MB, Canada
- 6Fisheries and Oceans Canada, Winnipeg, MB, Canada
Beluga whales (Delphinapterus leucas) are social animals that return to the same estuaries each summer season. The Western Hudson Bay belugas in the Churchill River estuary appear to be attracted to boats, resulting in whale-watching being a popular tourism activity for the area. However, regulators have raised concerns about whether this activity has negative consequences on the beluga population. We hypothesized that more vulnerable groups, such as groups containing calves, would avoid whale-watching vessels, while juvenile groups, who appear to have a more developed curiosity, would be more likely to interact with vessels. To test these hypotheses, behavioural observations were conducted in 2019, 2020, and 2021, in the estuary using a similar methodology to a study conducted in 2005 and 2006. An increase in tourism since that initial study offered a unique opportunity to compare potential change over time. In this study, a group of whales was randomly selected; the group type, initial behaviour type, distance to vessel, and observed behaviour with distance to vessel during three-minute focal follows were recorded. The same method was also applied from shore and we compared frequencies of whale behaviour during experimental (vessels present) and control (vessels greater than 300m away) periods. Results showed that there was a significant difference in the mean group size of belugas in the presence of vessels compared to those observed in the absence of vessels. There was also a significant difference in beluga behaviour as well as a significant behavioural difference between age class of belugas in the presence of vessels, compared to their behaviour in absence of vessels. During focal follows from vessels, belugas spent 63% of their time interacting with vessels, when within 25 m of a vessel, an increase from the 48% observed from the previous study conducted 15 years previous. Our results overall showed beluga behaviour differed in the experimental compared to control settings but suggest that those belugas’ reactions to vessels are not necessarily deleterious to them, in that direct interactions with wildlife are low and unlikely to carry risk, and that they may have habituated to whale-watching vessels over the last two decades.
Introduction
Tourism vessel traffic has increased in the Arctic, particularly since 2010 (Johnston et al., 2017). With “last-chance tourism” gaining momentum, people are drawn to the north by the notion of disappearing sea ice and ice associated species, which may shift their distribution under future climate scenarios (Chambault et al., 2022). Increases in vessel traffic are correlated with decreases in sea-ice concentrations (Pizzolato et al., 2016) and with ice-free summers in the Arctic expected as early as 2035 (Guarino et al., 2020), and before 2050 (Notz and SIMIP Community, 2020), marine mammals are being exposed to more marine vessel traffic with little understanding of the consequences (Hauser et al., 2018). In Canada, small vessel traffic, in the form of whale-watching boats, are also increasing in number, with the promise of close-range encounters and operating over a longer open-water season. The presence of whale-watching vessels has been shown to cause changes in whale behaviour (Lien, 2001), though the majority of these findings came from areas with high-density commercial and tourism vessel traffic, like that of the west coast of Canada (Malcolm, 2008). Tourism in the form of whale-watching is increasing in popularity in Nunavut, and in the Eastern Canadian Arctic and sub-Arctic. Recent federal changes to viewing regulations (Regulations Amending the Marine Mammal Regulations, SOR/2018-126) have brought focus to the small beluga (Delphinapterus leucas) whale-watching businesses in Churchill, Manitoba, on the coast of Western Hudson Bay.
Western Hudson Bay is home to the largest population of belugas in the world (Matthews et al., 2017), with summer distribution mainly in and around three estuaries of the Manitoba coast: the Nelson, the Churchill and the Seal estuaries (Richard, 2005) (Figure 1). Belugas feed, moult, and rear their young likely in the same estuaries each year (Brown Gladden et al., 1997; Brown Gladden et al., 1999), perhaps returning to these locations for social reasons more than any one environmental factor (Westdal et al., 2022). The Churchill estuary sees approximately 3,000 animals seasonally, moving in and out of the estuary daily, of the total estimated Western Hudson Bay beluga population of just over 54,000 (Matthews et al., 2017). Canadian populations of belugas are in varying states of health and the Western Hudson Bay population, although considered healthy and ‘Not At Risk’ as of 2020 under the Committee On the Status of Endangered Wildlife in Canada (COSEWIC) designations, is expected to be under increased pressure in the future from predicted sea ice reduction, increased shipping, climate change and increased ocean noise (COSEWIC, 2004; COSEWIC, 2020). With a predictable presence of belugas moving in and out of the Churchill estuary daily, and with such a large number of whales in a relatively small geographic area, this estuary has been popular for whale-watching. Commercial operations began in 1977 with small boats (zodiacs and aluminum small crafts) with up to 8 passengers. Two larger commercial vessels with up to 32 passengers were added over the last two decades as well as kayaks and paddleboards. Whales in the estuary approach vessels at close distance and appear to actively seek out interactions with vessels of all types and sizes, which is a unique behaviour for an Arctic cetacean. These established operations and unique behaviours provided an ideal setting to study beluga behaviour in response to these vessels.
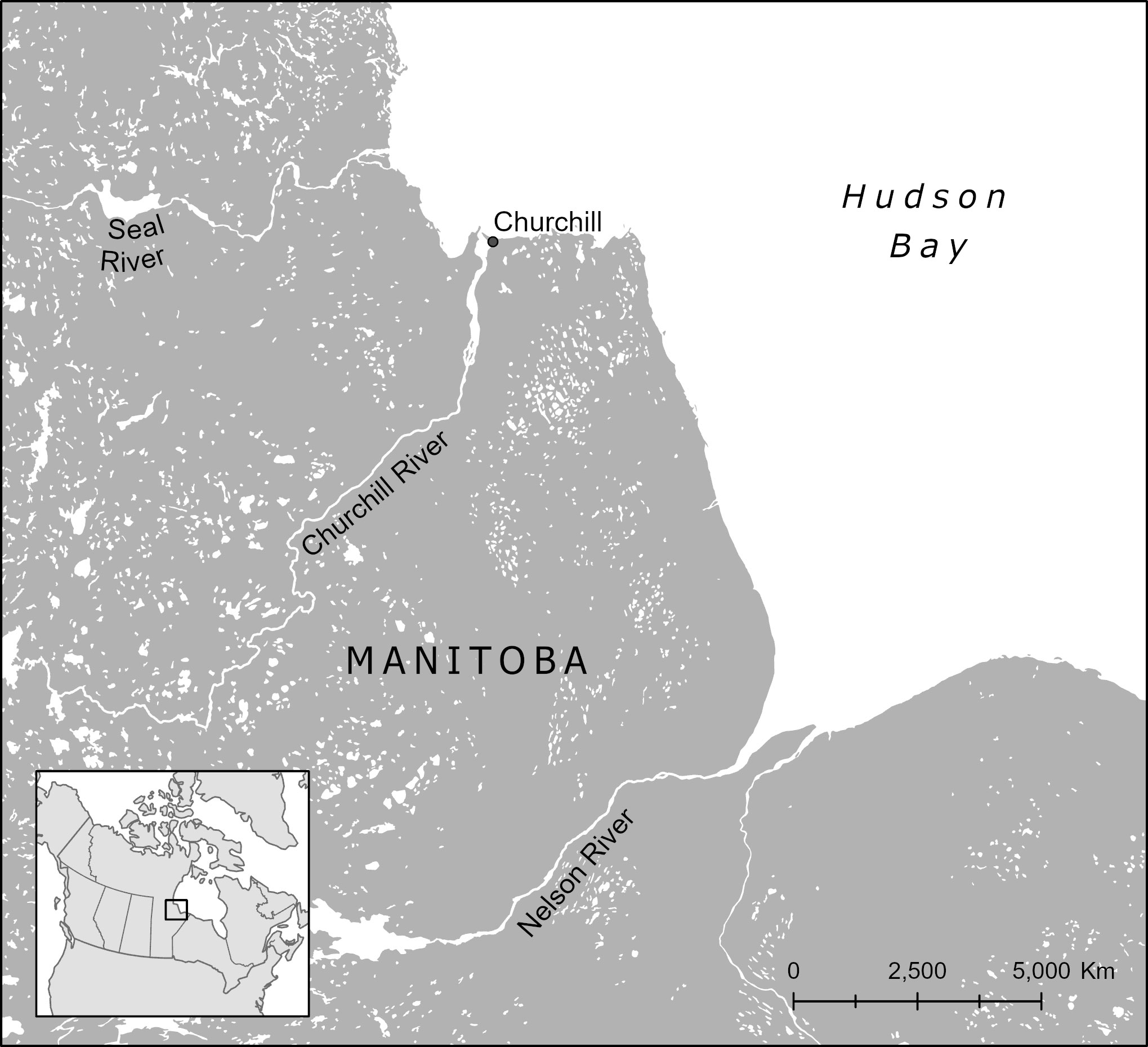
Figure 1 Map of the study area showing Churchill, as well as the Seal River to the north and the Nelson River to the south where larger aggregations of belugas are located in summer.
