 Hammoud El Vadhel1,2*†
Hammoud El Vadhel1,2*† Lene Buhl-Mortensen3
Lene Buhl-Mortensen3 Mohamed El Khalil Med Mahmoud1
Mohamed El Khalil Med Mahmoud1 Yeslem El Vally1
Yeslem El Vally1 Beyah Meissa1
Beyah Meissa1 François Le Loc’h2
François Le Loc’h2 Lenaick Menot4
Lenaick Menot4- 1Institut Mauritanien de Recherches Océanographiques et des Pêches (IMROP), Nouadhibou, Mauritania
- 2Univ. Brest, Institut de Recherche pour le Développement (IRD), Centre National de la Recherche Scientifique (CNRS), Institut Français de Recherche pour l’Exploitation de la Mer (Ifremer), Laboratoire des Sciences de l’Environnement Marin (LEMAR), Plouzané, France
- 3Institute of Marine Research (IMR), Bergen, Norway
- 4Univ Brest, Institut Français de Recherche pour l’Exploitation de la Mer, Biologie et Écologie des Écosystèmes Marins Profonds (BEEP), Plouzané, France
This study presents the megafauna related to coral mounds (coral remains) and cold-water coral (CWC) reefs off Mauritania. The mound-specific megafauna was documented by comparing 47 trawl catches from coral mounds with 47 catches from off-mound areas, sampled from 1982 to 2022. Coral reef megafauna were described from eleven ROV dives conducted during two surveys held in 2020 and 2021. The mound and reef environment were documented using data from 2159 CTD profiles collected between 1995 and 2022. A detailed annotation of the ROV recorded videos was undertaken, and megafauna habitat relations were identified using multivariate statistics. The coral ecosystems that supported a rich megafauna were situated in the oxygen minimum zone at depths of 400 to 650 meters, with an average temperature of 10.55°C. The coral mound hosted 31% more species than adjacent off-mound areas, and fish contributed 66% of the taxa of five groups, followed by crustaceans contributing with 20%. The five most common species were Helicolenus dactylopterus, Laemonema laureysi, Merluccius polli, Malacocephalus occidentalis and Hoplostethus cadenati. The CWC reefs hosted 120 taxa representing 11 different faunal groups, with fish as the most species rich group, contributing 39% of the recorded taxa, followed by cnidarians represented by several corals (18 taxa) and crustaceans (17 taxa). The most abundant taxa on the reefs, in addition to the reef-building coral Desmophyllum pertusum, were the anemone Synarachnactis cf. lloydii, the corals Acanthogorgia cf. hirsuta and Swiftia phaeton, the crustaceans Nematocarcinus africanus and Eumunida bella, the fish H. dactylopterus, the bivalve Acesta excavata and the sponge Cladorhiza corallophila. Live coral, sand and mud were environmental drivers of species distribution on the reefs, and separate communities were related to different reef habitats. This study provides the first comprehensive description of the megafauna related to the cold-water coral mounds and reefs off Mauritania. The rich megafauna associated with these ecosystems highlights the urgent need for conservation measures, particularly in light of increasing pressures from oil and gas activities, deep-sea fishing, and the impact of climate change.
1 Introduction
Cold-water corals (CWC) are widely distributed on continental margins and seamounts across most oceanic regions of the world, mainly at depths of 200–1500 m (Roberts et al., 2006, Roberts et al., 2009). In southwest Africa, they occur along the Angolan margin (Hebbeln et al., 2020; Le Guilloux et al., 2009), on the Angolan and Namibian margins (Hanz et al., 2019), on the Namibian shelf (Tamborrino et al., 2019), in the Gulf of Guinea (Buhl-Mortensen et al., 2017), along the Mauritania margin (Colman et al., 2005; Moctar et al., 2024; Ramos et al., 2017; Westphal et al., 2014; Wienberg et al., 2018), north of Senegal (Moctar et al., 2024), and south of Morocco (Buhl-Mortensen et al., 2024; Hebbeln et al., 2019). The recent discoveries of the different CWC reefs and mound provinces reveal that the West African margin is a coral hotspot area in the Atlantic Ocean (Wienberg et al., 2023). The habitat complexity and heterogeneity provided by CWC support a high diversity of benthic species and may also act as shelter and nurseries for many species, providing a wide range of habitats for invertebrates and fishes (Buhl-Mortensen et al., 2010; Capezzuto et al., 2018; Costello et al., 2005; D’Onghia et al., 2012; El Vadhel et al., 2024; Kutti et al., 2013; Mortensen & Fosså, 2006; Ross and Quattrini, 2007). Due to this important ecological role, CWC reef habitats are considered biodiversity hotspots in the deep sea (Henry and Roberts, 2017). Both live and dead coral structures can play an important role in increasing three-dimensionality and they provide a hard substratum in the deep sea that will add habitats for many species in an otherwise relatively homogeneous setting (Bongiorni et al., 2010; Buhl-Mortensen et al., 2010). Interestingly, the coral associated fauna is richer in the older and dead parts than in the live parts of actively growing reefs, and many reef organisms can be supported by the complex microhabitat provided by rubble, often with a clear role in ecosystem functioning (Mortensen et al., 1995; Wolfe et al., 2021).
Mauritanian waters (from 20°46’N to 16°04’N) are located in North Atlantic Africa at the meeting point of temperate and subtropical oceanic ecosystems and belong to the Canary Upwelling Current (CUC), making them one of the most productive areas worldwide (Camp et al., 1991; Carr and Kearns, 2003; Hagen, 2001). They support one of the oldest and most important West African fisheries (Brahim et al., 2021) that target pelagic and demersal resources, including deep fishing from 200 m to 1000 m depth. Marine biological studies in the Mauritanian waters have mainly focused on fish stocks of commercial interest on the continental shelf (< 200 m) (e.g., Braham et al., 2014; Braham, 2013; Meissa et al., 2013; Gascuel et al., 2007; Meissa and Gascuel, 2015). Since 2006, several scientific surveys have been conducted by German, Spanish, and Norwegian vessels in the deep waters (>200 m) with the aim of describing and characterizing the marine ecosystem with a focus on geology, fish communities, and more recently corals (Bridges et al., 2023; Eisele et al., 2011; Krastel et al., 2006; Ramos et al., 2017; Sanz et al., 2017; Westphal et al., 2014; Wienberg et al., 2018, Wienberg et al., 2023; Wienberg and Titschack, 2015; Buhl-Mortensen et al., 2024; Moctar et al., 2024). These scientific surveys and studies related to hydrocarbon exploration have greatly enhanced our understanding of the Mauritanian seabed and its diverse habitats. Among the most significant deep-sea ecosystems discovered in Mauritania waters are cold water corals (CWC) occurring as mounds (framework of coral remains) and active and live reefs. Coral mounds result from an interplay between coral growth, sediment input, and processes on the adjacent (off-mound) seafloor. The mounds are created by the framework-forming scleractinian Desmophyllum pertusum (formerly known as L. pertusa; see Addamo et al., 2016) and, to some degree, Madrepora oculata (Hebbeln et al., 2016; Wienberg and Titschack, 2015). Along the Mauritanian coast, coral mounds form an extensive ridge (~400 km long) at 400–550 m composed of coral remains with occasional live colonies (Wienberg et al., 2018, Wienberg et al., 2023). Large and actively growing CWC reefs have been documented in the region at 430–650 m as recently as in 2020 and 2021 (Buhl-Mortensen et al., 2024; Moctar et al., 2024). Both coral habitats occur within the prevailing oxygen minimum zone off Mauritania (Buhl-Mortensen et al., 2024; Moctar et al., 2024; Ramos et al., 2017; Westphal et al., 2014; Wienberg et al., 2018, Wienberg et al., 2023). Despite the progress made in recent years to study the deep-sea habitats off Mauritania, the available information on the fauna associated with CWC ecosystems is limited. Colman et al. (2005) and Westphal et al. (2014) published the first visual data on the coral habitat, using a Remotely Operated Vehicle (ROV). They reported the presence of some colonies of live CWC, mainly Desmophyllum pertusum and Madrepora oculata, associated with some benthic organisms (e.g., sponges, gorgonians, crustaceans, sea pens, fishes, molluscs etc.). Other studies have described new species associated with CWC habitats off Mauritania (e.g., Göcke et al., 2016; Hoffman et al., 2019; Sampaio et al., 2022). The relationship between Helicolenus dactylopterus and cold-water corals in Northwest Africa was described in detail (El Vadhel et al., 2024).
Despite these studies, there is still no comprehensive investigation of the megafauna associated with CWC reefs and mounds in Mauritanian waters and their contribution to ecosystem functioning and diversity. Our main objective is to describe the associated megafauna of two different CWC ecosystems occurring off Mauritania: the chain of coral mounds and the live coral reefs. The megafauna inhabiting coral mounds is characterized through a comparative analysis of trawl catches from coral mounds and those from adjacent off-mound areas conducted from 1982 to 2022 by four research vessels. The megafauna associated with CWC reef habitats is described through the analysis of videos from ROV dives undertaken as part of two seafloor habitat mapping surveys conducted by the R/V Dr. Fridtjof Nansen off Mauritania in 2020 and 2021. The results provide insight into the importance of these CWC habitats to the Mauritanian deep-sea ecosystem, information that is essential for the development of a national management plan for the use of marine resources and their ecosystems.
2 Materials and methods
2.1 Environmental setting
2.1.1 Oceanography
Situated within the Canary Current Large Marine Ecosystem (CCLME), the currents in Mauritanian waters represent one of the most complex transition systems in the world. This region serves as a convergence zone for subtropical water masses from the north and tropical water masses from the south (Peña-Izquierdo et al., 2015). The main water masses identified within the Mauritanian Exclusive Economic Zone (EEZ) include (i) surface waters within the mixed layer at depths ≤ 50 m, (ii) the North Atlantic and South Atlantic Central Waters (NACW, SACW), which occupy a permanent layer down to 600 m (Cianca et al., 2009), (iii) Antarctic Intermediate Waters (AAIW) found between 600 and 1000 m (Emery, 2001), and (iv) the North Atlantic Deep Waters (NADW), located between 1000 and 1600 m in depth (Tomczak and Godfrey, 1994).
The primary surface currents in this region are the Canary Current (CC) and the Mauritania Current (MC) (Zenk et al., 1991). The dominant current between 100 m and 300 m within the water column is the Poleward Undercurrent (PUC), responsible for the northward transport of the South Atlantic Central Waters (SACW) (Pelegrí et al., 2017). These relatively cold and less saline SACW are separated from the North Atlantic Central Waters (NACW) by the Cape Verde Frontal Zone (CVFZ), which shifts between 21°N and 16°N latitude (Peña-Izquierdo et al., 2015).
Mauritanian waters are influenced by the presence of oxygen minimum zones (OMZs), characterized by extremely low dissolved oxygen concentrations. Two well-known OMZs can be observed in the region. The Upper OMZ (~100 m deep) is primarily caused by intense biological productivity, which is driven by nutrient upwelling. The decomposition of sinking organic matter further depletes oxygen levels in the water. The Lower OMZ (~400 m deep) associated with advection of the anoxic South Atlantic Central Waters (SACW) by the Polar Undercurrent (PUC), which exacerbates the anoxic conditions.
2.2 CWC study sites
The study sites encompass mounds and reef areas at depths ranging from 400 to 650 m (Figure 1). The coral mounds extend from Cap Timiris to the border with Senegal, while the coral reefs are located north and south of the mounds. The southern reef site is composed of eight reefs, and the northern reef site comprises three reefs (Figure 1).
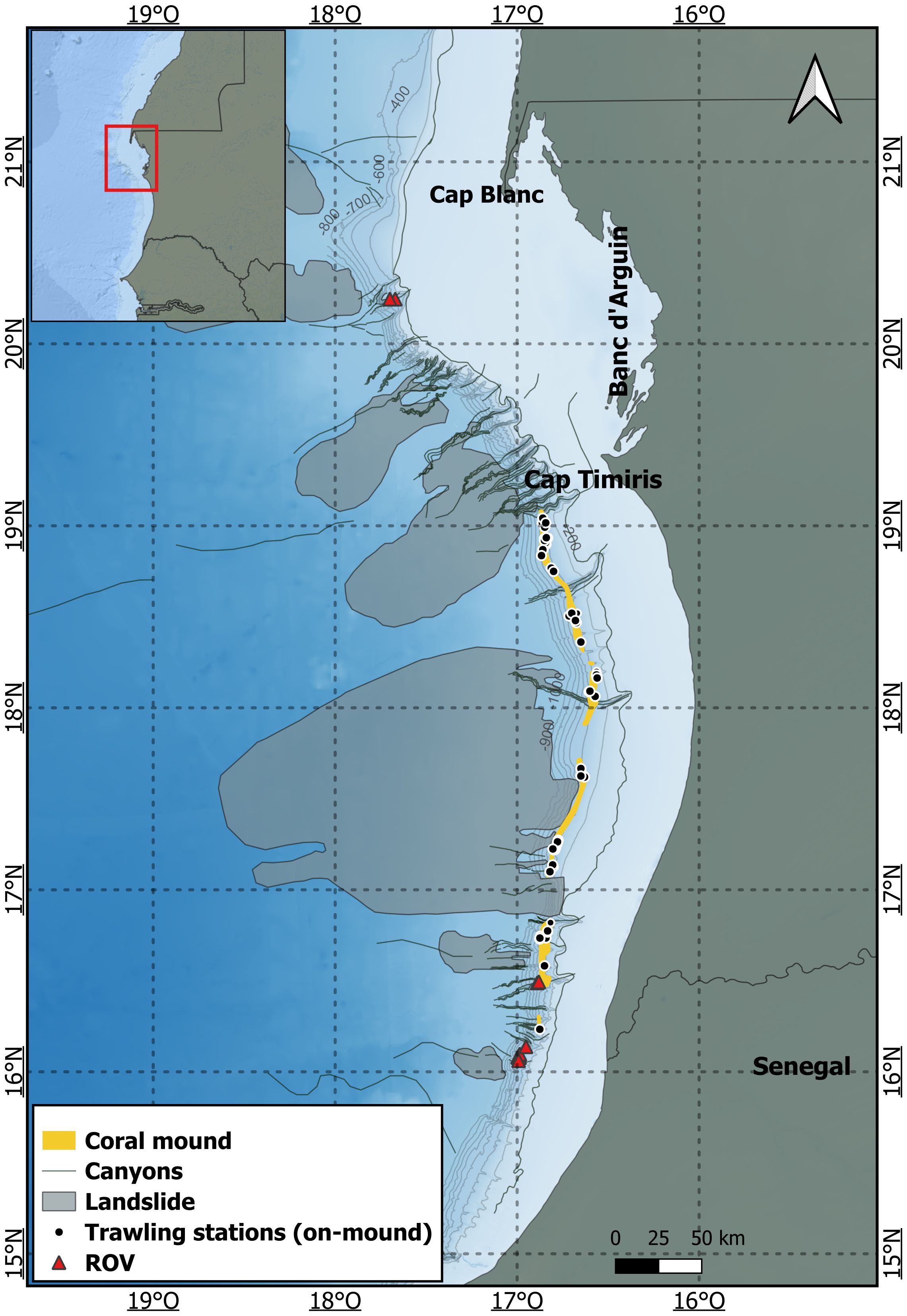
Figure 1. Map of the study area illustrating the distribution of coral mounds, canyons, and landslides along the Mauritian margins after (based on data from Colman et al., 2005; Ramos et al., 2017; Sanz et al., 2017; Westphal et al., 2014). The two CWC reef areas that were mapped and surveyed during 11 ROV dives are marked with red triangles. The 47 trawl surveys conducted in the coral mound zone are designated by black dots.
Here we discriminate between the coral mounds that are the remnants of former live coral reefs that in the region form a long barrier-like ridge. This old and basically dead structure formed by corals can host some live colonies. In contrast, a coral reef represents a single, well-defined structure built by the corals that have recently been actively growing, and reefs normally have actively growing parts.
2.2.1 The coral mounds
Colman et al. (2005) discovered a 190 km long succession of 100 m high coral mounds at 450–550 m depth, which later triggered several scientific surveys (Bridges et al., 2023; Eisele et al., 2011; Krastel et al., 2006; Ramos et al., 2017; Sanz et al., 2017; Westphal et al., 2014; Wienberg et al., 2018, Wienberg et al., 2023; Wienberg and Titschack, 2015). These coral mounds developed during the last 120,000 years, have an overall extension of ~400 km, and range between Cap Timiris (19°N) and the Senegalese border (16°N) (Ramos et al., 2017; Wienberg et al., 2018) (Figure 2). The mounds are cut by numerous canyons and submarine landslides. In some places, the mounds run in two parallel chains (Krastel et al., 2006; Sanz et al., 2017). The mounds are in a dormant state, with only scarce living coral colonies (Wienberg et al., 2018).
