 Verónica Iriarte*
Verónica Iriarte* Andreas Winter
Andreas Winter- Fisheries Department, Directorate of Natural Resources, Falkland Islands Government, Stanley, Falkland Islands
Seal exclusion devices (SED) fitted in trawls are recognized as the best technical measure to mitigate pinniped bycatch, but do not completely eliminate incidental mortality. In areas with high pinniped abundance and concentrated fishing effort, identifying cause of death of pinniped carcasses in different stages of decomposition is challenging, as the individuals could have died from a variety of causes. In the Falkland Islands, best practice to mitigate and monitor incidental mortality of South American fur seal (Arctocephalus australis) and South American sea lion (Otaria flavescens) in the bottom-trawl Patagonian squid (Doryteuthis gahi) fishery was introduced in 2017. From 2018 through 2023, best practice in the fishery included 100% dedicated observer coverage and the use of SEDs. We analyzed 29,812 trawls (5,748 without a SED and 24,054 with a SED) for pinniped bycatch (live SED escapes, live deck releases, and mortalities), corresponding catch and fishing effort, post-mortem gross examinations, and multimedia data in the Patagonian squid fishery. Mortalities in trawls without a SED included 45 drownings (fresh carcasses) and two unknown (carcasses in different stages of decomposition). Mortalities in trawls fitted with a SED included 53 drownings, 28 propeller strikes, and 26 unknown. Our results indicate that SEDs are efficient at reducing incidental mortality, although low levels of drownings continue during trawl hauling. These mortalities comprise mostly of males and are related to several operational variables and captain performance, which was found to be heterogenous in facilitating SED escapes. In addition, we link for the first time propeller strike mortality of South American otariids to a particular fishery, evidencing the existence of this type of cryptic mortality in offshore trawl fisheries in the Southwestern Atlantic. Since using SEDs, pinniped mortality levels in the Patagonian squid fishery have been negligible at their population levels. Results of this study can be used to work with captains to enhance best practice and contribute to the sustainable management of the fishery.
1 Introduction
Interactions of pinnipeds with fisheries have been widely documented (Wickens, 1995; Crespo et al., 1997; Reyes et al., 2013; Franco-Trecu et al., 2019a; Romero et al., 2024), with the accidental capture in fishing operations (bycatch), being the most common anthropogenic cause of pinniped incidental mortality (Read, 2008; Kovacs et al., 2012). Best practice for assessing and managing marine mammal bycatch includes the collection of incidental mortality and fishing effort data, estimating abundances of marine mammal populations, implementing mortality limits, and modifying the fishing gear (e.g., Wade, 1998; FAO, 2021; Moore et al., 2021; Wade et al., 2021; Good et al., 2024a).
Although fitting a seal exclusion device (SED) in trawls is recognized as the best technical measure to mitigate pinniped incidental mortality (Hamilton and Baker, 2015; FAO, 2021), it does not eliminate it (e.g., Lyle et al., 2016; Iriarte et al., 2020; Queirolo et al., 2025). Rough weather during fishing operations, loss of net tension during trawl maneuvering (i.e., shooting, hauling, and vessel turns), and entanglements/twists in the body of the trawl or the SED net extension, can block the SED escape passage (Allen et al., 2014; Santana-Garcon et al., 2018; Iriarte et al., 2020; Bonizzoni et al., 2022). Furthermore, hauling a trawl with a large catch may exert pressure and cause trauma on a trapped pinniped inside the SED net extension. Trauma caused by impact with the SED grid or other part of the trawl could also occur (Hamilton and Baker, 2015). Consequently, there has been a debate regarding SED efficiency and potential cryptic mortality of endangered species (Meyer et al., 2017; Roberts et al., 2018; Hamilton and Baker, 2019).
In areas with high pinniped abundance and concentrated fishing effort, identifying the cause of death of bycaught carcasses in different stages of decomposition is challenging, as the individual in question could have died from a variety of anthropogenic (drowning, vessel collision, contaminants, debris entanglement) or natural (parasitism, biotoxins, microbial infections) causes (Arbelo et al., 2013; Bernaldo de Quirós et al., 2018; Arregui et al., 2019).
South American fur seal (Arctocephalus australis, hereafter SAFS) and South American sea lion (Otaria flavescens, hereafter SASL) are the two otariid species from the Southwestern Atlantic. After the cessation of commercial exploitation in the 20th century, SAFS and SASL recovery has been continuous (Crespo, 2021; Grandi and Milano, 2024). This could be partially attributed to male-selective harvesting and their polygynous life history, with few males genetically contributing to their populations (e.g., Capozzo, 1995; Königson et al., 2013; Franco-Trecu et al., 2019b; Hindell et al., 2022). Following South Atlantic stocks’ genetic diversity and increasing trends, both species are globally listed as “Least Concern” by the International Union for the Conservation of Nature (IUCN) (Cárdenas-Alayza et al., 2016a, 2016b).
In the Falkland Islands, best practice to mitigate and monitor incidental mortality of SAFS and SASL in the bottom-trawl Patagonian squid (Doryteuthis gahi) fishery was introduced in 2017 (Iriarte et al., 2020). Best practice includes 100% dedicated observer coverage, the use of SEDs, discard management, and haul maneuvering to allow the safe escape of seals while the SED is still in the water (Iriarte et al., 2020). This paper analyzes six years (12 fishing seasons) of seal bycatch in the Patagonian squid fishery since SED implementation, from 2018 through 2023. We use data collected under the pinniped bycatch monitoring program aboard the fishery to assess SED efficiency and to disentangle cause of mortality in SED-fitted trawls. We analyze SAFS and SASL bycatch observed (i.e., live SED escapes, live deck releases, and mortalities) during trawls both fitted with SEDs and without SEDs, and corresponding catch and fishing effort data. We overlap the bycatch data with post-mortem gross examinations of fresh carcasses and multimedia (photos and video footage of carcasses, SED escaping individuals, live deck releases), to confirm causes of mortality. We demonstrate SED efficiency, and explore key variables during both SED live escapes and mortalities, to disentangle why drowned individuals were not able to escape through the SED hatch. Ultimately, we discuss the sustainability of the prevailing incidental mortality rate in the squid fishery.
2 Materials and methods
2.1 Bottom-trawl squid fishery
The Patagonian squid fishery is undertaken in separate first (February–April) and second (July–September) seasons each year, targeting the two annual spawning cohorts of D. gahi, comprising an overall annual fishing effort of up to 128 days (64 days per season). During the 2018–2023 study period, three of the fishing seasons were closed early for stock conservation: second season 2019, first season 2021, and second season 2023 (Skeljo and Winter, 2023). Participating vessels are factory stern trawlers with lengths overall ranging 64–100 m, using bottom trawls approximately 200 m long, and cod-ends with small-mesh liners. The fishery operates at depths between 90–300 m, at water temperatures of 6–10°C (first season) and 4.5–6°C (second season). The total fishing zone covers approximately 31,500 km2, extending from the north-northeast of East Falkland to the south of West Falkland (Figure 1). For management purposes, the fishing zone is divided into north and south sub-zones at latitude 52°S (Figure 1). Standard licensing allocation in this fishery is for 16 vessels at a time, but with temporary (mechanical repair) and permanent (fleet turnover) replacements, 21 different vessels operated in the fishery during the study period (Supplementary Table S1). Trawlers had between one and five different captains, although on all except two vessels more than 80% of fishing days were commanded by no more than two different captains. A total of 38 captains participated in the fishery throughout the study period, however 16 of them performed 86% of all trawls. Conversely, about a third of captains commanded more than one vessel (Supplementary Table S2). Following Falkland Islands Fisheries Department (FIFD) fishing license conditions, SEDs are fitted between the body of the trawl and the cod-end. Three SED configurations are authorized for use in the squid fishery, referred to as type B (inclusion of a mesh panel), type C (an internal funnel), and type A (neither) (see SED diagrams in Iriarte et al., 2020). Minimum escape hatch dimensions are identical for all three SED models. Captains can switch SED types at their discretion, but for the study period on average vessels used just one SED model for more than 80% of all their trawls. SED compliance is verified by FIFD fishery officers during port and at-sea inspections. Starting from 2018, compulsory SED use was triggered in either the north or south fishing sub-zones following two pinniped mortalities within a sub-zone. Starting from second season of 2021, SED use was compulsory at the onset of all second seasons, in both sub-zones. Starting from first season of 2023, SED use became compulsory at the onset of all first seasons. Further best practice included in the fishing license conditions mandate facilitating the escape of trapped seals during trawl hauling while the SED is still in the water, prohibiting discarding during all trawl maneuvers (i.e., shooting, hauling, turning), and mandatory net cleaning before shooting the trawl (Iriarte et al., 2020). Since January 2021 the squid fleet is fully equipped with discard storage tanks, their capacity ranging from five to 30 metric tons (t). To reduce net cleaning time between a haul and the following shoot, most vessels alternate the use of two trawls, with thorough net cleaning being carried out during trawling.
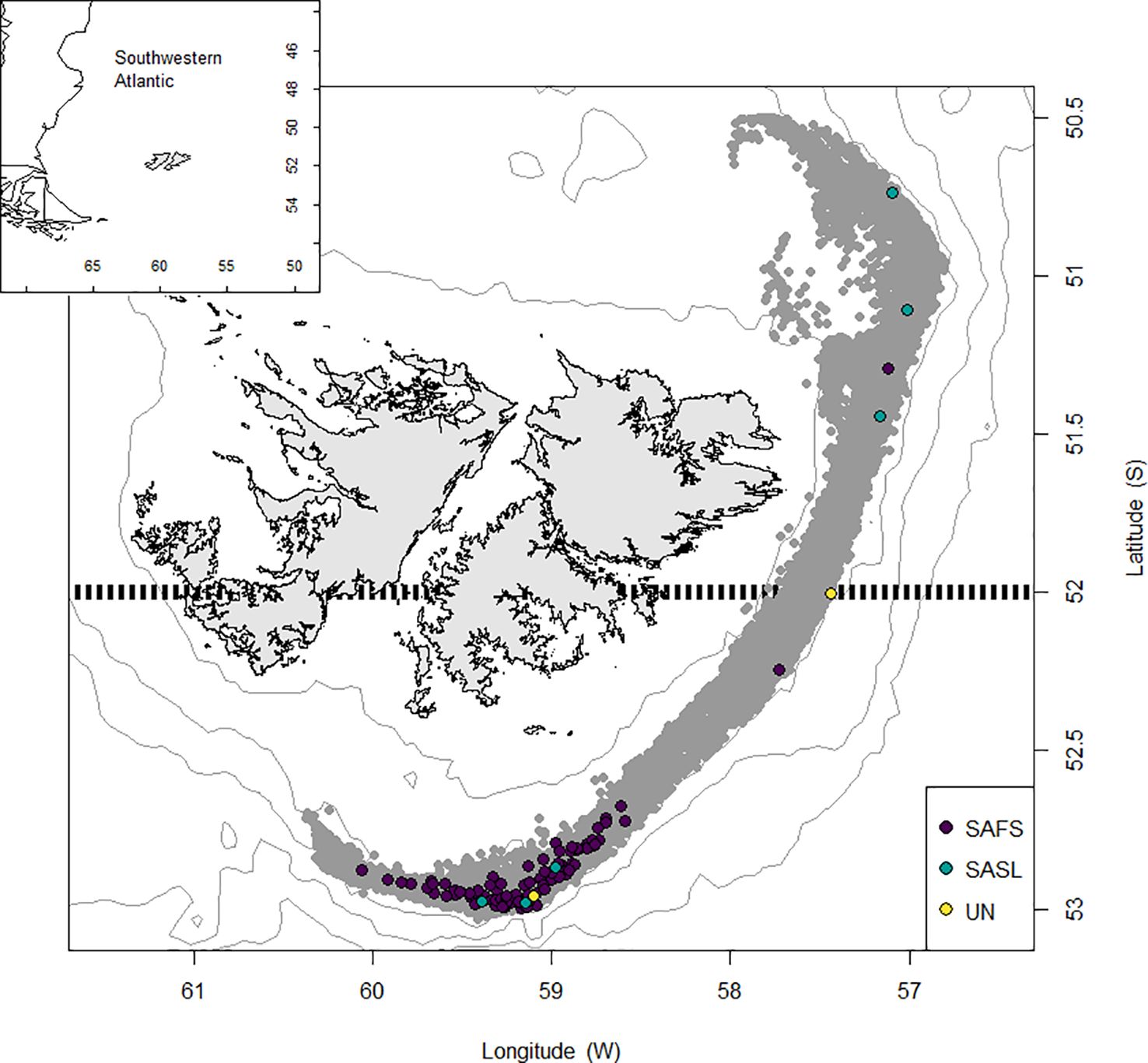
Figure 1. Distribution of observed D. gahi-target trawls using SEDs over the period 2018–2023. Grey circles: all trawls, indigo: South American fur seal (SAFS) mortalities, green: South American sea lion (SASL) mortalities, yellow: unknown species mortality. Broken horizontal line: the 52°S latitude parallel which divides north and south management sub-areas of the D. gahi fishery.
2.1.1 Fishery monitoring
Fishing information per trawl, reported by the vessel and by the observers, includes shoot and haul coordinate positions, maneuvering times, average trawl speed, Beaufort wind scale measure, and catch weight of trawls in kg. Vessels also record depth and their coordinate positions when changing direction, enabling calculation of the number of turns performed during trawls. Captains perform turns to return to large squid masses detected by their sensors. In rough weather, hauls may also be performed on a turn to maintain deck crew safety. During turns vessel speed is reduced to a minimum, trawl doors are hauled up to the surface (while the net remains midwater), and the vessel’s bearing is changed. Turns cause trawl meshes to lose tension, which could result in the blockage of the SED escape passage (Allen et al., 2014; Iriarte et al., 2020; Bonizzoni et al., 2022).
2.2 Pinniped bycatch monitoring program
During the study period, a total of 91 observers participated in at least one fishing season; however, the core observer team included 14 people, who covered 47% of the seasons (ranging from four to 10). Observer effort comprised of monitoring at least three trawls per day (both shooting and hauling) from the gantry and recording on each trawl maneuver observed, the numbers of attending seals (i.e., individuals who actively approach the vessel to be present during trawl maneuvering), species, behavior, interactions with the fishing gear/vessel, live SED escapes, deck releases, mortalities (i.e., carcasses in any stage of decomposition), and performing carcass marking to avoid recounts. Recording carcasses in decomposition ensured that cryptic mortalities were accounted for. Based on the ad libitum method for behavioral sampling which records all the observed behaviors (Altmann, 1974), a pinniped ethogram containing the full behavioral repertoire of both SAFS and SASL while attending vessels was developed. During trawl hauling observers carried out group scan sampling (Mann, 1999), taking a behavioral snapshot at the moment of maximum seal activity (i.e., whenever the SED and cod-end were on the surface), recording the number of individuals per species directly interacting with the trawl during the maneuver (i.e., foraging around the net, eating from the net, and eating on top of the net). Although observers also collected data during trawling and turning, these observations were opportunistic. On average, observers monitored 99% of the trawls carried out in at least one trawl maneuver (i.e., shoot, haul). Approximately 80% of trawls were monitored from the gantry. Gantry observation is particularly critical at hauls, as this position allows to best estimate the total number of attending seals, to record their surface behavior, and to efficiently see any trapped individuals, SED escapes, and mortalities. However, for safety reasons, gantry observations were not always practical during poor weather conditions or absence of daylight, and were then replaced by observation from the upper deck, bridge, or bridge wings. From second season 2019 to second season 2022, mortalities were immediately reported through instant messaging to one of us (VI), providing photographic evidence with close-ups views of eyes, nostrils, and genitalia. This allowed to identify the species, stage of decomposition, signs of cause of mortality, and sex. In addition, photographic evidence of carcass marking (i.e., cutting a portion/whole left pectoral fin) and total length measurement of fresh carcasses became a requirement. This immediate reporting schedule ensured rapid SED implementation, good data quality, and the opportunity to selectively preserve fresh carcasses for necropsy, including of species that do not attend squid fishing operations (i.e., southern elephant seal Mirounga leonina). The Falkland Islands are a hotspot for yearlings of this species, who are vulnerable to bycatch and can be incidentally caught during trawling (e.g., Brown et al., 2014; Barcenas-De La Cruz et al., 2017; Bernaldo de Quirós et al., 2018). Female carcasses were always preserved because they have a higher reproductive value and their mortality can cause greater impact at the population level (Crespo et al., 1997; Wallace et al., 2008; Riet-Sapriza et al., 2013; Hall, 2015; Hamilton and Baker, 2019). Males were selected for preservation based on the number of concurring mortalities reported within the fleet or by vessels of particular concern. Following biosecurity measures for zoonotic avian influenza (H5N1), no carcasses were collected after first season 2022.