Beluga behaviour was monitored and characterized in response to tourism vessels, following similar methods to a 2005-06 study (Malcolm and Penner, 2011). Our goal was to assess behavioural differences of beluga groups in the proximity of vessels compared to beluga groups distant from any vessels. Based on prior experience and anecdotal observations, as well as females with calves requiring more energy, we hypothesized that behaviour would be different overall in the presence (compared to control setting of vessels absent or greater than 300 m away) of vessels and that younger age classes of whales were likely to exhibit a greater behavioural response to distance from the whale-watching vessels. Specifically, we hypothesized that the presence of whale-watching vessels would result in females with calves of the year (termed Calf groups here) keeping greater distances to whale-watching vessels than the Adult or Juvenile groups. We also hypothesized that Juvenile groups would be least likely to display avoidance behaviour and more likely to approach and be found closer to whale-watching vessels.
Methods
Field methods
In July-August of 2019, 2020, and 2021 observations were made from (1) whale-watching tour boats (small inflatable zodiacs, 17’5” to 19’2” in length); (2) the Sea North II (a 32-passenger tour vessel, 42’ in length); and (3) the Port of Churchill’s port wall (approximately 5 meters above water level) (Figure 2). Different vessels were used over the three years due to change in boat availability. Observers were confident recording behaviour out to 300m on both vessel types and belugas were known to exhibited close interactions with all types of vessels, big or small, in this area.

Figure 2 The Sea North II passenger vessel (L) and zodiacs (R) used to provide tourists with views of beluga whales in the Churchill estuary (Photo credits: L: Sea North; R: Oceans North).
The port wall was used for control behaviour observations when no vessels were within 300 metres of our focal groups of beluga whales.
Using teams of two observers, a focal group of belugas was randomly selected by counting from left to right the number of groups visible in a 180-degree arc forward of the bow of the vessel (or the port wall) and within 150 meters of the observers. Having numbered each group within that range, a random number was chosen from a random number table and the group numbered as such was selected. The distance of the group to the vessel was determined using a laser range-finder (Nikon Aculon in 2019 and Nikon Prostaff 1000 in 2020 and 2021). Group size, composition (Table 1) and behaviour (Table 2) were recorded.
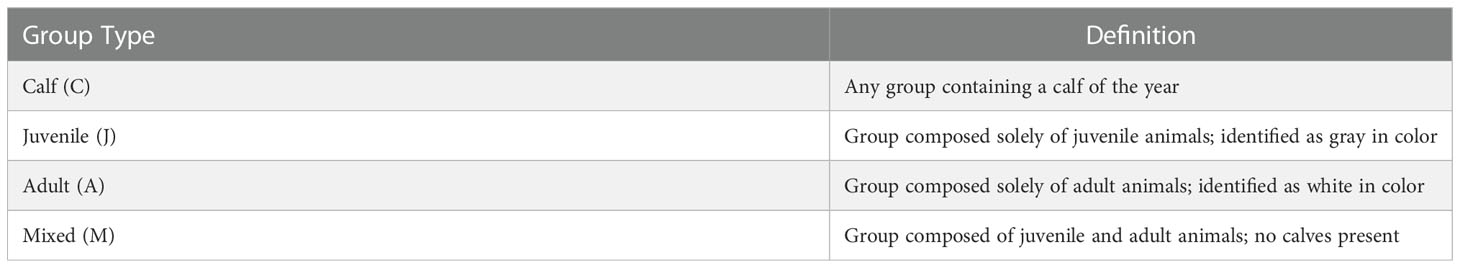
Table 1 Classification of beluga groups by age class.

Table 2 Classification of beluga behaviour types (adapted from Malcom and Penner, 2011).
During three-minute focal follows, behaviour and distance to observers were recorded. Behaviour types recorded included the following: travelling, socializing/milling, resting, feeding and interacting with vessels (Table 2). Note, although newborn calves were present, no nursing was observed. If observers lost track of a focal follow group, or the group split up or merged with another group, the focal follow was stopped and the data discarded. The same observation method was used in control observations from the port wall, when vessels were greater than 300 m from group being observed, with the addition of measuring distance to the group. If a vessel came within 300 meters of a group during control observations, the focal follow was stopped and the data was discarded. 300 meters was selected as the distance limit to make the data comparable to the Malcolm and Penner study (2011). In addition, the river near the mouth, where observation took place, is fairly narrow and there are few places where vessels could be further than 300 m.
Data was grouped by distance from the vessel following methodology used by Malcolm and Penner (2011): (0 to 25 m, 26 to 50 m, 51 to 75 m, 76 to 100 m, 101 to 125 m, 125 to 150 m and >150 m) and age class and behaviour frequencies were tabulated by distance zones and group age class. Observations were not corrected for height above sea level (less than 5 metres in vessel and shore observations) because the corrections were within than the laser range finder’s accuracy range (+/- 1 metre). If behaviour changed during a 3-minute focal follow, the behaviour that was observed the longest was retained as the dominant behaviour for that sample. Observations were not undertaken in sea state indices over 2 (i.e. no white caps).
Data analysis
Mean group size was calculated under experimental and control conditions and a t-test was used to determine if there was a significant difference between the two. Chi-Square Tests were used to compare beluga behaviour by age class in the experimental and control settings of vessels to determine if whales behaved differently when vessels were present. Beluga behaviour was then compared by age class to determine if the frequency of observed behaviours of age classes differed in the presence of vessels. To further examine the behaviour observed in the presence of vessels for each age class, the percentage of total frequency of behavioural classes and the mean observed distance was calculated for each behaviour by year.
To explore the probability of groups exhibiting a certain behaviour with distance from the vessel, a logistic regression was run in JASP (JASP Team, 2021), where the Behaviour category (coded Y for yes, the behaviour took place) is the dependent variable and Distance and Group classes are explanatory parameters. Distance was treated as a continuous dependent parameter while Group Class was treated as a factor. Since we have categorical variables (groups class and year) as predictors, the R code (run via JASP) for logistic regression uses one of these levels as the reference (Le, 2018). In our study, the group class Adult and the year 2019 were used as reference. The logistic regression output gives the difference between the other group classes over the reference class (Adult) on behaviour probability, as well as between 2019 and the other years when year was used as a factor in the regression.
Finally, observations of whale-vessel interactions, which were noted in the first distance category (0-25m), were tallied for each age class and overall interaction frequency near the vessel was calculated as a percent of total behaviours to test for differences among age classes. Results were compared to overall interaction time reported in the study by Malcolm and Penner (2011).
Results
Of the 258 focal follows (experimental and control) in 2019, 77 were discontinued. In 2020, 216 of 415 total follows (experimental and control) were discontinued and in 2021, 249 of 477 (experimental and control) total follows were discontinued due to either glare affecting the observations, too great a distance to the focal group, the observers losing sight of the group, or the focal follow pod merging with other groups of belugas in the area. Consequently, 181 successful follows (experimental and control) remained in 2019 aboard the Sea North II, 199 aboard zodiacs in 2020, and 228 in 2021.