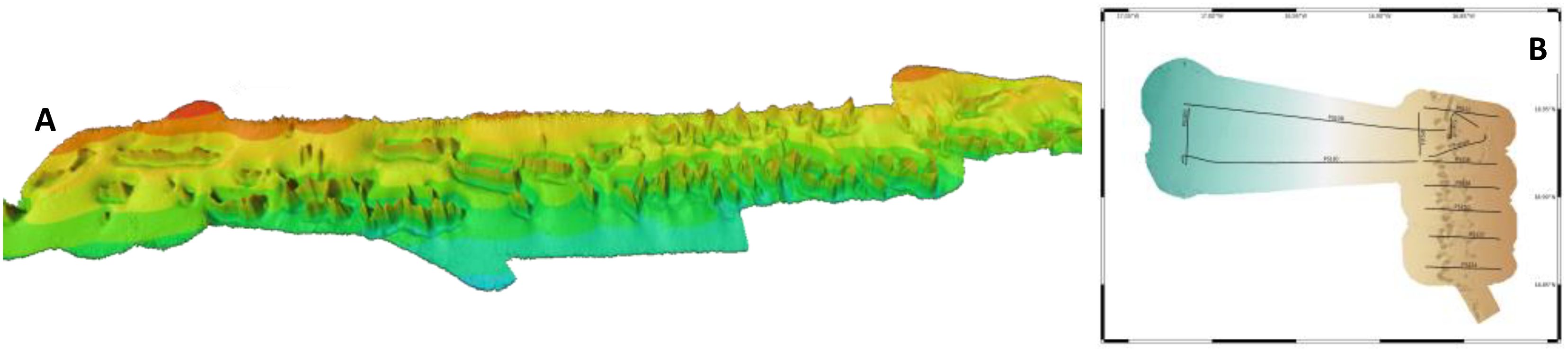
Figure 2. Bathymetric map showing the Mauritanian cold-water corals, (A) oblique view from W to E on a 30-nm-long strip off Cap Timiris coral mounds according to Freiwald et al. (2014), (B) Parasound profile of the Timiris mound complex (source: Bickert, T. and Shipboard Scientific Party of MSM127 (2024), unpublished data). Note that the two mound chains were developed.
2.2.2 The coral reefs
Previous studies on CWC reported mainly fossil corals based on sediment cores (Colman et al., 2005; Wienberg et al., 2018) and dredge samplings (Ramos et al., 2017), whereas ROV surveys reported only a few live colonies (Colman et al., 2005; Westphal et al., 2014).
However, recent investigations have recorded several healthy D. pertusum reefs in both the northern (Buhl-Mortensen et al., 2024) and southern regions of Mauritania (Moctar et al., 2024). Additionally, 13 (CWC) reefs were discovered, in the study area at the border between Mauritania and Senegal; six of these were healthy, with areas having 15%–50% cover of live colonies, and some reefs, such as the “Khairidine reef”, was approximately 70 m tall and 600 m long (Moctar et al., 2024).
2.3 Data sets
2.3.1 Oceanographic data
To describe the oceanographic setting of the CWC ecosystems, data from 2159 CTD profiles conducted within the Mauritanian EEZ were used. The profiles were conducted by 7 research vessels between 1995 and 2022 (Table 1) and were taken at depths between 0 and 1000 m, and 262 were from 400 to 650 m depth and positioned over the two CWC ecosystems. Data quality control and standardization were carried out using a Python script, which generated a standardized database of NetCDF files: (https://github.com/cmunozmas/nansen_pbgc_processing)
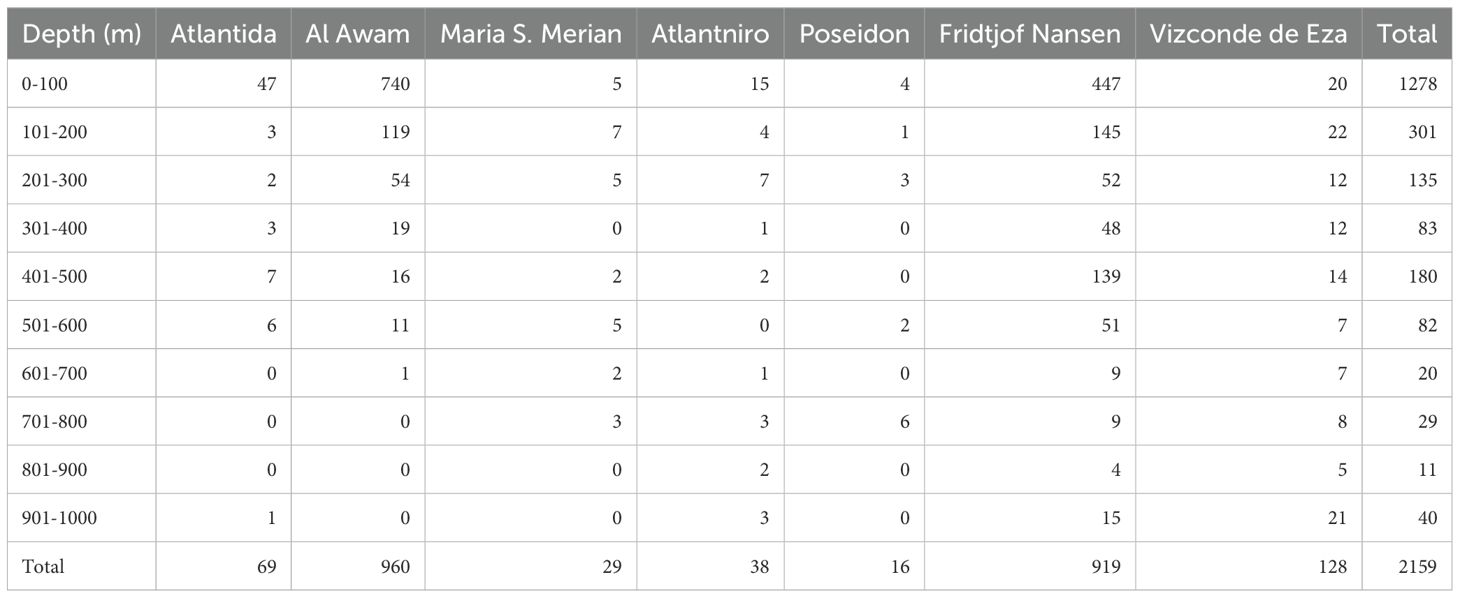
Table 1. Number of CTDs by depth and research vessel from 1995 to 2022.
2.3.2 Data from the coral mound
Data from deep trawling surveys were used to describe the megafauna inhabiting the coral mounds. The surveys were conducted by four research vessels (RV. N’Diago, RV. Al-Awam, R.V Vizconde de Eza and R.V Fridtjof Nansen) between 1982 and 2022 along the Mauritanian continental slope (>200 m). For selection of relevant trawl catches, trawling stations were plotted using QGIS software (version 3.26). Trawl stations starting on the mound but ending off the mound were excluded from the analysis to avoid mixing habitats. In total, 47 trawl stations were conducted at 400–600 m depth on the coral mounds and could be included in the analysis. For megafauna comparison, 47 off-mound trawl catches were selected adjacent to the mound barrier with a distance of at least 1.6 nautical miles from the mound to ensure habitat separation (Figure 3).
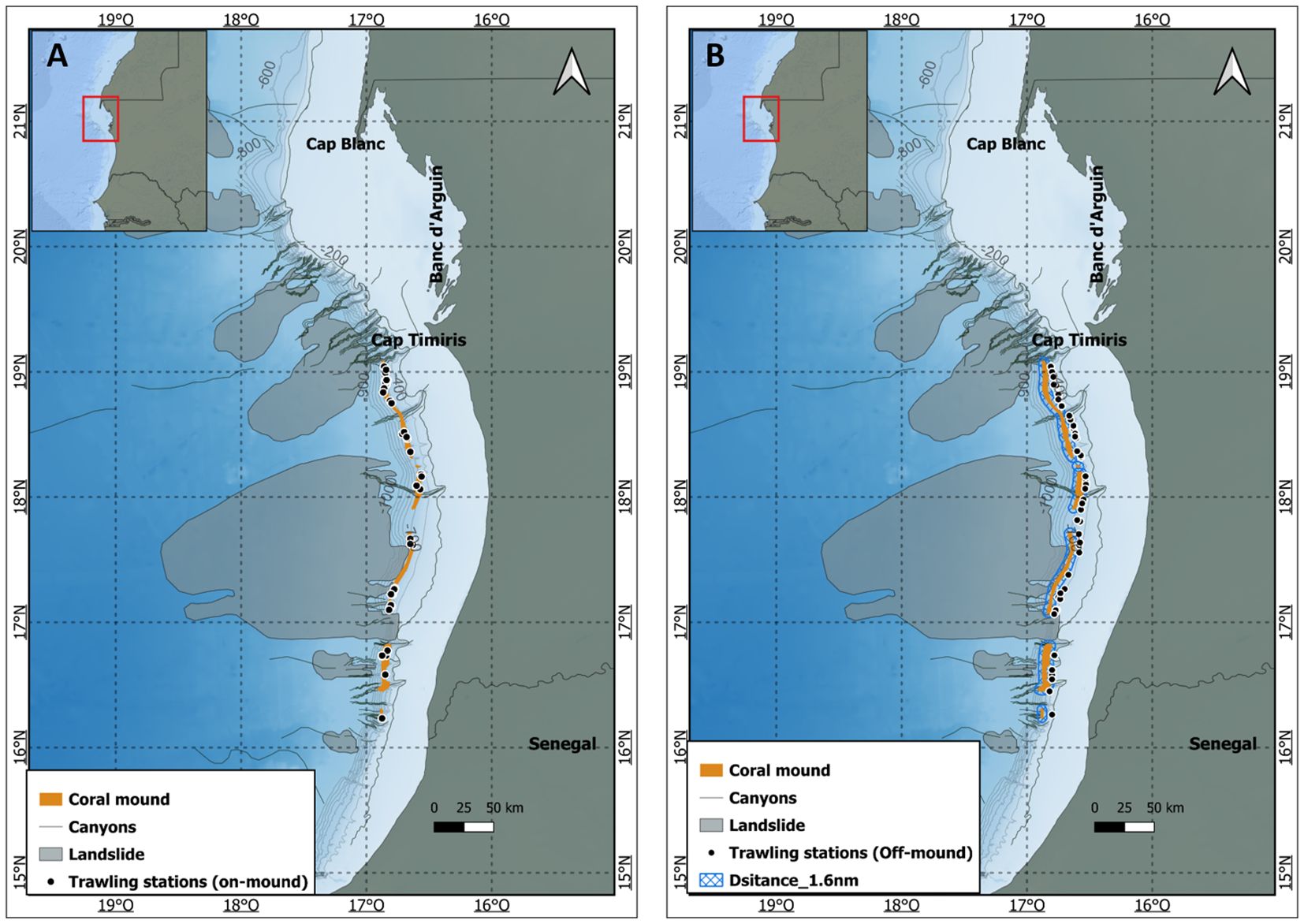
Figure 3. Map (A) shows the trawling stations conducted on-mound, and map (B) shows the trawling stations carried out in the off-mound area. The buffer zone of 1.6 nautical mile is indicated in blue.
The two Mauritanian research vessels, R/V Al Awam and R/V N’Diago, were using an “Irish-type” trawl with an average horizontal opening of 17 m and a vertical opening ranging from 2.8 to 3.5 m. The mesh size was 60 mm in the front part and 45 mm in the cod-end (pocket). The Spanish vessel R/V Vizconde de Eza used “Lofoten bottom trawl” during scientific surveys with a horizontal opening of 17.70 m and a vertical opening of 5.5 m, and a 35 mm mesh size at the codend (Ramos et al., 2017). The R.V Fridtjof Nansen used a “Gisund super demersal” for trawling, with a horizontal opening of about 18 m and a vertical opening of around 5.5 m, and a 20 mm mesh size in the codend and a 10 mm mesh size in the inner net.
Fauna data from the trawl catches included information on taxon name, latitude and longitude, depth, collection date, and number and weight of each specimen. Trawling times were 30 min for the three vessels (RV. N’Diago, RV. Al-Awam, R.V Fridtjof Nansen) and 60 min for the R.V Vizconde de Eza. However, due to the uneven and complex nature of the seabed, especially in the coral mound area, some trawling stations could not reach the standard trawling duration used by vessels (60 min for Vizconde de Eza and 30 min for the other three vessels). This was primarily due to obstacles such as coral structures and steep topography, which limited trawling efficiency. To ensure comparability, all trawling data were standardized by time. The number of trawling stations per vessel and zone (on-mound and off-mound) is the same (47 trawling stations) (Table 2).
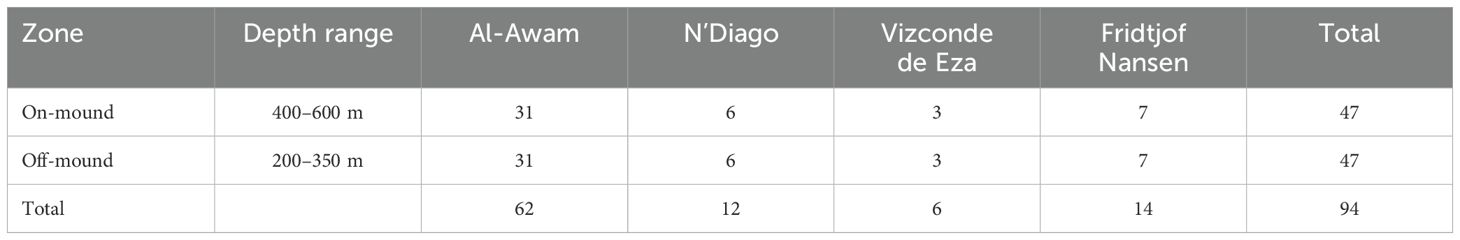
Table 2. Number of trawling stations by zone (on-mound and off-mound) and vessels.
2.3.3 Data from coral reefs
Video data from ROV surveys were collected as part of two habitat mapping surveys conducted by the R/V Dr. Fridtjof Nansen in the framework of the EAF-Nansen Programme. The first survey took place in 2020, in the northern part of Mauritania, while the second survey was carried out in 2021 in the southern part, at the border between Mauritanian and Senegal. The main objectives of both surveys were to document vulnerable seafloor habitats and their fauna, including CWC reefs, in this region (Table 3).
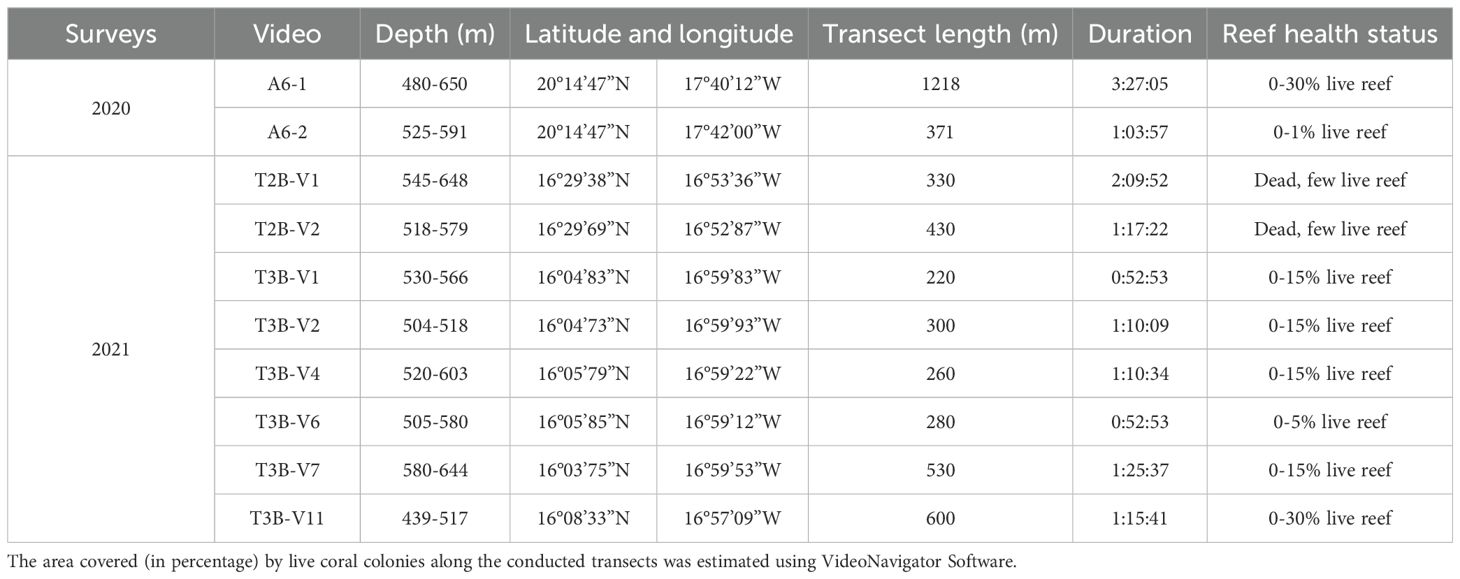
Table 3. Information on 11 ROV dives (A6–1 consists of 2 videos) including depth, position, transect length, duration, temperature, and observed % cover of live reef.
