 Johanna Paola Echeverry Hernandez
Johanna Paola Echeverry Hernandez José Ernesto Mancera Pineda
José Ernesto Mancera Pineda David Alejandro Sánchez Núñez
David Alejandro Sánchez Núñez- 1Department of Biology, Faculty of Sciences, Universidad Nacional de Colombia–Sede Bogotá, Bogotá, Colombia
- 2Dirección Académica, Universidad Nacional de Colombia-Sede de La Paz, La Paz, Cesar, Colombia
To understand the changes in the position of the mangrove forest fronts, migration trends of Rhizophora mangle along the coast, were analyzed in response to oceanographic, climatic variability and hydrological coast regulators. Due to the large biological, physical and climatic differences between Pacific and insular Caribbean coasts of Colombia, the working hypothesis proposed differences in migration drivers and mangrove responses between the coasts. Remote sensing and photointerpretation techniques were used to identify and quantify the spatial variation of deltaic forest types on the Pacific coast and open-water forest types in the Caribbean. Regression analysis was used to relate mangrove front changes with river water discharge, ENSO climate variability, and wave height and velocity coming from directions in interaction or not with the coral reef (in open water forest). It was found that in 13 years of observation (2010 to 2023), there were constant changes in the position of the mangrove front in both the Caribbean and the Pacific. Wave heights from directions that do not interact with the coral reef barrier, such as during periods of strong hurricane influence, along with ENSO variability, collectively explained the annual changes in the mangrove front of open waters (R² = 0.91). Freshwater discharge, which was linked to the size of a protective sediment bar in front of the mangroves, significantly accounted for the annual changes in mangrove coverage (R² = 0.70). During the period analyzed, the open-coast mangrove front exhibited a progradation of 0.45 to 1.02 yr⁻¹, while the deltaic mangrove in Bocagrande showed a retreat of 1.13 yr⁻¹. These findings provide valuable insights for marine spatial planning, supporting mangrove conservation efforts.
1 Introduction
Mangrove forests have been observed to modify their cover in response to physical changes in the environment, including climatic cycles and their variability (Friess et al., 2019). This adaptation is driven by their sensitivity and ability to thrive in the face of processes and phenomena occurring in the hydrosphere, lithosphere, and atmosphere (Spalding et al., 2014). One aspect of spatiotemporal variation in mangrove cover is the progradation and retreat of their outer edge. Such mangrove front changes are attributable to the response of mangroves (Lovelock et al., 2016) to abiotic factors or forcings (Hickey et al., 2021; Ghosh et al., 2020; Raw et al., 2019) at both local and regional scales (Spalding et al., 2014).
One of these factors is wave propagation, which causes fluctuations in the water level and wave energy, influencing mangrove forests (Pokhrel et al., 2022). Wave climate and its anomalies are a major energy forcing factor in much of the coastal zone, significantly affecting circulation, sediment transport, and coastal morphology (Babanin et al., 2019; Restrepo López et al., 2009). In addition, climate variability events associated with the El Niño Southern Oscillation (ENSO) constitute a key forcing factor. Changes in environmental conditions due to ENSO-associated variables, such as sea level pressure (SLP) and temperature, are common in many regions of the world, in both marine and terrestrial systems (Torres et al., 2016). These changes coincide with significant sediment accumulation and/or loss on coasts (Vos et al., 2021). These dynamics have implications for coastal vegetation cover. In northwestern Australia, for instance, a correlation has been identified between ENSO and alterations in the region’s mangroves, as demonstrated by a decrease in mangrove canopy cover during El Niño periods (Hickey et al., 2021).
The discharge of land-sea sediments through fluvial systems is a key factor in coastal change, as it serves as the primary mechanism for sediment transport and morphological evolution. This process is influenced by the horizontal advection of freshwater, which shapes the morphology of the fluvial plume and, in turn, affects sediment distribution along the coast (Vundavilli et al., 2021).
A recent Food and Agriculture Organization of the United Nations (2023) report highlights that the natural contraction and expansion of mangrove forests in South America are typical processes. In 2020, the FAO estimated a total mangrove area of 2.14 million ha for the region, with a loss of 89,400 ha and a gain of 91,300 ha from 2000 to 2020. In Colombia, the estimated mangrove area for 2020 is 280,754 ha, representing 56.1% of the country’s total coastline, following an estimated loss of 726,900 ha since 1996 (Global Mangrove Watch, 2024). Although the exact contribution of mangrove erosion and progradation to these changes remains unknown, it is evident that mangrove cover seaward is highly dynamic in deltaic and open coast systems (Restrepo and Cantera, 2013; Sánchez-Núñez et al., 2019)
In Colombia, mangrove forests are located along the coasts and islands of the Caribbean (27.3%) and Pacific (72.7%), covering an area of 2,891 km2 (Rodríguez-Rodríguez et al., 2021). This makes Colombia the sixteenth largest mangrove area globally (Bunting et al., 2022) and the first in terms of the tallest trees (Simard et al., 2025). However its significance lies not only in the extent of the forests, but also in their diversity. All mangrove types described in the latest biophysical typology are found in Colombia (Worthington et al., 2020). In the Pacific, characterized by very high annual rainfall and a mesotidal regime, deltaic and estuarine mangroves predominate, while in the Caribbean, with a microtidal regime and a mostly dry climate, lagoonal and open coast mangroves predominate (Mancera Pineda, 2022).
This research examines the migration of the R. mangle forest front in response to oceanographic (wave), climatic (ENSO), and hydrological (freshwater discharge) factors that drive coastal changes, with the aim of identifying and understanding site-specific migration trends. Remote sensing and photo-interpretation techniques were applied to identify and quantify the spatial variation of mangrove forests in the Bocagrande sector, Tumaco (on the Colombian Pacific coast), and in the Old Point Mangroves Regional Park on San Andrés Island (Colombian Caribbean), between 2010 and 2023. Given the large biological, physical and climatic differences between the Caribbean and the Colombian Pacific, the working hypothesis proposed variations in the migration factors and responses of R. mangle forests between these two coastal regions.
2 Materials and methods
2.1 Study area
The study was conducted in mangrove forests that were selected from distinct environments in terms of sedimentary origin, physiography, geology, and even conservation policies. Tumaco in the South Pacific and the island of San Andrés in the insular Caribbean (Figure 1). The mangroves of the insular Caribbean, the continental Caribbean, and the Colombian Pacific exhibit significant floristic differences, primarily attributable to three factors: 1. The strong climatic changes of the Pleistocene, marked by prolonged periods of drought and fluctuations in sea level, affected the vegetation of the Caribbean, while the Pacific coast maintained stable climatic conditions. The low tidal penetration, which oscillates between 40 and 60 centimeters in the insular Caribbean, causing the forests to be limited to narrow strips, contrasts with the tidal range of the Pacific, which oscillates between 2 and 3 meters in the low mesotidal zone. The range of the low mesotidal type is from 5 m to 3.5 m, and the range of the high mesotidal type is from 3.5 m to 5 m (Instituto de Investigaciones Marinas y Costeras José Benito Vives de Andréis, 2003). This enables the forests to extend inland, where the topography permits their settlement. The substantial productivity of the Pacific mangroves is attributable to the abundant supply of fresh water with inorganic nutrients, which are derived from the substantial quantity of hydric sources that flow into the Pacific (INVEMAR, 2003). This dynamic environment fosters the growth and proliferation of forests characterized by trees that attain heights exceeding 40 m, as previously documented by von Prahl (1989).
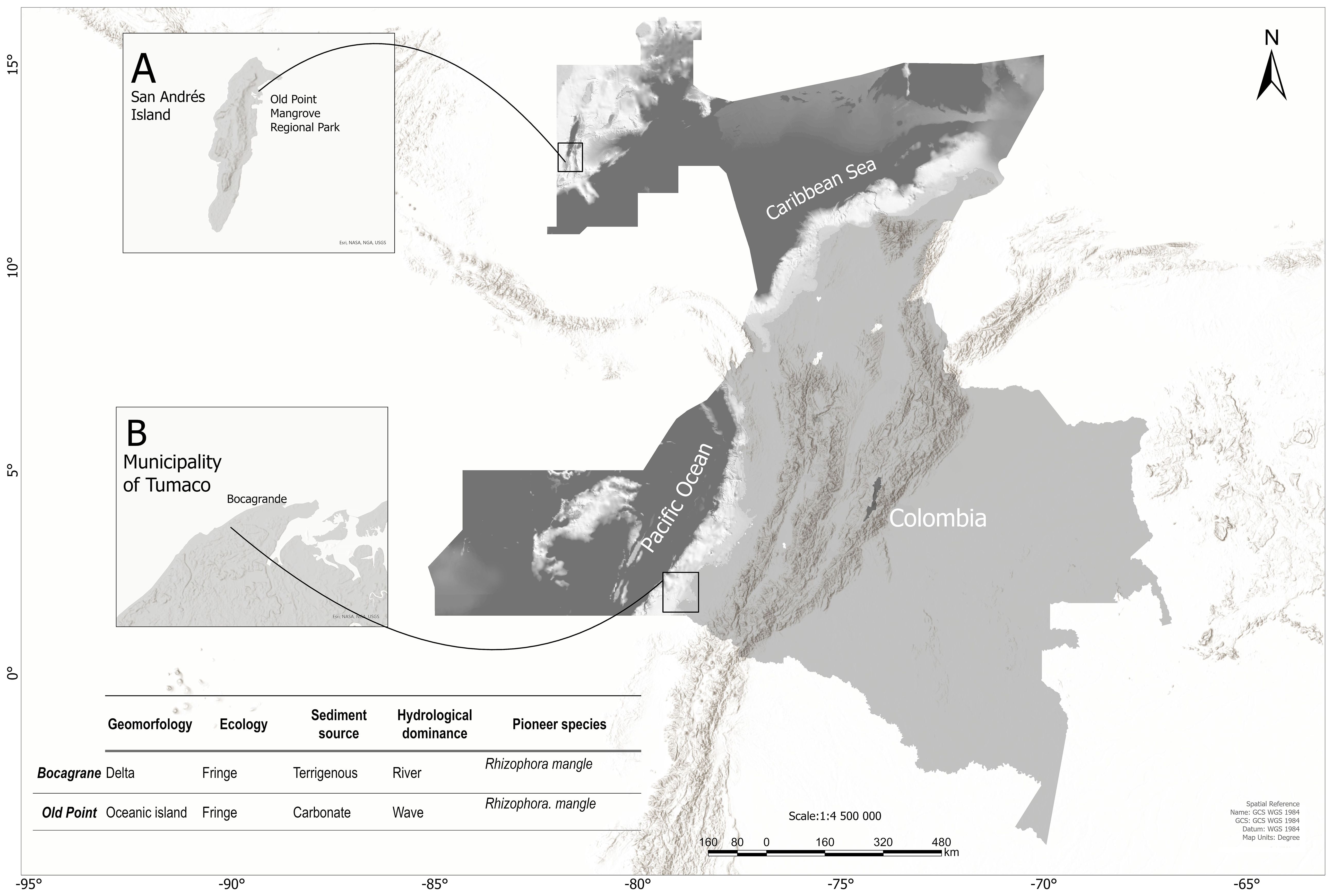
Figure 1. Location of the study area. (A) San Andrés Island, Colombian insular Caribbean. (B) San Andrés de Tumaco, Colombian Pacific.
Tumaco is located in one of the few global systems that remain without anthropic intervention: the Mira river Delta (Monroy et al., 2008), whose subaerial surface of 520 km2 is composed of intertidal plains, the delta front, and the prodelta (Restrepo López et al., 2008), an environment where sedimentation is highest (Syvitski et al., 1985). Its geomorphology is predominantly influenced by wave energy and is characterized by two major arms (Posada Posada et al., 2009) and eight distributary channels with a width of 521 m along a 50 km coastline. The estimated sediment discharge through the estuary is 9.77x106 t yr-1, with a maximum monthly streamflow of 3,270 m3 s-1 (López and Restrepo, 2007). According to Álvarez-Silva et al. (2022), the horizontal extension of the Mira river plume takes place parallel to the coast, symmetrically in the north and south directions. This phenomenon is attributed to the presence of the river, which generates significant currents directed towards the Bay of Tumaco. These currents are responsible for the transportation of sediments that nourish the beaches and bars, thereby safeguarding the municipal capital from the threat of marine hazards.