2.3 Pinniped incidental mortality
2.3.1 Identification of cause of mortality
2.3.1.1 Peracute underwater entrapment
Drowning of marine mammals in fishing gear consists of a peracute (brief and intense) underwater entrapment (PUE) (Moore et al., 2013; Epple et al., 2020). Signs consistent with PUE include whitish/blood-tinged froth in airways, heavy reddened lungs and eyes, pulmonary edema, fractures, edema/hemorrhage in more than one organ, and generalized congestion (Moore et al., 2013; Bernaldo de Quirós et al., 2018; Epple et al., 2020). The diagnosis of PUE can usually be made on gross examinations (Moore et al., 2013).
2.3.1.2 Vessel collision sharp trauma
Sharp trauma induced by vessel collisions with marine mammals include blunt-force impacts with a non-rotating feature of the vessel (e.g., hull, rudder, skeg), and sharp-force injuries such as incising and/or chopping wounds caused by the rotating propeller (Byard et al., 2012a; Moore et al., 2013). Traumatic lethal amputations may involve body portions or entire appendages, trunk, or head (Byard et al., 2012b; Arbelo et al., 2013; Moore et al., 2013).
2.3.2 Post-mortem gross examinations
All collected carcasses were identified with a waterproof label stating vessel call sign, date, and observer station number, placed inside the animal’s mouth. The carcass was then covered by plastic film and a secondary adhesive label (with the same information) placed on top. All carcasses were preserved in the fishing vessel’s hold, at a temperature lower than -18°C. Frozen carcasses were either brought ashore during vessel opportunistic port visits or at the end of the fishing season, and transported to a cold store operating at a temperature lower than -18°C. Post-mortem freezing period ranged from 4 to 77 days. Carcasses were weighed frozen and prior to necropsy, thawed for three days at room temperature (13–18°C). Carcasses were then examined and processed following standard marine mammal necropsy guidelines (i.e., Pugliares et al., 2007). Particular attention was given to health status and the presence of froth, blood and water in airways; lung condition (color, consistency, and presence of water, blood, and froth), and reproductive status if female (i.e., presence of a fetus or a corpus luteum, and presence of milk). The lung conditions described above were considered peri-mortem (Roe and Meynier, 2012), while hemorrhage from the nose/mouth and reddened eyes have not been definitively reported as post-mortem signs associated to the freezing-thawing process (Roe and Meynier, 2012; Roe et al., 2012). Photographs of all relevant organs were taken and shared with a veterinarian to confirm cause-of-death diagnosis. Age category classifications (i.e., juvenile, sub-adult, adult) were based on published total body length measurements at sex maturity: minimum 104 cm for female SAFS and 154 cm for male SAFS (Batallés et al., 1990; Capozzo, 1995; Ponce de León and Pin, 2006), mean 147 cm for female SASL and 212 cm for male SASL (Grandi et al., 2010).
2.3.3 Bycatch multimedia analysis
All information from first-hand observations, deployment reports, photographs and necropsies, were compiled in an electronic multimedia databank. The information was then used to add relevant details to the bycatch records of each mortality event (i.e., species, sex, stage of decomposition, age class (if total length available and/or secondary sexual characteristics visible), presence of froth exuding from nostrils/mouth, reddened eyes, carcass marked). From available evidence, cause of mortality was assigned as either PUE or vessel collision sharp trauma. Because decomposition in deep seawater takes longer than on land (Di Maio and Di Maio, 2001; Caruso, 2016), and the fact that recovered carcasses exhibited varying degrees of decomposition that could correspond to “fresh condition” (standard decomposition Code 1 in Geraci and Lounsbury, 1993), decomposition codes established for stranded marine mammals (Geraci and Lounsbury, 1993), could not be applied. Instead, based on eye features, facial appearance, and general status of the carcasses, stage of decomposition was defined as fresh (lubricated eye, black eyelids), in decomposition (dry/blurry eyes, white eyelids), or in advanced decomposition (absence of eyes, missing skin and fur) (Figure 2). In addition, photos of marked carcasses were matched to identify recaptures. SED escapes and deck releases were video-recorded whenever possible, and these video data were also reviewed to assess animal health status based on physical characteristics and behavior. All multimedia information was cross-checked against the raw observer data (Supplementary Table S3) and corrections were applied as necessary (Supplementary Table S4).
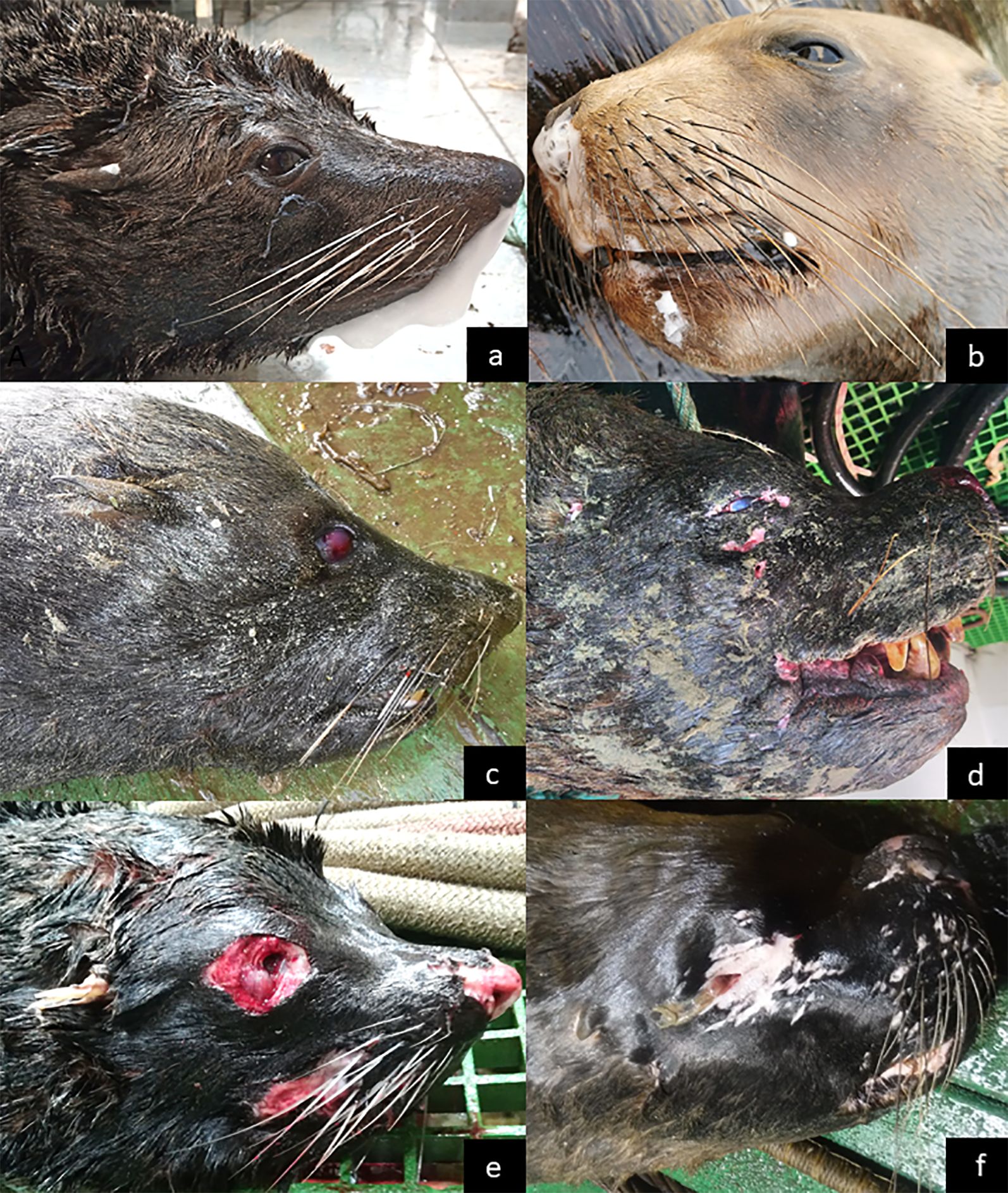
Figure 2. Decomposition stages: SAFS (left) and SASL (right). (a, b) Fresh: lubricated eye, black eyelids, froth exuding from nostrils and mouth. (c, d) In decomposition: dry/blurry eyes, white eyelids. (e, f) Advanced decomposition: absence of eyes, missing skin and fur.
2.4 Data exploration and preliminary statistical analysis
After data-cleaning erroneous or unverifiable entries, 29,812 observed trawls were retained, representing about 97% of all commercial squid trawls carried out for the study period 2018–2023. The biggest data collection deficit (only 88% of trawls observed) occurred during second season 2020, when COVID-19 regulations prevented observers from embarking for the first week of the season (Winter, 2020).
2.4.1 Pinniped attendance per fishing season
Because either fishing season targets different D. gahi cohorts (Arkhipkin et al., 2008), a first preliminary analysis was conducted to evaluate whether pinniped attendance to squid fishing vessel operations categorically differed between first and second fishing seasons.
Numbers of seals sighted and/or interacting with vessels were used as a proxy for attendance, sighting being defined as the maximum number of seals recorded within a trawl (i.e., in any of the observed maneuvers). A total of 55,232 seals were sighted interacting with vessels (Table 1), from about 40% of the 29,812 observed trawls. Each trawl was assigned a consecutive index integer, 1 to 29,812. To evaluate differences between first and second seasons, n = 55,232 integers (equal to the total number of observed seals, Table 1), were randomly sampled with replacement from the index and recorded as corresponding to either the first or second season. Random resampling was iterated 10,000×. Absolute differences between first season sightings per trawl and second season sightings per trawl were then calculated for the set of 10,000 iterations. The proportion of these resampled absolute differences that was greater than the observed absolute difference, represented the significance of difference between first and second fishing seasons’ sightings. Observed total sightings were 14,025/15,137 = 0.93 sightings per trawl in first seasons, versus 41,207/14,675 = 2.81 sightings per trawl in second seasons (Table 1). That difference measured by randomization was statistically significant (P < 0.001), confirming previous observations of higher pinniped attendance in second seasons (Iriarte et al., 2020; Iriarte, 2022). All subsequent analyses were therefore based on computing first and second fishing seasons separately.
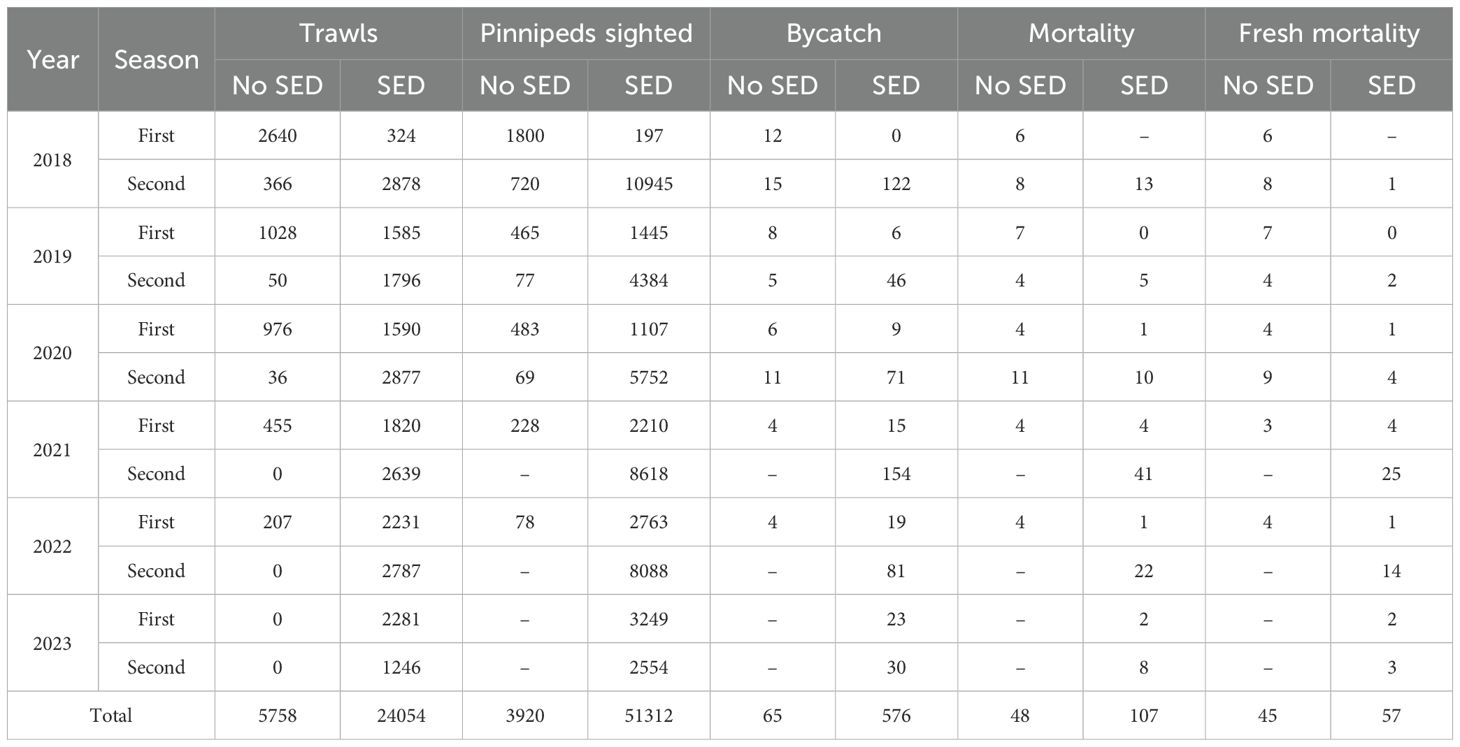
Table 1. Numbers of observed trawls, pinnipeds sighted, total bycatch, total mortalities and fresh mortalities; by year, season, and no SED versus SED.
2.4.2 Pinniped incidental mortality as a function of SED use
A second preliminary analysis was conducted to evaluate whether pinniped mortalities differed between trawls carried out with or without a SED fitted in the net. Only fresh mortalities were evaluated, as cause of mortality was confirmed and could be attributed to specific trawls from particular fishing vessels. A total of 102 fresh seal mortalities were recorded (Table 1), from 86 of the 29,812 observed trawls. Given the relatively low number of seal mortalities, Monte Carlo algorithms were used to evaluate differences. To evaluate fresh mortality differences between trawls fitted with or without SEDs, the same approach applied to pinniped attendance per fishing season was used, except that SED versus non-SED resamples were now randomized separately within first and second seasons, and proportions were based only on numbers of trawls with > 0 seal sightings. Differences by randomization obtained total fresh mortalities observed significantly higher (P < 0.001) in trawls without a SED than in trawls fitted with a SED in both first (n-SED: 0.019 mortalities per trawl with > 0 sightings, SED: 0.002 mortalities per trawl with > 0 sightings) and second fishing seasons (n-SED: 0.128, SED: 0.007 mortalities per trawl with > 0 sightings). Further analyses were therefore conducted only on trawls fitted with a SED, as its efficiency was demonstrated, and the main purpose of this study was to explore variables which could explain fresh mortalities recorded in SED-fitted trawls.
2.4.3 Pinniped incidental mortality as a function of depth
A third preliminary analysis was conducted to evaluate whether sea bottom depth was a factor in pinniped mortality, potentially confounding effects particular to the trawls. Total numbers of PUEs in first and second seasons, among trawls fitted with a SED, were computed as generalized additive models (GAM) with variables sea depth and haul latitude (latitude strongly correlates with proximity to the pinniped breeding areas). For both first and second seasons, sea bottom depth included in the GAM was not statistically significant, and was therefore not considered in further analyses.
2.5 Principal statistical analyses
Continuous explanatory variables trawl duration, average trawl speed, trawl catch, number of turns performed, and Beaufort scale were evaluated using GAM with package ‘mgcv’ (Wood, 2023) in R statistical software (R Core Team, 2022).