The Video Assisted Multi-Sampler (VAMS), developed by the Institute of Marine Research (IMR), was used to map the distribution of vulnerable ecosystems on the seafloor. It includes an ROV with a high-resolution video camera and two lasers mounted 5 cm apart, serving as a measuring scale, in addition it is equipped with five hydraulic operated grabs with a sampling area of 0.1 m2 (For description see Buhl-Mortensen et al., 2017). During transects, the VAMS was tethered by the vessel at a speed of approximately 0.3 knots with the ROV flying in front. A total of 11 dives were conducted during the two surveys, with 3 dives in the north and 8 in the south. The total area surveyed was approximately 6800 m2, covering a depth range of 440 m to 650 m.
2.4 Data analysis
2.4.1 Oceanography
A python script was used to process, analyse, and visualize oceanographic data to create profiles for the three environment parameters (temperature, salinity, and dissolved oxygen) in depth ranges between 0 to 1000 m. The analysis focused in particular on the depth zone of the two CWC ecosystems (400 to 650 m).
2.4.2 Fauna analysis
Data from trawls (94 stations) and ROV dives (11 videos) were organized separately in two different databases. We used the WoRMS Taxon Match Tool (available from https://www.marinespecies.org) to check the valid scientific name and complete the taxonomic classification for each taxon (e.g., phylum, class, order, family, etc.). For ROV data, species identification was based on videos and images. Taxonomists were consulted to confirm the identification of certain species (see Acknowledgments).
CWC Mounds: We analyzed the species diversity for on-mound and off-mound megafauna trawl catches. To describe and compare the diversity of both areas, species richness, Simpson index, Shannon-Weiner index, species accumulation curves, and Venn diagrams were calculated and drawn using the vegan package (Oksanen et al., 2022) in R software (version 4.2.1).
CWC Reefs: For the video analysis, we used the VideoNavigator software developed by the Institute of Marine Research (IMR) to identify, document and analyze the megafauna inhabiting the coral reef habitat. The software provides output files containing date, time, geographical position, depths, species names, abundances, substrate type, as well as observation of human impact (e.g., trawl marks, litter, etc.). In total, 7 substrates were listed (live coral: when live CWC represents more than 25%, coral rubble, rubble and live, coral block, sandy mud, sandy gravel, and bedrock).
For identification of community patterns and their relationship with environmental variables, the multivariate analysis Detrended Correspondence Analysis (DCA) and cluster analysis were performed using the PC-ORD software: Multivariate Analysis of Ecological Data version 5 (McCune and Mefford, 1999). This was conducted using a species composition matrix and an environmental variable matrix.
Before analysis, the 11 annotated video transects were divided into subsamples based on the dominant substrate (>50% cover). To maintain relatively equal sample sizes, subsamples were divided into equal lengths based on the length/record-time of the video sub-samples. Only taxa that occurred in three subsamples or more, and only subsamples that recorded at least two taxa, were retained in the input matrix for analysis. This resulted in a final dataset of 50 subsamples and 69 taxa and to reduce the influence of highly abundant species, data were square-root transformed before analysis.
After an initial run of DCA, only four categories of substratum were selected to ensure better representation of environmental relations: Live coral present (if live coral covers more than 5%), coral skeleton (coral rubble combined with blocks), sand & mud and gravel & bedrock if they cover > 50% of the substratum. Other environmental variables included were rugosity with three levels (low: flat seafloor, medium: thick rubble and some coral colonies, and high: high-relief areas with many coral blocks and live coral), depth, and site position (degrees north and south).
A correlation matrix in R was used to investigate the correlation between the main CWC reef building species (D. pertusum, M. oculata) and the other species in the reef community, and a heatmap was generated using the ggplot2 package in R (Wickham, 2016) to illustrate the relative abundance of species across different substrate types. For spatial distribution of D. pertusum, we used the QGIS software (version 3.26) to show the occurrence of this species in the study area. This was based on data from the two surveys conducted in 2020 and 2021, as well as literature available on the presence of live D. pertusum in Mauritanian waters (Westphal et al., 2014).
3 Results
3.1 Oceanographic environment
The analysis of 2159 CTD vertical profiles shows the properties of water masses, the Figure 4 presents three profiles of temperature (°C), salinity (PSU), and dissolved oxygen (ML/L) plotted as functions of depth from 0 to 1000 m in the Mauritanian Exclusive Economic Zone (EEZ). These profiles are based on all the historical CTD data collected from 1995 to 2022.
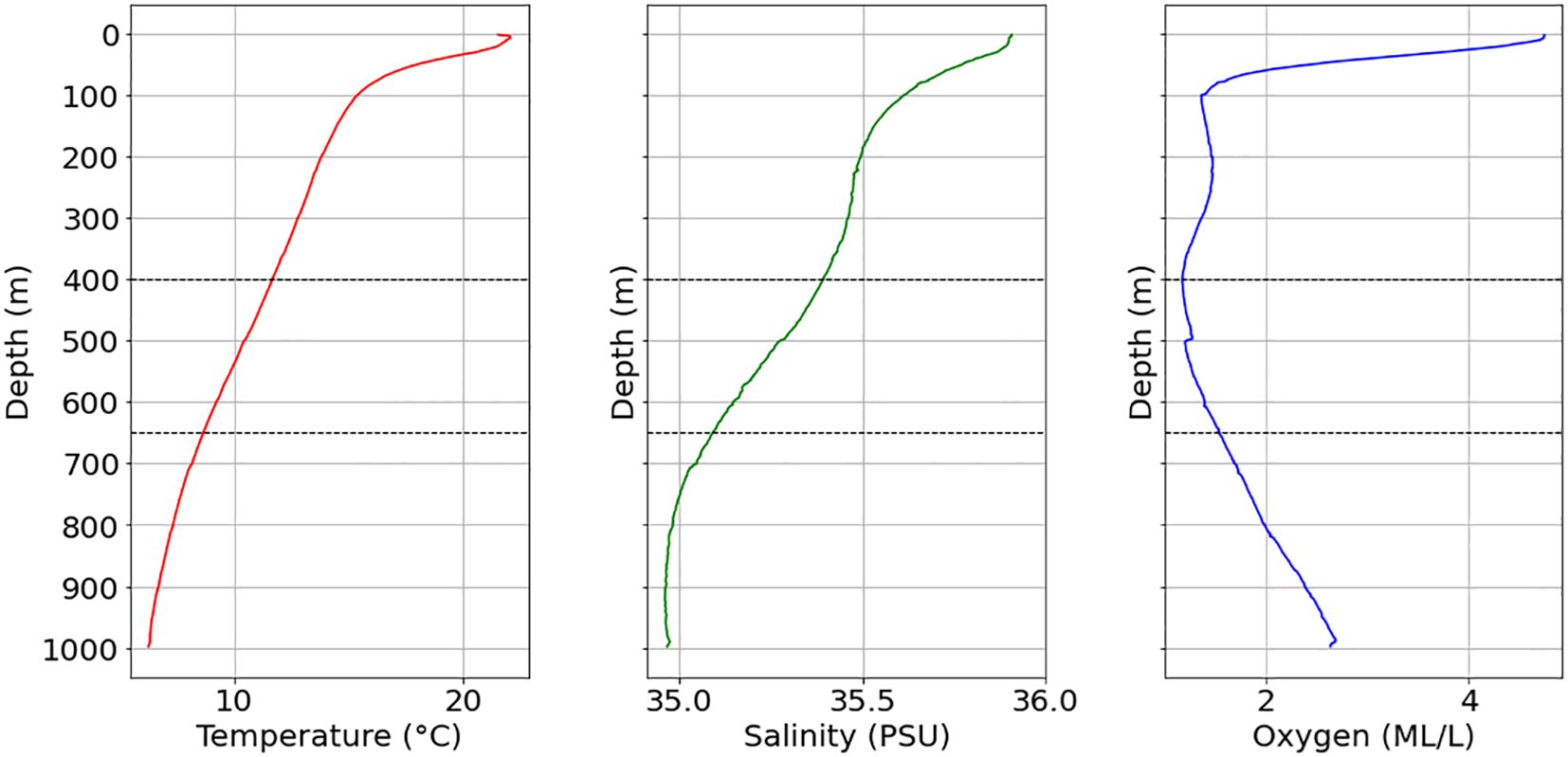
Figure 4. Temperature, salinity, and dissolved oxygen profiles of the CTD deployment in the Mauritanian EEZ between 1995 and 2022. The depth zone of mounds and reefs (400–650 m) is indicated by the dotted lines.
3.1.1 Temperature profile
The temperature profile demonstrates a typical thermocline structure, with warm surface waters reaching an average temperature of 22°C. Below this surface layer, temperatures decrease sharply through the thermocline, which spans from the surface to 100 m, with temperatures dropping to around 10°C at 500 m, beyond this depth, the temperature decline is more gradual, reaching around an average temperature of 5°C at 1000 m.
3.1.2 Salinity profile
Salinity remained relatively constant throughout the water column, ranging between 35 and 36 PSU, suggesting minimal variation and offering a consistent environment that can support marine life.
3.1.3 Dissolved oxygen profile
The dissolved oxygen profile reveals two prominent Oxygen Minimum Zones (OMZs): a shallow one at around 100 m and a deeper one near 400 m. The shallow OMZ exhibits a rapid drop in average oxygen concentrations from surface levels of approximately 4–5 ml L−1 to below 1 ml L−1at 100 m. After that, the oxygen content levels out, averaging between 1 and 1.5 ml L−1down to around 300 m, followed by a second, more pronounced decline at 400 m, where oxygen reaches its lowest average concentration. This depth corresponds to the onset of two CWC ecosystems. Below 400 m, average oxygen levels start to increase in the intermediate layer, reaching over 2.5 ml L−1 at 1000 m.
A focus on the area of occurrence of coral mounds and CWC reefs (400–650 m) through the analysis of more than 260 CTDs carried out in this depth range shows that the CWC ecosystems occur in waters characterized by an average temperature of 10.55°C, an average salinity of 35.28 PSU and an average dissolved oxygen of 1.24 ml L−1. At this depth, dissolved oxygen concentrations could reach a minimum of 0.77 ml L−1 and the temperature could be as low as 7.58°C. (Table 4).
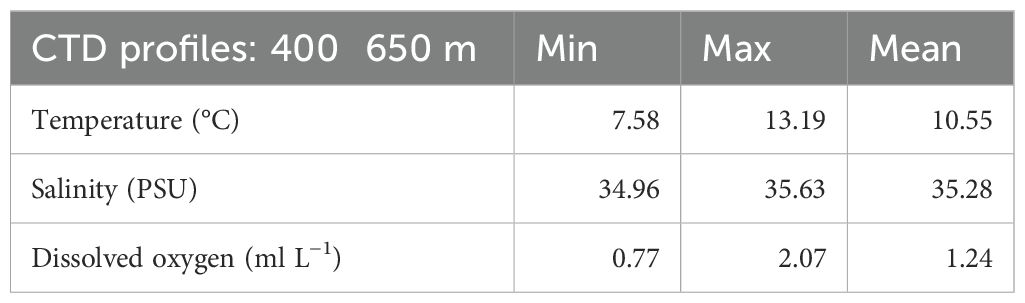
Table 4. Temperature, salinity and dissolved oxygen CTD profiles (400 to 650 m).
3.2 Trawling data analysis
The analysis of 47 trawl catches from coral mounds revealed a rich megafauna, including 282 recorded taxa belonging to the 5 taxonomic groups: Fish (187), Crustacea (56), Mollusca (31), Cnidaria (5) and Echinodermata (3).
The most common fish species was the blackbelly rosefish Helicolenus dactylopterus recorded in 41 out of 47 trawl catches. Other frequently reported species (found in more than 26 stations) included the guinean codling (Laemonema laureysi), black hake (Merluccius polli), western softhead grenadier (Malacocephalus occidentalis), black slimehead (Hoplostethus cadenati), Darwin’s slimehead (Gephyroberyx darwini), Blackbellied angler (Lophius budegassa), Senegalese hake (Merluccius senegalensis), and pink frogmouth (Chaunax pictus). The most common of the crustaceans was the box crab, Paromola cuvieri occurring in 16 trawl catches, followed by the deep-water pink shrimp Parapenaeus longirostris and the African mud shrimp Solenocera africana, both encountered in more than 10 trawl catches.
The Mollusca group was dominated by the European flying squid Todarodes sagittatus, this species was reported in 21 trawl stations, followed by the shortfin squid Illex coindetii with 14 observations. The last two taxonomic groups (Cnidaria and Echinodermata) were poorly represented in data from demersal trawling surveys.
For the 30 most common species in both habitats, H. dactylopterus comes first in on-mound areas and second in off-mound adjacent areas. Other commercial species were also abundant in both areas (e.g., M. polli, Parapenaeus longirostris, M. senegalensis, I. coindetii and L. budegassa). (Table 5)
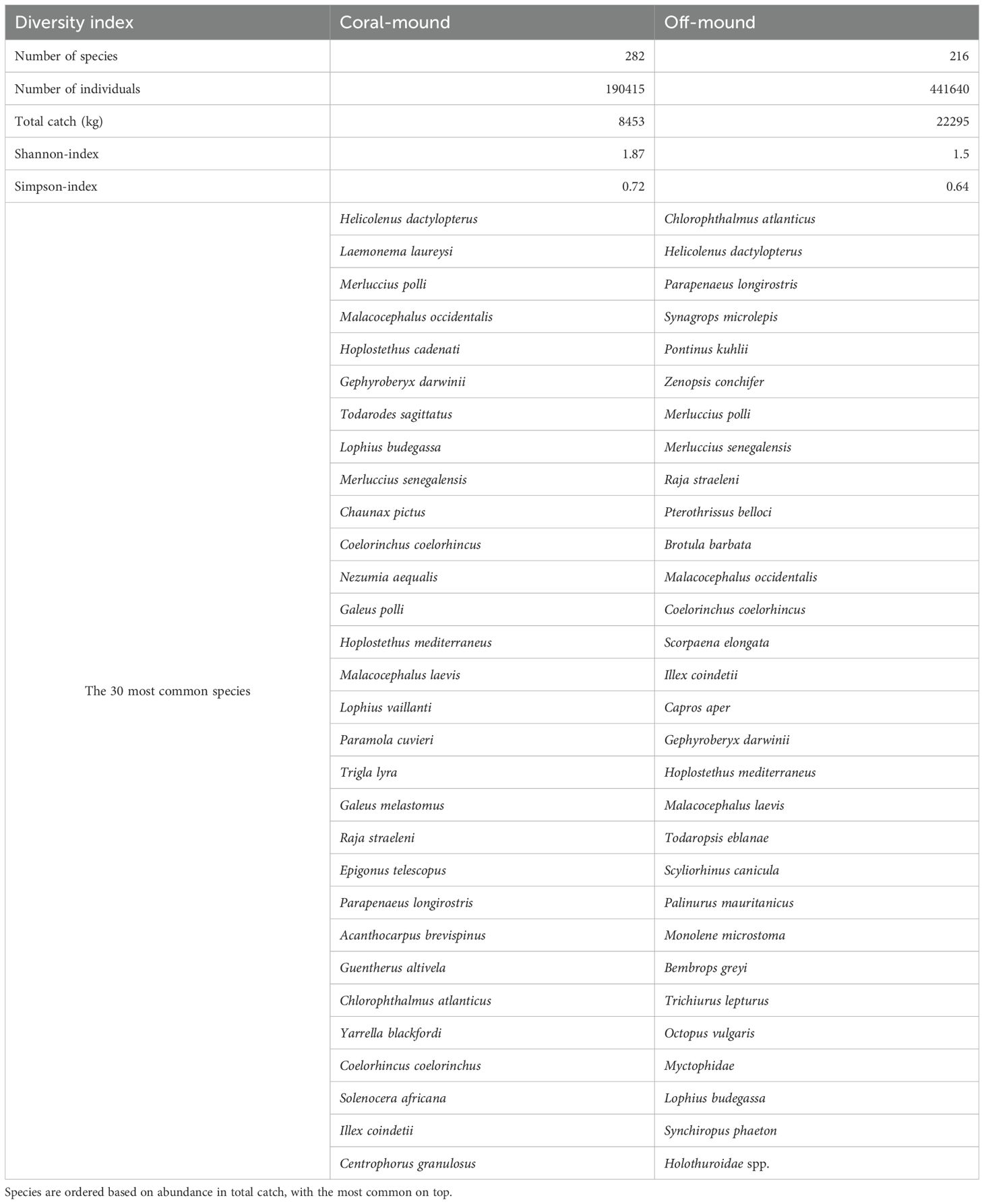
Table 5. Species diversity and most common species in trawl catches from coral mound and off-mound environment.
