Abstract
Since the advent of recreational and scientific SCUBA diving in the 1950s, coral reefs have become one of the most studied and appreciated marine ecosystems. Direct human observations have been essential to many scientific discoveries. However, most in-water experiences are limited to short durations (tens of minutes to ~1 hour), shallow depths (surface to ~30 m), and calm sea conditions, leaving much of the reef environment underexplored—particularly deeper zones (to ~80 m) and with periods of challenging environmental conditions undersampled. Over a 20-year period, we employed research approaches centered on extended human observation through advanced modes of scientific diving, combined with in situ instrumentation. These efforts revealed extensive heterogeneity in reef physical conditions driven by internal waves, varying across time (minutes to seasons) and space (meters to 100s of km). Technological advances in autonomous sensors, microprocessors, and memory allowed for continuous, distributed, and high-resolution environmental sampling and 3D data visualization over time. Our work was enabled by enriched oxygen (nitrox) and helium-oxygen (trimix) technical diving, as well as saturation diving supported by the U.S. National Undersea Research Center and Aquarius Reef Base in the Florida Keys. Early direct observations of rapid changes in temperature, salinity, and current flow—and associated behavioral responses of reef organisms—led to new understanding of internal wave dynamics, including the formation of bores or “internal surf,” as persistent features of reef environments. These observations prompted further studies into nutrient and plankton transport, and revealed trophic connections between shallow reefs and deeper macroalgal communities (~50–80 m). Targeted deployments of high-resolution instruments helped characterize small-scale variability and link local dynamics to regional internal wave forcing by the Florida Current. A late season tropical cyclone arriving during a saturation diving mission in 1994 further highlighted extreme reef environmental dynamics rarely observed directly by humans. Together, these findings show how direct human observations, combined with continuous environmental monitoring, are essential for understanding complex reef processes—offering insights into ecosystem connectivity and coral thermal refugia under climate change.
Introduction
The long history of human interactions with coral reefs includes reliance on food resources, protection of coastal and island habitats, navigational guides and hazards to navigation for pan-island Pacific peoples and during the age of European sailing exploration (Dobbs, 2009), and more recently marine science and conservation in a new era of global human impacts on reef environments (Bellwood et al., 2019). At least since the work of Darwin and other European naturalists, coral reef formation was considered from geological perspectives with the view of reefs forming over long periods of time through continuous calcification and coral growth, with gradual accumulation of the substratum facilitated by occasional wave disturbance producing fine materials cemented into the matrix, all occurring in a context of tectonic subsidence, and occasional uplift (Darwin, 1842; Yonge, 1963; Stoddart, 1976; Dobbs, 2009). These early efforts and more recent and contemporary scientific efforts have been influenced by human perceptions of coral reef ecosystems as seemingly quiescent environments, with limited thermal and other environmental variability, and appearing to develop slowly under stable conditions with only occasional large disturbances caused by intense, but locally rare, storms including hurricanes and cyclones (Goreau, 1959; Yonge, 1963; Connell, 1978; Wing and Leichter, 1997). By contrast, more recent understanding of physical and biological processes in reef environments have revealed and underscored the extensive, and sometimes rapid fluctuations in conditions also inherent to reef environments.
Our research on coral reefs in the Florida Keys began in the early 1990s associated with a research initiative of the U.S. National Oceanic Administration’s National Undersea Research Program (NOAA NURP) to develop a focus for coral reef research at a long-term site for the Aquarius Habitat saturation diving platform at Conch Reef, Florida Keys, USA (24.983° N, 80.416° W) (Figure 1). The initial motivation for a project led by M. Patterson (then of UC Davis) was to investigate effects of variation in water velocity at the reef surface on diffusional boundary layers and coral photo-physiology linked to coral growth and observations of coral bleaching. A goal of the initial work was to gather information on current velocities and hydrographic conditions at the Conch Reef site across depth gradients from ~10 to 30 m depth, and through time in conjunction with the plans for a long-term research program and deployment of the Aquarius Habitat, later termed the Aquarius Reef Base (Benjamin, 2021). In the following examples from that initial project and multiple subsequent endeavors we illustrate how direct human observations, often made possible by incremental advances in diving capabilities, were combined with instrumentation and more detailed biological and physical sampling to build a multi-factorial view of the underlying physical dynamics. In all cases, due to the highly dynamic oceanographic conditions at this site, we found previously unknown levels of environmental variability for a coral reef. We then built upon the observations and new understanding of the highly variable physical environment to investigate the biological and ecological impacts of the environmental forcing. We highlight how access to scientific diving techniques enabling enhanced bottom times and extending the depth of investigation were central to direct observations. Direct observations were enhanced by developments of in situ instrumentation for continuous collection of key environmental data and analyses aimed at identifying the specific driving mechanisms of the environmental variability.
Figure 1
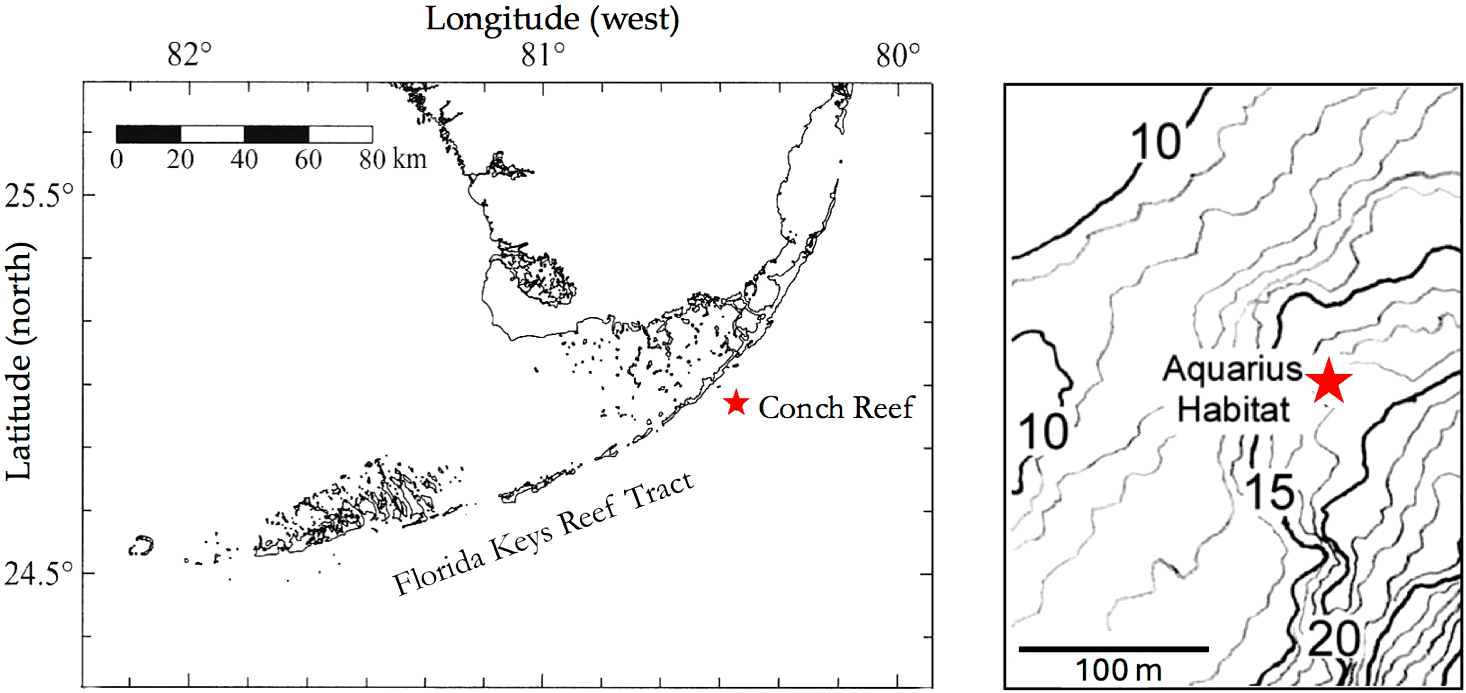
The Florida Keys Reef Track and bathymetry at the site of the Aquarius Habitat Reef Base.
One motivation for this sequence of projects was that in the early 1990s there was limited prior evidence of the high levels of oceanographic variability and dynamic physical conditions that could be observed on coral reefs. This was especially true of the then sparse and short-duration observations of internal waves impacting coral reefs (Wolanski and Pickard, 1983; Wolanski and Delesalle, 1995). Our initial work (Leichter et al., 1996) led to growing understanding that turbulence and mixing resulting from breaking internal waves could play significant roles in redistributing materials from the water column offshore to the temporally and spatially variable ecosystems on the reefs (Leichter et al., 1996, 1998, 2003).
The combination of developments in diving approaches, microelectronics and sensor technology, and a vision within the NOAA National Undersea Research Program for sustained competitive funding opportunities to support field scientists working underwater led to approximately two decades of scientific discovery. Thus, from our initial goals of measuring physical conditions, primarily water temperature and near bottom flow conditions likely to contribute to spatial heterogeneity of coral bleaching and adaptation to bleaching susceptibility, we were able to combine direct observations and instrumentation to reveal previously unknown dynamic conditions not just on Conch Reef, but throughout the Florida Keys reef tract and beyond in other reef systems. Many of the processes discovered through our work subsequently motivated research programs showing similar dynamics to be widespread on coral reefs at global scales across ocean basins. The results have become highly relevant in the current context of ocean climate change and anthropogenic stresses affecting coral reefs.