The oceanic island of San Andrés, the largest of a group of islands that form the archipelago of San Andrés, Providencia and Catalina, located in the northwest of the Colombian Caribbean Sea, was declared an International Biosphere Reserve by UNESCO in 2000. The island’s topography is distinguished by karstic features, featuring Tertiary limestone formations and recent Quaternary deposits. The island receives an average annual rainfall of 1973 millimeters, with temperatures ranging from 25°C to 30°C. The island’s climate is classified into three distinct seasons: a rainy season, which occurs from June to November and accounts for 80% of the annual precipitation; a dry season that extends from January to April; and two transitional months of May and December. This seasonal pattern has been documented by Medina-Calderón et al. (2021).
The Pacific study area is located south of the urban area, in the sector of Bocagrande, at coordinates 78°50’18.065”W, 1°48’45.318”N. There, the mangrove forest is of deltaic type, according to geomorphology, and of fringe type, according to its physiognomy, and is influenced by a very dynamic barrier beach due to the action of marine currents and the discharge of sediments from the Mira river. Furthermore, ENSO exerts a substantial influence on the hydrology and oceanography of the region, affecting variables such as precipitation rates, streamflow, and mean sea level (Álvarez-Silva et al., 2022).
The study area of the insular Caribbean is located within the confines of the Old Point Mangroves Regional Park, situated northeast of San Andrés Island at precise coordinates 81°42’3.918” W and 12°33’56.397” N. Given the absence of permanent water courses on San Andrés, the hydrological cycle of this mangrove is predominantly dictated by tidal fluctuations and precipitation (Sánchez-Núñez and Mancera-Pineda, 2011). According to their geomorphology, the mangroves on the island are of the open coastal type with karstic sediments (Worthington et al., 2020). The total area covered by the six forests on the island amounts to 1.51 km2 (Global Mangrove Watch, 2024). Of particular interest are the inland forests of Sound Bay and Smith Channel, which are not connected to the sea, and the edge and basin forests of Salt Creek, Cocoplum Bay, El Cove, and Old Point Regional Mangrove Park, which are connected to the sea (Medina-Calderón et al., 2021).
In both study areas, the pioneer species is R. mangle, which, according to the mangrove species zonation along the salinity gradient, is located in the lower estuarine zones (Costa et al., 2015). There are structural differences between the mangroves in the two environments: In Bocagrande, the Diameter at Breast Height (DBH) averages 31.11 ± 1.93 cm, with a density of 450 ± 54.09 ind. ha−1 and a relative abundance of 75% (Gomez Garcia, 2024), while in Old Point, the average DBH is 6.8 ± 0.1 cm, the density is 2520 ± 2.3 ind. ha−1, and the relative abundance is 68% (Quintero, 2025). Additionally, the volumetric biomass density calculated in cubic meters from the ground level (Horstman et al., 2014) is 0.085 in Bocagrande, while in San Andrés it is 0.022 (Echeverry Hernández, 2025) (Figure 2).

Figure 2. Structural differences of Rhizophora mangle trees in (A) Bocagrande and (B) Old Point.
2.2 Changes in the position of the mangrove front
A multitemporal analysis was conducted to characterize and analyze the morphological evolution and spatial changes in the position of the mangrove front, specifically of the pioneer species R. mangle. Remote sensing technology was employed due to its accessibility and cost-effectiveness. Satellite images from 2010 to 2023, obtained by Planet’s Education and Research Program (Planet Labs, 2023), were utilized (Figures 3, 4). This program provides daily imagery from the Planet Scope satellite constellation (2016-2023) and the Rapideye satellite constellation (2010–2014) (European Space Agency, 2024). The RapidEye images possess a spatial resolution of 5 meters, while the Planet Scope images have a spatial resolution of 3 m. The complete set of images corresponds to multispectral panchromatic images, comprising green, blue, red, and near-infrared bands, with a temporal resolution of one day for the equatorial zones (Table 1).

Figure 3. Satellite images of the Bocagrande sector in Tumaco used for the multitemporal analysis. Rapideye (2010, 2012, 2013, 2014) and Planet Scope (2016, 2017, 2018, 2019, 2020, 2021, 2022, 2023).

Figure 4. Satellite images of the Old Point sector on San Andrés Island used for the multitemporal analysis. Rapideye (2011, 2012, 2013, 2014,2015) and Planet Scope (2016, 2017, 2018, 2019, 2020, 2021, 2022, 2023).
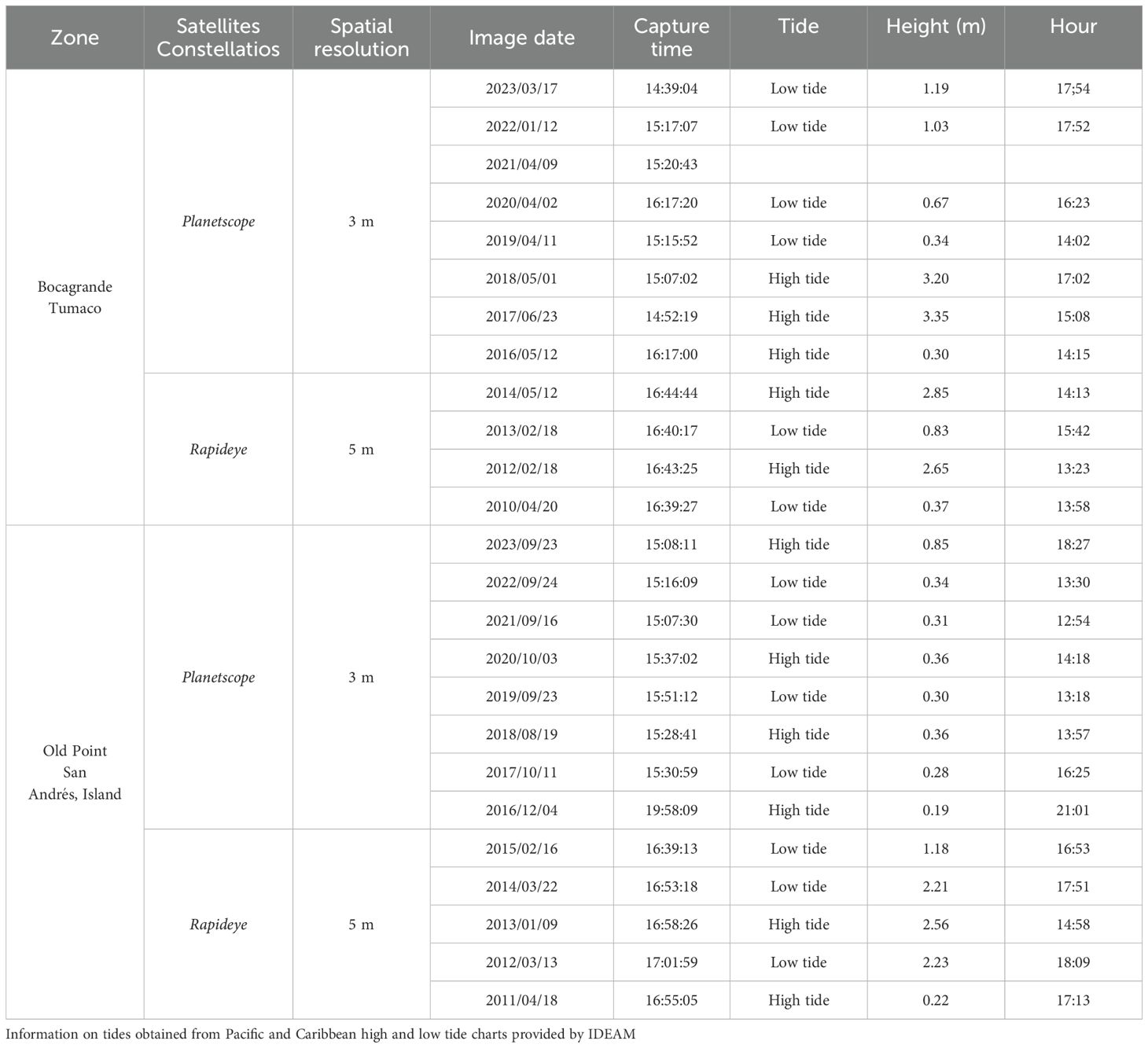
Table 1. Satellite images used in the analysis of the change in the position of the mangrove front line.
The satellite images were selected and subsequently processed using the visual analysis technique. This technique entailed the identification of the boundaries between the vegetation front, beaches, bars, or water bodies by shape, size, tones, and pattern (Lillesand et al., 2015). In each of the multitemporal images, all elements of the boundary line between the vegetation and the adjacent cover element or geoform (water or sediment) were vectorized. Subsequent to this, the alterations in the position of the mangrove front were measured. The utilization of the spatial position of the mangrove forest front as a bioindicator of coastal processes in the photointerpretation of images possesses the advantage that it is not influenced by the tide height at the time of image acquisition, since the spatial analysis is performed with a zenithal view of the forest canopy, in contrast to the analysis of changes in the coastline based on the interpretation of the land-water boundary, which depends on the time of image acquisition and the position of the tide at each instant.
The degree of accuracy of the position of the analyzed vegetation front line is determined by the spatial resolution of the images or the size of the pixels that compose it. The precision of the spatial positioning of the photo-interpreted details is contingent on the precision of the manual digitization, the spatial resolution of the images used (3 and 5 m), and the precision of their georeferencing (Gairin et al., 2021; Pogson and Smith, 2015).
The changes in the spatial configuration of the mangrove forest front were determined through the utilization of ArcMap 10.8 software, employing the Digital Shoreline Analysis System instrument developed by the U.S. Geological Survey. This instrument facilitates the automated estimation of change rate statistics from a sequence of coastal vector positions (Himmelstoss et al., 2021). The Digital Shoreline Analysis System is a widely adopted tool in research endeavors concerning erosion and coastal variability (Mishra et al., 2020; Oyedotun, 2014). The methodology employed is referred to as baseline transects (Federal Emergency Management Agency and US Department of Homeland Security, 2022) involves the establishment of equidistant transects perpendicular to a baseline, which follows the trajectory of the coastline. Each transect intersects each shoreline, enabling the measurement of the distance between them in meters. This approach is founded on a statistical technique known as the end point rate (EPR), which calculates the distance between two coastlines in meters by dividing the elapsed time in years between them (Jonah et al., 2016).
2.3 Wave parameters
The dataset under consideration in this study encompasses the ECMWF fifth generation reanalysis data for global climate over the past eight decades, designated as ERA5, which was obtained from the Climate Data Store (European Commission et al., 2023). These data are available since 1940, with hourly estimates of various variables, including oceanic variables. The data corresponding to the variables significant wave height (H), wave direction (D), and wave period (T) were obtained for specific pixels defined between the coordinates 1.5°N to 3°N latitude and 80°W to 78°W longitude for Tumaco, and 12.1°N to 13°N latitude and 81.5°W to 80.5°W for San Andrés. The data were downloaded in GRIB format and converted to.txt format using a Matlab routine. Consequently, the subsequent conversion facilitated the derivation of additional variables, including significant wave height, wavelength in deep water, number of waves, wave energy, group velocity, and wave power.
Considering the wave attenuation role attributed to mangroves, reducing wave energy by an average of 97% and dissipating approximately 86% of their energy (Ferrario et al., 2014), the changes in the vegetation front were analyzed in three sectors of Old Point, one in the northern area, protected by a bay in front of the Coast Guard station, another in the central area, parallel to the reef barrier, and finally one in the southern area, in front of Haines Bigth. Since San Andrés Island is protected by a coral reef about 8.5 kilometers long, located on the eastern side of the island, surrounding it from Johnny Cay at the northern tip to the sector of Rocky Cay, about 2.5 kilometers from the coast, a filtering of the wave data was performed, selecting those coming from the E, SE, S and SW, between angles 90° and 192.5° in direction, since this is the one that directly affects the southern area of the Old Point mangrove forest, given the absence of a coral reef in this sector. In the same sense, in the study area of Bocagrande, Tumaco, the data corresponding to waves coming from the NE, N, NW, W, SW and S, from directions 45° to 180° (counterclockwise), were selected, since these are the directions incident on the coastline, according to its orientation.
2.4 El Niño-Southern Oscillation
Climate variability events associated with El Niño have been shown to cause massive natural and social impacts around the world (Zebiak et al., 2014). In order to prepare responses to climate variability, observations, records, and forecasts are needed. To this end, several indices have been developed to monitor ENSO events based on sea surface temperature (SST) anomalies averaged over a given region (van Oldenborgh et al., 2021), such as those located between 170°W-120°W longitudes and 5°S-5°N latitudes (Bartholomew and Menglin, 2013).
