 Cristina González-Andrés
Cristina González-Andrés Beatriz Guijarro
Beatriz Guijarro Eduardo Ramírez-Romero
Eduardo Ramírez-Romero Enric Massutí1
Enric Massutí1- 1Centre Oceanogràfic de les Balears (COB-IEO), Consejo Superior de Investigaciones Científicas (CSIC), Palma, Spain
- 2Instituto de Ciencias Marinas de Andalucía (ICMAN), Consejo Superior de Investigaciones Científicas (CSIC), Puerto Real, Cadiz, Spain
- 3Department of Ecology and Geology, Faculty of Sciences, University of Malaga, Málaga, Spain
In the context of global change, studying the synergic effects of climate and marine resources is key to understanding and predicting their impact on exploited stocks and improving adaptive fisheries management. In the western Mediterranean, European hake (Merluccius merluccius) and deep-water rose shrimp (Parapenaeus longirostris) are two key demersal species with distinct ecological preferences. This study models long-term projection of their population dynamics at two geographical subareas (GSAs) established by the General Fisheries Commission for the Mediterranean (FAO-GFCM), Balearic Islands (GSA 5) and Northern Spain (GSA 6), under different management strategies and climatic scenarios. The methodological approach followed three steps. First, recruitment and spawning stock biomass temporal series of these stocks were obtained from fisheries assessment models, developed within the framework of the FAO-GFCM. Then the influence of parental stock and environmental drivers on their recruitment was modelled. In GSA 5, European hake recruitment was mainly driven by winter sea surface temperature, while in GSA 6, additional factors included chlorophyll-a and mean salinity. For deep-water rose shrimp, bottom temperature was the key driver in both GSAs. Lastly, we projected the population dynamics of the stocks along the whole 21st century by combining five potential fishing management strategies, including those currently applied in the investigated area (European Union Multiannual Plan) and the models of main explanatory drivers developed concurrently with an ensemble of four Regional Climate Models under two climatic scenarios (RCP4.5 and RCP8.5) adopted by the Intergovernmental Panel on Climate Change, which describe medium and extreme heating. From the combination of these management strategies and climatic scenarios, projections of the population parameters of European hake and deep-water rose shrimp and its catches were carried out up to 2100. The projections revealed a decline in European hake recruitment, population, and catches across all climate scenarios, regardless of fishery management strategies. In contrast, deep-water rose shrimp showed an overall improvement in population parameters and catches under all conditions. Our results may enhance scientific advice for implementing an adaptive fisheries management and highlight the need of integrating climatic effects in the assessment and management processes to enhance stock sustainability under global change.
1 Introduction
Fishing exploitation is a well-known stressing factor for marine ecosystems, modifying not only the population structure of the main target and by-catch species, but also producing indirect effects in populations and communities (Branch et al., 2010). Aside from this, the influence of environmental variability on the population dynamics of exploited species has been a key point in fisheries oceanography since its beginnings (Hjort, 1914). Authors such as Duffy-Anderson et al. (2005) and Harley et al. (2006) pointed out that local, regional and large-scale physical factors are of great importance in the communities and the population dynamics, highlighting the influence of climate on the production patterns of marine ecosystems.
In the current context of global change, both climate change and overfishing has been demonstrated as responsible factors of a quick restructuration of highly productive marine ecosystems, with effects in the pelagic and benthic domains (Kirby et al., 2009). The current vision is that it is not possible to separate the effects induced by both climate and fishing (e.g. Planque et al., 2010), as they probably act in a synergic way (Kirby et al., 2009). Fishing exploitation can produce demographic changes in exploited populations, which make them less resilient and more vulnerable in front of changes of environmental conditions (Stenseth and Rouyer, 2008). Hence, the study of synergic effects of climate and fishing in exploited communities is basic for the improvement of fisheries oceanography. This allowed in the last years developing of different approaches to predict the consequences of climate change in fisheries, their main threats and opportunities, as well as to assess different strategies for adapting to these changes (Daw et al., 2009).
In the case of fisheries, it is also important to know in advance any change and its potential effects, for providing scientific advice to manage the fisheries at medium and long term. For that, it is needed to develop predictions and projections, which are considered among the most valuables products of applied fisheries research (Brander et al., 2013), as they provide a quantitative basis for assessing strategies and adapting measures. Such predictions and projections should be made at a regional and local scale, where historical data can contribute to improve complex models, as global models have a limited value (Brander, 2015).
European hake (Merluccius merluccius) and deep-water rose shrimp (Parapenaeus longirostris) are two key fishery resources in the Mediterranean, each with distinct distributions and ecological preferences. Catches and abundance of both species have shown interannual changes off north-eastern Iberian Peninsula and the Balearic Islands (western Mediterranean), suggesting the influence of environmental factors on their distribution and population dynamics (Massutí et al., 2008; Hidalgo et al., 2011; Maynou, 2011; Sbrana et al., 2019; González-Andrés et al., 2024; Mingote et al., 2024, among others). These studies indicate that higher temperatures enhance the abundance and distribution of deep-water rose shrimp, a thermophilic species, but in the case of European hake, which is a cold adapted Atlantic species (Lloris et al., 2003), its variability is more related to productivity changes. Thus, the role of environmental drivers on their recruitment and population dynamics have been analyzed, but they did not project future scenarios under combined climate and management conditions. In fact, there is limited work combining climate projections with alternative fishing management strategies to simulate long-term stock dynamics of these species under climate change, particularly using ensembles of regional climate models. For instance, in the Adriatic Sea (Central Mediterranean), Panzeri et al. (2024) predicted the future spatial distribution of some demersal species, including European hake, under a warming scenario, but without considering potential changes from fishing management measures.
This study aims to fill this gap by modelling mid- and long-term projections of the population dynamics of European hake and deep-water rose shrimp at the two geographical subareas established by the General Fisheries Commission for the Mediterranean (FAO-GFCM), Balearic Islands (GSA 5) and Northern Spain (GSA 6) (Figure 1). For that, we integrated environmental drivers, derived from regional climate models and fishing management strategies, inspired by the current European Union Multiannual Plan (EU-MAP). From the available knowledge on both species, we hypothesize that the thermophilic species P. longirostris will be more positively affected by warmer waters than European hake and that the recovery of this latter species may be influenced by its current state of exploitation, as it is considered to be the most overexploited species in the Mediterranean (GFCM, 2024).
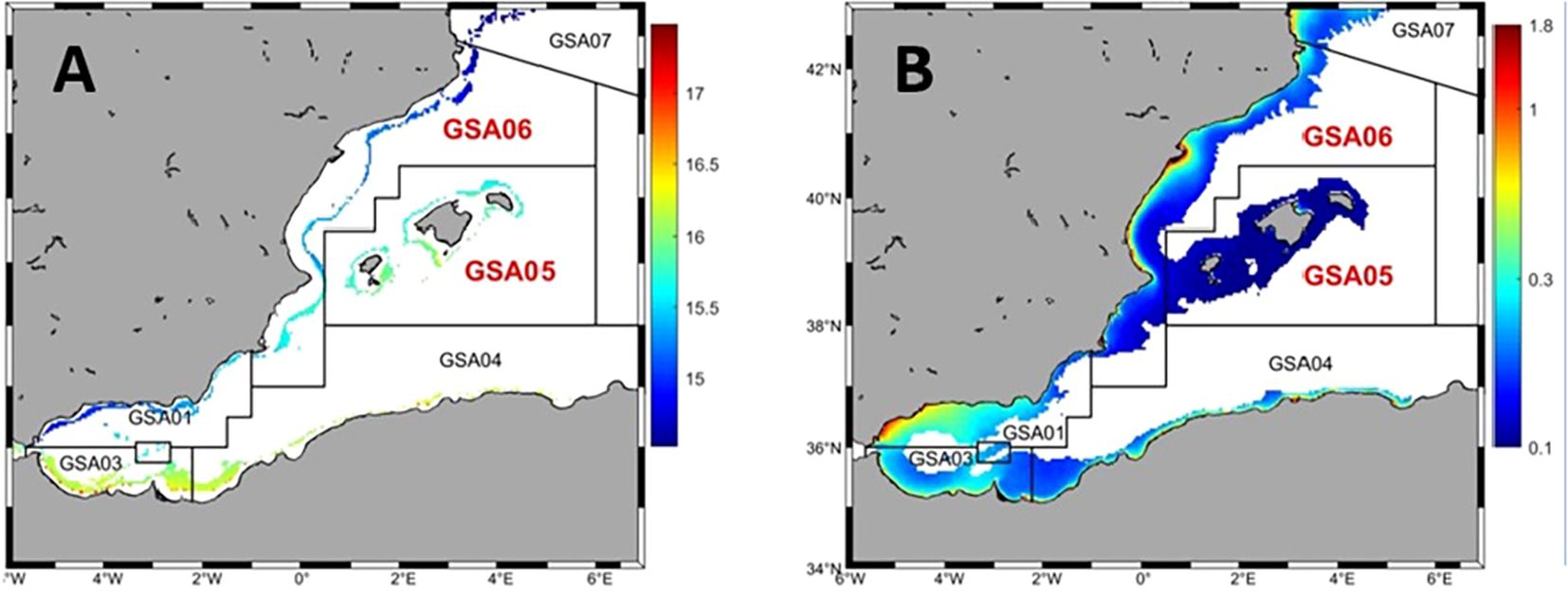
Figure 1. Maps of the western Mediterranean showing the climatology of annual values of sea bottom temperatura (°C) (A; between 150 and 400 m depth) and surface chlorophylln (mg*m-3) (B; log10 transformed satellite data). The FAO-GFCM geographical sub-areas considered in the present study, GSA 5 (Balearic Islands) and GSA 6 (Northern Spain), both in red, are also shown.
2 Study area and species
Within the general oligotrophic environment of the Mediterranean, the north-eastern Iberian Peninsula and the Gulf of Lions stand out as highly productive zones, due to several oceanographic features. These include a wide continental shelf, significant land run-off (e.g. from Rhône and Ebro rivers) and intense vertical water mixing in winter, induced by strong, dry, and cold north-westerly winds, which occasionally lead to coastal upwelling. These factors collectively enhance nutrient availability in the marine ecosystem, resulting in increased planktonic production (Estrada, 1996; Bosc et al., 2004). This high productivity benefits various fish and invertebrate fishing stocks, including European hake (Lloret et al., 2001).
In contrast, the waters around the Balearic Islands show a more pronounced oligotrophy, due to the lack of the previous oceanographic features (Estrada, 1996; Bosc et al., 2004). Frontal meso-scale events between Mediterranean and Atlantic waters (Pinot et al., 2002) and input of cold northern water into the Balearic channels (Fernández de Puelles et al., 2004) can act as external fertilization mechanisms that enhance productivity off the Archipelago. This cold water is originated in the north-easternmost Iberian Peninsula and the Gulf of Lions by deep convection (Pinot et al., 2002), producingthe Northern Current which flows southwards along the continental shelf edge and upper slope of the Iberian Peninsula. This current bifurcates when reaches the Ibiza Channel: one part forms the Balearic Current, which flows along the northern coast of the Balearic Islands (Monserrat et al., 2008). The major or minor formation of this cold water affects the interannual variability of hydrodynamism and productivity around the Archipelago (Massutí et al., 2014) influencing also biology and population dynamics of deep-water species, including European hake and deep-water rose shrimp (Massutí et al., 2008; Quetglas et al., 2012; Rueda et al., 2014). Such regional oceanographic differences likely influence the life history strategies of key demersal species such as the longer lifespan and protracted spawning period of European hake versus the faster growth and shorter lifespan of deep-water rose shrimp thereby modulating their population responses to environmental variability.
Due to its geomorphological, oceanographic and fishing specificities, the Balearic Islands have been considered as an individualized area for the assessment and management of fisheries in the western Mediterranean (Quetglas et al., 2013). According to these authors, the demersal resources and the fishing exploited benthic ecosystems in GSA 5 are in a healthier state than in the adjacent Iberian Peninsula (GSA 6), in part because the trawl fishing effort in GSA 5 is much lower than in GSA 6.
The European hake (M. merluccius) is a demersal gadoid fish species that prefers cold and temperate waters (Olivar et al., 2003; Maynou et al., 2003), with a wide geographic distribution covering the north-eastern Atlantic, from Norway and Iceland to Mauritania, including the Mediterranean (Casey and Pereiro, 1995; Oliver and Massutí, 1995). It has great ecological and economic importance both in the Atlantic and the Mediterranean (Alheit and Pitcher, 1995). In this last area, it is distributed between 30 and 1000m depth, although the largest abundance and biomass and optimal depth range occur between 100 and 400m (Relini et al., 2002; Sion et al., 2019). It is one of the main target species of the Mediterranean fisheries, mainly exploited by bottom trawling, but also by other gears such as bottom longlines and gillnets (Oliver and Massutí, 1995). This species historically shows the highest levels of overexploitation in the area (Colloca et al., 2014; GFCM, 2024).
The deep-water rose shrimp (P. longirostris) is a demersal decapod crustacean with a wide geographic distribution along the Atlantic Ocean. In its eastern part it is found from northern Iberian Peninsula to southern Angola, including the Mediterranean Sea (Sobrino et al., 2005). It is a fast-growing species, with a thermophilic preference, that inhabits muddy and sandy bottoms mainly of the deep shelf and upper slope between 100 and 400m depth (Fortibuoni et al., 2010). Deep-water rose shrimp is also a key fisheries resource in the Mediterranean (Sbrana et al., 2006; Guijarro et al., 2009). Its bathymetric distribution in the Spanish Mediterranean coast (GSAs 1, 5 and 6) has historically been defined between 20 and 750m, with the greatest densities in terms of biomass recorded between 100 and 300m (Sobrino et al., 2005), although the depth at which this maximum is detected can be different even at short-spatial scales (Guijarro et al., 2009). It is more abundant in the eastern and central basins (Abelló et al., 2002; Sbrana et al., 2019). The current catches of this species, exclusively provided by bottom trawling, shown a clear increasing trend at Mediterranean level since 2005 (FAO, 2023). Juveniles and early life stages share a similar bathymetric distribution with adults, occurring within the same depth range and often showing continuous recruitment throughout the year, which results in the coexistence of multiple cohorts on the fishing grounds (Sobrino et al., 2005; Guijarro et al., 2009; Druon et al., 2015).
In the Balearic Islands, catches of European hake have shown interannual changes during the last decades, suggesting a possible influence of climatic factors on this species. This relationship was suggested in the past by Astudillo and Caddy (1988) and Oliver (1993), confirmed more recently by Massutí et al. (2008) and Hidalgo et al. (2009, 2011). In this area, macro and meso-scale climate regimes can influence the population dynamics of hake recruitment, which seems to be enhanced during colder-than-normal winters and and increase of productivity in the area, that may be favorable to hake. The influence of climate indices on the population dynamics of European hake has been also suggested in the north-eastern Iberian Peninsula, also related with an enhanced productivity (Hidalgo et al., 2011; Maynou, 2011).
As regards deep-water rose shrimp, several environmental factors have been suggested to be influencing its dynamics. Thus, its abundance enhances and its distribution expands to northern both in the western and central Mediterranean, with the increase of sea temperature and salinity (Colloca et al., 2014; Sbrana et al., 2019; Quattrocchi et al., 2020; González-Andrés et al., 2024; Mingote et al., 2024). A decrease of wind circulation seems to be linked to an increase of potential preys for this species, which may be also positive for its life cycle and abundance (Ligas et al., 2011).
3 Material and methods
The main milestones raised have been: (i) exploration of the temporal trends during the last two decades in the Recruitment (R) and Spawning Stock Biomass (SSB) of European hake and deep-water rose shrimp, coming from stock assessments of the two species in GSAs 5 and 6, developed within the framework of the FAO-GFCM; (ii) modelling the influence of environment and SSB on R, using an aprioristic set of environmental variables as potential predictors in both species; and (iii) projection of R, SSB and fishing catches by species and GSA along the 21st century, using concurrently the main explanatory drivers with an ensemble of Regional Climate Models, under two climatic scenarios adopted by the Intergovernmental Panel on Climate Change (IPCC) and five fishing management strategies in each climatic scenario (Figure 2).
