Abstract
Documented biodiversity associated with offshore oil and gas platforms has led to growing interest in their potential conversion to artificial reefs under Rigs-to-Reefs (RTR) programmes following decommissioning. In this study, we assessed fish assemblages associated with offshore platforms in Qatar’s Al Khalij oilfield for the first time, using remotely operated vehicle (ROV) video surveys. Species diversity, abundance, and biomass were quantified across depth gradients by surveying the full water column around two platforms. A total of 4,474 fishes were recorded, representing 46 taxa, with Carangidae and Serranidae as the most diverse families, and Pinjalo pinjalo, Scomberoides commersonnianus, and Lutjanus argentimaculatus as the most abundant species. Estimated biomass reached 6,184 kg per platform. A clear vertical zonation in species composition was observed, with Carangidae dominating surface layers and Serranidae more prevalent near the seafloor. Structurally complex areas, such as crossbeams and base zones, were identified as local biodiversity hotspots. Notably, Cheilinus trilobatus was recorded for the first time in the Arabian Gulf. Comparisons with previous studies from Qatar’s Al Shaheen oilfield revealed differences in species composition, likely due to variation in platform structure, geographic location, and sampling design. These findings demonstrate the role of platforms as artificial reefs supporting diverse fish communities and highlight the limited knowledge of biodiversity associated with artificial structures in the region. They provide a valuable baseline to inform future RTR decisions in the Arabian Gulf.
1 Introduction
Similar to other seas worldwide, the Arabian Gulf (the Gulf) is experiencing a decline in marine biodiversity (Sheppard et al., 2010). This decline is mainly caused by a loss of coral reef habitats due to recurring coral heat-related bleaching events (Rezai et al., 2004; Riegl and Purkis, 2012; Hereher, 2020; Aeby et al., 2024) and the extensive coastal development of Gulf countries in recent years (Bartholomew and Feary, 2012; Burt and Bartholomew, 2019).
Artificial offshore structures, such as oil and gas platforms, are increasingly recognized for their habitat value for marine species, and some have been well studied due to their high diversity and abundance of both sessile and pelagic marine organisms (Claisse et al., 2014; Vidal et al., 2022). While the degradation of natural coral reefs leads to a loss of both structural and biological complexity, submerged platform jackets offer a large, hard, and complex substrate for a wide variety of benthic invertebrates (Friedlander et al., 2014; Love et al., 2019). The vertical orientation of these platforms, extending from the seafloor through the surface, promotes conditions for benthic growth across the entire water column, facilitating the depth zonation of marine organisms and thereby supporting greater diversity (Torquato et al., 2017, 2021). Fish assemblages on artificial structures, including offshore platforms, often display distinct vertical zonation patterns, with certain species preferring specific depths or structural features throughout the water column. Studies on other artificial reefs have shown that predatory and reef-associated species often aggregate at midwater crossbeams, while juvenile and small-bodied fish may be more abundant in the shallow or structurally complex zones (e.g., around jackets and cross-bracings) that offer protection from predators (Consoli et al., 2013; van Elden et al., 2022). This zonation can mirror natural reef patterns, where habitat complexity and light availability influence fish distribution, suggesting that platforms could functionally mimic natural reef habitats in supporting biodiversity across different depth strata.
Studies across the world’s oceans, in both tropical (Rilov and Benayahu, 2000; Alexander et al., 2022; van Elden et al., 2022) and temperate waters (Birt et al., 2024; Karwowski et al., 2024), have revealed that oil and gas platforms host fish assemblages with higher diversity and abundance compared to surrounding areas, similar to the way natural reefs support increased diversity. To some extent, the high diversity and abundance of fishes associated with offshore platforms maybe attributed to the strict security measures enforced in offshore oil and gas extraction areas. These security protocols include a complete ban on fishing within the oilfields. Thus, the offshore platforms inadvertently act as de facto no-take marine reserves (Love et al., 2003; Kashubsky and Morrison, 2013; Cordes et al., 2016). Offshore platforms may have the potential to contribute significantly to local fish productivity and support fisheries management goals. Research has shown that platforms can support both juvenile and adult fish populations, potentially boosting local fish productivity (Fujii, 2016). Thus, due to their high productivity and moderate site fidelity, platform-associated fish communities may contribute to the supply of fish to fisheries in surrounding natural habitats. Moreover, platforms may offer essential nursery habitats, particularly in regions where natural reefs are degraded, thereby contributing to the recruitment and sustainability of key fish species (Fowler and Booth, 2012). These potential benefits align well with regional fisheries management strategies and underscore the value of incorporating platforms into broader conservation and fisheries frameworks in the Gulf region. In this context, the Rigs-to-Reefs (RTR) programme, which involves converting decommissioned oil platforms into permanent artificial reefs, has risen in popularity as a viable strategies for enhancing and supporting marine biodiversity (Dauterive, 1999; Schroeder and Love, 2004; Cordes et al., 2016; Riera et al., 2023).
The vast majority of economic income for the southern Gulf states (Bahrain, Kuwait, Saudi Arabia, UAE, and Qatar) is derived from offshore oil and gas extraction (Peterson, 2009). The Gulf hosts thousands of oil and gas industry structures, ranging from loading terminals to pipelines (Feary et al., 2011), including approximately 800 offshore platforms in 2010 (Sheppard et al., 2010) and more than 2000 by 2019 (Burt and Bartholomew, 2019). The benthic growth on the submerged structures of oil and gas platforms in the Gulf is dominated by filter and suspension-feeding invertebrates, mainly barnacles, bryozoans, and bivalves, with coral growth also observed on the structures (Stachowitsch et al., 2002; Torquato et al., 2021). From platform jackets in Qatar, Range et al. (2018) described the first observation of an azooxanthellate scleractinian (possibly of the Caryophylliidae family) in the Gulf. Offshore platforms are suggested to act as important stepping stones for the dispersal and connectivity of marine organisms in an increasingly fragmented network of natural coral reefs (Burt et al., 2009; Bartholomew and Feary, 2012; Friedlander et al., 2014; Schutter et al., 2019). However, such connectivity have been invoked to increase the risk of spreading invasive or non-native species (Schulze et al., 2020). Other studies have shown that platforms in the southern part of the Gulf might act as nurseries for juvenile fishes (Rabaoui et al., 2015). A major oilfield in the central Gulf, Al Shaheen oilfield, lies at the heart of an important feeding ground for one of the world’s largest annual aggregations of whale sharks (Rhincodon typus) (Robinson et al., 2013; Bach et al., 2016). However, the potential connection between the platforms and the availability of tuna eggs, a key food source for the whale sharks, remains unclear.
The platforms of Al Shaheen oilfield have been the focus of biodiversity studies, particularly concerning their associated fish and invertebrate fauna. Torquato et al. (2017) identified 83 taxonomic groups of fish, including two species (Cyclichthys orbicularis and Lutjanus indicus) recorded for the first time in the area. The study also described patterns in the vertical zonation of fish species, with the greatest species richness found between 20 m and 50 m, while overall abundance decreased with depth. Strong vertical stratification has also been observed for invertebrates on these platforms, and the benthic community composition has shown considerable variation over time, with older platforms harboring significantly different invertebrate communities compared to newer ones (Torquato et al., 2021). Both studies of the Al Shaheen platforms reported high diversity and abundance of marine organisms, with species richness of fishes exceeding that of surrounding natural reefs (Torquato et al., 2017). However, Riera et al. (2023) argue that these platforms cannot be considered as direct surrogates for natural reefs, as the species compositions and functional diversity are significantly different.
The studies of the platforms in Al Shaheen were conducted using footage from remotely operated vehicles (ROVs) employed by the company inspecting the structural integrity of the platforms (Torquato et al., 2017, 2021). Consequently, the recordings were not originally intended for biodiversity studies. As a result, the survey design and camera resolution had limitations, affecting the observation and accurate identification of certain species. In this study, we surveyed the fish fauna associated with platforms in a different Qatari oilfield, Al Khalij. We employed high-resolution 4K cameras attached to ROVs, following a survey protocol specifically aimed at fish species identification and quantification. The full water column, from the surface to the bottom, was surveyed along the legs of platforms, with recordings taken during both day and night. While documenting the fish fauna of the Al Khalij oilfield for the first time, we hypothesize that the improved sampling design implemented in this biodiversity study will be more efficient and enable more accurate species identification than previous ROV studies conducted in the Gulf.
2 Methods
2.1 Study area
The Gulf is a shallow, semi-enclosed sea with low species richness due to extreme environmental conditions and a short geological history (Price, 2002; Coles, 2003; Sheppard et al., 2010). Covering 240,000 km² with an average depth of 35 m, it experiences extreme temperatures (15–36°C), high salinity (40–50 ppt), and limited freshwater input, connected to the Arabian Sea only via the narrow Strait of Hormuz (Price et al., 1993; Sheppard et al., 1992).
Al Khalij oilfield is located in the central part of the Gulf, within the Exclusive Economic Zone (EEZ) of Qatar, approximately 130 km east of the mainland (Figure 1). The oilfield was discovered in 1991, and production began in 1997 (QatarEnergy, 2020). The seabed is sandy and depth ranges from 55 m to 60 m. The nearest island, Halul, and the nearest known natural coral reefs are located approximately 50 km south of the oilfield (Rezai et al., 2004; Burt et al., 2017). Al Khalij consists of eight individual platforms interconnected by submerged cables and pipelines along the seabed. The fish fauna associated with two platforms, DP3 and DP4, located 10 km apart, was surveyed in the present study. DP4 is situated in the northern part of the oilfield, while DP3 is located centrally. Both platforms are similar in size, with approximately 25 m between each leg at the base. Horizontal and diagonal crossbeams connect the legs at both platforms, with attachments at around 12 m and 35 m, increasing the structural complexity at these depths.
Figure 1
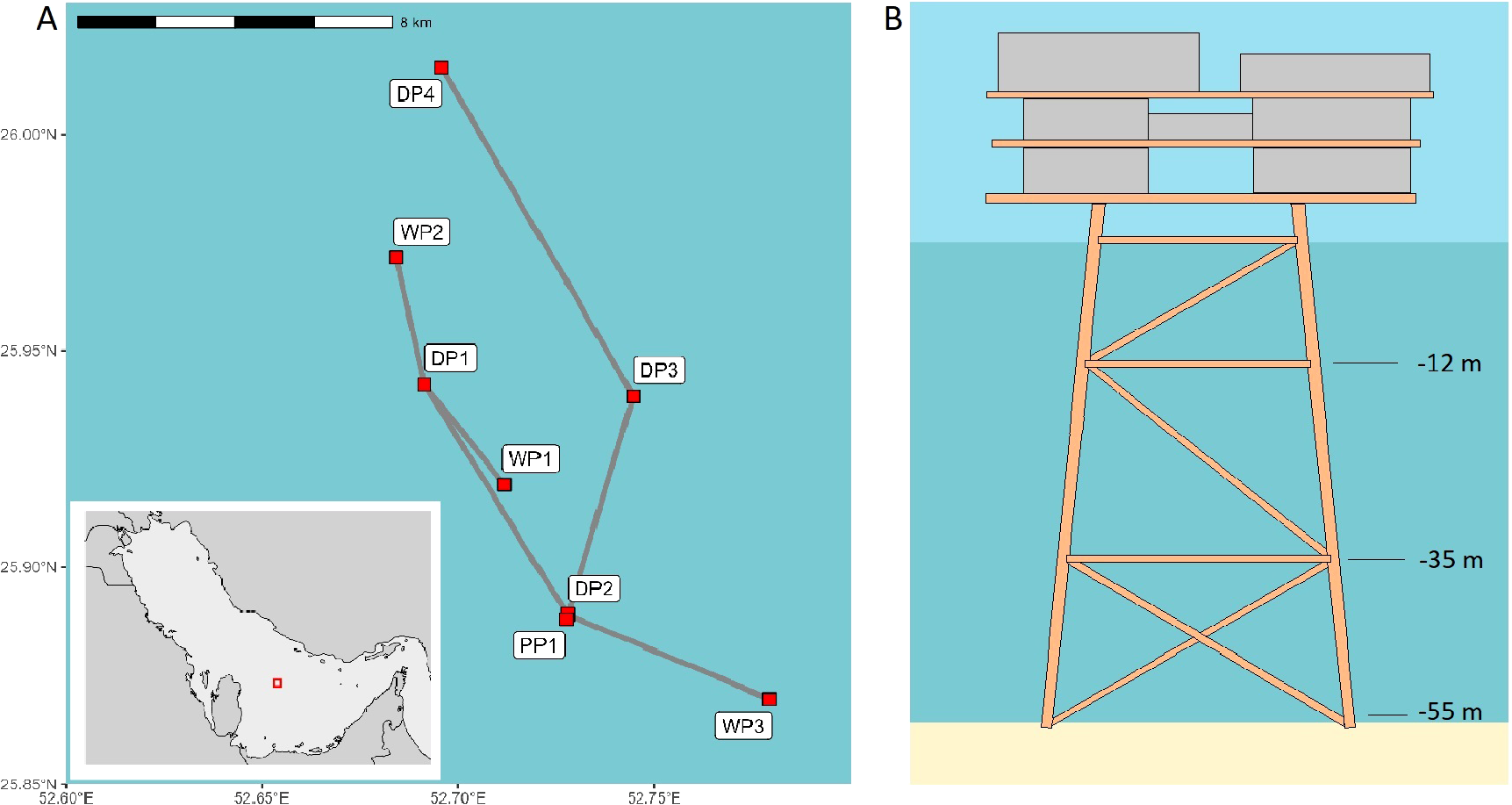
(A) Map of Al Khalij oilfield located 110 km west of Qatar, illustrating the positions of each platform (red squares) and their connections via submerged pipelines (grey lines). (B) Diagram depicting a simplified platform jacket layout of DP3 and DP4, showing crossbeam connections at depths of approximately 12 m and 35 m.