One such index is the Oceanic Niño Index (ONI), which was developed in the 1990s by the Space Weather Prediction Center (NWS) of the National Oceanic and Atmospheric Administration (NOAA) (Webb & Magi, 2022). The ONI is designed to monitor changes in sea surface temperature (SST) in the tropical regions of the Pacific Ocean. The 3-month moving average (December–February, January–March, February–April, etc.) is employed to predict the onset and duration of the anomaly. The classification of El Niño or La Niña phenomena is determined by the extent of anomalies that exceed +0.5°C or -0.5°C for a minimum of five consecutive months. Typically, anomalies are calculated relative to a 30-year base period that is updated every five years (Glantz and Ramirez, 2020). Historical data concerning El Niño/La Niña events from 1950 to the present can be found on the NOAA’s ONI website (NOAA, 2023). When the threshold is reached for at least five consecutive overlapping seasons, periods of above-normal SST are shown in red and cool or below-normal periods are shown in blue, based on the Extended Reconstructed Sea Surface Temperature (ERSST) analysis. The values for the study years (2009-2023) were obtained from the ONI.
2.5 Streamflow
The large deltas and estuaries of the Colombian Pacific coast are mainly the result of large sedimentation from the Western Cordillera (Posada Posada et al., 2009) and the semidiurnal tidal range, with average heights between 2 and 4 m. A total river discharge to the Pacific Ocean of 254 km3 per year has been calculated, and a suspended sediment transport rate of 96 x 106 t yr-1 has been determined, corresponding to a sediment production of 1260 t km2 yr-1. The three rivers that contribute the most sediment to the Colombian Pacific coast are the Patía, the San Juan, and the Mira, accounting for over 40% of the total load (López and Restrepo, 2007).
For the Pacific study area, the Mira river was selected as the hydrological element forcing changes in the coastline due to the geographic location and distribution of its delta, of which the Tumaco study area is a part. The main drainage system is located at Cape Manglares and has seven distributaries or estuaries, smaller to the south and north of the main channel, which control the stability and evolution of the coastline into which they streamflow (Restrepo López and Otero Díaz, 2007) (Figure 5). Given the absence of rivers on San Andrés Island, the present analysis was conducted exclusively for Bocagrande, Tumaco.
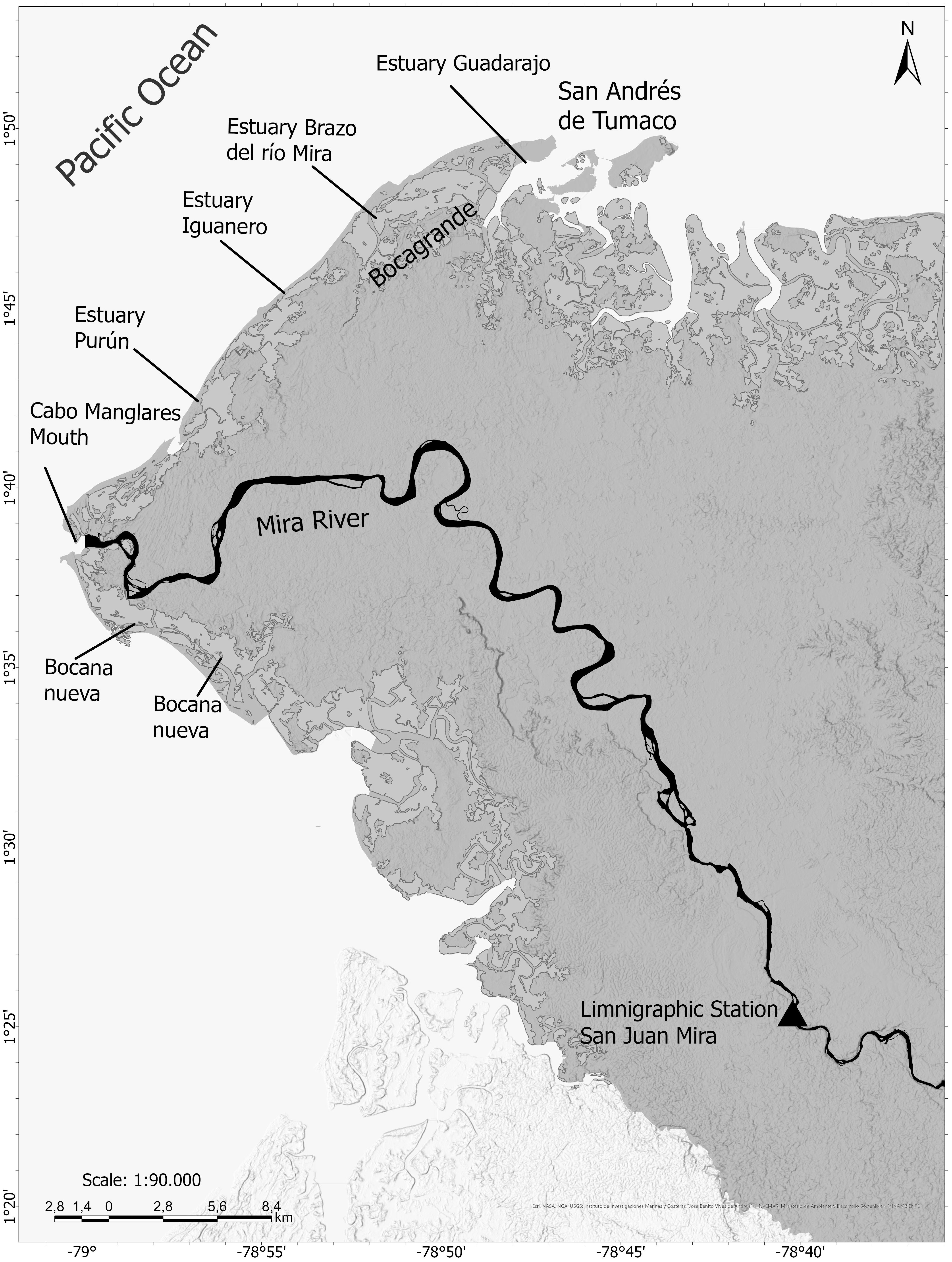
Figure 5. Mira River Delta, of which the Bocagrande sector is a part, and location of IDEAM’s San Juan Mira station from where the Mira River streamflow data was obtained.
The daily mean streamflow data for the present study were obtained from the Institute of Hydrology, Meteorology and Environmental Studies (IDEAM), specifically from the automatic technology limnigraphic station with telemetry called SAN JUAN MIRA - AUT [51027060], located at latitude 1.4239 N and longitude 78.6703 W, at 2 m above mean sea level.
The mean streamflow has a daily temporal resolution; consequently, total averages were calculated for the periods between the acquisition dates of each pair of consecutive satellite images used in the multitemporal analysis of changes in the mangrove front.
2.6 Data analysis
Simple and multiple linear regression analyses were performed between the rates of change for each pair of shorelines in consecutive years as a function of wave height and velocity, ONI, and mean river streamflow. Given that, under natural conditions, mangroves depend on propagule rooting to maintain their populations and colonize new areas, and since propagule establishment and growth into seedlings can take between 17 and 30 days depending on nutrient availability (Tovilla Hernández and Orihuela Belmonte, 2002), the rapid growth observed during the first three to four months after recruitment likely results from the use of nutrient reserves stored in the propagules and the contribution of foliar photosynthesis (de Oliveira Lima et al., 2018). Linear and multiple correlations were performed with averages of six months of data, before the start of each period. This approach was taken to consider the response time of R. mangle, according to the methodology proposed by Duke et al. (2022) (Figure 6).
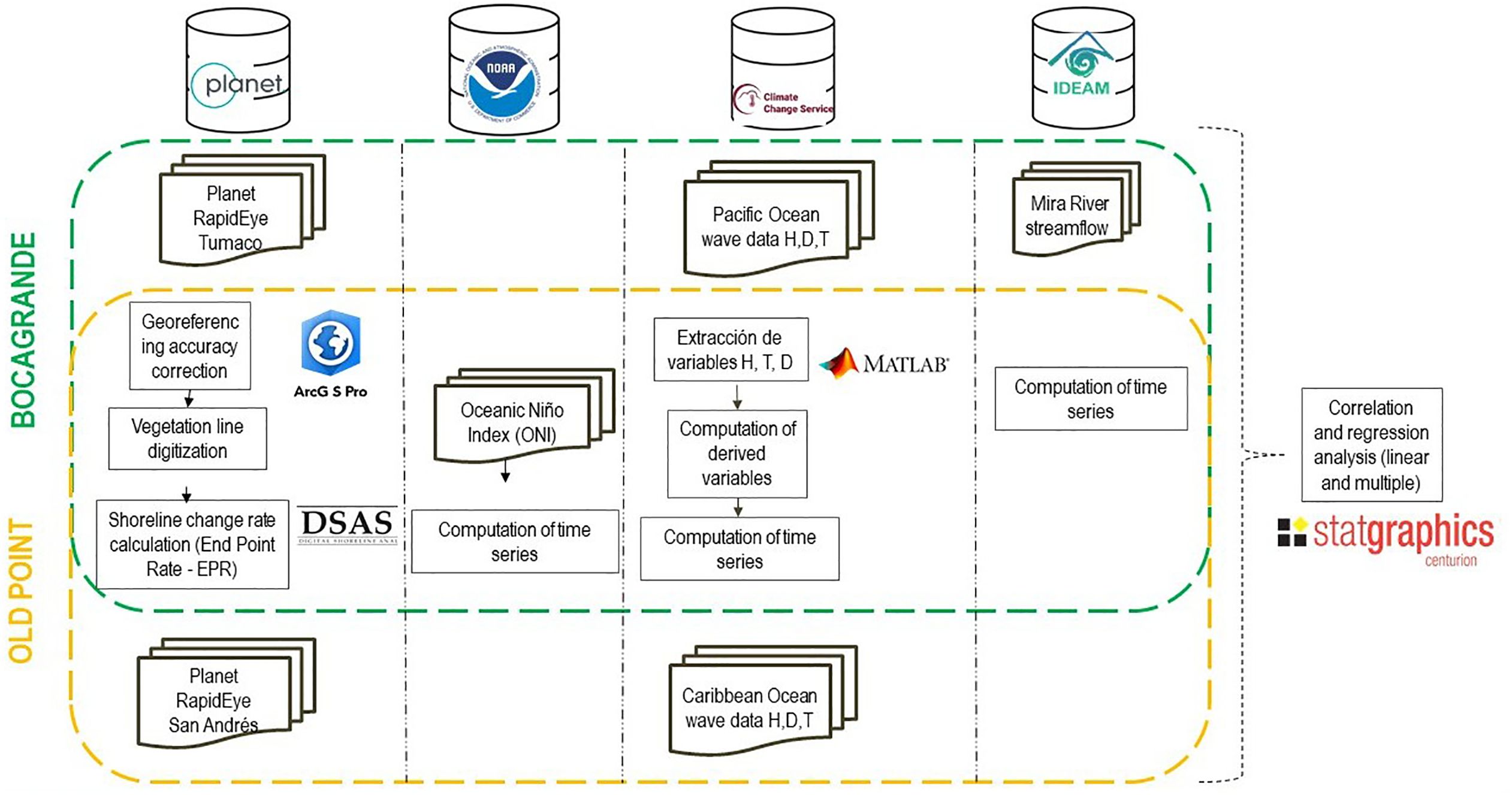
Figure 6. Methodological process applied to the data for each evaluated variable.
3 Results
3.1 Changes in forest front position
A multitemporal analysis of the mangrove front reveals high spatial variability over a short period. This variability in the forest front was observed in both regions studied (Figures 7A, B). Overall, from 2010 to 2023, the rate of change in mangrove front in the Bocagrande study area was negative, indicating an average retreat of 1.13 m yr-1. However, when analyzing each annual period, the rate of change showed great variability across all the periods evaluated, with a maximum retreat of the mangrove front of 19.75 m yr-1 between 2017 and 2018, and 18.05 m yr-1 between 2019 and 2020. The periods 2014–2015 and 2021–2022 had the greatest stability, while the periods 2016-2017, 2018-2019, and 2022–2023 showed the greatest progradation with rates of 9.18, 9.93, and 11.69 m yr-1, respectively. The results of the overall endpoint rates are shown in Table 2.