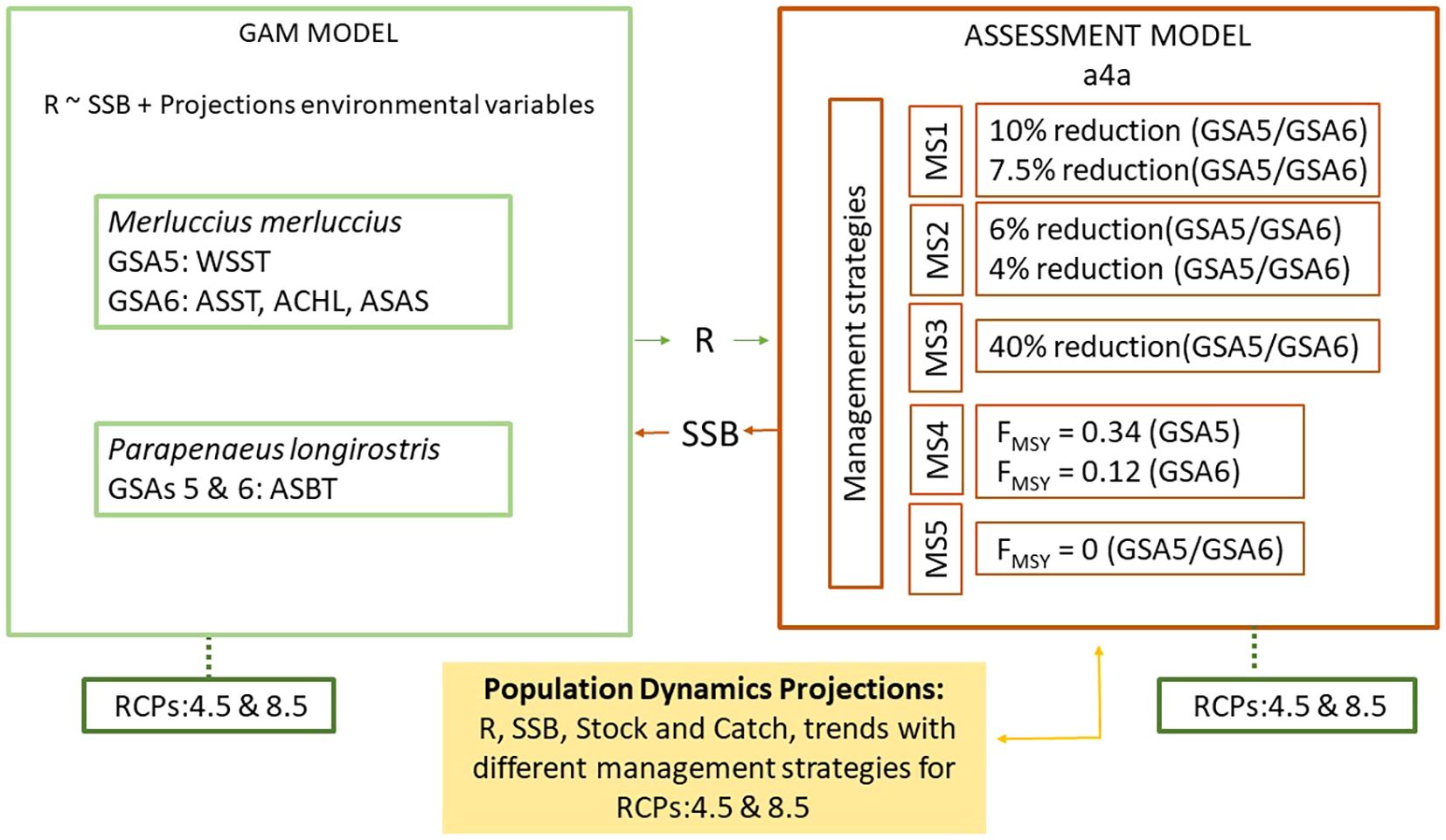
Figure 2. Schemes of the procedure applied for projecting recruitment (R) and spawning stock biomass (SSB) of European hake (Merluccius merluccius) and deep-water rose shrimp (Parapenaeus longirostris) along the 21st century in the FAO-GFCM geographical sub-areas GSA 5 (Balearic Islands) and GSA 6 (Northern Spain). WSST, Winter Sea Surface Temperature; ASST, Annual Sea Surface Temperature; ASAS, Annual Sea Average Salinity; ACHL, Annual Surface Chlorophyll; ASBT, Annual Sea Botton Temperature. MS1, MS2, MS3, MS4 and MS5 correspond to the fishing management strategies 1, 2, 3, 4 and 5, respectively. RCP, Regional Climate Models.
3.1 Biological data
Time series of R (age 0) and SSB for European hake and deep-water rose shrimp stocks in GSAs 5 and 6 were obtained from the GFCM Working Group on Stock Assessment of Demersal Species, available at https://www.fao.org/gfcm/data/star/en/. Times series were 1998–2020 in GSA5 and 2002–2020 in GSA 6 for European hake (Supplementary Figure S1) and 2001–2021 for deep-water rose shrimp in both GSAs (Supplementary Figure S2).
Stock assessments were carried out using the catch-at-age model a4a (Jardim et al., 2014), considering as input data: (i) age composition of catches, estimated from monthly length frequency sampling on board the fishing fleet and/or in port, depending on the year; (ii) official data on landings and discards; and (iii) biological parameters of the species. These parameters were: length-weight relationship and maturity ogive, estimated in each area from the analysis of biological samples of the two species coming from the European Union Data Collection Framework (https://dcf.ec.europa.eu/general-information_en), the von Bertalanffy growth parameters obtained from tagging experiments (Mellon-Duval et al., 2010) and the natural mortality (M) estimated using Prodbiom program (Abella et al., 1998). The assessment model was calibrated with age-structured abundance indices coming from the MEDITS bottom trawl surveys (Spedicato et al., 2019).
The a4a model uses three major components, called submodels: fishing mortality (F), survey catchability and recruitment (Jardim et al., 2014). They are essentially log-linear models, although the full model is still non-linear. The final model for provision on advice was selected during the GFCM Working Group on Stock Assessment of Demersal Species (Online, 17–21 January 2022), considering the best combination of submodels taking into account the fit of the models versus observed data, the model comparison (AIC, BIC, GCV) and the absence of patterns in the retrospective analysis. The model adjustment was carried out using the Fisheries Libraries in R (FLR). The results of the assessment showed an overexploitation for the four stocks, with a ratio between current fishing mortality and the reference point (F0.1) around 2 for the two stocks of deep-water rose shrimp (with relatively high biomass in both cases), 4 for the European hake in GSA 5 and 11 for the European hake in GSA 6, with relatively low biomass in both cases (FAO, 2022, 2024).
3.2 Environmental data
The initial selection of variables was based on available bibliographic information and include hydrological variables, such as water temperature and salinity, indicators of hydrodynamic conditions which had been previously identified as potential drivers of European hake recruitment in GSA 5 (Massutí et al., 2008; Hidalgo et al., 2011) and deep-water rose shrimp (Sbrana et al., 2006; Fortibuoni et al., 2010). Additionally, sea temperature and salinity are related to marine productivity, as they may affect nutrient availability, metabolic rates and water stratification, which act as potential drivers of many demersal species (Katsanevakis et al., 2009). Another factor related to productivity is chlorophyll, whose concentration is considered as a potential index of primary production in the ecosystem and markers of the trophic state of the water column (Richardson and Schoeman, 2004; Piroddi et al., 2017). Several studies have shown that primary production can be an important factor driving the recruitment of European hake (Bartolino et al., 2008; Druon et al., 2015; Coll et al., 2016) and deep-water rose shrimp (Guijarro et al., 2009).
Data were mostly extracted from Copernicus Marine Environment Monitoring Services (CMEMS; Table 1), which use remote sensing observations and include reanalysis products specifics for the Mediterranean. These variables were extracted restricting the habitat of European hake recruits and deep-water rose shrimp (150–400 and 50–1000 m depth, respectively; Oliver and Massutí, 1995; Sobrino et al., 2005; Druon et al., 2015; Sion et al., 2019). For deep-water rose shrimp, the larger bathymetric range was selected due to potential differences in the distribution of the species among even nearby areas (Guijarro et al., 2009) and the known spatial changes of this species in the area, which has spread northwards in the two last decades (González-Andrés et al., 2024; Mingote et al., 2024), and whose depth distribution range increases with temperature (Quattrocchi et al., 2020). Subsequently, the variables were spatially averaged over each GSA using shapefiles provided by the GFCM (https://www.fao.org/gfcm/data/maps/gsas/es/). Monthly time series were averaged with annual and winter basis (JFM) for both species and summer averages (JJA) for European hake, since the recruitment of this last species mainly occurs during this season. The annual environmental variables were included because, in the study area, recruitment of deep-water rose shrimp occurs year-round without marked seasonality (Guijarro et al., 2009). Winter season was also considered because of particular oceanographic circulation in this sub-basin, due to the intensified mixing processes and changes in water mass distribution, driving nutrient availability and primary productivity (Estrada, 1996; Monserrat et al., 2008).
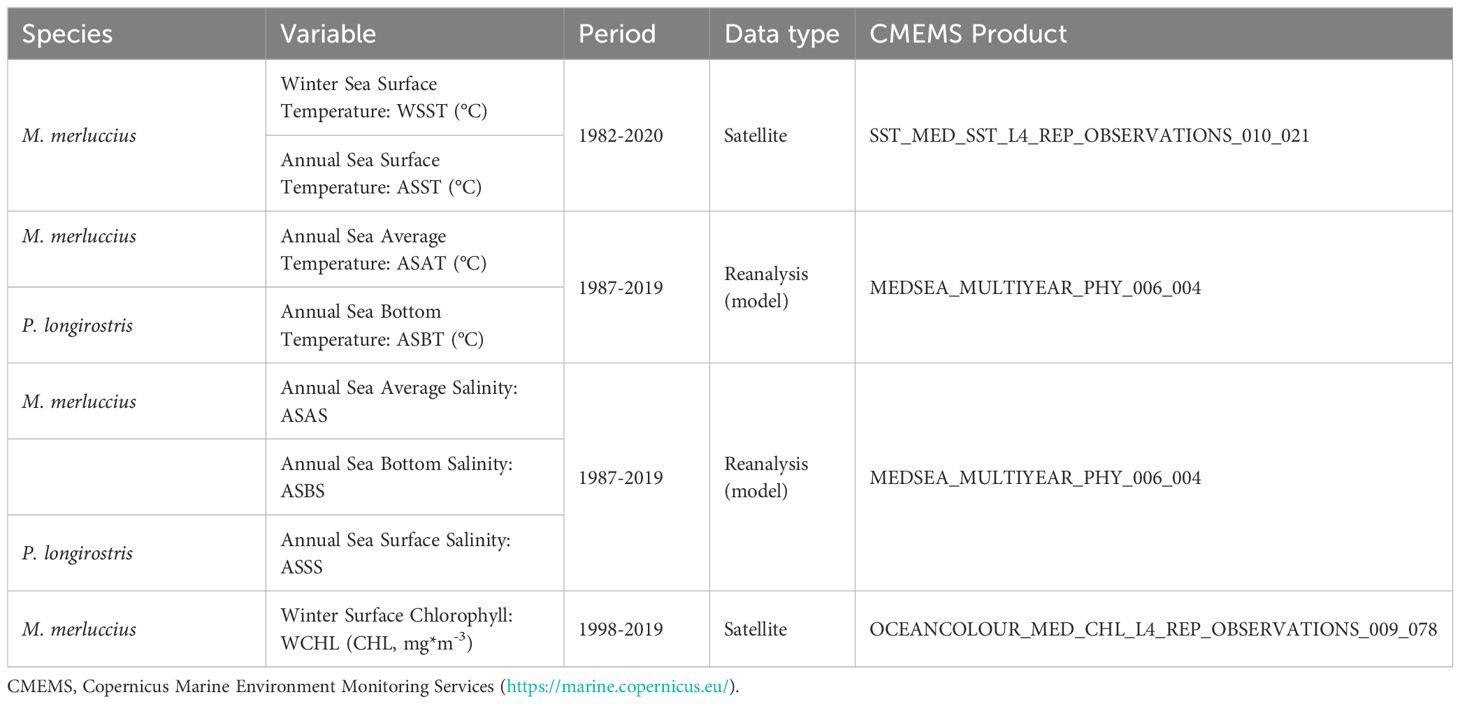
Table 1. Summary of the environmental variables considered as potential predictors to develop population dynamics models of European hake (M. merluccius) and deep-water rose shrimp (P. longirostris) in the western Mediterranean.
3.3 Climate change projections
To describe the potential future conditions in the western Mediterranean, a set of simulations (ensemble) were collected ad hoc for this study from four downscaled Regional Climate Models (RCM), under the frame of Coupled Model Intercomparison Project Phase 5 (CMIP5) to assess the certainty and plausibility of the projections. The methodology applied is well established in regional climate modeling studies. The use of downscaled CMIP5 RCMs, bias correction via the delta method and ensemble modeling to account for uncertainty are widely accepted approaches (van Vuuren et al., 2011; Zappa et al., 2013). RCM provide high-resolution simulations that allow capturing the particular and complex oceanographic characteristics, such as the formation of deep waters, the presence of fronts or oceanic eddies that occur in this sub-basin. The main characteristics of these simulations have been summarized and compiled in Supplementary Table S1. The methodology used to extract the variables projected into the future was similar to the extraction of the present variables, with the aforementioned bathymetric restrictions for the habitat of recruits for European hake and deep-water rose shrimp.
We used projections for the greenhouse gas concentration scenarios adopted by the Intergovernmental Panel on Climate Change (IPCC) of the two Representative Concentration Pathways: RCP 4.5 (average warming) and RCP 8.5 (extreme warming), based on moderate and high concentrations of greenhouse gases, which by the end of the 21st century increase radiative forcing on Earth by 4.5 and 8.5 Wm-2, respectively.
The RCM ensemble simulations (Supplementary Figure S14) were bias-corrected using a delta correction method. Specifically, the mean deviation between simulated and observed values during the overlapping historical period (2006–2020) was calculated and subtracted from future projections to align them with present-day conditions. This method preserves the projected climate change signal while correcting systematic biases in model outputs. To compare the simulations with the current values of the variables (Table 1), different sources have been used. When available, satellite observations were extracted and, otherwise, information was obtained from reanalysis-type models with data assimilation. Although confidence intervals were not graphically displayed in the projection figures due to the high number of scenarios and the need to maintain figure readability, model uncertainty was explicitly addressed by incorporating multiple Regional Climate Models (RCMs) and contrasting fishing management strategies. This approach allowed us to explore a plausible range of future outcomes but are not discussed in this article to simplify the interpretation of the results.
3.4 Fishing management strategies
Although some of the fishing management strategies considered (e.g. MS2 and MS3) reflect actual management measures implemented under the EU-MAP since 2020, others represent plausible alternative scenarios (e.g. MS4 and MS5) used to explore future stock responses under different fishing pressure levels. Within each climate scenario, different fishing management strategies were considered, taking into account both the scientific advice on the state of exploitation of European hake stocks in GSAs 5 and 6 and the EU-MAP currently in force (Regulation 2019/1022 of the European Parliament and of the Council of 20 June 2019) that establishes a multiannual plan for the fisheries exploiting demersal stocks in the western Mediterranean. This regulation considers some target species, including European hake, which shows the highest levels of overfishing in the Mediterranean (FAO, 2022) and deep-water rose shrimp, which has shown an important increasing trend in the western Mediterranean during the last years. Following Article 7 of the EU-MAP, a fishing effort regime to all trawling vessels was applied since 1 January 2020, with an effort reduction of 10, 7.5 and 6% of the fishing effort (expressed as fishing days) in 2020, 2021 and 2022, respectively, considering as baseline the average value of the period 2015-2017. According to the EU-MAP, the maximum allowable fishing effort shall be reduced by a maximum of 40% during its implementation period.
According to this, the following five fishing management strategies were considered: MS1) fishing mortality (F) in the status quo (considering the status quo as the average of the last three years of the time series; 2019-2021); MS2) the F reduction considered in EU-MAP of 10, 7.5, 6 and 4% in 2020, 2021, 2022 and 2023, respectively; MS3) the maximum reduction of 40% F in 2024, according to the EU-MAP; MS4) fishing mortality reaching values of the target reference point F0.1 in 2025, which corresponds to 0.34 for GSA 5 and 0.12 for GSA 6 in the case of European hake and to 0.57 for GSA 5 and 0.78 for GSA 6 in the case of deep-water rose shrimp, according to the stock assessment results (FAO, 2022); and MS5) strategies without fishing activity (F=0), through gradually reducing the F value. This gradual reduction was modelled based on the current decreasing trend of bottom trawlers in both GSAs, adjusting a linear model in its number for each GSA, which predicted that the bottom trawl fleet will disappear in 2039 and 2061 in GSA 5 and GSA 6, respectively (Supplementary Figure S3). In order to limit the total biomass in the model results, twice the maximum SSB found in the series studied was considered as the carrying capacity of the system. This decision aimed to prevent biologically unrealistic exponential growth projections, as historical biomass increases likely reflect recovery from reduced fishing pressure. A linear regression was used to project the disappearance of the fleet (in 2039 for GSA 5 and 2061 for GSA 6), allowing us to assess the potential effects of a gradual reduction in fishing impact under extreme conservation conditions. The approach, supported by demersal species modelling (Colloca et al., 2017), adopts a conservative cap aligned with Mediterranean oligotrophic ecosystem carrying capacity. The projections were made using the free software R. Although some of the fishing strategies considered (MS1- MS3) reflect actual management measures implemented under the EU Multiannual Plan since 2020, others (MS4 and MS5) represent plausible alternative scenarios used to explore future stock responses under different fishing pressure levels.
3.5 Statistical analysis
The selection of the environmental variables was carried out in two stages: i) Pearson’s correlations of recruitment series were carried out in each GSA for European hake and deep-water rose shrimp, with a total of 20 and 12 environmental variables, respectively (Supplementary Table S2). More information about this process is included in Supplementary Information; ii) the predictor variables selected in the first stage were filtered based on two metrics to avoid overfitting and reduce correlation and collinearity between variables. First, all predictor covariates were examined using Pearson’s correlation (Wood, 2006) and pairs of variables with high correlation values (r>0.7) were identified and only one of the correlated pairs was included in the modelling process (Lezama-Ochoa et al., 2017). Subsequently, multicollinearity between predictor variables was tested by calculating the variance inflation factor (VIF) with a cutoff value of 5 and using the corvif function from the AED package in R (Zuur et al., 2009). As a result, highly correlated and multicollinearity variables were tested separately during the modelling, and only the one with the highest total explained variance was included in the final models.
To study the effect of environmental variability on European hake and deep-water rose shrimp recruitment, Generalized Additive Models (GAM; Hastie and Tibshirani, 1990; Wood, 2006) were adjusted for each GSA, considering the SSB of the previous year (SSBi-1) and the environmental variables of the corresponding year. The models were fitted through a Gamma distribution with logarithmic link function. The GAM models were fitted in the R software (Wood, 2017) using the mgcv package (Wood, 2004), where the degrees of freedom of the smoothing functions were limited to 3 (K=3) for the splines of the different explanatory variables. Cubic regression splines (Wood, 2006) were used for all explanatory variables.
To establish the best models for each stock, a forward stepwise variable selection procedure was applied. In each case, the best model was selected based on Akaike Information Criterion (AIC), the unbiased risk estimator (UBRE; Burnham and Anderson, 2004), the significant explanatory variables and the highest explained deviance. To evaluate the predictive model for R, the Pearson correlation coefficient was calculated between the values predicted by the model and the observed values. In addition, diagnostic plots including those derived from Generalized Cross Validation (GCV) using the mgcv package are provided as Supplementary Material. These plots offer further insight into model behavior and contribute to transparency in the fitting process. To ensure a robust and ecologically interpretable model selection, we did not rely exclusively on minimizing the Akaike Information Criterion (AIC). Instead, model choice was based on a combination of multiple metrics, including explained deviance, statistical significance of predictor variables, the Pearson correlation coefficient (r) between observed and predicted values, and inspection of residuals to assess model adequacy. This approach aims to balance predictive accuracy with parsimony and biological interpretability. For instance, in GSA 5 for European hake, the selected model explained 52% of the deviance and had a moderate predictive fit (r= 0.5), although it did not yield the lowest AIC. Its selection was supported by its simplicity, significant predictors, and ecological coherence. In GSA 6, a model with a slightly higher AIC but fewer variables (four instead of five) and 79% deviance explained was preferred over a more complex alternative, aligning with the principle of parsimony. This multi-criteria selection strategy helps to avoid overfitting and improves the ecological relevance of projections, particularly when environmental predictors are intercorrelated or show strong interannual variability.