The survey was conducted in April 2022. During this month, sea surface temperatures in the area remain below 25°C (Riegl and Purkis, 2012). A seasonal thermocline and halocline develop between 15 m and 30 m in the area during the summer, beginning in April (Al-Ansari et al., 2022).
2.2 Sampling design
Four survey transects were performed at each of the platforms, one transect at each platform leg. All transects were sampled within a 24-hour period. One leg at platform DP3 was recorded during daylight in the afternoon of 22 April 2022, while the remaining three legs were recorded the following night. All four legs of DP4 were recorded in daylight on the morning of 23 April 2022. The survey transects were conducted using an ROV equipped with a GoPro Hero10 Black camera recording in 4K at 30 frames per second. For night-time transects, the ROV was equipped with lights to ensure proper visibility. Positioned three meters away from the platform leg, the ROV captured footage as it descended from the surface to the bottom, keeping the structures centered in the field of view. Each descent took approximately 5–10 min. At one of the DP3 legs recorded at night, the ROV was unable to capture the surface layers (0–15 m) due to rough weather conditions at the time.
Each transect recording was segmented into 5 m depth intervals and analyzed independently. Using the SeaGIS EventMeasure (v. 6.43) software, fishes observed within each depth interval were counted and identified to the lowest taxonomical unit possible. The maximum number of individuals for each taxon observed in a single frame (MaxN) was determined for each depth interval (Ellis and DeMartini, 1995).
Total biomass for each observed taxon was estimated by assigning 10% of randomly selected individuals, or at least one individual per taxon, to predefined size classes. The average mass for each taxon’s size class was primarily determined using total lengths of fishes caught at the Al Khalij platforms. Most of the fish caught were near the maximum recorded length for their species, which was accounted for when assigning individuals to size classes, with the majority placed in the maximum size class. Additionally, size class biomass was estimated using relevant literature, length-weight relationship functions available for most species in the FishBase database (FishBase.se ver. 06/2024), and measurements from museum specimens at the Natural History Museum of Denmark. The total biomass of each taxon at each depth was then extrapolated from the average mass of the 10% sample. The data on fish caught at the platforms used for biomass estimations are provided in the Supplementary Material Table 1), along with the estimated biomass of each species Supplementary Material Table 2.
To account for the decreasing visibility with decreasing light levels at greater depths and to test the effect of visibility on species richness and abundance, the volume of water in the field of view (FoV) of the camera was calculated for each segment. The volume of the camera’s pyramid-shaped FoV was computed using known vertical () and horizontal () angles of the camera, along with the estimated visibility length (H), according to the formula:
Additionally, the volume of the triangular prism-shaped FoV formed by the vertical movement of the ROV (L) was determined by:
The total recorded volume was then calculated as the sum of the volumes of the pyramid FoV and the triangular prism FoV:
The percentage of the FoV obstructed by the platform structure was estimated and subtracted from the total volume, resulting in the actual surveyed volume of water. The total FoV calculation is illustrated in Figure 2.
Figure 2
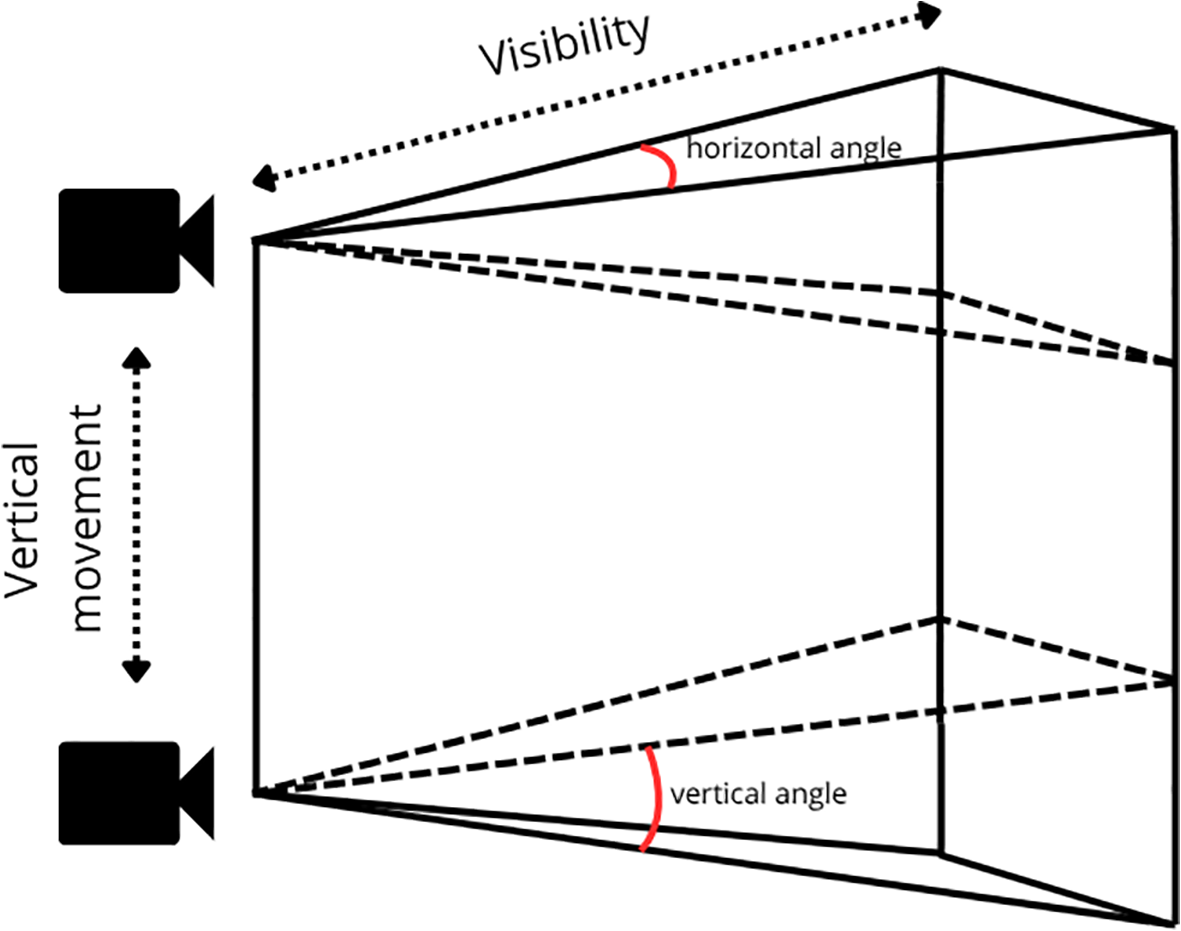
Illustration of the surveyed volume of water within the camera’s field of view. The total volume for each sample consists of a pyramid-shaped field of view defined by the camera’s vertical and horizontal angles and the recorded visibility distance. The ROV’s five-metre vertical movement adds a triangular prism to the total surveyed volume.
2.3 Data analysis
Several analyses of fish assemblages associated with the offshore platforms were conducted, focusing on patterns in diversity, relative abundance, and biomass. Species accumulation plots were generated to illustrate how species richness accumulates over the total observed FoV and time. A cut-off of 95% of the total observed species richness was used to determine when sampling effort was sufficient to capture the majority of the diversity. These plots offer insights into the sampling effort required to capture the full diversity present, highlighting the efficiency of the survey design in documenting species richness.
The median, interquartile range, and outliers of the depth distribution were calculated for each taxon to highlight species’ depth preferences around the platforms.
The patterns of species composition across different depths and platforms were investigated using non-metric multidimensional scaling (NMDS) based on Bray-Curtis dissimilarity, implemented in the vegan package in R (metaMDS, distance = “bray”, K = 3). The genera contributing to the observed variation were visualized using principal component analysis (PCA), performed with the base R function prcomp. For PCA, genus abundance data were log-transformed (log1p) and standardized, with genera showing variance below 2.4 excluded. For NMDS, all taxa at the lowest taxonomic level were included, while species were merged into their respective genera for PCA. Both analyses were performed exclusively on daytime samples, excluding unidentified individuals. Additionally, one daytime sampling point (DP3, Leg A1, 45m–50m) was omitted due to the recording of only one individual (Carcharhinus sp.), which distorted the visualization of the main patterns.
Biomass and total MaxN were summarized at each depth interval to compare biomass and relative abundance across depths, platforms, and times of day. Generalized additive models (GAMs) and linear regression models were applied to explore relationships between depth, FoV, abundance, and species diversity in daytime transects. GAMs, implemented with the gam function in the mgcv package in R, examined non-linear relationships. When relationships involving FoV were found to be linear, linear regression models were applied using the lm function in R. These analyses provided insights into how depth affects abundance and species diversity and how sampling efficiency (including visibility and surveyed water volume) influences species detection and abundance.
The average biomass per platform jacket was calculated by summing the biomass of all daytime transects and dividing by the number of transects. The estimated biomass per platform was then determined by multiplying the average jacket biomass by four.
3 Results
Combining all depth intervals of all transects at both platforms resulted in a total of 67 min of analyzed recordings, with the FoV estimation totaling 28,296 m³ surveyed. The species accumulation curves peaked after 25 min or 25,000 m³ of water surveyed, as shown in Figure 3. The total MaxN for all species across both platforms, transects, and depth intervals was 4,474 individuals, with an estimated total biomass of 8,270kg, equivalent to 292 g per m³ surveyed. During the daytime transects, the average biomass per leg was 1,546 kg (SE = 179 kg), resulting in an estimated 6,184 kg of fish biomass per platform and 103 kg per meter of submerged vertical structure (60 m).