The diversity of species that inhabit the coral mound area (400 to 600 m) was compared to the adjacent off-mound habitat (200 to 350 m). Species richness was higher on-mound than off-mound (282 versus 216 taxa; Shannon index 1.87 versus 1.5; Simpson index 0.72 versus 0.64). Abundance (i.e., number of individuals, total catch) was more than two times higher off-mound than on-mound.
3.2.1 Diversity by family
The diversity of the megafauna on the mounds is high on family level, with 116 families representing 282 taxa found on coral mounds. The top 16 families are illustrated in Figure 5.
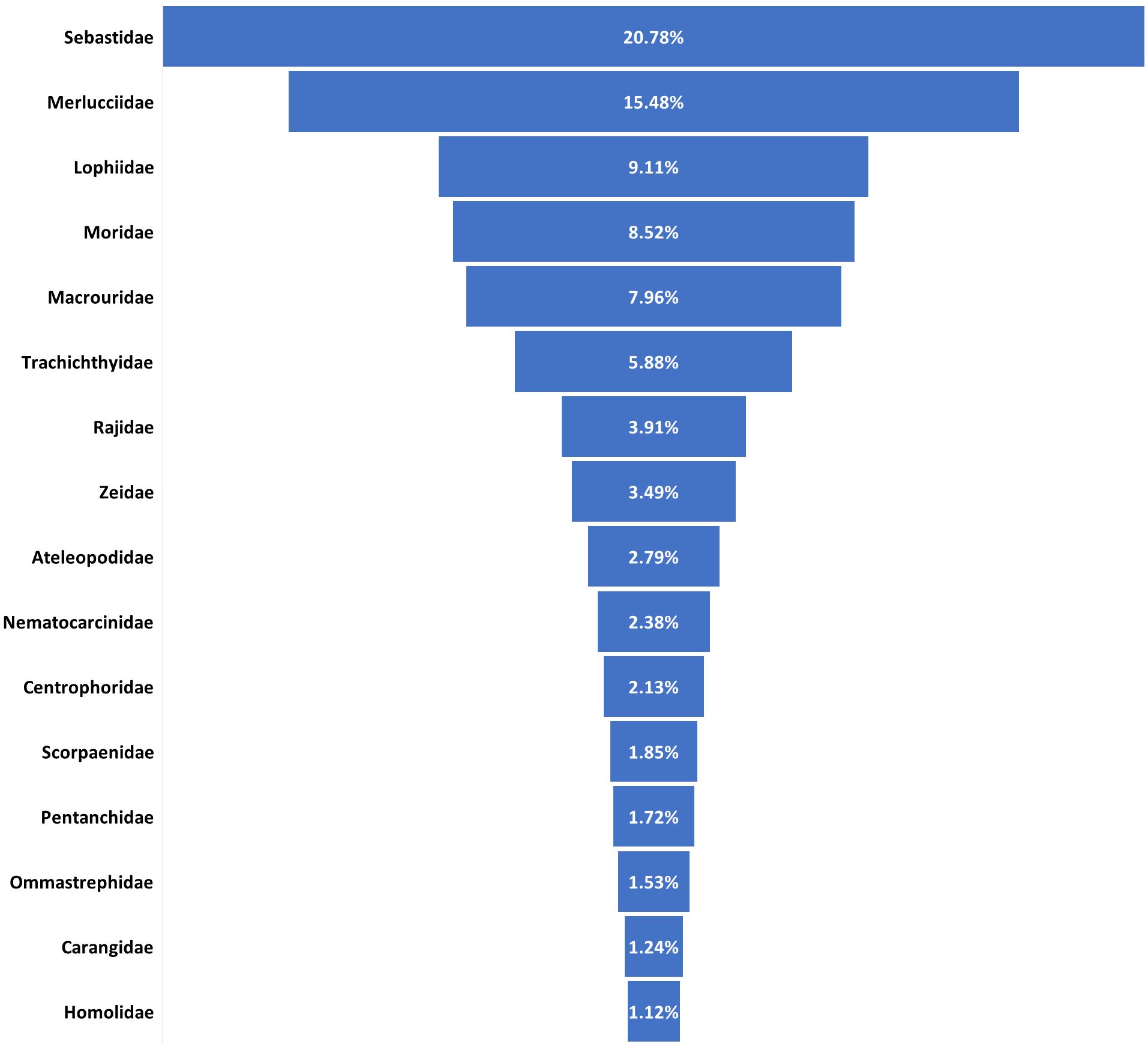
Figure 5. The sixty families of species that contribute significantly to the total catch on the mound.
In terms of species richness, the most represented family is the deep-sea fish family Macrouridae, with 13 species, followed by the Soleidae family of flatfishes and the Centrophoridae family of squaliform sharks, each with 9 species. The Congridae family, which includes conger and garden eels, and the Pandalidae family of pandalid shrimps both have 8 species.
When considering the total catch, the Sebastidae family, which is represented by two fish species (H. dactylopterus and Trachyscorpia cristulata) account for more than 20%. The Merlucciidae family, including both species of black hake (M. polli and M. senegalensis), is second with over 15%. The Lophiidae family, which contains L. budegassa, Lophius vaillanti and Lophius sp., contributes more than 9%. Moreover, the Moridae family, which includes Gadella imberbis, Gadella Maraldi, L. laureysi and Laemonema yarellii as well as the Macrouridae family, which includes many species (Bathygadus macrops, Coelorinchus braueri, Coelorinchus caelorhincus, Coryphaenoides sp., Hymenocephalus italicus, Malacocephalus laevis, Malacocephalus occidentalis, Malacocephalus sp.,Nezumia aequalis, Nezumia micronychodon, Nezumia sp., Trachyrincus scabrus, Trachyrincus sp.) also contribute notably to the total catch on mound, with 8% and 7%, respectively.
3.2.2 Diversity between the on-mound and off-mound
A total of 349 species were identified from 94 trawls carried out in on-mound and off- mound sites. Of these species, 42% occurred in both habitats (overlapping circle), and over one third (38%) are found only on-mound (Figure 6). These results confirm the notable specific richness observed in the on-mound habitat.
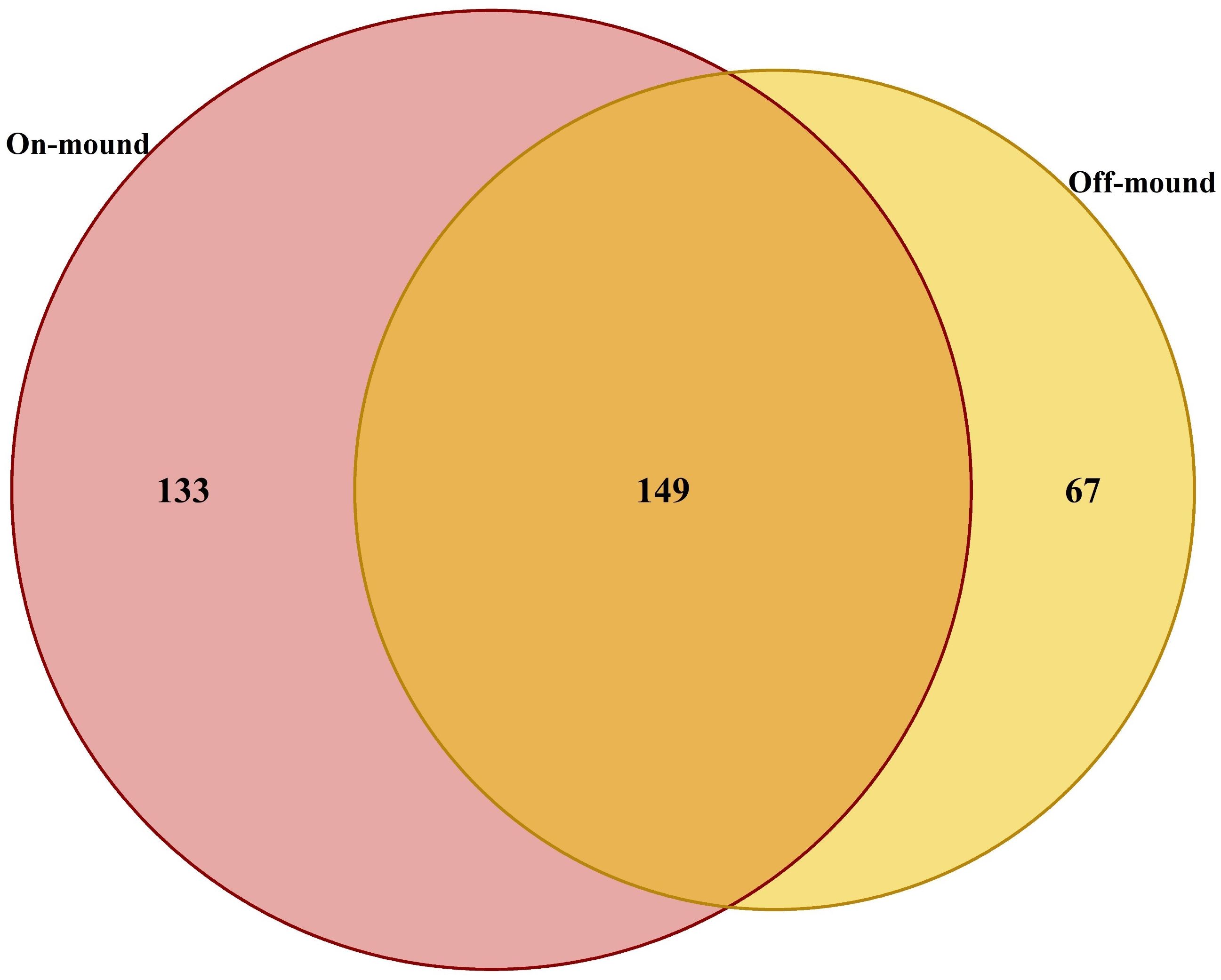
Figure 6. Venn diagram illustrates the number of shared and unique species between on- and off-mound areas.
3.2.3 Species accumulation curves
Species accumulation curves do not reach an asymptote, suggesting both sites are under sampled. However, the rate of species detection with increasing area sampled was faster on-mound than off-mound, thus highlighting that species richness is higher on-mound compared to off-mound habitat (Figure 7).
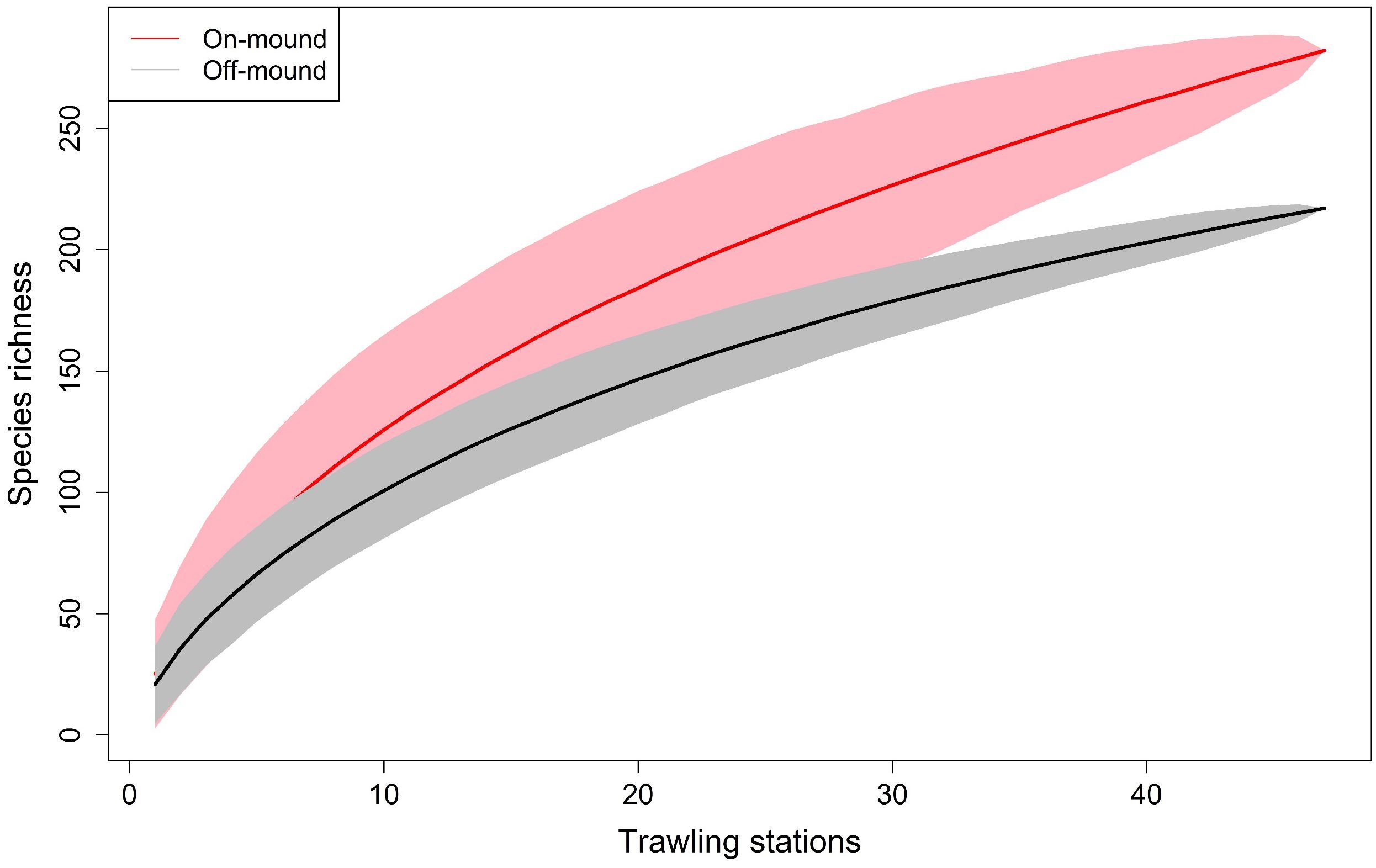
Figure 7. Species accumulation curves considering species richness per trawling stations: on-mound in pink and off-mound in gray.
3.3 ROV dives data analysis
3.3.1 Presence of live CWC
Eight species of CWC were identified. These included the numerically dominant Acanthogorgia cf. hirsuta followed by D. pertusum, Swiftia phaeton, Thesea talismani, M. oculata, Tanacetipathes cf. spinescens, Anthomastus cf. grandifloras and Clavularia borealis. The highest concentration of living CWC was found in the far north (20°N), where D. pertusum predominated, and in the south, at the boundary between Mauritania and Senegal, where D. pertusum and M. oculata together formed reef structures (Figure 8).

Figure 8. Map showing the presence of live CWC along the Mauritanian margin at the two CWC reef areas: (A) Northern reefs are constituted mainly by D. pertusum and, (B) southern reefs, which are constructed by D. pertusum and, to a lesser degree, by M. oculata, the bivalve Acesta excavata and the squat lobster Eumunida bella were abundant among the reef community, where they find food and shelter. Between the two reefs, few living corals are indicated (black dots) according to Westphal et al., 2014.
3.3.2 Megafauna associated with CWC
The reefs offer habitat for many species. A total of 120 taxa belonging to 68 families were identified and divided into 11 groups. Fishes were diverse with 47 taxa, followed by Cnidaria (18 taxa), Crustacea (17 taxa), Mollusca and Echinodermata with 11 taxa for each group, and Porifera with 7 taxa. H. dactylopterus was the most observed fish species, with over 371 individuals. Cnidarians were dominated by Synarachnactis cf. lloydii (> 13000 individuals). While Nematocarcinus africanus was the most common crustacean (>900 individuals). Among mollusks, the bivalve Gigantidas mauritanicus was most frequent, with more than 1800 individuals. And the sponge Cladorhiza corallophila was prevalent among porifera (Table 6).
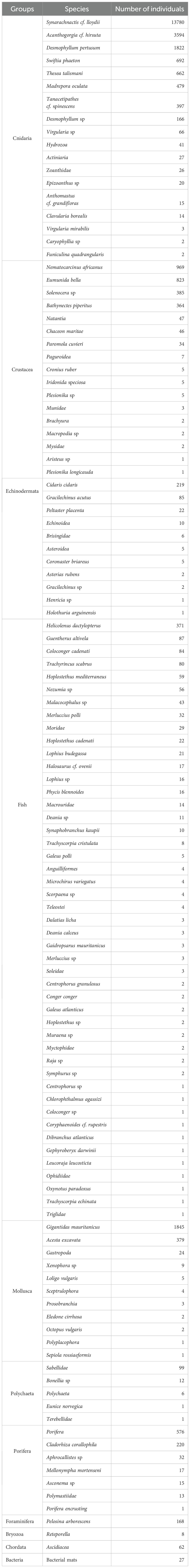
Table 6. A list of all taxa by group with the number of individuals annotated for the 11 ROV dives in the study area.