To implement the population dynamics forecasting model, we used the FLR (Fisheries Library in R) framework, integrating the empirical relationships obtained between spawning stock biomass (SSB), recruitment (R), and the selected environmental variables through Generalized Additive Models (GAMs). For each year of the projection period (2021–2100), recruitment was estimated based on the SSB from the previous year and the environmental conditions projected by Regional Climate Models (RCMs) under the RCP 4.5 and RCP 8.5 scenarios. From the projected recruitment, the SSB for the following year was calculated using constant biological parameters —growth, natural mortality and maturity— averaged over the period 2019–2021 (Figure 2).
The structural model implemented in FLR follows a production model logic with recruitment dependence, incorporating selectivity and catchability functions derived from the stock assessments conducted using the a4a model developed by the GFCM. Fishing mortality rates (F rates) were simulated under five different management strategies, ranging from status quo to progressive reductions and complete fishing cessation (F=0). In all scenarios, a maximum carrying capacity was assumed to constrain population growth in the absence of fishing, defined as twice the highest SSB observed in the historical time series.
4 Results
4.1 Trends in population dynamics and effect of environmental variables
In the period 2003-2020, the R and SSB data series of European hake showed a pronounced decrease in GSA 6, while this decreasing trend was smaller in GSA 5. Despite these trends, both R and SSB values were much higher in GSA 6 than in GSA 5 (Supplementary Figure S1). On the contrary, R and SSB of deep-water rose shrimp showed a continuous increase since 2015, reaching higher values in GSA 6 (Supplementary Figure S2). These contrasting trends between species and GSAs highlight the importance of region-specific dynamics, which may be influenced by differing environmental and fishing conditions to be explored in the discussion.
In the first stage, Pearson correlations between recruitment series for European hake and deep-water rose shrimp in each GSA and environmental variables (20 in GSA 5 and 12 in GSA 6) indicated 5 and 9 of them, respectively as potential predictors for European hake recruitment, while 6 environmental variables determined the deep-water rose shrimp recruitment in both GSAs (Table 1, Supplementary Table S2):
- European hake in GSA 5: Winter Surface Chlorophyll (WCHL) and Annual Surface Chlorophyll (ACHL), both in mg*m-3; Winter Sea Surface Temperature (WSST), Annual Sea Surface Temperature (ASST) and Annual Sea Average Temperature (ASAT), all of them in °C.
- European hake in GSA 6: WCHL, ACHL, WSST, ASST, ASAT and Annual Sea Bottom Temperature (ASBT) in °C; and Annual Sea Surface Salinity (ASSS), Annual Sea Average Salinity (ASAS) and Annual Sea Bottom Salinity (ASBS).
- Deep-water rose shrimp in both GSAs: ACHL, ASAT, ASBT, ASAS, ASBS and ASST.
In the second stage, the environmental variables selected to model European hake R differed between GSAs. According to the cross-correlation matrix of the 5 potential predictor variables in the GSA 5, WSST was highly correlated (Pearson correlation; r>0.7) with ACLH, while WSST was highly correlated with ASAT. Most of the variance inflation factor (VIF) values were between 1.4 and 4.9, which was considered as an indication of the absence of multicollinearity between the explanatory variables. The variable showing the presence of multicollinearity was ASAT, with a value of 6.68 (Supplementary Figure S4). As a result, highly correlated variables with multicollinearity were tested separately in the GAMs and only those with the greatest total variance explained were included in the final models. In GSA 5, the WSST was selected as the main environmental driver of European hake R, with an explained deviation of 52%, an AIC value of 8633 and a correlation between observed and predicted of 0.5 (Supplementary Table S3). The smooth functions of the final model indicated that the probability of R was lower as WSST and SSB increase (Figure 3a). This suggests that temperature-related factors may play a predominant role in recruitment variability in GSA 5, in contrast with GSA 6, where other variables are more relevant.
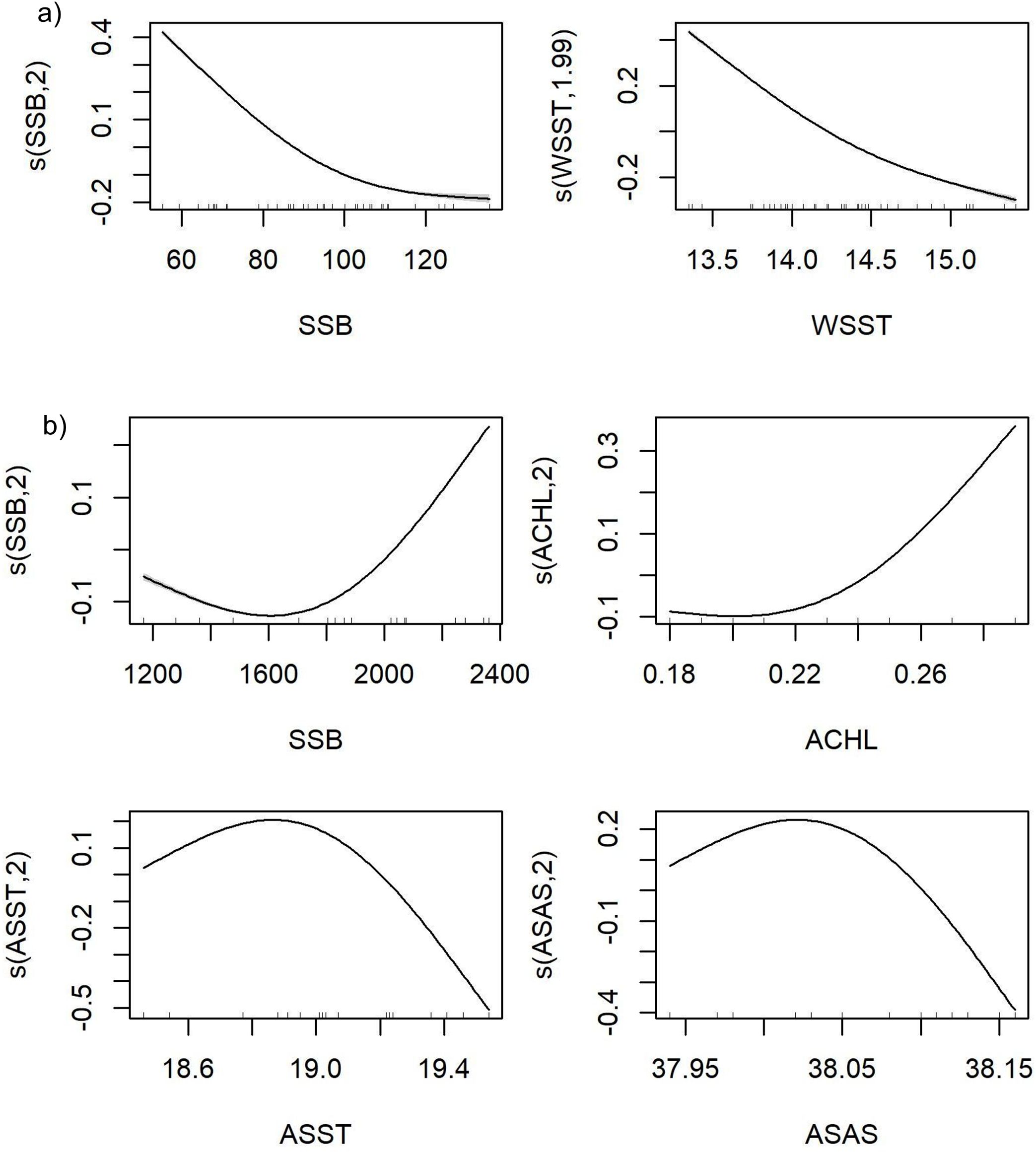
Figure 3. GAM partial plots for the best recruitment models of European hake (Merluccius merluccius) in the FAO-GFCM geographical sub-areas: (a) GSA 5 (Balearic Islands); and (b) GSA 6 (Northern Spain). The Y axis (logit scale) represents the recruitment probability and the X axis represents the range of values of the predictor variables SSB, Spawning Stock Biomass; WSST, Winter Sea Surface Temperature; SSB, Spawning Stock Biomass; ACHL, Annual Surface Chlorophyll; ASST, Annual Sea Surface Temperature; ASAS, Annual Sea Average Salinity.
Among the 9 explanatory variables included in the GSA 6 models of European hake R, the most highly correlated were the following (Supplementary Figure S5): SBB with ASBT; WCHL with ACHL; WSST and WSST with WCHL, ACHL and ASAT; ASBT with SSB ASAT and ASBS; and ASSS with ASAS. The variables without the presence of multicollinearity were SSB, WCHL, ACHL, ASST, ASBT and ASAS, with values of 3.7, 4.6, 5.0, 4.9, 5.0 and 4.50, respectively. Finally, ASST, ACHL and ASAS were selected as the main environmental drivers of European hake R in the GSA 6, with an explained deviation of 79%, an AIC value of 88489 and a correlation between observed and predicted of 0.9 (Supplementary Table S3). The smooth functions of the response variables for the final model indicated that there was greater R with ASST of 18.9-19°C and that R decreased when ASAS and ASST increased. Finally, for SSB and ACHL there was a positive linear relationship with R (Figure 3b). These results point to a broader suite of environmental drivers influencing European hake R in GSA 6, particularly involving productivity and salinity.
For deep-water rose shrimp, among the 7 explanatory variables included in GSAs 5 and 6 models, the highest correlation in GSA 5 were between SSB, ASAT and ASBT (Supplementary Figure S6). In GSA6, the variables with higher correlation were ASAT and ASBT (Supplementary Figure S7). Regarding collinearity, ASAT presents a value >5 in both GSAs: 8.82 and 12.22, respectively (Supplementary Figures S6, S7).
ASBT was selected as the main environmental driver for European hake and deep-water rose shrimp R in GSAs 5 and 6, with respectively an explained deviation of 82.62 and 95.1%, an AIC value of 371.13 and 450.04, and a correlation between observed and predicted of 0.87 and 0.83 (Supplementary Table S4). The smooth functions of the final model indicated that probability of R was greater as SSB and ASBT increased (Figure 4). The consistent importance of ASBT across GSAs for deep-water rose shrimp, compared to the more variable drivers in European hake, indicates a potentially stronger link between benthic thermal regimes and the life cycle of this species. The diagnostic plots derived from GCV, suggested that the selected models are generally acceptable, showing reasonable fit and residual patterns for the recruitment models of both species across GSAs (Supplementary Figures S8-S11).
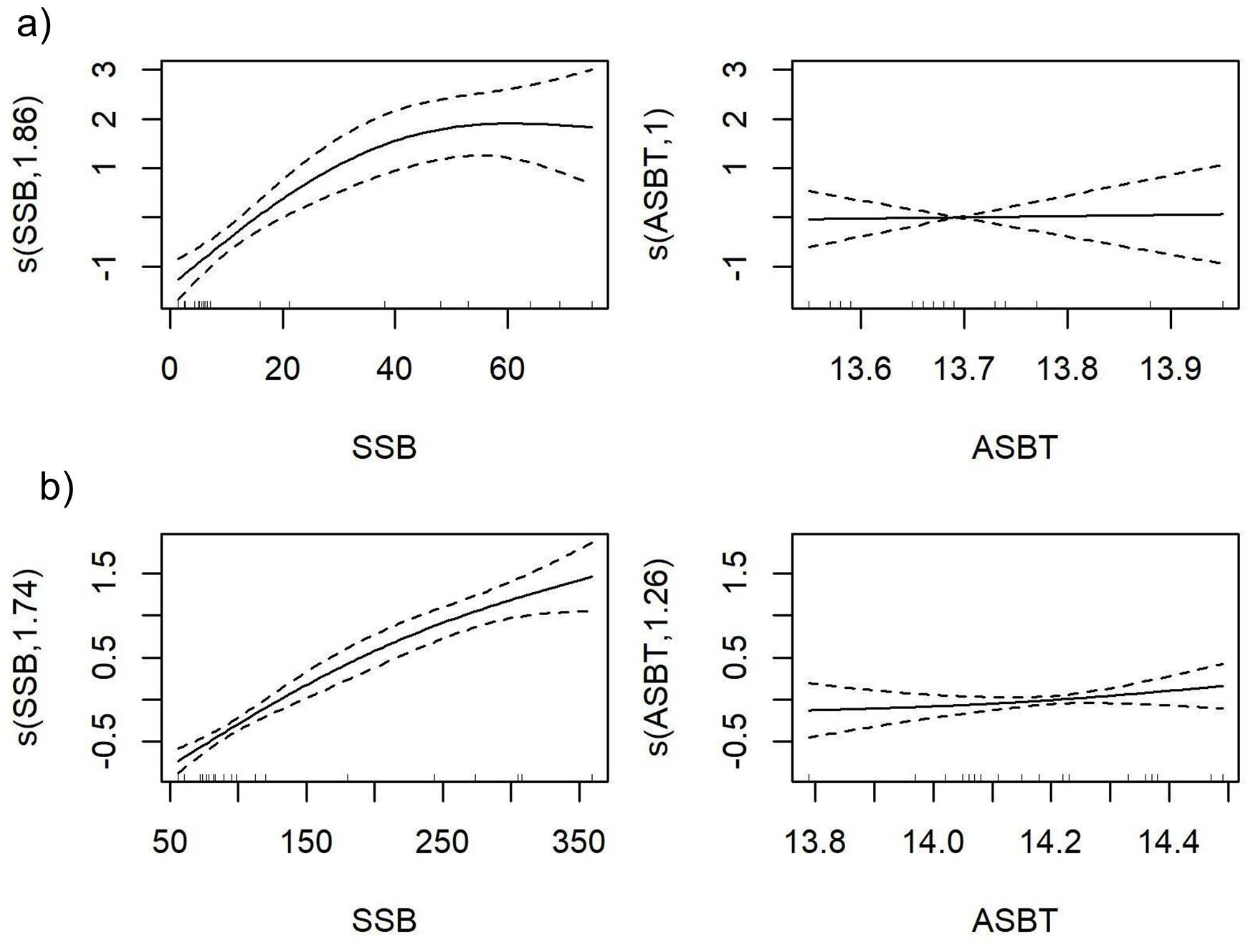
Figure 4. GAM partial plots for the best recruitment models of the deep-water rose shrimp (Parapenaeus longirostris) in the FAO-GFCM geographical sub-areas: (a) GSA 5 (Balearic Islands); and (b) GSA 6 (Northern Spain). The Y axis (logit scale) represents the recruitment probability and the X axis represents the range of values of the predictor variables SSB, Spawning Stock Biomass; ASBT, Annual Sea Bottom Temperature.
4.2 Projections
4.2.1 Environmental variables
The results of the projections along the 21st century of the selected environmental variables for European hake and deep-water rose shrimp in GSAs 5 and 6 are shown in Supplementary Figures S12-S14. In GSA 5, the WSST will increase up to 1.5 or 3°C in 2100, depending on the emission scenario (Supplementary Figure S12). In GSA 6, the ASST is expected to reach up to 21.5 or 23°C at the end of the century, depending on the RCP (Supplementary Figure S13).
In the GSA 6, the models also predicted a slight increase in mean ACHL levels and a slight decrease in ASAS, more evident for RCP 8.5 and from 2050 onwards (Supplementary Figure S13). These two variables presented a greater dispersion in the predictions compared to ASST. For ASBT the model predicted an increase of 1 or 2°C and 1.5 or 3°C in 2100, respectively, depending on the emission scenario (RCP4.5 and 8.5). In general, the RCP4.5 scenarios presented a greater dispersion in the predictions compared to RCP 8.5 (Supplementary Figure S14).
4.2.2 Population dynamics of European hake and deep-water rose shrimp and their catches
Although some common patterns in the population dynamics of European hake were observed in both GSAs under fishing management strategies (MSs) 1, 2 and 3 and climatic scenarios RCPs 4.5 and 8.5 (Figures 5–8), no consistent patterns were observed in R and SSB between GSAs under the more extreme MSs 4 and 5. In general, MSs 1–3 showed decreasing trends in R, SSB and catches along the whole time period under both RCPa 4.5 and 8.5, although greater fluctuations were observed in GSA 6 under RCP 8.5 compared to RCP 4.5.