PUE and fresh propeller sharp trauma mortalities were modelled separately, as they represent a different outcome of interacting seals with the fishing gear/vessel. SED escapes and mortalities were both modelled as absolute numbers per trawl, to encompass the total encounter of pinnipeds with vessels, and as numbers per seals sighted attending a trawl, to encompass more specifically the proportional effect of a vessel when encountered. GAMs for SED escapes and mortalities per trawl were fitted using a Poisson distribution, and GAMs for SED escapes and mortalities per sightings, being fractional values, were fitted using a Tweedie distribution, suitable for zero-inflated data (Shono, 2008). Continuous variables were scaled by subtracting the mean and dividing by the standard deviation. Continuous variables were then ranked and added by forward stepwise selection to the final GAM if they improved pseudo-R2 (1 – fitted deviance/null deviance; Swartzman et al., 1992) by at least 1%.
Categorical variables vessel and captain were examined separately from the continuous variables, as they were too numerous (21 different vessels, 38 different captains) to include effectively in GAMs. Because of the high correspondence between vessel and captain, the analysis was focused only on captains, given that operational decisions and maneuvering style are known to influence marine mammal bycatch and its outcome (Hall et al., 2000, 2003; Hall, 2015). Monte Carlo algorithms were again used to randomly reassign PUE and propeller sharp trauma fresh mortalities in first and second fishing seasons, and calculate which captains had mortality numbers greater or lesser than the 95% quantiles of 10,000 random iterations. In addition, post-release mortalities, SED escapes, and total bycatch including only PUE mortalities were evaluated, as these can be affected by the operation of the captain. Mortality rates and bycatch rates (bycatch per sighting) were compared between first and second seasons for captains that worked on both fishing seasons, using a bootstrap Spearman’s rank test (Curran, 2015) to weight differences in trawl numbers among captains.
Because of the high correspondence between vessels, captains, and the SED types that they used, fresh mortality differences between SEDs were analyzed case-by-case whenever one captain, on the same vessel, used more than one SED type, within either first or second fishing seasons. Mortalities were restricted to PUE mortalities, and were analyzed using Kruskal-Wallis tests.
3 Results
3.1 Pinniped bycatch
Of the total trawls carried out in the period 2018–2023, 24,054 were performed with a SED and 5,748 without a SED, with 55,232 seals observed from the gantry directly interacting with the fishing gear. Total seal bycatch for the study period included 641 individuals, of which 155 were mortalities, comprised of carcasses in different stages of decomposition (Table 1). Bycatch in trawls without a SED comprised of 65 individuals (16 live deck releases, 48 mortalities, one recaptured marked carcass) (Table 1). Stage of decomposition of carcasses recorded in trawls without a SED included 45 fresh individuals (all PUE mortalities), two in decomposition, and one in unknown stage of decomposition. Bycatch in trawls fitted with a SED included 576 individuals (351 SED live escapes, 104 live deck releases, 107 mortalities, 14 recaptured carcasses) (Table 1; Supplementary Figure S1; Supplementary Table S3). Of the carcasses recorded in trawls fitted with a SED, 53 exhibited PUE signs (Figure 3; Supplementary Table S5), 28 had injuries which corresponded to vessel collision sharp trauma (Figure 4; Supplementary Table S6), while 26 could not be allocated to either of the previous categories, being cause of mortality unknown (Figure 5; Supplementary Table S7). All SED mortalities attributed to PUE comprised of fresh carcasses, while mortalities attributed to vessel collision sharp trauma included one direct observation of the collision, three fresh carcasses, 22 in decomposition, and two in advanced decomposition (Figure 5; Supplementary Tables S5, S6).
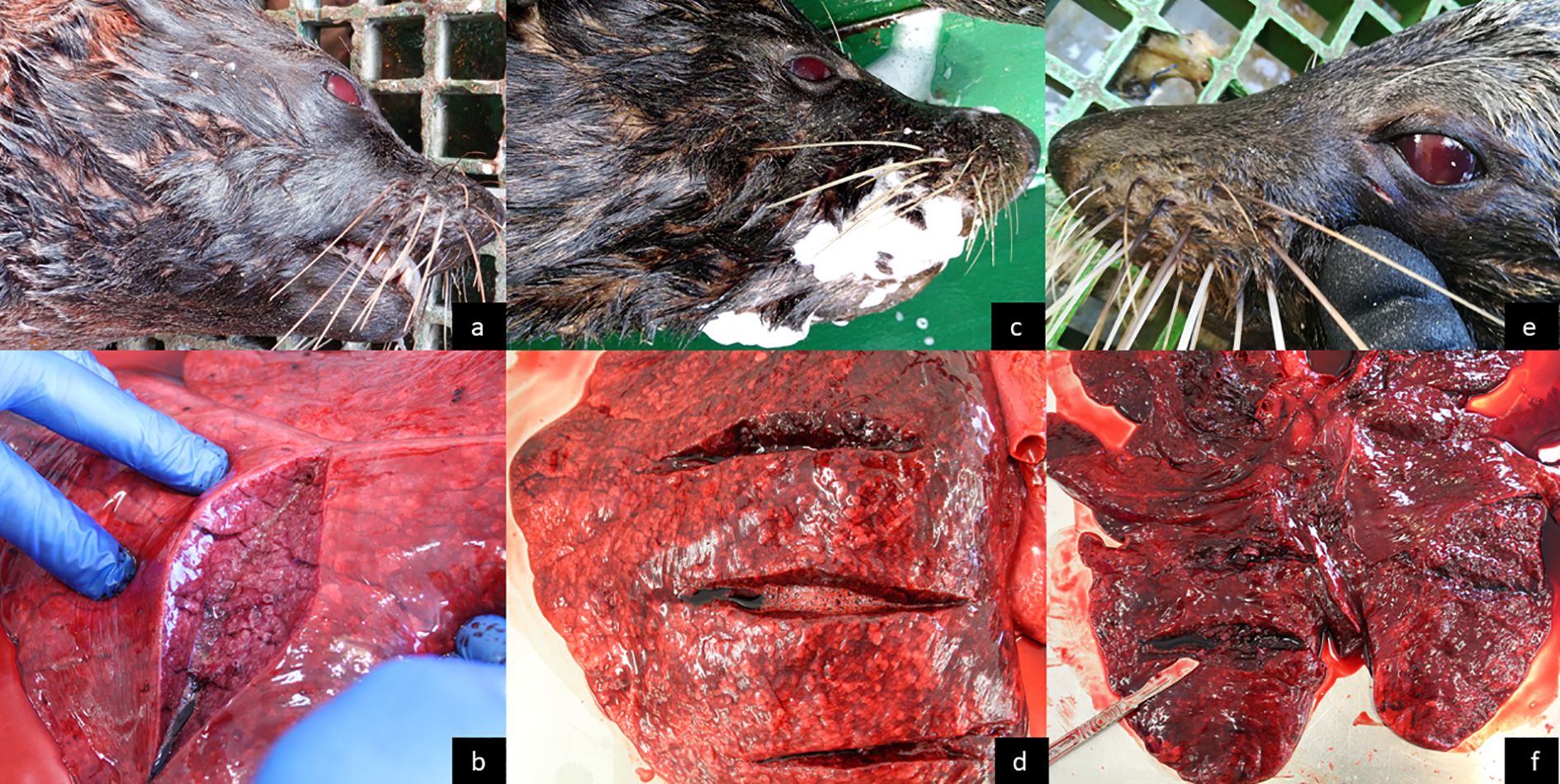
Figure 3. SAFS gross post-mortem examinations and different PUE syndrome lung appearances. (a, b) Lactating female; reddened eyes and moderate pulmonary hemorrhage with edema. (c, d) Adult male; froth exuding from nostrils and mouth, reddened eyes (changed from brown to red after ~ 50 min), severe hemorrhagic lungs with edema. (e, f) Adult female; reddened eyes, massive pulmonary hemorrhage, and consolidated lungs.

Figure 4. Vessel collision sharp trauma. (a) SASL adult female with incised wound; carcass in decomposition. Tip of left pectoral missing due to carcass marking. (b) SAFS adult male with two incised wounds; carcass in decomposition. (c, d) SAFS adult male with chopped body. Carcass in decomposition, note white eyelids. (e) SAFS adult male with two incised wounds. Fresh carcass, note dark lubricated eye and black eyelids.
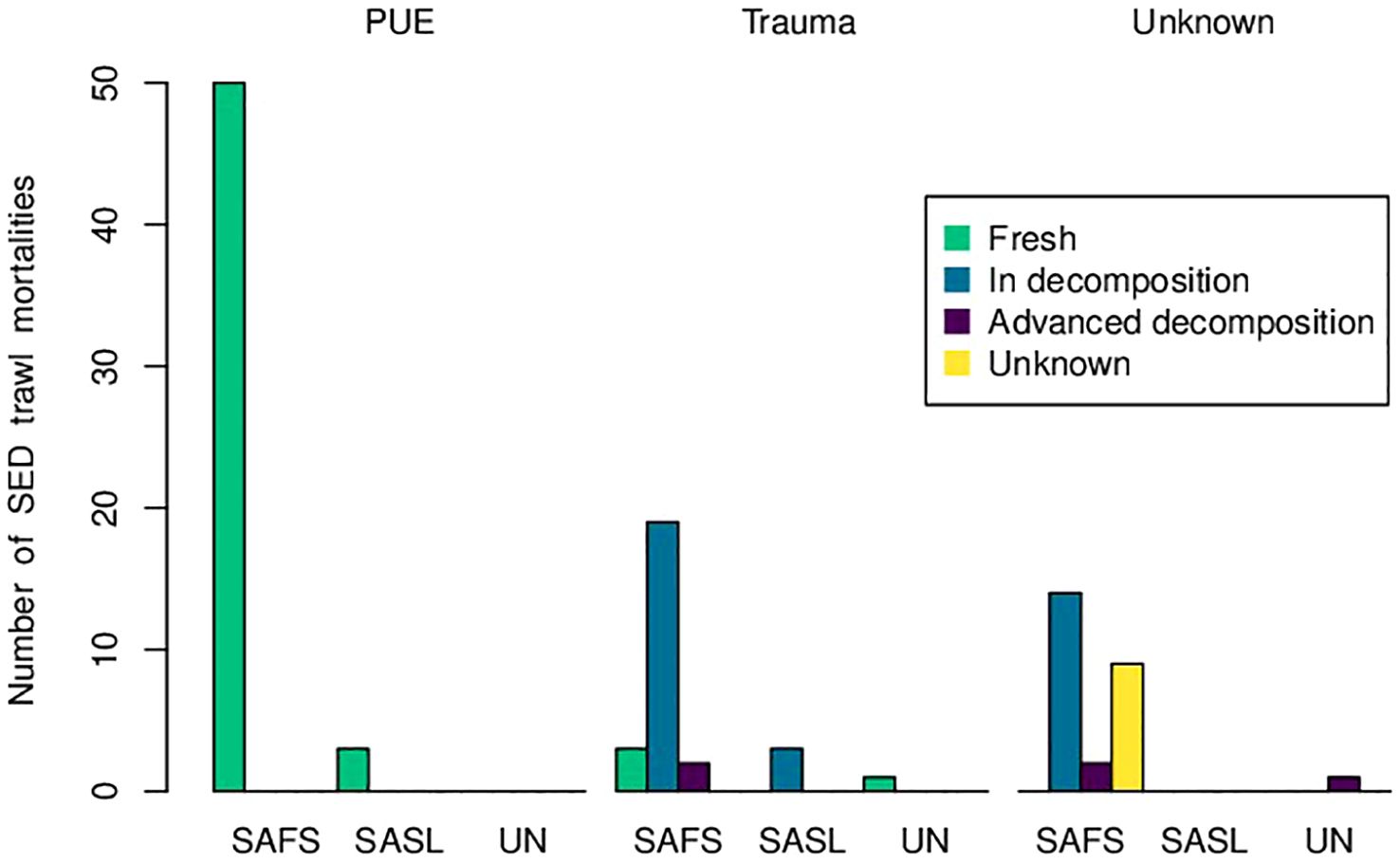
Figure 5. Pinniped mortality numbers by cause and carcass condition, in trawls fitted with a SED.
3.2 SED live escapes
Of the 351 total SED escapes recorded in this study, 350 were observed at the surface of the water during trawl hauling (53 in first seasons, 298 in second seasons). One SASL was observed inside the body of the trawl during a shoot. Based on previous observations (Iriarte et al., 2020), it is assumed to have safely escaped through the SED (Supplementary Table S4).
During first seasons trawl duration had a significant (P < 0.05) positive relationship with the number of SED escapes per trawl, over the trawling interval of about 2.8 to 5.2 hours (h) (Table 2; Figure 6, top row).
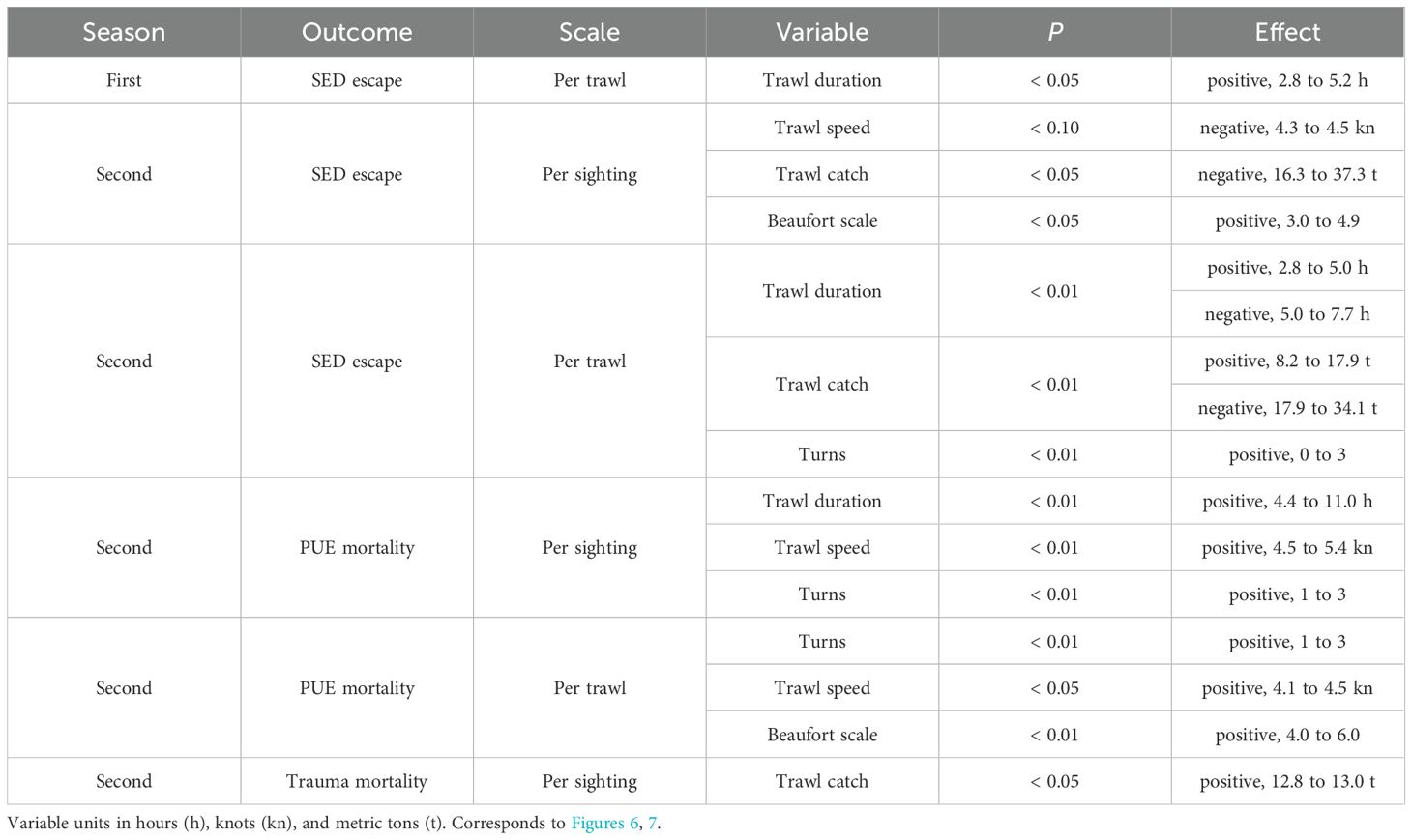
Table 2. Summary of statistically significant variables predicting numbers of SED escapes and fresh mortalities from GAMs, by fishing season.