Observations of internal waves at Conch Reef
Ocean internal waves – gravity waves occurring within water columns that are stratified with respect to the vertical density distribution – were known from the 1940s (Lamb, 1945; Sverdrup et al., 1942; Ewing, 1950) and studied extensively by physical oceanographers over subsequent decades (e.g. see Pond and Pickard, 1983; Baines, 1986; Holloway, 1991; Lamb, 2014). Internal waves can form, propagate and subsequently break in density stratified water columns by a variety of complex mechanisms (Lamb, 2014). A geographically widespread mechanism of formation is the displacement of isopycnal layers by tidally-varying currents in the vicinity of abrupt bathymetric features such as underwater ridges, seamounts, and shelf breaks. After propagating into shallow systems near coast or islands, internal wave breaking dynamics become complex and strongly influenced by the extent of water column density stratification and slope angles of the local seafloor (Lamb, 2014). In regions of critical slope angle relative to the strength of density stratification, internal wave breaking can lead to strong intensification of near-bottom currents, with resulting sediment resuspension and potential erosion of the seabed (Cacchione and Drake, 1986; Cacchione et al., 2002; Lamb, 2014). However, while internal waves continue to be extensively studied, especially by physical oceanographers, in the early 1990s knowledge of internal wave impacts on coastal ecosystems was almost entirely from temperate locations with strong seasonal water column stratification (e.g. Sandstrom and Elliott, 1984; Pineda, 1991). Only a few short-term observations had been made of internal waves on coral reefs at the margin of the Coral Sea and at Tahiti (Wolanski and Pickard, 1983; Wolanski and Delesalle, 1995). Most direct experiences by scientists using surface and SCUBA diving on coral reefs indicated environments characterized by warm and stable thermal conditions, limited water column thermal and density stratification, and relatively constrained seasonal variation in temperature between approximately 22 to 30°C (Jokiel and Coles, 1977). The existence of internal waves impacting coral reefs was largely unknown in the 1990s, most likely due to limited continuous observations, and perhaps also due to inaccurate impressions by scientists working in shallow water, close to reefs, that water column thermal and density stratification would be limited in tropical settings.
In general, corals are known to be stenotopic – having a narrow range of adaptability to changes in environmental conditions – especially compared to organisms in other extensively studied shallow water marine environments such as temperate rocky intertidal habitats (e.g. Jokiel and Coles, 1977, 1990). Associated with the limited ability of corals to adapt to high temperatures, from the mid 1980s on there was growing recognition of widespread and frequently lethal coral bleaching in response to elevated water temperatures, e.g. T > 30°C, (reviewed in Brown, 1997, but without recognition of high frequency thermal events). Some investigations had also started to focus not just on static temperatures but on hydrodynamics and bottom boundary layer dynamics impacting the flux of heat, and other environmentally relevant variables such as dissolved oxygen, over coral colonies as potentially important factors influencing and perhaps mitigating the effects of high temperatures on coral physiological stress (e.g. Patterson et al., 1991; Patterson, 1992).
Our initial direct observations were facilitated by extended bottom times associated with two developments in SCUBA technology: the use of 36% oxygen “nitrox” gas mixtures as well as the growing ability to rely not only on traditional dive tables but on reliable dive computers capable of calculating safe, multilevel, non-decompression dive profiles. For example, dive times on 36% nitrox could be in excess of 60 min at 15 to 20 m depths and approximately 30 to 40 min at 25 to 30 m depths. Subsequently, a series of saturation diving campaigns utilizing the Aquarius Habitat provided greatly extended diver excursion times, approaching 9 hours of excursion time per diver per day for a team of 4 divers and with depths to 40 m easily attainable within swimming distance from the Aquarius base. Also important in our ability to detect and interpret the impacts of internal waves were increasing availability and decreasing costs of stand-alone, housed data logging environmental sensors, especially temperature sensors with capabilities of sampling at 1 to 10 min intervals and recording data for multiple months while costing < US $200 including compact individual pressure housings. By coupling the direct observations to instrument deployments we were able to confirm that rapid changes in water temperature on the reef slope were recurring, periodic events. The advent of relatively low-cost instruments was a key element of the research by enabling our ability to deploy not just individual sensors but arrays of synchronized, individually recording, temperature recorders (e.g. Ryan Tempmentors, and Onset Hobo Stowaway loggers) across depths from 10 to 35 m on the reef slope. By modifying the thermistors of these sensors to be in direct contact with fast response stainless steel pins we were able to increase the response time for the sensors from minutes to a few seconds. These deployments during our Aquarius Habitat expeditions were further augmented by access to recording Conductivity-Temperature-Depth (CTD) sensors, recording current meters (InterOcean S4 electromagnetic current meters) and acoustic Doppler current profilers (RDI ADCPs) many of which were made available via shared resources of the NOAA program (see Figure 2). The collection of continuous data over months to multiple years then facilitated the important analytical capability to apply basic time series and signal processing analyses to multi-season temperature records. The work was also facilitated by the growing availability in the mid 1990s of low-cost computers with increasing processor capabilities and increasing availability of computer languages and scripting packages such as C+, Systat, and Matlab. Analyses revealed clear patterns of periodicity in the fluctuating temperature and water current velocity time series. Specifically, the relative ease of calculating and plotting power spectra allowed us to delineate statistically significant peaks at semi-diurnal frequencies indicative of tidal-period forcing associated with incident internal wave fields, and showed predictable return times for temperature change events (Leichter et al., 1996; Leichter and Miller, 1999). The collection of continuous long-term data and the application of spectral analysis were critical to resolving how and why the temperature changes directly observed by divers swimming over the reef were variable on longer time frames. The initial time series also revealed an important and surprising observation of increasing magnitude of variation with greater cooling and increasing temperature variation with increasing depth from 10 to 30 m, as well as marked variation in internal wave forcing across seasons and even within the summer season of greatest variability.
Figure 2

(A) Schematic view of instrumentation approach to measure changes in temperature, salinity and currents near the reef surface and through the water column at Conch Reef, Florida Keys, USA, site of the Aquarius Habitat (B). (C) Acoustic Doppler Current Meter, (D) early design of individual recording temperature sensors in customized pressure housings, (E) Conductivity-Temperature-Depth sensor deployed immediately below an electromagnetic current meter.
Striking features of our initial diver observations at Conch Reef were the strong and variable currents especially in the along-shore direction, and particularly occasional bouts of strong near bottom flows in the cross-shore, up-slope direction corresponding to dramatic step-like declines in temperature and increases in salinity. The upslope flows were often associated with rapid, drops in temperature that could be directly felt by divers as well as observed as the relatively dense water transported by internal bores diffracted light resulting in shimmering density discontinuities and blurring of visual images. The temperature fluctuations were quantified as ~2 to 5°C drops in temperature in continuous data recorded by sensors on the reef surface distributed across depths on the reef slope (Leichter et al., 1996) (See Figure 3). Power spectra calculated on time series data at the surface and at depths of 10, 20, and 30 m on Conch Reef consistently showed differences among seasons and depth, with prominent peaks at semi-diurnal frequencies with greatest magnitudes at the deepest depths and in the summer seasons when water column stratification is greatest (see also Leichter et al., 2014). These direct observations, coupled with examination of temperature and salinity relationships for the offshore water column clearly showed that subthermocline water was arriving on the reef slope and likely transported upwards from deeper depths onto the reef slope (Leichter et al., 1996). On numerous occasions we also directly observed near-bottom flows of cool water with marked decreases in visibility as water was moving in the onshore direct near the reef surface.
Figure 3
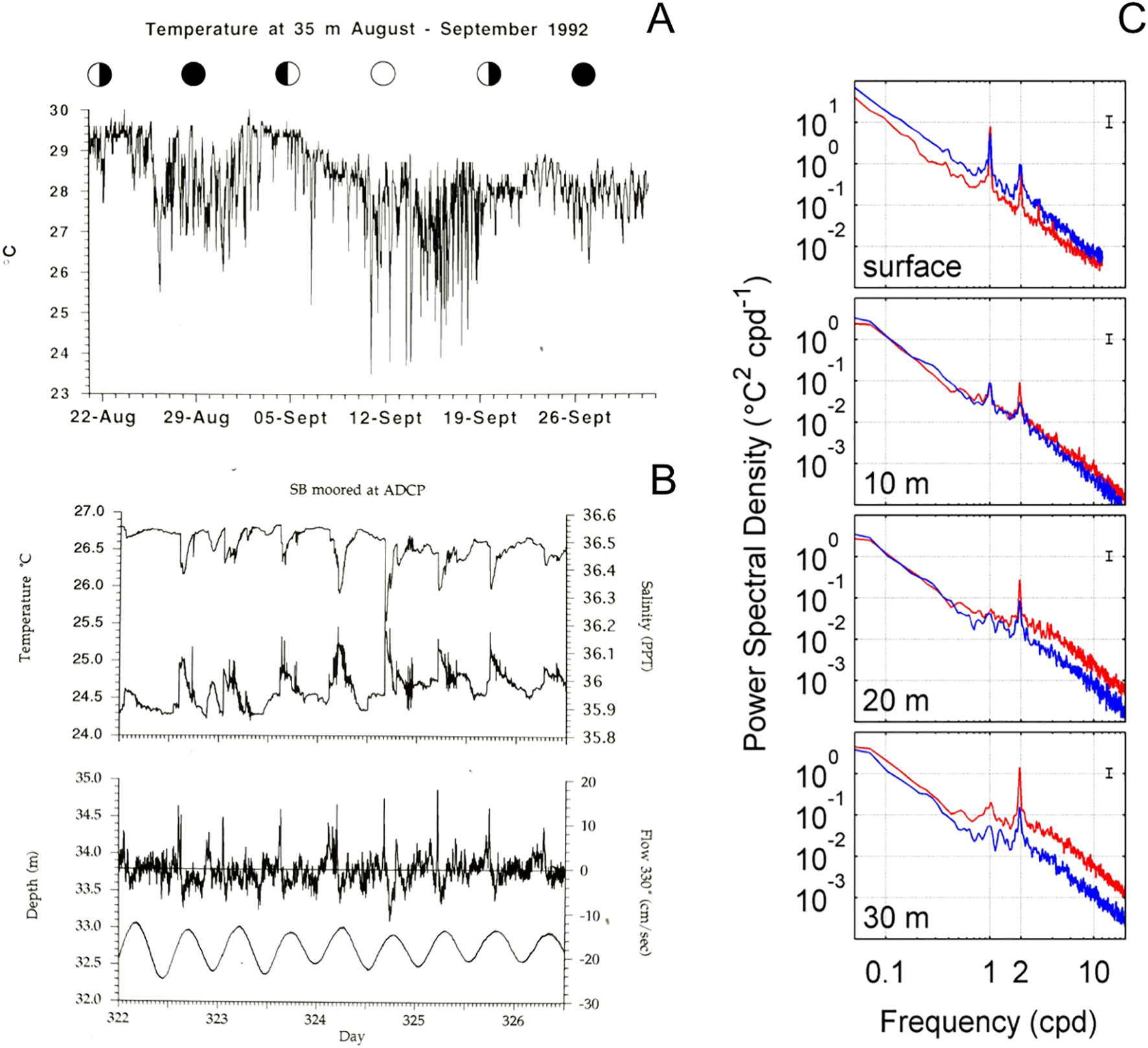
(A) First measurements of temperature variability caused by internal waves at Conch Reef 35 m depth. (B) Evidence of co-occurring changes in temperature, salinity, and upslope flow (positive velocity at 330°), and tidal depth. Data adapted from Leichter et al., 1996. (C) Power spectra for temperature data at the surface, at 1 m above the bottom at 10 m, 20 m, and 30 m depths at Conch Reef. Spectra are shown in two seasons, summer May 1 – Sep 30 (red), and winter Oct 1 – Apr 30 (blue) for data collected from 1997 through 2006, adapted from Leichter et al., 2014.