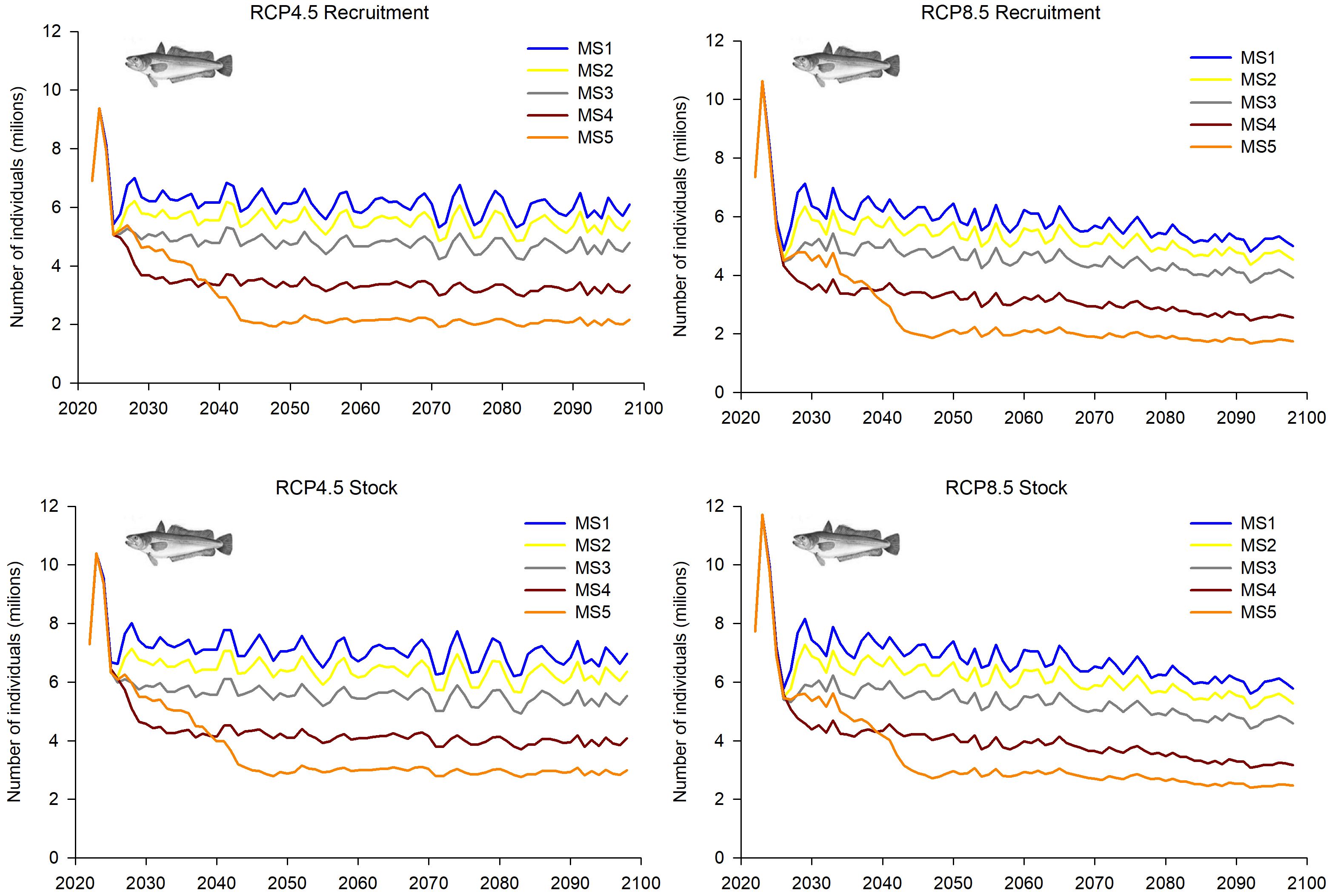
Figure 5. Predictions and projections along the 21st century of the recruitment and the overall stock of European hake (Merluccius merluccius) in the FAO-GFCM geographical sub-area GSA 5 (Balearic Islands), under the five fishing management strategies considered (see Material and methods and Figure 2 for more details) and the Regional Climatic Models under two climatic scenarios (RCP 4.5 and RCP 8.5).
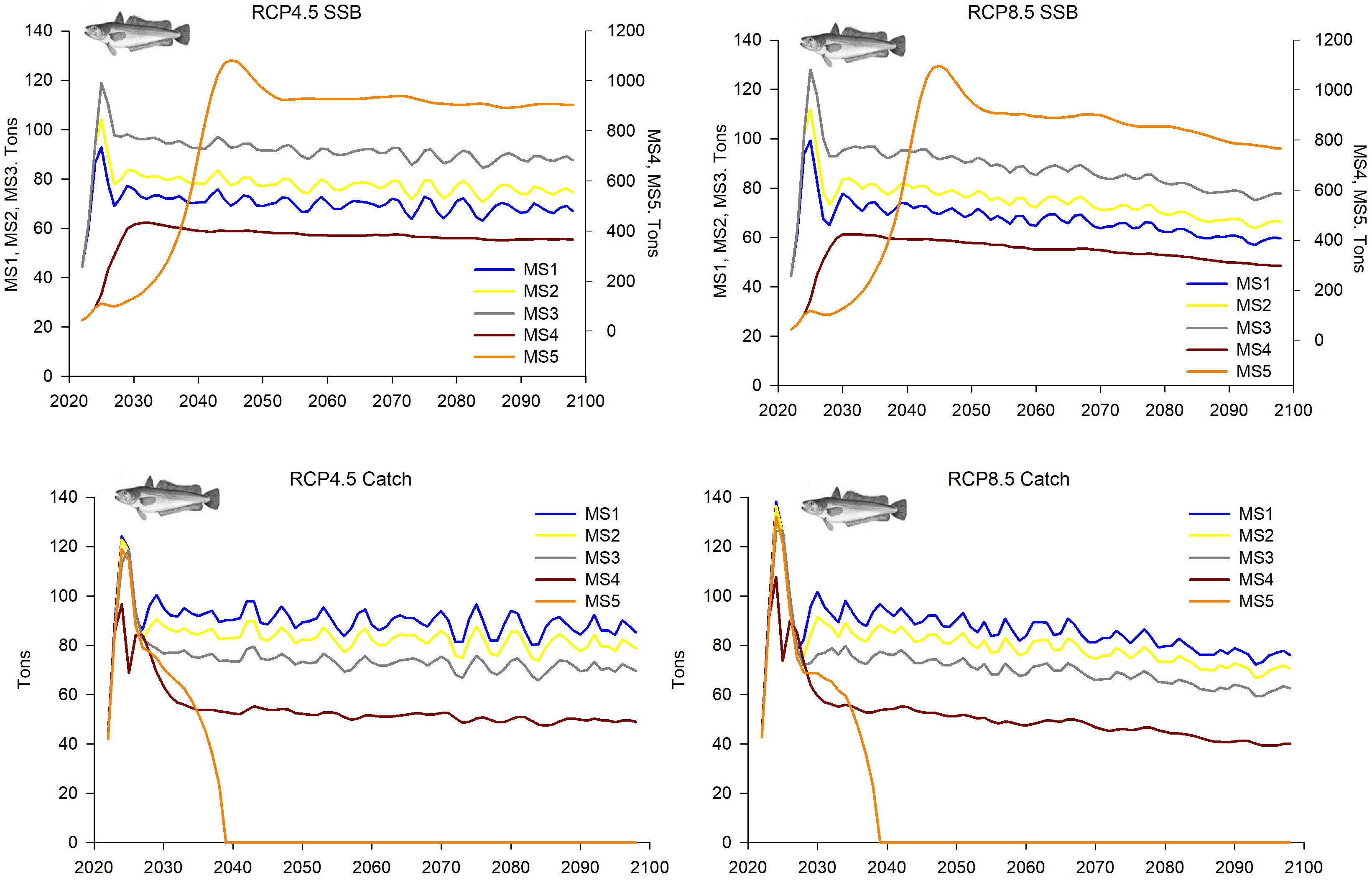
Figure 6. Predictions and projections along the 21st century of the SSB and catches of European hake (Merluccius merluccius) in the FAO-GFCM geographical sub-area GSA 5 (Balearic Islands), under the five management strategies considered (see Material and methods and Figure 2 for more details) and the Regional Climatic Models under two climatic scenarios (RCP 4.5 and RCP 8.5).
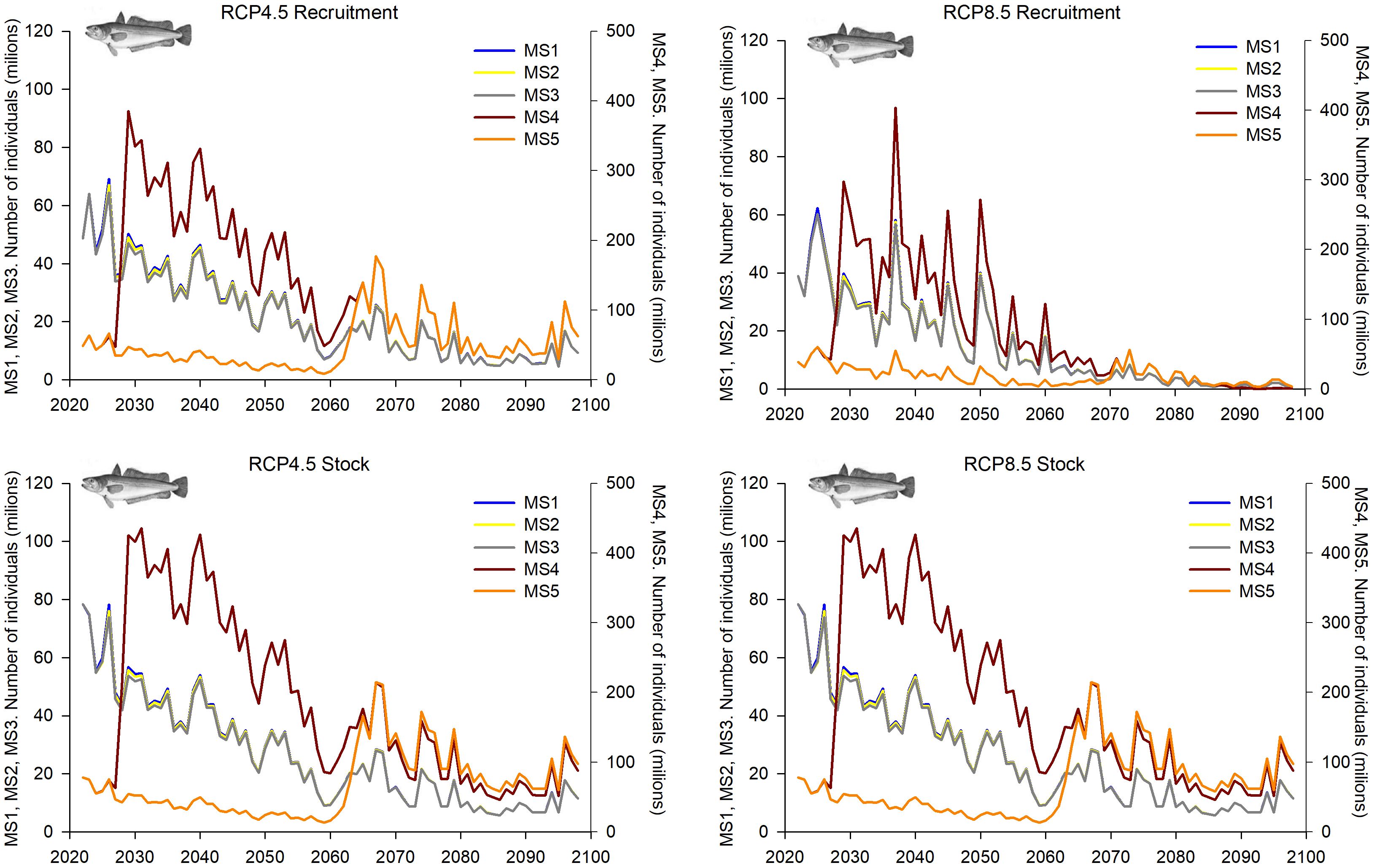
Figure 7. Predictions and projections along the 21st century of the recruitment and overall stock of European hake (Merluccius merluccius) in the FAO-GFCM geographical sub-area GSA 6 (Northern Spain), under the five fishing management strategies considered (see Material and methods and Figure 2 for more details) and the Regional Climatic Models under two climatic scenarios (RCP 4.5 and RCP 8.5).
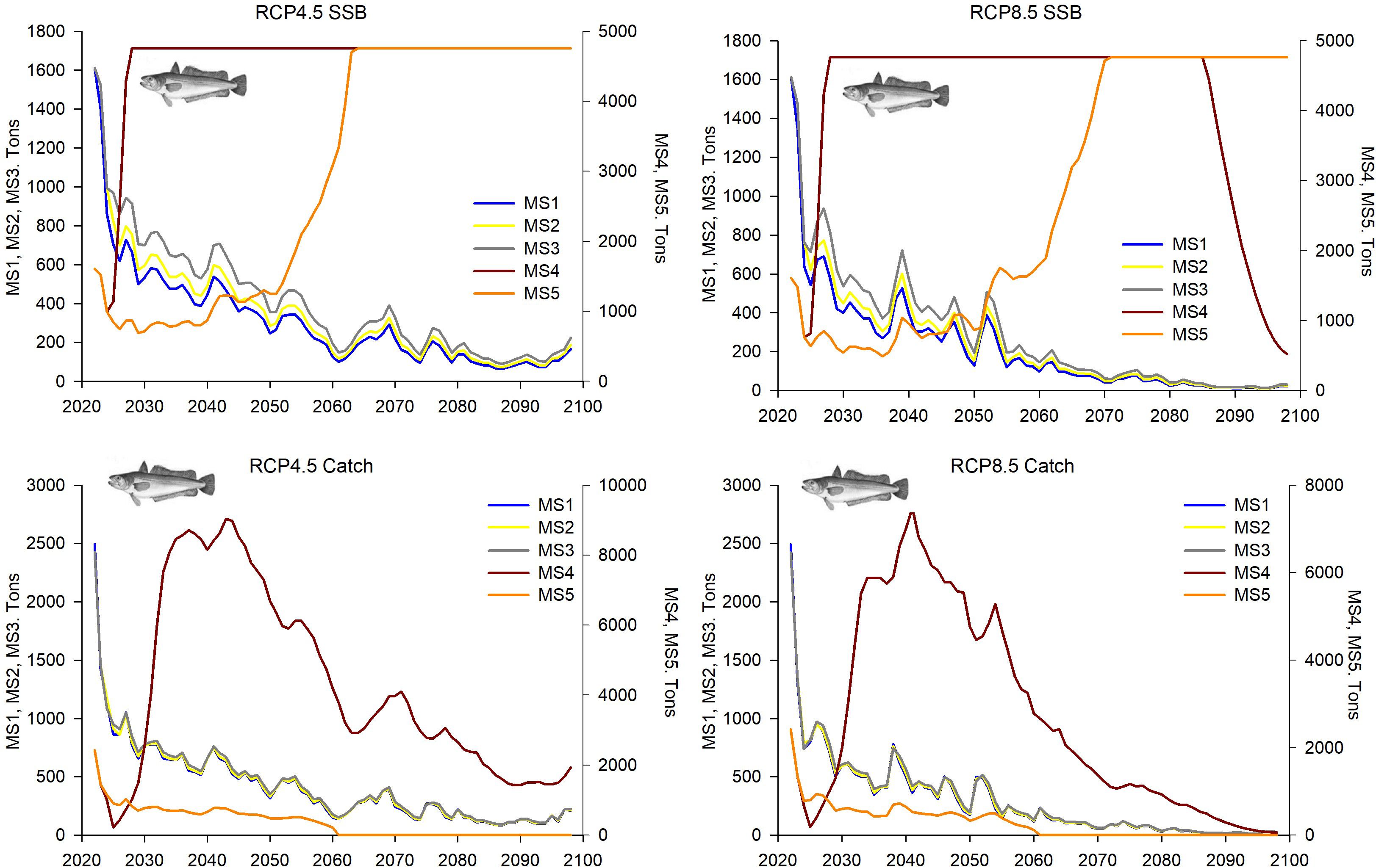
Figure 8. Predictions and projections along the 21st century of the SSB and catches of European hake (Merluccius merluccius) in the FAO-GFCM geographical sub-area GSA 6 (Northern Spain), under the five fishing management strategies considered (see Material and methods and Figure 2 for more details) and the Regional Climatic Models under two climatic scenarios (RCP 4.5 and RCP 8.5).
In GSA 5, the trends in R and the overall stock under MSs 1–3 were similar, showing relative stability or a slight decrease throughout the projection period. This decline was more pronounced under the climatic scenario RCP 8.5 compared to RCP 4.5 (Figure 5).
This decrease was more evident in MSs 4 and 5, especially during the first projected decade. By contrast, the SSB showed an increase during the first years or one-two projected decades under the fishing management scenarios MSs 1–3 and 4-5, respectively, with certain stability or slight decrease afterwards (Figure 6). Trends of catches along the 21th century were similar to those observed in R, except for MS 5 (F=0), in which there are no catches (Figure 6).
The population dynamics of European hake in GSA 6 showed absolute values of parameters much higher than those of GSA 5 and also quite different trends (Figures 7, 8). Both R and the overall stock decreased along the whole projected period within MSs 1-3, while increments of both population parameters were projected within MSs 4 and 5 (Figures 7, 8). However, this increment was at the beginning of the time series within MS 4, followed by a slight decrease, while within MS 5 the increment was during the 2060s. In all cases, greater fluctuations in the values were estimated in the climatic scenario RCP 8.5 than in RCP 4.5. The SSB also showed a continuous decrease along the whole projected period under SCMSs 1–3 and a progressive increase after the middle of 2030s within MS 5, while the MS 4 showed abrupt increment and decrease at the beginning and the end of the time series, respectively (Figure 8). Catches showed similar trends, with a decrease along the whole projected period under MSs 1-3 (Figure 8) and zero catches for MS 5, since the moment F is set to zero (beginning of 2060s) (Supplementary Figure S3). Within MS 4, values increased at the beginning of the time series, followed by a continuous decrease, which in that fishing management strategy was more pronounced than in the others. In MS 4, the differences in the fluctuations of values between the climatic scenarios RCP 4.5 and 4.8 were not as large as those observed in the other fishing management strategies.
For the deep-water rose shrimp, some common increasing patterns in the population dynamics were observed in both GSAs within all MSs considered and both climatic scenarios (Figures 9–12). In GSA 5, MSs 1–3 showed increasing trends in R, overall stock, SSB and catches along the whole time period under both climatic scenarios, which was more pronounced in RCP 8.5 than in RCP 4.5 (Figures 9, 10). This growth becomes more notable in MSs 4 and 5 for the SSB, while MS 5 showed greater growth in the second decade. For R, overall stock and catches, these scenarios show lower values than MSs 1-3, although the trend, like the rest of fishing management strategies, showed a gradual increase throughout the time series (Figures 9, 10).