Not all behaviour types were observed across all age classes during experimental and control observations. The most common group type observed was Mixed (43%) (Table 3) and the most common behaviour seen across all years was Travelling (64%) (Table 4). Resting was observed once under experimental conditions and once under control conditions in 2019; was not observed in 2020; and only under control conditions in 2021 and was thus removed from the analysis. Socializing/Milling occurrences were few in both years. An examination of frequency of behaviour between group types indicated that Calf groups were more often observed Feeding, and at a higher percentage of overall focal follows, than other age classes in 2019 (Table 4). In all three years, when Feeding was observed in groups containing Calves, it was at a closer distance to vessels on average than other age classes. Calf groups were also observed Travelling less frequently than other age classes in all years (Table 4).
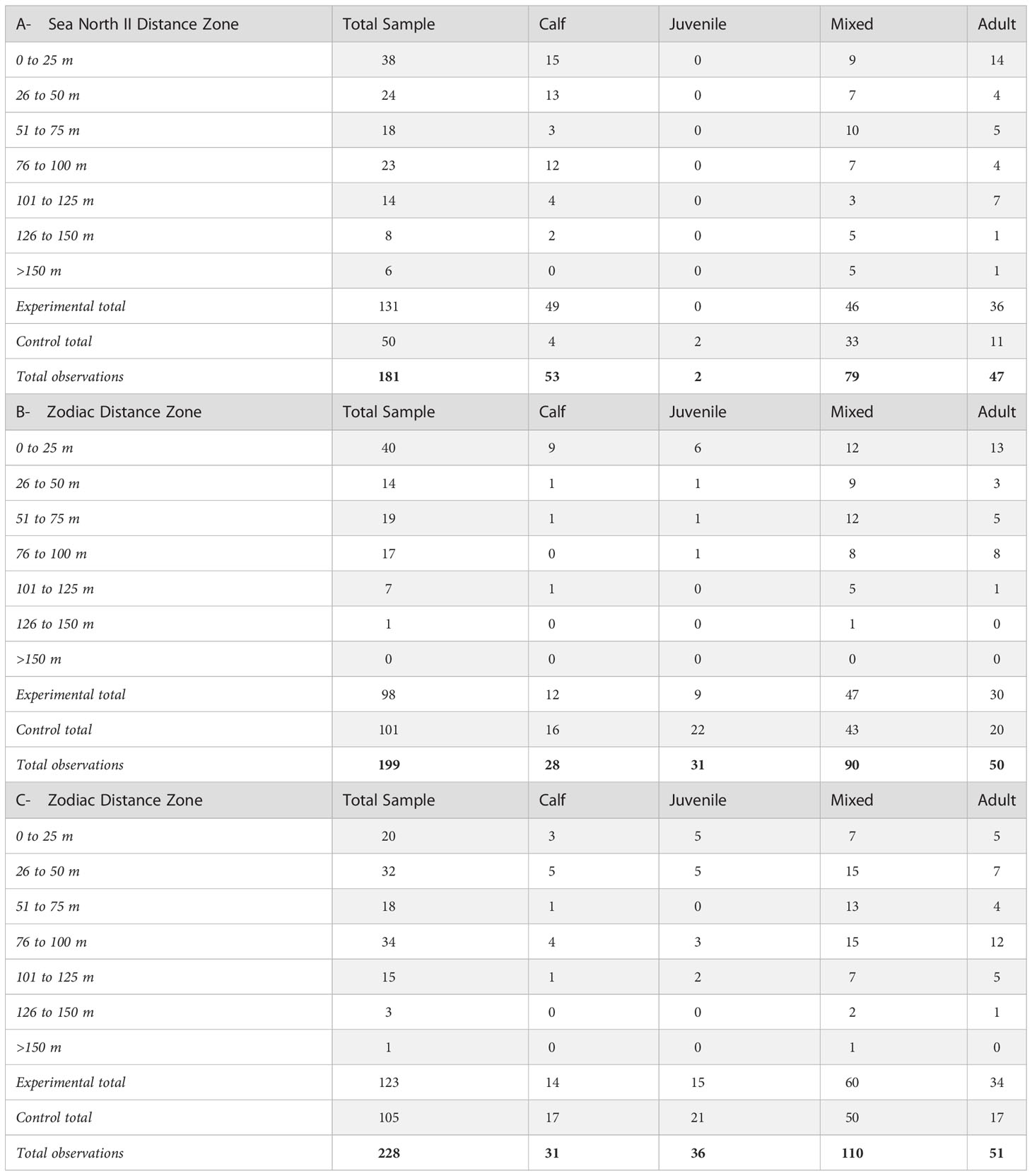
Table 3 Number of distance/behaviour observations of beluga whales in the Churchill estuary, Hudson Bay: A- from the Sea North II passenger vessel in 2019; B - from Zodiacs in 2020; C - from Zodiacs in 2021, and in all years from the control location.
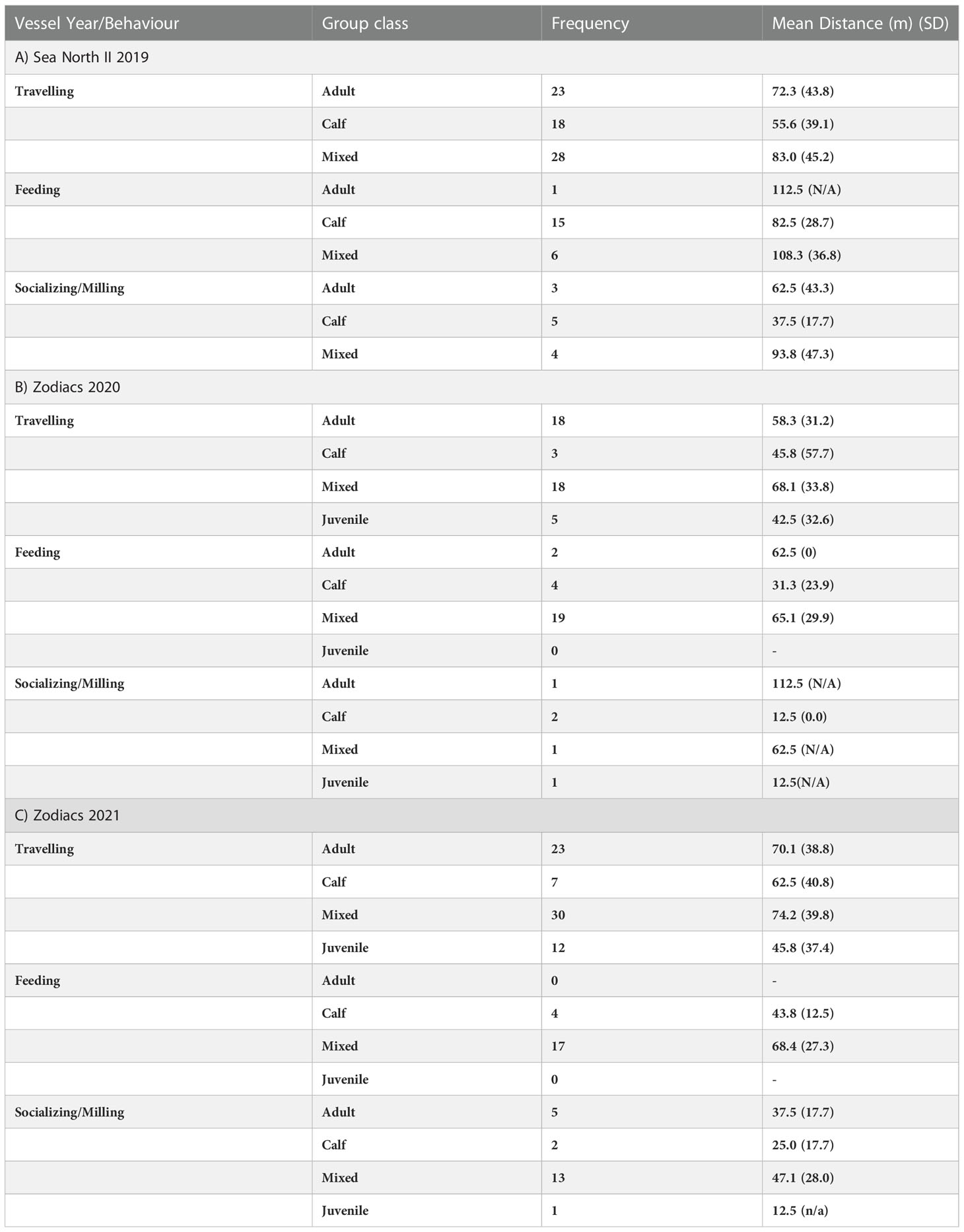
Table 4 Frequencies and mean distances of observed travelling, feeding and socializing/milling behaviour by age class observed from: A) Sea North II vessel in 2019; B) zodiacs in 2020; and C) zodiacs in 2021.