The occurrence of increasing temperature variability marked by greater cooling with increasing depth, and of lags in the onset of cooling at successive positions up the reef slope indicated that the temperature changes were produced by moving fronts of cool water traveling up the reef slope from deeper to shallower depths. Overall, the combination of extended dive durations with direct human observations, particularly during saturation diving excursions, and the deployments of continuously recording instruments were critical to both directly observing and quantitatively measuring the changes in temperature, salinity, and near bottom velocities as internal bores formed by broken internal waves were traveling shoreward. Current meter data indicated that the variability was not just in temperature, but also in near bottom and water column flow speeds and flow directions (Leichter et al., 1996; Figure 4). The initial goals of collecting detailed environmental and hydrodynamic observations to investigate hypotheses around forced convection and mass transport limited systems led to an entirely new understanding of processes governing the physical environment at Conch Reef.
Figure 4
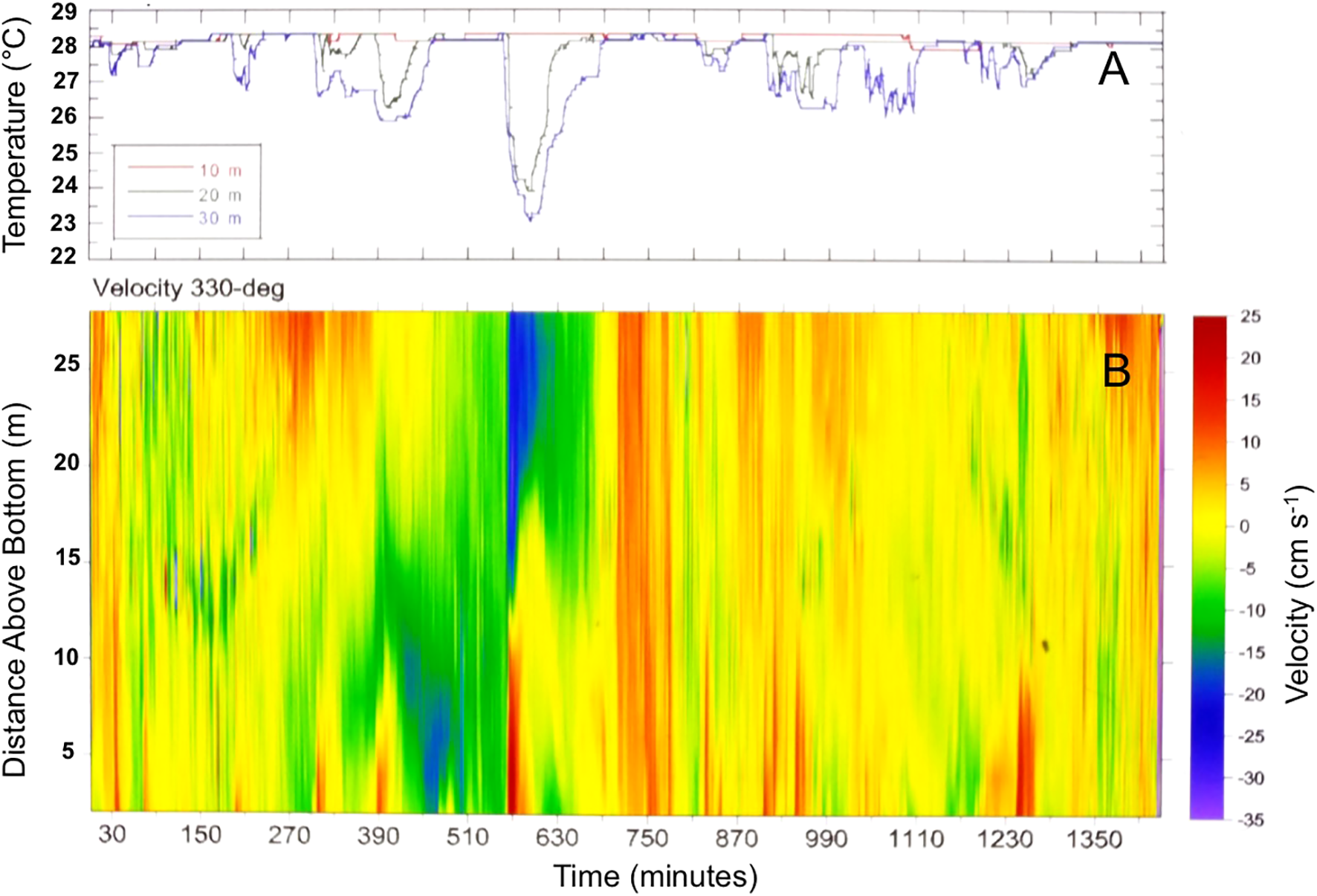
(A) Evidence of rapid decreases in temperature at the reef surface at 20, 30 m depth, with (B) surges of cross shore upslope flow (positive flows at 330°C) in the bottom 5 to 10 m of the water column measured by profiling acoustic current meter at 33 m depth. Data adapted from Leichter et al., 1996.
The experiences of seeing and feeling sustained flows of cold water onto a reef along were coupled with observations of the behavior of animals across levels of complexity from sponges spawning to soft and hard coral polyps reorienting in flows, and fish orienting and feeding at moving fronts (Figure 5). These strongly suggested that the observed internal waves were likely to have organismal to ecosystem consequences for the reef ecosystem. While the initial study was motivated by an interest in temperature and hydrodynamics and their potential impact on coral bleaching, the serendipitous observations made possible by being on the reef for extended periods of time led to new scientific appreciation of the importance of the physical forcing causing sustained cooling and associated strong upslope flow velocities. Simultaneous deployments of ADCPs and vertical temperature strings also revealed periods of downslope, return flow, of the water that had moved onshore and then subsequent reversals of the flow down the reef slope. Upslope flow velocities associated with the arrival of internal bores were measured to be ~10 to 30 cm s-1 (Leichter et al., 1996 and see Figures 3, 4). The ADCP data were also important in revealing shear within the water column with water moving opposite directions top and bottom of the ~ 30 m water column above the reef. Notably the availability of sophisticated ADCPs and vertical thermistor-strings were critical to measuring and interpreting dynamics that could not be directly observed only by divers.
Figure 5

Reef organisms observed to be affected by and respond to changing flow speeds and temperatures associated with internal waves on Conch Reef. (A, B) gorgonians, (C) Queen Angelfish, Holacantus cilaris, (D) Atlantic Spadefish, Chaetodipterun faberl, (E) Giant Barrel Sponge, Xestospongia muta (photographed in a 0.25 m2 quadrat).
Also notable and quite intriguing were observations of surface slicks corresponding to events on the bottom (personal observations). Bands of surface slicks in coastal waters are linked to internal waves and provided a surface manifestation and confirmation of the subsurface physical forcing impacting the reef slope. Thus, multiple types of evidence, ranging from direct human diver observations to time series instrument measurements across multiple iterations, were all critical to interpreting the environmental dynamics on the reef. The combination of direct human observations and autonomous sampling instruments provided far more than either approach could independently. The novelty of the observations in this environment provided the interest and subsequent funding to sample in greater detail especially with focus on biological consequences of internal waves at Conch Reef and beyond.
Highly variable plankton concentrations and ‘plankton storm’
Our initial direct visual observations as divers on the reef included rapid changes of visibility in the water column and the advance of dark water with high concentrations of particles and zooplankton that appeared to attract large numbers of zooplanktivorous fishes. On multiple occasions we observed shimmering density discontinuities moving upslope, with dense plankton concentrations, and fishes including Blue Chromis, Chromis cyaneus, Queen Angelfish, Holacantus ciliaris, and Atlantic Spadefish, Chaetodipterus faber, orienting to intercept the incident, moving fronts associated with internal bores. Divers described the experience to being in the underwater equivalent of a sudden snow storm – a “plankton storm” on the reef slope. While recording transmissometers can provide valuable time series on water column particle concentrations and cloudiness, these were not initially available, nor would we have been likely to anticipate the potential signal to sample without initial direct visual observations. Further, it would have been unlikely to make these direct observations of rapidly changing water clarity characteristics, the moving density interfaces, and changing fish behavior from instrumentation alone and without divers on the reef over periods of tens of minutes to hours. SCUBA diving provides opportunities for spatially extensive, adaptive visual observations in ways not available through instrumentation alone. The extended dive durations associated with saturation diving from the Aquarius Habitat allowed for up to 9 hours of dive excursion time per day. Thus we were able to observe and sample changes in plankton abundance repeatedly throughout multiple days.