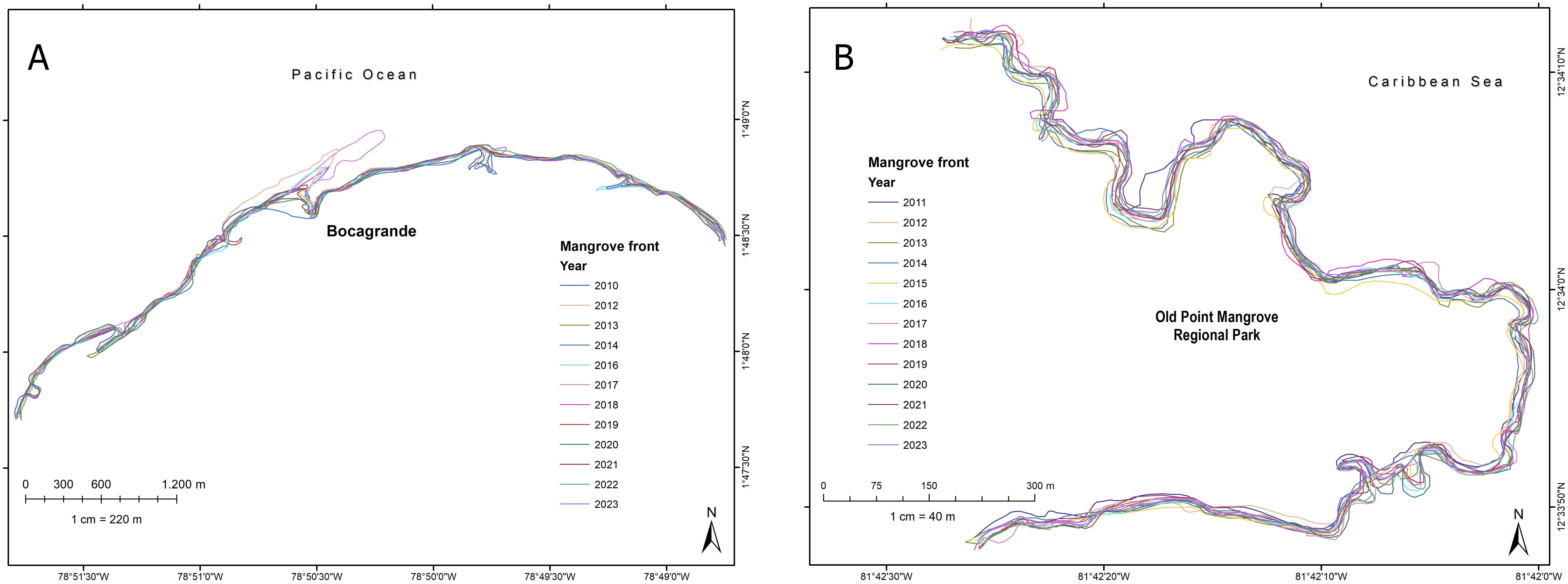
Figure 7. Vegetation front lines digitized from images of the Bocagrande coast (A) and the Old Point coast (B).
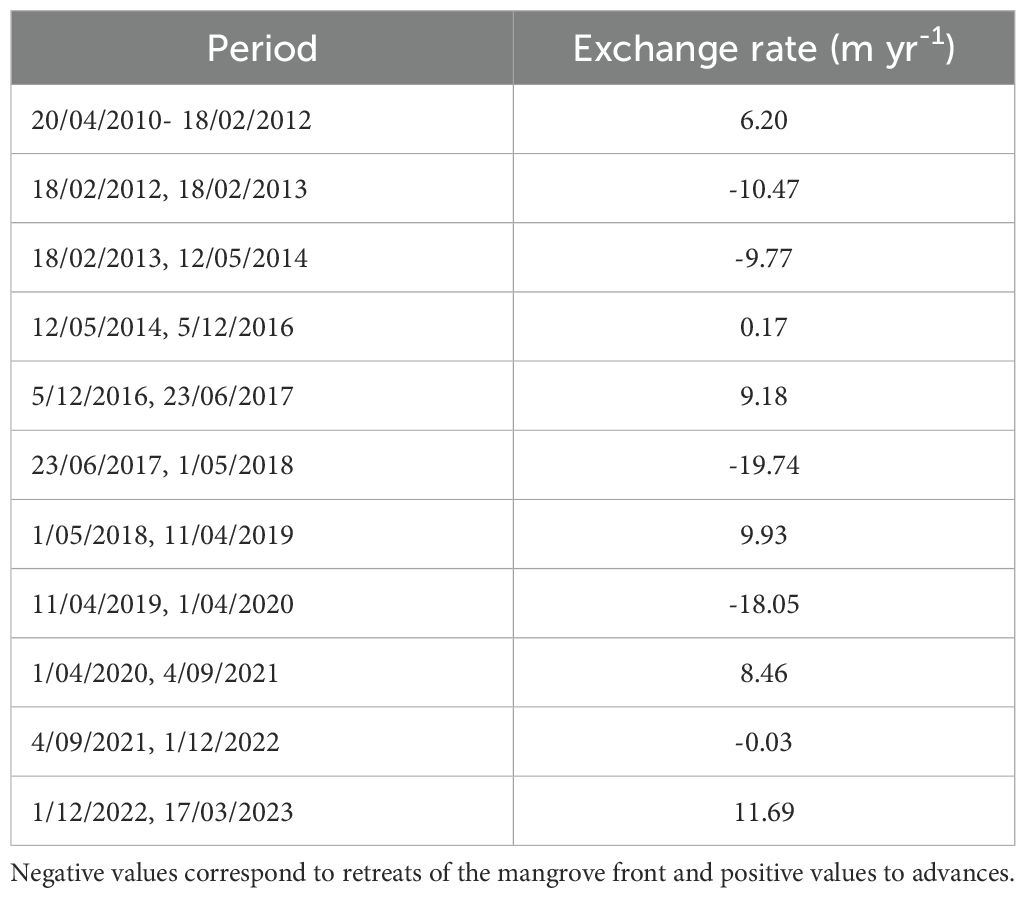
Table 2. Rate of movement or change in position of the mangrove front calculated for Bocagrande, Tumaco, from the spatial position of the mangrove front in different years.
The mangrove front changes in the insular Caribbean Island were similar to those of Bocagrande in terms of annual variability. However, unlike the Pacific, the overall trend from 2011 to 2023 was forest accretion in the three sectors, with a positive rate of change of 0.45 m yr-1 in the center; 0.05 m yr-1 in the north and 1.02 m yr-1 in the south. The southern zone showed the greatest variation despite being within a bay, while the central and particularly the northern zone showed much less variation in the forest front.
The years with the greatest forest retreat at Old Point were 2013–2014 and 2022–2023 in the central sector with 6.6 and 6.42 m yr-1 respectively; 2012–2013 in the northern area with 8.8 m yr-1; and 2020–2021 in the southern area with 6.76 m yr-1. The largest positive migrations occurred in 2012–2013 for the central area (7.57 m yr-1), 2020–2021 for the northern area (10.65 m yr-1), and 2012–2013 for the southern area (6.02 m yr-1). These results show that each sector of the same forest, depending on its orientation with respect to the incident waves or degree of exposure, exhibited variations with different patterns that did not follow a general trend (Table 3).
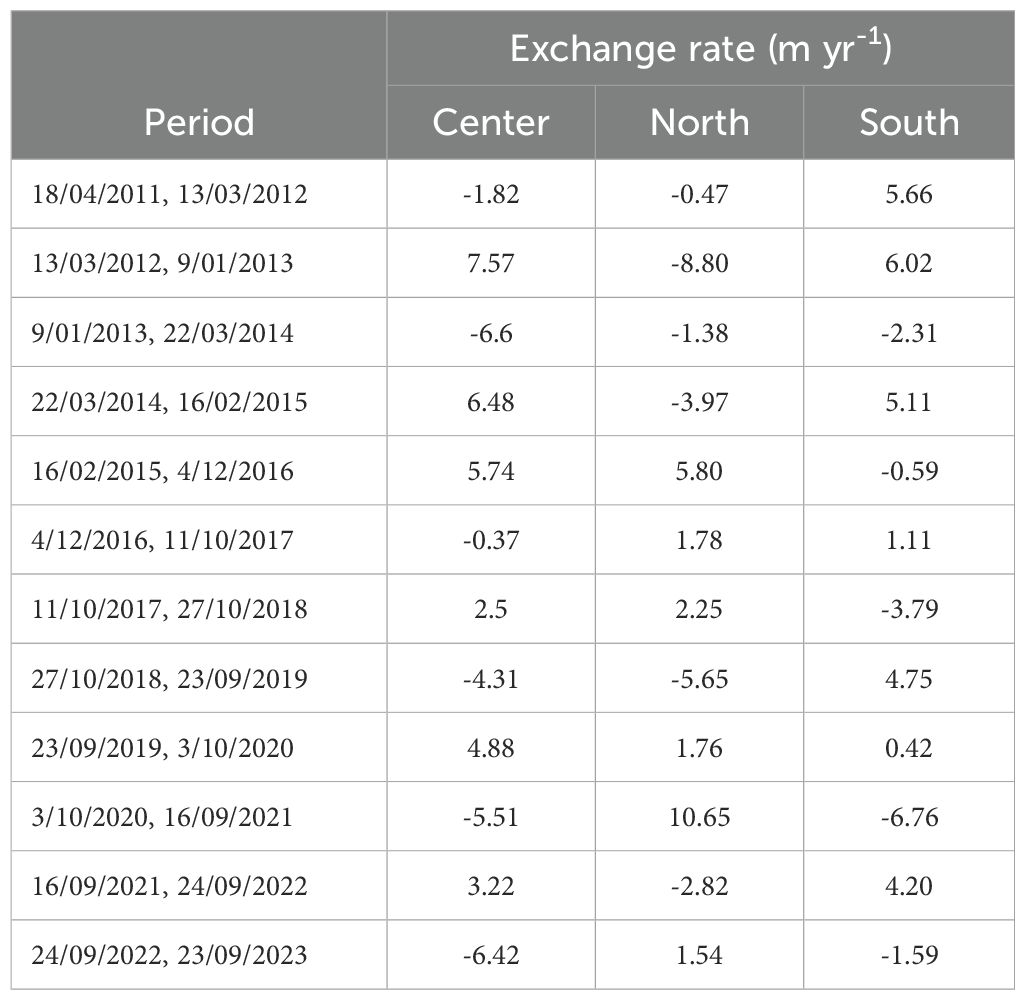
Table 3. Rate of movement or change of position of mangrove front calculated for Old Point, San Andrés Island, from the spatial position of the mangrove front in different years.
3.2 El Niño – Southern Oscillation
The linear correlation between the mangrove front shift in Bocagrande and ENSO yielded a p-value of 0.16, indicating that there is no statistically significant relationship at the 95.0% confidence level. Furthermore, the R-squared statistic indicates that the fitted model explains only 22.44% of the variability in mangrove front position and the correlation coefficient is equal to -0.47, indicating an inverse but relatively weak relationship between mangrove front migration and ENSO (Figure 8A). On San Andrés Island, the R-squared statistic indicates that the fitted model explains 22.16% of the variability in the forest front. The correlation coefficient is 0.16, indicating a relatively weak relationship between the variables. (Figure 8B).