Overall, the reef areas supported a rich megafauna associated with the CWC coral reef. Figure 9 shows images of several common species found on the CWC reefs.

Figure 9. Representative ROV images of common species in the CWC reefs off Mauritania across the different taxonomic groups. (A) three specimens of H. dactylopterus with live D. pertusum in high densities of S. lloydii. (B) L. budegassa with live D. pertusum, T. spinescens, S. phaeton, A. hirsuta, C. corallophila and unidentified sponge. (C) M. polli and C. cidaris with mainly dead corals. (D) several individuals of the Mediterranean slimehead H. mediterraneus. (E) a school of jellynose fish (G. altivela) at the edge of the reef; this species is red-listed by the (IUCN). (F) a large West African geryon C. maritae resting in sandy mud sediment. (G) the large crab P. cuvieri. (H) tow specimens of the spring crab E. bella with B. piperitus. (I) the African spider shrimp N. africanus. (J) M. oculata. (K) a close-up of D. pertusum polyps. (L) intense presence of the bivalve G. mauritanicus, on top of bacteria mats with evidence of methane seep. (M) A. excavata attached to hard substrata with D. pertusum. (N) the sponge M. mortenseni. (O) the sponge Aphrocallistes sp.
3.3.3 Heatmap analysis of species abundance across substrate types
To make a heatmap that displays the relative abundance of the 25 most abundant species across different substrate types, five species were selected from each type of substrate (Figure 10).
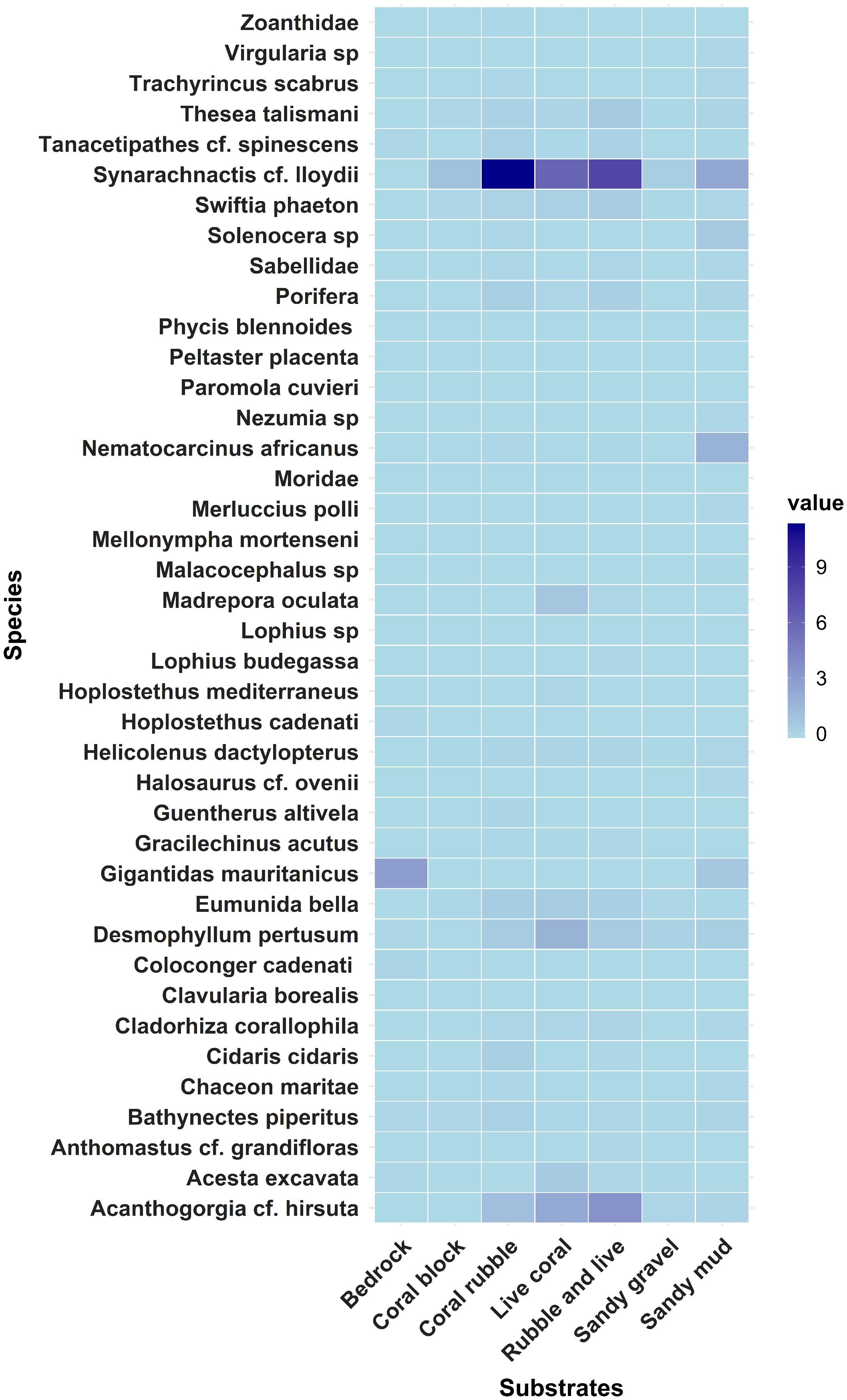
Figure 10. The heatmap shows species abundance across different substrates, the blue color gradients in this heatmap visually show the various levels of species abundance in relation to the seven substrates with dark blue indicating a high presence of the species on each substrate.
Species that demand more stable and substantial foundations, such as the bivalve G. mauritanicus and the deep-water-dwelling eel C. cadenati, are supported by bedrock substrates. C. cadenati uses rock crevices and bedrock formations as refuges. Notably, bacterial mats were frequently linked to the high abundance of G. mauritanicus. Species such as the fish H. dactylopterus, the crab E. bella, and the sea anemone S. cf. lloydii are common on coral blocks, indicating a predilection for intricate structures that provide food and refuge for these species. It is important to note that S. cf. lloydii was prevalent on all seven substrate types, demonstrating its broad range, habitat preference, and adaptability.
Coral rubble hosts a diverse range of species, underscoring the importance of this habitat type. The most abundant species included S. cf. lloydii, A. cf. hirsuta, and B. piperitus. Live coral is primarily inhabited by two CWC species, the gorgonian A. cf. hirsuta and the stony coral D. pertusum, along with S. cf. lloydii, which are the most dominant in this habitat. Similar species, including A. cf. hirsuta, S. cf. lloydii, and S. phaeton, also dominate in the combination of rubble and live coral, indicating that these species require hard substrate for survival.
The fish H. dactylopterus, the CWC species D. pertusum, and the S. phaeton have a slight presence in the sandy gravel substrate. Finally, sandy mud is dominated by two shrimp species, N. africanus and Solenocera sp., as well as S. cf. lloydii, indicating that softer, muddier substrates support a distinct species assemblage when compared to more solid or rocky settings.
3.3.4 Species-by-species correlation matrix
A correlation matrix between the 40 most frequent species is provided (Figure 11) to detect positive (in blue) and negative (in red) correlations between species in terms of occurrence (ranging from -1 to 1). A focus on the main reef building CWC D. pertusum and M. oculata shows that for D. pertusum, four species showed a strong positive correlation (0.9) with this species: E. bella, P. placenta, Lophius sp, and C. borealis. This was followed by a group of species with a correlation greater than 0.7: A. excavata, M. oculata, P. cuvieri, A. cf. hirsuta, S. phaeton, H. dactylopterus, H. mediterraneus and C. corallophila. For M. oculata, three species showed a strong positive correlation (greater than 0.9): Acesta excavata, Lophius sp., and H. mediterraneus, followed by D. pertusum (0.8), P. placenta, and E. bella (0.7).
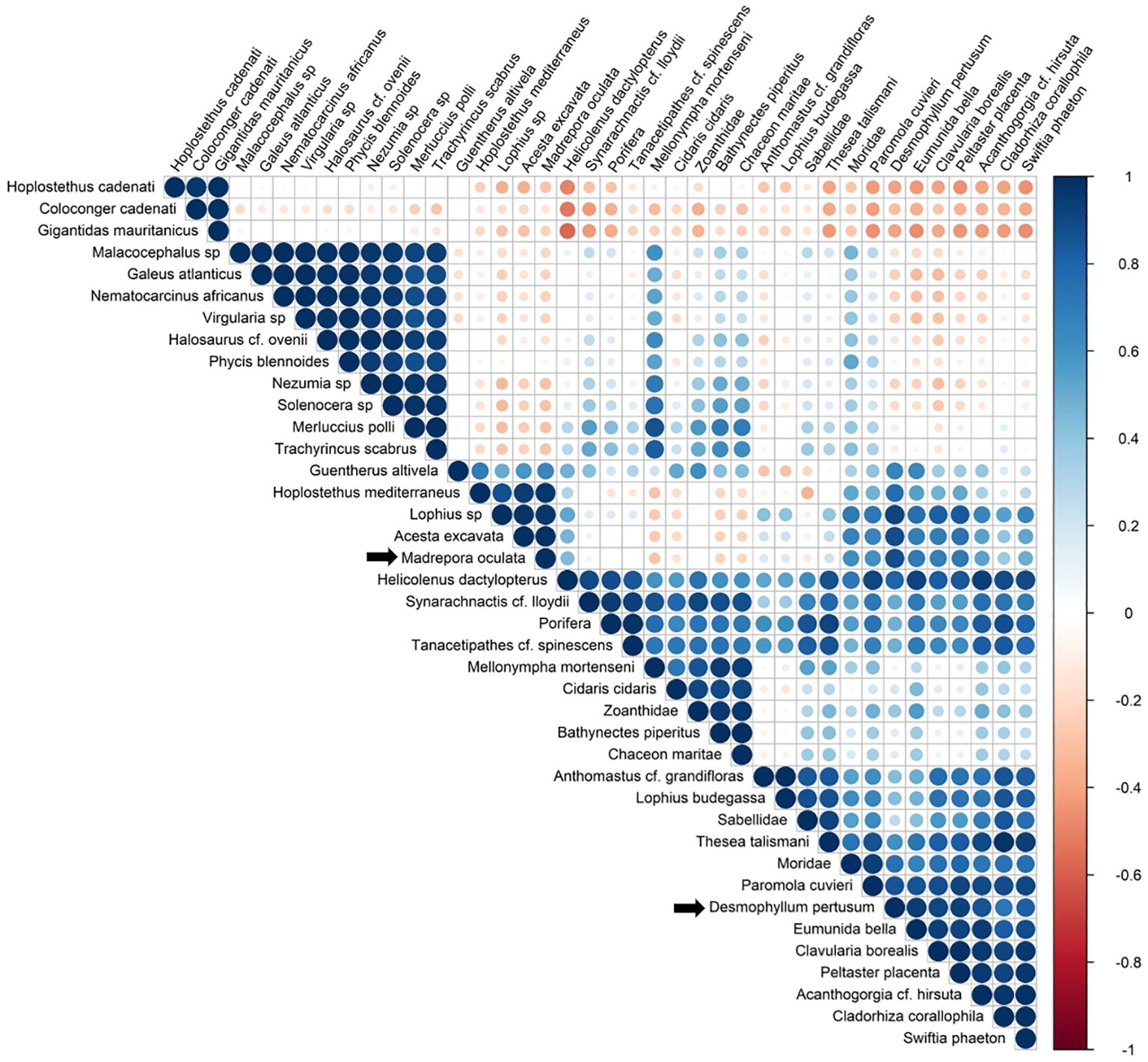
Figure 11. The correlation matrix of the different species associated with CWC. The legend color shows the correlation gradient from -1 (negative correlations) to 1 (positive correlations). The size of the circles and color intensity are proportional to the correlation coefficients. The main CWC species, D. pertusum and M. oculata are indicated by the black arrow.
3.3.5 Community patterns
The cluster analysis identified seven groups of organisms inhabiting CWC reefs, with one additional group (8) isolated from the arrangement (Figure 12).
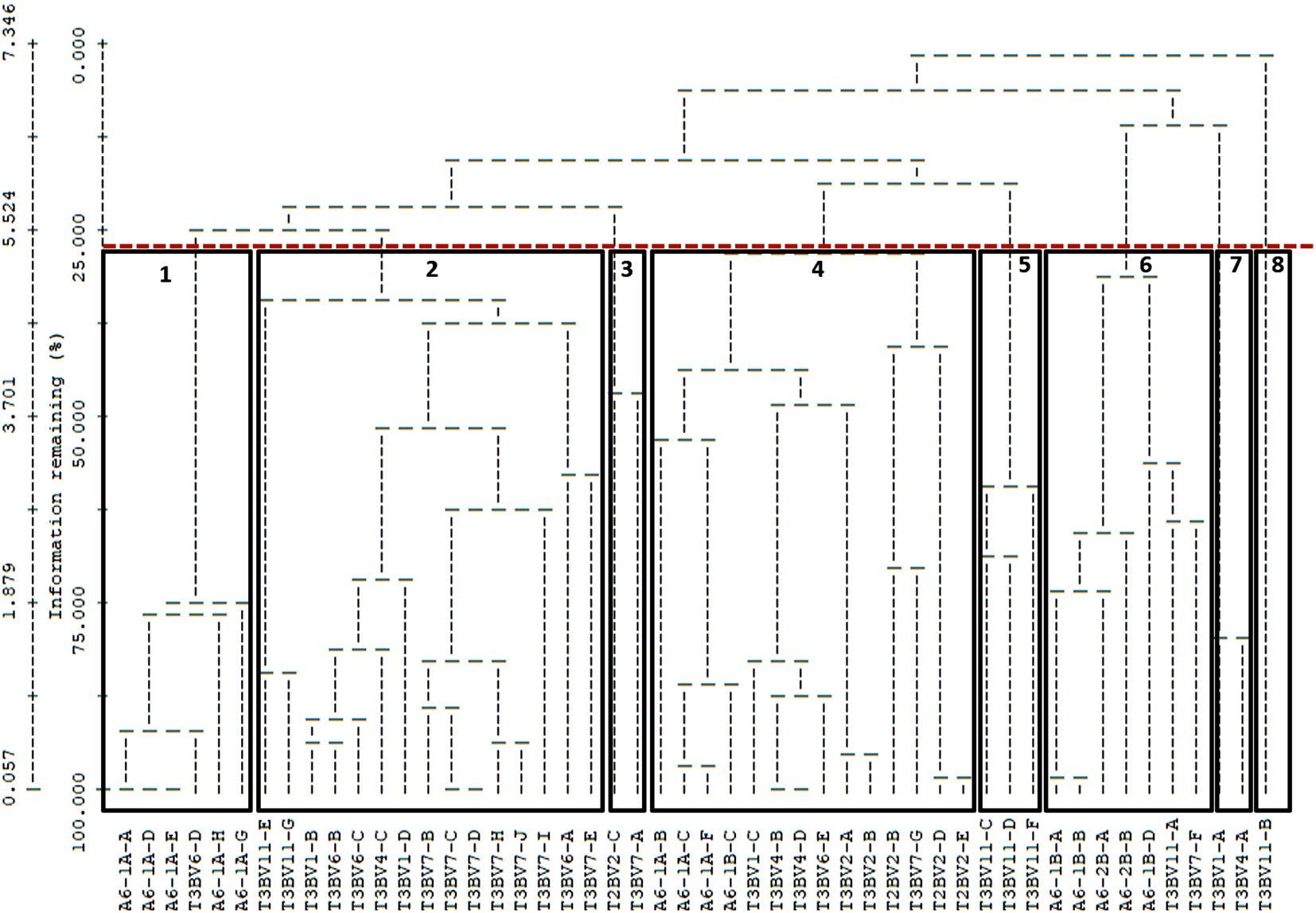
Figure 12. A dendrogram generated from hierarchical clustering based on species composition illustrates the similarities between samples, with eight primary community groups highlighted in different colors at a ~ 28% similarity levels.
Group 1 is characterized by the sea anemone S.cf. lloydii and two CWC species, D. pertusum and S. phaeton. The predominant substrates in this group are a mixture of different substrata (coral skeleton (37%), gravel & bedrock (28%), live coral present (19%), and Sand & mud (16%).