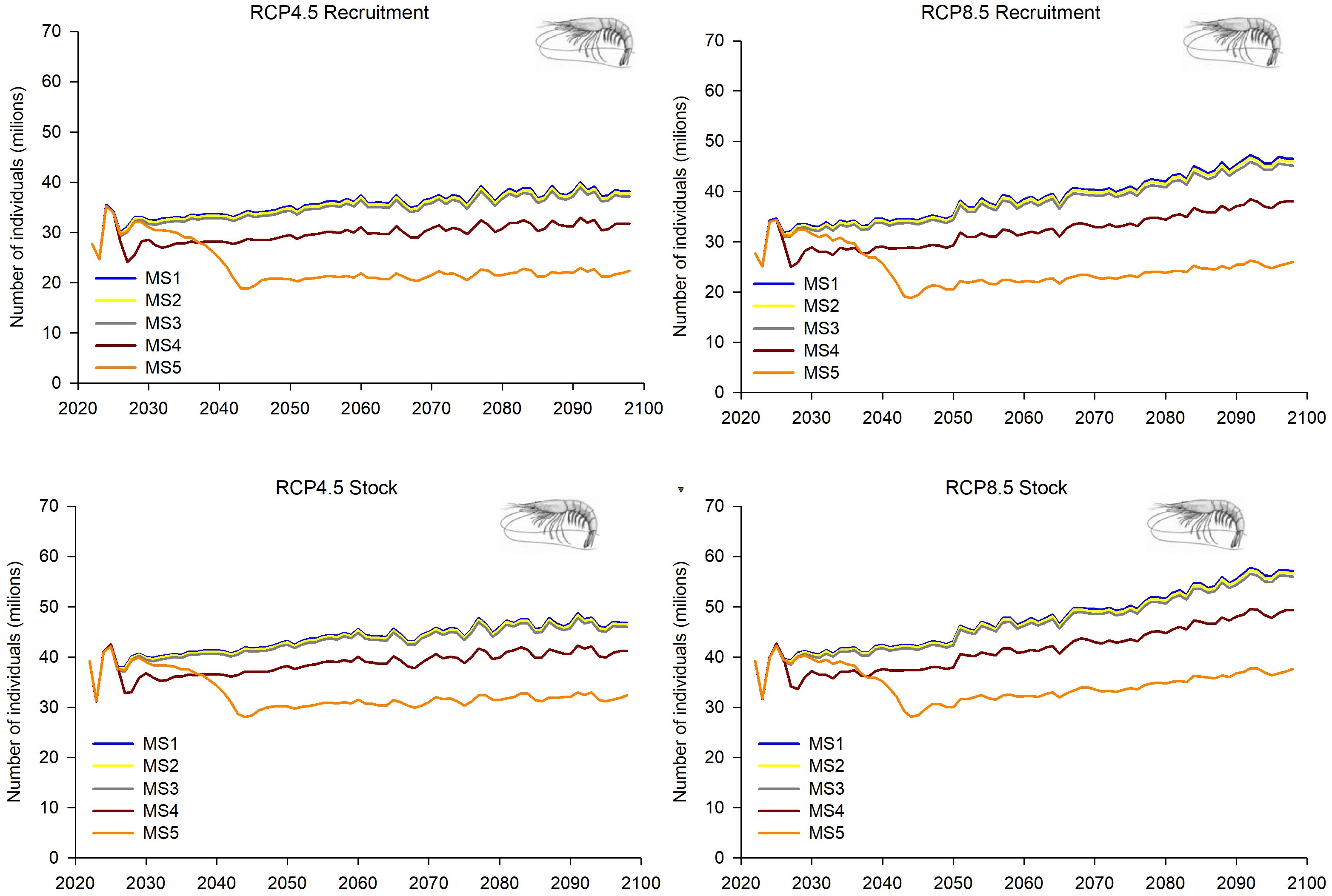
Figure 9. Predictions and projections along the 21st century of the recruitment and the overall stock of deep-water rose shrimp (Parapenaeus longirostris) in the FAO-GFCM sub-area GSA 5 (Balearic Islands), under the five fishing management strategies considered (see Material and methods and Figure 2 for more details) and the Regional Climatic Models under two climatic scenarios (RCP 4.5 and RCP 8.5).
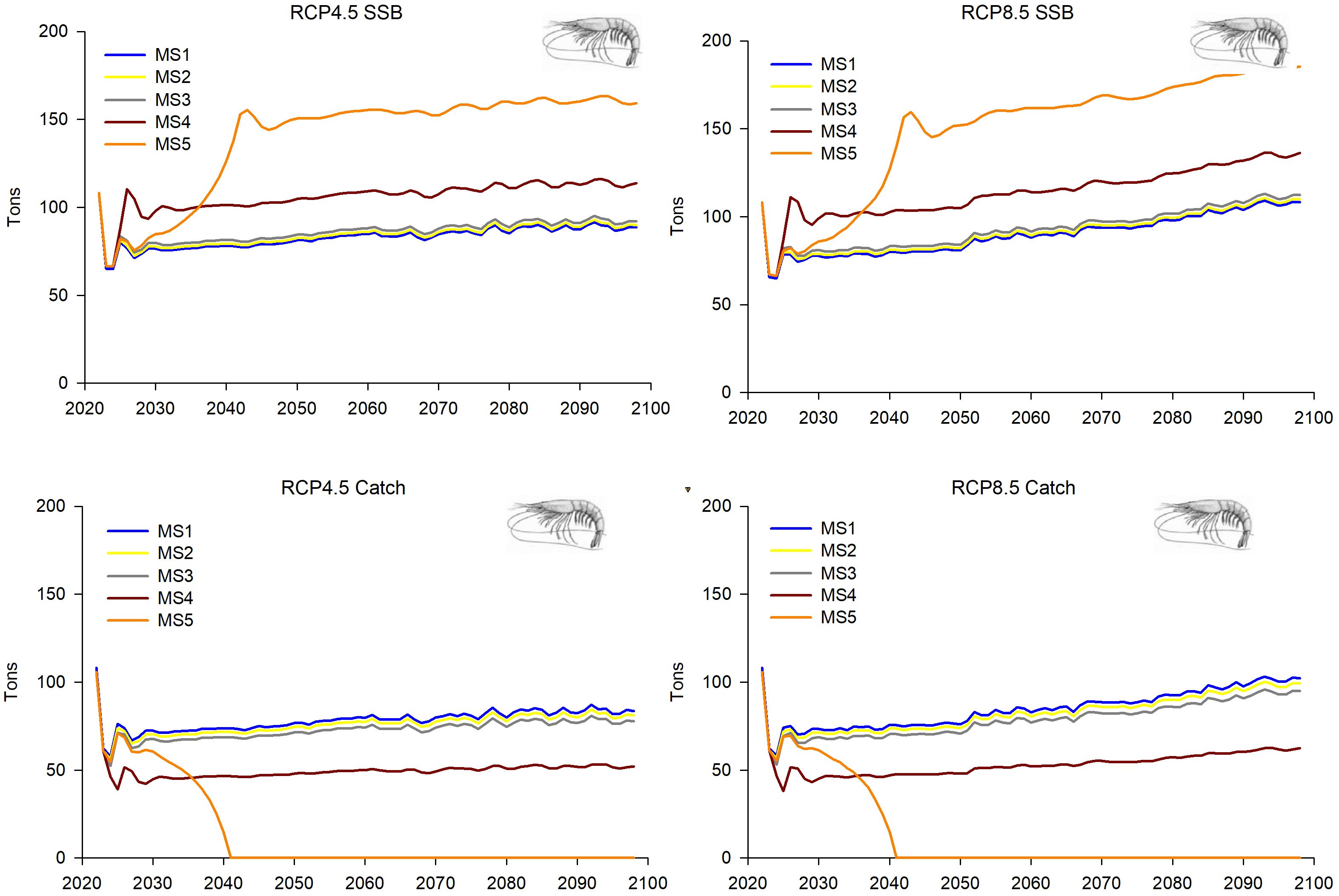
Figure 10. Predictions and projections along the 21st century of the SSB and the catches of the deep-water rose shrimp (Parapenaeus longirostris) in the FAO-GFCM sub-area GSA 5 (Balearic Islands), under the five fishing management strategies considered (see Material and methods and Figure 2) and the Regional Climatic Models under two climatic scenarios (RCP 4.5 and RCP 8.5).
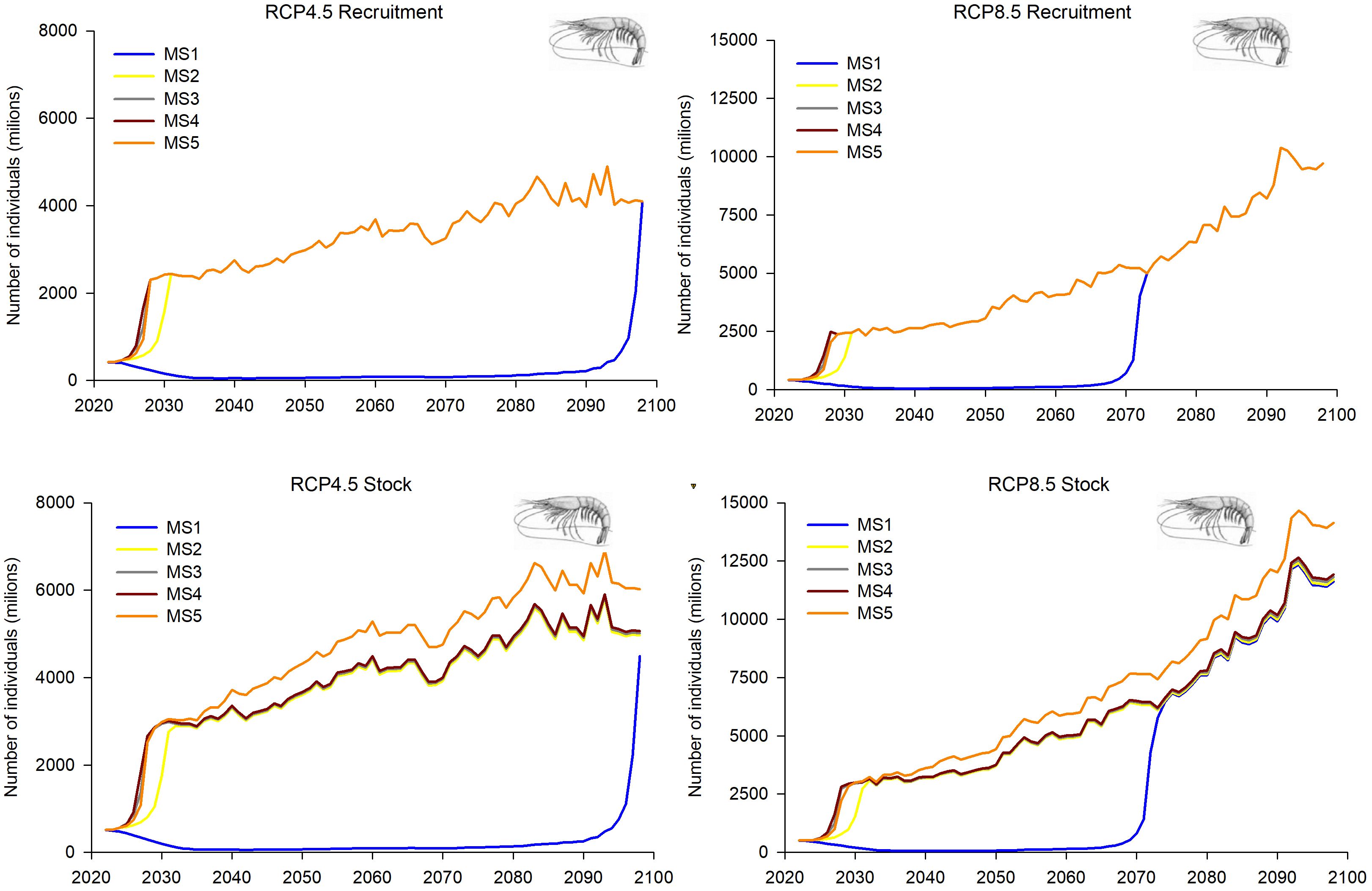
Figure 11. Predictions and projections along the 21st century of the recruitment and the overall stock of deep-water rose shrimp (Parapenaeus longirostris) in the FAO-GFCM geographical sub-area GSA 6 (Northern Spain), under the five fishing management strategies considered (see Material and methods and Figure 2 for more details) and the Regional Climatic Models under two climatic scenarios (RCP 4.5 and RCP 8.5).
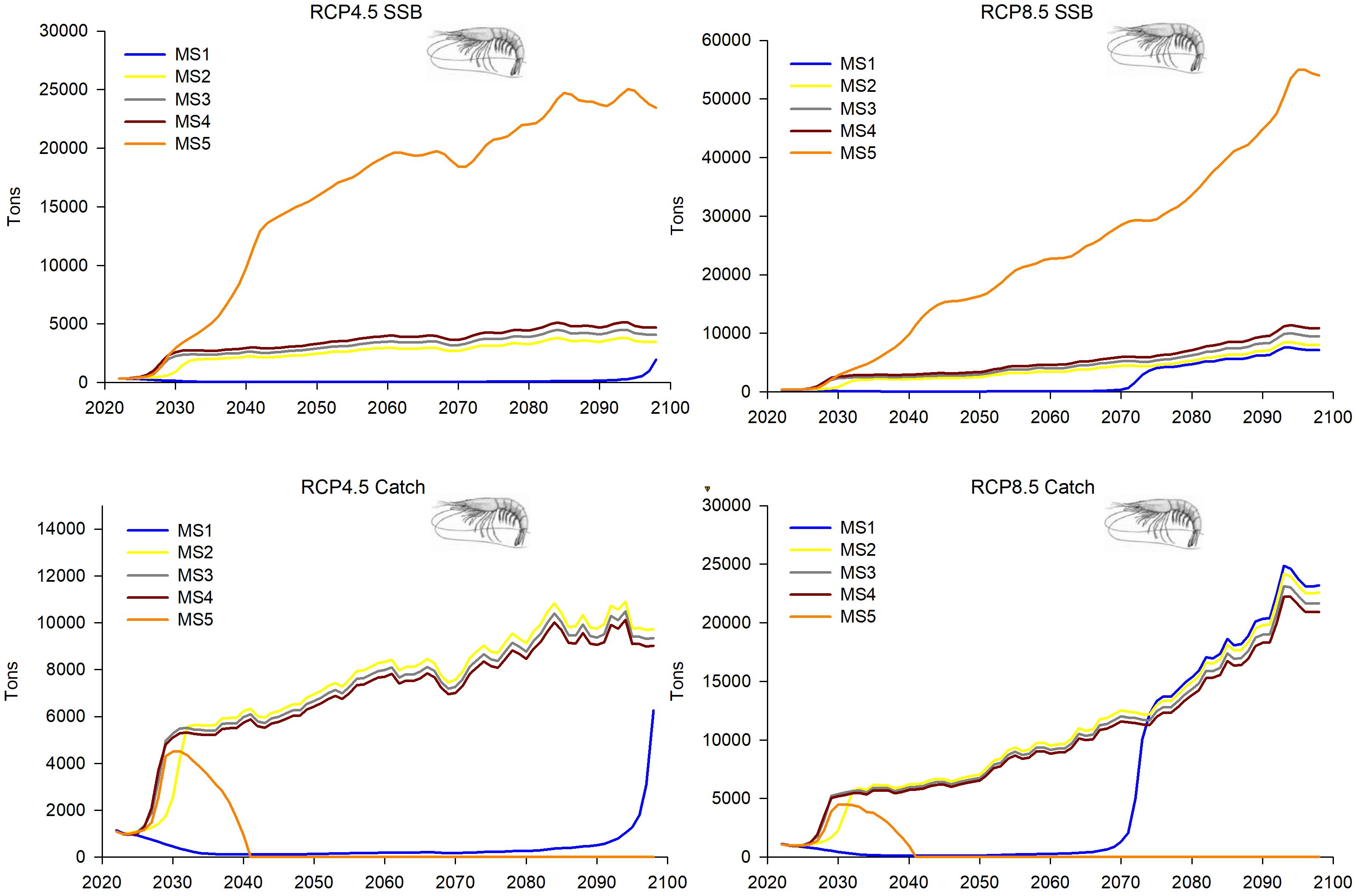
Figure 12. Predictions and projections along the 21st century of the SSB and the catches of the deep-water rose shrimp (Parapenaeus longirostris) in the FAO-GFCM sub-area GSA 6 (Northern Spain), under the five fishing management strategies considered (see Material and methods and Figure 2 for more details) and the Regional Climatic Models under two climatic scenarios (RCP4.5 and RCP8.5).
GSA 6 showed quite similar trends in the population dynamics of deep-water rose shrimp within the MSs 2–5 for R, overall stock and SSB, reaching higher values in the most severe climatic scenario (RCP8.5). These projections showed a greater increase in the first decade, followed by gradual growth (Figures 11, 12). The management strategy with the greatest fishing restriction, MS 4, presents the lowest values of R, overall stock, SSB and catches, although its trend is one of growth, being more notable in the RCP 8.5 climate scenario, where the greatest growth appears in 2070 (Figures 11, 12).
5 Discussion
Within the current context of global warming, it is important to highlight the relationship between oceanographic dynamics and the dynamics of fish populations, as fisheries management will be improved by accounting for changing productivity in a warmer ocean (Beamish et al., 2000; Free et al., 2019). However, the environmental impact on fishery resources and our knowledge of the underlying mechanisms can vary greatly between areas and stocks (e.g. Bakun, 1996; Brander, 2010) and will likely affect fishing activities and seasonal patterns, due to changes in their spatial distribution (Panzeri et al., 2024). According to Brander et al. (2013), the predictions and projections on the impact of climate on fisheries must be made at regional and local scales, where historical data series can contribute to improving complex models, being global models of limited value (Payne et al., 2016; Rutterford et al., 2015). This was the approach considered in the present study, focused on species relevant for the fishing sector in the western Mediterranean and currently subjected to a management plan (EU-MAP), which has brought important management measures for the demersal fisheries.
In this study, we modelled the effects of environmental variables on the recruitment (R) of European hake and deep-water rose shrimp, two key demersal resources in the western Mediterranean, under various fishing management strategies and climate scenarios projected throughout the 21st century. We integrated long-term fisheries data (15–20 years) from stock assessment models and oceanographic information from platforms with adequate spatial and temporal resolution. This approach enables the exploration of mesoscale environmental influences on population dynamics and supports improved stock assessments and fisheries advice in the context of global change (Hidalgo et al., 2019; van der Sleen et al., 2022).