Figure 3
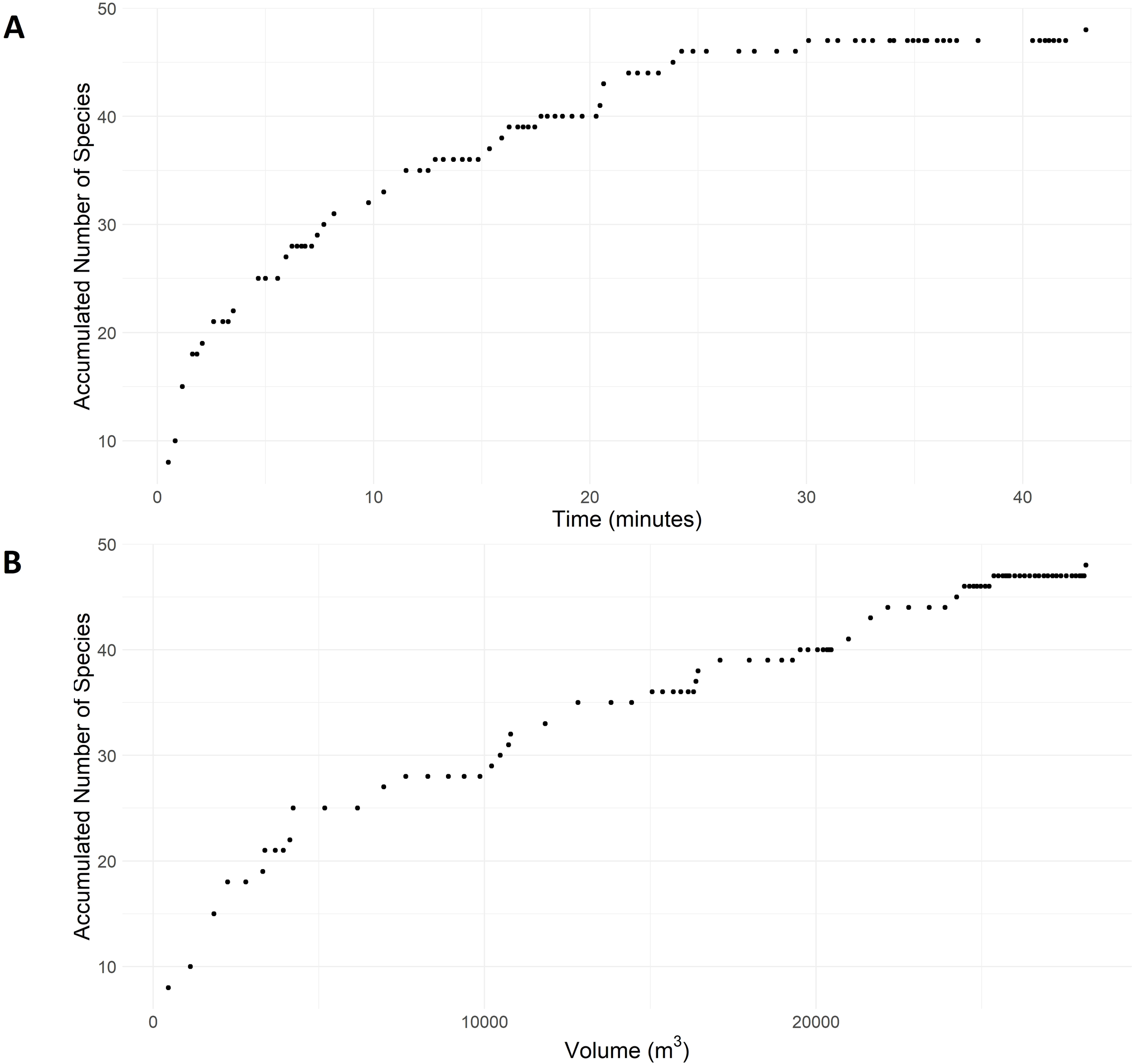
Species accumulation curves showing accumulated number of species as a function of time (A) and volume of water (B).
3.1 Species composition
Out of the 4,474 observed fishes, 3,549 were identified to at least family level, while 925 individuals remained unidentified. In total, 46 different taxa were observed, comprising 32 species, 30 genera, and 19 families. Carangidae (8 genera, 10 species) and Serranidae (2 genera, 4 species) were the most diverse families. The most abundant species identified was Pinjalo pinjalo (Lutjanidae), with a total MaxN of 486. Other abundant species included Scomberoides commersonnianus (Carangidae), Lutjanus argentimaculatus (Lutjanidae), Heniochus acuminatus (Chaetodontidae), Flavocaranx bajad (Carangidae), Pomacanthus maculosus (Pomacanthidae), and Platax teira (Ephippidae). During one of the night transects, a large school of Encrasicholina sp. (Engraulidae) was observed, with a total MaxN of 1,123. The genus Platycaranx was also prevalent across all transects, with a total MaxN of 199, but it was rarely possible to identify individuals to the species level. All observed taxa, along with size class proportions and estimated biomass, are presented in Table 1.
Table 1
| Family | Species | Common Name | Total MaxN | Percentage in Small Size Class | Percentage in Medium Size Class | Percentage in Maximum Size Class | Total Biomass (g) | Average Weight (g) |
|---|---|---|---|---|---|---|---|---|
| Carangidae | Alepes vari | Herring scad | 62 | 0.0% | 0.0% | 100% | 119692 | 1931 |
| Carangidae | Caranx sexfasciatus | Bigeye trevally | 12 | 0.0% | 100% | 0.0% | 76735 | 6395 |
| Carangidae | Caranx spp. | Jacks | 3 | 0.0% | 100% | 0.0% | 16250 | 5417 |
| Carangidae | Caranx ignobilis | Giant trevally | 2 | 0.0% | 100% | 0.0% | 12012 | 6006 |
| Carangidae | Caranx heberi | Blacktip trevally | 1 | 0.0% | 100% | 0.0% | 3849 | 3849 |
| Carangidae | Elagatis bipinnulata | Rainbow runner | 1 | 100% | 0.0% | 0.0% | 1112 | 1112 |
| Carangidae | Flavocaranx bajad | Yellowfin jack | 174 | 0.0% | 17.6% | 82.4% | 186206 | 1070 |
| Carangidae | Gnathanodon speciosus | Golden trevally | 1 | 100% | 0.0% | 0.0% | 3129 | 3129 |
| Carangidae | Platycaranx spp. | Trevallies | 141 | 28.6% | 71.4% | 0.0% | 504840 | 3580 |
| Carangidae | Platycaranx chrysophrys | Longnose trevally | 58 | 33.3% | 66.7% | 0.0% | 201434 | 3473 |
| Carangidae | Scomberoides commersonnianus | Talang queenfish | 356 | 0.0% | 72.2% | 27.8% | 1581559 | 4443 |
| Carangidae | Seriola dumerili | Greater amberjack | 1 | 0.0% | 100% | 0.0% | 9794 | 9794 |
| Carangidae | Carangidae unknown | Jacks and pompanos | 55 | 0.0% | 100% | 0.0% | 269660 | 4903 |
| Carcharhinidae | Carcharhinus spp. | Requiem sharks | 2 | 0.0% | 0.0% | 100% | 24000 | 12000 |
| Chaetodontidae | Chaetodon nigropunctatus | Blackspotted butterflyfish | 1 | 0.0% | 0.0% | 100% | 55 | 55 |
| Chaetodontidae | Heniochus acuminatus | Longfin bannerfish | 218 | 0.0% | 78.3% | 21.7% | 34985 | 160 |
| Echeneidae | Echeneis naucrates | Live sharksucker | 1 | 0.0% | 0.0% | 100% | 1267 | 1267 |
| Engraulidae | Encrasicholina sp. | Anchovies | 1123 | 0.0% | 100% | 0.0% | 8445 | 8 |
| Ephippidae | Platax teira | Longfin batfish | 106 | 0.0% | 0.0% | 100% | 95400 | 900 |
| Ephippidae | Platax spp. | Batfishes | 15 | 0.0% | 50.0% | 50.0% | 6750 | 450 |
| Ephippidae | Platax orbicularis | Orbicular batfish | 3 | 0.0% | 0.0% | 100% | 2700 | 900 |
| Labridae | Cheilinus trilobatus | Tripletail wrasse | 2 | 0.0% | 0.0% | 100% | 4800 | 2400 |
| Labridae | Thalassoma lunare | Moon wrasse | 4 | 0.0% | 100% | 0.0% | 448 | 112 |
| Lethrinidae | Lethrinus spp. | Emperors | 3 | 0.0% | 100% | 0.0% | 6287 | 2096 |
| Lethrinidae | Lethrinus nebulosus | Spangled emperor | 1 | 0.0% | 100% | 0.0% | 2096 | 2096 |
| Lutjanidae | Pinjalo pinjalo | Pinjalo | 486 | 0.0% | 32.7% | 67.3% | 654612 | 1347 |
| Lutjanidae | Lutjanus argentimaculatus | Mangrove red snapper | 352 | 0.0% | 100% | 0.0% | 1204477 | 3422 |
| Lutjanidae | Lutjanus spp. | Snappers | 8 | 0.0% | 100% | 0.0% | 27374 | 3422 |
| Ostraciidae | Ostracion cyanurus | Blue boxfish | 1 | 0.0% | 0.0% | 100% | 93 | 93 |
| Ostraciidae | Ostracion cubicum | Yellow boxfish | 1 | 0.0% | 100% | 0.0% | 188 | 188 |
| Pomacanthidae | Pomacanthus maculosus | Yellowbar angelfish | 113 | 0.0% | 81.8% | 18.2% | 117818 | 1043 |
| Pomacentridae | Chromis spp. | Damselfishes | 13 | 0.0% | 100% | 0.0% | 139 | 11 |
| Scaridae | Scarus spp. | Parrotfishes | 7 | 0.0% | 0.0% | 100% | 17819 | 2546 |
| Scaridae | Scarus persicus | Persian parrotfish | 1 | 0.0% | 0.0% | 100% | 2546 | 2546 |
| Scombridae | Scomberomorus commerson | Narrow-barred Spanish mackerel | 5 | 0.0% | 100% | 0.0% | 113689 | 22738 |
| Scorpaenidae | Pterois miles | Devil firefish | 7 | 0.0% | 100% | 0.0% | 3955 | 565 |
| Serranidae | Cephalopholis hemistiktos | Yellowfin hind | 10 | 0.0% | 0.0% | 100% | 4700 | 470 |
| Serranidae | Epinephelus spp. | Groupers | 35 | 0.0% | 100% | 0.0% | 167293 | 4780 |
| Serranidae | Epinephelus chlorostigma | Brownspotted grouper | 33 | 0.0% | 100% | 0.0% | 37633 | 1140 |
| Serranidae | Epinephelus coioides | Orange-spotted grouper | 7 | 0.0% | 100% | 0.0% | 58935 | 8419 |
| Serranidae | Epinephelus areolatus | Areolate grouper | 1 | 0.0% | 0.0% | 100% | 960 | 960 |
| Siganidae | Siganus sp. | Rabbitfish | 3 | 0.0% | 0.0% | 100% | 578 | 193 |
| Sparidae | Acanthopagrus bifasciatus | Twobar seabream | 55 | 0.0% | 100% | 0.0% | 107304 | 1951 |
| Sphyraenidae | Sphyraena qenie | Blackfin barracuda | 21 | 0.0% | 100% | 0.0% | 18450 | 879 |
| Sphyraenidae | Sphyraena jello | Pickhandle barracuda | 5 | 0.0% | 0.0% | 100% | 51101 | 10220 |
| Sphyraenidae | Sphyraena spp. | Barracudas | 2 | 0.0% | 0.0% | 100% | 20440 | 10220 |
| Unknown | Unknown | 764 | 5.6% | 56.8% | 37.6% | 2484575 | 3252 | |
| Unknown | Cryptobenthic | 162 | 0.0% | 100% | 0.0% | 1727 | 11 |
Observed species, including abundance (Total MaxN), percentage of individuals within each size class (Percentage in Small Size Class, Percentage in Medium Size Class, and Percentage in Maximum Size Class), estimated total biomass (Total Biomass), and average weight per taxon (Average Weight). Size class definitions for each taxon are provided in the Supplementary Material. All weight estimates are presented in grams (g).
3.2 Diversity, abundance, and biomass across depth gradients
A clear vertical zonation in species assemblage was observed with increasing depth. The vertical zonation is illustrated in Figure 4. At shallow depths near the surface, a greater diversity of Carangidae species was noted. Particularly species of the Caranx genus showed a clear preference for the surface layers. Scomberoides commersonnianus was also predominantly associated with the surface. Lutjanus argentimaculatus, Flavocaranx bajad, and Pomacanthus maculosus were observed throughout the entire water column, but the majority of individuals of these species were found at shallower depths. The Platax and Sphyraena genera were mainly observed in the top layers, but their distribution was deeper than the aforementioned species, primarily between 5 m and 20 m.