Group 2 shares similarities in terms of substratum with group 1 (49% of coral skeleton, 34% of sand and mud and 17% live coral present), it is also dominated by the sea anemone S.cf. lloydii, with the gorgonian A. cf. hirsuta and the sea fan T. talismani.Group 3 composed mainly of sand and mud (52%) with live coral present (48%), is dominated by the sea anemone S. cf. lloydii and crustaceans such as shrimps, N. africanus.
Group 4 has the highest diversity and abundance of species and features the sea anemone S. cf. lloydii and CWC species such as A. cf. hirsuta, D. pertusum, S. phaeton, T. talismani, and the crustacean E. bella along with the fish H. dactylopterus. This group is dominated by live coral substratum (60%) and coral skeleton (40%).
Group 5 is characterized by CWC species such as A. cf. hirsuta, D. pertusum, M. oculata, T. talismani, and the crustacean E. bella the bivalve A. excavata. The substrate is primarily live coral present (100%).
Group 6 is dominated by sand and mud (76%) with 24% of coral skeleton. This group is characterized by S. cf. lloydii, and crustaceans inhabiting soft substrates such as N. africanus, E. bella and B. piperitus, the fish H. dactylopterus and T. scabrus.
Group 7 is characterized by shrimp’s species such as N. africanus and Solenocera sp, sea anemone S. cf. lloydii and the sea pen Virgularia sp, which dominate this soft substrate community. This group is dominated by sand and mud (100%), (Table 7).
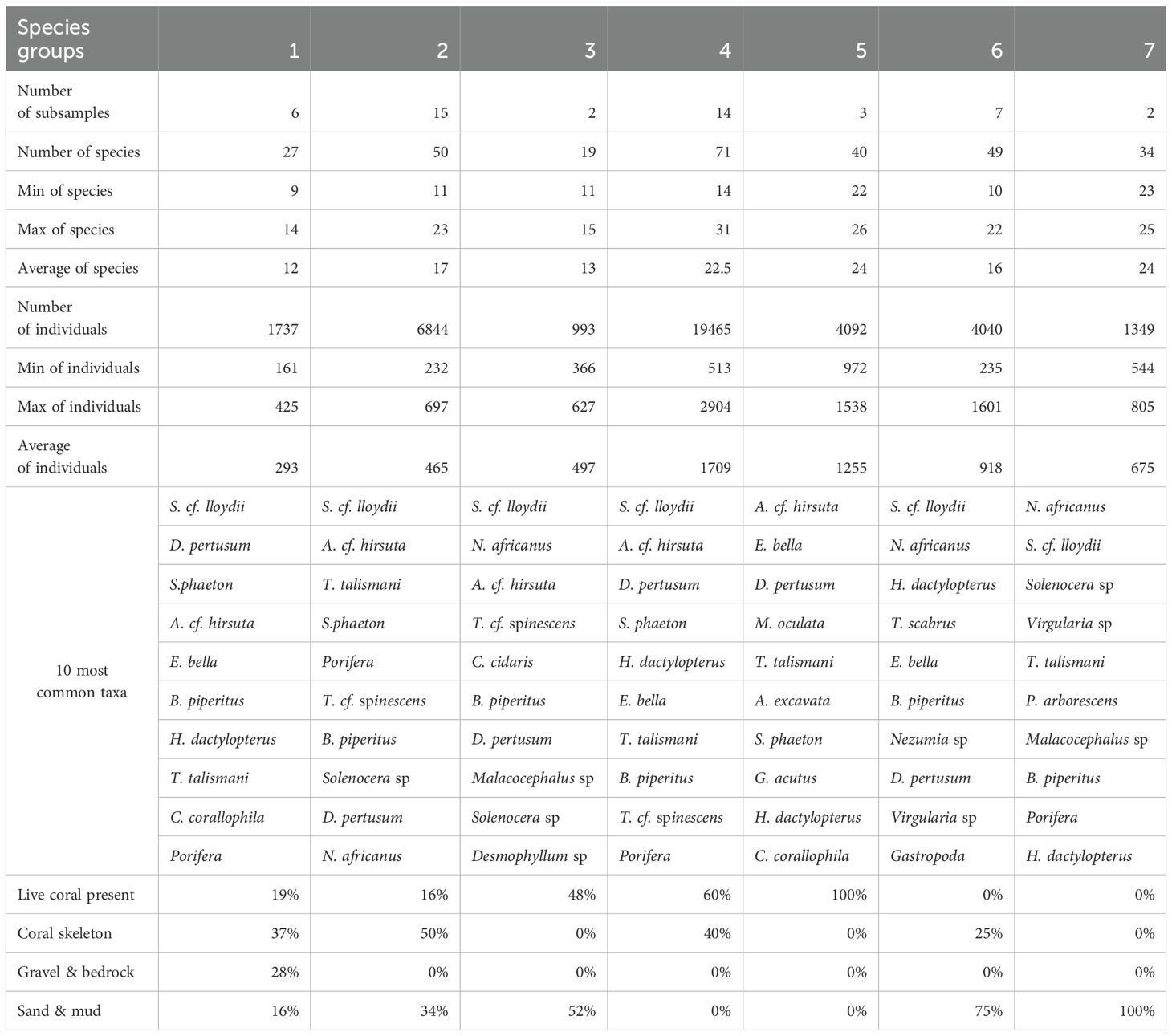
Table 7. The main features of the seven groups include 10 common species, the number of species, individuals, the average of species and individuals by subsamples, the number of subsamples, and substrate types (%).
The DCA analysis considered seven environmental variables, including substrate types, rugosity levels, depth, and site (north and south). The DCA graph illustrates species distribution patterns in relation to key environmental gradients (Figure 13).
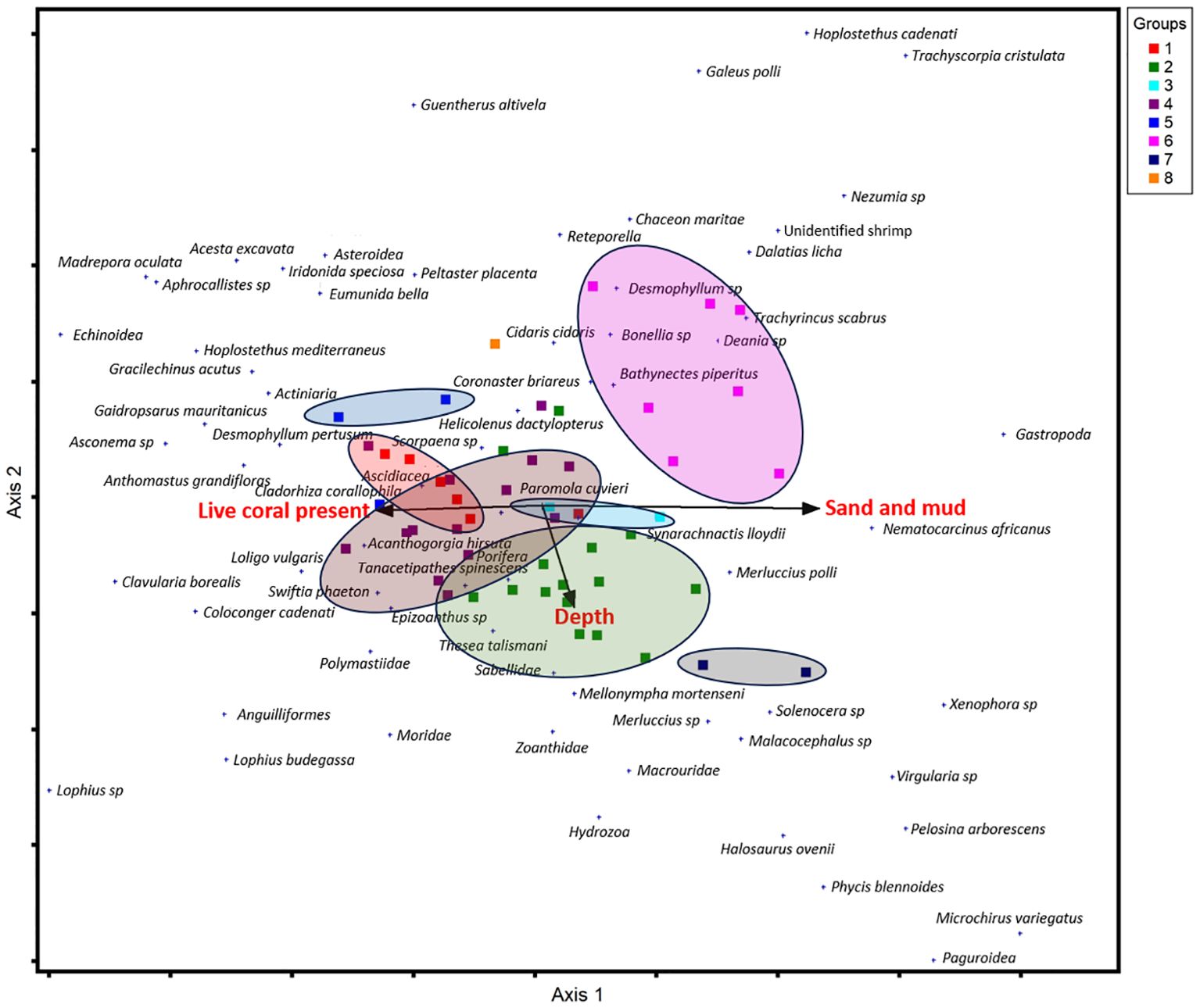
Figure 13. DCA ordination results for both axes (1 and 2), displaying the species community groups (1 to 7) colored according to the cluster results (Figure 12). The black line indicates the main environmental variables (live coral present, sandy mud and depth) related to the community composition.
Axis 1 of the DCA shows a clear substrate gradient, with living coral habitats on the left and soft substrate habitats (sand and mud) on the right. Axis 2 appears to have a lesser association with depth. These major environmental variables, denoted by black arrows, shed light on how environmental gradients influence different species compositions, as seen in the spatial clustering of species groups. The groups on the left side of the plot are substantially influenced by live corals, implying that these species benefit from the structural complexity of coral ecosystems. In contrast, species groups on the right side of the plot are mostly influenced by sand and mud, showing a preference for places dominated by soft sediments.
The DCA indicated that the depth environmental gradient may exert a weaker influence on species distribution compared to other variables. The DCA analysis did not show any patterns for the other environmental variables, such as rugosity and site.
Species in groups 1, 4 and 5, are all associated with the presence of live coral. Group 1 is strongly associated with live coral presence, while group 4 lies between the live coral presence and depth gradient, suggesting that transitional species benefit from coral habitats and are tolerant of deeper conditions. Although Group 5 is not directly overlapping with the live coral presence cluster, its special placement and species composition indicate that live coral may still play a role in shaping its distribution. In terms of common species in different groups, it is worth noting that S. cf. lloydii was abundant across the seven substrate types, indicating a broad distribution. Common species in these groups (Groups 1, 4, and 5) include several coral species such as D. pertusum, M. oculata, A. cf. hirsuta, S. phaeton, T. talismani, the crustacean E. bella, and the fish H. dactylopterus, suggesting a preference for complex structures provided by coral habitat that offer shelter and food for these species.
The sandy mud arrow points aligns with Group 3, 6 and 7, indicating an influence on species distribution. Group 6 shows the strongest association with this substrate, suggesting a preference for soft sediments. Although Group 3 is situated near live coral, slightly away from the live coral presence gradient, this Groupe includes species that are commonly associated with the structural complexity provided by live coral. Their presence in areas where coral declines and transition into sandy mud suggests a degree of tolerance to soft sediments.
Group 7 is located close to the depth gradient but shows some affinity for sandy mud substrates. Its species distributions suggested a potential influence for both depth and soft sediment. These groups (2,3 and 7) are dominated by species typically found on soft substrates, such as shrimp species, N. africanus and Solenocera sp., and sea pen Virgularia sp., and the fish T. scabrus.
Group 2 is located in the central part of the DCA, close to the transition zone between different habitats. It aligns moderately with the depth gradient. This position suggests that species of this group tolerate deeper conditions compared to live coral groups (e.g., Group 1), although they are not strict deep-water specialists. The two dominant species of this group are S. cf. lloydii and N. africanus.
One subsample (8) does not belong to a group and constitutes a single outlier sample. It contains very fewer species and doesn’t represent a specific affinity.
Overall, the DCA highlights that the two substrate types (live coral presence and sand and mud) appear to be the most important drivers of species distribution, while the depth gradient exerts a weaker influence on species distribution compared to other variables. The absence of some other environmental variables (e.g., site) from the DCA results may indicate that the site variable (north and south) does not influence the distribution of species groups, suggesting that the two reefs zone show similarities in their species communities. This is supported by the fact that the northern reef video samples (A6) are mixed with the southern samples (T2B and T3B) in the cluster dendrograms (Figure 12).
The correlation matrix for the environmental variables (Table 8) shows that live coral is positively related to rugosity (0.60) and negatively related to sand & mud (-0.47). The strong correlation between live coral and rugosity may explain why the rugosity was excluded from the DCA as a key environmental variable. Skeleton is positively related to live coral (0.33) and negatively related to sand and mud (-0.44), which is positively related to depth (0.25).
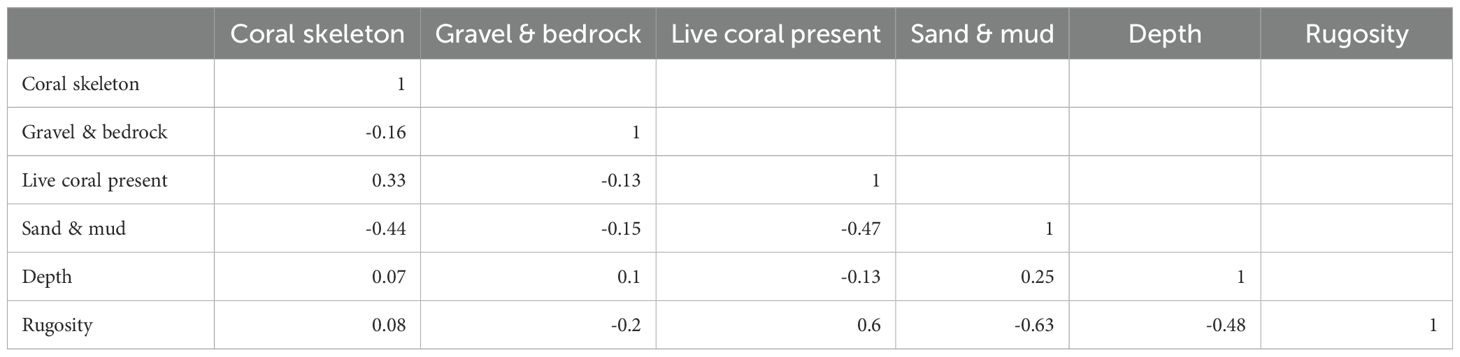
Table 8. Correlation table for environmental variables with the Pearson correlation coefficient.
4 Discussion
4.1 Methodology
To compare coral mound megafauna with areas without mounds, we used data from trawls with a mesh size ranging from 20 to 45 mm, depending on the vessels, which cannot provide representative samples of smaller taxa (< 2 cm). This limitation was evident in the low representation of certain taxa within Cnidaria and Echinodermata, and the diversity of the communities was underestimated. We compared the mound community occurring at 400 to 600 meters depth with off-mound communities at 200 to 350 meters. Thus, in addition to the differences in the presence of coral mounds, they also differed in depth-related environmental settings (e.g.; temperature, salinity, and dissolved oxygen). In the study area, the difference in oceanographic settings within the depth zone covered is not pronounced, which makes the presence of coral mounds with their effect on terrain, substratum, currents, etc., the main driver behind the observed faunal differences.
The megafauna associated with CWC reefs were studied using 11 ROV video transects, covering approximately, 6800 m2. Three transects were from the northern parts of the region and eight from the south. That is a few and disparate transects. The statistical analysis showed that the rich megafauna associated with the Mauritanian coral reefs is not different for reefs in the north and south. Even though the material is limited to 11 reef sites, the length of each transect ensures that the megafauna is well documented, and in addition, the present study is based on more ROV transects compared to the two previous studies of coral fauna in the region (Colman et al., 2005; Westphal et al., 2014).
4.2 Oceanography
Our results show the presence of two prominent oxygen minimum zones (OMZs): a shallower one at around 100 meters and a deeper, more pronounced one, near 400 meters. The deeper minimum zone coincides with the depth zone (400–650 m) in which coral mounds and CWC reefs occur in Mauritanian waters. In the depth zone of the CWC reefs, the recorded temperature ranged from 8.66 to 11.56°C, measured salinities ranged from 35.09 to 35.38 PSU, and oxygen concentration was from 1.17 to 1.53 ml L−1. This is in line with other studies of CWC mound off Mauritania; Ramos et al., 2017; Westphal et al., 2014). On the West African continental margin, cold water coral mounds, typically having only coral skeletons or a few living colonies, have been reported from areas with low oxygen concentrations (<1.5 mL L-1) off Angola and Namibia (Hanz et al., 2019; Le Guilloux et al., 2009).