Group sizes ranged from one to >100 animals (observers were not able to estimate group sizes larger than 100), with means ranging from 4 to 12 depending on the year and observation platform (Table 5). There was a significant difference in the mean group size between animals in the presence of vessels and those observed in the absence of vessels (t(4)=3.64, p=0.02). Average group size was larger in the presence of vessels (Table 5).
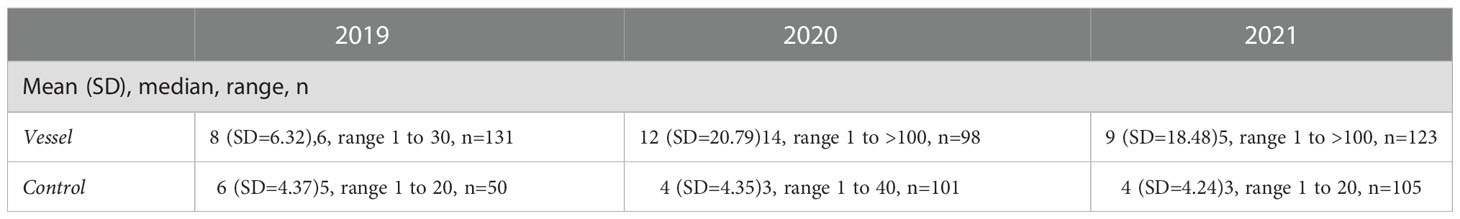
Table 5 Mean and median group size during focal follows in 2019, 2020 and 2021.
There were significant differences for most comparisons of the frequencies of beluga behaviours in the presence of vessels compared to the frequencies of behaviours in the control. This was observed in all age classes, with the exception of juveniles (Table 6). A significant difference was also observed when comparing the frequencies of behaviours of each age class in the presence of vessels (Table 7).
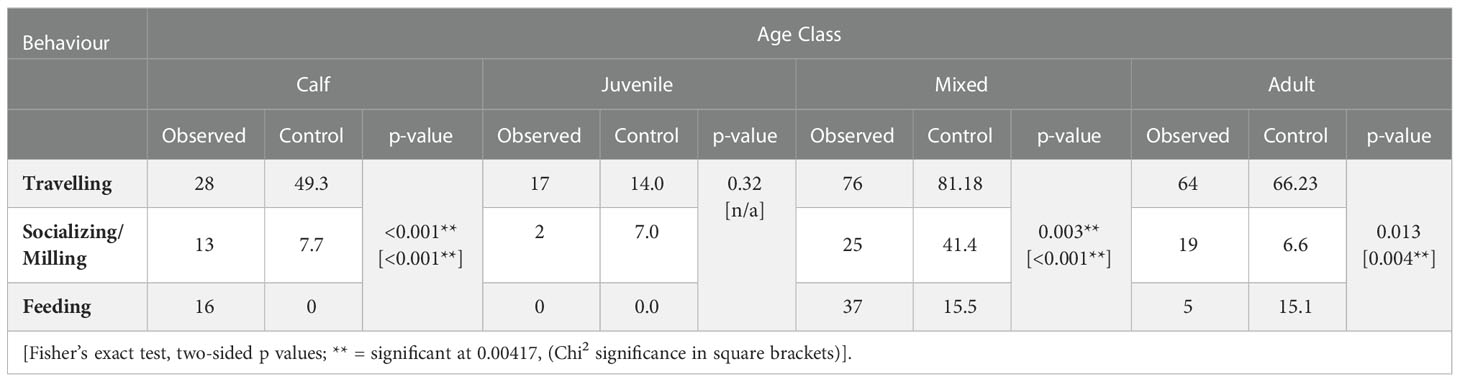
Table 6 Comparison of the frequency of beluga behaviour by age class in the presence (observed) and absence (control) of vessels for all years combined.
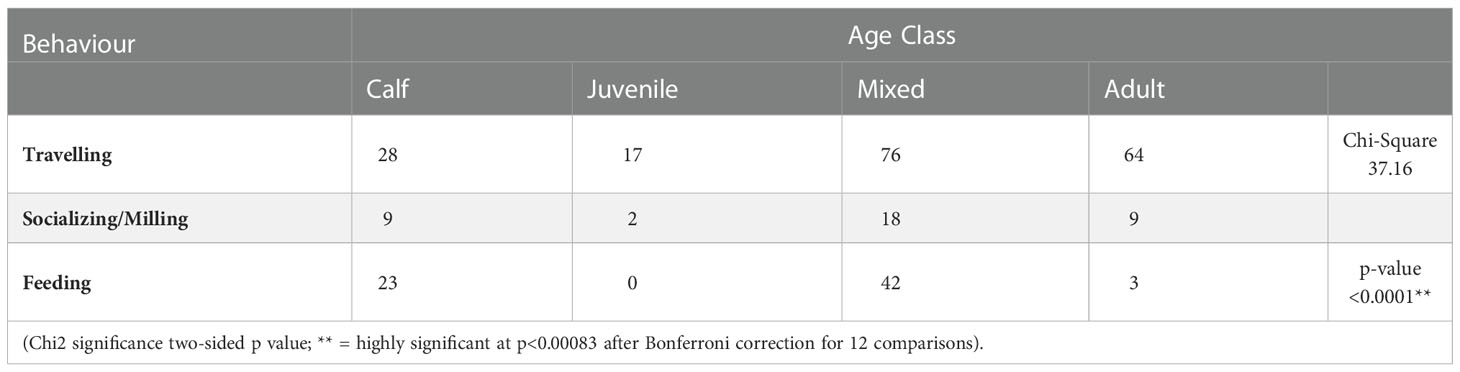
Table 7 Comparison of observed frequency of beluga behaviour in the presence of vessels by age class.
The logistic regressions indicated that the probability of Feeding and Travelling with distance from a vessel was highly significant for the Adult reference group in 2019 (Tables 8A, B). Calf groups had a significantly lesser effect on the probability of Travelling than the Adult reference group, while other age groups did not significantly change it (Table 8A). Calf and Mixed groups had a higher effect on the probability of Feeding with distance compared to the Adult reference class (Table 8B). The probability of Travelling with distance was significant smaller in 2020 and 2021, compared to the 2019 reference year (Table 8A). The Feeding probability with distance was significantly smaller in 2020 than the reference year (2019), but not in 2021. There were few observations of Socializing/Milling and none of the parameters of the logistic regression for Socializing/Milling behaviours produced any significant relationship with distance (Table 8C).