Figure 4
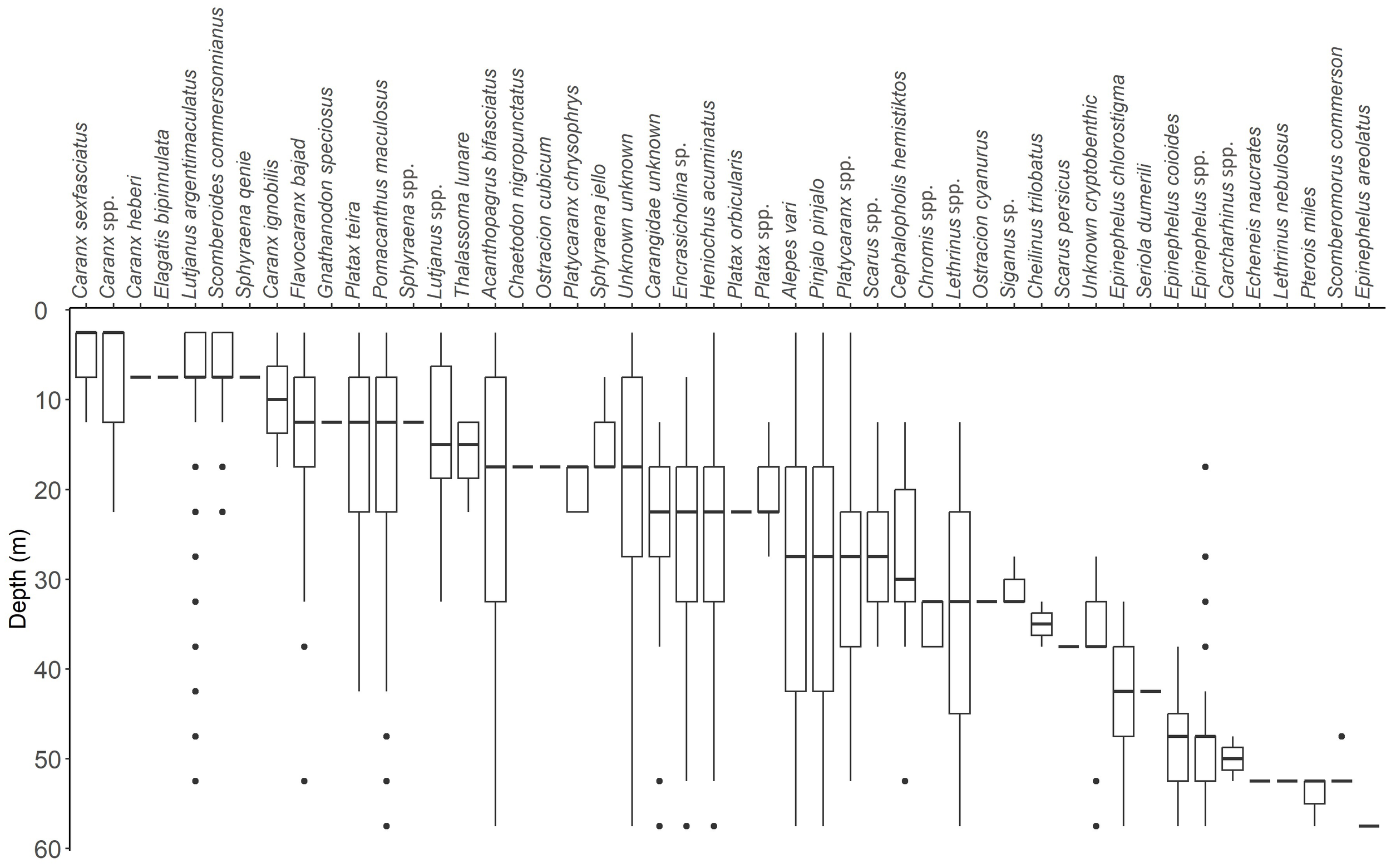
Box plots illustrating the depth distribution of each observed taxon. The box plots display the data distribution with the whiskers extending to 1.5 times the interquartile range (IQR) from the first and third quartiles. Points outside this range are shown as outliers.
At the seafloor, fishes of the Epinephelus genus were dominant, though some members of the genus were also observed at shallower depths, within the platform structure, occasionally reaching up to 20 m. Pterois miles (Scorpaenidae), Scomberomorus commerson (Scombridae), and Carcharhinus spp. (Carcharhinidae) were only recorded near the seafloor, although Carcharhinus spp. were frequently observed near the surface by the authors but not captured in the recordings.
Alepes vari (Carangidae), Acanthopagrus bifasciatus (Sparidae), Encrasicholina sp., Heniochus acuminatus, Pinjalo pinjalo, and Platycaranx spp. (Carangidae), which were all among the most abundant taxa, were recorded throughout the entire water column with no clear indication of a preferred depth.
A number of taxa were primarily recorded in areas of increased structural complexity. These areas were primarily where horizontal and diagonal crossbeams connected the platform legs (at 12 m and 35 m), and at the seafloor. The recorded taxa included Chaetodon nigropunctatus (Chaetodontidae), Cheilinus trilobatus (Labridae), Chromis spp. (Pomacentridae), Ostracion cubicum, O. cyanurus (Ostraciidae), Scarus persicus (Scaridae), Siganus sp. (Siganidae), and unidentified small benthic species, presumably mainly species of Chromis and Pseudanthias. Coral growth were also observed in some of these areas of increased structural complexity.
All depth intervals showed similar species richness, with 8 to 14 species observed at each interval. There was no clear pattern of decreasing or increasing species number correlated to depth.
The NMDS analysis of all daytime sampling points (except DP3, Leg A1, 45m-50m) demonstrated a clear pattern of species assemblage changing with depth. These patterns are illustrated in Figure 5. Shallow points clustered together at one end, distinctly separated from the deep samples, with intermediate depths positioned in between. Bray-Curtis dissimilarity calculations revealed greater similarity among shallow sampling points, indicating that deep sampling points were more varied from each other. PCA analysis revealed that the genera contributing most to the observed variance were Epinephelus, Flavocaranx, Lutjanus, Pinjalo, Platax, Pomacanthus, Scomberoides, and the cryptobenthic group. Flavocaranx, Pinjalo, Platax, and Scomberoides were indicative of the surface layers at both platforms, while deep water samples, particularly from DP3, were dominated by the Epinephelus genus. Midwater layers around 35 m on DP4 were characterized by the presence of unidentified cryptobenthic species.
Figure 5
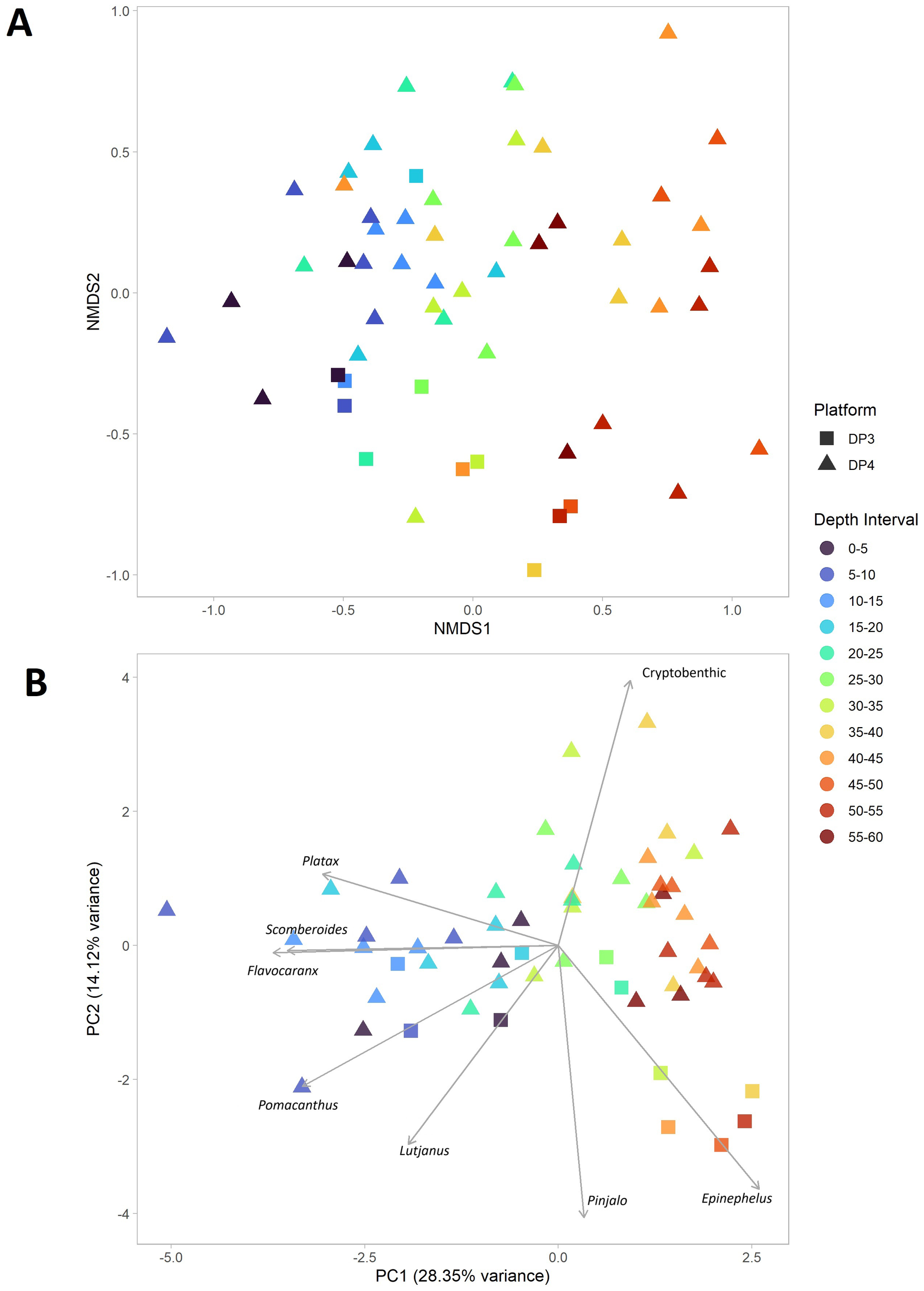
NMDS plot (A) based on Bray-Curtis dissimilarity (K = 3, stress = 0.16) illustrating species composition patterns across all daytime sampling points. PCA plot (B) derived from daytime genus abundance data, following variance thresholding, log transformation, and outlier removal. Both plots exclude DP3, Leg A1, 45m-50m, as this sample, consisting of only one observation, distorted the visualization of the main dynamics.
Both total abundance and biomass of all observed species combined decreased with depth, as shown in Figure 6. At shallow depths (0m-25m), above the potential thermocline or halocline, the average MaxN per depth interval was 580.4 (SE = 43.7), decreasing to 324.7 (SE = 27.4) at the mid layers (25m-40m), and further decreasing to 140.5 (SE = 16.3) at the bottom layers (40m-60m). However, an increase in MaxN was noted at the seafloor level and adjacent (50m-60m) compared to the immediately overlying level (40m-50m). The daytime transect conducted at DP3 deviated from the general pattern, with peaks in abundance and biomass observed around 10 m, 35 m, and at the seafloor.
Figure 6
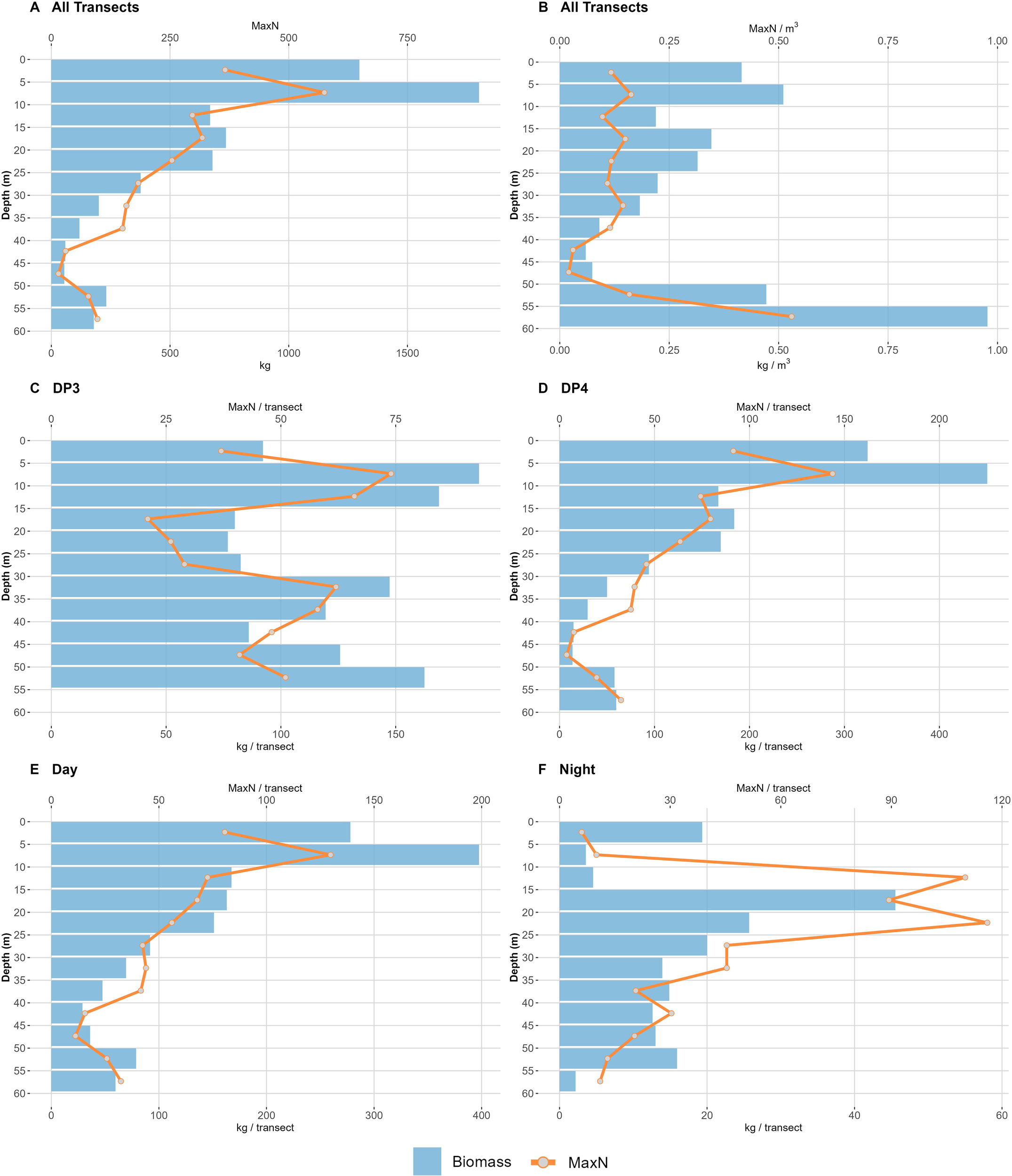
Bar plots illustrating (A) total biomass and total abundance across all transects combined, (B) biomass per m³ and abundance per m³ for all transects combined, (C) biomass per transect and abundance per transect for the DP3 daytime transect, (D) biomass per transect and abundance per transect for DP4 daytime transects, (E) biomass per transect and abundance per transect for all daytime transects, and (F) biomass per transect and abundance per transect for all night transects.