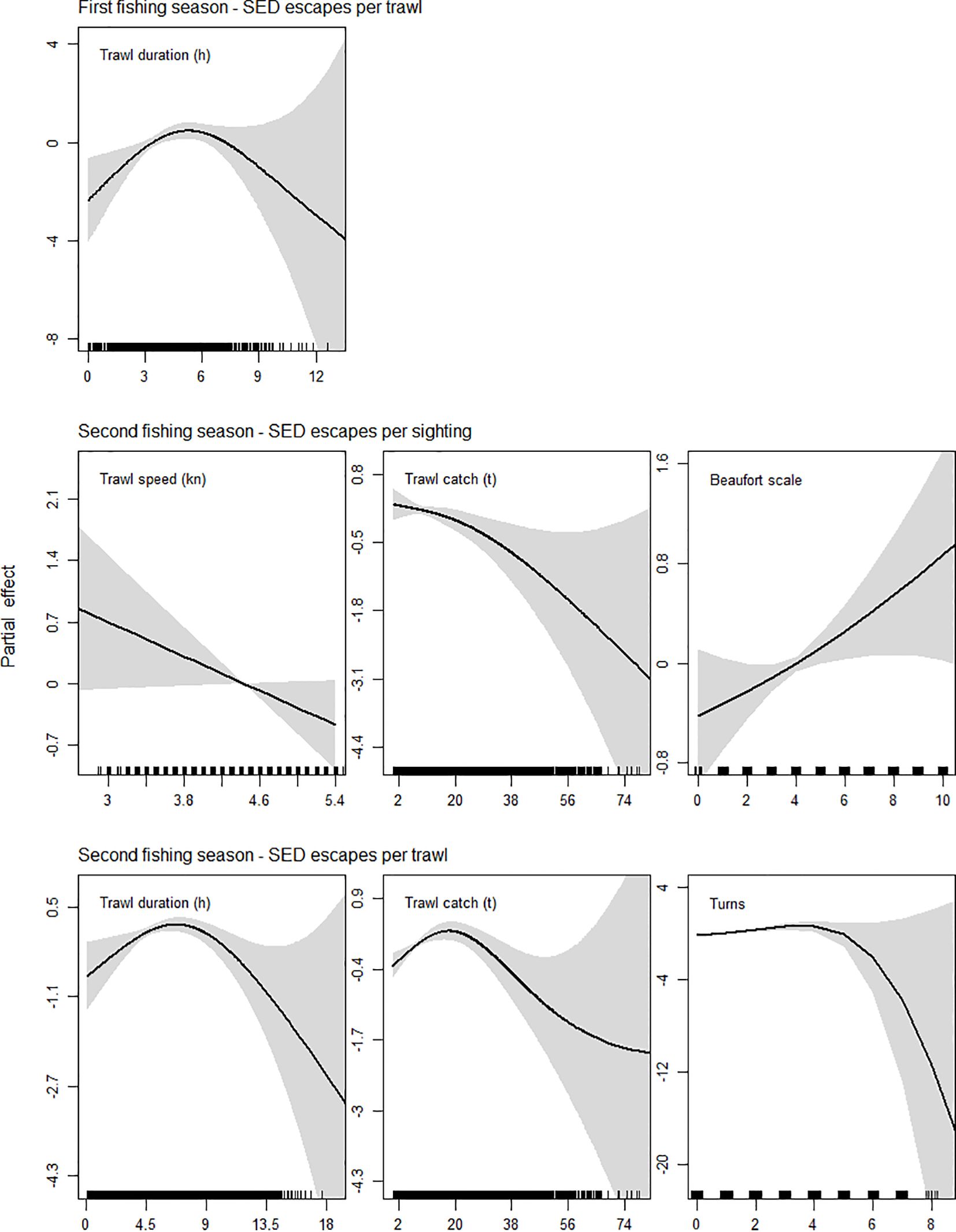
Figure 6. GAM smooth partial effect plots of significant explanatory variables for SED escapes as a proportion of sightings per trawl, and by total numbers per trawl, in first and second fishing seasons. Variable units in hours (h), knots (kn), and metric tons (t). Black lines: model fit, grey shading: 95% confidence interval.
During second seasons SED escapes per sighting were significantly negatively related to trawl speed around 4.4 knots (kn), negatively related to trawl catch weight between 16.3 and 37.3 t, and positively related to the Beaufort scale between 3 and 4.9 (Table 2; Figure 6, middle row). SED escapes per trawl were positively related to trawl duration between 2.8 and 5 h, negatively related to trawl duration between 5 and 7.7 h, positively related to trawl catches from 8.2 to 17.9 t, negatively related to trawl catch weights from 17.9 to 34.1 t, and positively related to to the performance of up to three turns (Table 2; Figure 6, bottom row).
3.3 SED fresh mortalities
A total of 57 pinniped fresh mortalities (53 PUE, four propeller sharp trauma) were recorded in 24,054 SED trawls, of which eight occurred in first seasons and 49 in second seasons (Table 1; Supplementary Table S4). On two PUE mortalities, eye color change from brown to red was recorded occurring approximately 50 min after being recovered from the trawl (Figure 3c; Supplementary Table S4). Based on the recorded timing and personal at-sea observations from one of us (VI), we concluded lubricated brown eyes to be indicative of a short time lapse since death (i.e., < 1 h). Through the bycatch multimedia analysis, we were able to confirm that 40 out of 53 PUE mortalities had lubricated brown eyes (Figures 2a, b; Supplementary Table S5). In addition, the bycatch multimedia analyses evidenced that only nine out of 53 PUE mortalities had reddened eyes (Figures 3a, c, e), with seven of them occurring in trawls with one to four turns and two recovered from trawls without any turn (Supplementary Table S5). Of the total pinniped fresh mortalities, 53 were SAFS, three SASL, and one unknown species (the observer reporting presumably a SAFS). Regarding the SAFS mortalities, seven were categorized as post-release mortalities: one individual died on deck, while six were released alive (Supplementary Table S4). However, these six SAFS arrived aboard unconscious and with PUE signs. Although they were left to rest on deck for some time (max ~ 2 h), their symptoms included froth/water exuding from nostrils/mouth, difficulty breathing, mouth open and tongue exposed, and were judged irrecoverable (Supplementary Video S1). Concerning the underlying known causes of PUE of some individuals, two SAFS mortalities consisted of neck entanglements in the mouth/wing of the trawl meshes, one male juvenile SASL came entangled in a trawl door, while eight SAFS could not escape through the SED as a result of either a twisted trawl or a haul maneuver carried out with a simultaneous turn (Supplementary Tables S4, S5). Observed fresh mortalities were always species-segregated except for a single trawl (during second season 2022) that had both one SAFS and one SASL mortalities.
3.3.1 First fishing season
The eight pinniped SED fresh mortalities recorded during first seasons were all PUE mortalities, seven SAFS (all males: two juveniles, one sub-adult, four adults) and one SASL (adult female) (Supplementary Figure S2). No continuous variables were statistically significant in explaining either absolute mortality numbers or mortalities per sighting.
3.3.2 Second fishing season
The 49 pinniped SED fresh mortalities recorded during second seasons included 45 PUE and four propeller sharp trauma mortalities: 46 SAFS (39 males (10 juvenile, three sub-adults, 26 adults), five adult females, one juvenile of unknown sex, one individual of unknown sex and age class), two juvenile male SASL, and one unknown species/sex/age class (Table 1; Supplementary Figure S2; Supplementary Table S4).
3.3.2.1 PUE
Like SED escapes analyses, PUE mortality showed positive correlations with continuous variables, except trawl catch (Table 2; Figure 7). PUE mortalities per sighting were significantly positively (P < 0.01) related to trawl duration between 4.4 and 11 h, and positively related to trawl speed between 4.5 to 5.4 kn (Table 2; Figure 7, top row). PUE mortalities per trawl were positively (P < 0.01) related to the performance of 1–3 turns, positively related (P < 0.05) to trawl speed between 4.1 and 4.5 kn, and positively related (P < 0.01) to Beaufort scale between 4 and 6 (Table 2; Figure 7, middle row). Trawl duration was a significant explanatory variable for PUE mortalities per sighting, however it was not significant for PUE mortalities per trawl.
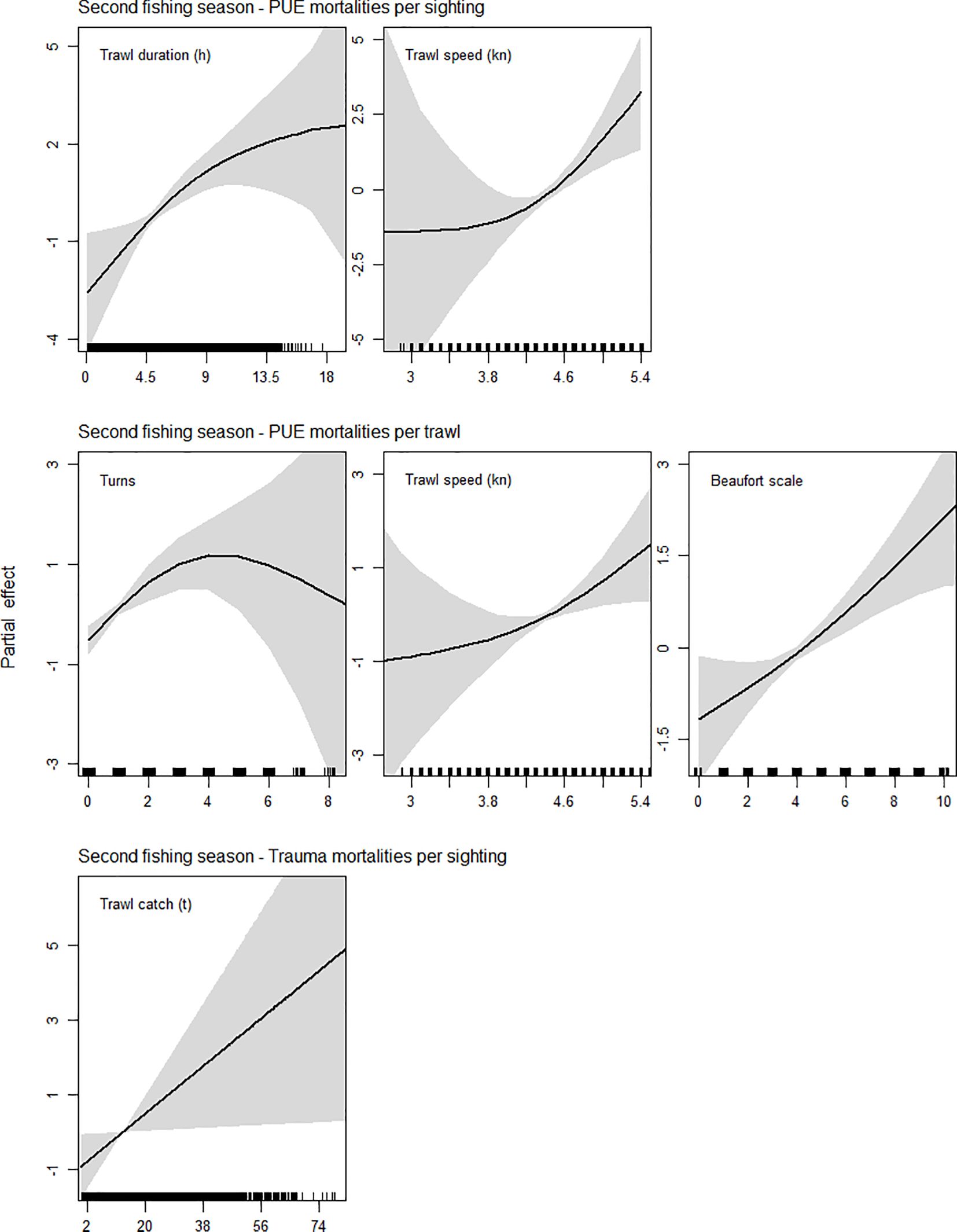
Figure 7. GAM smooth partial effect plots of significant explanatory variables for fresh mortalities as a proportion of sightings per trawl, and by total numbers per trawl, in second fishing seasons (none were significant in first seasons). Variable units in hours (h), knots (kn), and metric tons (t). Black lines: model fit, grey shading: 95% confidence interval.
3.3.2.2 Propeller sharp trauma
A total of four vessel collision sharp trauma fresh mortalities were recorded and quantitatively evaluated, one being directly observed from the gantry during trawling, and three comprising recovered carcasses. The fresh mortality observed from the gantry was an unknown species (presumably SAFS); strong vessel shuddering occurred and blood in the water was seen astern the vessel. However, in this study a proportionally high number of non-fresh trauma-injured seals was additionally recorded, indicating greater prevalence of propeller strike incidental mortality in the fishery (Supplementary Table S6). The propeller sharp trauma fresh mortalities per sighting were positively related (P < 0.05) to trawl catch weight, the statistically significant inflection occurring around 12.9 t (Table 2; Figure 7, bottom row).
3.4 Post-mortem gross examinations
Seventeen fresh frozen carcasses were examined, five originating from trawls without a SED and 12 from trawls with a SED (Supplementary Table S8). All carcasses exhibited good nutritional condition and signs in lungs, airways, and eyes, consistent with PUE. At the time of the post-mortems, different PUE lung appearances were noticed (Figures 3b, d, f), which could be related to different depths at death (Moore et al., 2013). Because in this study no underwater cameras were used, the exact time and depth at death of PUE individuals remains unknown. At the time of recovery from the SED-fitted trawl, nine of the carcasses had exhibited lubricated brown eyes and only three had reddened eyes (Figure 3; Supplementary Table S5), while by the time of post-mortem all had reddened eyes (Supplementary Table S8). As reddened eyes have been found diagnostic of PUE (Moore et al., 2013; Bernaldo de Quirós et al., 2018; Epple et al., 2020), disregarded a byproduct of freezing (Roe and Meynier, 2012), and the fact that eye color change was observed in situ in this study, we assumed the pre and post-mortem differences in eye color consist of a natural process of the carcass. PUE diagnosis in the examined individuals was supported by the lung and airway conditions observed during the post-mortems (Supplementary Table S8). In two SED carcasses, chronic active ulcerative gastritis associated with nematode infection was found (Supplementary Table S8).
3.5 SED mortalities in decomposition
Fifty mortalities in different stages of decomposition were recorded (36 in decomposition, five in advanced decomposition, nine in unknown stage of decomposition) (Figure 5; Supplementary Table S4). Of these, 24 corresponded to propeller sharp trauma, comprised of 19 male SAFS (18 adults, one juvenile), three SASL (two adult males, one adult female), and two unknown species/sex (Figure 5; Supplementary Table S6). All the mortalities in decomposition attributed to sharp trauma were exsanguinated and only one had reddened eyes. Injuries included incised wounds, chop wounds, and amputations of head and entire body sections (Figure 4; Supplementary Table S6). The remaining 26 of 50 mortalities in decomposition could not be assigned to any particular cause of mortality (Supplementary Table S7). Fourteen of these were male SAFS (two juveniles, 12 adults), 11 were SAFS of unknown sex (one juvenile, 10 unknown), while one corresponded to an unknown species and sex (Figure 5; Supplementary Table S7).
3.6 Captain effect on pinniped bycatch
Twenty-one of the captains worked both first and second seasons (Tables 3, 4). During first seasons, one captain had a significantly higher than random number of deck releases (Ca21, Table 3).