Plankton dynamics on coral reefs have long been known to influence food availability to reef invertebrate suspension feeders and fishes (e.g. Glynn, 1973; Hobson and Chess, 1978; Gerber and Marshall, 1982; Erez, 1990). The dense aggregations of suspension feeders on reefs, termed a “wall of mouths” (Hamner et al., 1988) are dependent on supply of energy resources in the form of plankton and other suspected particles, with the linkage constituting a direct connection between water column production and benthic communities of coral reefs (Birkeland, 1997; Hatcher, 1997; Wyatt et al., 2010). In the water column seaward of reefs there can be marked increases in suspended chlorophyll-a concentration indicative of phytoplankton abundance at the subsurface chlorophyll maximum layer and we observed strong vertical stratification of the offshore water column seaward of the Florida Keys with a distinct subsurface chlorophyll-a maximum (see Figure 3 shown in Leichter et al., 1998). We therefore speculated that transport associated with internal wave bores could create a temporally variable link to shared offshore zooplankton and potentially planktonic larval pools implying potential for important meta-population connectivity between Florida Keys reefs and a large offshore region of the stratified pycnocline/thermocline (Leichter et al., 1998).
For the project reported in Leichter et al., 1998 we initially used CTD sampling from a surface boat to measure the water column temperature, salinity, density and chlorophyll-a structure from the reef crest to stations offshore. Having established the water column structure and location of the subsurface chlorophyll-a maxium layer at ~45 to 60 m depth offshore, we then focused hydrographic and biological sampling on the reef surface. Following directly from the initial observations and knowledge of the water column hydrographic and chlorophyll structure, we attempted to gather quantitative information on the transport of suspended particles including fish eggs, holoplankton such as copepods, and planktonic larvae onto the reef slope. Here again multiple direct human observations, facilitated by the long duration of dives on the reef slope provided both motivation and direct evidence. A combination of physical sampling equipment was deployed across the reef surface and the abundance and diversity of plankton was sampled by divers during a 10-day saturation diving mission (Figure 6). This sampling scheme, while unusual and certainly effort intensive compared to, for example, more traditional boat-based plankton net tows, provided for adaptive sampling very close to the reef surface during periods of cooler water transiently moved onto the reef surface by upslope flows associated with internal bores (Leichter et al., 1998). Here, the possibility for adaptive sampling in response to direct observations of changing environmental conditions proved a great advantage of saturation diving.
Figure 6
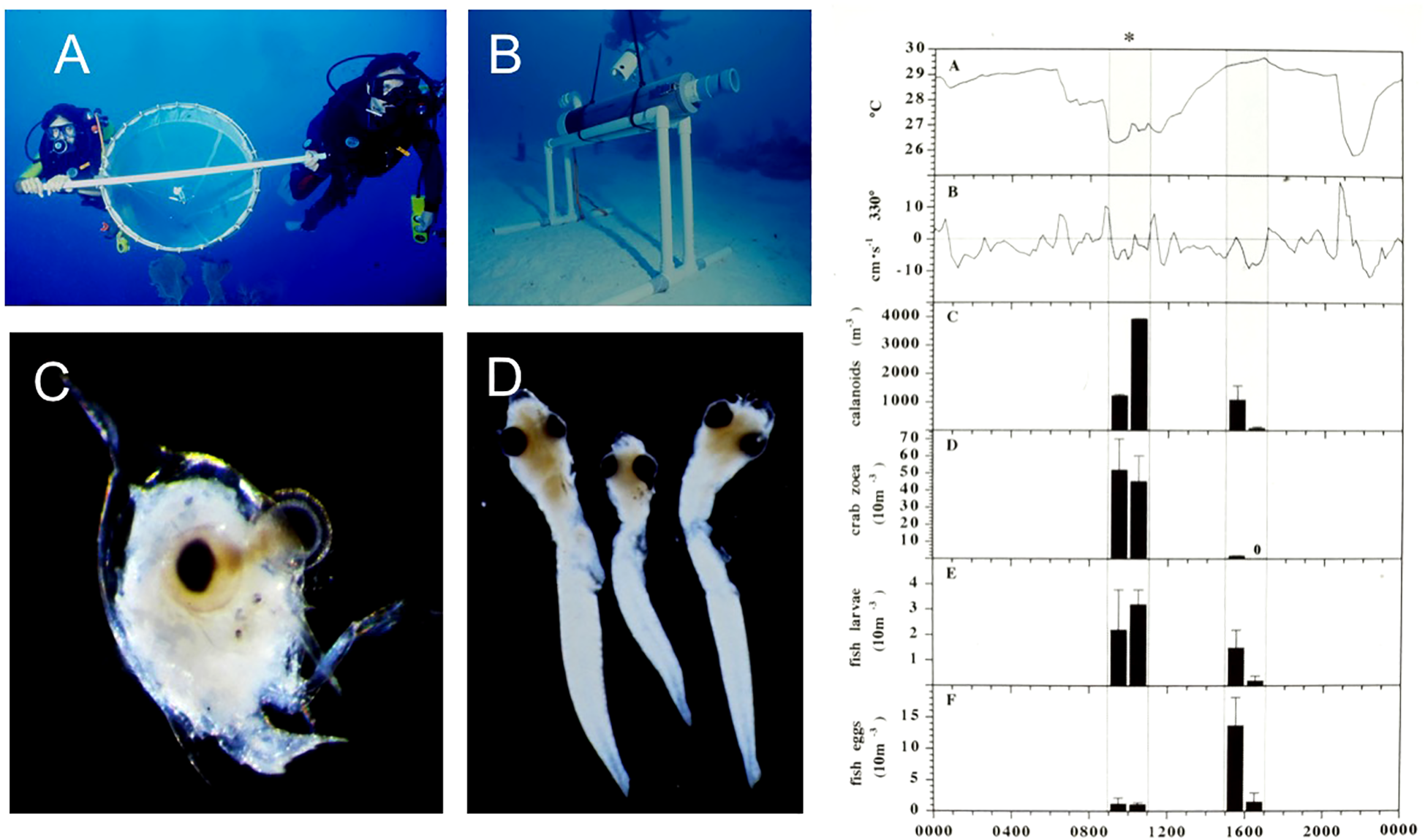
Efforts to sample zooplankton near the reef surface by (A) divers swimming a 1 m2 plankton net, (B) plankton pumps powered from the Aquarius Habitat. (C) Crab zooea and (D) fish larvae found in significant higher abundance during cool water and onshore flow periods associated with internal waves. Zooplankton data (right column) adapted from Leichter et al., 1998.
During 6 days in August 1995 we conducted swimming diver plankton ‘tows’ in the morning and evening while temperature recorders, ADCPs, and CTDs sampled physical conditions. This focused sampling and subsequent quantification of the resulting plankton samples revealed multiple order of magnitude differences in zooplankton concentrations, especially for calanoid copepods and fish and crab larvae that were higher associated with the upslope flow of cool water than the ambient warmer water conditions on the reef outside of the times of internal bore arrival (Leichter et al., 1998). On multiple occasions divers noted rapid decreases in water column visibility, high concentrations of zooplankton sufficiently large to be visible to the naked eye, as well as large aggregations of fish feeding at the moving internal bore front. On one occasion while swimming several hundred meters out to the research site from the Aquarius Habitat, divers also noted mass spawning of the Giant Barrel Sponge, Xestospongia muta in conjunction of the arrival of cool water internal bores on the reef slopes.
To examine whether the transport of zooplankton over the reef surface might impact growth of suspension feeding corals we also conducted a coral growth rate experiment in which replicate branch tips of the suspension feeding pocilloporide coral Madracis mirabilis (sensuWells, 1973 but later reclassified M. aurentenra by Locke et al., 2007) were positioned on racks at multiple depths across the reef slope. Coral growth was estimated from change in skeletal mass before and after the 15 month experiment and revealed a pattern of highest growth at 30 m relative to growth at 20 or 15 m depth (Figure 7). The resulting pattern suggested that provisioning of plankton associated with greater incidence and duration of internal bores at the deeper sites on the reef could offset the effect of decreased light availability with depth for this suspension feeding coral and perhaps generally for suspension feeders in mesophotic reef environments (Leichter et al., 1998). In a subsequent project we conducted at a site on the north shore of Jamaica also impacted by internal waves, we found a similar pattern of enhanced growth of suspension feeding M. aurentenra (neé mirabilis) with increasing depth (Leichter and Genovese, 2006). Our sampling of plankton variability and coral growth pointed to multiple ways in which internal waves impacting reef slopes can modify currents, temperature, dissolved nutrients, and particle concentrations all influencing reef organisms. The observations of highly variable plankton concentrations also suggested internal waves might influence fish and invertebrate recruitment dynamics. Specifically, periods of enhanced internal wave activity could result in quasi-synchronous plankton and larval availability among sites across the Florida Keys (see Sponaugle et al., 2005).