When accounting for the decreasing FoV due to reduced visibility at greater depths, biomass still decreased between the surface and 50 m. However, abundance counts remained relatively consistent from the surface to 35 m, before declining between 35 m and 50 m, when evaluated in the context of the reduced volume of water surveyed. Both MaxN/m³ and kg/m³ peaked at the bottom.
Transects conducted at night recorded both fewer taxa and a lower abundance and biomass than the daytime transects. The total MaxN for all species recorded at night was 1,404, of which 1,349 were identified to at least the family level, while only 55 individuals could not be identified. In total, 16 different taxa were observed, comprising 10 species, 13 genera, and 10 families. Only one taxon, Encrasicholina sp., was not observed on daytime transects. Encrasicholina sp. was by far the most abundant taxon recorded, with a total MaxN of 1,123 (79.9% of all individuals recorded at night). Despite the peak in abundance and biomass caused by schooling Encrasicholina sp., night transects displayed a more evenly distributed abundance and biomass, with no clear pattern of increasing or decreasing values with depth, unlike the daytime transects.
3.3 The effect of depth and visibility
The relationships between depth, surveyed water volume (FoV), species diversity (number of species), and abundance (MaxN) revealed significant non-linear relationships (GAM edf < 1.5, p < 0.05) for MaxN and number of species as a function of depth, as well as for species diversity as a function of MaxN (Table 2; Figure 7). Although both were significant, number of species was better explained by MaxN (72.9% deviance explained) compared to depth (40% deviance explained). The GAMs indicated linearity in all cases where FoV was either the dependent or independent variable, with effective degrees of freedom (edf) and reference degrees of freedom (Ref.df) close to 1. A clear trend of decreasing visibility with depth was demonstrated by the highly significant negative correlation between FoV and depth (slope = -14.18, p < 0.001, R² = 0.78) (Table 3). Although very low p-values suggested a statistical relationship between FoV and species diversity or MaxN, the slope estimates near zero and low R²-values indicate that visibility had a minimal impact on the observed number of species and individuals.
Table 2
| Dependent Variable | Independent Variable | edf | Ref.df | F-statistic | p-value | Adjusted R² | Dev. Explained |
|---|---|---|---|---|---|---|---|
| FoV | Depth | 1.000 | 1.000 | 165.5 | < 0.001 | 0.743 | 74.7% |
| MaxN | Depth | 1.671 | 2.080 | 17.16 | < 0.001 | 0.380 | 39.9% |
| Number of species | Depth | 5.488 | 6.643 | 4.736 | < 0.001 | 0.336 | 40.0% |
| Number of species | FoV | 1.000 | 1.000 | 5.740 | 0.020 | 0.077 | 9.3% |
| MaxN | FoV | 1.301 | 1.540 | 18.15 | < 0.001 | 0.305 | 32.1% |
| Number of species | MaxN | 6.503 | 7.517 | 17.52 | < 0.001 | 0.694 | 72.9% |
Generalized Additive Models (GAMs) showing the relationships between Depth, surveyed water volume (FoV), species diversity (number of species), and abundance (MaxN). The table includes effective degrees of freedom (edf), reference degrees of freedom (Ref.df), F-statistics, p-values, adjusted R², and deviance explained (Dev. Explained).
Table 3
| Dependent Variable | Independent Variable | Estimate (Slope) | Std. Error | t-value | p-value | R² | Adjusted R² |
|---|---|---|---|---|---|---|---|
| FoV | Depth | -14.183 | 1.103 | -12.86 | < 0.001 | 0.747 | 0.748 |
| Number of species | FoV | 0.003 | 0.001 | 2.396 | 0.02 | 0.093 | 0.077 |
| MaxN | FoV | 0.09 | 0.018 | 5.044 | < 0.001 | 0.312 | 0.300 |
Linear regression models showing the relationship between variables with linearity proven by the GAMs. Linear relationships between Depth, surveyed water volume (FoV), species diversity (number of species), and abundance (MaxN) are presented with the estimated coefficient (Estimate (Slope)), standard errors (Std. Error), t-values, p-values, R², and adjusted R².
Figure 7
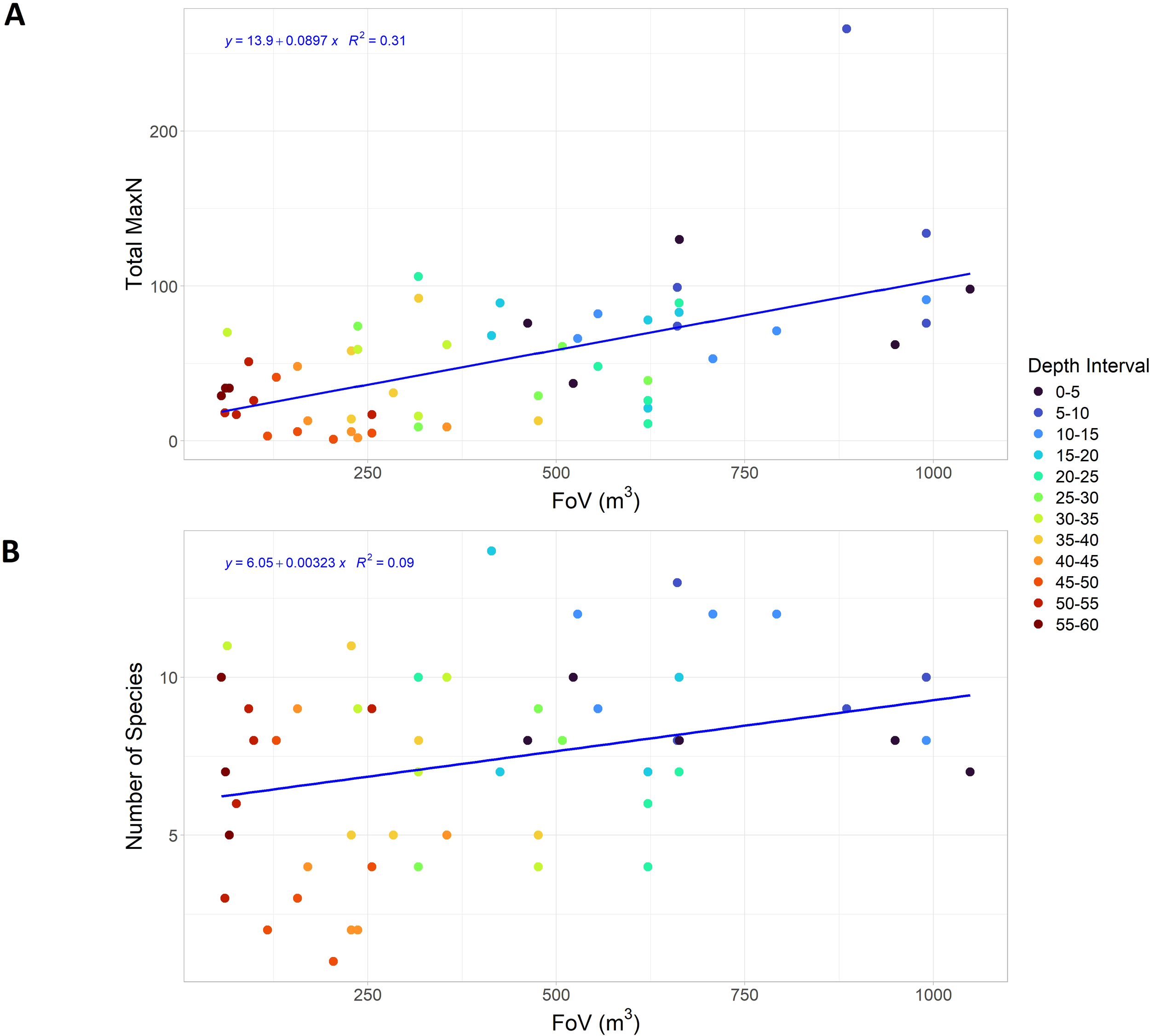
Linear regression plots showing the relationship between abundance (A) and species diversity (B) as a function of surveyed water volume.
3.4 Other observations
Beside species identifications, abundance counts, and biomass estimates, several other noteworthy observations were made during the analysis of the video footage. Strong suggestions of spawning behavior in Caranx sexfasciatus, Caranx ignobilis and Platycaranx spp. (Carangidae) was frequently observed, indicated by the color changes in the males (Graham and Castellanos, 2005; Allen and Erdmann, 2012; Madgett et al., 2022). A spawning pair of Caranx sexfasciatus was recorded near the surface of DP4, and a spawning male Caranx ignobilis was observed at DP3 during a night transect. Groups of Platycaranx spp., each with a spawning male, were observed multiple times throughout the entire water column at both platforms.
In addition to spawning behavior, predation was observed on multiple occasions. Direct instances of predation were particularly evident during the night transects, where Pinjalo pinjalo and Epinephelus spp. (Epinephelidae) were recorded feeding on Encrasicholina sp. During a daytime transect at 10 m depth around DP4, a group of 21 Sphyraena qenie (Sphyraenidae) was recorded actively chasing Flavocaranx bajad and other Carangidae species.
One individual of Cheilinus trilobatus was observed near the platform crossbeam connections at a depth of 35 m on DP3. The highest coverage of coral growth was also noted at the corner of this crossbeam connection. Although C. lunulatus is the only species of the genus previously recorded within the Gulf, the distinctive white band across the peduncle confirmed the identification as C. trilobatus. This observation represents the first recorded occurrence of C. trilobatus in the Gulf, marking a potentially significant extension of its known range (FishBase.se ver. 06/2024; GBIF.org December 2024).
Video segments showing spawning behavior, predation, hard coral growth, and the observed Cheilinus trilobatus are provided in the Supplementary Materials.
4 Discussion
This study presents the first assessment of the fish assemblages associated with offshore platforms in the Al Khalij oilfield, Qatar. Utilizing a survey protocol specifically designed for biodiversity detection, and employing high-resolution cameras attached to ROVs, the study identified diverse fish assemblages at the Al Khalij platforms and described a clear stratification of species with depth. Additionally, this research provides the first estimation of fish biomass at different depths around offshore platforms in the Gulf.
4.1 Fish fauna of Al Khalij
Among the 46 taxa observed around the platforms, Carangidae and Serranidae were the most diverse families. These two families exhibit notable differences in functional traits, behavior, and lifestyle. The pelagic species of Carangidae were predominantly found in the surface levels, where they dominated the assemblages, while the bentho-pelagic Serranidae, particularly species from the Epinephelus genus, were observed near the seafloor, shaping the species composition of the deeper layers. Both families are heavily targeted by fisheries (Allen and Erdmann, 2012; Al-Abdulrazzak et al., 2015), and may therefore benefit from the protection offered by the platforms, which provide both physical shelter and legal fishing restrictions.