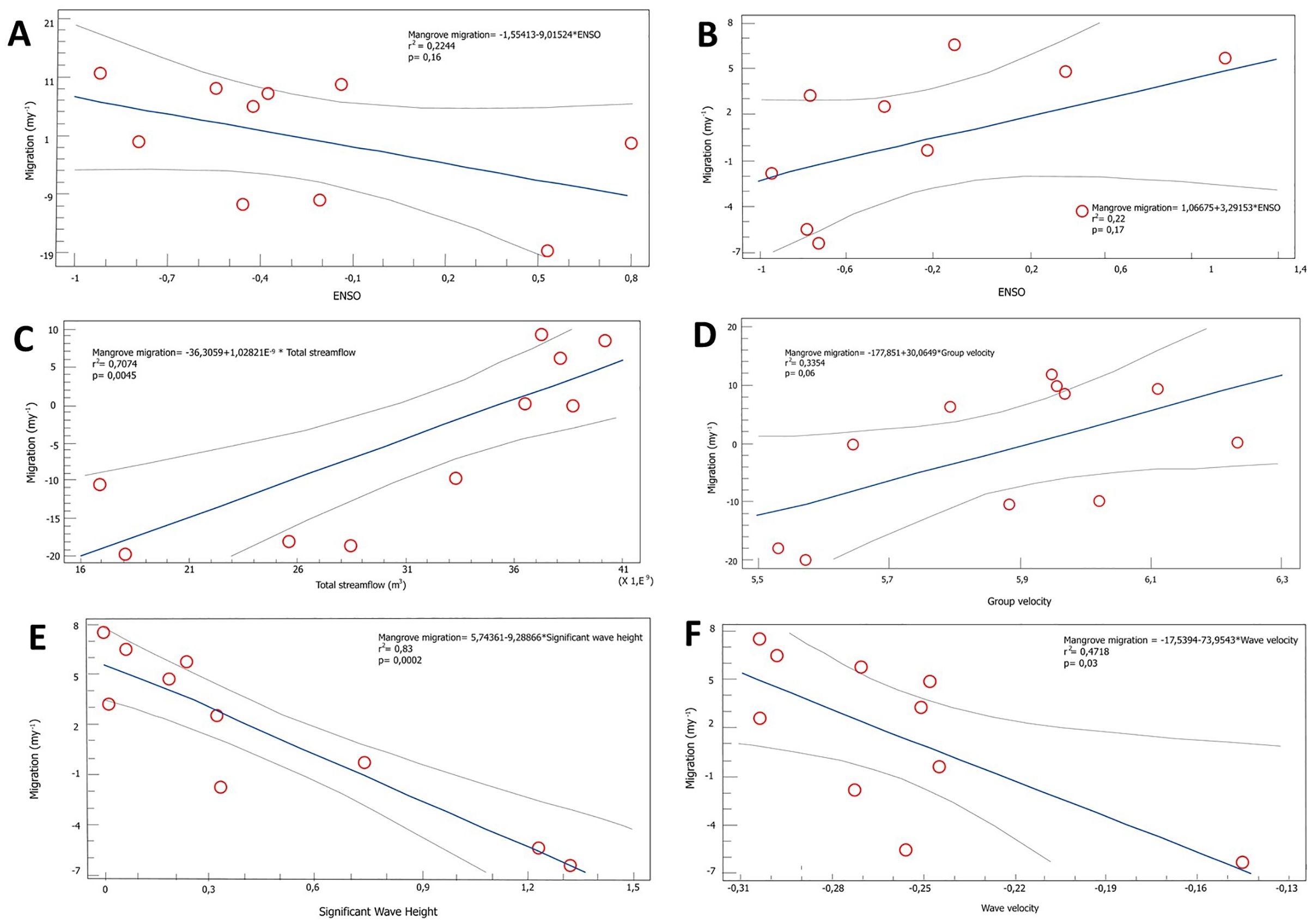
Figure 8. Results of the linear regression between the forcing variables and the response of the mangrove edge: (A) Bocagrande mangrove migration - ENSO, (B) Old Point mangrove migration - ENSO, (C) Bocagrande mangrove migration - Mira river total streamflow, (D) Bocagrande mangrove migration - Group velocity, (E) Old Point mangrove migration - Significant Wave Height, (F) Old Point mangrove migration - Wave velocity.
3.3 Mira river water discharge
In Bocagrande, the linear regression between the mangrove front change and the total streamflow (in m3) of the Mira River had a statistically significant relationship (p-value= 0.0045). The R-squared statistic indicates that the fitted model explains 70.74% of the variability in the position of the mangrove front, and the correlation coefficient of 0.84 indicates a moderately strong direct relationship between the two variables (Figure 8C).
3.4 Wave-related variables
It was found that wave velocity, group velocity, and significant wave height differentially explained changes in the mangrove front at the study sites (Table 4). In Bocagrande, the most explanatory variable was group velocity of waves, while in San Andrés, it was the wave velocity and significant wave height. There was no statistically significant relationship between changes in mangrove front position and group velocity of waves in Bocagrande. However, the p-value=0.06 and the R-square value indicate that the fitted model explains 33.54% of the variability in mangrove front changes. The correlation coefficient of 0.58 indicates a moderately strong relationship between the variables, with a significance level just above the threshold to reject the null hypothesis (Figure 8D).
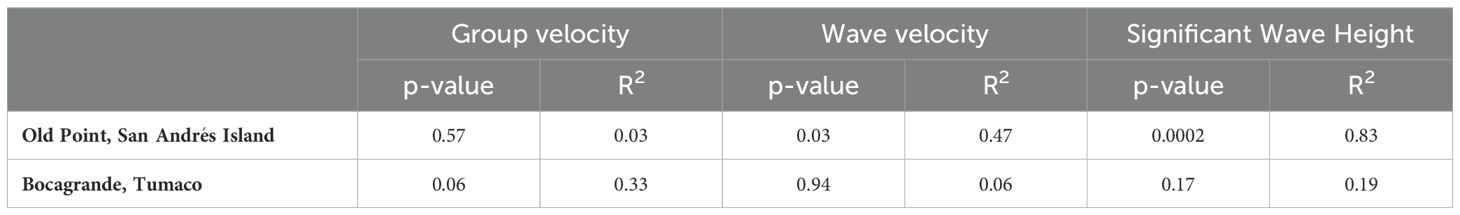
Table 4. Results of correlation between mangrove forest migration and wave-associated variables.
In San Andrés Island, the relationship between the change in the position of the mangrove front and the wave height was statistically significant with a confidence level of 95.0% and a p-value=0.0002. The R-squared statistic indicates that the fitted model explains 83.38% of the variability in the central side, and the correlation coefficient is equal to -0.91, indicating a relatively strong inverse relationship between the variables (Figure 8E).
On the other hand, the relationship between mangrove front change and wave velocity was statistically significant, indicating a moderately strong inverse relationship between the variables (95.0% confidence level; P=0.03; R-square = 47.18%) (Figure 8F).
3.5 Main drivers of mangrove front change
In San Andrés, significant wave height (P=0.0002) and ENSO (P=0.04) collectively explained 91.1% of the variability in mangrove front change in the central sector of Old Point, according to multiple linear regression analysis. Including wave velocity in the model (P = 0.54) did not increase considerably its explanatory power (R-squared = 91.7%). In Bocagrande, the explanatory power of the model reached 85.3% by including ENSO (P = 0.14), group velocity of waves (P = 0.14), and freshwater discharge as explanatory variables. However, only freshwater discharge significantly accounted for the observed variability (P = 0.02).
4 Discussion
The services provided by mangroves vary with forest type and location (Walters et al., 2008; Gilman et al., 2008; Ewel et al., 1998), and changes in mangrove cover have important implications for coastal areas, regardless of forest type. Mangrove loss can increase vulnerability to hazards such as erosion, flooding, waves and storm surges (Danielsen et al., 2005; Kathiresan and Rajendran, 2005; Dahdouh-Guebas et al., 2005); lead to a loss of biodiversity, as mangroves serves as refuge and breeding habitats for many species from different taxonomic groups (Malik et al., 2015); affect coastal water quality (Gilman et al., 2008); and, in general, generate significant negative impacts on human communities that depend on them directly and indirectly (Nagelkerken et al., 2008; Walters et al., 2008; Mumby et al., 2004; Ewel et al., 1998). This is likely the case for communities in the Colombian Pacific (López-Angarita et al., 2016). Therefore, the analysis of mangrove forest migration as a bioindicator of coastal processes is of great interest for both scientific understanding and environmental management.
4.1 Terrigenous sediments and mangrove frontal shifts
The supply of sediment by rivers regulates the relief and is considered one of the most dynamic geomorphological variables in the coastal landscape (Hooke, 1977; Gupta and Fox, 1974; Coleman, 1969; Turnbull et al., 1966). Thus, understanding fluvial processes is essential for comprehending geomorphological processes, such as erosion and progradation (Hooke, 1979). Deltaic mangroves are closely linked to their basins; the contributions of water, sediment and nutrients strongly influence ecosystem processes as well as the composition, structure and function of biological communities (Poff et al., 2006; Resh et al., 1988).
In Bocagrande, Tumaco, the greatest retreat of the vegetation front occurred during periods with the lowest freshwater discharge from the Mira River. Although the sources, sinks, and sediment fluxes in river systems vary greatly in both time and space (Trimble, 1999), reduced river discharge leads to lower sediment inputs (Florsheim et al., 2011). Between 2010 to 2023, the temporary reduction in the sediment load of the Mira river resulted in episodes of coastal erosion at its mouth, increasing the vulnerability of the mangroves and leading to the loss of cover due to the retreat of the front. During years of low freshwater discharge, sediment bars in front of mangroves are smaller (e.g. 2012). As a result, protection from waves and tidal currents diminishes, and mangrove cover contracts seaward probably due to defoliation and stress at minor erosion rates and by tree uprooting and death at strong erosion rates, as observed in other locations (Sánchez-Núñez et al., 2019). In contrast, mangrove progradation on Bocagrande coast was influenced by the availability of sediments discharged by the Mira River and by waves parallel to the coast that transport and distribute the sediments. During years of high discharge, sediment bars become larger, providing greater protection, and allowing mangrove foliage to recover and expand. Furthermore, sediment accumulation during favorable years aids in the development of propagules and seedlings, which eventually mature into juveniles and enhance sediment retention.
The position of the mangrove forest front on Bocagrande showed high variability, with periods of both progradation and retreat in response to the discharge of the Mira river and the redistribution of sediments. Similar findings have been observed in other Pacific regions, such as southern Thailand, where coasts near river mouths and sheltered bays experience higher rates of forest expansion, while erosion increases in areas when river contributions are reduced (Thampanya et al., 2006). River modification can have catastrophic consequences for mangroves, as recorded in a sector of the Colombian Pacific north of Tumaco, where approximately 5,200 ha of mangroves died due to the diversion of water from the Patía River to the Sanquianga river (Restrepo and Cantera, 2013).
The predominance of erosion over progradation in Bocagrande in terms of magnitude (34% larger), but not in the number of events (five versus six), indicates that while vegetation retreat processes can occur in very short periods of time, such as days, recovery, colonization and development processes require months to years (Teutli-Hernández et al., 2020).
On San Andrés, on the other hand, there are no permanent surface water flows, and only during rainy periods small drainage areas are formed on the western side of the island and some small channels drain the eastern side (Vargas Cuervo, 2004). In addition, the island corresponds to an open coast geomorphic setting (Hubach, 1956), characterized by oligotrophic waters with few nutrients (Geister and Díaz, 2007). Under these conditions, other factors other than the availability of terrigenous sediments are more relevant, as discussed in the following sections.
4.2 ENSO and changes in mangrove fronts
According to the annual average streamflow of the Mira river, from April 2021 to January 2022 the river reached its highest flow of 1,025.46 m3 s-1, coinciding with the La Niña phase of ENSO. In Colombia, this phase is characterized by intense rainfall, river flooding, avalanches, and flooding of plains in the Andean and Caribbean regions. Conversely, the lowest flow of the Mira river occurred between April 2009 and April 2010 with 634.35 m3 s-1, coinciding with the El Niño phase of ENSO, during which droughts and forest fires affect most of the territory, except for the southern Pacific region (Sánchez et al., 2001). A similar response was observed in the Atrato river Delta, where periods of erosion and progradation coexist in different regions, with a delayed coupling to the transitions between La Niña and El Niño phases, but stronger associations with the dry and rainy seasonal cycles. Erosion often coincides with dry seasons (Vásquez-Salazar et al., 2024). In the Gulf of Carpentaria, mangroves are more vulnerable during El Niño conditions due to droughts caused by reduced rainfall (Duke et al., 2022). The mangrove migration in response to the El Niño phase of ENSO in Bocagrande aligns with findings by Vos et al. (2021) on sandy beaches in the Pacific basin, where erosion rates predominated during El Niño periods in western North America, South America, Japan, and the Hawaiian Islands. Despite trends found between ENSO phases and mangrove front changes in this and other studies, the seasonal linear regression between the ONI index and the daily Mira river flows showed that ENSO does not have a significant relationship with the Mira river freshwater discharge.
The lack of a strong correlation between ENSO and Mira river discharge suggests that other climate variability drivers not assessed in this study may contribute to the river’s hydrological variability. low-intensity climatic variability processes, such as the Pacific Decadal Oscillation (PDO) or sea surface pressure (SLP) anomalies, could be such drivers, as found in the Magdalena (Restrepo López, 2014), Amazon (Amarasekera et al., 1997), Orinoco, Sao Francisco, and Tocantis rivers (García and Mechoso, 2009). Therefore, the weak correlation between mangrove front changes in Bocagrande and the ONI index in this study may be explained by a delayed response to ENSO events.
The contrasting results observed between Bocagrande and Old Point in the relationship between ENSO and mangrove front changes suggest different underlying mechanisms. In the open-water mangrove forest of Old Point, the cold phase of ENSO (La Niña) caused a retreat of the mangrove front, while in Bocagrande there was a progradation trend. During the La Niña phase, Old Point experiences higher rainfall, which leads to wet sediments that facilitate wave erosion. Similarly, in Cispatá Bay, higher erosion rates were found during La Niña compared to El Niño in fringe mangroves with low sediment availability (Data not published).