Figure 7
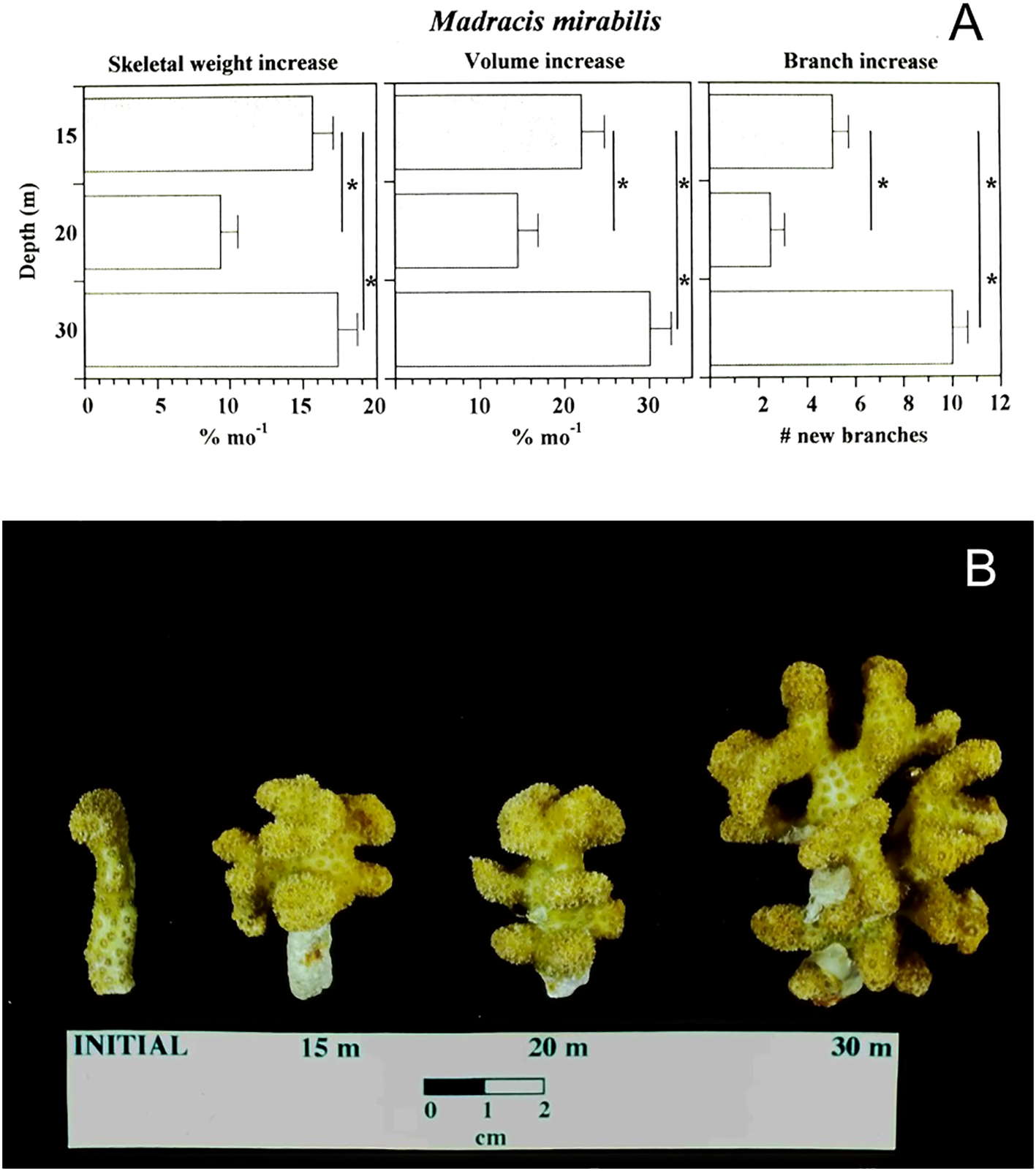
(A) Growth of the suspension feeding coral Madracis mirabilis (later suggested to be M. aurentenra) across depths in a 16-month growth rate experiment. Bars represent within-depth means and standard errors. ( * ) Significant differences at p < 0.05. (B) Photograph of representative individual coral nubbins at initial and final size across depths 15, 20, and 30 m. Data adapted from Leichter et al., 1998.
Nutrient supply to the reef slope and deeper water benthic algae
One impetus for our research into internal waves at Conch Reef was to better understand natural variation in nutrient dynamics as an important context for evaluating potential anthropogenic sources of nutrients associated with agricultural runoff, coastal development and rapid growth in human populations in the Florida Keys (Lapointe 1997; Leichter et al., 2003). This motivated another project that combined adaptive diver sampling made possible by saturation diving from the Aquarius Habitat, with real-time environmental sampling of temperature and hydrographic conditions we were able to measure directly the fluxes of major dissolved nutrients, nitrate and phosphate, in relation to internal bores on the reef slope. Hydrodynamic and nutrient conditions varied markedly during the sampling period which was timed to correspond with seasonal periods of high internal wave activity in long-term time series data from the reef. We found that nutrient fluxes (the product of concentrations and flow speed) increased by up to 40 fold during internal bore events compared with warmer, non-bore periods (Leichter et al., 2003). The nutrients exhibited isotopic signatures of the deep water nutrient pool offshore below the thermocline (Leichter et al., 2007) and exposure times to cool water were significantly correlated with tissue nutrient concentrations in branching alga Codium isthmocladum sampled across depths on the reef slope. An additional surprising direct observation associated with the arrival of cool water internal bores on the reef slope was the transport onto the reef of large fragments and at times whole blades of benthic algae not found growing in the shallow coral reef environment (Figure 8). These direct and fortuitous observations, which again were only possible by having divers in the water then led to an investigation of water column and benthic processes in deeper water and offshore of the reef tract. In a ship-based project of intensive hydrographic sampling throughout the Florida Keys we were able to confirm high concentrations of dissolved nutrients, especially nitrate and phosphate in the offshore subthermocline waters and marked changes in nutrient concentrations on the reef slope in water samples hand collected by divers with syringes in response to temperature change events (Leichter et al., 2003) and with distinct isotopic values of Δ15N that could be used to infer sources of nutrients supporting algal growth on the reef slope (Leichter et al., 2003). Subsequently we also endeavored to sample macro algal communities both on the reef and offshore at bottom depths of 40 to 80 m.
Figure 8

Examples of large macro algae found at depths of 30 to 40 m seaward of the reef slope at Conch Reef and observed as fragments transported onto the reef slope by internal bores. (A)Kallymenia sp. (B, C)Codium spp. (D) Schools of Atlantic Blue Tang, Acanthurus coeruleus exhibiting positive rheotaxis while foraging on macroalgae fragments at 30 to 40 m depth seaward of the reef slope.
An essential component of our work on algal communities in deeper water was NURC’s development of deeper diving training and research capabilities based on open circuit “trimix” decompression diving. During a series of field campaigns, we utilized an 18% oxygen, 50% helium, 32% nitrogen breathing gas mixture to access research sites at 40 to 80 m depths seaward of reef slopes in the upper Florida Keys (Leichter et al., 2008) (Figure 9). Photographic and diver hand collections identified abundant and diverse assemblages of macroalgae with 26, 23, and 9 identified taxa of chlorophytes, rhodophytes, and phaeophytes respectively (Leichter et al., 2008). The capability of scientific diving at significantly greater depths than previously feasible allowed biological sampling of species composition and abundance and revealed extensive benthic algal communities seaward of the reef slope. Typical total dive times were on order of 90 minutes with 25 minutes of time available for work on the bottom at 40 to 80 m depth. In addition to revealing zones of high macroalge abundance at the seafloor, the research suggested a further trophic linkage to the coral reef community in the form of fish and invertebrate herbivory of algae material transported on to the reef slope. The ability to dive to ~80 m and to relocate specific sites on the seafloor allowed for deploying and later resampling settlement blocks for macroalgal recruitment, and revealed patterns of increasing algal recruitment with depth (Leichter et al., 2008).
Figure 9

Efforts to sample benthic algae at 50 to 80 m depth utilizing open circuit “Trimix” (A-E), high abundance of benthic algae at 60 m depth sample photographically, images adapted from Leichter et al., 2008.
As part of another Aquarius Habitat project in 2003, we deployed a network of ~100 synchronized temperature sensing nodes to map heterogeneity of internal wave exposure at Conch Reef. The array was deployed using SCUBA and the locations of all nodes were carefully mapped on the reef surface during a 10-day saturation diving mission. The sensor network array and deployed over an area of roughly 180 x 50 m and ranging in depths from 10 to 30 m and recorded at 10 s intervals from May through August 2003. The resulting data revealed an unprecedented picture of the time varying structure of temperatures on the benthos (Leichter et al., 2005), and data were also used in analyses of spatial variation in algal isotopic composition again related to internal wave exposure and transient delivery of subthemocline water and nutrients onto the reef surface (Stokes et al., 2011; Stokes et al., 2020) (Figure 10).
Figure 10
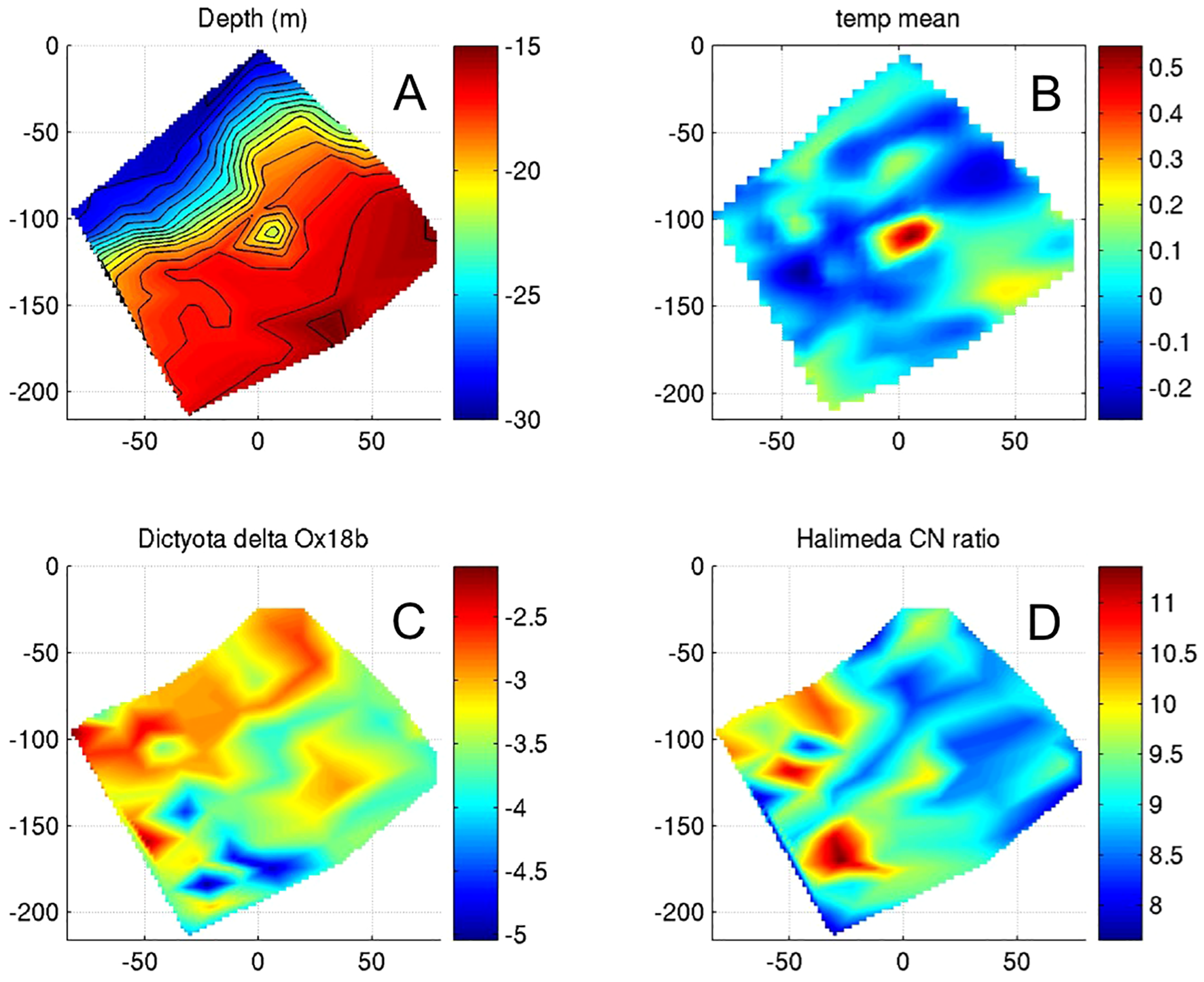
Results of sampling spatial variability in internal wave impacts on Conch Reef via distributed, synchronous sampling array. (A) depth, and (B) mean temperature (color scales) across temperature array. Oxygen isotopic signatures and carbon: nitrogen ratio of benthic algae Dictyota (C), and Halimedia (D). X and Y axes are distance (m). Adapted from Leichter et al., 2005, and Stokes et al., 2011.