Aside from several species within the Carangidae family, some of the most abundant species observed were Pinjalo pinjalo, Lutjanus argentimaculatus (Lutjanidae), and Platax teira (Ephippidae). As a member of the Lutjanidae family, typically composed of bottom-associated species (Smith, 2003), Pinjalo pinjalo was the most abundant species recorded throughout the entire water column around the platforms. Although Pinjalo pinjalo may be less benthic than other Lutjanidae species, its broad depth distribution, from surface to bottom, suggests that the platforms provide suitable habitat for bentho-pelagic species across all layers in an otherwise open-water environment. Similarly, Lutjanus argentimaculatus, though commonly known as the mangrove red snapper, is often found at offshore reefs as fully grown adults (Allen and Erdmann, 2012). In Al Khalij, more than 100 km from the nearest coast, this species was observed forming large schools near the surface. Like species from the Carangidae and Serranidae families, Lutjanus argentimaculatus is a prized food fish and is heavily targeted by fisheries. Platax teira, frequently found around artificial reefs and floating rafts (Heemstra et al., 2022), is omnivorous and was primarily recorded in the surface layers, where it was feeding on leftover food and other organic waste discarded by platform workers. Encrasicholina sp. (Engraulidae) accounted for the highest number of individuals recorded but was observed only at night, likely due to a large school being attracted to the ROV’s light, which reduced the FoV across multiple depth intervals.
Other characteristic species frequently observed around the platforms included Acanthopagrus bifasciatus (Sparidae), Heniochus acuminatus (Chaetodontidae), and Pomacanthus maculosus (Pomacanthidae). All three species are common at both natural and artificial reef sites in the Gulf (Carpenter et al., 1997; Torquato et al., 2017; Sigsgaard et al., 2020). Some observations included species commonly associated with natural coral reef sites, such as Chaetodon nigropunctatus (Chaetodontidae), Chromis spp. (Pomacentridae), Ostracion cubicum, O. cyanurus (Ostraciidae), Pterois miles (Scorpaenidae), Scarus persicus (Scaridae), Siganus sp. (Siganidae), and Thalassoma lunare (Labridae). Although none of these species were abundant, with a total MaxN of less than 7 for all except Chromis spp., their presence indicates a habitat of higher complexity capable of supporting a large biodiversity.
The first record of Cheilinus trilobatus (Labridae) within the Gulf, along with the first records of two other species observed by Torquato et al. (2017), underscores the limited exploration of the Gulf’s platforms as artificial reefs. With two independent studies of two different Qatari oilfields recording three new species in the Gulf, it is reasonable to hypothesize that more extensive studies of these artificial habitats could lead to the discovery of additional previously unrecorded species in the region. These rarely observed species may also indicate that platforms act as stepping stones for the dispersal and connectivity of species in an area where natural reefs are becoming increasingly fragmented (Friedlander et al., 2014; Schutter et al., 2019).
A number of observations could not be identified to the species level, either due to limited visibility in the footage or the possible presence of several closely related species with no distinguishable morphological characteristics visible. In most cases, individuals identified only to the genus level with likely belonged to one of the species already observed within the same genus. For instance, Epinephelus spp. (Epinephelidae) were likely E. areolatus, E. chlorostigma, or E. coioides. In some cases, direct observations made in the area by the authors can help qualify a reasonable assumption of the correct species observed in the video footage. Carcharhinus melanopterus and C. limbatus (Carcharhinidae) have frequently been observed near the surface at the platforms, making these the likely species for the unidentified Carcharhinus spp. The same applies to Lethrinus nebulosus (Lethrinidae), which was caught by anglers under a rare, research-permitted allowance to fish from the platforms, making it the most likely species among the identified Lethrinus spp. However, L. borbonicus, L. lentjan, and L. microdon cannot be ruled out. In some instances, species identification can only be inferred based on previously observed species in the area. The abundant Platycaranx spp., P. chrysophrys, P. malabaricus, and P. talamparoides (Carangidae) have all been previously recorded in the region. These three species exhibit very similar morphology, making confident species identification challenging based on video footage alone. The Platycaranx spp. observations may belong to any one or a combination of these species. Encrasicholina sp., which dominated the night transects, could potentially belong to E. devisi, E. gloria, E. heteroloba, or E. punctifer. Scarus persicus was directly observed and identified from the footage, but the Scarus spp. may also include S. ferrugineus, S. fuscopurpureus, S. ghobban, or S. psittacus, all of which are known to occur in the Gulf. The most likely identification of Siganus sp. is either S. sutor or S. luridus. The observed Chromis spp. could potentially belong to either C. flavaxilla or C. xanthopterygia. However, making meaningful inferences about the identity of the observed cryptobenthic species is challenging due to the limited morphological information obtained from the video footage of these small species.
4.2 Fish assemblages associated to depth and structural complexity
A distinct shift in species composition was observed with increasing depth, despite the total number of species remaining relatively consistent across different levels of the water column (Figure 4). Pelagic Carangidae and Lutjanidae species dominated the surface layers, accounting for the highest abundance and biomass, while benthopelagic fishes such as Epinephelus and Pterois miles characterized the seafloor. Other benthopelagic species, such as Pomacanthus maculosus and Acanthopagrus bifasciatus, were observed throughout most of the water column but predominantly in the surface layers. This affirms the platforms’ function as artificial reefs, providing habitats for bottom-associated species in the layers with higher light penetration within a deep-water environment. The Bray-Curtis dissimilarity visualized in the NMDS plot (Figure 5) revealed greater dissimilarity between seafloor samples compared to surface samples, likely due to the stronger site affiliation of benthic species, such as Epinephelus coioides, in contrast to the more mobile pelagic species observed at the surface, such as Scomberoides commersonnianus and Alepes vari (Carangidae).
The overall trend in abundance and biomass showed a decline in both fish counts and biomass estimates with increasing depth. This pattern may be influenced by several factors, including higher primary production in the surface layers due to greater light penetration, which supports a larger abundance of organisms at higher trophic levels. The discharge of organic waste from platform workers may also contribute to the high abundance of fish near the surface. Platax teira, along with other species, were frequently observed by the authors rapidly feeding on food waste discarded from the platforms. Both light penetration and organic discharges during working hours may also explain why this depth-related decline in abundance and biomass was less pronounced at night. However, differences between day and night samples, both in the number of samples and in fish behavior, should be considered when interpreting this trend. Diurnal changes in fish activity levels may influence how easily they are detected on video, which can introduce bias in visual survey data. These limitations underline the importance of using more balanced sampling designs in future studies. Notably, the observed pattern represents only a temporal snapshot, as the survey included data spanning just 24 hours. Abundance and biomass patterns are known to be influenced by both seasonal variations (Martin and Lowe, 2010) and anthropogenic activities (Martin et al., 2012). For instance, increased water temperatures during the summer and platform activities, such as jacket “hydrocleaning” events that remove sessile invertebrates, are expected to impact these patterns. Additional surveys conducted across different seasons are needed to fully understand the temporal changes in fish depth distribution around these platforms. Understanding the relationship between depth, abundance, and biomass is crucial for potential RTR scenarios following platform decommissioning, particularly since some proposals advocate for removing the platform’s top layers (Dauterive, 1999). Similar conclusions have been drawn in the Gulf of Mexico, where standing platforms have been shown to support higher fish biomass and biodiversity by retaining habitat in the upper water column. Partial removal as part of RTR efforts risks the loss of these shallow-water communities, although reefed platforms may still preserve essential ecological functions at deeper strata (Stunz and Coffey, 2020).
Although the overall trend showed a negative correlation between abundance and biomass with increasing depth, the relationship was not strictly linear, as certain depth layers exhibited peaks in fish counts and biomass estimates. At both platforms, the bottom layers showed significantly higher abundance than the levels directly above, likely due to a number of species strictly associated with the seafloor. At DP3, samples taken around 10 m and 35 m also revealed clear peaks in abundance and biomass, likely influenced by the presence of crossbeam structures, which increases habitat complexity. This pattern of heightened diversity and abundance around structurally complex areas of platforms is consistent with findings by Ajemian et al. (2015); Consoli et al. (2018), and Rilov and Benayahu (2002). As with the relationship between depth and abundance, the importance of high-complexity areas on platforms should be considered in future RTR decision-making processes. However, due to differences in the number of daytime transects conducted at each platform, the deviations observed at DP3 cannot be interpreted as general differences between the two platforms.
These biomass patterns are inherently linked to the methods used for biomass estimation. The use of a single camera in the ROV setup limits the accuracy of biomass estimates, whereas stereo camera setups provide more precise size measurements and reduce uncertainty. However, because many of the fish caught at the platforms were near the maximum recorded length for their species, assigning individuals to the maximum size class likely improved biomass estimates, reducing the risk of underestimation compared to using average size values.
4.3 Comparison to Al Shaheen oilfield
Similar to this study, the diversity and abundance of fishes around another Qatari oilfield, Al Shaheen, have been previously documented (Torquato et al., 2017). The study conducted in Al Shaheen utilized a substantially larger dataset compared to the present study of Al Khalij. Torquato et al. (2017) analyzed 20 more hours of footage, collected over a six-year period, whereas the present study spanned only 24 hours. Additionally, nine different platforms were surveyed in Al Shaheen, compared to just two platforms in Al Khalij. As a result of the larger data collection, 8,348 more fish from 37 more taxa, belonging to 12 more families, were recorded in Al Shaheen. Unlike the present study, the ROV survey of Al Shaheen was not conducted with the specific purpose of biodiversity assessment, and as such, the survey was not optimized for species detection and quantification. Differences in the results of ROV surveys from platforms, depending on whether the sampling design was specifically aimed at biodiversity assessment, have been previously described (Love et al., 2020). With a targeted sampling design, 66.8 fish per minute were recorded in Al Khalij, compared to 10.2 fish per minute in Al Shaheen, where the ROV footage was not specifically aimed at biodiversity surveys. Additionally, the high-resolution cameras deployed in the present study allowed for identification of lower taxonomical units. Our observations were identified to species, genus, and family levels at 70%, 28%, and 2%, respectively, compared to Torquato et al. (2017), who reported 60%, 24%, and 16%. Despite the targeted sampling design being more effective overall, a larger number of rare species, such as Stegostoma tigrinum and Taeniurops meyeni, were reported from Al Shaheen, likely due to the larger sample size.
In both oilfields, the Carangidae family was the most diverse. However, substantial differences in species composition within the family were detected between the oilfields. The abundant species in Al Khalij, such as Alepes vari and Platycaranx spp., were absent in Al Shaheen, and only one individual of Scomberoides commersonnianus was recorded in Al Shaheen compared to 356 in Al Khalij. Species common to both oilfields from the diverse Carangidae family were Caranx sexfasciatus, Caranx ignobilis, Flavocaranx bajad, Gnathanodon speciosus, Scomberoides commersonnianus, and Seriola dumerili. Torquato et al. (2017) also reported Labridae (including Scaridae) and Serranidae (referred to as Epinephelidae) as the other most diverse families, similar to the present study. Among the most abundant species in both oilfields were Heniochus acuminatus, Pomacanthus maculosus, and Acanthopagrus bifasciatus. However, differences were noted, with Abudefduf vaigiensis being among the most abundant in Al Shaheen but absent in Al Khalij, where Pinjalo pinjalo was highly abundant, while only three individuals of this species were observed in Al Shaheen. A negative correlation between abundance and depth was also observed at Al Shaheen.
Differences in sampling design may explain some of the variations in the fish assemblages around the platforms of Al Khalij and Al Shaheen, but other factors undoubtedly also play an important role in these dissimilarities. Though both oilfields are located in the central Gulf there are around 100 km between them with Al Shaheen located more northwest. The Al Shaheen survey covered a larger geographic area, with its nine platform sites spread across distances of up to 20 km, compared to the two platforms surveyed at Al Khalij, only 10 km apart. The platform sites at Al Shaheen are much larger, often comprising multiple large, interconnected structures, including wellhead, process, and accommodation platforms, whereas the two platforms at Al Khalij are relatively small and standalone. This configuration at Al Shaheen provides a significantly larger surface area of hard substrate and increased structural complexity. The documented differences in species diversity and abundance, influenced by structural size, complexity, and geographic location, should be taken into account by future RTR decision makers.