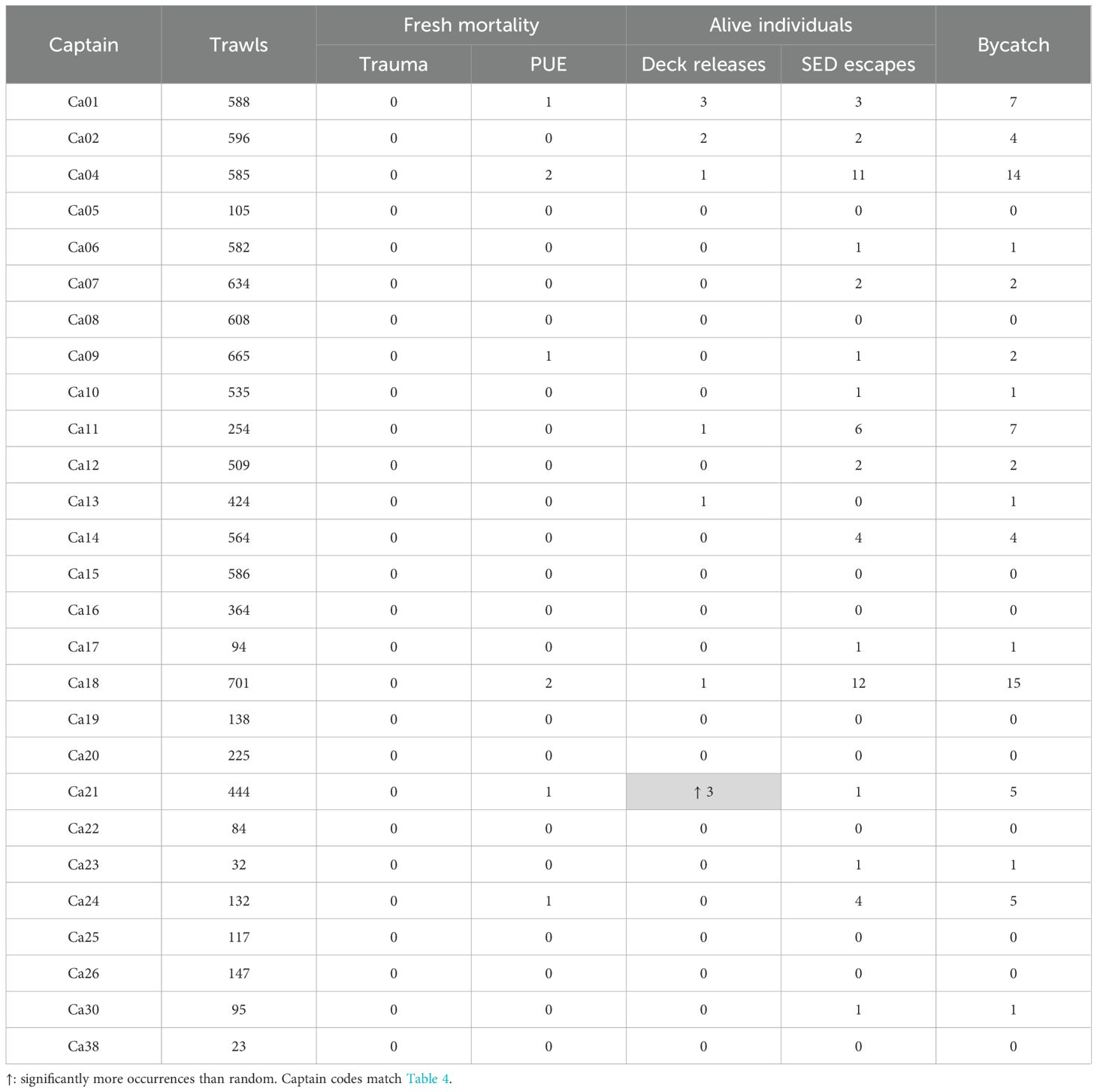
Table 3. Pinniped mortalities and live deck releases/SED escapes per captain during first seasons.
During second seasons, three captains had significantly more than random bycatch (Ca02, Ca04, Ca11, Table 4). The first of these captains (Ca02) also had significantly more than random live deck releases (Table 4). The second and third of these captains (Ca04, Ca11, Table 4), also had significantly more than random SED escapes. Another captain (Ca12, Table 4) had significantly fewer than random bycatch. Two different captains (Ca13, Ca37) had significantly more than random PUE mortalities, without correspondingly having significant numbers of total bycatch (Table 4).
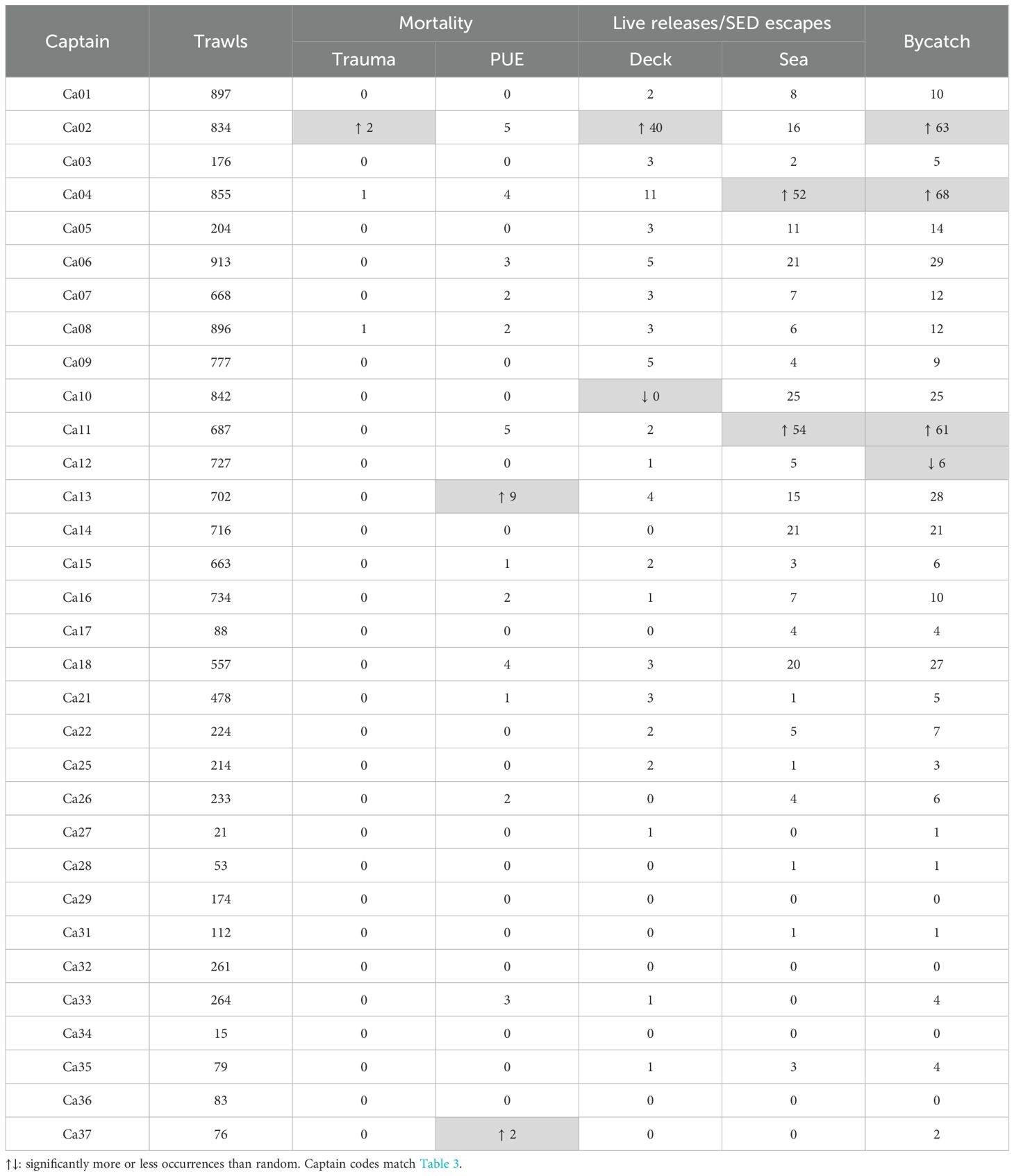
Table 4. Pinniped fresh mortalities and live deck releases/SED escapes per captain during second fishing seasons.
Overall pinniped bycatch or mortality rates per captain were not significantly correlated between first and second seasons (95% confidence interval of bootstrap Spearman’s rank coefficient ρ encompassing zero). Nevertheless, the three captains with significantly more than random bycatch in second seasons (Ca02, Ca04, Ca11, Table 4) also accounted for 34.2% of all pinniped captures in first seasons (25 of 73), versus 14.6% of all trawls in first seasons (1,435 of 9,831) (Table 3).
The captain that had significantly more than random deck releases (Ca02), also had significantly more than random fresh propeller mortalities (two out of four total) (Table 4).
3.7 SED effect on pinniped bycatch
Nineteen combinations of captain/vessel/season included the use of more than one SED model (Table 5). Fourteen of these combinations were not statistically comparable for fresh mortality differences between SEDs, as they included zero PUE mortalities. Mortality differences of the other five combinations (Ca01/Ve02/first season; Ca07/Ve07/second season; Ca08/Ve08/second season; Ca09/Ve09/first season; Ca13/Ve13/second season) were individually not statistically significant (Kruskal-Wallis test, P > 0.1). However, all five of these combinations obtained higher PUE mortalities in type B SEDs (one–two) than in type C SEDs (zero) (Table 5). The random probability of five binary outcomes being unidirectional, given the respective trawl numbers, was P < 0.05, suggesting marginal evidence that type B SEDs are less effective at facilitating the live escape of seals than type C SEDs.
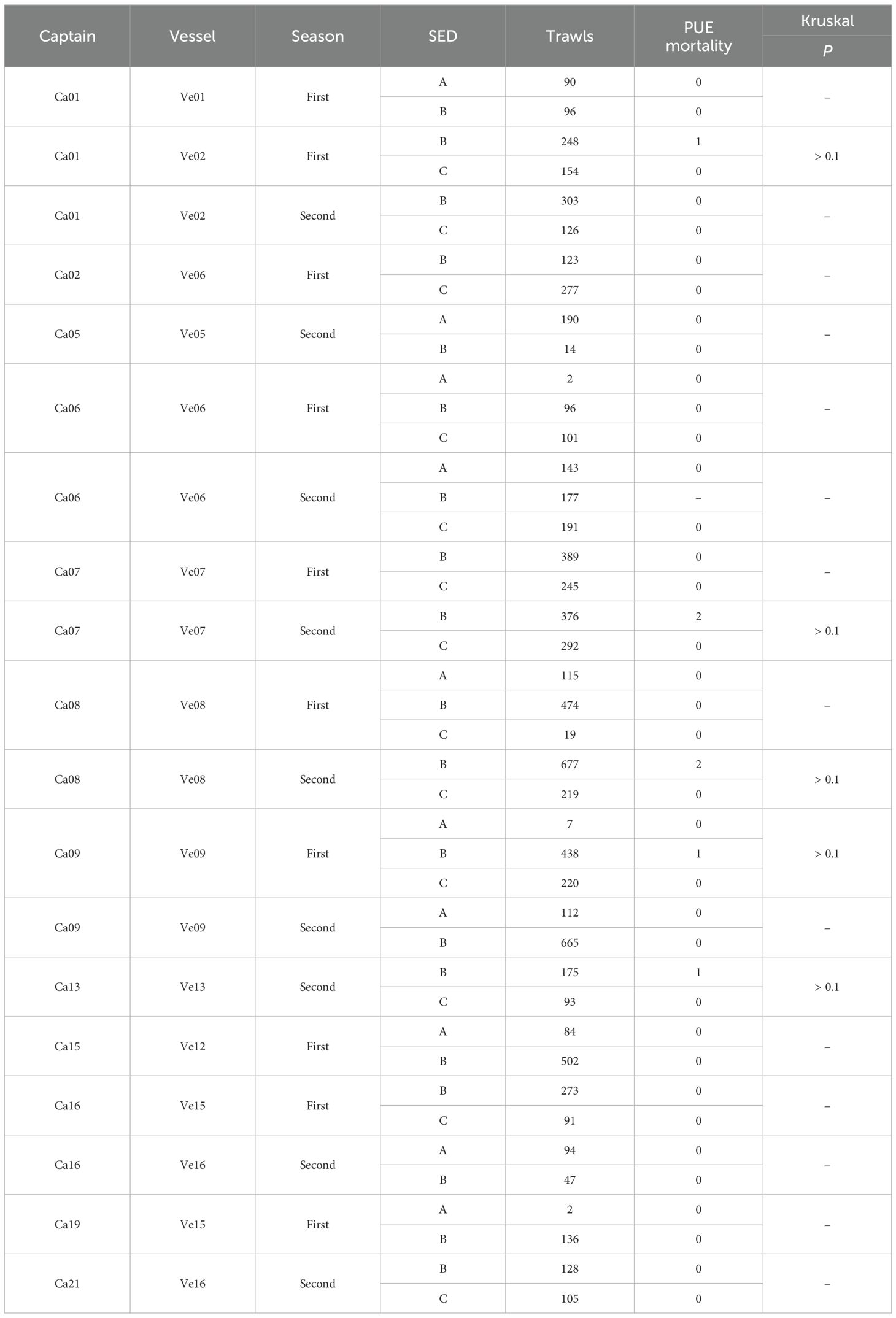
Table 5. PUE mortalities by captains that used two or three different SEDs on the same vessel in either first or second fishing seasons, compared by Kruskal-Wallis tests.
4 Discussion
In this paper we have presented six years (12 fishing seasons) of pinniped bycatch management through monitoring post-SED implementation. Following a sudden observed increase in pinniped interactions, SEDs were initially implemented in the Falkland Islands D. gahi bottom-trawl fishery during second season 2017 (Iriarte et al., 2020), and for the study period 2018–2023 a team of dedicated observers provided 100% vessel coverage. This fishery management response resulted in an important decrease of pinniped mortalities: from 143 reported incidental mortalities (fresh carcasses) for a single fishing season in 2017 (Iriarte et al., 2020), to 107 observed mortalities (in different stages of decomposition) within the next 12 fishing seasons (this study). The applied management was thus successful, mitigating pinniped bycatch without affecting squid yields (Iriarte et al., 2020; Falkland Islands Government, 2024). However, incidental mortalities were not eliminated completely. Through the integration of different research approaches (fisheries, observer, and forensic data), our results confirmed that seal mortality in the squid fishery is mostly comprised of SAFS males, typically occurs during trawl hauling, and have highlighted the critical role captains play during trawl maneuvering to ensure SED efficiency for mortality mitigation. Because the observer protocol was designed to record cryptic mortality, we were able to identify and report for the first time, otariid incidental mortality due to propeller strikes in an industrial offshore fishery operating in the Southwestern Atlantic.
4.1 Pinniped bycatch and SED live escapes
This study confirms previous reports that higher pinniped attendance to the Patagonian squid fishery occurs during second fishing seasons and primarily within the hauling maneuver, SAFS being the most abundant interacting species (Iriarte et al., 2020; Iriarte, 2022). Of the 55,232 live seals observed directly interacting with the fishing gear in this study, 508 (0.91%) were bycaught in SED trawls, of which 455 (89.5%) were live individuals, while 53 (10.4%) drowned, either as consequence of blockage of the SED escape path, or as a result of an entanglement with some part of the trawl. Of the live individuals, 77% of seals trapped in the trawl safely escaped through the SED, while 23% were brought aboard and released from deck. Because no cameras were installed in the trawls, recorded SED escapes were exclusively observed at the surface, during trawl hauling. The number of underwater SED escapes during the remaining trawl maneuvers is unknown. Nevertheless, in the Patagonian squid fishery it has been previously reported that SAFS can enter in the trawl during shooting and safely escape through the SED hatch (Iriarte et al., 2020).
The positive relationship of trawl duration with the number of SED escapes per trawl during first seasons, could imply the existence of a minimum time frame during which seals would be attracted to aggregate astern and follow a vessel until its hauling maneuver. Considering that the fleet is fully equipped with discard storage tanks, in hauls of long duration trawls and/or with low floating squid availability (i.e., small catches and/or carried out in good weather conditions), seals might maximize their energy cost-benefit of following a vessel for hours by taking the risk of entering the trawl during the haul, then escaping through the SED.
High trawl speed during hauling can catch an animal more quickly in front of the mouth of the net and lower its chance of evasion by swimming, increasing the probability of bycatch. In addition, higher speed might prevent seals from escaping in time through the SED hatch. Results of this study evidenced that faster hauling speeds of certain captains not only can increase the risk of a seal being caught, but can reduce the time frame for the trapped seal to reach the SED, which is consistent with what has been found in Australian marine mammal-finfish trawl fishery interactions (Hamer and Goldsworthy, 2006; Allen et al., 2014). In the Patagonian squid fishery, live seals which end up on deck not only represent a potential hazard for crew, but their release from deck usually causes a delay on the subsequent trawl shoot.