Florida Keys-wide variability, Florida current eddies, storm impact
Our research from 1997 through 2006 included collecting synchronous near-bottom temperature data at six sites throughout the Florida Keys. This led to an ability to detect the timing of events occurring at a regional scale on the Florida Keys reef tract. This is a ~180 km span of discontinuous reefs spanning the zones from Key West in the southwest to Carysfort Reef in the northeast. As a result of the sampling efforts we were able to analyze ~ 10-year temperature records we collected at multiple sites throughout the Florida Keys along with regional scale observations of sea surface height anomalies from satellite altimetry. Significant observations of the time series included quasi synchronous events of markedly enhanced temperature variability occurring at multiple reef sites especially in summer. Earlier efforts at Conch Reef had identified roles of the Florida Current in modulating the observed internal wave dynamics at that site (Davis et al., 2008). Analyses of the Keys-wide data indicated apparent lag times in the onset of multi-day cooling events with sharp increases in semi-diurnal temperature variability. Coupling the direct observations on the reef crest with satellite altimetry at the scale of the entire Florida Current revealed that current meanders with embedded eddies were the mostly likely cause of changes in temperature and density stratification depth offshore of the reef slopes and associated eddies in the Florida Current traveling north-east along the reef tract (Leichter et al., 2014), Figure 11. This research revealed how meso-scale current meanders could result in quasi synchronous enhancement of internal wave activity at the scale of entire regions within the Florida Keys Reef Tract.
Figure 11
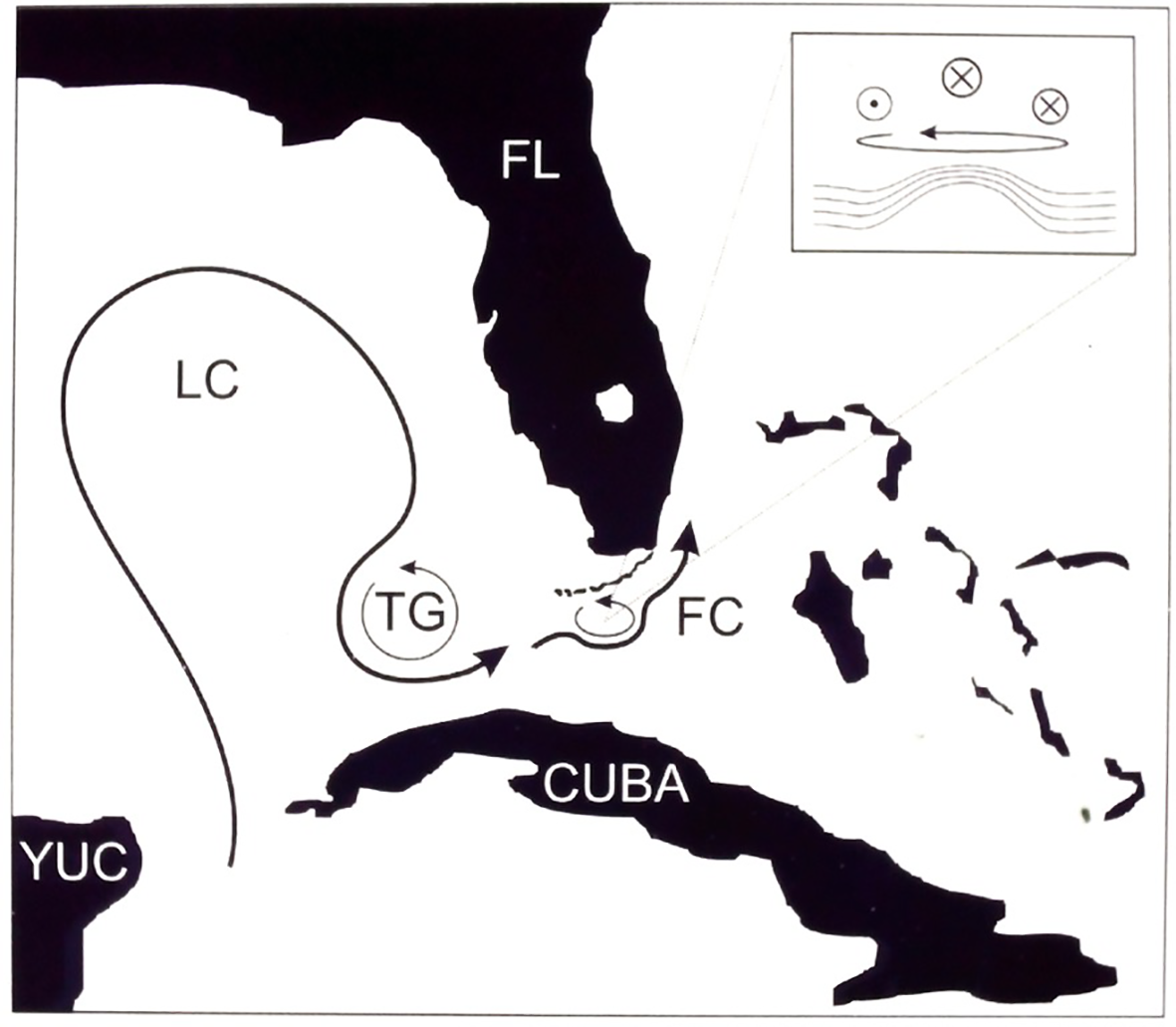
Schematic representation of the influence of the Florida Current (FC) cyclonic eddies likely to cause local uplift of isopycnals and enhanced internal wave activity as they travel along the Florida Keys Reef Tract. Tortugas Gyre and Loop Current shown upstream (West) of the Florida Current.
An unplanned event early in our research in 1994 also allowed direct observations and appreciation of conditions on a reef slope that can be quite different than experienced during normal dive excursions during favorable conditions. During Nov 1994, a locally intense tropical cyclone, Gordon, passed over the Florida Keys after the start of an Aquarius Habitat mission. Gordon was a late season storm with catastrophic impacts in regions of the Caribbean and Central America. NOAA meteorological buoy #42037 near Isla Morada (24.508° N, 81.873° W) recorded hourly significant wave heights between 2 and 3 m on Nov 16 and 17, 1994. Saturation and surface supported divers had completed deployment of a suite of instruments intended to sample water near-bottom and water column variability associated with internal waves, but instead observed the impacts of a large storm surge with extensive resuspension of sediments, high near bottom velocities sufficient to cause movement of the Aquarius Habitat, and local reductions in temperature (Figure 12). The planned 10-day saturation diving mission was terminated with manually controlled decompression utilizing onboard gas storage tanks following the loss of primary life support due to an electrical fire on the surface support barge. While unplanned, the direct observations by scientists inside the Aquarius Habitat during extreme weather with data collected by sampling instruments offered a direct view of environmental conditions on a coral reef that would have been impossible through normal boating or diving operations. By contrast a great majority of studies of storm impacts on coral reefs were necessarily initiated after, rather than before or during, the passage of large storms (e.g. Woodley et al., 1981).