4.4 Production versus attraction
Offshore platform artificial reefs have often been central to the ‘attraction vs. production’ debate (Layman and Allgeier, 2020). The question of whether the diversity and abundance associated with artificial reefs result from high local production of biomass (Powers et al., 2003) or the attraction of marine organisms from surrounding natural habitats (Bohnsack and Sutherland, 1985; Bohnsack, 1989) has been widely discussed. Recent studies suggest that established artificial reefs contribute to local production and do not necessarily attract fish from neighboring natural habitats (Logan and Lowe, 2018). Our observations of multiple spawning Carangidae species align with the findings of Madgett et al. (2022) who also documented spawning behavior of Caranx sexfasciatus at offshore platforms in the Gulf of Thailand. These observations confirm that at least some local production is occurring at the platforms. Both Madgett et al. (2022) and the present study observed spawning behavior exclusively in members of the Carangidae family. This may suggest that Carangidae particularly benefit from artificial habitats provided by platforms, potentially explaining the high diversity within the family. However, a simpler explanation could be that spawning behavior in Carangidae is more easily observed due to the distinct color changes in males (Graham and Castellanos, 2005) while the spawning of other taxa may be less conspicuous and, therefore, unrecorded through visual observations. Although spawning behavior was confirmed only for Carangidae species, a large aggregation of Lutjanus argentimaculatus observed near the surface at DP4 may also indicate spawning of this species, as large spawning aggregations are commonly reported for Lutjanus species (Gibson et al., 2004; Russell and McDougall, 2008).
4.5 Limitations of visual observations
ROV-assisted video recording has proven effective for biodiversity studies, not only in this study but also at offshore platforms in other regions (Schutter et al., 2019; Love et al., 2020; Alexander et al., 2022; Birt et al., 2024). However, as the method relies exclusively on visual observations, it is inherently limited by visibility and provides only a temporal snapshot of the surveyed area. Although ROV surveys have been successful in low-visibility environments (López-Garrido et al., 2020), the significant negative linear correlation between depth and FoV suggests that the method may be biased towards surface layers, where visibility is generally better. However, accounting for the effective survey water volume, the seafloor levels accounted for the highest numbers per m³ in both abundance and estimated biomass. Although accounting for FoV might provide a more accurate representation of the relative abundance in the deeper levels where visibility is low, this approach risks highly underestimating the potentially fully saturated samples from the surface layers due to the aggregated distribution of fishes around the platform legs. Linear relationships between FoV and both species count and abundance were identified, yet the slope of the regressions indicates that these relationships hold little practical significance in this study.
Using ROV-assisted video observations, we were able to reach a plateau in the number of species recorded, both in terms of accumulated survey time and the effective volume surveyed. Although this plateau was reached after 25 minutes of analyzed recordings, the observed number of taxa should not be interpreted as the total number present at the platforms. Rather, this species count reflects the number of taxa that could be detected using this method at the specific time of the survey. Beyond visibility limitations, diurnal variation may further affect species detectability. The behavior and activity levels of fish can vary significantly between day and night, influencing the likelihood of detection via visual methods. For example, some species may be more active and visible during specific times of day, while others may be more cryptic or rest in sheltered areas, escaping detection. This temporal variation, coupled with the imbalance in day and night transects between platforms, introduces additional uncertainty in comparing observations across sites. The ROV itself may also cause some disturbance. While the deployment of an ROV likely causes less disturbance than direct visual observations using SCUBA divers, it was evident from the recordings that at least Encrasicholina sp. was attracted to the ROV’s lights during night surveys. Conversely, it is plausible that other species may be repelled by the ROV’s rotors and movement.
The addition of complementary observation methods would likely increase the number of species detected (Pearman et al., 2016; Alexander et al., 2022). Thus, incorporating eDNA metabarcoding is recommended for monitoring marine diversity associated with platforms in the Gulf. While being even less invasive than ROV surveys, eDNA observations could help detect smaller species not visible through traditional visual methods and offer more accurate species identification with higher taxonomic resolution, relying on genetic data rather than solely on visible morphological traits.
5 Conclusion
This study contributes to the growing body of research supporting the RTR strategy by providing specific insights into the ecological roles of offshore platforms within the unique environmental conditions of the Arabian Gulf. For the first time, we have documented the fish fauna associated with the offshore platforms in Qatar’s Al Khalij oilfield. 4,474 individual fishes were counted, with an estimated biomass of 4,135 kg per platform, and 46 taxa were identified, highlighting the substantial ecological role of these artificial structures in supporting marine biodiversity. Additionally, numerous instances of spawning behavior were observed, underscoring the platforms’ role as habitats for reproductive activity.
Furthermore, our hypothesis that the designated sampling design would enhance the effectiveness of species identification and data collection was validated. The use of high-resolution cameras and a targeted sampling approach facilitated a more accurate quantification of fish abundance, biomass, and diversity compared to previous surveys.
These findings emphasize the importance of offshore platforms as refuges for marine life, particularly in the context of declining natural reef habitats in the Arabian Gulf. By documenting the fish assemblages in the Al Khalij oilfield, this study provides critical insights into the ecological functions of offshore platforms. The observed patterns of species distribution, abundance, and biomass, alongside the detection of spawning behavior, provide valuable insights for future RTR decision-making and reflect the platforms’ capacity to function as effective artificial reefs, contributing significantly to regional marine ecosystem sustainability.
Statements
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
Ethical approval was not required for the study involving animals in accordance with the local legislation and institutional requirements because the data collection was purely observational.
Author contributions
JS: Data curation, Visualization, Writing – original draft, Writing – review & editing, Formal analysis, Investigation, Methodology. FT: Methodology, Writing – review & editing. LF: Validation, Writing – review & editing. CL: Validation, Writing – review & editing, Funding acquisition, Project administration. MC: Project administration, Resources, Writing – review & editing. AG: Project administration, Resources, Writing – review & editing. FA-J: Funding acquisition, Writing – review & editing. RB-H: Conceptualization, Funding acquisition, Methodology, Supervision, Writing – review & editing, Project administration. PM: Conceptualization, Supervision, Validation, Writing – review & editing, Funding acquisition, Project administration. PR: Conceptualization, Funding acquisition, Methodology, Supervision, Writing – review & editing, Project administration.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This project is funded by a NPRP-Standard (NPRP-S) Thirteenth (13th) Cycle grant # NPRP13S-0208-200302, from the Qatar National Research Fund (a member of Qatar Foundation), with co-funding from TotalEnergies. Open Access funding provided by the Qatar National Library.
Acknowledgments
We thank Anne Basseres and Marc Ecoffet of TotalEnergies for their invaluable support with the logistics and planning of the study, and Kevin Cailleaud of TotalEnergies for his helpful scientific review of the manuscript. We are also grateful to the three reviewers for their constructive feedback, which greatly improved the quality of the manuscript.
Conflict of interest
Authors MC and AG were employed by TotalEnergies EP Qatar.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors declare that this study received funding from TotalEnergies EP Qatar. The funder had the following involvement in the study: study design, data collection, decision to publish, and preparation of the manuscript.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Author disclaimer
The findings herein reflect the work, and are solely the responsibility of the authors.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2025.1589927/full#supplementary-material
References
1
Aeby G. S. Ben-Hamadou R. Burt J. A. Padierna M. L. Range P. A. G. Riegl B. et al . (2024). “Coral reefs in the world’s warmest sea: The paradoxical vulnerability of the world’s most robust corals,” in Coral Reefs and Associated Marine Fauna around the Arabian Peninsula (London, England: CRC Press).
2
Ajemian M. J. Wetz J. J. Shipley-Lozano B. Shively J. D. Stunz G. W. (2015). An analysis of artificial reef fish community structure along the Northwestern Gulf of Mexico shelf: potential impacts of “Rigs-to-reefs” Programs. PloS One10, e0126354. doi: 10.1371/journal.pone.0126354 , PMID:
3
Al-Abdulrazzak D. Zeller D. Belhabib D. Tesfamichael D. Pauly D. (2015). Total marine fisheries catches in the Persian/Arabian Gulf from 1950 to 2010. Regional Stud. Mar. Sci.2, 28–34. doi: 10.1016/j.rsma.2015.08.003
4
Al-Ansari E. M. A. S. Husrevoglu Y. S. Yigiterhan O. Youssef N. Al-Maslamani I. A. Abdel-Moati M. A. et al . (2022). Seasonal variability of hydrography off the east coast of Qatar, central Arabian Gulf. Arab J. Geosci15, 1659. doi: 10.1007/s12517-022-10927-4
5
Alexander J. B. Marnane M. J. Elsdon T. S. Bunce M. Songploy S. Sitaworawet P. et al . (2022). Complementary molecular and visual sampling of fish on oil and gas platforms provides superior biodiversity characterisation. Mar. Environ. Res.179, 105692. doi: 10.1016/j.marenvres.2022.105692 , PMID:
6
Allen G. R. Erdmann M. (2012). Reef Fishes of the East Indies: Andaman Sea, Myanmar, Thailand, Indonesia, Christmas Island, Singapore, Malaysia, Brunei, Philippines, Papua New Guinea, Solomon Islands (Arlington, Virginia, USA: Tropical Reef Research).
7
Bach S. S. Robinson D. P. Abdulrahman A. A. Al-Jaidah M. (2016). “Testing the affinity of whale sharks, Rhincodon typus, to offshore oil platforms in a newly discovered aggregation site off the cost of Qatar in the Arabian Gulf,” in The 4th International Whale Shark Conference. Hamad bin Khalifa University Press (HBKU Press) doi: 10.5339/qproc.2016.iwsc4.6
8
Bartholomew A. Feary D. (2012). Man-made structures as artificial reefs in the gulf. (Dordrecht, the Netherlands: Springer) 171–186. doi: 10.1007/978-94-007-3008-3_10
9
Birt M. McLean D. L. Case M. Jaworski S. Speed C. W. Pygas D. et al . (2024). Contribution of offshore platforms and surrounding habitats to fish production in the Bass Strait, south-east Australia. Continental Shelf Res.274, 105209. doi: 10.1016/j.csr.2024.105209
10
Bohnsack J. A. (1989). Are high densities of fishes at artificial reefs the result of habitat limitation or behavioral preference? Bull. Mar. Sci.44, 631–645.
11
Bohnsack J. A. Sutherland D. L. (1985). Artificial reef research: A review with recommendations for future priorities. Bull. Mar. Sci.37, 11–39.
12
Burt J. A. Bartholomew A. (2019). Towards more sustainable coastal development in the Arabian Gulf: Opportunities for ecological engineering in an urbanized seascape. Mar. pollut. Bull.142, 93–102. doi: 10.1016/j.marpolbul.2019.03.024 , PMID:
13
Burt J. Bartholomew A. Usseglio P. Bauman A. Sale P. (2009). Are artificial reefs surrogates of natural habitats for corals and fish in Dubai, United Arab Emirates? Coral Reefs28, 663–675. doi: 10.1007/s00338-009-0500-1
14
Burt J. Ben-Hamadou R. Abdel-Moati M. Fanning L. Kaitibie S. Al-Jamali F. et al . (2017). Improving management of future coastal development in Qatar through ecosystem-based management approaches. Ocean Coast. Manage.148, 171–181. doi: 10.1016/j.ocecoaman.2017.08.006
15
Carpenter K. Krupp F. Jones D. Zajonz U. (1997). Living marine resources of Kuwait, eastern Saudi Arabia, Bahrain, Qatar, and the United Arab Emirates. Available online at: https://www.semanticscholar.org/paper/Living-marine-resources-of-Kuwait%2C-eastern-Saudi-Carpenter-Krupp/d363526e259e5b6eb522c29eaffb71045d340776 (Accessed February 8, 2023).
16
Claisse J. T. Pondella D. J. Love M. Zahn L. A. Williams C. M. Williams J. P. et al . (2014). Oil platforms off California are among the most productive marine fish habitats globally. Proc. Natl. Acad. Sci.111, 15462–15467. doi: 10.1073/pnas.1411477111 , PMID:
17
Coles S. L. (2003). Coral species diversity and environmental factors in the Arabian Gulf and the Gulf of Oman: a comparison to the Indo-Pacific Region. Atoll Res. Bull.507, 1–19. doi: 10.5479/si.00775630.507.1
18
Consoli P. Mangano M. C. Sarà G. Romeo T. Andaloro F. (2018). The influence of habitat complexity on fish assemblages associated with extractive platforms in the central Mediterranean Sea. Adv. Oceanography Limnology9. doi: 10.4081/aiol.2018.7918
19
Consoli P. Romeo T. Ferraro M. Sarà G. Andaloro F. (2013). Factors affecting fish assemblages associated with gas platforms in the Mediterranean Sea. J. Sea Res.77, 45–52. doi: 10.1016/j.seares.2012.10.001
20
Cordes E. E. Jones D. O. B. Schlacher T. A. Amon D. J. Bernardino A. F. Brooke S. et al . (2016). Environmental impacts of the deep-water oil and gas industry: A review to guide management strategies. Front. Environ. Sci.4. doi: 10.3389/fenvs.2016.00058
21
Dauterive L. (1999). Rigs-to-Reefs Policy, Progress, and Perspective (Austin, Texas, USA: OnePetro). doi: 10.2118/52709-MS
22
Ellis D. M. DeMartini E. E. (1995). Evaluation of a video camera technique for indexing abundances of juvenile pink snapper, Pristipomoides filamentosus, and other Hawaiian insular shelf fishes. Fishery Bull.93, 67–77.