Our results show there is a negative relationship between large catches and seal bycatch/SED escapes, which is coherent with a higher availability of squid becoming loose from the trawl meshes, giving seals the opportunity to forage effectively around the net without entering the trawl (Supplementary Video S2). This indicates that seals attend haul maneuvers to opportunistically forage around the net on unmeshed squid from the wings and body of the trawl, consistent with marine mammal behavior in fisheries elsewhere (Wickens and Sims, 1994; Hamer and Goldsworthy, 2006; Greenman and McFee, 2014; Hamilton and Baker, 2015; Lyle et al., 2016; Bonizzoni et al., 2022). At hauls with a high availability of floating squid (i.e., large catches and/or carried out under the forces of rough weather), seals might cover their energy cost-benefit by foraging around the net without entering the trawl. A recent study in this fishery found that overall seal attendance to vessels (i.e., including direct interactions without any bycatch) associated with high catch quantities (Riaz et al., 2024). Nevertheless, our results are consistent with research carried out in other fisheries, which suggest that in rough weather foraging dives may be more energetically demanding, with seals therefore more attracted to opportunistically attend fishing vessel maneuvers (Hamer and Goldsworthy, 2006; Hamilton and Baker, 2015), but without a direct relationship with higher bycatch (Hamer and Goldsworthy, 2006).
The positive relation of SED escapes to Beaufort scale (across moderate levels) might be indicative that better sea conditions prevented the fishing gear from bouncing and losing tension during haul maneuvering, favoring seal escapes. However, less net bouncing could also be translated into a reduced number of enmeshed squid in the wings/body of the trawl becoming loose, seals entering the fishing gear to maximize their opportunistic foraging, and then safely escaping through the SED. The positive relationship between SED escapes and Beaufort scale may also reflect sea conditions facilitating captains’ maneuvering and compliance with mandatory best practice during haul maneuvers (i.e., working cables and bridles back and forth to facilitate the escape of seals while the SED is still in the water), or just that in good weather conditions observers had gantry access and could monitor SED escapes during hauling more efficiently.
The positive relation between SED escapes and the performance of 1–3 turns during trawling, resembles the relation between SED escapes and trawl duration seen for first seasons; seals balancing the cost-benefit of following a vessel for several hours, maximizing their opportunistic foraging at the haul while entering the trawl.
4.2 PUE
The positive relationship of trawl duration with PUE mortalities per sighting is explained by hauls being not only the maneuver that seals attend the most, but also the maneuver when mortalities can actually be recorded. The absence of a relationship between trawl duration and PUE mortalities per trawl, is evidence that PUE mortalities are most likely to occur during hauling, irrespective of trawl duration. This result matches the bycatch multimedia analysis, which confirmed that 40 out of 53 PUE mortalities had lubricated brown eyes (Supplementary Table S5), evidencing a short time lapse since death (i.e., < 1 h). The significant positive relationship between PUE mortalities and the performance of turns also corresponded with the results of the bycatch multimedia analyses. We confirmed that seven out of nine PUE mortalities with reddened eyes were recovered from trawls with one to four turns carried out at least 50 min before the haul (Supplementary Table S5). These mortalities attributed to vessel turning might have occurred while the rigidity of the trawl was relaxed, creating folds or bulges that could have blocked the SED escape path. The two individuals with reddened eyes recovered from trawls without any turn could have died either during shooting or trawling; mortality after SED net extension entanglements during shooting has been reported to occur in the fishery (Iriarte et al., 2020). The positive relationship of absolute PUE mortality numbers per trawl—but not per sighting—with the Beaufort scale, implies that higher wind and sea state attracted more seals to trawls; however this did not have a proportionally more lethal effect. Trawl bouncing and associated water flux in bad weather might increase the number of squid detaching off the trawl wings and body, balancing the cost-benefit for seals to attend vessels without entering the net and risk being killed. Although mortalities due to trawl door and neck entanglements in the net meshes cannot be prevented, the outcome of a bycatch event can be influenced by different factors. Bad weather could increase the probability of entanglements and twists in the trawl system, while captain decisions can influence trawl speed and the maneuvering of the fishing gear. Even though in Australia 50% of fur seal incidental mortalities were reported to take place during trawl shooting (Hamer and Goldsworthy, 2006) or trawling (Lyle et al., 2016), in the Patagonian squid fishery 81.6% of PUE mortality occurred during trawl hauling, implicating the expertise of the captain in performing this maneuver. While it may be surprising that mortality risk is highest just before the end of a trawl, hauling is the maneuver when tons of bulk and hard metal abruptly collide and which typically has the highest pinniped attendance. Not coincidentally, our results indicate that hauling is the maneuver that may represent the most variance among vessel captains’ operational styles.
4.3 Pinniped attendance to vessels and propeller strikes
Like most industrial trawlers, the Patagonian squid fleet uses ducted propellers with controllable pitch, which enhance efficiency for highly loaded propellers at low speeds (Koh et al., 2015). A heavy cod-end may increase the risk of suction into the duct, by requiring the vessel to generate more thrust when hauling (Wickens and Sims, 1994). A heavy cod-end may also push pinnipeds performing high-speed foraging dives at the stern of the vessel, into closer proximity with the propeller. This could particularly occur during the haul of large catches, when there might be a higher availability of unmeshed squid; seals foraging around the net and performing high-speed dives very close to the stern being more vulnerable to propeller strikes (Wickens and Sims, 1994). Marine mammal-vessel collisions are known from multiple environments, but are mostly reported from stranded individuals (Bexton et al., 2012; Barcenas-De La Cruz et al., 2017; Schoeman et al., 2020; Olson et al., 2021). Ducted propeller damage due to fur seal interactions has been reported for a South African offshore demersal trawl fishery (Wickens and Sims, 1994). In the Patagonian squid fishery dead seals have sometimes required to be removed from propellers by divers, but we are not aware of any damage caused to propellers. Although the three fresh carcasses recorded were exsanguinated, it is believed the exsanguination occurs in 3–4 h (Di Maio and Di Maio, 2001; Moore et al., 2013). This time range allows the possibility of the fresh carcass being recovered in the trawl of the same vessel that caused the sharp trauma mortality with its propeller. Because seals forage in productive areas where also fishing vessels operate (Riet-Sapriza et al., 2013; Hamilton and Baker, 2015; Riaz et al., 2023), seals foraging in the vicinity of a trawling vessel could become distracted/confused in the wake and risk being pulled towards the propeller (Wickens and Sims, 1994; Bexton et al., 2012). To our knowledge, this study is the first to link otariid vessel collision sharp trauma directly to a large-vessel trawl fishery operating in the Southwestern Atlantic. All except one of the carcasses with traumatic injury were recorded in a remote area in the south of the fishing zone, where other vessel traffic does not occur. Although diagnostic muscle samples were not taken to confirm that the strikes occurred pre-mortem (see Sierra et al., 2014), it is unlikely for a non-bloated carcass to engage the propeller of an offshore fishing vessel by drifting. Neither of the trauma-injury mortalities in this study had bloating nor was marked, and only one exhibited reddened eyes (Supplementary Table S6). We presume the type of injury depends on the shape of the propeller blades, their type of rotation, and the movement and speed of the swimming seal. It is important to note that in a bottom-trawl fishery such as this, pinniped carcasses —which presumably sink similarly to cetaceans (Moore et al., 2020) —are more likely to be retrieved and recorded than in pelagic fisheries, where marine mammal bycatch interactions are higher (Crespo et al., 1997; Hall et al., 2000; Lyle et al., 2016) and many lethal propeller interactions might remain cryptic.
4.4 Undefined cause of mortality
One of the constraints on pinniped carcass collection for post-mortem examinations is the state of decomposition. Given that the Patagonian squid fleet processes and freezes seafood for human consumption, health and safety protocols required that only freshly bycaught seals could be preserved. The 12 unmarked carcasses in decomposition/unknown stage of decomposition from 2018–2019 could have originated from observer errors (i.e., mismarking of carcasses), while the five unmarked carcasses in decomposition recorded in second season 2020 could correspond to unreported mortalities that occurred during the first fishing week without observer coverage (Table 1). It is unknown whether the remaining nine carcasses represented natural mortalities, unrecorded/unreported drownings, SED cryptic mortality (i.e., lost fresh carcasses), or post-deck release mortalities. Because most of the fishing effort is carried out in an area with high occurrence of SAFS, natural mortality is not ruled out. Two of the necropsied individuals presented ulcerative gastritis associated to nematode infection, which has been linked to stranding and deteriorating health conditions in pinnipeds (Jacobus et al., 2016). Though the necropsied individuals were robust, it is presumed the severity of their lesions would have led them to natural mortality (Supplementary Table S8). Marine mammals with a debilitated health status and young individuals have been reported to have higher vulnerability to bycatch (Brown et al., 2014; Barcenas-De La Cruz et al., 2017; Bernaldo de Quirós et al., 2018; IJsseldijk et al., 2021). In addition, unobserved mortalities occurring during observer resting times could have remained unreported by the vessel. While the top-open SEDs proved to be efficient for retaining carcasses, losses could have occurred if the SED flipped during hauling, remaining unreported if the observer was not carrying out their work from the gantry.
4.5 Captain effect on pinniped bycatch
The significantly higher number of deck releases for specific captains during first and second seasons, suggests fast hauling that left seals trapped inside the SED net extension, bringing them aboard. The two captains who had significantly more than random SED escapes, suggests their trawl maneuvering during hauling allowed seals to escape through the SED hatch while still in the water (e.g., hauling slowly, keeping trawl meshes extended, avoiding trawl bends, and working cables back and forth before hauling the SED aboard). Captains who performed hauls at low speed and worked warp cables back and forth, facilitated seal SED escapes. In contrast, captains who did not haul in a way that kept trawl meshes extended and avoided trawl bends, impeded seal escapement and hindered their survival. The fact that three of the captains (Ca02, Ca04, Ca11), who commanded more than one vessel (Supplementary Tables S1, S2) accounted for 34.2% of all pinniped captures in first seasons and also had significantly more than random bycatch in second seasons, supports the existence of a captain effect on the bycatch outcome. Vessel effects have been previously reported on megafauna-fisheries interactions elsewhere (e.g., Gilman et al., 2007; Dietrich et al., 2009; Allen et al., 2014). Considering that PUE mortalities in the Patagonian squid fishery are most likely to occur during hauling, and the positive relationship of PUE mortalities—without significant numbers of total bycatch—with two of the captains (Ca13, Ca37), supports that in the Patagonian squid fishery captains’ way of maneuvering the trawl can affect SED efficiency. However, in rough weather captains might be forced to perform the haul on a turn to maintain deck crew safety. The statistically significant clustering of pinniped bycatch and mortality shown in this study has been corroborated from other fisheries which have found vessel and/or captain performance heterogeneities in mitigating megafauna bycatch (Hall et al., 2000, 2003; Gilman et al., 2007; Dietrich et al., 2009; Allen et al., 2014; Hall, 2015). Regarding Ca02 fresh propeller mortalities, although the sample was too small for statistical conclusion, vessel maneuvering through a ducted propeller could generate a stream of fast water to draw a seal swimming in the water column (see Wickens and Sims, 1994; Bexton et al., 2012).
4.6 Sustainability of prevailing pinniped mortality
The overlap of fisheries with megafauna foraging areas is well-established (Crespo et al., 1997; Kirkwood et al., 2006; Romero et al., 2011; Reyes et al., 2013; Riet-Sapriza et al., 2013; Franco-Trecu et al., 2019a; Carneiro et al., 2022), however this overlap does not necessarily translate into bycatch and incidental mortality (e.g., Hamer and Goldsworthy, 2006; Carneiro et al., 2022). The risk of bycatch of an individual might depend on its behavioral response to the presence of a fishing vessel, which can be influenced by sex, age, sexual maturity, and health status (Crespo et al., 1997; Kirkwood et al., 2006; Riet-Sapriza et al., 2013; Brown et al., 2014; Bernaldo de Quirós et al., 2018; IJsseldijk et al., 2021; Carneiro et al., 2022). The results of this study provide detailed information on sex and age class of the individuals interacting with the Falkland Islands Patagonian squid fishery, filling a knowledge gap on intrinsic pinniped factors (see Riaz et al., 2023, 2024). The Falkland Islands D. gahi fishing area has been defined as an important foraging area for female SAFS during second fishing seasons (Riaz et al., 2023), and seal-fishery interactions have been described as a significant and consistent problem (Riaz et al., 2024). The results of this study support that recommended best practice (e.g., FAO, 2021; Moore et al., 2021; Wade et al., 2021; Good et al., 2024a) implemented in the fishery has been successful at mitigating the pre-SED pinniped-fishery agonistic interactions recorded in 2017 (Iriarte et al., 2020). The total number of pinniped mortalities recovered in the SED-fitted Patagonian squid fishery has not only been low (107 observed carcasses in different stages of decomposition in six years), but we confirmed that 78% comprised of adult and immature males and only 7% were females (Supplementary Tables S4-S7). This male bias in seal-fishery interactions and bycatch is consistent with previous reports from fisheries elsewhere (Crespo et al., 1997; Kirkwood et al., 2006; Goetz et al., 2008; Königson et al., 2013; Rivadeneyra-Villafuerte, 2021; Sepúlveda et al., 2023; Westphal et al., 2025). Both SAFS and SASL are polygynous species, and few dominant males monopolize breeding females (Capozzo, 1995; Ponce de León and Pin, 2006; Franco-Trecu et al., 2014, 2015). Because few males actually contribute to the genetic pool of their populations, their average reproductive value is low and their removal from the population is less important than the removal of adult females (e.g., Crespo et al., 1997; Wallace et al., 2008; Königson et al., 2013; Hall, 2015; Hindell et al., 2022). Furthermore, male-selective commercial harvesting of SAFS has not affected their post-harvesting recovery (Franco-Trecu et al., 2019b). However, SASL have lower resilience than SAFS and have required longer recovery after harvest pressure (Romero et al., 2017; Franco-Trecu et al., 2019b). Although endangered pinniped populations can be impacted by incidental mortality of few individuals (Alava and Salazar, 2006; Gobush et al., 2016), for healthy pinniped stocks such losses are not necessarily unsustainable (Wickens and Sims, 1994; Crespo et al., 1997; Read, 2008; Underwood et al., 2008; Romero et al., 2024). Both SAFS and SASL are listed as global level “Least Concern” by the IUCN (Cárdenas-Alayza et al., 2016a, 2016b). The South Atlantic population of SAFS is genetically diverse and increasing (Crespo et al., 2015; Cárdenas-Alayza et al., 2016a; Grandi and Milano, 2024), and its minimum population size is estimated at 280,000 individuals (Grandi and Milano, 2024). The Falkland Islands archipelago holds the largest South American fur seal population, with a minimum production of 36,425 pups per year (Baylis et al., 2019). The South Atlantic population of SASL is genetically diverse and stable, with total population size estimated at 143,000 individuals (Feijoo et al., 2011; Cárdenas-Alayza et al., 2016b). Minimum estimate of SASL pup production in the Falkland Islands is 4,443 (Baylis et al., 2015).
Considering the reproductive strategy, population sizes, and conservation status of both SAFS and SASL, the 107 mortalities (in different stages of decomposition) recorded between 2018–2023 in the Falkland Islands bottom-trawl-SED-fitted squid fishery, are negligible. However, in the Falkland Islands marine mammals are protected species. The Marine Mammals Ordinance prohibits the intentional kill and take of marine mammals (Falkland Islands Government, 1992), while the Fisheries (Conservation and Management) Ordinance sanctions the implementation of measures to prevent or mitigate mortality or injury of marine mammals in fishing operations (Falkland Islands Government, 2005). In recent years, values related to the protection of megafauna have created demand for market-based initiatives of sustainable fisheries management, product certification and ecolabelling (Wessells et al., 2001; Hilborn et al., 2023; Good et al., 2024a, 2024b), promoting consumers’ expectation of corporate social responsibility (Peiró-Signes et al., 2022). Continuous observer monitoring of pinniped interactions in the squid fishery is necessary to better understand the factors that affect even the small numbers of prevailing incidental mortalities, principally SED efficiency and the occurrence of propeller strikes.