4.3 Waves, coral reef protection and changes in mangrove fronts
Hydrodynamic processes on the eastern side of San Andrés Island, driven by offshore waves, have different effects depending on the configuration of the reef barrier. The reef dissipates a large part of the incident wave energy coming from the east and northeast. The southeast side is less protected because the reef is smaller and even has an opening at the coordinates 81°41’40.35”W-12°32’13.859”N (Olarte Cáceres, 2019). Hurricanes can pass at relatively far distances from San Andrés island or relatively close as during November 2020. They can modify the usual wave energy and direction patterns around the island (Bernal et al., 2016). Thus, a mangrove retreat in the central sector of Old Point is expected if a hurricane increases wave energy from the south and south east as waves from these directions do not interact with the main reef barrier and its dissipation power.
In San Andrés, waves were more closely associated with mangrove front changes, compared to the result from the Pacific. Wave energy is one of the main drivers of erosion in mangrove environments with low sediment availability (Sánchez-Núñez et al., 2019). The impact of waves leads to the loss of finer, less dense particles and the accumulation of the heavier, coarser particles. However, these changes depend on the physical properties of the sediments, such as grain mass, density, size, morphology, as well as porosity, cohesion and fabric (Robert, 2009). Thus, the behavior of the wave-sediment relationship is different in the two study areas owing to the availability, origin and properties of their sediments.
On the other hand, the seaward colonization of propagules is challenging in Old Point due to the presence of a cliff at the mangrove front, which limits propagule establishment under conditions of high inundation and relatively high wave energy. The cliff results from sediment retention by prop roots. Its gradual, long-term expansion seaward may be attributed to the transport and retention of skeletal sediments of coral origin, non-skeletal sediments composed of grains produced by physicochemically induced carbonate precipitation, and, to a much lower extent, to sediments derived from terrestrial sources
Wave variables were a key driver of mangrove front changes in both study areas. On the one hand, on San Andrés Island, wave height drove forest edge expansion or retreat, while in Bocagrande, the opposite response occurred, given the high availability of sediments provided by the discharge of the Mira River; the greater the group velocity of waves, the more sediments accumulated, which may have favored R. mangle foliage recovery and colonization. These different behaviors between the Caribbean and Pacific forests are consistent with the results found by Sánchez-Núñez et al. (2019) in the mangrove forest of the Sinú river delta in the Colombian continental Caribbean at sites with low sediment availability. Low wave energy triggered accretion, while relatively high wave energy caused erosion.
In San Andrés, the expansion of mangroves is not linked to the growth of sediment bars in front of the mangroves fed by allochthonous sediments of fluvial, anthropogenic or shallow marine origin, as in the case of Bocagrande or other areas of the Colombian continental Caribbean, such as the Gulf of Urabá (Suárez et al., 2015). In Old Point, mangroves are directly exposed to waves, and mangrove expansion episodes observed in this study can be attributed to foliage recovery. Years of mangrove front retraction, caused by high wave energy from the south — which does not interact with the reef — are typically followed by years of expansion when wave energy decreases. Mangrove front expansion occurs by the horizontal growth of new branches generated from edge trees. Branches growing close to the sea surface can also produce prop roots, which help gain ground towards the sea. During strong wave events or years, leaves are more affected than prop roots, and during favorable years leaves can quickly recover from colonizing branches and prop roots. For instance between 2020-2021, the time interval when Hurricane Eta and Iota affected San Andrés, mangrove cover contracted by 5.5 m. The following year cover expanded by 3.2 m. Mangrove cover is assessed through the greenness signal, which reveals strong changes in the mangrove canopy due to wave action, despite the higher resilience of roots.
4.4 Geomorphic types, coastal environments, and coastal change
The geomorphic settings of the coasts studied undoubtedly influences the responses of the mangrove front. The mangrove forest of Old Point in San Andrés is protected from the direct action of the sea by a reef barrier, while in Bocagrande (Tumaco) the mangrove is protected by a sandbar that forms and disappears cyclically. The interaction between sediment availability, properties, accommodation space and the capacity of morphodynamic agents to transport and distribute them is the fundamental basis that conditions the evolution of coasts, determining their type, shape, structure, as well as their stability (Restrepo López, 2018). Moreover, these determinants largely depend on the origin of the sediments, whether fluvial, as in the case of Tumaco, whose sediments are transported by terrestrial drainage, or biological, as in the case of San Andrés, whose carbonate sediment is mainly of coral origin (Jackson and Short, 2020).
The results of this study show that different environmental factors influence the spatial arrangement of mangrove forests and that they respond relatively quickly to variations in these forces. The response varies according to the type of forest, its characteristic physiography and, in the case of San Andrés, according to the interaction of waves, extreme events, and coral reef heterogeneity.
Based on these findings, it is possible to conclude that the response of mangrove forests to the regulating and forcing factors of coastal change is diametrically different between the Pacific coast and the oceanic Caribbean islands (Figures 9A, B), so that prevention, response, and adaptation plans must be designed according to the type of forest and the environmental conditions that define it. The study, management, protection and effective restoration of mangrove ecosystems must take into account the biological, geographic, and physical differences of each region, as well as its specific responses to the dynamic and changing environment, in order to ensure their integrity and the provision of ecosystem services.
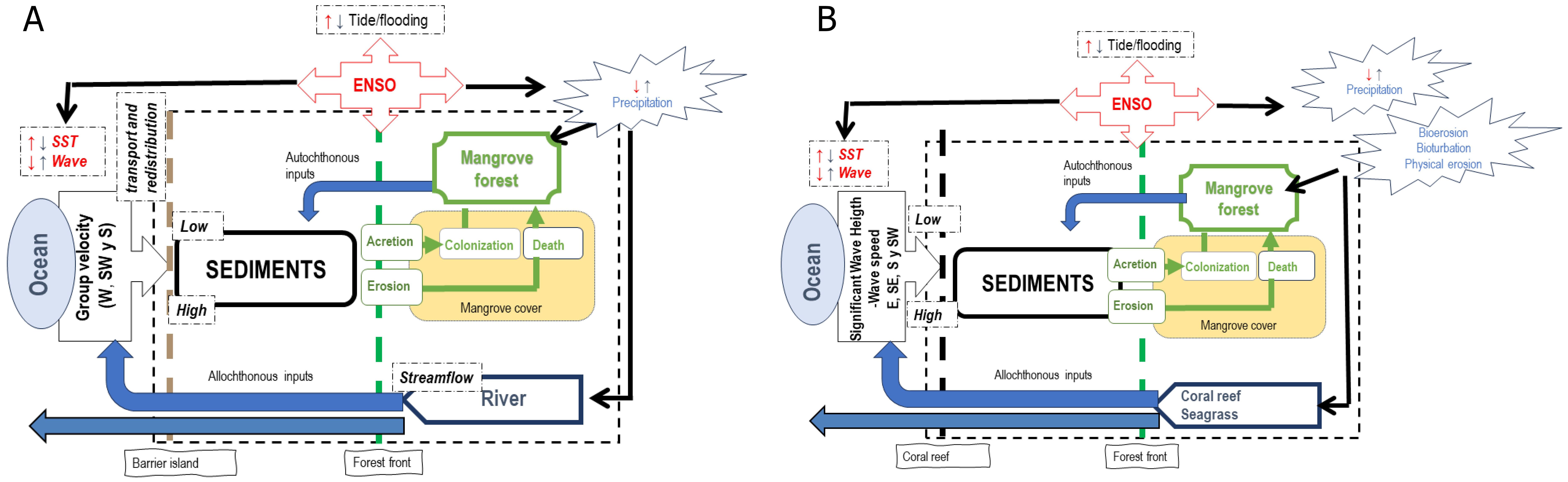
Figure 9. Diagram showing the ecosystem connectivity between mangroves and the environment. (A) Bocagrande, (B). Old Point.
Understanding the specific mechanisms that influence the adaptations of this ecosystem in different sedimentary environments and disturbance regimes will allow more accurate projections of the long-term impacts of forcing factors that influence both their ecosystem integrity (Krauss et al., 2013) and the provision of ecosystem services. Based on site-specific knowledge, it is possible to develop successful restoration strategies in ecosystem-based adaptation projects to mitigate mangrove loss due to erosion associated with sea-level rise, impacts resulting from changes in coastal sediment availability, wave climate and extreme events. Monitoring, using high-resolution imagery to map forest cover and early stages of colonization in rapidly changing landscapes, together with time-series data of environmental variables, can support evidence-based policy decisions on coastal migration in response to ecosystem forcing.
5 Final considerations
In the present study, it was found that in only 13 years of observation, there were constant changes in the position of the mangrove front, both open-water karst type in the insular Caribbean and of the deltaic type in the Colombian Pacific, which receives water from the Mira river. Although the net magnitudes of change were similar, the predominant direction of migration was not. While in the open water forest, the mangrove migrated towards the sea and gained cover, in the deltaic mangrove, there was a retreat.
The results of this study showed that while it is true that environmental drivers shape mangrove front changes, the magnitude and direction of such changes varies between regions and depends on local hydro-morphodynamic conditions. The documented changes in the migration of the two types of mangroves support the hypothesis and showed that there is no single mangrove response trend and that different types of mangroves respond quickly and differently to coastal disturbance factors.
The mechanisms involved in the response of R. mangle forests were varied in this study. The sediment dynamics provided by the Mira river is the main driver of the migration of the deltaic mangrove in Tumaco, dominating the mesotidal regime and the high rainfall characteristic of the Colombian Pacific region. In contrast, waves and ENSO are the main drivers of the migration dynamics of the open-water mangrove on San Andrés Island, a microtidal mangrove without river input, where sediments come directly from the sea or are of autochthonous origin.
Just as mangrove forest cover is affected by drivers of change, so too is the provision of ecosystem services. The service of coastal erosion mitigation varies in response to the changes the forest undergoes in response to these drivers. Similarly, the ecosystem service of carbon capture and storage is directly affected. The degradation of the areas covered by these forests not only reduces the density of carbon to be stored but also releases into the atmosphere large reserves of blue carbon that were previously sequestered mainly in sediments.
Analyzing, understanding, and monitoring these responses and identifying local, site-specific trends in mangrove forest front change is necessary to understand coastal processes and define timely and successful management strategies in the areas of the country where this ecosystem is present, together with their coastal landscape and seascape. For example, if the Bocagrande mangrove depends directly on the sediment supply from the Mira river, a reduction in these due to natural or anthropogenic changes in the river will reduce the resilience of the forest. In addition, coral reef health, openings and unprotected coral reef areas drive changes in the mangrove front, given their interactions with wave energy. The results of this study provide a positive contribution to mangrove ecosystem restoration plans in the phases of prioritization of areas with restoration potential, identification of stressors and disturbance regimens that cause ecosystem degradation, and identification and implementation of restoration actions that address stressor and disturbances. They could also be considered in broader public policies such as coastal marine management, marine spatial planning and multipurpose cadastre.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
JE: Conceptualization, Data curation, Formal Analysis, Investigation, Methodology, Writing – original draft, Writing – review & editing. JM: Conceptualization, Investigation, Methodology, Supervision, Validation, Writing – original draft, Writing – review & editing. DS: Conceptualization, Data curation, Investigation, Methodology, Supervision, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research was funded by the Ministry of Science and Technology of Colombia -MinCiencias. Project “ Reguladores del servicio de control de la erosión provisto por manglares en diferentes ambientes costeros continentales e insulares del Caribe y Pacífico Colombiano” (Hermes code 46804 and MinCiencias code 71299). Program “ Evaluación comparativa de servicios ecosistémicos proporcionados por diferentes tipos de manglar en Colombia” (code 110685270796).