Figure 12
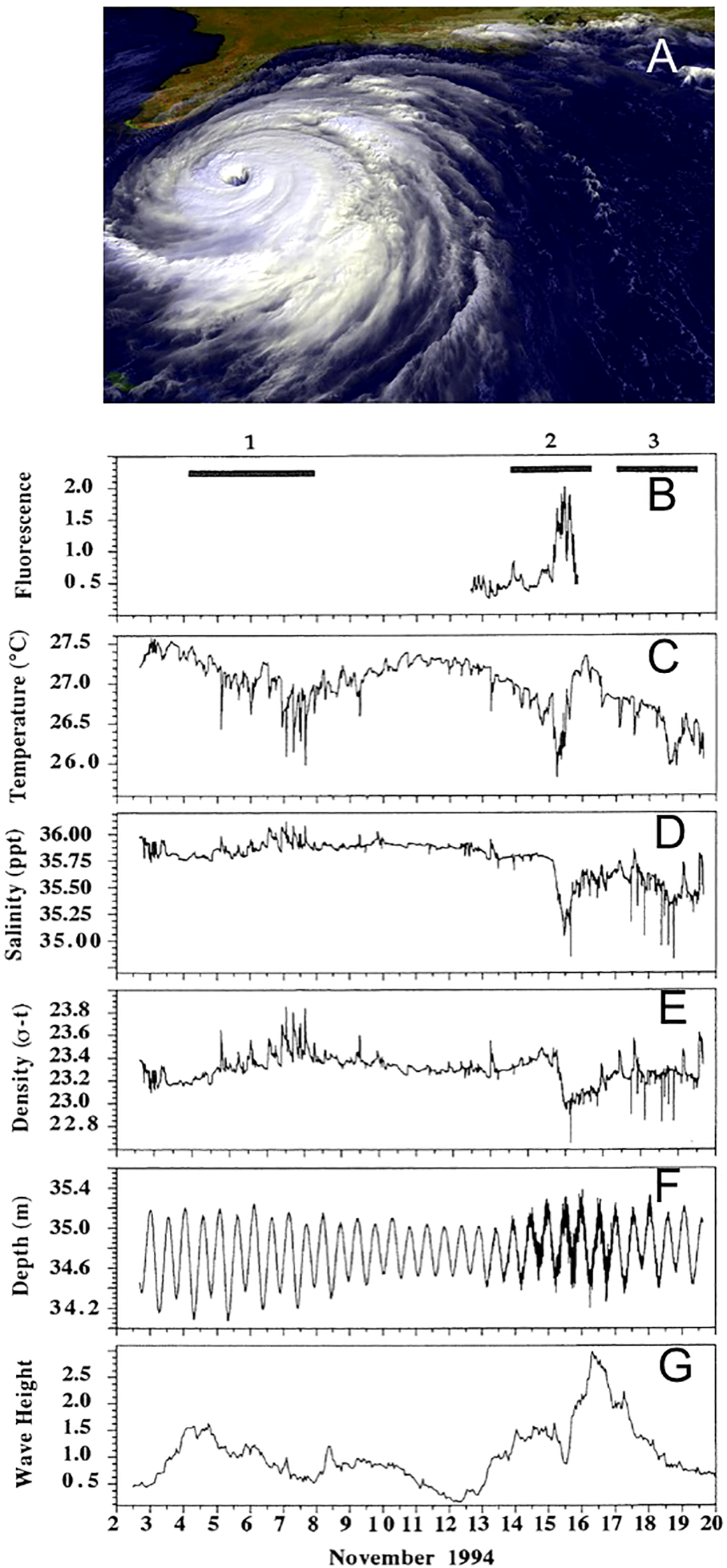
Influences of Tropical Storm Gordon (A) as it approached and passed over the Florida Keys during an Aquarius Habitat saturation mission, Nov 1994. Associated with the arrival and passage of the storm chlorophyll-a fluorescence (B) increase (prior to instrument failure), water temperature (C) salinity (D), and (E) density decreased with rapid fluctuations across the tidal signal (F), while significant wave height (G) increased up > 2.5 m. The large significant wave heights and wave periods caused high water velocities on the seafloor, significant movement of the occupied Aquarius Habitat, loss of surface support power, and very limited visibility. Data from Wing and Leichter, 1997.
Global observations and future directions
Scientific observations of the impact of internal waves on coral reefs now span the tropical ocean basins. Internal waves have been identified as major contributors to the physical dynamics of reefs, and particularly to reefs in deeper water (e.g. depth > 15 m) where research effort has typically been far less than in shallower reef environments. Overall, environmental cooling brought about by internal waves is known to be a key element of heat stress on reefs at widely varying geographic locations (e.g. Wall et al., 2015;Gove et al., 2016; Wyatt et al., 2020). Recent and future research could also benefit greatly by incorporating remote sensing observations, for example of sea surface thermal features and internal wave slicks, as well as Lagrangian observations from autonomous gliders and profiling drifters such as Argo floats, and from continuous observations made by coastal observing stations. For example, recent work utilizing remote sensed data and autonomous drifter data has identified ocean eddy dynamics similar to those we observed earlier in the Florida Keys, playing an important cooling role affecting the outcome of extreme heating events and coral bleaching in the central Pacific (Wyatt et al., 2023). Recent empirical and modeling results suggest that the propagation of internal waves is increasing with changes in ocean stratification driven by climate change (Zhao, 2023; Gong et al., 2025), and internal wave impacts on coral reefs may be increasingly important in mitigating heat stress under warming sea surface conditions (Wyatt et al., 2020).
While there have been significant recent advances in remotely operated and autonomous instrumentation and vehicles, a strong case can be made for the continued and in fact expanding value of direct human presence and observations in marine environments. Among the benefits of direct human operations facilitated by scientific diving are our ability to observe the broad community structure of multi-species assemblages, identify patches and length scales of variability, apply intuition to the design of long-term sampling strategies, pose hypotheses regarding community dynamics in response to physical environmental forcing, and conduct manipulative experiments efficiently. Each of these capabilities of divers in situ provide enhanced human perception to identify environmental and natural history surprises and mismatches from expectations. Continued developments in diving approaches and technologies, including closed circuit rebreathers, safe multi-depth decompression algorithms and dive computers, and potentially a new era of prolonged duration saturation diving all offer new ways to expand the duration of diver excursion times and depths of safe operations.
Many marine scientists remain optimistic that there are compelling reasons to expand the capabilities for scientists to work directly in underwater environments and to continue developing sensor networks that compliment and extend diver observations. However, it is also important to note that such capabilities are costly with respect to equipment, training time, and logistics and that these costs can represent a disproportionate burden on scientists from smaller institutions and in countries with limited science budgets. Collaborative efforts to share resources across programs and to support inclusion across economic levels can build a stronger base of diving scientists. In addition, the unfortunate trend in many scientific institutions of increasing restrictions on diving depths and operations well within levels proven to be safe through years of scientific and recreational diving will reduce opportunities for future exploration and new discoveries, and perhaps most importantly, training of future generations of graduate students and early career scientists. In efforts particularly by universities to manage and eliminate risk, an overlooked and hidden category of risk may be that overly restrictive operational constraints can prevent new observations and scientific discoveries that could have been safely made.
Exciting new opportunities exist at the interfaces of sensor and microelectronics design, human diving capabilities, and advanced computing for large, multi-parameter datasets and image processing. We advocate for actively pushing the frontiers of scientific discoveries at these interfaces of sensor development, human observations, and computation. The growing ability to combine direct human underwater activities with technology deployment increases our scientific ability to sample at large numbers of sites, synchronously, and over long-term measurement periods. Large distributed sensor networks and multi-function sensors incorporating, for example, state-of-the-art transmissometers, fluorometers, and current meters could be connected to a future saturation diving station. Combining human presence in underwater environments with sophisticated instrumentation can lead to new modes of adaptive sampling to understand biological responses to the dynamic physical environments of coral reefs.
Statements
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The manuscript presents research on animals that do not require ethical approval for their study. Written informed consent was obtained from the individual(s) for the publication of any identifiable images or data included in this article.
Author contributions
JL: Writing – original draft, Writing – review & editing. SW: Writing – original draft, Writing – review & editing. MS: Writing – original draft, Writing – review & editing. SG: Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. NOAA National Undersea Research Program grants U.S. National Science Foundation Grants OCE-9986547, OCE-0242157, and OCE-0927448 to JL.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1
Baines P. G. (1986). Internal tides, internal waves and near-inertial motions. In: Mooers ed. Baroclinic processes continental shelves3, 19–31. (Berlin, Springer-Verlag) doi: 10.29/C0003
2
Bellwood D. R. Pratchett M. S. Morrison T. H. Gurney G. G. Hughes T. P. Álvarez-Romero J. G. et al . (2019). Coral reef conservation in the Anthropocene: Confronting spatial mismatches and prioritizing functions. Biol. Conserv.236, 604–615. doi: 10.1016/j.biocon.2019.05.056
3
Benjamin S. (2021). “Aquarius habitat,” in Handbook of life support systems for spacecraft and extraterrestrial habitats. Eds. SeedhouseE.ShaylerD. J. (Springer, Cham). doi: 10.1007/978-3-319-09575-2_164-1
4
Birkeland C. (Ed.) (1997). Life and death of coral reefs. (New York: Chapman & Hall).
5
Brown B. E. (1997). Coral bleaching: causes and consequences. Coral reefs16, S129–S138. doi: 10.1007/s003380050249
6
Cacchione D. A. Drake D. E. (1986). Nepheloid layers and internal waves over continental shelves and slopes. Geo-Marine Lett.6, 147–152. doi: 10.1007/BF02238085
7
Cacchione D. A. Pratson L. F. Ogston A. S. (2002). The shaping of continental slopes by internal tides. Science296, 724–727. doi: 10.1126/science.1069803
8
Connell J. H. (1978). Diversity in tropical rain forests and coral reefs: high diversity of trees and corals is maintained only in a nonequilibrium state. Science199, 1302–1310. doi: 10.1126/science.199.4335.1302
9
Darwin C. R. (1842). “The structure and distribution of coral reefs,” in Being the first part of the geology of the voyage of the Beagle, under the command of Capt. Fitzroy, R.N. during the years 1832 to 1836 (Smith and Elder, London).
10
Davis K. A. Leichter J. J. Hench J. L. Monismith S. G. (2008). Effects of western boundary current dynamics on the internal wave field of the Southeast Florida shelf. J. Geophysical Research: Oceans113. C09010. doi: 101029/2007JC004699
11
Dobbs D. (2009). Reef madness: Charles Darwin, Alexander Agassiz, and the meaning of coral (New York: Pantheon).
12
Erez J. (1990). On the importance of food sources in coral-reef ecosystems. In Dubinsky ed. Ecosystems of the World, Coral Reefs, 25, 411–418 (Amsterdam: Elsevier).
13
Ewing G. C. (1950). Relation between band slicks at the surface and internal waves in the sea. Science111, 91–94. doi: 10.1126/science.111.2874.91.b
14
Gerber R. P. Marshall N. (1982). Characterization of the suspended particulate organic matter and feeding by the lagoon zooplankton at Enewetak atoll. Bull. Mar. Sci.32, 290–300.