Our results have shown that environmental changes affect differently the two species analyzed. The results of European hake also suggested different relationships between the population parameters and the environmental variables, depending on the GSA. While in GSA 5 R is lower when SSB and sea temperature increases (Winter Sea Surface Temperature), in GSA 6 a positive relationship between R and SSB with chlorophyll (Annual Surface Chlorophyll), and a negative relationship with temperature (Annual Sea Surface Temperature) and salinity (Annual Sea Average Salinity) has been detected. For deep-water rose shrimp, a positive relationship has been observed between R and SSB and sea temperature (Annual Sea Bottom Temperature) in both GSAs.
To understand this pattern, we can consider the differences in productivity between these two areas. The north-eastern Iberian Peninsula (GSA 6) and the Gulf of Lions (GSA 7) are two of the most productive areas in the Mediterranean. Although GSA 6 has historically been considered one of the most productive areas in the western Mediterranean due to nutrient inputs from the Ebro River and vertical mixing processes (Estrada, 1996; Bosc et al., 2004), recent studies confirm a sustained decline in riverine nutrient inputs over last decades, linked to reduced river discharge and anthropogenic interventions (Cozzi et al., 2018). Consequently, a decrease in vertical nutrient transport, particulate carbon flux and net productivity has been observed on the continental shelf of the Gulf of Lions, suggesting a progressive loss of productivity in the GSA 6 (Many et al., 2021; Ser-Giacomi et al., 2020). Nevertheless, both areas remain productivity hotspots overall, though they now face intensifying thermal stratification and nutrient limitation under climate change. The historical high productivity stemmed from nutrient availability supporting planktonic production and fisheries such as European hake (Lloret et al., 2001) and deep-water rose shrimp (Guijarro et al., 2009).
On the contrary, the Balearic Islands (GSA 5), where there is no supply of nutrients from land runoff, show a more pronounced oligotrophy (Estrada, 1996; Bosc et al., 2004). In this area, the input of the northern waters, a system of fronts generated by deep convention driven by intense sea-atmosphere heat exchanges and the subsequent temperature and buoyance lost of surface waters during winter induced by cold, dry and persistent northern and north-western winds during winter in the more productive northern coasts of the western Mediterranean (Monserrat et al., 2008) can act as an external fertilization mechanism that enhances the productivity (Darmaraki et al., 2019), along with the frontal meso-scale events between the Mediterranean and Atlantic waters (Pinot et al., 2002; Barceló-Llull et al., 2019). The major or minor presence of these cold waters determines the interannual variability in the hydrodynamism and productivity around the Archipelago (Massutí et al., 2014; Menna et al., 2022) and influences recruitment, somatic condition and fishing yields of demersal fishing resources (Hidalgo et al., 2008; Massutí et al., 2008; Quetglas et al., 2012; Rueda et al., 2014). Recent studies confirm that these physical mechanisms are highly sensitive to climate change. Since 2000, the depth of winter mixing has decreased by approximately 3.5% per decade, limiting the vertical transport of nutrients to surface waters and potentially constraining primary production (Darmaraki et al., 2019). At the same time, intensified marine heatwaves are destabilizing frontal systems, which are critical for nutrient retention and larval transport (Garrabou et al., 2022). These changes in oceanographic conditions, particularly in winter mixing and stratification dynamics, have been shown to significantly influence the recruitment success of key demersal species such as European hake (Lloret et al., 2001).
These differences in productivity between GSAs 5 and 6, and the different level of European hake overfishing in these areas () is almost three times higher in GSA 6 than in GSA 5) could explain the opposite effect of SSB on R. In oligotrophic GSA 5, density-dependent mortality likely stabilizes hake recruitment near carrying capacity. In contrast, the greater productivity of GSA 6, jointly with recruitment overfishing of European hake in this area, which has resulted in a progressive decrease of spawning stock biomass during the last decade (Ordines et al., 2019), would explain the positive effect of SSB on R. These authors reported that in GSA 6, lower competition for food among recruits allows individuals to achieve better somatic condition than in other GSAs, such as the Balearic Islands. This mechanistic framework like integrating productivity gradients (Many et al., 2021; Hidalgo et al., 2022b), climate-sensitive physical processes (Darmaraki et al., 2019), and fishing impacts (Ordines et al., 2019), ultimately explains the observed negative temperature-R relationship for hake in both GSAs.
Water temperature and salinity, among other environmental factors, affect the distribution and production of plankton (e.g. Edwards, 2017; Benedetti et al., 2021) and, subsequently, the food for larvae, juveniles and adults of fishing resources (e.g. Brander, 2017). European hake in the Mediterranean is not an exception, being temperature a key factor for its larval survival, through the influence on growth and mortality (Palomera et al., 2005). Specifically, several studies have demonstrated a negative effect of elevated water temperatures on hake recruitment, where summer thermal anomalies were associated with lower autumn recruit abundance in the Central Mediterranean (Bartolino et al., 2008). Similar patterns were found in the Galician shelf (García-Fernández et al., 2021), where winter and spring temperature increases were shown to reduce growth in younger hake, potentially impacting population trajectories (Vieira et al., 2020). We have observed similar results both in GSAs 5 and 6, where an increase in temperature (Winter and Annual Sea Surface Temperature, respectively) influenced negatively on European hake recruitment. This aligns with evidence that warmer winters reduce deep-water formation and nutrient upwelling in the western Mediterranean, limiting productivity (González-Gil et al., 2018; Habib et al., 2022). As established by Estrada et al. (1985) and further supported by recent studies, years with reduced winter cooling tend to be less productive in the western Mediterranean, due to weaker convective mixing and diminished formation of nutrient-rich deep waters (Pastor et al., 2019; Macías et al., 2015; Von Schuckmann et al., 2022). In contrast, colder-than-average winters promote deep winter convection in the Gulf of Lions, enhancing the upward transport of nutrients and fueling spring phytoplankton blooms (Estrada et al., 2014; Conan et al., 2018; Fourrier et al., 2022). These events contribute to the formation of Western Mediterranean Intermediate Waters, which flow southwards and can reach the Balearic Channels in spring, potentially increasing productivity in these regions and creating favorable conditions for the recruitment of European hake (Massutí et al., 2008; Estrada et al., 2014; Colin et al., 2021).
Physiologically, European hake, a cold adapted Atlantic species (Lloris et al., 2003), faces thermal stress in warming Mediterranean waters. While early twenty first century populations showed resilience (Hidalgo, 2007), recent studies confirm that chronic overfishing has eroded adaptive capacity, exacerbating climate vulnerability (Bastardie et al., 2022). Thus, its recruitment decline reflects both Mediterranean warming (Vieira et al., 2020) and fishing impact.
Warming favors smaller zooplankton, which have shown declining biomass and size in areas like the Bay of Marseille (north-western Mediterranean), particularly during warmer winters with reduced nutrient input (Garcia et al., 2023). Biochemical shifts in these communities (such as decreased lipid content despite stable energy density due to higher protein and carbohydrate levels) suggest reduced nutritional quality for higher trophic levels (Garcia et al., 2024). These bottom-up changes impair energy transfer to small pelagics, the main prey of European hake (Palomera et al., 2007; Coll et al., 2013), and illustrate a “trophic squeeze” that may intensify with continued warming (Archibald et al., 2022).
Parras-Berrocal et al. (2022) have studied the effect of global warming under the RCP 8.5 climatic scenario and predicted a possible collapse of the deep water formation in the Gulf of Lions during the period 2040-2050, with impacts on the overturning circulation and even in the heat and salinity exchange between the Mediterranean Sea and the Atlantic Ocean. Taking into account the importance of deep water formation in the productivity of the north-western Mediterranean (Estrada et al., 1985) and its effect on demersal species (Massutí et al., 2008; Quetglas et al., 2012; Rueda et al., 2014), this change in oceanographic dynamics could impact significantly on fish populations and fishing resources. While our models did not incorporate this collapse scenario, they project declining European hake recruitment, population biomass, and catches throughout the 21st century under medium/extreme warming trends largely unaffected by fishing management strategies. Using the same global warming scenario for spatial distribution projections, Panzeri et al. (2024) found spatial changes for hotspot areas of juveniles and adults in the Adriatic Sea for certain demersal species, including European hake. This aligns with established synergies between environmental stress and overexploitation (Rosenberg, 2003; Hidalgo et al., 2011), now quantified for Mediterranean hake (Moullec et al., 2016). Critically, even high intrinsic resilience (Hidalgo et al., 2011) cannot compensate for compound climate-fishing pressures under projected warming, as evidenced by population declines under conservative (MS 4) and zero-fishing (MS 5) scenarios. Our results also show that this future decline in western Mediterranean European hake populations, due to climate change, will be more pronounced in the Balearic Islands (GSA 5) than in the Iberian Peninsula (GSA 6), even if the current state of exploitation is better in the former than in the later. This can be due not only to the greater oligotrophy of the waters surrounding the Archipelago, but also because its productivity depends on the greater or lesser reinforcement of the shelf-break fronts, as a consequence of the deep water formation in the northern coast of the north-western Mediterranean (Massutí et al., 2008; Richon et al., 2019). On the contrary, off Iberian Peninsula, in addition to the greater productivity of its waters, there are other factors such as runoff from rivers that can contribute to increasing it, although these will also be negatively affected by climate change (Navarro-Ortega et al., 2012; Alvarez et al., 2024). This shows the important role climatic conditions play in the dynamics of European hake, even higher than management measures, and how much relevant is the incorporation of its influence in the current process of assessment and management of this resource.
The case of deep-water rose shrimp represents the opposite scenario, as the abundance of this thermophilic species is enhanced by the increment of water temperature, as shown in our results, with the positive influence of temperature (Annual Sea Bottom Temperature) in R both in GSAs 5 and 6. These results corroborated those reported by Abelló et al. (2002) and Fortibuoni et al. (2010) and are reinforced by recent studies documenting significant range expansions of this species under Mediterranean warming (Sbrana et al., 2019; Quattrocchi et al., 2020; Mingote et al., 2024; González-Andrés et al., 2024). Colloca et al. (2014) also pointed out a positive relationship between its abundance and the increase in water temperature in the Ligurian, Ionian and Tyrrhenian seas, a pattern now observed throughout the western Mediterranean. The mechanistic basis for this response involves improved survival of planktonic stages. As larvae predominantly inhabit upper slopes and deep continental shelves (Torres et al., 2013), temperature-mediated acceleration of larval metamorphosis and growth enhances recruitment (Zeng et al., 2020). The SSB of deep-water rose shrimp also shows a positive relationship with temperature. In this short-lived species, the increase in temperature can accelerate the growth and sexual maturation, thus increasing the spawning biomass (Sobrino et al., 2005).
In contrast to European hake, an increase in the abundance and fishing yields of the thermophilic species deep-water rose shrimp along the 21st century has been detected. In GSA 5, this species also shows lower R and overall stock in the most restrictive fishing strategies (MSs 5 and 6). This may be due to the pronounced oligotrophy of the Balearic Islands, and the lower carrying capacity of their ecosystems, which increases competition for food and does not allow an increase in the abundance due to a more limiting carrying capacity. Recent empirical and theoretical research supports density-dependent regulation in marine decapods: reductions in individual condition and size under high population densities have been observed in juvenile penaeid shrimps (Blanco-Martínez et al., 2020). Although specific studies focused on P. longirostris in the Mediterranean are still limited, bio-ecological assessments of crustacean fisheries in the western Mediterranean provide ecologically consistent evidence that intra- and inter-specific competition likely modulate stock biomass at higher densities (Awadh and Aksissou, 2020).
Given the multispecificity of the Mediterranean demersal fisheries (Lleonart and Maynou, 2003), it is necessary to balance the economic losses and gains from changes in the catchability of different species in the context of climate change (Cheung et al., 2010). That is the case of European hake and deep-water rose shrimp, among the most important target species for the Mediterranean bottom trawl fleets, and currently subjected to a management plan (EU-MAP). The impacts of climate change on fisheries can manifest in various ways. Several studies suggest the idea that an increase in water temperature could lead to greater availability of thermophilic (warm-water) species, potentially boosting catches of some commercial species. For instance, research indicates that as ocean temperatures rise, species adapted to warmer conditions may expand their range, becoming more abundant and available to fisheries in certain areas (Pecl et al., 2017; González-Andrés et al., 2024). Conversely, species that thrive in colder waters may see their populations decline or migrate to deeper, cooler areas, reducing their availability to fisheries. This redistribution of species can significantly impact the economic outcomes for fisheries, depending on their ability to adapt to these changes (Barange et al., 2018; Pecl et al., 2017; Payne et al., 2021). The overall commercial and economic impacts of these shifts can be complex and unpredictable (Panzeri et al., 2024). While some regions might experience increased profitability from higher catches of thermophilic species, others might face economic challenges due to the decreased availability of traditionally targeted cold-water species. Furthermore, changes influenced by climate change can also affect the trophic networks and, consequently, the availability to fisheries of some target species (Gambaiani et al., 2009; Hidalgo et al., 2022a). These insights underscore the needs to adopt an ecosystem-based approach to fisheries management taking into account the multifaceted impacts of climate change on marine ecosystems and the socioeconomic systems dependent on them (Hidalgo et al., 2022a; Ofir et al., 2023).
Expanding our knowledge on how oceanographic variables influence the population dynamics of European hake, deep-water rose and other species of fishing interest, as well as knowing their evolution in response to different management measures is fundamental in the current context of global warming. The two species analyzed in the present study, jointly with other demersal resources (red mullet Mullus barbatus, blue and red shrimp Aristeus antennatus, giant red shrimp Aristaeomorpha foliacea and Norway lobster Nephrops norvegicus) are currently subjected to a multiannual plan for fisheries exploiting demersal stocks in the western Mediterranean (EU-MAP). This management plan focuses on the short-term biological impact of fisheries on these populations and the application of certain management measures to reach their sustainable exploitation in the long term. The assessment models in which the management measures from the EU-MAP are applied, do not consider the influence of global warming on the population dynamics of the target stocks. Thus, the results of this study highlight the need of considering these aspects as they show that the forecast of the population dynamics of two of the main demersal resources in the western Mediterranean will depend more on the influence of the climate than on the current and future fishing management.
It is important to acknowledge several limitations in the current modeling approach. First, the model assumes static relationships between environmental variables and species recruitment over the period studied (up to 80 years), which may overlook potential temporal changes due to evolutionary adaptations or shifting environmental regimes. Additionally, multispecies interactions, habitat shifts, and abrupt ecosystem changes such as regime shifts have not been incorporated. These factors can significantly affect population dynamics and may introduce additional uncertainty into projections. Explicitly recognizing these limitations is essential, and future research should aim to integrate these complex ecological processes to improve model accuracy and applicability.
Furthermore, the transferability of the model results to other Mediterranean subareas (GSAs) and demersal species with similar ecological and fishery characteristics should be done with caution. While the combination of long-term fisheries data, regional climate models, and adaptive management scenarios provides a valuable tool, its effectiveness depends heavily on the availability and quality of local data, as well as on appropriate validation efforts. Thus, specific studies from each species and subareas should be carried out to obtain realistic results.
6 Conclusions
The present study has shown good predictive results that highlight how environmental factors can affect the recruitment of European hake and deep-water shrimp in the western Mediterranean. The projections along the 21st century indicate that climate change is a dominant long-term driver of the population dynamics of these two species, with similar outcomes regardless of the fishing management strategies applied. The contrasting temporal evolution observed is due to the opposite thermal affinities of the two species. These results emphasize the critical importance of integrating environmental and climate change scenarios into fishery assessments, in line with the Ecosystem Approach to Fisheries. We strongly advocate for the incorporation of climate influence and projections into existing scientific advisory and management bodies, such as those of the FAO- GFCM and the EU-STECF (Scientific, Technical and Economic Committee for Fisheries), to enhance adaptive strategies and ensure sustainable exploitation of demersal resources under changing climatic conditions.
Data availability statement
Publicly available datasets were analyzed in this study. This data can be found here: GFCM Working Group on Stock Assessment of Demersal Species https://www.fao.org/gfcm/data/star/en/.
Author contributions
CG: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. BG: Funding acquisition, Project administration, Conceptualization, Formal analysis, Methodology, Software, Writing – original draft, Writing – review & editing. ER: Formal analysis, Methodology, Software, Writing – review & editing. EM: Conceptualization, Funding acquisition, Investigation, Project administration, Resources, Supervision, Visualization, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This study has been developed within the FishClim project “Scientific knowledge for adaptation to climate change in the Spanish fishing sector” (MAPA-04-03), funded by the European Union-NextGenerationEU through the Axis 4 of the “Agreement between the Ministry of Agriculture, Fisheries and Food and the State Agency Superior Council for Scientific Research, M.P. –through the Spanish Institute of Oceanography– to promote fishing research as a basis for sustainable fisheries management”. It has also received support from the SosMed Project (MAPA-01-01), included in Axis 1 of the same agreement. MEDITS survey, the collection of commercial data and the development of stock assessment models are co-financed by the European Union through the European Maritime Fisheries and Aquaculture Fund (EMFAF) within the National Program of collection, management and use of data in the fisheries sector and support for scientific advice regarding the Common Fisheries Policy. We acknowledge support of the publication fee by the CSIC Open Access Publication Support Initiative (PROA) through its Unit of Information Resources for Research (URICI).
Acknowledgments
We would like to thank the Spanish General Secretariat of Fisheries, the Spanish Institute of Oceanography (IEO-CSIC) and the Institute of Marine Sciences of Andalucía (ICMAN-CSIC) for their help in the creation and development of the FishClim project.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2025.1587297/full#supplementary-material
References
Abella A., Serena F., and Caddy J. F. (1998). Estimation of the parameters of the Caddy reciprocal M-at-age model for the construction of natural mortality vectors. Cahiers Options Mediterraneennes (CIHEAM) 35, 191–200.
Abelló P., Abella Á., Adamidou A., Jukic-Peladic S., Maiorano P., and Spedicato M. T. (2002). Geographical patterns in abundance and population structure of Nephrops norvegicus and Parapenaeus longirostris (Crustacea: Decapoda) along the European Mediterranean coasts. Scientia Marina 66, 125. doi: 10.3989/scimar.2002.66s2125
Alheit J. and Pitcher T. J (Eds.) (1995). Hake: fisheries, ecology and markets (London: Chapman and Hall), 478 p.