Acknowledgments
We thank professors Adriana Santos-Martínez and Jairo Medina from the Universidad Nacional de Colombia, Caribbean campus and Botanical Garden. We also thank Professors Jaime Cantera and Enrique Peña of the Universidad del Valle, directors of the program. Our appreciation for all the support provided by the Universidad Nacional de Colombia, Caribbean and Bogotá campuses.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Álvarez-Silva O., Saavedra V., Otero L., and Restrepo J. C. (2022). On the mechanisms controlling near-coast circulation in the southern Colombian Pacific at tidal, seasonal, and interannual time scales. J. Marine Systems. 236, 1–15. doi: 10.1016/j.jmarsys.2022.103804
Amarasekera K. N., Lee R. F., Williams E. R., and Eltahir E. A. (1997). ENSO and the natural variability in the flow of tropical rivers. J. Hydrology. 200, 24–39. doi: 10.1016/S0022-1694(96)03340-9
Babanin A. V., Rogers W. E., de Camargo R., Doble M., Durrant T., Filchuk K., et al. (2019). Waves and swells in high wind and extreme fetches, measurements in the southern ocean. Front. Marine Science. 6. doi: 10.3389/fmars.2019.00361
Bartholomew H. and Menglin S. J. (2013). ENSO effects on land skin temperature variations: A global study from satellite remote sensing and NCEP/NCAR reanalysis. Climate 1, 53–73. doi: 10.3390/cli1020053
Bernal G., Osorio A. F., Urrego L., Peláez D., Molina E., Zea S., et al. (2016). Occurrence of energetic extreme oceanic events in the Colombian Caribbean coasts and some approaches to assess their impact on ecosystems. J. Marine Syst. 164, 85–100. doi: 10.1016/j.jmarsys.2016.08.007
Bunting P., Rosenqvist A., Hilarides L., Lucas R. M., Thomas N., Tadono T., et al. (2022). Global mangrove extent change 1996–2020: global mangrove watch version 3.0. Remote Sens 14, (3657). doi: 10.3390/rs14153657
Coleman J. M. (1969). Brahmaputra River: channel processes and sedimentation. Sedi- ment. Geol. 3, 129–239. doi: 10.1016/0037-0738(69)90010-4
Costa P., Dórea A., Mariano-Neto E., and Barros F. (2015). Are there general spatial patterns of mangrove structure and composition along estuarine salinity gradients in Todos os Santos Bay? Estuarine Coastal Shelf Sci. 166, 1–9. doi: 10.1016/j.ecss.2015.08.014
Dahdouh-Guebas F., Jayatissa L. P., Di Nitto D., Bosire J. O., Lo Seen D., and Koedam N. (2005). How effective were mangroves as a defence against the recent tsunami? Curr. Biol. 15, R443–R447. doi: 10.1016/j.cub.2005.06.008
Danielsen F., Sørensen M. K., Olwig M. F., Selvam V., Parish F., Burgess N. D., et al. (2005). The Asian tsunami: a protective role for coastal vegetation. Science. 310, 643–643. doi: 10.1126/science.1118387
de Oliveira Lima K. O., Pereira Tognella M. M., Cunha S. R., and de Andrade H. A. (2018). Growth models of Rhizophora mangle L. seedlings in tropical southwestern atlantic. Estuarine Coastal Shelf Sci. 207, 1–29. doi: 10.1016/j.ecss.2018.03.021
Duke N. C., Mackenzi J. R., Canning A. D., Hutley L. B., Bourke A. J., Kovacs J. M., et al. (2022). ENSO-driven extreme oscillations in mean sea level destabilise critical shoreline mangroves-An emerging threat. PloS Climate. 1, 1–23. doi: 10.1371/journal.pclm.0000037
Echeverry Hernández J. P. (2025). “Tesis de Maestría en Ciencias-Biología,” in Vulnerabilidad del litoral Pacífico Colombiano a la erosión costera: Bosques de manglar como ecosistemas mitigadores (Bogotá, Colombia: Universidad Nacional de Colombia). (Accessed 12 January 2025).
European Commision, Copernicus, European Centre for Medium-Range Weather Forecasts, and Climate Change Service (2023). ERA5 hourly data on single levels from 1940 to present. Available online at: https://cds.climate.copernicus.eu/cdsapp!/dataset/reanalysis-era5-single-levels?tab=overview (Accessed October 28, 2023).
European Space Agency. (2024). Earth Online. Available at: About RapidEye: https://earth.esa.int/eogateway/missions/rapideye (Accessed August 10, 2024).
Ewel K., Twilley R., and Ong J. (1998). Different kinds of mangrove forests provide different goods and services. Global Ecol. Biogeography Letters. 7, 83–94. doi: 10.2307/2997700
Federal Emergency Management Agency and US Department of Homeland Security (2022). FEMA Region I Coastal Erosion Study – Rockingham County (Arlington, Virginia: Compass).
Ferrario F., Beck M. W., Storlazzi C. D., Micheli F., Shepard C. C., and Airoldi1 L. (2014). The effectiveness of coral reefs for coastal hazard risk reduction and adaptation. Nat. Communications. 5, 1–9. doi: 10.1038/ncomms4794
Florsheim J. L., Pellerin B. A., Oh N. H., Ohara N., Bachand P. A. M., Bachand S. M., et al. (2011). From deposition to erosion: Spatial and temporal variability of sediment sources, storage, and transport in a small agricultural watershed. Geomorphology 132, 272–286. doi: 10.1016/j.geomorph.2011.04.037
Food and Agriculture Organization of the United Nations (2023). The World’s Mangroves 2000-2020 (Rome).
Friess D. A., Rogers K., Lovelock C. E., Krauss K. W., Hamilton S. E., Lee S. Y., et al. (2019). The state of the world’s mangrove forests: past, present, and future. Annu. Rev. Environ. Resources. 44, 89–115. doi: 10.1146/annurev-environ-101718-033302
Gairin E., Collin A., James D., Maueau T., Roncin Y., Lefort L., et al. (2021). Spatiotemporal trends of Bora Bora’s shoreline classification and movement using high-resolution imagery from 1955 to 2019. Remote sensing. 13, 2–18. doi: 10.3390/rs13224692
García N. O. and Mechoso C. R. (2009). Variability in the discharge of South American rivers and in climate. Hydrological Sci. J. 50, 459–478. doi: 10.1623/hysj.50.3.459.65030
Geister J. and Díaz J. (2007). Reef environments and Geology of an Oceanic Archipelago: San Andrés, Old Providence and Santa Catalina (Caribbean Sea, Colombia). Available online at: https://boris.unibe.ch/id/eprint/85537 (Accessed January 20, 2025).
Ghosh S., Bakshi M., Gupta K., Mahanty S., Bhattacharyya S., and Chaudhuri P. (2020). A preliminary study on upstream migration of mangroves in response to changing environment along River Hooghly, India. Marine Pollution Bull. 151, 1–14. doi: 10.1016/j.marpolbul.2019.110840
Gilman E. L., Ellison J., Duke N. C., and Field C. (2008). Threats to mangroves from climate change and adaptation options: A review. Aquat. Botany. 89, (2)237–(2)250. doi: 10.1016/j.aquabot.2007.12.009
Glantz M. H. and Ramirez I. (2020). Reviewing the Oceanic Niño Index (ONI) to enhance societal readiness for El Niño’s impacts. Int. J. Disaster Risk Science. 11, 394–403. doi: 10.1007/s13753-020-00275-w
Global Mangrove Watch (2024). Global Mangrove Watch. Available online at: https://www.globalmangrovewatch.org/ (Accessed 15 July 2024).
Gomez Garcia L. F. (2024). “Tesis de Maestría en Ciencias-Biología,” in Reservas de carbono orgánico en manglares del Pacífico Colombiano y su relación con el grado de intervención antrópica (Universidad Nacional de Colombia, Bogotá).
Gupta A. and Fox H. (1974). Effects of high-magnitude floods on channel forms - a case study in Maryland Piedmont. Water Resour. Res. 10, 499–509. doi: 10.1029/WR010i003p00499
Hickey S., Radford B., Callow J., Phinn S., Duarte C., and Lovelock C. (2021). ENSO feedback drives variations in dieback at a marginal mangrove site. Sci. Rep. 11, 8130. doi: 10.1038/s41598-021-87341-5
Himmelstoss E. A., Henderson R. E., Kratzmann M. G., and Farris A. (2021). Digital Shoreline Analysis System (DSAS) version 5.1 user guide. Open-File Report 2021–109 (Reston, Virginia: U.S. Geological Survey).
Hooke J A. (1979). An analysis of river bank erosion processes. Journal of Hydrology 42 (1-2), 39–62. doi: 10.1016/0022-1694(79)90005-2
Hooke J. M. (1977). “The distribution and nature of changse in river channel patterns,” in River Channer Changes. Ed. Gregory K. J. (Wiley, London), 265–280.
Horstman E., Dohmen-Janssen C., Narra P., van den Berg N., Siemerink M., and Hulscher S. (2014). Wave attenuation in mangroves: A quantitative approach to field observations. Coastal Eng. 94, 47–62. doi: 10.1016/j.coastaleng.2014.08.005
Hubach E. (1956). Aspectos geográficos y geológicos y recursos de las islas de San Andrés y Providencia (Bogotá: Sociedad Geográfica de Colombia).
Instituto de Investigaciones Marinas y Costeras José Benito Vives de Andréis (2003). Programa holandés de asistencia para estudios de cambio climático: Colombia. Definición de la vulnerabilidad de los sistemas bio-geofísicos y socioeconómicos debido a un cambio en el nivel del mar en la zona costera Colombiana y adaptación (Santa Marta: INVEMAR).
Jonah F., Boateng I., Osman A., Shimba M., Mensah E., Adu-Boahen K., et al. (2016). Shoreline change analysis using end point rate and net shoreline movement statistics: An application to Elmina, Cape Coast and Moree section of Ghana’s coast. Regional Stud. Marine Sci. 7, 19–31. doi: 10.1016/j.rsma.2016.05.003
Kathiresan K. and Rajendran N. (2005). Coastal mangrove forests mitigated tsunami. Estuarine Coastal Shelf Science. 65, 601–606. doi: 10.1016/j.ecss.2005.06.022
Krauss K. W., McKee K. L., Lovelock C. E., Cahoon D., Saintilan N., Reef R., et al. (2013). How mangrove forests adjust to rising sea level. New Phytologist. 202, 19–34. doi: 10.1111/nph.12605
Lillesand T., Kiefer R., and Chipman J. (2015). Remote Sensing and Image Interpretation (Medellín, Colombia: Wiley).
López S. A. and Restrepo J. D. (2007). Modelos morfodinámicos de los deltas fluviales Colombianos. Boletín Geología 29, 103–113. Available at: https://revistas.uis.edu.co/index.php/revistaboletindegeologia/article/view/845 (Accessed November 15, 2024).
López-Angarita J., Roberts C. M., Tilley A., Hawkins J. P., and Cooke R. G. (2016). Mangroves and people: Lessons from a history of use and abuse in four Latin American countries. For. Ecol. Management. 368, 151–162. doi: 10.1016/j.foreco.2016.03.020
Lovelock C. E., Krauss K. W., Osland M. J., Reef R., and Ball M. C. (2016). “The physiology of mangrove trees with changing climate,” in Tropical Tree Physiology. Adaptations and Responses in a Changing Environment. Eds. Goldstein G. and Santiago L. S. (Springer, Switzerland), 149–179. doi: 10.1007/978-3-319-27422-5_7
Malik A., Fensholt R., and Mert O. (2015). Mangrove exploitation effects on biodiversity and ecosystem services. Biodiversity Conserv. 24, 3543–3557. doi: 10.1007/s10531-015-1015-4
Mancera Pineda J. E. (2022). “Bosques de manglar,” in En: Colombia país de bosques. Eds. Rodríguez M. and Valdés M. F. (Alphaeditorial, CIENCIA Y TECNOLOGÍA, Bogotá, D.C), 426, ISBN: 9789587787368.
Medina-Calderón J. H., Mancera-Pineda J. E., Castañeda-Moya E., and Rivera-Monroy V. H. (2021). Hydroperiod and salinity interactions control mangrove root dynamics in a karstic oceanic island. Front. Marine Science. 7. doi: 10.3389/fmars.2020.598132
Mishra M., Sudarsan D., Kar D., Naik A. K., Das P. P., Santos C., et al. (2020). The development and research trend of using DSAS tool for shoreline change analysis: A scientometric analysis. J. Urban Environ. Engineering. 14, 69–77. doi: 10.4090/juee.2020.v14n1.069077
Monroy L. ,. C., Restrepo J. D., and López Isaza S. A. (2008). “Morfodinámica del delta del río Mira,” in Deltas de Colombia: morfodinámica y vulnerabilidad ante el Cambio Global. Ed. Restrepo Ángel J. D. (Fondo Editorial Universidad EAFIT, Medellín), 181–205.
Mumby P. J., Edwards A. J., Arias-González J. E., Lindeman K. C., Blackwell P. G., Gall A., et al. (2004). Mangroves enhance the biomass of coral reef fish communities in the Caribbean. Nature. 427, 533–536. doi: 10.1038/nature02286
Nagelkerken I., Blaber S., Bouillon S., Green P., Haywood M. D., Kirton L. G., et al. (2008). The habitat function of mangroves for terrestrial and marine fauna: A review. Aquat. Botany. 89, 155–185. doi: 10.1016/j.aquabot.2007.12.007
NOAA (2023). Cold & Warm Episodes by Season. Available online at: https://origin.cpc.ncep.noaa.gov/products/analysis_monitoring/ensostuff/ONI_v5.php (Accessed October 28, 2023).