Our study shows that a rich megafauna is associated with CWC mounds and reefs in the OMZ that clearly can tolerate the low oxygen concentrations. Recently, other studies have discovered live and healthy reefs at oxygen concentrations between 1.0 and 1.2 ml L−1 (Buhl-Mortensen et al., 2024) concluding that low oxygen concentrations are not hindering the growth of CWC reefs. The first alive and large CWC reef off Ghana was recorded from within the OMZ zone (Buhl-Mortensen et al., 2017), and live reefs from the Angola have been reported from the OMZ there (Hanz et al., 2019). Recently, healthy reefs were recorded in OMZ at the border between Mauritania and Senegal (Moctar et al., 2024). Our study shows that a rich megafauna is associated with CWC mounds and reefs in the OMZ that clearly also can tolerate low oxygen concentrations. Hebbeln et al. (2020) speculate that CWC thrive under these extreme conditions because they have developed certain adaptive metabolic strategies and that the negative effects of hypoxia could be compensated by the availability of enhanced food supply. On the other hand, Portilho-Ramos et al. (2022) demonstrated that low oxygen concentrations in deep waters can act as an additional stressor, while the food supply exerts the strongest effect on coral vitality.
4.3 Megafauna associated with CWC habitats
In this study, we use data from demersal trawling surveys to describe the species diversity related to coral mounds off Mauritania; this has not been done earlier in the region. Ramos et al. (2017) described the fauna colonizing coral mounds based on dredged material (13 rock dredge samplings), which cannot be compared directly with our observations due to the difference in methodology.
Diversity, as indicated by species richness, Shannon and Simpson indexes, and species accumulation curves, shows a richer community on the CWC mounds compared to the off-mound zone, and our results show a high-diversity community inhabiting coral mound including 282 taxa. The fish fauna on the coral mounds was dominated by the blackbelly rosefish H. dactylopterus, a species that is known to thrive in CWC habitats, and both juveniles and adults are frequently observed in CWC habitats in Northwest Africa (El Vadhel et al., 2024). Our observation confirms this relation to CWC, and other abundant fish species on the mound were L. laureysi, M. polli, C. caelorhincus, H. cadenati, M. occidentalis, G. darwinii and H. mediterraneu, of which several are known to be associated with CWC reefs (e.g., Capezzuto et al., 2018; Costello et al., 2005; Milligan et al., 2016).
In contrast to diversity, the biomass (total catch) and abundance (number of individuals) were twice as high in the off-mound zone compared to the mounds. The lower values obtained on-mound could be related to the coral mound structure acting as an obstacle that negatively affects the catch efficiency of the fishing gear. Another reason could be that in the non-coral zone, the species have gregarious behavior, which facilitates their capture by trawling in significant quantities.
The ROV transects documented that the CWC reefs had a rich associated megafauna of 120 taxa, belonging to 11 fauna groups. The most abundant species on the reef by group were: fish (H. dactylopterus, G. altivela, C. cadenati, T. scabrus, H. mediterraneus, Nezumia sp, Malacocephalus sp, M. polli, Moridae, H. cadenati, and L. budegassa), Cnidaria (S. cf. lloydii, A. cf. hirsuta, D. pertusum, S. phaeton, T. talismani, M. oculata and T. cf. spinescens), Crustacea (N. africanus, E. bella, Solenocera sp, B. piperitus and C. maritae), Mollusca, (G. mauritanicus and A. excavata), Echinodermata (C. cidaris, G. acutus), and Porifera (C. corallophila, Aphrocallistes sp, and M. mortenseni).
In comparison, Westphal et al. (2014) reported from the same region a total of 39 taxa belonging to 7 fauna groups (Fish (16), Crustacea (9), Cnidaria (6), Mollusca (5), Polychaeta, Echinodermata and Annelida were represented by 1 taxon each). More than 22 taxa (56%) from this study were also documented in the present study. The most common fishes documented by Westphal et al. (2014) (T. scabrus, Hoplostethus sp., H. dactylopterus, L. budegassa) were also common in our study. And a highlight observation was an aggregation of the G. altivela, the gregarious behavior of this species near CWC was observed also in the present study. Some other species reported by Westphal et al. (2014) that were also common in our study include the crustaceans (P. cuvieri, B. piperitus, E. bella, C. maritae), mollusks (A. excavata).
Westphal et al. (2014) recorded several coral species off Mauritania, mainly D. pertusum and M. oculata, Dendrophyllia cornigera and Acanthogorgia sp., of these, D. cornigera was not observed in this study, however, we recorded in addition the corals; S. phaeton, T. talismani, T. cf. spinescens, A. cf. grandifloras and C. borealis.
Many of the recorded species are known to be associated with CWCs from other studies (e.g., Buhl-Mortensen et al., 2017; Costello et al., 2005; Henry and Roberts, 2017). It is interesting that even when the studied reefs are in an oxygen minimum zone, they still host many of the same associates.
Our study has documented species that are new to the region from CWC reefs; an example is Cladorhiza corallophila, a carnivorous sponge species that was described by Göcke et al. (2016). This sponge was common in association with D. pertusa and M. oculata in our study. The octocoral Swiftia (Swiftia phaeton) was described by Sampaio et al. (2022) from the upper bathyal off Mauritania. This species was in the present study very common and the fourth most recorded species. Finally, specimens of a new fish of Gadiformes (Gaidropsarus mauritanicus) that was discovered by Knorrn et al. (2024) were recorded three times in our study (Figure 14).

Figure 14. ROV images showing the 3 species new to science recently discovered in the deep sea off Mauritania: (A) C. corallophila associated with D. pertusum and the fish T. scabrus, (B) S. phaeton, (C) G. mauritanicus with A. cf. hirsuta.
Our results reveal that both CWC mounds and reefs share several common species, despite the use of different sampling methods. These species, such as the fish H. dactylopterus, M. polli, H. cadenati, G. darwinii, L. budegassa, H. mediterraneus, and G. altivela, as well as the crustaceans P. cuvieri and S. africana occurred in both habitats. This overlap in species composition suggests that CWC mounds and reefs are ecologically linked and could act as a continuous essential habitat for some of the associated megafauna they have in common.
The megafauna associated with CWC is found in different sub habitats, including the coral related sub habitats (live coral, coral rubble, rubble and live), soft substrate (sandy mud) and hard substrate (bedrock, coral blocks, sandy gravel). Our results highlight the importance of coral substrates (coral rubble, rubble and live) for diversity. Indeed, more than 56% of the megafauna associated with CWC have been reported in this substrate. These results agree with Mortensen et al. (1995) and Wolfe et al. (2021), who found that the coral associated fauna is more abundant in the older and dead areas than in the live parts of actively growing reefs, where coral rubble creates complex microhabitats supporting diverse reef organisms.
The heatmap analysis of species abundance across the different substrate types, as well as the species-by-species correlation matrix, further reinforces the importance of habitat heterogeneity in structuring benthic communities on the CWC reefs off Mauritania. Species such as the fish H. dactylopterus, the crab E. bella and the bivalve A. excavata exhibited a strong preference for coral blocks and live corals, suggesting that these structures provide critical habitat complexity and refuge for these species (Buhl-Mortensen et al., 2010). Our results further showed the functional importance of the frameworks of the two corals, D. pertusum and M. oculata, in sustaining a range of faunal assemblages. The species-by-species correlation matrix showed strong correlations between these important reef-building corals and the megafauna that they are linked with. Overall, these findings emphasize the critical role of various substrate types in supporting a wide range of species, emphasizing the importance of preserving diverse substrate types in order to maintain biodiversity and ecosystem function.
According to the diversity-by-family analysis, the majority of total catches (61%) on coral mounds are found in the families Sebastidae, Merlucciidae, Lophiidae, Moridae, and Macrouridae, which include many commercially important species like M. polli, M. senegalensis, L. budegassa, L. vaillanti, H. dactylopterus, and others. This is in line with the findings of Fernandez-Peralta and Gonzalez (2017), who examined the demersal ichthyofaunal assemblages in Mauritanian deep waters (80 to 1860 m) and discovered that Sebastidae and Merlucciidae are among the main demersal fish families in biomasses, contributing significantly to the total catch with over 25%.
The detrended correspondence analysis (DCA) revealed that the main factors influencing species distribution in CWC reefs are the presence of live coral colonies and sandy mud substrate. The DCA highlighted the importance of habitat complexity provided by live corals in shaping species composition; this is aligning with (Buhl-Mortensen et al., 2010), who emphasized how species diversity is impacted by coral structure. This finding is further supported by De Haast et al. (2024), who highlighted that the live coral substratum has an impact on taxa assemblages and is the most important factor in determining species distributions, further supporting this finding.
Our study generally revealed a rich and diverse megafauna inhabiting the two CWC ecosystems off Mauritania: the chain of mounds and the live reefs. These valuable and vulnerable habitats are facing various challenges, including deep-sea trawling and hydrocarbon exploitation, in addition to the impacts of climate change. The southern reefs, located on the border between Mauritania and Senegal (Moctar et al., 2024), support a remarkable biodiversity, including a significant presence of live D. pertusum in an area of gas resource development and exploitation. This situation requires special attention to protect this rich and vulnerable habitat.
5 Conclusion
This study provides, for the first time, a comprehensive description of the megafauna associated with cold-water coral (CWC) reefs and mounds off Mauritania. The data used include demersal trawling surveys conducted over four decades on board research vessels in the Mauritanian deep sea, as well as ROV video dives conducted as part of two seafloor habitat mapping surveys in 2020 and 2021. Our study documented a rich diversity of species associated with these CWC ecosystems. The megafauna inhabiting coral mounds consists of 282 taxa from five species groups, dominated by fishes. This megafauna is more diverse compared to the off-mound area. The CWC reefs provide habitat for many species (120 taxa) divided into 11 groups, including certain species recently discovered in the deep sea off Mauritania, such as Cladorhiza corallophila, Swiftia phaeton, and Gaidropsarus mauritanicus, which are relatively new to science.
The most notable observation is that the two CWC ecosystems thrive in the extreme environmental condition of the oxygen minimum zone, yet they still offer habitat for several taxa, many of which are globally associated with CWC ecosystems in other regions. Characteristic species of CWC habitats include the corals Acanthogorgia cf. hirsuta, Desmophyllum pertusum, Swiftia phaeton, Thesea talismani, Madrepora oculata, Tanacetipathes cf. spinescens, the sea anemone Synarachnactis cf. lloydii, the fish Helicolenus dactylopterus, Laemonema laureysi, Merluccius polli, Hoplostethus cadenati, Trachyrincus scabrus, Hoplostethus mediterraneus and Gephyroberyx darwinii, the crustaceans Nematocarcinus africanus, Eumunida bella, Bathynectes piperitus, the echinoderm Cidaris cidaris, the mollusks Gigantidas mauritanicus and Acesta excavata and the poriferan Cladorhiza corallophila.
Our findings underscore the ecological significance of CWC habitats off Mauritania and offer essential insights for developing a national marine resource management plan. Given rising anthropogenic pressures such as oil and gas exploration, deep-sea fishing, and climate change, these ecosystems require urgent conservation efforts.
Data availability statement
The data underlying our findings are available upon request to FAO and the Mauritanian Institute of Oceanographic Research and Fisheries (IMROP).
Ethics statement
Ethical approval was not required for the study involving animals in accordance with the local legislation and institutional requirements because the study is based on existing datasets.
Author contributions
HE: Conceptualization, Data curation, Formal analysis, Methodology, Visualization, Writing – original draft, Writing – review & editing. LB-M: Conceptualization, Formal analysis, Methodology, Supervision, Writing – review & editing. MM: Data curation, Formal analysis, Methodology, Writing – review & editing. YE: Data curation, Formal analysis, Methodology, Writing – review & editing. BM: Conceptualization, Writing – review & editing. FL: Supervision, Writing – review & editing. LM: Conceptualization, Formal analysis, Methodology, Supervision, Writing – review & editing.
Funding
The author(s) declare that no financial support was received for the research and/or publication of this article.
Acknowledgments
The authors are grateful for the invaluable support of many taxonomists for the identification of certain species from ROV images. We would like to think Samuel P. IGLESIAS from the Muséum national d’Histoire naturelle in Concarneau, France, and Peter Nick Psomadakis from FAO for their identification of fish species. We also thank Alexander H. Knorrn from Senckenberg am Meer, Germany for identifying the new fish species Gaidropsarus mauritanicus. Tina Molodtsova from the P.P. Shirshov Institute of Oceanology, Moscow, Russia for identifying black corals, Íris Sampaio from Portuguese Blue Biobank Manager (CIIMAR) for identifying the new octocoral Swiftia phaeton and Diana Zaera-Perez from the Institute of Marine Research, Norway for identifying the shark species. This paper uses data collected during two surveys with the research vessel Dr. Fridtjof Nansen as part of the collaboration between the Food and Agriculture Organization of the United Nations (FAO) on behalf of the EAF-Nansen Programme. The EAF-Nansen Programme is a partnership between the FAO, the Norwegian Agency for Development Cooperation (Norad), and the Institute of Marine Research (IMR) in Norway for sustainable management of the fisheries in partner countries and regions. We are grateful to all survey participants for their help and support during the data collection.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Author’s note
The views expressed in this publication are those of the author(s) and do not necessarily reflect the views or policies of the Food and Agriculture Organization of the United Nations and the Institute of Marine Research.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Addamo A. M., Vertino A., Stolarski J., García-Jiménez R., Taviani M., and Machordom A. (2016). Merging scleractinian genera: The overwhelming genetic similarity between solitary Desmophyllum and colonial Lophelia. BMC Evolutionary Biol. 16, 108. doi: 10.1186/s12862-016-0654-8, PMID: 27193263
Bongiorni L., Mea M., Gambi C., Pusceddu A., Taviani M., and Danovaro R. (2010). Deep-water scleractinian corals promote higher biodiversity in deep-sea meiofaunal assemblages along continental margins. Biol. Conserv. 143, 1687–1700. doi: 10.1016/j.biocon.2010.04.009
Braham C. B. (2013). “Les pêcheries pélagiques de la ZEE mauritanienne: nouvelles méthodes pour de nouvelles connaissances,” in Thesis, montpellier. (Montpellier, France: Université de Montpellier 2), 2. Available at: http://www.theses.fr/2013MON20248/document (Accessed August 2024).