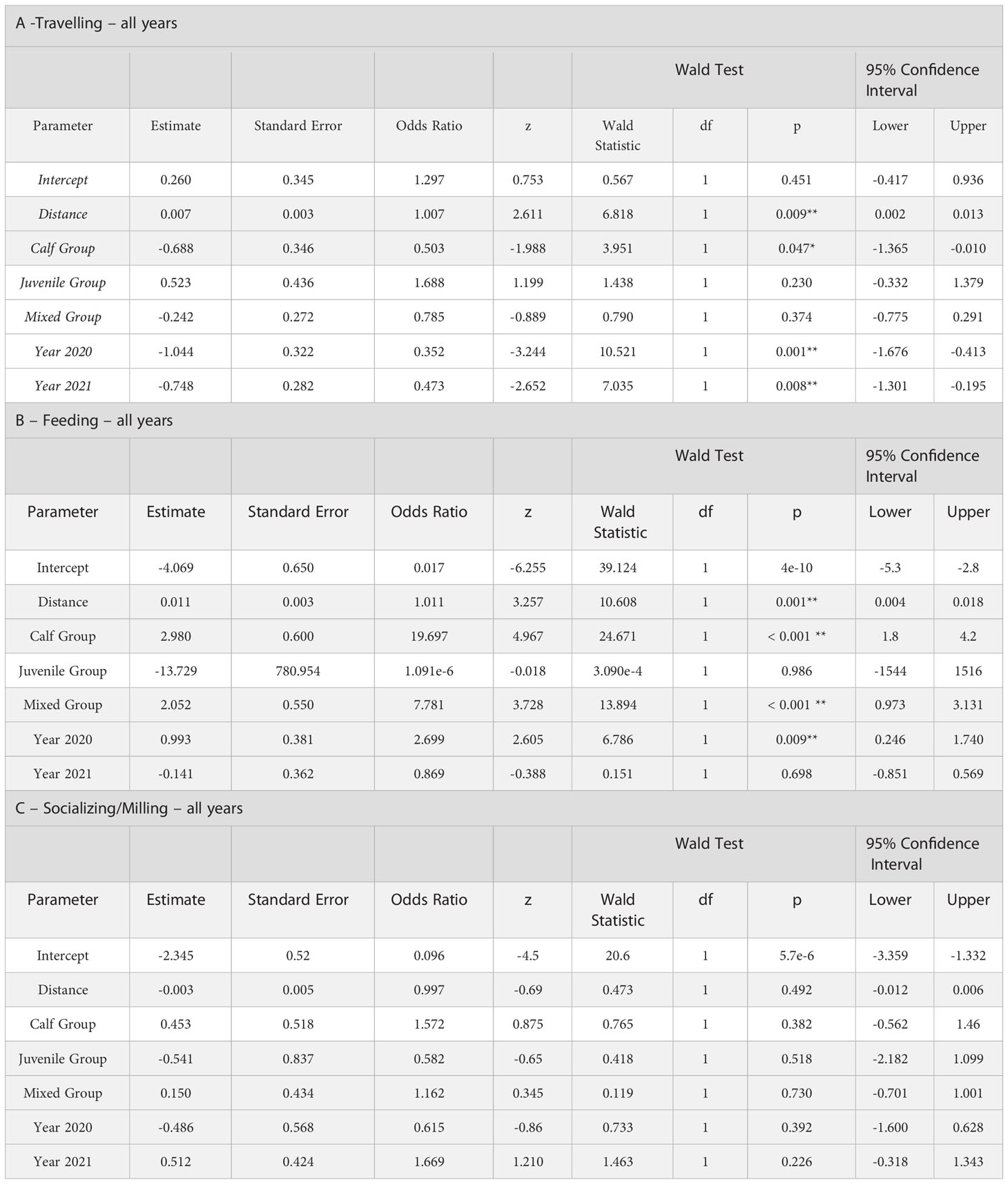
Table 8 Logistic regressions of behavioural frequencies observed from observation vessels, with Distance, Year and Age Group as explanatory variables: A -Travelling seen from Sea North II in 2019 and Zodiacs in 2019, 2020 & 2021; B- Feeding Sea North II and Zodiacs in 2019, 2020 & 2021; C- Socializing/Milling Sea North II and Zodiacs in 2019, 2020 & 2021 (* =significant at p<0.05).
Plots of behaviour probability with distance from vessels indicate a larger effect of distance for Travelling than for Feeding, but the slope of Feeding increased exponentially at higher distances (Figures 3A, B). The regression line for Socializing/Milling did not show a significant effect of that behaviour with distance from vessels (Figure 3C).
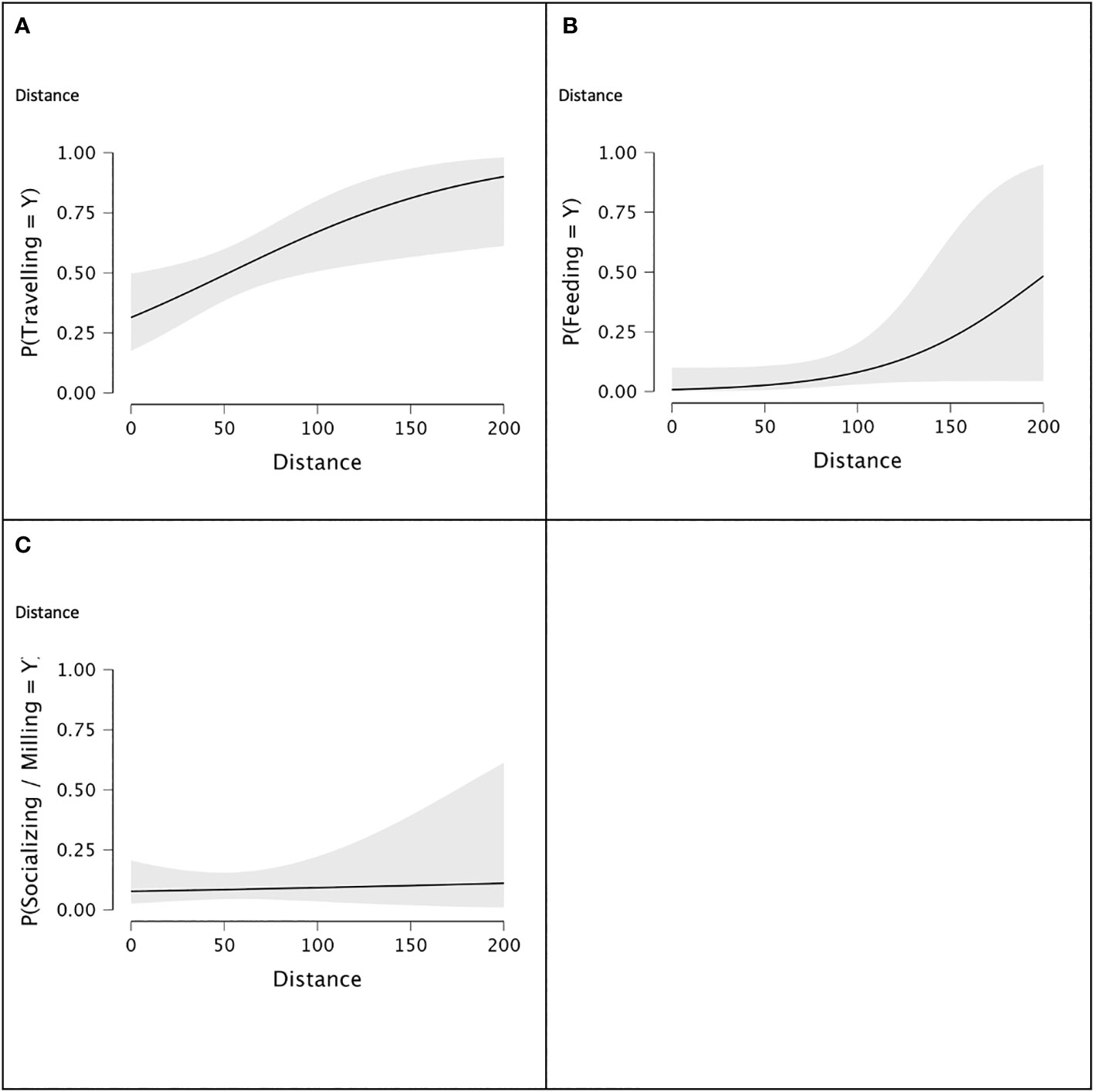
Figure 3 Probability of beluga behaviours with Distance with Year and Group class as logistic regression factors: (A) - Travelling seen from Sea North II vessel or zodiacs in 2019, 2020 and 2021; (B) - Feeding seen from Sea North II vessel or zodiacs in 2019, 2020 and 2021; (C) – Socializing/Milling seen from Sea North II vessel or zodiacs in 2019, 2020 and 2021, where P(Behaviour = Y) means the probability of a behaviour occurring, i.e., Y means “Yes”). Shaded areas around the regression line represent the 95% confidence. .