4.7 Recommendations
Our results can be used to improve best practice, particularly while vessels are targeting squid south of 52°S, where SAFS attendance to the fishery is higher. In order to ensure the safe escape of seals through the SED hatch during both shooting and hauling, closing the mouth of the trawl should be avoided and trawl meshes should be kept extended, preventing bends. During bycatch events, hauling speed should be low enough to give time to the trapped seal/s to arrive to the SED net extension on time. In rough weather conditions, it would be ideal to carry out shorter trawls to avoid hauling full cod-ends, which not only adds a safety risk for the vessel and its crew, but also increases the probability of entanglement and occurrence of twists in the trawl meshes. During second seasons, in the south of the fishing area the performance of turns should be replaced by consecutive shorter trawls. In addition, Falkland Islands Fisheries Department (i.e., fishery managers, scientists, fishery officers) and industry should strengthen collaboration to ensure pinniped interactions reporting and SED efficiency. Direct outreach from scientists to captains, increased fishery officers’ at-sea inspections for SED compliance, and industry incentives to their captains, could ensure a continuous contribution to the sustainable management of the fishery.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author/s.
Ethics statement
Seal carcasses were collected under Research License No: R12/2022 issued by the Falkland Islands Environment Department.
Author contributions
VI: Conceptualization, Data curation, Investigation, Methodology, Project administration, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. AW: Data curation, Formal analysis, Methodology, Software, Validation, Visualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. The pinniped bycatch monitoring program was funded by the Loligo Producers Group (LPG) (2017–2019), LPG and Falkland Islands Government (FIG) (2020–2021), and FIG (2021–2023). This research and its publication have been financially supported by the Directorate of Natural Resources, FIG.
Acknowledgments
We are indebted to all the dedicated observers, particularly to the core team: Beatríz Rodríguez, Dália Neves, Eva Burgos, Félix Morales, Fernando Anido, Gonçalo Álvares, Hugo Dias, Jon Ander Marín, Konstantinos Papadopoulos, Matea Haggia, Nuno Carrilho, Pedro de Jesus, Rodolfo Curralo, and Vedrana Vukasin. Photo credits: N. Carrilho (Figure 2a), Pere Hernández (Figures 2b, 4c, d), H. Dias (Figure 2c), E. Burgos (Figure 2d), R. Curralo (Figures 2e, 3e, 4a), P. de Jesus (Figures 2f, 3a), Gala Serrano (Figure 3c), B. Rodríguez (Figure 4b), and Paloma del Mar Carrasco (Figure 4e). Video credits: P. de Jesus (V1), J.A Marín (V2). Special thanks to all the captains and crews for fruitful discussions. Fortuna and Polar Seafish kindly provided space in their cold stores. Dale Evans and several FIFD observers assisted with post-mortems; Dr Vet. Med. Phillip Van der Riet provided expert advice; Alex Blake prepared trawl, depth, and positional data. We are thankful to the editor and the reviewers, who provided suggestions and questions that allowed us to improve the manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2025.1588956/full#supplementary-material
References
Alava J. J. and Salazar S. (2006). “Status and conservation of otariids in Ecuador and the Galápagos Islands,” in Sea Lions of the World. Eds. Trites A. W., Atkinson S. K., DeMaster D. P., Fritz L. W., Gelatt T. S., Rea L. D., et al (Alaska Sea Grant College Program, Fairbanks), 495–520.
Allen S. J., Tyne J. A., Kobryn H. T., Bejder L., Pollock K. H., and Loneragan N. R. (2014). Patterns of dolphin bycatch in a North-Western Australian trawl fishery. PloS One 9, e93178. doi: 10.1371/journal.pone.0093178
Altmann J. (1974). Observational study of behavior: sampling methods. Behavior 49, 227–267. doi: 10.1163/156853974X00534
Arbelo M., Espinosa de los Monteros A., Herráez P., Andrada M., Sierra E., Rodríguez F., et al. (2013). Pathology and causes of death of stranded cetaceans in the Canary Islands, (1999-2005). Dis. Aquat. Org. 103, 87–99. doi: 10.3354/dao02558
Arkhipkin A. I., Middleton D. A., and Barton J. (2008). Management and conservation of a short-lived fishery resource: Loligo gahi around the Falkland Islands. Am. Fish. Soc Symp. 49, 1243–1252.
Arregui M., Bernaldo de Quirós Y., Saavedra P., Sierra E., Suárez-Santana C. M., Arbelo M., et al. (2019). Fat embolism and sperm whale ship strikes. Front. Mar. Sci. 6. doi: 10.3389/fmars.2019.00379
Barcenas-De La Cruz D., DeRango E., Johnson S. P., and Simeone C. A. (2017). Evidence of anthropogenic trauma in marine mammals stranded along the central California coast 2003-2015. Mar. Mamm. Sci. 34, 330–346. doi: 10.1111/mms.12457
Batallés L. M., Pin O., and Lima M. (1990). Estudio del crecimiento del lobo fino Sudamericano (Arctocephalus australis) en Isla del Lobos, Uruguay. Frente Marítimo 7, 69–73.
Baylis A. M. M., Orben R. A., Arnould J. P. Y., Christiansen F., Hays G. C., and Staniland I. J. (2015). Disentangling the cause of a catastrophic population decline in a large marine mammal. Ecology 96, 2834–2847. doi: 10.1890/14-1948.1
Baylis A. M. M., Orben R. A., Arkhipkin A. A., Barton J., Brownell R. L., Staniland I. J., et al. (2019). Re-evaluating the population of South American fur seals and conservation implications. Aquat. Conserv.: Mar. Freshw. Ecosyst. 29, 1988–1995. doi: 10.1002/aqc.3194
Bernaldo de Quirós Y., Hartwick M., Rotstein D. S., Garner M. M., Bogomolni A., Greer W., et al. (2018). Discrimination between bycatch and other causes of cetacean and pinniped stranding. Dis. Aquat. Org. 127, 83–95. doi: 10.3354/dao03189
Bexton S., Thompson D., Brownlow A., Barley J., Milne R., and Bidewell C. (2012). Unusual mortality of pinnipeds in the United Kingdom associated with helical (corkscrew) injuries of anthropogenic origin. Aquat. Mamm. 38, 229–240. doi: 10.1578/AM.38.3.2012.229
Bonizzoni S., Hamilton S., Reeves R. R., Genov T., and Bearzi G. (2022). Odontocete cetaceans foraging behind trawlers, worldwide. Rev. Fish. Biol. Fish. 32, 827–877. doi: 10.1007/s11160-022-09712-z
Brown S., Reid D., and Rogan E. (2014). Characteristics of fishing operations, environment and life history contributing to small cetacean bycatch in the Northeast Atlantic. PloS One 9, e104468. doi: 10.1371/journal.pone.0104468
Byard R. W., Winskog C., Machado A., and Boardman W. (2012a). The assessment of lethal propeller strike injuries in sea mammals. J. Forensic Leg. Med. 19, 158–161. doi: 10.1016/j.jflm.2011.12.017
Byard R. W., Machado A., Woolford L., and Boardman W. (2012b). Symmetry: the key to diagnosing propeller strike injuries in sea mammals. Forensic Sci. Med. Pathol. 9, 103–105. doi: 10.1007/s12024-012-9335-0
Capozzo H. L. (1995). Comportamiento reproductivo en dos especies de Otáridos de América del Sur. [PhD thesis]. [Buenos Aires] (Universidad de Buenos Aires: Facultad de Ciencias Exactas y Naturales).
Cárdenas-Alayza S., Oliveira L., and Crespo E. (2016a). Arctocephalus australis. The IUCN Red List of Threatened Species 2016a: e.T2055A45223529. https://www.researchgate.net/publication/319077319_Arctocephalus_australis_South_American_fur_seal_The_IUCN_Red_List_of_Threatened_Species_2016 (Accessed November 7, 2024).
Cárdenas-Alayza S., Crespo E., and Oliveira L. (2016b). Otaria byronia. The IUCN Red List of Threatened Species 2016b: e.T41665A61948292. http://dx.doi.org/10.2305/IUCN.UK.2016-1.RLTS.T41665A61948292.en (Accessed November 7, 2024)
Carneiro A. P. B., Clark B. L., Pearmain E. J., Clavelle T., Wood A. G., and Phillips R. A. (2022). Fine-scale associations between wandering albatrosses and fisheries in the Southwest Atlantic Ocean. Biol. Conserv. 276, 109796. doi: 10.1016/j.biocon.2022.109796
Caruso J. L. (2016). Decomposition changes in bodies recovered from water. Acad. Forensic Pathol. 6, 19–27. doi: 10.23907/2016.003
Crespo E. A. (2021). “Exploitation and recovery of the South American sea lion in the Southwestern Atlantic,” in Ethology and behavioral ecology of otariids and the odobenid. Eds. Campagna C. and Harcourt R. (Springer, Cham), 521–537. doi: 10.1007/978-3-030-59184-7_24
Crespo E. A., Pedraza S. N., Dans S. L., Koen Alonso M., Reyes L. M., García N. A., et al. (1997). Direct and indirect effects of the highseas fisheries on the marine mammal populations in the northern and central Patagonian coast. J. Northw. Atl. Fish. Sci. 22, 189–207. doi: 10.2960/J.v22.a15
Crespo E. A., Schiavini A. C. M., García N. A., Franco-Trecu V., Goodall R. N. P., Rodríguez D., et al. (2015). Status, population trend and genetic structure of South American fur seals, Arctocephalus australis, in Southwestern Atlantic waters. Mar. Mamm. Sci. 31, 866–890. doi: 10.1111/mms.12199
Curran P. A. (2015). Monte Carlo error analyses of Spearman’s rank test. Instrumentation and Methods for Astrophysics. arXiv e-prints 979, 1–5. doi: 10.48550/arXiv.1411.3816
Dietrich K. S., Parrish J. K., and Melvin E. F. (2009). Understanding and addressing seabird bycatch in Alaska demersal longline fisheries. Biol. Conserv. 142, 2642–2656. doi: 10.1016/j.biocon.2009.06.013
Di Maio V. J. and Di Maio D. (2001). “Forensic pathology,” in CRC series in practical aspects of criminal and forensic investigations (CRC Press, Boca Raton, FL).
Epple A. L., Daniel J. T., Barco S. G., Rotstein D. S., and Costidis A. M. (2020). Novel necropsy findings linked to peracute underwater entrapment in bottlenose dolphins (Tursiops truncatus). Front. Mar. Sci. 7. doi: 10.3389/fmars.2020.00503
Falkland Islands Government (1992). The Marine Mammals Ordinance. Available online at: https://www.falklands.gov.fk/mineralresources/jdownloads/Legislation/Environmental/marine_mammals_ordinance_1992_updated.pdf (Accessed June 13, 2025).
Falkland Islands Government (2005). Fisheries (Conservation and Management) Ordinance. Available online at: https://www.legislation.gov.fk/view/html/inforce/2019-03-17/fiord-2005-14 (Accessed June 13, 2025).
Falkland Islands Government (2024). Fisheries Department Fisheries Statistics. 28, 2023. Available online at: https://www.falklands.gov.fk/fisheries/publications/fishery-statistics?task=download.send&id=333:fisheries-statistics-28-2023&catid=7 (Accessed June 13, 2025).
FAO (2021). “Fishing operations. Guidelines to prevent and reduce bycatch of marine mammals in capture fisheries,” in FAO Technical Guidelines for Responsible Fisheries No.1, vol. Suppl. 4. (Rome: Food and Agriculture Organisation of the United Nations). doi: 10.4060/cb2887en
Feijoo M., Lessa E. P., Loizaga de Castro R., and Crespo E. A. (2011). Mitochondrial and microsatellite assessment of population structure of South American sea lion (Otaria flavescens) in the Southwestern Atlantic Ocean. Mar. Biol. 158, 1857–1867. doi: 10.1007/s00227-011-1697-4
Franco-Trecu V., Costa P., Schramm Y., Tassino B., and Inchausti P. (2014). Sex on the rocks: reproductive tactics and breeding success of South American fur seal males. Behav. Ecol. 25, 1513–1523. doi: 10.1093/beheco/aru145
Franco-Trecu V., Costa-Urrutia P., Schramm Y., Tassino B., and Inchausti P. (2015). Tide line versus internal pools: mating system and breeding success of South American sea lion males. Behav. Ecol. Sociobiol. 69, 1985–1996. doi: 10.1007/s00265-015-2010-1
Franco-Trecu V., Szephegyi M. N., Doño F., Forselledo R., Reyes F., Passadore C., et al. (2019a). Marine mammal bycatch by the industrial bottom trawl fishery at the Río de la Plata Estuary and the adjacent Atlantic Ocean. Lat. Am. J. Aquat. Res. 47, 89–101. doi: 10.3856/vol47-issue1-fulltext-10
Franco-Trecu V., Drago M., Grandi M. F., Soutullo A., and Crespo E. A. (2019b). Abundance and population trends of the South American fur seal (Arctocephalus australis) in Uruguay. Aquat. Mamm. 45, 48–55. doi: 10.1578/AM.45.1.2019.48
Geraci J. R. and Lounsbury V. J. (1993). Marine Mammals Ashore. A field guide for strandings. (Galveston, TX: Texas A&M University Sea Grant publications).
Gilman E., Kobayashi D., Swenarton T., Brothers N., Dalzell P., and Kinan-Kelly I. (2007). Reducing sea turtle interactions in the Hawaii-based longline swordfish fishery. Biol. Conserv. 139, 19–28. doi: 10.1016/j.biocon.2007.06.002
Gobush K. S., Wurth T. A., Henderson J. R., and Becker B. L. (2016). Prevalence of interactions between Hawaiian monk seals (Nemonachus schauinslandi) and nearshore fisheries in the main Hawaiian Islands. Pac. Conserv. Biol. 23, 25–31. doi: 10.1071/PC15029
Goetz S., Wolff M., Stotz W., and Villegas M. J. (2008). Interactions between the South American sea lion (Otaria flavescens) and the artisanal fishery off Coquimbo, northern Chile. ICES J. Mar. Sci. 65, 1739–1746. doi: 10.1093/icesjms/fsn152
Good S. D., McLennan S., Gummery M., Lent R., Essington T. E., Wallace B. P., et al. (2024a). Updating requirements for Endangered, Threatened and Protected species MSC Fisheries Standard v3.0 to operationalize best practices. Mar. Pol. 163, 106117. doi: 10.1016/j.marpol.2024.106117
Good S. D., Dewar K., Burns P., Sainsbury K., Phillips R. A., Wallace B. P., et al. (2024b). Adapting the Marine Stewardship Council risk-based framework to estimate impacts on seabirds, marine mammals, marine turtles and sea snakes. Mar. Pol. 163, 106118. doi: 10.1016/j.marpol.2024.106118
Grandi M. F., Dans S. L., García N. A., and Crespo E. A. (2010). Growth and age at sexual maturity of South American sea lions. Mamm. Biol. 75, 427–436. doi: 10.1016/j.mambio.2009.09.007
Grandi M. F. and Milano V. N. (2024). Recovery of South American fur seals in the central South Atlantic Ocean. Mar. Mamm. Sci. 41, e13176. doi: 10.1111/mms.13176
Greenman J. T. and McFee W. E. (2014). A characterization of common bottlenose dolphin (Tursiops truncatus) interactions with the commercial shrimp trawl fishery of South Carolina, USA. J. Cet. Res. Manage. 14, 69–79. doi: 10.47536/jcrm.v14i1.525
Hall M. A. (2015). “More on bycatches: changes, evolution, and revolution,” in Fisheries bycatch: global issues and creative solutions. Eds. Kruse G. H., DiCosimo J., Eischens C. A., Gislason G. S., McBride D. N., Rose C. S., et al (Alaska Sea Grant, University of Alaska, Fairbanks, AK), 203–225. doi: 10.4027/fbgics.2015.1
Hall M., Alverson D. L., and Metuzals K. I. (2000). By-catch: problems and solutions. Mar. Pol. Bull. 41, 204–219. doi: 10.1016/S0025-326X(00)00111-9
Hall M. A., Campa M., and Gómez M. (2003). “Solving the tuna-dolphin problem in the Eastern Pacific purse-seine Fishery,” in Ocean Yearbook 17. Eds. Mann Borgese E., Chircop A., and McConell M. (The University of Chicago Press, Chicago, IL), 60–92.