Braham C.-B., Fréon P., Laurec A., Demarcq H., and Bez N. (2014). New insights in the spatial dynamics of sardinella stocks off Mauritania (North-West Africa) based on logbook data analysis. Fisheries Res. 154, 195–204. doi: 10.1016/j.fishres.2014.02.020
Brahim K., Taleb H., Abou E., Meissa B., Braham C.-B., Bouzouma M., et al. (2021). Rapport de la neuvième édition du groupe de travail de l’IMROP Nouadhibou, du 11 au 14 février 2020: Aménagement des ressources halieutiques et gestion de la biodiversité au service du développement durable. (Nouadhibou, Mauritania: IMROP). doi: 10.13140/RG.2.2.18840.93445
Bridges A. E. H., Howell K. L., Amaro T., Atkinson L., Barnes D. K. A., Bax N., et al. (2023). “Review of the central and south atlantic shelf and deep-sea benthos: science, policy, and management,” in Oceanography and marine biology, 1st ed. Eds. Hawkins S. J., Russell B. D., and Todd P. A. (Boca Raton, Florida, USA: CRC Press), 127–218. doi: 10.1201/9781003363873-5
Buhl-Mortensen L., Houssa R., M’bengue B., Nyadjro E. S., Cervantes D., Idrissi M., et al. (2024). Lophelia reefs off North and West Africa–Comparing environment and health. Mar. Biol. 171, 29. doi: 10.1007/s00227-023-04344-8
Buhl-Mortensen L., Serigstad B., Buhl-Mortensen P., Olsen M. N., Ostrowski M., Błażewicz-Paszkowycz M., et al. (2017). First observations of the structure and megafaunal community of a large Lophelia reef on the Ghanaian shelf (the Gulf of Guinea). Deep Sea Res. Part II: Topical Stud. Oceanography 137, 148–156. doi: 10.1016/j.dsr2.2016.06.007
Buhl-Mortensen L., Vanreusel A., Gooday A. J., Levin L. A., Priede I. G., Buhl-Mortensen P., et al. (2010). Biological structures as a source of habitat heterogeneity and biodiversity on the deep ocean margins: Biological structures and biodiversity. Mar. Ecol. 31, 21–50. doi: 10.1111/j.1439-0485.2010.00359.x
Camp L., Nykjaer L., Mittelstaedt E., and Schlittenhardt P. (1991). Upwelling and boundary circulation off Northwest Africa as depicted by infrared and visible satellite observations. Prog. In Oceanography 26, 357–402. doi: 10.1016/0079-6611(91)90012-B
Capezzuto F., Sion L., Ancona F., Carlucci R., Carluccio A., Cornacchia L., et al. (2018). Cold-water coral habitats and canyons as Essential Fish Habitats in the southern Adriatic and northern Ionian Sea (central Mediterranean). Ecol. Questions 29, 9. doi: 10.12775/EQ.2018.019
Carr M.-E. and Kearns E. J. (2003). Production regimes in four Eastern Boundary Current systems. Deep Sea Res. Part II: Topical Stud. Oceanography 50, 3199–3221. doi: 10.1016/j.dsr2.2003.07.015
Cianca A., Santana R., Marrero J. P., Rueda M. J., and Llinás O. (2009). Modal composition of the central water in the North Atlantic subtropical gyre. Ocean Sci. Discuss. 6, 2487–2506. doi: 10.5194/osd-6-2487-2009
Colman J., Gordon D., Lane A., Forde M., and Fitzpatrick J. (2005). Carbonate mounds off Mauritania, Northwest Africa: Status of deep-water corals and implications for management of fishing and oil exploration activities. In Cold-water Corals Ecosyst. pp, 417–441). doi: 10.1007/3-540-27673-4_21
Costello M. J., McCrea M., Freiwald A., Lundälv T., Jonsson L., Bett B. J., et al. (2005). “Role of cold-water Lophelia pertusa coral reefs as fish habitat in the NE Atlantic,” in Cold-water corals and ecosystems. Eds. Freiwald A. and Roberts J. M. (Berlin, Germany: Springer Berlin Heidelberg), 771–805. doi: 10.1007/3-540-27673-4_41
D’Onghia G., Maiorano P., Carlucci R., Capezzuto F., Carluccio A., Tursi A., et al. (2012). Comparing deep-sea fish fauna between coral and non-coral “Megahabitats” in the santa maria di leuca cold-water coral province (Mediterranean sea). PloS One 7, e44509. doi: 10.1371/journal.pone.0044509, PMID: 23028551
De Haast J. A., Moloney C. L., Buhl-Mortensen L., and Karenyi N. (2024). Relationships between taxa assemblages and substratum types on cold-water coral reefs. Estuarine. Coast. Shelf Sci. 313, 109101. doi: 10.1016/j.ecss.2024.109101
Eisele M., Frank N., Wienberg C., Hebbeln D., López Correa M., Douville E., et al. (2011). Productivity controlled cold-water coral growth periods during the last glacial off Mauritania. Mar. Geology 280, 143–149. doi: 10.1016/j.margeo.2010.12.007
El Vadhel H., Buhl-Mortensen L., Babou D. A., Dridi A., Balde B. S., Bouzouma M. E. M., et al. (2024). Do cold water corals provide an essential habitat for Helicolenus dactylopterus (Delaroch) in the Northwest Africa? Mar. Environ. Res. 198, 106538. doi: 10.1016/j.marenvres.2024.106538, PMID: 38782662
Emery W. J. (2001). “Water types and water masses,” in Encyclopedia of ocean sciences (Amsterdam, Netherlands: Elsevier), 3179–3187. doi: 10.1006/rwos.2001.0108
Fernandez-Peralta L. and Gonzalez M. A. (2017). “Demersal ichthyofaunal assemblages in Mauritanian deep-waters,” in Deep-sea ecosystems off Mauritania: research of marine biodiversity and habitats in the northwest african margin, (Dordrecht, Netherlands: Springer), 155–199. doi: 10.1007/978-94-024-1023-5_4
Freiwald A., Beuck L., Lundälv T., and Wienberg C. (2014).General morphology and preliminary biological inventory of the Mauritanian cold-water coral habitats in Phaeton—Paleoceanographic and paleo-climatic record on the Mauritanian ShelfEdsWestphal H., Beuck L., Braun S., Freiwald A., Hanebuth T., Hetzinger S., Klicpera A., Kudrass H., Lantzsch H., Lundälv T., Mateu-Vicens G., Preto N., Reumont J., Schilling S., Taviani M., and Wienberg C.. (Hamburg, Germany: DFG Senatskommission für Ozeanographie). doi: 10.2312/cr_msm16_3
Gascuel D., Labrosse P., Meissa B., Taleb Sidi M., and Guénette S. (2007). Decline of demersal resources in North-West Africa: An analysis of Mauritanian trawl-survey data over the past 25 years. Afr. J. Mar. Sci. 29, 331–345. doi: 10.2989/AJMS.2007.29.3.3.333
Göcke C., Hestetun J., Uhlir C., Freiwald A., BEUCK L., and Janussen D. (2016). Cladorhiza corallophila sp. Nov., a new carnivorous sponge (Cladorhizidae, Demospongiae) living in close association with Lophelia pertusa and Madrepora oculata (Scleractinia). Zootaxa 4168, 512. doi: 10.11646/zootaxa.4168.3.4, PMID: 27701324
Hagen E. (2001). Northwest african upwelling scenario. Oceanologica Acta 24, 113–128. doi: 10.1016/S0399-1784(00)01110-5
Hanz U., Wienberg C., Hebbeln D., Duineveld G., Lavaleye M., Juva K., et al. (2019). Environmental factors influencing benthic communities in the oxygen minimum zones on the Angolan and Namibian margins. . Biogeosciences 16, 4337–4356. doi: 10.5194/bg-16-4337-2019
Hebbeln D., Bender M., Gaide S., Titschack J., Vandorpe T., Van Rooij D., et al. (2019). Thousands of cold-water coral mounds along the Moroccan Atlantic continental margin: Distribution and morphometry. Mar. Geology 411, 51–61. doi: 10.1016/j.margeo.2019.02.001
Hebbeln D., Van Rooij D., and Wienberg C. (2016). Good neighbours shaped by vigorous currents: Cold-water coral mounds and contourites in the North Atlantic. Mar. Geology 378, 171–185. doi: 10.1016/j.margeo.2016.01.014
Hebbeln D., Wienberg C., Dullo W.-C., Freiwald A., Mienis F., Orejas C., et al. (2020). Cold-water coral reefs thriving under hypoxia. Coral Reefs 39, 853–859. doi: 10.1007/s00338-020-01934-6
Henry L. A. and Roberts J. (2017). “Global biodiversity in cold-water coral reef ecosystems,” in Marine animal forests: the ecology of benthic biodiversity hotspots. (Cham, Switzerland: Springer International Publishing AG), 235–256. doi: 10.1007/978-3-319-21012-4_6
Hoffman L., Monsecour K., and Freiwald A. (2019). Columbellidae (Gastropoda) from deep-water coral habitats off Mauritania. Miscellanea Malacologica 8, 41–49.
Knorrn A. H., Beuck L., Barros-García D., Fernández-Peralta L., and Freiwald A. (2024). Gaidropsarus mauritanicus (Gadiformes, Gaidropsaridae) a new three-bearded rockling from a deep-water coral ecosystem with a genetically verified biogeographical distribution of the genus and notes to its ecology and behavior. J. Fish Biol. 105, jfb.15859. doi: 10.1111/jfb.15859, PMID: 39150971
Krastel S., Wynn R., Hanebuth T., Henrich R., Holz C., Meggers H., et al. (2006). Mapping of seabed morphology and shallow Sediment structure of the Mauritania continental margin, Northwest Africa: Some implications for geohazard potential. Norwegian J. Geology 86, 163–176. doi: 10.1007/s00531-006-0112-8
Kutti T., Bergstad O., Fossa J., and Helle K. (2013). Cold-water coral mounds and sponge-beds as habitats for demersal fish on the Norwegian shelf. Deep Sea Res. Part II Topical Stud. Oceanography. 99, 122–133. doi: 10.1016/j.dsr2.2013.07.021
Le Guilloux E., Olu K., Bourillet J. F., Savoye B., Iglésias S. P., and Sibuet M. (2009). First observations of deep-sea coral reefs along the Angola margin. Deep Sea Res. Part II: Topical Stud. Oceanography 56, 2394–2403. doi: 10.1016/j.dsr2.2009.04.014
McCune B. and Mefford M. J. (1999). “PC-ORD multivariate analysis of ecological data,” in Version 4. MjM software design. (Gleneden Beach, Oregon, USA: MjM Software Design).
Meissa B. and Gascuel D. (2015). Overfishing of marine resources: Some lessons from the assessment of demersal stocks off Mauritania. ICES J. Mar. Science: J. Du Conseil 72, 414–427. doi: 10.1093/icesjms/fsu144
Meissa B., Gascuel D., and Rivot E. (2013). Assessing stocks in data-poor African fisheries: a case study on the white grouper (Epinephelus aeneus) of Mauritania. Afr. J. Mar. Sci. 35, 253–267. doi: 10.2989/1814232X.2013.798244
Milligan R., Spence G., Roberts J., and Bailey D. (2016). Fish communities associated with cold-water corals vary with depth and substratum type. Deep Sea Research Part I: Oceanographic Research Papers. 114, 43–54. doi: 10.1016/j.dsr.2016.04.011
Moctar S. M. M., Buhl-Mortensen L., El Vadhel H., Kide S. O., Kutti T., Sarre A., et al. (2024). Lophelia reefs (Desmophyllum pertusum (Linnaeu)) in the oxygen minimum zone of the Mauritania/Senegal region – Distribution and health status. Deep Sea Res. Part I: Oceanographic Res. Papers 208, 104322. doi: 10.1016/j.dsr.2024.104322
Mortensen P. B. and Fosså J. H. (2006). Species diversity and spatial distribution of invertebrates on deep-water Lophelia reefs in Norway. Proceedings of the 10th International Coral Reef Symposium, 1849–1868. Japan: International Society for Reef Studies, Japanese Coral Reef Society.
Mortensen P. B., Hovland M., Brattegard T., and Farestveit R. (1995). Deep water bioherms of the scleractinian coral Lophelia pertusa (L.) at 64° n on the Norwegian shelf: Structure and associated megafauna. Sarsia 80, 145–158. doi: 10.1080/00364827.1995.10413586
Oksanen J., Simpson G., Blanchet F., Kindt R., Legendre P., Minchin P., et al. (2022). “vegan: community ecology package,” in R package version 2.6-4. (Vienna, Austria: Comprehensive R Archive Network (CRAN)). Available at: https://CRAN.R-project.org/package=vegan (Accessed October 2024).
Pelegrí J. L., Peña-Izquierdo J., Machín F., Meiners C., and Presas-Navarro C. (2017). “Oceanography of the cape verde basin and Mauritanian slope waters,” in Deep-sea ecosystems off Mauritania. Eds. Ramos A., Ramil F., and Sanz J. L. (Dordrecht, Netherlands: Springer Netherlands), 119–153. doi: 10.1007/978-94-024-1023-5_3
Peña-Izquierdo J., Van Sebille E., Pelegrí J. L., Sprintall J., Mason E., Llanillo P. J., et al. (2015). Water mass pathways to the N orth A tlantic oxygen minimum zone. J. Geophysical Research: Oceans 120, 3350–3372. doi: 10.1002/2014JC010557
Portilho-Ramos R., Titschack J., Wienberg C., Siccha M., Yokoyama Y., and Hebbeln D. (2022). Major environmental drivers determining life and death of cold-water corals through time. PloS Biol. 20, e3001628. doi: 10.1371/journal.pbio.3001628, PMID: 35587463
Ramos A., Sanz J., Ramil F., Agudo Bravo L. M., and Presas-Navarro C. (2017). “The giant cold-water coral mounds barrier off Mauritania,” in Deep-sea ecosystems off Mauritania: research of marine biodiversity and habitats in the northwest african margin. (Dordrecht, Netherlands: Springer), 481–525. doi: 10.1007/978-94-024-1023-5_13
Roberts J., Wheeler A., and Freiwald A. (2006). Reefs of the deep: the biology and geology of cold-water coral ecosystems. Science (New York, N.Y) 312, 543–547. doi: 10.1126/science.1119861, PMID: 16645087
Roberts J. M., Wheeler A., Freiwald A., and Cairns S. (2009). Cold-water corals: the biology and geology of deep-sea coral habitats (Cambridge, United Kingdom: Cambridge University Press). doi: 10.1017/CBO9780511581588
Ross S. W. and Quattrini A. M. (2007). The fish fauna associated with deep coral banks off the southeastern United States. Deep Sea Res. Part I: Oceanographic Res. Papers 54, 975–1007. doi: 10.1016/j.dsr.2007.03.010
Sampaio Í., Beuck L., and Freiwald A. (2022). A new octocoral species of Swiftia (Holaxonia, Plexauridae) from the upper bathyal off Mauritania (NE Atlantic). ZooKeys 1106, 121–140. doi: 10.3897/zookeys.1106.81364, PMID: 36760817
Sanz J., Maestro A., and Agudo Bravo L. M. (2017). “The Mauritanian margin,” in Bathymetric and geomorphological characteristics. In deep-sea ecosystems off Mauritania: research of marine biodiversity and habitats in the northwest african margin. (Dordrecht, Netherlands: Springer), 53–117. doi: 10.1007/978-94-024-1023-5_2
Tamborrino L., Wienberg C., Titschack J., Wintersteller P., Mienis F., Schröder-Ritzrau A., et al. (2019). Mid-Holocene extinction of cold-water corals on the Namibian shelf steered by the Benguela oxygen minimum zone. Geology 47, 1185–1188. doi: 10.1130/G46672.1
Tomczak M. and Godfrey J. S. (1994). “Antarctic oceanography,” in Regional oceanography (Amsterdam, Netherlands: Elsevier), 67–87. doi: 10.1016/B978-0-08-041021-0.50010-2
Westphal H., Beuck L., Braun S., Freiwald A., Hanebuth T., Hetzinger S., et al. (2014). Phaeton—Paleoceanographic and paleo-climatic record on the Mauritanian Shelf. (Hamburg, Germany: DFG Senatskommission für Ozeanographie). doi: 10.2312/cr_msm16_3
Wickham H. (2016). ggplot2: elegant graphics for data analysis (New York: Springer-Verlag). doi: 10.1007/978-3-319-24277-4_9
Wienberg C., Freiwald A., Frank N., Mienis F., Titschack J., Orejas C., et al. (2023). “Cold-water coral reefs in the oxygen minimum zones off west africa,” in Cold-water coral reefs of the world, vol. 19 . Eds. Cordes E. and Mienis F. (Cham, Switzerland: Springer International Publishing), 199–235. doi: 10.1007/978-3-031-40897-7_8
Wienberg C. and Titschack J. (2015). “Framework-forming scleractinian cold-water corals through space and time: A late quaternary north atlantic perspective,” in Marine animal forests. Eds. Rossi S., Bramanti L., Gori A., and Orejas Saco Del Valle C. (Cham, Switzerland: Springer International Publishing), 1–34. doi: 10.1007/978-3-319-17001-5_16-1
Wienberg C., Titschack J., Freiwald A., Frank N., Lundälv T., Taviani M., et al. (2018). The giant Mauritanian cold-water coral mound province: Oxygen control on coral mound formation. Quaternary Sci. Rev. 185, 135–152. doi: 10.1016/j.quascirev.2018.02.012
Wolfe K., Kenyon T. M., and Mumby P. J. (2021). The biology and ecology of coral rubble and implications for the future of coral reefs. Coral Reefs 40, 1769–1806. doi: 10.1007/s00338-021-02185-9
Keywords: cold water corals (CWC), coral mounds, coral reefs, megafauna associated, Mauritania
Citation: El Vadhel H, Buhl-Mortensen L, Mahmoud MEKM, El Vally Y, Meissa B, Le Loc’h F and Menot L (2025) Cold-water coral mounds and reefs off Mauritania and associated megafauna. Front. Mar. Sci. 12:1575636. doi: 10.3389/fmars.2025.1575636
Received: 12 February 2025; Accepted: 19 June 2025;
Published: 16 July 2025.
Edited by:
Elva G. Escobar-Briones, National Autonomous University of Mexico, MexicoReviewed by:
Anna Maria Addamo, Nord University, NorwayJacek Raddatz, Helmholtz Association of German Research Centres (HZ), Germany
Ana Colaço, University of the Azores, Portugal
Copyright © Food and Agriculture Organization of the United Nations and Institute of Marine Research 2025. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY), which permits unrestricted use, adaptation (including derivative works), distribution, and reproduction in any medium, provided the original work is properly cited. In any reproduction or adaptation of this article there should not be any suggestion that FAO/IMR or this article endorse any specific organisation or products. The use of the FAO/IMR logo is not permitted. This notice should be preserved along with the article’s original URL.
*Correspondence: Hammoud El Vadhel, aGFtbW91ZGVsdmFkZWxAZ21haWwuY29t
†ORCID: Hammoud El Vadhel, orcid.org/0009-0001-0707-1595