An attempt to run logistic regressions with Group Class and Year and interactions between these factors and Distance was not possible in R, so we ran regressions for each year separately with the interaction terms between Group Class and Distance. The probability of Travelling with distance was significant for the Adult reference group class in all three years, but the term interaction terms were not significant (Tables 9A–C). Other age groups and the interaction terms did not change the Travelling relationship with distance significantly. Regressions for Feeding and Socializing/Milling did not result in significant relationships with any one of the factor or interactions.
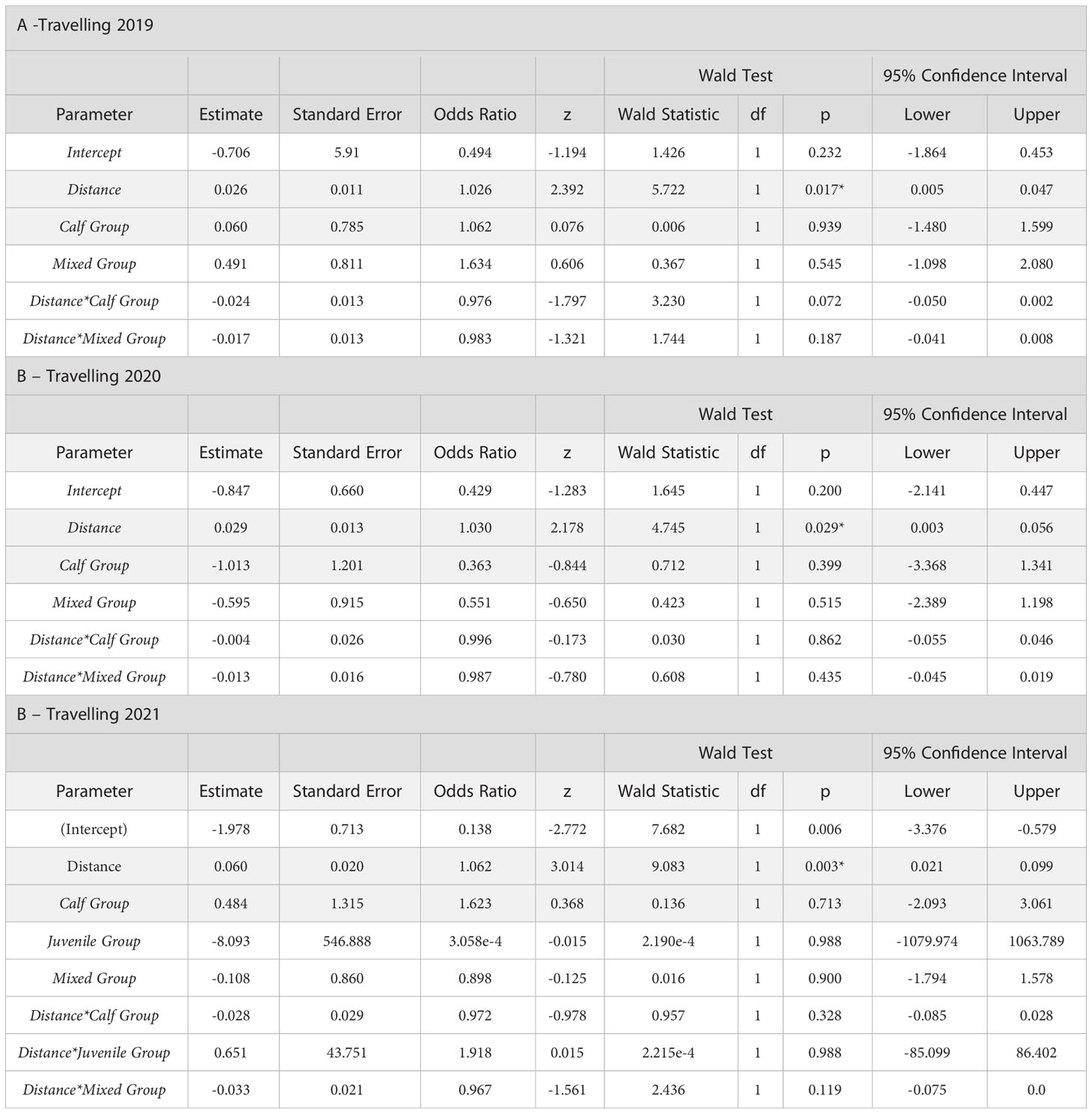
Table 9 Logistic regressions of Travelling frequencies observed from observation vessels, with Distance and Age Group as explanatory variables: A -Travelling seen from Sea North II in 2019; B- Travelling seen from Zodiacs in 2020; C- Travelling seen from Zodiacs in 2021 (* = significant at p<0.05).
Observations of whale-vessel interactions which were noted in the nearest distance category (0-25m) showed that all beluga groups interacted with vessels and they did so almost as frequently across all age groups (Table 10). On average, observed belugas of all age classes and over all three years spent 63% of their time (SD=0.167, range 25% to 89%), interacting with vessels (Table 10).
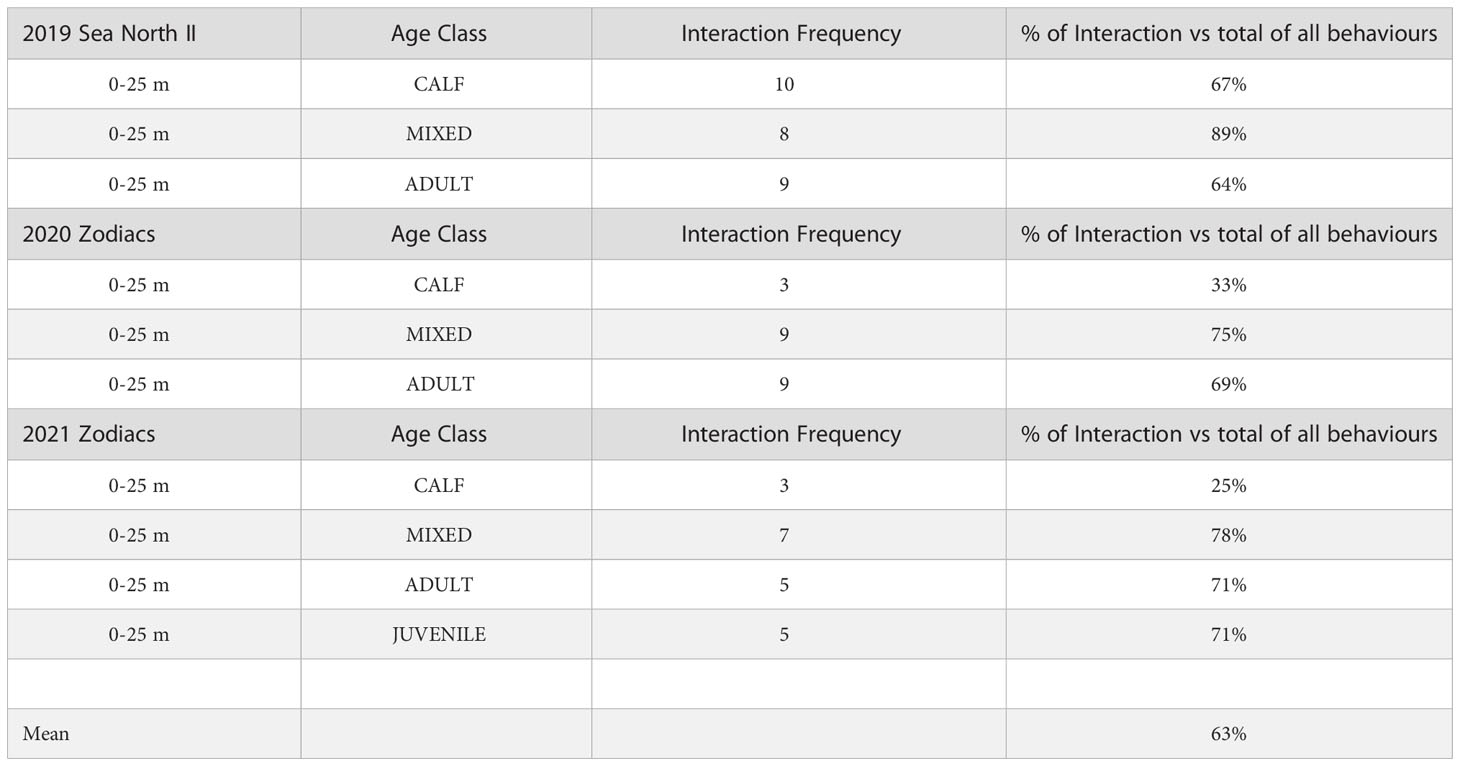
Table 10 Frequency of beluga interactions with whale-watching vessels by year and age group.