Olarte Cáceres P. M. (2019). Tesis de Maestría en Ingeniería de Recursos Hidráulicos: Climatología del transporte potencial de sedimentos costeros inducido por oleaje en la Isla San Andrés (Medellín: Universidad Nacional de Colombia, Sede Medellín). Available at: https://repositorio.unal.edu.co/handle/unal/77571 (Accessed July 30, 2024).
Oyedotun T. (2014). Shoreline Geometry: DSAS as a Tool for Historical Trend Analysis. Geomorphological Techniques, Chap. 3, Sec. 2.2, 1-12. British Society for Geomorphology.
Planet Labs (2023). Planet Analytic Imagery and Archive. Available online at: https://www.planet.com/products/planet-imagery/ (Accessed 23 September 2024).
Poff N. L., Bledsoe B. P., and Cuhaciyan C. O. (2006). Hydrologic variation with land use across the contiguous United States: geomorphic and ecological consequences for stream ecosystems. Geomorphology 79, 264–285. doi: 10.1016/j.geomorph.2006.06.032
Pogson M. and Smith P. (2015). Effect of spatial data resolution on uncertainty. Environ. Modelling Software 63, 87–96. doi: 10.1016/j.envsoft.2014.09.021
Pokhrel P., Ioup E., Simeonov J., Hoque M. T., and Abdelguerfi M. (2022). A transformer-based regression scheme for forecasting significant wave heights in oceans. IEEE J. Oceanic Eng. 47 (4), 1010–1023. doi: 10.1109/JOE.2022.3173454
Posada Posada B. O., Henao Pineda W., and Guzman Ospitia G. (2009). Diagnóstico de la erosión y sedimentación en la zona costera del Pacífico Colombiano (Santa Marta: INVEMAR). (Serie de publicaciones especiales No. 17 ed.).
Quintero A. P. (2025). “Tesis de Maestría en Ciencias-Biología,” in Influencia de la salinidad en la acumulación de carbono en bosques de manglar sin subsidios externos de materia orgánica (Universidad Nacional de Colombia, San Andrés, Isla).
Raw J. L., Godbold J. A., van Niekerk L., and Adams J. B. (2019). Drivers of mangrove distribution at the high-energy, wave-dominated, southern African range limit. Estuarine Coastal Shelf Sci. 226, 1–8. doi: 10.1016/j.ecss.2019.106296
Resh V. H., Brown A. V., Covich A. P., Gurtz M. E., Li H. W., Minshall G. W., et al. (1988). Role of disturbance in stream ecology. J. North Am. Benthological Soc. 7, 433–455. doi: 10.2307/1467300
Restrepo J. D. and Cantera J. R. (2013). Discharge diversion in the Patía River delta, the Colombian Pacific: Geomorphic and ecological consequences for mangrove ecosystems. J. South Am. Earth Sci. 46, 183–198. doi: 10.1016/j.jsames.2011.04.006
Restrepo López J. C. (2014). Tesis doctoral en Ciencias del Mar: Dinámica Sedimentaria en Deltas Micromareales – Estratificados de Alta Descarga: Delta del Rio Magdalena (Colombia – Mar Caribe) (Barranquilla: Universidad del Norte). Available at: https://manglar.uninorte.edu.co/handle/10584/7422 (Accessed April 30, 2023).
Restrepo López J. C. (2018). “Las montañas construyen las costas y los deltas,” in Arrastrando la Montaña al mar. Ed. Restrepo Ángel J. D. (Agenda del Mar), 81–91.
Restrepo López J. C. and Otero Díaz L. J. (2007). Aplicación de modelos hidrodinámicos para evaluar la dinámica del oleaje y el nivel del mar en el sistema deltaico del río Mira (Colombia): Bases para la gestión costera. Boletín Científico CCCP. 14, 31–48. doi: 10.26640/01213423.14.31_48
Restrepo López J. C., Otero Díaz L., and López Isaza S. A. (2009). Clima de oleaje en el Pacífico sur de Colombia, delta del río Mira: Comparaciones estadísticas y aplicación a procesos costeros. Rev. la Academia Colombiana Cienc. Exactas Físicas y Naturales. 33, 357–376. doi: 10.18257/raccefyn.33(128).2009.2369
Restrepo López J. C., Otero Díaz L., López Isaza S. A., and Monroy L. ,. C. (2008). “Los deltas y el oleaje: estadísticas, clima, corrientes, energía y procesos de asomeramiento en el delta del río Mira,” in Deltas de Colombia: morfodinámica y vulnerabilidad ante el Cambio Global, vol. 33 . Ed. Restrepo Ángel J. D. (Fondo Editorial Universidad EAFIT, Medellín), 181–205.
Robert C. M. (2009). Global sedimentology of the ocean and interplay between geodynamics and paleoenvironment Vol. 3 (Hungary: Elsevier).
Rodríguez-Rodríguez J. A., Mancera-Pineda J. E., and Tavera H. (2021). Mangrove restoration in Colombia: Trends and lessons learned. For. Ecol. Manage. 496, 1-11. doi: 10.1016/j.foreco.2021.119414
Sánchez R., Vargas G., González H., and Pabón D. (2001). Los fenómenos cálido del Pacífico (EL Niño) y frío del Pacífico (La Niña) y su incidencia en la estabilidad de las laderas en Colombia. Available online at: https://repository.agrosavia.co/handle/20.500.12324/18814 (Accessed January 15, 2024).
Sánchez-Núñez D. A., Bernal G., and Mancera Pineda J. E. (2019). The relative role of mangroves on wave erosion mitigation and sediment properties. Estuaries Coasts. 42, 2124–2138. doi: 10.1007/s12237-019-00628-9
Sánchez-Núñez D. A. and Mancera-Pineda J. E. (2011). Flowering patterns in three neotropical mangrove species: Evidence from a Caribbean Island. Aquat. Botany. 94, 177–182. doi: 10.1016/j.aquabot.2011.02.005
Simard M., Fatoyinbo L., Thomas N., Stovall A., Parra A., Barenblitt A., et al. (2025). A new global mangrove height map with a 12 meter spatial resolution. Nature 12 (15), 1–13. doi: 10.1038/s41597-024-04213-z
Spalding M., McIvor A., Tonneijck F., Tol S., and van Eijk P. (2014). Mangrove for coastal defense. Guidelines for coastal managers & policy makers (Netherlands: Wetlands International and The Nature Conservancy). Available at: https://www.wetlands.org/publication/mangroves-for-coastal-defence/ (Accessed February 18, 2022).
Suárez J. A., Urrego L. E., Osorio A., and Ruiz H. Y. (2015). Oceanic and climatic drivers of mangrove changes in the Gulf of Urabá, Colombian Caribbean. Latin Am. J. Aquat. Res. 43, 972–985. doi: 10.3856/vol43-issue5-fulltext-17
Syvitski J. P., Asprey K. W., Clattenburg D. A., and Hodge G. D. (1985). The prodelta environment of a fjord: suspended particle dynamics. Sedimentology. 32, 83–107. doi: 10.1111/j.1365-3091.1985.tb00494.x
Teutli-Hernández C., Herrera-Silveira J. A., Cisneros-de la Cruz D. J., and Roman-Cuesta. R. (2020). “Guía para la restauración ecológica de manglares: Lecciones aprendidas,” in Proyecto, Mainstreaming Wetlands into the Climate Agenda: A multi-level approach (SWAMP) México: SWAMP. doi: 10.17528/cifor/007794
Thampanya U., Vermaat J. E., Sinsakul S., and Panapitukkul N. (2006). Coastal erosion and mangrove progradation of Southern Thailand. Estuarine. Coastal Shelf Science. 68, 75–85. doi: 10.1016/j.ecss.2006.01.011
Torres M. V., Federico G., and Collins P. A. (2016). ‘La Niña’ phenomenon and the relationship between decapod populations and fishes in temporarily isolated shallow lakes. Marine Freshwater Res. 68, 1010–1022. doi: 10.1071/MF16035
Tovilla Hernández C. and Orihuela Belmonte D. E. (2002). Supervivencia de Rhizophora mangle L. en el manglar de Barra de Tecoanapa (Guerrero, México: Madera y Bosques), 89–102. Available at: https://www.redalyc.org/articulo.oa?id=61780105 (Accessed August 28, 2024).
Trimble S. W. (1999). Decreased rates of alluvial sediment storage in the Coon Creek basin, Wisconsin 1975–93. Science 285, 1244–1246. doi: 10.1126/science.285.5431.1244
Turnbull W. J., Krinitsky M., and Weaver F. J. (1966). Bank erosion in soils of the Lower Mississippi Valley. J. Soil Moisture Div. Proc. Am. Soc Civ. Eng. 92, 121–136. doi: 10.1061/JSFEAQ.000082
van Oldenborgh G. J., Hendon H., Stockdale T., L´Heureux M., Coughlan de Perez E., Singh R., et al. (2021). Defining El Niño indices in a warming climate. Environ. Res. Lett. 16, 1–9. doi: 10.1088/1748-9326/abe9ed
Vargas Cuervo G. (2004). Geología y aspectos geográficos de la Isla de San Andrés, Colombia. Geología Colombiana. 29, 73–89. Available at: https://revistas.unal.edu.co/index.php/geocol/article/view/31915 (Accessed March 18, 2023).
Vásquez-Salazar R. D., Cardona-Mesa A. A., Valdés-Quintero J., Olmos-Severiche C., Gómez L., Travieso-González C. M., et al. (2024). Detection of coastal erosion and progradation in the Colombian ‘Atrato river’ Delta by using sentinel-1 synthetic aperture radar data. Remote Sens. 16, 1–21. doi: 10.3390/rs16030552
von Prahl H. (1989). Manglares de Colombia (Bogotá: Banco de Occidente). Available at: https://comunidadplanetaazul.com/ecolibros/manglares-de-Colombia/ (Accessed June 15, 2023).
Vos K., Harlley M. D., Turner I. L., and Splinter K. D. (2021). Pacific shoreline erosion and accretion patterns controlled by El Niño/Southern Oscillation. Nat. Geoscience. 16, 140–146. doi: 10.1038/s41561-022-01117-8
Vundavilli H., Mullarney J. C., MacDonald I. T., and Bryan. K. R. (2021). The interaction of buoyant coastal river plumes with mangrove vegetation and consequences for sediment deposition and erosion in a tidal environment. Continental Shelf Res. 222, (104417). doi: 10.1016/j.csr.2021.104417
Walters B. B., Rönnbäck P., Kovacs J. M., Crona B., Hussain S. A., Badola R., et al. (2008). Ethnobiology, socio-economics and management of mangrove forests: A review. Aquat. Bot. 89, 220–236. doi: 10.1016/j.aquabot.2008.02.009
Webb E. J. and Magi B. I. (2022). The ensemble oceanic niño index. Int. J. climatology 42 (10), 5321–5341. doi: 10.1002/joc.7535
Worthington T. A., Zu Ermgassen P. S., Friess D. A., Krauss K. W., Lovelock C. E., Thorley, et al. (2020). A global biophysical typology of mangroves and its relevance for ecosystem structure and deforestation. Sci. Rep. 10, 14652. doi: 10.1038/s41598-020-71194-5
Keywords: coastal erosion, neotropical mangroves, geomorphic setting, Pacific, Caribbean, Rhizophora mangle
Citation: Echeverry Hernandez JP, Mancera Pineda JE and Sánchez Núñez DA (2025) Migration of deltaic and open water mangrove forests in response to environmental factors. Front. Mar. Sci. 12:1569857. doi: 10.3389/fmars.2025.1569857
Received: 02 February 2025; Accepted: 22 April 2025;
Published: 20 May 2025.
Edited by:
Rina Kumari, Central University of Gujarat, IndiaReviewed by:
Luiz Drude Lacerda, Federal University of Ceara, BrazilRadhika Bhargava, National University of Singapore, Singapore
Copyright © 2025 Echeverry Hernandez, Mancera Pineda and Sánchez Núñez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Johanna Paola Echeverry Hernandez, anBlY2hldmVycnloQHVuYWwuZWR1LmNv; José Ernesto Mancera Pineda, amVtYW5jZXJhcEB1bmFsLmVkdS5jbw==