Alvarez I., Pereira H., Lorenzo M. N., Picado A., Sousa M. C., Taboada J. J., et al. (2024). Drought projections for the NW Iberian Peninsula under climate change. Climate Dynamics 62, 4775–4791. doi: 10.1007/s00382-023-07084-z
Archibald K. M., Dutkiewicz S., Laufkötter C., and Moeller H. V. (2022). Thermal responses in global marine planktonic food webs are mediated by temperature effects on metabolism. J. Geophysical Research: Oceans 127, e2022JC018932. doi: 10.1029/2022JC018932
Astudillo A. and Caddy J. (1988). “Periodicidad de los desembarcos de merluza (Merluccius merluccius) y salmonete (Mullus sp.) en la isla de Mallorca,” Wyatt T. and Larrañeta M.G. in International Symposium on Long Term Changes in Marine Fish Populations. 221–234. Imprenta Real, Vigo (Spain).
Awadh H. and Aksissou M. (2020). Population dynamics and stock assessment of deep water rose shrimp Parapenaeus longirostris (Lucas 1846) from the Mediterranean coast of Morocco. Indian J. Fisheries 67, 8–14. doi: 10.21077/ijf.2019.67.2.93423-02
Bakun A. (1996). Patterns in the Ocean: Ocean Processes and Marine Population Dynamics (La Jolla, CA: California Sea Grant College System, NOAA), 323 pp.
Barange M., Bahri T., Beveridge M. C. M., Cochrane K. L., Funge-Smith S., Poulain F., et al, eds. (2018). Impacts of climate change on fisheries and aquaculture: synthesis of current knowledge, adaptation and mitigation options. FAO Fisheries and Aquaculture Technical Paper No. 627. FAO, Rome. 628.
Barceló-Llull B., Pascual A., Ruiz S., Escudier R., Torner M., and Tintoré J. (2019). Temporal and spatial hydrodynamic variability in the Mallorca channel (western Mediterranean Sea) from 8 years of underwater glider data. J. Geophysical Research: Oceans 124, 2769–2786. doi: 10.1029/2018JC014636
Bartolino V., Colloca F., Sartor P., and Ardizzone G. (2008). Modelling recruitment dynamics of hake, Merluccius merluccius, in the central Mediterranean in relation to key environmental variables. Fisheries Res. 92, 277–288. doi: 10.1016/j.fishres.2008.01.007
Bastardie F., Feary D. A., Brunel T., Kell L. T., Döring R., Metz S., et al. (2022). Ten lessons on the resilience of the EU common fisheries policy towards climate change and fuel efficiency-A call for adaptive, flexible and well-informed fisheries management. Front. Mar. Sci. 9, 947150. doi: 10.3389/fmars.2022.947150
Beamish R. J., McFarlane G. A., and King J. R. (2000). Chapter 5 Fisheries climatology: understanding decadal scale processes that naturally regulate British Columbia fish populations. Editors: Paul J. Harrison, Timothy R. Parsons. In book: Fisheries Oceanography: An Integrative Approach to Fisheries Ecology and Management, 94–145. Blackwell Science, University of British Columbia, Canada.
Benedetti F., Vogt M., Elizondo U. H., Righetti D., Zimmermann N. E., and Gruber N. (2021). Major restructuring of marine plankton assemblages under global warming. Nat. Commun. 12, 5226. doi: 10.1038/s41467-021-25385-x
Blanco-Martínez Z., Pérez-Castañeda R., Sánchez-Martínez J. G., Benavides-González F., Rábago-Castro J. L., de la Luz Vázquez-Sauceda M., et al. (2020). Density-dependent condition of juvenile penaeid shrimps in seagrass-dominated aquatic vegetation beds located at different distance from a tidal inlet. PeerJ 8, e10496. doi: 10.7717/peerj.10496
Bosc E., Bricaud A., and Antoine D. (2004). Seasonal and interannual variability in algal biomass and primary production in the Mediterranean Sea, as derived from 4 years of SeaWiFS observations. Global Biogeochemical Cycles 18, 1–17. doi: 10.1029/2003GB002034
Branch T. A., Watson R., Fulton E. A., Jennings S., McGilliard C. R., Pablico G. T., et al. (2010). The trophic fingerprint of marine fisheries. Nature 468, 431–435. doi: 10.1038/nature09528
Brander K. (2010). Impacts of climate change on fisheries. J. Mar. Syst. 79, 389–402. doi: 10.1016/j.jmarsys.2008.12.015
Brander K. (2015). Improving the reliability of fishery predictions under climate change. Curr. Climate Change Rep. 1, 40–48. doi: 10.1007/s40641-015-0005-7
Brander K. (2017). “Plankton and fisheries,” in Marine Plankton: A practical guide to ecology, methodology, and taxonomy. Eds. Castellani C. and Edwards M. (Oxford (United Kingdom): Oxford University Press), 81–88.
Brander K., Neuheimer A., Andersen K. H., and Hartvig M. (2013). Overconfidence in model projections. ICES J. Mar. Sci. 70, 1065–1068. doi: 10.1093/icesjms/fst055
Burnham K. P. and Anderson D. R. (2004). Multimodel inference: understanding AIC and BIC in model selection. Sociological Methods Res. 33, 261–304. doi: 10.1177/0049124104268644
Casey J. and Pereiro J. (1995). “European hake (M. merluccius) in the North-east Atlantic,” in Hake: Biology, fisheries and markets (Springer Netherlands, Dordrecht), 125–147.
Cheung W. W., Lam V. W., Sarmiento J. L., Kearney K., Watson R. E. G., Zeller D., et al. (2010). Large-scale redistribution of maximum fisheries catch potential in the global ocean under climate change. Global Change Biol. 16, 24–35. doi: 10.1111/j.1365-2486.2009.01995.x
Colin C., Duhamel M., Siani G., Dubois-Dauphin Q., Ducassou E., Liu Z., et al. (2021). Changes in the intermediate water masses of the Mediterranean Sea during the last climatic cycle—New constraints from neodymium isotopes in foraminifera. Paleoceanography Paleoclimatology 36, e2020PA004153. doi: 10.1029/2020PA004153
Coll M., Navarro J., Olson R. J., and Christensen V. (2013). Assessing the trophic position and ecological role of squids in marine ecosystems by means of food-web models. Deep Sea Res. Part II: Topical Stud. Oceanography 95, 21–36. doi: 10.1016/j.dsr2.2012.08.020
Coll M., Steenbeek J., Sole J., Palomera I., and Christensen V. (2016). Modelling the cumulative spatial–temporal effects of environmental drivers and fishing in a NW Mediterranean marine ecosystem. Ecol. Model. 331, 100–114. doi: 10.1016/j.ecolmodel.2016.03.020
Colloca F., Mastrantonio G., Lasinio G. J., Ligas A., and Sartor P. (2014). Parapenaeus longirostris (Lucas 1846) an early warning indicator species of global warning in the central Mediterranean Sea. J. Mar. Syst. 138, 29–39. doi: 10.1016/j.jmarsys.2013.10.007
Colloca F., Scarcella G., and Libralato S. (2017). Recent trends and impacts of fisheries exploitation on Mediterranean stocks and ecosystems. Front. Mar. Sci. 4, 244. doi: 10.3389/fmars.2017.00244
Conan P., Testor P., Estournel C., d’Ortenzio F., and de Madron X. D. (2018). Observing Winter Mixing and Spring Bloom in the Mediterranean Vol. 99 (Eos, Transactions American Geophysical Union).
Cozzi S., Ibáñez C., Lazar L., Raimbault P., and Giani M. (2018). Flow regime and nutrient-loading trends from the largest South European watersheds: Implications for the productivity of mediterranean and Black Sea’s Coastal Areas. Water 11, 1. doi: 10.3390/w11010001
Darmaraki S., Somot S., Sevault F., Nabat P., Cabos Narvaez W. D., Cavicchia L., et al. (2019). Future evolution of marine heatwaves in the Mediterranean Sea. Climate Dynamics 53, 1371–1392. doi: 10.1007/s00382-019-04661-z
Daw T. M., Adger N., Brown K., and Badjeck M. C. (2009). “Climate change and capture fisheries. Climate change implications for fisheries and aquaculture. Overview of current scientific knowledge,” in FAO Fisheries and Aquaculture Technical Paper. Eds. Cochrane K., Young C. D., Soto D., and Bahri T. (Food and Agricultural Organization, Rome), 95–135.
Druon J. N., Fiorentino F., Murenu M., Knittweis L., Colloca F., Osio C., et al. (2015). Modelling of European hake nurseries in the Mediterranean Sea: an ecological niche approach. Prog. oceanography 130, 188–204. doi: 10.1016/j.pocean.2014.11.005
Duffy-Anderson J. T., Bailey K., Ciannelli L., Cury P., Belgrano A., and Stenseth N. C. (2005). Phase transitions in marine fish recruitment processes. Ecol. Complexity 2, 205–218. doi: 10.1016/j.ecocom.2004.12.002
Edwards M. (2017). “Plankton and global change,” in Marine Plankton: A practical guide to ecology, methodology, and taxonomy. Eds. Castellani C. and Edwards M. (Oxford (United Kingdom): Oxford University Press), 67–80.
Estrada M., Latasa M., Emelianov M., Gutiérrez-Rodríguez A., Fernández-Castro B., Isern-Fontanet J., et al. (2014). Seasonal and mesoscale variability of primary production in the deep winter-mixing region of the NW Mediterranean. Deep Sea Res. Part I: Oceanographic Res. Papers 94, 45–61. doi: 10.1016/j.dsr.2014.08.003
Estrada M., Vives F., and Alcaraz M. (1985). Chapter 6: Life and the Productivity of the Open Sea. In: Margalef R. (ed). Western Mediterranean (pp. 148–197). Pergamon Press, Oxford, United Kingdom.
FAO (2022). General Fisheries Commission for the Mediterranean – Report of the twenty-third session of the Scientific Advisory Committee on Fisheries, FAO headquarters, Rome, Italy, 21–24 June 2022 (Rome: FAO Fisheries and Aquaculture Report No. 1395). doi: 10.4060/cc3109en
FAO (2023). The State of Mediterranean and Black Sea Fisheries 2023 – Special edition (Rome: General Fisheries Commission for the Mediterranean). doi: 10.4060/cc8888en
FAO. (2024). General Fisheries Commission for the Mediterranean – Report of the twenty-fourth session of the Scientific Advisory Committee on Fisheries, FAO headquarters, Rome, Italy, 20–23 June 2023. FAO Fisheries and Aquaculture Report, No. 1421. Rome. doi: 10.4060/cc8652en
Fernández de Puelles M. L., Valencia J., Jansá J., and Morillas A. (2004). Hydrographical characteristics and zooplankton distribution in the Mallorca channel (Western Mediterranean): spring 2001. ICES J. Mar. Sci. 61, 654–666. doi: 10.1016/j.icesjms.2004.03.031
Fortibuoni T., Bahri T., Camilleri M., Gristina M., Garofalo G., and Fiorentino F. (2010). Nursery and spawning areas of deep-water rose shrimp, Parapenaeus longirostris (Decapoda: Penaeidae), in the strait of Sicily (Central Mediterranean sea). J. Crustacean Biol. 30, 167–174. doi: 10.1651/09-3167.1
Fourrier M., Coppola L., D’ortenzio F., Migon C., and Gattuso J. P. (2022). Impact of intermittent convection in the northwestern Mediterranean Sea on oxygen content, nutrients, and the carbonate system. J. Geophysical Research: Oceans 127, e2022JC018615. doi: 10.1029/2022JC018615
Free C. M., Thorson J. T., Pinsky M. L., Oken K. L., Wiedenmann J., and Jensen O. P. (2019). Impacts of historical warming on marine fisheries production. Science 363, 979–983. doi: 10.1126/science.aau1758
Gambaiani D. D., Mayol P., Isaac S. J., and Simmonds M. P. (2009). Potential impacts of climate change and greenhouse gas emissions on Mediterranean marine ecosystems and cetaceans. J. Mar. Biol. Assoc. United Kingdom 89, 179–201. doi: 10.1017/S0025315408002476
Garcia T., Bănaru D., Guilloux L., Cornet V., Gregori G., and Carlotti F. (2023). Temporal changes in zooplankton indicators highlight a bottom-up process in the Bay of Marseille (NW Mediterranean Sea). PloS One 18, e0292536. doi: 10.1371/journal.pone.0292536
Garcia T., Carlotti F., Lepoint G., Guilloux L., Tesán-Onrubia J. A., Grassi B., et al. (2024). The decadal variability of elemental, isotopic, and biochemical compositions of coastal Mediterranean zooplankton responds to environmental changes. Mar. Biol. 171, 119. doi: 10.1007/s00227-024-04430-5
García-Fernández C., Suca J. J., Llopiz J. K., Álvarez P., Domínguez-Petit R., and Saborido-Rey F. (2021). Spatial and temporal variability in the occurrence and abundance of European Hake larvae, Merluccius merluccius, on the Galician shelf (NE Atlantic). Front. Mar. Sci. 8, 696246. doi: 10.3389/fmars.2021.696246
Garrabou J., Gómez-Gras D., Medrano A., Cerrano C., Ponti M., Schlegel R., et al. (2022). Marine heatwaves drive recurrent mass mortalities in the Mediterranean Sea. Global Change Biol. 28, 5708–5725. doi: 10.1111/gcb.16301
GFCM (2024).Report of the twenty-fifth session of the Scientific Advisory Committee on Fisheries, Marseille, France, 24–27 June 202/Rapport de la vingt-cinquième session du Comité scientifique consultatif des pêches, Marseille, France, 24–27 juin 2024. Available online at: https://www.fao.org/gfcm/statutory-meetings/detail/en/c/1712734/ (Accessed December 2, 2024).
González-Andrés C., Ramírez-Romero E., Guijarro B., Farre M., Macias D., and Massutí E. (2024). Warming promotes expansion of a key demersal fishing resource of the western Mediterranean. Front. Mar. Sci. 11, 1494177. doi: 10.3389/fmars.2024.1494177
González-Gil R., González Taboada F., Cáceres C., Largier J. L., and Anadón R. (2018). Winter-mixing preconditioning of the spring phytoplankton bloom in the Bay of Biscay. Limnology Oceanography 63, 1264–1282. doi: 10.1002/lno.10769
Guijarro B., Massutí E., Moranta J., and Cartes J. E. (2009). Short spatio-temporal variations in the population dynamics and biology of the deep-water rose shrimp Parapenaeus longirostris (Decapoda: Crustacea) in the western Mediterranean. Scientia Marina 73, 183–197. doi: 10.3989/scimar.2009.73n1183
Habib J., Ulses C., Estournel C., Fakhri M., Marsaleix P., Pujo-Pay M., et al. (2022). Seasonal and interannual variability of the pelagic ecosystem and of the organic carbon budget in the Rhodes Gyre (Eastern Mediterranean): influence of winter mixing. Biogeosciences Discussions 2022, 1–45. doi: 10.5194/bg-20-3203-2023
Harley C. D., Randall Hughes A., Hultgren K. M., Miner B. G., Sorte C. J., Thornber C. S., et al. (2006). The impacts of climate change in coastal marine systems. Ecol. Lett. 9, 228–241. doi: 10.1111/j.1461-0248.2005.00871.x
Hastie T. J. and Tibshirani R. J. (1990). Generalized additive models Vol. 43. Eds. Cox, Hinkley, Rubin, and Silverman (Boca Rato (United States of America): CRC press), 1–329.
Hidalgo M. (2007) in Recruitment process and population dynamics of European hake (Merluccius merluccius L.) off the Balearic Islands: seasonal and interannual approach (Doctoral dissertation, PhD dissertation, University of Vigo, Vigo).
Hidalgo M., El-Haweet A. E., Tsikliras A. C., Tirasin E. M., Fortibuoni T., Ronchi F., et al. (2022a). Risks and adaptation options for the Mediterranean fisheries in the face of multiple climate change drivers and impacts. ICES J. Mar. Sci. 79, 2473–2488. doi: 10.1093/icesjms/fsac185
Hidalgo M., Massutí E., Guijarro B., Moranta J., Ciannelli L., Lloret J., et al. (2009). Population effects and changes in life history traits in relation to phase transitions induced by long-term fishery harvesting: European hake (Merluccius merluccius) off the Balearic Islands. Can. J. Fisheries Aquat. Sci. 66, 1355–1370. doi: 10.1139/F09-081
Hidalgo M., Massutí E., Moranta J., Cartes J., Lloret J., Oliver P., et al. (2008). Seasonal and short spatial patterns in European hake (Merluccius merluccius L.) recruitment process at the Balearic Islands (western Mediterranean): the role of environment on distribution and condition. J. Mar. Syst. 71, 367–384. doi: 10.1016/j.jmarsys.2007.03.005
Hidalgo M., Rossi V., Monroy P., Ser-Giacomi E., Hernández-García E., Guijarro B., et al. (2019). Accounting for ocean connectivity and hydroclimate in fish recruitment fluctuations within transboundary metapopulations. Ecol. Appl. 29, e01913. doi: 10.1002/eap.1913
Hidalgo M., Rouyer T., Molinero J. C., Massutí E., Moranta J., Guijarro B., et al. (2011). Synergistic effects of fishing-induced demographic changes and climate variation on fish population dynamics. Mar. Ecol. Prog. Ser. 426, 1–12. doi: 10.3354/meps09077
Hidalgo M., Vasilakopoulos P., García-Ruiz C., Esteban A., López-López L., and García-Gorriz E. (2022b). Resilience dynamics and productivity-driven shifts in the marine communities of the Western Mediterranean Sea. J. Anim. Ecol. 91, 470–483. doi: 10.1111/1365-2656.13648
Hjort J. (1914). Fluctuations in the great fisheries of northern Europe viewed in the light of biological research. Rapport Proces-Verbaux Des. Réunion Conseil Int. pour l’Exploration la Mer 20, 1–228.