Hamer D. J. and Goldsworthy S. D. (2006). Seal-fishery operational interactions: Identifying the environmental and operational aspects of a trawl fishery that contribute to by-catch and mortality of Australian fur seals (Arctocephalus pusillus doriferus). Biolog. Conserv. 130, 517–529. doi: 10.1016/j.biocon.2006.01.014
Hamilton S. and Baker G. B. (2015). Review of research and assessments on the efficacy of sea lion exclusion devices in reducing the incidental mortality of New Zealand sea lions Phocartos hookeri in the Auckland Islands squid trawl fishery. Fish. Res. 161, 200–206. doi: 10.1016/j.fishres.2014.07.010
Hamilton S. and Baker G. B. (2019). Population growth of an endangered pinniped–the New Zealand sea lion (Phocartos hookeri)–is limited more by high pup mortality than fisheries bycatch. ICES J. Mar. Sci. 76, 1794–1806. doi: 10.1093/icesjms/fsz039
Hilborn R., Amoroso R., Collie J., Hiddink J. G., Kaiser M. J., Mazor T., et al. (2023). Evaluating the sustainability and environmental impacts of trawling compared to other food production systems. ICES J. Mar. Sci. 80, 1567–1579. doi: 10.1093/icesjms/fsad115
Hindell M. A., McMahon C. R., Guinet C., Harcourt R., Jonsen I. D., Raymond B., et al. (2022). Assessing the potential for resource competition between the Kerguelen Plateau fisheries and southern elephant seals. Front. Mar. Sci. 9. doi: 10.3389/fmars.2022.1006120
IJsseldijk L. L., Scheidat M., Siemensma M. L., Couperus B., Leopold M. F., Morell M., et al. (2021). Challenges in the assessment of bycatch: postmortem findings in harbor porpoises (Phocoena phocoena) retrieved from gillnets. Vet. Pathol. 58, 405–415. doi: 10.1177/0300985820972454
Iriarte V. (2022). “LOL 2021-X MMO monitoring report,” in Fisheries Department, Directorate of Natural Resources, Falkland Islands Government. Available online at: https://www.falklands.gov.fk/fisheries/publications/megafauna-bycatch-mitigation?task=download.send&id=251:mmo-monitoring-program-report-2021-season-2&catid=19 (Accessed November 7, 2024).
Iriarte V., Arkhipkin A., and Blake D. (2020). Implementation of exclusion devices to mitigate seal (Arctocephalus australis, Otaria flavescens) incidental mortalities during bottom-trawling in the Falkland Islands (Southwest Atlantic). Fish. Res. 227, 105537. doi: 10.1016/j.fishres.2020.105537
Jacobus K., Marigo J., Gastal S. B., Taniwaki S. A., Ruoppolo V., Catão-Dias J. L., et al. (2016). Identification of respiratory and gastrointestinal parasites of three species of pinnipeds (Arctocephalus australis, Arctocephalus gazella, and Otaria flavescens) in southern Brazil. J. Zoo Wildl. Med. 47, 132–140. doi: 10.1638/2015-0090.1
Kirkwood R., Lynch M., Gales N., Dann P., and Sumner M. (2006). At-sea movements and habitat use of adult male Australian fur seals (Arctocephalus pusillus doriferus). Can. J. Zool. 84, 1781–1788. doi: 10.1139/z06-164
Koh K. K., Omar Y., Azreen E., and Nurhaslinda K. (2015). The study of ducted propeller in propulsion performance of a Malaysia fishing boat. J. Teknol. 74, 39–43. doi: 10.11113/jt.v74.4639
Königson S., Fjälling A., Berglind M., and Lunneryd S. G. (2013). Male gray seals specialize in raiding salmon traps. Fish. Res. 148, 117–123. doi: 10.1016/j.fishres.2013.07.014
Kovacs K. M., Aguilar A., Aurioles D., Burkanov V., Campagna C., Gales N., et al. (2012). Global threats to pinnipeds. Mar. Mamm. Sci. 28, 414–436. doi: 10.1111/j.17478-7692.2011.00479.x
Lyle J. M., Willcox S. T., and Hartmann K. (2016). Underwater observations of seal-fishery interactions and the effectiveness of an exclusion device in reducing bycatch in a midwater trawl fishery. Can. J. Fish. Aquat. Sci. 73, 436–444. doi: 10.1139/cjfas-2015-0273
Mann J. (1999). Behavioral sampling methods for cetaceans: a review and critique. Mar. Mamm. Sci. 15, 102–122. doi: 10.1111/j.1748-7692.1999.tb00784.x
Meyer S., Robertson B. C., Chilvers B. L., and Krkošek M. (2017). Marine mammal population decline linked to obscured by-catch. PNAS 114, 11781–11786. doi: 10.1073/pnas.1703165114
Moore J. E., Heinemann D., Francis T. B., Hammond P. S., Long K. J., Punt A. E., et al. (2021). Estimating bycatch mortality for marine mammals: concepts and best practices. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.752356
Moore M. J., Mitchell G. H., Rowles T. K., and Early G. (2020). Dead cetacean? Beach, bloat, float, sink. Front. Mar. Sci. 7. doi: 10.3389/fmars.2020.00333
Moore M. J., van der Hoop J., Barco S. G., Costidis A. M., Gulland F. M., Jepson P. D., et al. (2013). Criteria and case definitions for serious injury and death of pinnipeds and cetaceans caused by anthropogenic trauma. Dis. Aquat. Org. 103, 229–264. doi: 10.3354/dao02566
Olson J. K., Lambourn D. M., Huggins J. L., Raverty S., Scott A. A., and Gaydos J. K. (2021). Trends in propeller strike-induced mortality in harbor seals (Phoca vitulina) of the Salish Sea. J. Wildl. Dis. 57, 689–693. doi: 10.10.7589/JWD-D-20-00221
Peiró-Signes A., Miret-Pastor L., Galati A., and Segarra-Oña M. (2022). Consumer demand for environmental, social, and ethical information in fishery and aquaculture product labels. Front. Mar. Sci. 9. doi: 10.3389/fmars.2022.948437
Ponce de León A. and Pin O. D. (2006). “Distribución, reproducción y alimentación del lobo fino Arctocephalus australis y del león marino Otaria flavescens en Uruguay,” in Bases para la conservación y el manejo de la costa Uruguaya. Eds. Menafra R., Rodríguez-Gallego L., Scarabino F., and Conde D. (Vida Silvestre Uruguay, Montevideo), 305–313.
Pugliares K. R., Touhey K. M., Bogomolni A. L., Herzig S. M., and Moore M. J. (2007). Marine mammal necropsy: an introductory guide for stranding responders and field biologists (Massachusetts: Woods Hole Oceanographic Institution). doi: 10.1575/1912/1823
Queirolo D., Ahumada M., San Martín M., Román C., Araya-Schmidt T., Apablaza P., et al. (2025). Testing and implementation of an exclusion device for mitigating the incidental capture of sea lions in the Chilean small-scale trawl hake fishery. Fish. Res. 281, 107205. doi: 10.1016/j.fishres.2024.107205
R Core Team (2022). R: A language and environment for statistical computing (Vienna, Austria: R Foundation for Statistical Computing). Available online at: https://www.R-project.org/ (Accessed November 1, 2024).
Read A. J. (2008). The looming crisis: interactions between marine mammals and fisheries. J. Mammal. 89, 541–548. doi: 10.1644/07-MAMM-S-315R1.1
Reyes P., Hucke-Gaete R., and Torres-Florez J. P. (2013). First observations of operational interactions between bottom-trawling fisheries and South American sea lion, Otaria flavescens in south-central Chile. J. Mar. Biol. Assoc. U.K. 93, 489–494. doi: 10.1017/S0025315412001282
Riaz J., Orben R. A., Jones K. A., Shapiro M., Winter A., Brickle P., et al. (2023). Spatial overlap between South American fur seal foraging effort and commercial trawl fisheries in the Falkland Islands. Glob. Ecol. Conserv. 46, e02615. doi: 10.1016/j.gecco.2023.e02615
Riaz J., Büring T., van der Grient J., Winter A., Lee B., Brickle P., et al. (2024). Seal-fishery interactions in the Falkland Islands – operational and environmental factors drive resource competition. ICES J. Mar. Sci. 0, 1–12. doi: 10.1093/icesjms/fsae161
Riet-Sapriza F. G., Costa D. P., Franco-Trecu V., Marín Y., Chocca J., González B., et al. (2013). Foraging behavior of lactating South American sea lions (Otaria flavescens) and spatial-temporal resource overlap with the Uruguayan fisheries. Deep Sea Res. II. 88, 106–119. doi: 10.1016/j.dsr2.2012.09.005
Rivadeneyra-Villafuerte S. (2021). Interaction between the South American sea lion Otaria byronia and industrial vessels in Pisco (autumn 2013). Bol. Inst. Mar. Perú. 36, 282–303. doi: 10.53554/boletin.v36i1.331
Roberts J., Childerhouse S., Roe W., Baker G. B., and Hamilton S. (2018). No evidence of cryptic bycatch causing New Zealand sea lion population decline. PNAS 115, e8330–e8331. doi: 10.1073/pnas.1806136115
Roe W. D. and Meynier L. (2012). “Review of necropsy records for bycaught NZ sea lions (Phocartos hookeri), 2000–2008,” in New Zealand aquatic environment and biodiversity report 98. Available online at: https://webstatic.niwa.co.nz/library/AEBR_98.pdf (Accessed June 13, 2025).
Roe W. D., Gartrell B. D., and Hunter S. A. (2012). Freezing and thawing of pinniped carcasses results in artefacts that resemble traumatic lesions. Vet. J. 194, 326–331. doi: 10.1016/j.tvjl.2012.03.028
Romero M. A., Dans S., González R., Svendsen G., García N., and Crespo E. (2011). Solapamiento trófico entre el lobo marino de un pelo Otaria flavescens y la pesquería de arrastre demersal del golfo San Matías, Patagonia, Argentina. Lat. Am. J. Aquat. Res. 39, 344–358. doi: 10.3856/vol39-issue2-fulltext-16
Romero M. A., Grandi M. F., Koen-Alonso M., Svendsen G., Ocampo Reinaldo M., García N. A., et al. (2017). Analyzing the natural population growth of a large marine mammal after a depletive harvest. Sci. Rep. 7, 5271. doi: 10.1038/s41598-017-05577-6
Romero M. A., Flórez L., Ocampo Reinaldo M., Machado R., Coller M., and Crespo E. A. (2024). Bycatch in the balance: the trawl fisheries impact on South American sea lions in San Matias Gulf, with comments on other species. Mar. Mamm. Sci. 41, e13181. doi: 10.1111/mms.13181
Santana-Garcon J., Wakefield C. B., Dorman S. R., Denham A., Blight S., Molony B. W., et al. (2018). Risk versus reward: interactions, depredation rates, and bycatch mitigation of dolphins in demersal fish trawls. Can. J. Fish. Aquat. Sci. 75, 2233–2240. doi: 10.1139/cjfas-2017-0203
Schoeman R. P., Patterson-Abrolat C., and Plön S. (2020). A global review of vessel collisions with marine animals. Front. Mar. Sci. 7. doi: 10.3389/fmars.2020.00292
Sepúlveda M., Szteren D., Alfaro-Shigueto J., Crespo E. A., Durán L. R., Guerrero A. I., et al. (2023). Sea lion and fur seal interactions with fisheries and aquaculture in South American waters: threats and management perspectives. Mamm. Rev. 53, 116–131. doi: 10.1111/mam.12311
Shono H. (2008). Application of the Tweedie distribution to zero-catch data in CPUE analysis. Fish. Res. 93, 154–162. doi: 10.1016/j.fishres.2008.03.006
Sierra E., Fernández A., Espinosa de los Monteros A., Arbelo M., Díaz-Delgado J., Andrada M., et al. (2014). Histopathological muscle findings may be essential for a definitive diagnosis of suspected sharp trauma associated with ship strikes in stranded cetaceans. PloS One 2, e88780. doi: 10.1371/journal.pone.0088780
Skeljo F. and Winter A. (2023). “Falkland calamari Doryteuthis gahi stock assessment–2023 2nd season,” in Fisheries Department, Directorate of Natural Resources, Falkland Islands Government. Available online at: https://www.falklands.gov.fk/fisheries/publications/all-catergories?task=download.send&id=321:loligo-stock-assessment-second-season-2023&catid=3 (Accessed November 7, 2024).
Swartzman G., Huang C., and Kaluzny S. (1992). Spatial analysis of Bering sea groundfish survey data using generalized additive models. Can. J. Fish. Aquat. Sci. 49, 1366–1378. doi: 10.11397/f92-152
Underwood J. G., Hernandez Camacho C. J., Aurioles-Gamboa D., and Gerber L. R. (2008). Estimating sustainable bycatch rates for California sea lion populations in Gulf of California. Conserv. Biol. 22, 505–809. doi: 10.1111/j.1523-1739.2008.00919.x
Wade P. R. (1998). Calculating limits to the allowable human-caused mortality of cetaceans and pinnipeds. Mar. Mamm. Sci. 14, 1–37. doi: 10.1111/j.1748-7692.1998.tb00688.x
Wade P. R., Long K. J., Francis T. B., Punt A. E., Hammond P. S., Heinemann D., et al. (2021). Best practices for assessing and managing bycatch of marine mammals. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.757330
Wallace B. P., Heppel S. S., Lewison R. L., Kelez S., and Crowder L. B. (2008). Impacts of fisheries bycatch on loggerhead turtles worldwide inferred from reproductive value analyses. J. Appl. Ecol. 45, 1076–1085. doi: 10.1111/j.1365-2664.2008.01507.x
Wessells C. R., Cochrane K., Deere C., Wallis P., and Willmann R. (2001). “Product certification and ecolabelling for fisheries sustainability,” in FAO Fisheries Technical Paper 422. (Rome: Food and Agriculture Organisation of the United Nations).
Westphal L., von Vietinghoff V., Moritz T., von Nordheim H., Schaarschmidt T., Teifke J. P., et al. (2025). By-catch of grey seals in fish traps in the German Baltic Sea – from incidents to mitigation and fisheries regulation. Aquat. Conserv. Mar. Freshw. Ecosyst. 35, e70147. doi: 10.1002/aqc.70147
Wickens P. A. (1995). “A review of operational interactions between pinnipeds and fisheries,” in FAO Fisheries Technical Paper 346. (Rome: Food and Agriculture Organisation of the United Nations).
Wickens P. A. and Sims P. F. (1994). Trawling operations and South African (Cape) fur seals, Arctocephalus pusillus pussilus. Mar. Fish. Rev. 56, 1–12. Available online at: https://spo.nmfs.noaa.gov/sites/default/files/pdf-content/MFR/mfr563/mfr5631.pdf (Accessed June 13, 2025).
Winter A. (2020). “Falkland calamari Doryteuthis gahi stock assessment–2020 2nd season,” in Fisheries Department, Directorate of Natural Resources, Falkland Islands Government. Available online at: https://www.falklands.gov.fk/fisheries/publications/all-catergories?task=download.send&id=204:loligo-stock-assessment-second-season-2020&catid=3 (Accessed November 7, 2024).
Wood S. (2023). mgcv: Mixed GAM Computation Vehicle with Automatic Smoothness Estimation (Vienna: R Package). Available online at: https://cran.r-project.org/web/packages/mgcv/index.html (Accessed May 4, 2025).
Keywords: bottom-trawl, bycatch, drowning, incidental mortality, pinniped, propeller strike, seal exclusion device
Citation: Iriarte V and Winter A (2025) Disentangling pinniped incidental mortality in a bottom-trawl fishery with seal exclusion devices. Front. Mar. Sci. 12:1588956. doi: 10.3389/fmars.2025.1588956
Received: 06 March 2025; Accepted: 02 July 2025;
Published: 19 August 2025.
Edited by:
Gregory Skomal, Massachusetts Division of Marine Fisheries, United StatesReviewed by:
Juan Pablo Torres-Florez, Buro Happold, Saudi ArabiaAndrea Bogomolni, University of Massachusetts Boston, United States
Copyright © 2025 Iriarte and Winter. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Verónica Iriarte, dmlyaWFydGVAbmF0dXJhbHJlc291cmNlcy5nb3YuZms=