15
Glynn P. W. (1973). Ecology of a Caribbean coral reef. The Porites reef-flat biotope: Part II. Plankton community with evidence for depletion. Mar. Biol.22, 1–21. doi: 10.1007/BF00388905
16
Gong Y. Chen Z. Xu J. Yao Y. Wang C. Cai S. (2025). Accelerated internal tides in a warming climate. Sci. Adv.11, eadq4577. doi: 10.1126/sciadv.adq4577
17
Goreau T. F. (1959). The ecology of Jamaican coral reefs I. Species composition and zonation. Ecology40, 67–90. doi: 10.2307/1929924
18
Gove J. M. McManus M. A. Neuheimer A. B. Polovina J. J. Drazen J. C. Smith C. R. et al . (2016). Near-island biological hotspots in barren ocean basins. Nat. Commun.7, 10581. doi: 10.1038/ncomms10581
19
Hamner W. M. Jones M. S. Carleton J. H. Hauri I. R. Williams D. M. (1988). Zooplankton, planktivorous fish, and water currents on a windward reef face: Great Barrier Reef, Australia. Bull. Mar. Sci.42, 459–479.
20
Hatcher B. G. (1997). Organic production and decomposition. In Birkland, ed. Life and Death of Coral Reefs (New York: Chapman & Hall), 140–174.
21
Hobson E. S. Chess J. R. (1978). Trophic relationships among fishes and plankton in the lagoon at Enewetak Atoll, Marshall Islands. Fish Bull.76, 133–153.
22
Holloway P. E. (1991). On the dissipation of internal tides. Tidal hydrodynamics449, 466.
23
Jokiel P. L. Coles S. L. (1977). Effects of temperature on the mortality and growth of Hawaiian reef corals. Mar. Biol.43, 201–208. doi: 10.1007/BF00402312
24
Jokiel P. L. Coles S. L. (1990). Response of Hawaiian and other Indo-Pacific reef corals to elevated temperature. Coral reefs8, 155–162. doi: 10.1007/BF00265006
25
Lamb H. (1945). Hydrodynamics. (New York: Dover).
26
Lamb K. G. (2014). Internal wave breaking and dissipation mechanisms on the continental slope/shelf. Annu. Rev. Fluid Mechanics46, 231–254. doi: 10.1146/annurev-fluid-011212-140701
27
Leichter J. J. Deane G. B. Stokes M. D. (2005). Spatial and temporal variability of internal wave forcing on a coral reef. J. Phys. Oceanography35, 1945–1962. doi: 10.1175/JPO2808.1
28
Leichter J. J. Genovese S. J. (2006). Intermittent upwelling and subsidized growth of the scleractinian coral Madracis mirabilis on the deep fore-reef slope of Discovery Bay, Jamaica. Mar. Ecol. Prog. Ser.316, 95–103. doi: 10.3354/meps316095
29
Leichter J. J. Miller S. L. (1999). Predicting high-frequency upwelling: Spatial and temporal patterns of temperature anomalies on a Florida coral reef. Continental Shelf Res.19, 911–928. doi: 10.1016/S0278-4343(99)00004-7
30
Leichter J. J. Shellenbarger G. Genovese S. J. Wing S. R. (1998). Breaking internal waves on a Florida (USA) coral reef: a plankton pump at work? Mar. Ecol. Prog. Ser.166, 83–97. doi: 10.3354/meps166083
31
Leichter J. J. Stewart H. L. Miller S. L. (2003). Episodic nutrient transport to Florida coral reefs. Limnology Oceanography48, 1394–1407. doi: 10.4319/lo.2003.48.4.1394
32
Leichter J. J. Stokes M. D. Genovese S. J. (2008). Deep water macroalgal communities adjacent to the Florida Keys reef tract. Mar. Ecol. Prog. Ser.356, 123–138. doi: 10.3354/meps07230
33
Leichter J. J. Stokes M. D. Vilchis L. I. Fiechter J. (2014). Regional synchrony of temperature variation and internal wave forcing along the Florida Keys reef tract. J. Geophysical Research: Oceans119, 548–558. doi: 10.1002/2013JC009371
34
Leichter J. J. Wankel S. Paytan A. Hanson K. Miller S. Altabet M. A. (2007). Nitrogen and oxygen isotopic signatures of subsurface nitrate seaward of the Florida Keys reef tract. Limnology Oceanography52, 1258–1267. doi: 10.4319/lo.2007.52.3.1258
35
Leichter J. J. Wing S. R. Miller S. L. Denny M. W. (1996). Pulsed delivery of subthermocline water to Conch Reef (Florida Keys) by internal tidal bores. Limnology Oceanography41, 1490–1501. doi: 10.4319/lo.1996.41.7.1490
36
Locke J. M. Weil E. Coates K. A. (2007). A newly documented species of Madracis (Scleractinia: Pocilloporidae) from the Caribbean. Proc. Biol. Soc. Washington120, 214–226. doi: 10.2988/0006-324X(2007)120[214:ANDSOM]2.0.CO;2
37
Patterson M. R. (1992). A chemical engineering view of cnidarian symbioses. Am. Zoologist32, 566–582. doi: 10.1093/icb/32.4.566
38
Patterson M. R. Sebens K. P. Olson R. R. (1991). In situ measurements of flow effects on primary production and dark respiration in reef corals. Limnology Oceanography36, 936–948. doi: 10.4319/lo.1991.36.5.0936
39
Pineda J. (1991). Predictable upwelling and the shoreward transport of planktonic larvae by internal tidal bores. Science253, 548–549. doi: 10.1126/science.253.5019.548
40
Pond S. Pickard G. L. (1983). Introductory dynamical oceanography (Oxford: Butterworth-Heinemann).
41
Sandstrom H. Elliott J. A. (1984). Internal tide and solitons on the Scotian Shelf: A nutrient pump at work. J. Geophysical Research: Oceans89, 6415–6426. doi: 10.1029/JC089iC04P06415
42
Sponaugle S. Lee T. Kourafalou V. Pinkard D. (2005). Florida Current frontal eddies and the settlement of coral reef fishes. Limnology Oceanography50, 1033–1048. doi: 10.4319/lo.2005.50.4.1033
43
Stoddart D. R. (1976). Darwin, Lyell, and the geological significance of coral reefs. Br. J. History Sci.9, 199–218. doi: 10.1017/S0007087400014783
44
Stokes M. D. Leichter J. J. Wing S. R. (2020). Spatial variations in the stable isotope composition of the benthic algae, Halimeda tuna, and implications for paleothermometry. Sci. Rep.10, 16276. doi: 10.1038/s41598-020-73283-x
45
Stokes M. D. Leichter J. J. Wing S. Frew R. (2011). Temperature variability and algal isotopic heterogeneity on a Floridian coral reef. Mar. Ecol.32, 364–379. doi: 10.1111/j.1439-0485.2011.00469.x
46
Sverdrup H. U. Johnson M. W. Fleming R. H. (1942). The Oceans: Their physics, chemistry, and general biology (New York: Prentice-Hall).
47
Wall M. Putchim L. Schmidt G. M. Jantzen C. Khokiattiwong S. Richter C. (2015). Large-amplitude internal waves benefit corals during thermal stress. Proc. R. Soc. B: Biol. Sci.282, 20140650. doi: 10.1098/rspb.2014.0650
48
Wells J. W. (1973). Coral reef project—papers in memory of Dr. Thomas F. Goreau. 2. New and old scleractinian corals from Jamaica. Bull. Mar. Sci.23, 16–58.
49
Wing S. R. Leichter J. J. (1997). Storm-generated physical variability on Conch reef, Florida Keys. Proc. 8th Int. Coral Reef Symposium.2, 1097–1102.
50
Wolanski E. Delesalle B. (1995). Upwelling by internal waves, Tahiti, French Polynesia. Continental shelf Res.15, 357–368. doi: 10.1016/0278-4343(93)E0004-R
51
Wolanski E. Pickard G. L. (1983). Upwelling by internal tides and Kelvin waves at the continental shelf break on the Great Barrier Reef. Mar. Freshw. Res.34, 65–80. doi: 10.1071/MF9830065
52
Woodley J. D. Chornesky E. A. Clifford P. A. Jackson J. B. C. Kaufman L. S. Knowlton N. et al . (1981). Hurricane Allen’s impact on Jamaican coral reefs. Science214, 749–755. doi: 10.1126/science.214.4522.749
53
Wyatt A. S. Leichter J. J. Toth L. T. Miyajima T. Aronson R. B. Nagata T. (2020). Heat accumulation on coral reefs mitigated by internal waves. Nat. Geosci.13, 28–34. doi: 10.1038/s41561-019-0486-4
54
Wyatt A. S. Leichter J. J. Washburn L. Kui L. Edmunds P. J. Burgess S. C. (2023). Hidden heatwaves and severe coral bleaching linked to mesoscale eddies and thermocline dynamics. Nat. Commun.14, 25. doi: 10.1038/s41467-022-35550-5
55
Wyatt A. S. Lowe R. J. Humphries S. Waite A. M. (2010). Particulate nutrient fluxes over a fringing coral reef: relevant scales of phytoplankton production and mechanisms of supply. Mar. Ecol. Prog. Ser.405, 113–130. doi: 10.3354/meps08508
56
Yonge C. M. (1963). The biology of coral reefs. In Adv. Mar. Biol.1, 209–262. doi: 10.1016/S0065-2881(08)60260-6
57
Zhao Z. (2023). Satellite evidence for strengthened M2 internal tides in the past 30 years. Geophysical Res. Lett.50, e2023GL105764. doi: 10.1029/2023GL105764
Summary
Keywords
internal waves, coral reefs, mass transport, nutrient and plankton dynamics, thermal refugia, climate change, ecosystem connectivity
Citation
Leichter JJ, Wing SR, Stokes MD and Genovese SJ (2025) Being there: scientific saturation and technical diving with in situ instrumentation in an internal surf zone reveal driving physical dynamics of coral reefs. Front. Mar. Sci. 12:1548608. doi: 10.3389/fmars.2025.1548608
Received
19 December 2024
Accepted
19 May 2025
Published
19 June 2025
Volume
12 - 2025
Edited by
John D. Coley, Northeastern University, United States
Reviewed by
Charles Reid Nichols, Marine Information Resources Corporation, United States
Alejandra Hernandez-Agreda, California Academy of Sciences, United States
Updates
Copyright
© 2025 Leichter, Wing, Stokes and Genovese.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: James J. Leichter, jleichter@ucsd.edu
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.