Discussion
Published accounts of behavioural changes of cetaceans in the presence of small vessels are not uncommon. Changes include vessel avoidance (Blane and Jaakson, 1994), increased travel speeds (Kruse, 1991; Williams et al., 2002; Matsuda et al., 2011), increased active surface behavior (Trites and Bain, 2000); shorter surfacing times (Blane and Jaakson, 1994), increased feeding dive durations (Bass, 2000), and less direct and erratic traveling routes (Williams et al., 2002; Mattson et al., 2005; Schuler et al., 2019). In general, in our study, beluga behaviour differed in the experimental compared to control settings and this was observed for all age classes. Beluga behaviour also differed with age class in the presence of vessels. Group size in the presence of vessels was also significantly different from observations in the absence of vessels.
We hypothesized that whale-watching vessels might affect Calf group behaviour the most, but Calf groups occasionally interacted with vessels in all three years. Also, there is no clear evidence, based on our data, that there was an effect of vessel proximity on Calf groups. In fact, in the presence of boats, there was a noticeably high occurrence of interactions of those groups with vessels, which contradicts our initial hypothesis.
Regression results showed that the probability of Travelling and Feeding with Distance regressions were not consistently significant for a differing effect on Calf groups compared to the Adult reference group, depending on the factors used (Tables 8, 9). Nevertheless, it should be noted that, because of their small size and dark colour, calves are not as likely to be detected in a group at long distances, especially in the frenzied activity of a feeding group. It is conceivable that we could have obtained different results had we been able to confidently detect calves at long distances.
Based on prior observations of curious juvenile whales and anecdotal observations from whale-watching operators, it was hypothesized that juveniles would be most likely to interact with vessels or less likely to move away or stay away from the vessels. There appears to be no clear support for this hypothesis either. Juveniles were no more likely to interact with vessels than other groups classes. The low number of juvenile groups did not result in clear effects in the behaviour-distance regressions.
In summary, impacts of interactions are context specific in beluga groups of all classes, as other literature has suggested. Groups that were within the 0 to 25 metre observation zone spent an average of 63% of their time interacting with vessels (moving with the vessel, gliding under, rubbing against, and facing the prop wash). Interestingly, the similar study by Malcolm and Penner (2011), conducted 15 years prior, found that belugas spent less time, approximately 48%, interacting with vessels. Habituation of belugas to tour boats over the last two decades therefore seems quite likely.
Others have observed decreases in whale behavioural responses over time (Krasnova et al., 2020). Blane and Jaakson (1994) however, observed that although the majority of animals disturbed by whale-watching vessels returned to some pre-disturbance behaviour, feeding and travelling behaviours were not resumed. This does not appear to be the case in our study. Frequency of interactions between different age groups when close to an observation vessel show that all groups within the 0-25 m distance class interacted to a sizable extent with whale-watching boats. Calf groups had the lowest frequencies in two of the three years but those observations still accounted for a sizable proportion (≥ 25%) of their behaviour in the 0-25 m distance class. These results again contradict our initial hypothesis that females with calves would avoid the boats. Juvenile groups had interactions with the vessels as expected and so did Mixed groups which contain juveniles. Adult groups showed a lower probability of feeding near a vessel, but few observations of adult groups feeding were recorded at any distance and the effect could be largely be in part due to vessel operators standing back during a feeding event.
This research begins to address some of the gaps in our current understanding of beluga behaviour with respect to vessels and suggests that vessels may change beluga behaviour in the Churchill estuary; whether the overall effect is negative in the long term could not be determined with our study design. For other species, in different locations, there may be clear responses and consequences to human disturbances (Senigaglia et al., 2016), but that does not appear to be the case here with respect to vessels. Belugas in the Churchill estuary appear to have become habituated to the presence of tourism vessels over the last fifteen years, and interact frequently with them, however interaction times are limited (likely less than 20 min per 24 hour period). In addition, tourism vessels have adopted a code of conduct over the last decade which renders their activities more predictable to the whales, and a familiar part of their summer estuarine environment. If there are impacts, they could be more subtle than what can be isolated by these focal follows, or other recent studies (Ausen et al., 2022) and might require a detailed analysis of effects at the pod or individual level. Despite a dramatic decrease in tourism in 2020 due to the COVID-19 pandemic, as many as 17 boats (including kayaks which tended to be in large groups) were still seen at one time on the water. This research did not assess the potential effect of number of boats on beluga behaviour but could be included in future work as this may affect behaviour (Blane and Jaakson, 1994; Richardson and Wursig, 1997). Other observational methods may be required to document beluga behavioural time budgets in the estuary and compare them with observations in estuaries with no boat traffic. The effects of vessel noise also remain to be studied. Nonetheless, results of this research are useful for conservation and management planning. Exploring the response of cetaceans to human activity is important to understand larger ecosystem impacts and to predict the effect of future increased development and vessel activity in the North.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Ethics statement
Ethical review and approval was not required for the animal study because Animals in this study were observed in their natural environment only. No review was required.
Author contributions
KW designed the research, assisted in gathering and directing the field research, conducted the analysis and wrote the draft manuscript. PR made a significant contribution to the research design, direction of field research, analysis plan, analysis, and the writing of this manuscript. JS and EG assisted in gathering and directing the field research. SF provided guidance on research design and comments throughout the writing process. All authors contributed to the article and approved the submitted version.
Funding
Survey data was collected and analysis was funded by Oceans North Canada.
Acknowledgments
Thank you to Danielle Chiasson, Ben Verzijlenzerg and Terry Palmer in 2020, and Joni Reimer and Jordan Stewart in 2021 for field work assistance including behavioural observations and data entry. A special thanks to the Churchill Northern Studies Centre for logistical and technician support and to Sea North Tours for accommodating researchers on daily tours. And finally, special thanks to Dr. Cortney Watt and Dr. Attahiru Alfa for statistical support.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
(2018). Regulations amending the marine mammal regulations: SOR/2018-126. Can. Gazette Part II 152, 14. Available at: https://gazette.gc.ca/rp-pr/p2/2018/2018-07-11/html/sor-dors126-eng.html.