Jardim E., Millar C. P., Mosqueira I., Scott F., Osio G. C., Ferreti M., et al. (2014). What if stock assessment is as simple as a linear model? The a4a initiative. ICES J. Mar. Sci. 72, 232–236. doi: 10.1093/icesjms/fsu050
Katsanevakis S., Maravelias C. D., Damalas D., Karageorgis A. P., Tsitsika E. V., Anagnostou C., et al. (2009). Spatiotemporal distribution and habitat use of commercial demersal species in the eastern Mediterranean Sea. Fisheries Oceanography 18, 439–457. doi: 10.1111/j.1365-2419.2009.00523.x
Kirby R. R., Beaugrand G., and Lindley J. A. (2009). Synergistic effects of climate and fishing in a marine ecosystem. Ecosystems 12, 548–561. doi: 10.1007/s10021-009-9241-9
Lezama-Ochoa N., Murua H., Hall M., Román M., Ruiz J., Vogel N., et al. (2017). Biodiversity and habitat characteristics of the bycatch assemblages in fish aggregating devices (FADs) and school sets in the eastern Pacific Ocean. Front. Mar. Sci. 4, 265. doi: 10.3389/fmars.2017.00265
Ligas A., Sartor P., and Colloca F. (2011). Trends in population dynamics and fishery of Parapenaeus longirostris and Nephrops norvegicus in the Tyrrhenian Sea (NW Mediterranean): the relative importance of fishery and environmental variables. Mar. Ecol. 32, 25–35. doi: 10.1111/j.1439-0485.2011.00440.x
Lleonart J. and Maynou F. (2003). Fish stock assessments in the Mediterranean: state of the art. Scientia Marina 67, 37–49. doi: 10.3989/scimar.2003.67s137
Lloret J., Lleonart J., Solé I., and Fromentin J. M. (2001). Fluctuations of landings and environmental conditions in the north-western Mediterranean Sea. Fisheries Oceanography 10, 33–50. doi: 10.1046/j.1365-2419.2001.00151.x
Lloris D., Matallanas J., and Oliver P. (2003). Merluzas del mundo (Familia Merlucciidae). Catálogo comentado e ilustrado de las merluzas conocidas. FAO Cat. Esp. Pesc. 2, 1–57.
Macías García-Gorriz D. M. E. and Sips A. (2015). Productivity changes in the Mediterranean Sea for the twenty-first century in response to changes in the regional atmospheric forcing. Front. Mar. Sci. 2, 79. doi: 10.3389/fmars.2015.00079
Many G., Ulses C., Estournel C., and Marsaleix P. (2021). Particulate organic carbon dynamics in the Gulf of Lion shelf (NW Mediterranean) using a coupled hydrodynamic–biogeochemical model. Biogeosciences 18, 5513–5538. doi: 10.5194/bg-18-5513-2021
Massutí E., Monserrat S., Oliver P., Moranta J., López-Jurado J. L., Marcos M., et al. (2008). The influence of oceanographic scenarios on the population dynamics of demersal resources in the western Mediterranean: Hypothesis for hake and red shrimp off Balearic Islands. J. Mar. Syst. 71, 421–438. doi: 10.1016/j.jmarsys.2007.01.009
Massutí E., Olivar M. P., Monserrat S., Rueda L., and Oliver P. (2014). Towards understanding the influence of environmental conditions on demersal resources and ecosystems in the western Mediterranean: Motivations, aims and methods of the IDEADOS project. J. Mar. Syst. 138, 3–19. doi: 10.1016/j.jmarsys.2014.01.013
Maynou F. (2011). “Impact of NAO on Mediterranean fisheries,” in Hydrological, Socioeconomic and Ecological Impacts of the North Atlantic Oscillation in the Mediterranean Region, vol. 46 . Eds. Vicente-Serrano S. and Trigo R. (Springer, Dordrecht). Advances in Global Change Research. doi: 10.1007/978-94-007-1372-7_7
Maynou F., Lleonart J., and Cartes J. E. (2003). Seasonal and spatial variability of hake (Merluccius merluccius L.) recruitment in the NW Mediterranean. Fisheries Res. 60, 65–78. doi: 10.1016/S0165-7836(02)00062-0
Mellon-Duval C., De Pontual H., Métral L., and Quemener L. (2010). Growth of European hake (Merluccius merluccius) in the Gulf of Lions based on conventional tagging. ICES J. Mar. Sci. 67, 62–70. doi: 10.1093/icesjms/fsp215
Menna M., Gačić M., Martellucci R., Notarstefano G., Fedele G., Mauri E., et al. (2022). Climatic, decadal, and interannual variability in the upper layer of the Mediterranean Sea using remotely sensed and in-situ data. Remote Sens. 14, 1322. doi: 10.3390/rs14061322
Mingote M. G., Galimany E., Sala-Coromina J., Bahamon N., Ribera-Altimir J., Santos-Bethencourt R., et al. (2024). Warming and salinization effects on the deep-water rose shrimp, Parapenaeus longirostris, distribution along the NW Mediterranean Sea: Implications for bottom trawl fisheries. Mar. pollut. Bull. 198, 115838. doi: 10.1016/j.marpolbul.2023.115838
Monserrat S., López-Jurado J. L., and Marcos M. (2008). A mesoscale index to describe the regional circulation around the Balearic Islands. J. Mar. Syst. 71, 413–420. doi: 10.1016/j.jmarsys.2006.11.012
Moullec F., Benedetti F., Saraux C., van Beveren E., and Shin Y. J. (2016). Sub-chapter 2.1.2 Climate change induces bottom-up changes in the food webs of the Mediterranean Sea. (IRD Éditions) In: Thiébault S. and Médiène S. (eds). The Mediterranean Region under Climate Change: A Scientific Update (pp. 219–228). Institut de Recherche pour le Développement, Marseille, France.
Navarro-Ortega A., Acuña V., Batalla R. J., Blasco J., Conde C., Elorza F. J., et al. (2012). Assessing and forecasting the impacts of global change on Mediterranean rivers. The SCARCE Consolider project on Iberian basins. Environ. Sci. pollut. Res. 19, 918–933. doi: 10.1007/s11356-011-0566-5
Ofir E., Corrales X., Coll M., Heymans J. J., Goren M., Steenbeek J., et al. (2023). Evaluation of fisheries management policies in the alien species-rich Eastern Mediterranean under climate change. Front. Mar. Sci. 10, 1155480. doi: 10.3389/fmars.2023.1155480
Olivar M. P., Qulez G., and Emelianov M. (2003). Spatial and temporal distribution and abundance of European hake, Merluccius merluccius, eggs and larvae in the Catalan coast (NW Mediterranean). Fisheries Res. 60, 321–331. doi: 10.1016/S0165-7836(02)00139-X
Oliver P. (1993). Analysis of fluctuations observed in the trawl fleet landings of the Balearic Islands. Sci. Mar. 57, 219–222.
Oliver P. and Massutí E. (1995). “Biology and fisheries of western Mediterranean hake (M. merluccius),” in Hake: Biology, fisheries and markets (Springer Netherlands, Dordrecht), 181–202.
Ordines F., Lloret J., Tugores P., Manfredi C., Guijarro B., Jadaud A., et al. (2019). A new approach to recruitment overfishing diagnosis based on fish condition from survey data. Scientia marina 83, 223–233. doi: 10.3989/scimar.04950.03A
Palomera I., Olivar M. P., and Morales-Nin B. (2005). Larval development and growth of the European hake Merluccius merluccius in the northwestern Mediterranean. Scientia Marina 69, 251–258. doi: 10.3989/scimar.2005.69n2251
Palomera I., Olivar M. P., Salat J., Sabatés A., Coll M., García A., et al. (2007). Small pelagic fish in the NW Mediterranean Sea: an ecological review. Prog. Oceanography 74, 377–396. doi: 10.1016/j.pocean.2007.04.012
Panzeri D., Reale M., Cossarini G., Salon S., Carlucci R., Spedicato M. T., et al. (2024). Future distribution of demersal species in a warming Mediterranean sub-basin. Front. Mar. Sci. 11. doi: 10.3389/fmars.2024.1308325
Parras-Berrocal I. M., Vázquez R., Cabos W., Sein D. W., Álvarez O., Bruno M., et al. (2022). Surface and intermediate water changes triggering the future collapse of deep water formation in the North Western Mediterranean. Geophysical Research Letters 49, e2021GL095404. 10.1029/2021GL095404.
Pastor F., Valiente J. A., and Palau J. L. (2019). “Sea surface temperature in the Mediterranean: Trends and spatial patterns, (1982–2016),” in Meteorology and climatology of the Mediterranean and Black Seas (Springer International Publishing, Cham), 297–309.
Payne M. R., Barange M., Cheung W. W., MacKenzie B. R., Batchelder H. P., Cormon X., et al. (2016). Uncertainties in projecting climate-change impacts in marine ecosystems. ICES J. Mar. Sci. 73, 1272–1282. doi: 10.1093/icesjms/fsv231
Payne M. R., Kudahl M., Engelhard G. H., Peck M. A., and Pinnegar J. K. (2021). Climate risk to European fisheries and coastal communities. Proc. Natl. Acad. Sci. 118, e2018086118. doi: 10.1073/pnas.2018086118
Pecl G. T., Araújo M. B., Bell J. D., Blanchard J., Bonebrake T. C., Chen I. C., et al. (2017). Biodiversity redistribution under climate change: Impacts on ecosystems and human well-being. Science 355, eaai9214. doi: 10.1126/science.aai9214
Pinot J. M., López-Jurado J. L., and Riera M. (2002). The CANALES experiment, (1996-1998). Interannual, seasonal, and mesoscale variability of the circulation in the Balearic Channels. Prog. Oceanography 55, 335–370. doi: 10.1016/S0079-6611(02)00139-8
Piroddi C., Coll M., Liquete C., Macias D., Greer K., Buszowski J., et al. (2017). Historical changes of the Mediterranean Sea ecosystem: modelling the role and impact of primary productivity and fisheries changes over time. Sci. Rep. 7, 44491. doi: 10.1038/srep44491
Planque B., Fromentin J. M., Cury P., Drinkwater K. F., Jennings S., Perry R. I., et al. (2010). How does fishing alter marine populations and ecosystems sensitivity to climate? J. Mar. Syst. 79, 403–417. doi: 10.1016/j.jmarsys.2008.12.018
Quattrocchi F., Fiorentino F., Lauria V., and Garofalo G. (2020). The increasing temperature as driving force for spatial distribution patterns of Parapenaeus longirostris (Lucas 1846) in the Strait of Sicily (Central Mediterranean Sea). J. Sea Res. 158, 101871. doi: 10.1016/j.seares.2020.101871
Quetglas A., Guijarro B., Ordines F., and Massutí E. (2012). Stock boundaries for fisheries assessment and management in the Mediterranean: the Balearic Islands as a case study. Sci. Mar. 76 (1), 17–28. 10.3989/scimar.2012.76n1017.
Quetglas A., Ordines F., Hidalgo M., Monserrat S., Ruiz S., Amores Á., et al. (2013). Synchronous combined effects of fishing and climate within a demersal community. ICES J. Mar. Sci. 70, 319–328. doi: 10.1093/icesjms/fss181
Relini L. O., Papaconstantinou C., Jukic-Peladic S., Souplet A., de Sola L. G., Piccinetti C., et al. (2002). Distribution of the Mediterranean hake populations (Merluccius merluccius smiridus Rafinesque 1810) (Ostheichthyes: Gadiformes) based on six years monitoring by trawl-surveys: some implications for management. Scientia Marina 66, 21–38. doi: 10.3989/scimar.2002.66s221
Richardson A. J. and Schoeman D. S. (2004). Climate impact on plankton ecosystems in the Northeast Atlantic. Science 305, 1609–1612. doi: 10.1126/science.1100958
Richon C., Dutay J. C., Bopp L., Le Vu B., Orr J. C., Somot S., et al. (2019). Biogeochemical response of the Mediterranean Sea to the transient SRES-A2 climate change scenario. Biogeosciences 16, 135–165. doi: 10.5194/bg-16-135-2019
Rosenberg A. A. (2003). Managing to the margins: the overexploitation of fisheries. Front. Ecol. Environ. 1, 102–106. doi: 10.1890/1540-9295(2003)001[0102:MTTMTO]2.0.CO;2
Rueda L., Moranta J., Abelló P., Balbín R., Barberá C., de Puelles M. F., et al. (2014). Body condition of the deep water demersal resources at two adjacent oligotrophic areas of the western Mediterranean and the influence of the environmental features. J. Mar. Syst. 138, 194–202. doi: 10.1016/j.jmarsys.2013.11.018
Rutterford L. A., Simpson S. D., Jennings S., Johnson M. P., Blanchard J. L., Schön P. J., et al. (2015). Future fish distributions constrained by depth in warming seas. Nat. Climate Change 5, 569–573. doi: 10.1038/nclimate2607
Sbrana M., Viva C., and Belcari P. (2006). Fishery of the deep-water rose shrimp Parapenaeus longirostris (Lucas 1846) (Crustacea: Decapoda) in the northern Tyrrhenian sea (Western Mediterranean). Hydrobiologia 557, 135–144. doi: 10.1007/s10750-005-1316-9
Sbrana M., Viva C., Belcari P., and Sartor P. (2019). Spatial distribution and abundance of European hake (Merluccius merluccius) in the northern Tyrrhenian and Ligurian seas. Scientia Marina 83, 187–196. doi: 10.1007/s10750-005-1316-9
Ser-Giacomi E., Jordá-Sánchez G., Soto-Navarro J., Thomsen S., Mignot J., Sevault F., et al. (2020). Impact of climate change on surface stirring and transport in the Mediterranean Sea. Geophysical Res. Lett. 47, e2020GL089941. doi: 10.1029/2020GL089941
Sion L., Zupa W., Calculli C., Garofalo G., Hidalgo M., Jadaud A., et al. (2019). Spatial distribution pattern of European hake, Merluccius merluccius (Pisces: Merlucciidae), in the Mediterranean Sea. Scientia Marina 83, 21–32. doi: 10.3989/scimar.04988.12A
Sobrino I., Silva C., Sbrana M., and Kapiris K. (2005). A review of the biology and fisheries of the deep-water rose shrimp, Parapenaeus longirostris, in European Atlantic and Mediterranean waters. Crustaceana 78, 1153–1184.
Spedicato M. T., Massutí E., Mérigot B., Tserpes G., Jadaud A., and Relini G. (2019). The MEDITS trawl survey specifications in an ecosystem approach to fishery management. Sci. Mar. 83, 9–20. doi: 10.3989/scimar.04915.11X
Stenseth N. C. and Rouyer T. (2008). Destabilized fish stocks. Nature 452, 825–826. doi: 10.1038/452825a
Torres A. P., Dos Santos A., Alemany F., and Massutí E. (2013). Larval stages of crustacean species of interest for conservation and fishing exploitation in the western Mediterranean. Scientia Marina 77, 149–160. doi: 10.3989/scimar.03749.26D
van der Sleen P., Zuidema P. A., Morrongiello J., Ong J. L. J., Rykaczewski R. R., Sydeman W. J., et al. (2022). Interannual temperature variability is a principal driver of low-frequency fluctuations in marine fish populations. Commun. Biol. 5, 28. doi: 10.1038/s42003-021-02960-y
van Vuuren D. P., Edmonds J., Kainuma M., Riahi K., Thomson A., Hibbard K., et al. (2011). The representative concentration pathways: an overview. Climatic Change 109, 5–31. doi: 10.1007/s10584-011-0148-z
Vieira A. R., Dores S., Azevedo M., and Tanner S. E. (2020). Otolith increment width-based chronologies disclose temperature and density-dependent effects on demersal fish growth. ICES J. Mar. Sci. 77, 633–644. doi: 10.1093/icesjms/fsz243
Von Schuckmann K., Le Traon P. Y., Menna M., Martellucci R., Notarstefano G., Mauri E., et al. (2022). Copernicus Ocean state report, issue 6. J. Operational Oceanography 15, 1–220. doi: 10.1080/1755876X.2022.2095169
Wood S. N. (2006). On confidence intervals for generalized additive models based on penalized regression splines. Aust. New Z. J. Stat 48, 445–464. doi: 10.1111/j.1467-842X.2006.00450.x
Wood S. N. (2017). Generalized additive models: an introduction with R (New York (United States of America): CRC press).
Wood S.N. (2004). Stable and efficient multiple smoothing parameter estimation for generalized additive models. Journal of the American Statistical Association 99, 673–686. 10.1198/016214504000000980.
Zappa G., Shaffrey L. C., and Hodges K. I. (2013). The ability of CMIP5 models to simulate North Atlantic extratropical cyclones. J. Climate 26, 5379–5396. doi: 10.1175/JCLI-D-12-00501.1
Zeng C., Rotllant G., Giménez L., and Romano N. (2020). Effects of environmental conditions on larval growth and development. Dev. Biol. larval ecology Natural history Crustacea 7, 195–222. doi: 10.1093/oso/9780190648954.003.0007
Keywords: demersal stocks, population dynamics, projections, climate change, fisheries management, western Mediterranean
Citation: González-Andrés C, Guijarro B, Ramírez-Romero E and Massutí E (2025) Long term predictions on the population dynamics of two contrasting demersal fishing resources in the western Mediterranean. Front. Mar. Sci. 12:1587297. doi: 10.3389/fmars.2025.1587297
Received: 04 March 2025; Accepted: 06 October 2025;
Published: 06 November 2025.
Edited by:
Laura Recasens, Institute of Marine Sciences, Spanish National Research Council (ICM-CSIC), SpainReviewed by:
Taner Yildiz, Istanbul University, TürkiyePaolo Sartor, Centro Interuniversitario di Biologia Marina ed Ecologia Applicata “G. Bacci” di Livorno (CIBM), Italy
Copyright © 2025 González-Andrés, Guijarro, Ramírez-Romero and Massutí. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Cristina González-Andrés, Y3Jpc3RpbmEuZ29uemFsZXouY2FwODBAZ21haWwuY29t; Beatriz Guijarro, YmVhdHJpei5ndWlqYXJyb0BpZW8uY3NpYy5lcw==
†These authors have contributed equally to this work