Ausen E. L., Marcoux M., Chan W. S., Barber D. G. (2022). Beluga (Delphinapterus leucas) response to personal watercraft and motorized whale watching vessels in the Churchill river estuary. Front. Mar. Sci. 9. doi: 10.3389/fmars.2022.837425
Bass J. (2000). Variations in grey whale feeding behavior in the presence of whale-watching vessels in clayoquot sound (University of Victoria: 1993–1995 [Unpublished PhD Dissertation]).
Blane J. M., Jaakson R. (1994). The impact of ecotourism boats on the St. Lawrence beluga whales. Environ. Conserv. 21 (3), 267–269.
Brown Gladden J. G., Ferguson M. M., Clayton J. W. (1997). Matriarchal genetic population structure of north American beluga whales delphinapterus leucas (Cetacea: Monodontidae). Mol. Ecol. 6, 1033–1046. doi: 10.1046/j.1365-294X.1997.00275.x
Brown Gladden J. G., Ferguson M. M., Friesen M. K., Clayton J. W. (1999). Population structure of north American beluga whales (Delphinapterus leucas) based on nuclear DNA microsatellite variation and contrasted with the population structure revealed by mitochondrial DNA variation. Mol. Ecol. 8, 347–363. doi: 10.1046/j.1365-294X.1998.00559.x
Chambault P., Kovacs K. M., Lydersen C., Shpak O., Teilmann J., Albertsen C. M., et al. (2022). Future seasonal changes in habitat for Arctic whales during predicted ocean warming. Sci. Adv. 8 (29), 1–9. doi: 10.1126/sciadv.abn2422
COSEWIC (2004). COSEWIC assessment and update status report on the beluga whale delphinapterus leucas in Canada (Ottawa: Committee on the Status of Endangered Wildlife in Canada) 70.
COSEWIC (2020). COSEWIC assessment and status report on the Beluga Whale Delphinapterus leucas, Eastern High Arctic - Baffin Bay population, Cumberland Sound population, Ungava Bay population, Western Hudson.
Guarino M.-V., Sime L. C., Schroeder D., Malmierca-Vallet I., Rosenblum E., Ringer M., et al. (2020). Sea Ice-free Arctic during the last interglacial supports fast future loss. Nat. Climate Change 10, 928–932. doi: 10.1038/s41558-020-0865-2
Hauser D. D. W., Laidre K. L., Stern H. L. (2018). Vulnerability of Arctic marine mammals to vessel traffic in the increasingly ice-free Northwest passage and northern Sea route. PNAS 115 (29), 7617–7622. doi: 10.1073/pnas.1803543115
Johnston M., Dawson J., De Souza E., Stewart E. J. (2017). Management Challenges for the fastest growing marine shipping sector in the Arctic: pleasure crafts. Polar Rec. 53 (1), 67–78. doi: 10.1017/S0032247416000565
Krasnova V. V., Prasolova E. A., Belikov R. A., Chemetsky A. D., Panova E. M. (2020). Influence of boat tourism on the behaviour of solovetskiy beluga whales (Delphinapterus leucas) in Onega bay, the white Sea. Aquat. Conservation: Mar. Freshw. Ecosyst. 2020, 1–12. doi: 10.1002/aqc.3369
Kruse S. (1991). “Dolphin societies: discoveries and puzzles,” in The interactions between killer whales and boats in Johnstone strait, B.C. Eds. Pryor K., Norris K. S. (Berkeley, CA: University of California Press), (pp.149–159).
Lien J. (2001). The conservation basis for the regulation of whale-watching in Canada by the department of fisheries and oceans: A precautionary approach. can. tech. rep. fish. Aquat. Sci. 2363, vi + 38 p.
Malcolm C. D. (2008). The current state and future prospects of whale-watching management, with special emphasis on whale-watching in British Columbia, Canada [Unpublished PhD dissertation]. Univ. Victoria.
Malcolm C. D., Penner H. C. (2011). “Polar tourism: Human, environmental and governance dimensions,” in Behaviour of belugas in the presence of whale-watching vessels in Churchill, Manitoba and recommendations for local beluga-watching activities. Eds. Maher P. T., Stewart E. J., Luck M. (Elmsford: Cognizant Communication Corporation), 54–79.
Matsuda N., Shirakihara M., Shirakihara K. (2011). Effects of dolphin-watching boats on the behaviour of indo-pacific bottlenose dolphins off amakusa-shimoshima island, Japan. Nippon Suisan Gakkaishi. 77 (1), 8–14.
Matthews C. J. D., Watt C. A., Asselin N. C., Dunn J. B., Young B. G., Montsion L. M., et al. (2017). Estimated abundance of the Western Hudson bay beluga stock from the 2015 visual and photographic aerial survey. DFO can. Sci. Advis. Sec. Res. Doc. 2017/061. v, + 20 p.
Mattson M. C., Thomas J. A., St. Aubin D. (2005). Effects of boat activity on the behaviour of bottlenose dolphins (Tursiops truncates) in waters surrounding Hilton head island, south Carolina. Aquat. Mammals 31 (1), 133–140. doi: 10.1578/AM.31.1.2005.133
Notz D., SIMIP Community (2020). Arctic Sea ice in CMIP6. Geophysical Res. Lett. 47 (10), 1–11. doi: 10.1029/2019GL086749
Pizzolato L., Howell S. E. L., Dawson J., Laliberte F., Copland L. (2016). The influence of declining sea ice on shipping activity in the Canadian Arctic. Geophys. Res. Lett. 43 (23), 146–154. doi: 10.1002/2016GL071489
Richard P. R. (2005). An estimate of the western Hudson bay beluga population size in 2004. Can. Sci. Advisory Secretariat Res. Document 2005/017, 29p.
Richardson J. W., Wursig B. (1997). Influences of man-made noise and other human actions on cetacean behaviour. Mar. Fresh. Behav. Physiol. 29, 183–209. doi: 10.1080/10236249709379006
Schuler A. R., Piwetz S., Di Clemente J., Steckler D., Mueter F., Pearson H. C. (2019). Humpback whale movements and behaviour in response to whale-watching vessels in Juneau, AK. Front. Mar. Sci. 6, 710. doi: 10.3389/fmars.2019.00710
Senigaglia V., Christiansen F., Bejder L., Gendron D., Lundquist D., Noren D. P., et al. (2016). Meta-analyses of whale-watching impact studies: comparisons of cetacean responses to disturbance. Mar. Ecol. Prog. Ser. 542, 251–263. doi: 10.3354/meps11497
Trites A. W., Bain D. E. (2000). Short-term and long-term effects of whale-watching on killer whale (Orcinus orca) in British Columbia (Vancouver, BC: University of British Columbia) 10 pp.
Westdal K. H., Davies J., Ferguson S. H. (2022). Assessing seasonal and spatial segregation by age class of beluga whales (Delphinapterus leucas) in Western Hudson bay estuaries. PLoS One 17 (11), e0255756. doi: 10.1371/journal.pone.0255756
Keywords: behavioural response, beluga, vessels, tourism, Hudson Bay
Citation: Westdal KH, Richard PR, Sportelli J, Gillis E and Ferguson SH (2023) Beluga whale (Delphinapterus leucas) behaviour in the presence of whale-watching vessels. Front. Mar. Sci. 9:891003. doi: 10.3389/fmars.2022.891003
Received: 07 March 2022; Accepted: 05 December 2022;
Published: 06 January 2023.
Edited by:
Vitor H. Paiva, University of Coimbra, PortugalReviewed by:
Candice Emmons, National Oceanic and Atmospheric Administration (NOAA), United StatesEvan Richardson, Environment and Climate Change Canada (ECCC), Canada
Copyright © 2023 Westdal, Richard, Sportelli, Gillis and Ferguson. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Kristin H. Westdal, a3dlc3RkYWxAb2NlYW5zbm9ydGguY2E=