23
Feary D. A. Burt J. A. Bartholomew A. (2011). Artificial marine habitats in the Arabian Gulf: Review of current use, benefits and management implications. Ocean Coast. Manage.54, 742–749. doi: 10.1016/j.ocecoaman.2011.07.008
24
Fowler A. M. Booth D. J. (2012). Evidence of sustained populations of a small reef fish on artificial structures. Does depth affect production on artificial reefs? J. Fish Biol.80, 613–629. doi: 10.1111/j.1095-8649.2011.03201.x , PMID:
25
Friedlander A. M. Ballesteros E. Fay M. Sala E. (2014). Marine communities on oil platforms in Gabon, west africa: high biodiversity oases in a low biodiversity environment. PloS One9, e103709. doi: 10.1371/journal.pone.0103709 , PMID:
26
Fujii T. (2016). Potential influence of offshore oil and gas platforms on the feeding ecology of fish assemblages in the North Sea. Mar. Ecol. Prog. Ser.542, 167–186. doi: 10.3354/meps11534
27
Gibson R. N. Atkinson R. J. A. Gordon J. D. M. (2004). Oceanography and Marine Biology: An annual review Vol. 42 (Boca Raton, Florida, USA: CRC Press).
28
Graham R. Castellanos D. (2005). Courtship and spawning behaviors of carangid species in Belize. Fishery Bulletin- Natl. Oceanic Atmospheric Administration103, 426–432.
29
Heemstra P. C. Heemstra E. Ebert D. A. Holleman W. Randall J. E. (2022). Coastal Fishes of the Western Indian Ocean (Makhanda, South Africa: South African Institute for Aquatic Biodiversity).
30
Hereher M. E. (2020). Assessment of climate change impacts on sea surface temperatures and sea level rise—The arabian gulf. Climate8, 50. doi: 10.3390/cli8040050
31
Karwowski H. Heitmann H. Ibanez-Erquiaga B. Coolen J. W. P. Svendsen J. C. (2024). Oil and gas platforms in the North Sea: Do they benefit commercial fish species and biodiversity? Habitat28, 70–77.
32
Kashubsky M. Morrison A. (2013). Security of offshore oil and gas facilities: exclusion zones and ships’ routeing. Aust. J. Maritime Ocean Affairs5, 1–10. doi: 10.1080/18366503.2013.10815725
33
Layman C. A. Allgeier J. E. (2020). An ecosystem ecology perspective on artificial reef production. J. Appl. Ecol.57, 2139–2148. doi: 10.1111/1365-2664.13748
34
Logan R. K. Lowe C. G. (2018). Residency and inter-reef connectivity of three gamefishes between natural reefs and a large mitigation artificial reef. Mar. Ecol. Prog. Ser.593, 111–126. doi: 10.3354/meps12527
35
López-Garrido P. H. Barry J. P. González-Gordillo J. I. Escobar-Briones E. (2020). ROV’s Video Recordings as a Tool to Estimate Variation in Megabenthic Epifauna Diversity and Community Composition in the Guaymas Basin. Front. Mar. Sci.7. doi: 10.3389/fmars.2020.00154
36
Love M. S. Nishimoto M. M. Clark S. Kui L. Aziz A. Palandro D. (2020). A comparison of two remotely operated vehicle (ROV) survey methods used to estimate fish assemblages and densities around a California oil platform. PloS One15, e0242017. doi: 10.1371/journal.pone.0242017 , PMID:
37
Love M. S. Nishimoto M. M. Snook L. Kui L. (2019). An analysis of the sessile, structure-forming invertebrates living on California oil and gas platforms. Bull. Mar. Sci.95, 583–596. doi: 10.5343/bms.2017.1042
38
Love M. S. Schroeder D. M. Nishimoto M. M. (2003). The Ecological Role of Oil and Gas Production Platforms and Natural Outcrops on Fishes in Southern and Central California: A Synthesis of Information. Available online at: https://www.govinfo.gov/app/details/GOVPUB-I-51cf6a08a3bcabd863d066fb8f807ebc (Accessed December 6, 2024).
39
Madgett A. S. Harvey E. S. Driessen D. Schramm K. D. Fullwood L. A. F. Songploy S. et al . (2022). Spawning aggregation of bigeye trevally, Caranx sexfasciatus, highlights the ecological importance of oil and gas platforms. Estuarine Coast. Shelf Sci.276, 108024. doi: 10.1016/j.ecss.2022.108024
40
Martin C. J. B. Allen B. J. Lowe C. G. (2012). Environmental impact assessment: detecting changes in fish community structure in response to disturbance with an asymmetric multivariate BACI sampling design. soca111, 119–131. doi: 10.3160/0038-3872-111.2.119
41
Martin C. J. B. Lowe C. G. (2010). Assemblage structure of fish at offshore petroleum platforms on the san pedro shelf of southern california. Mar. Coast. Fisheries2, 180–194. doi: 10.1577/C09-037.1
42
Pearman J. K. Anlauf H. Irigoien X. Carvalho S. (2016). Please mind the gap – Visual census and cryptic biodiversity assessment at central Red Sea coral reefs. Mar. Environ. Res.118, 20–30. doi: 10.1016/j.marenvres.2016.04.011 , PMID:
43
Peterson J. E. (2009). Life after oil: economic alternatives for the arab gulf states. Mediterr. Q.20, 1–18. doi: 10.1215/10474552-2009-011
44
Powers S. P. Grabowski J. H. Peterson C. H. Lindberg W. J. (2003). Estimating enhancement of fish production by offshore artificial reefs: uncertainty exhibited by divergent scenarios. Mar. Ecol. Prog. Ser.264, 265–277. doi: 10.3354/meps264265
45
Price A. R. G. (2002). Simultaneous “hotspots“ and “coldspots” of marine biodiversity and implications for global conservation. Mar. Ecol. Prog. Ser.241, 23–27. doi: 10.3354/meps241023
46
Price A. Sheppard C. Roberts C. (1993). The Gulf: Its biological setting. Mar. pollut. Bull.27, 9–15. doi: 10.1016/0025-326X(93)90004-4
47
QatarEnergy (2020). QatarEnergy Annual Review (not applicable: TotalEnergies.com). Available online at: https://totalenergies.com/media/news/press-releases/Qatar-petroleum-et-total-signent-un-nouvel-accord-prolongeant-de-25-ans-leur-partenariat-sur-le.
48
Rabaoui L. Lin Y.-J. Qurban M. A. Maneja R. H. Franco J. Joydas T. V. et al . (2015). Patchwork of oil and gas facilities in Saudi waters of the Arabian Gulf has the potential to enhance local fisheries production. ICES J. Mar. Sci.72, 2398–2408. doi: 10.1093/icesjms/fsv072
49
Range P. Riera R. Omerspahic M. Bouwmeester J. Bach S. S. Møller P. R. et al . (2018). Fouling assemblages on offshore oil platforms: diverse vertical reefs in the Arabian Gulf (Doha, Qatar: Hamad bin Khalifa University Press). doi: 10.5339/qfarc.2018.EEPP894
50
Rezai H. Wilson S. Claereboudt M. Riegl B. (2004). 5. Coral reef status in the ropme sea area: Arabian/Persian gulf, gulf of Oman and Arabian sea. Status Coral Reefs World1, 155–170.
51
Riegl B. Purkis S. (2012). “Coral Reefs of the Gulf: Adaptation to Climatic Extremes in the World’s Hottest Sea,” in Marine & Environmental Sciences Faculty Books and Book Chapters. (Dordrecht, Netherlands: Springer Science+Business Media) Available online at: https://nsuworks.nova.edu/occ_facbooks/26.
52
Riera R. Torquato F. Range P. Ben-Hamadou R. Møller P. R. Tuset V. M. (2023). Are offshore platforms a good candidate to restore functional diversity of reef fish communities in the Arabian Gulf? Regional Stud. Mar. Sci.66, 103171. doi: 10.1016/j.rsma.2023.103171
53
Rilov G. Benayahu Y. (2000). Fish assemblage on natural versus vertical artificial reefs: the rehabilitation perspective. Mar. Biol.136, 931–942. doi: 10.1007/s002279900250
54
Rilov G. Benayahu Y. (2002). Rehabilitation of coral reef-fish communities: The importance of artificial-reef relief to recruitment rates. Bull. Mar. Sci.70, 185–197.
55
Robinson D. P. Jaidah M. Y. Jabado R. W. Lee-Brooks K. El-Din N. M. N. Malki A. A. A. et al . (2013). Whale Sharks, Rhincodon typus, Aggregate around Offshore Platforms in Qatari Waters of the Arabian Gulf to Feed on Fish Spawn. PloS One8, e58255. doi: 10.1371/journal.pone.0058255 , PMID:
56
Russell D. J. McDougall A. J. (2008). Reproductive biology of mangrove jack (Lutjanus argentimacuiatus) in northeastern Queensland, Australia. New Z. J. Mar. Freshw. Res.42, 219–232. doi: 10.1080/00288330809509950
57
Schroeder D. M. Love M. S. (2004). Ecological and political issues surrounding decommissioning of offshore oil facilities in the Southern California Bight. Ocean Coast. Manage.47, 21–48. doi: 10.1016/j.ocecoaman.2004.03.002
58
Schulze A. Erdner D. L. Grimes C. J. Holstein D. M. Miglietta M. P. (2020). Artificial reefs in the northern gulf of Mexico: community ecology amid the “Ocean sprawl. Front. Mar. Sci.7. doi: 10.3389/fmars.2020.00447
59
Schutter M. Dorenbosch M. Driessen F. M. F. Lengkeek W. Bos O. G. Coolen J. W. P. (2019). Oil and gas platforms as artificial substrates for epibenthic North Sea fauna: Effects of location and depth. J. Sea Res.153, 101782. doi: 10.1016/j.seares.2019.101782
60
Sheppard C. Al-Husiani M. Al-Jamali F. Al-Yamani F. Baldwin R. Bishop J. et al . (2010). The Gulf: A young sea in decline. Mar. pollut. Bull.60, 13–38. doi: 10.1016/j.marpolbul.2009.10.017 , PMID:
61
Sheppard C. Price A. Roberts C. (1992). Marine ecology of the Arabian region: patterns and processes in extreme tropical environments (London: Academic Press). Available online at: http://www.gbv.de/dms/bowker/toc/9780126394900.pdf.
62
Sigsgaard E. E. Torquato F. Frøslev T. G. Moore A. B. M. Sørensen J. M. Range P. et al . (2020). Using vertebrate environmental DNA from seawater in biomonitoring of marine habitats. Conserv. Biol.34, 697–710. doi: 10.1111/cobi.13437 , PMID:
63
Smith J. L. B. (2003). Smiths’ Sea Fishes (Johannesburg, South Africa: Penguin Random House South Africa).
64
Stachowitsch M. Kikinger R. Herler J. Zolda P. Geutebrück E. (2002). Offshore oil platforms and fouling communities in the southern Arabian Gulf (Abu Dhabi). Mar. pollut. Bull.44, 853–860. doi: 10.1016/S0025-326X(02)00085-1 , PMID:
65
Stunz G. W. Coffey D. M. (2020). A Review of the Ecological Performance and Habitat Value of Standing versus Reefed Oil and Gas Platform Habitats in the Gulf of Mexico. Available online at: https://hdl.handle.net/1969.6/89073 (Accessed June 17, 2025).
66
Torquato F. Jensen H. M. Range P. Bach S. S. Ben-Hamadou R. Sigsgaard E. E. et al . (2017). Vertical zonation and functional diversity of fish assemblages revealed by ROV videos at oil platforms in The Gulf. J. Fish Biol.91, 947–967. doi: 10.1111/jfb.13394 , PMID:
67
Torquato F. Omerspahic M. H. Range P. Bach S. S. Riera R. Ben-Hamadou R. (2021). Epibenthic communities from offshore platforms in the Arabian Gulf are structured by platform age and depth. Mar. pollut. Bull.173, 112935. doi: 10.1016/j.marpolbul.2021.112935 , PMID:
68
van Elden S. Meeuwig J. J. Hobbs R. J. (2022). Offshore platforms as novel ecosystems: A case study from Australia’s Northwest Shelf. Ecol. Evol.12, e8496. doi: 10.1002/ece3.8496 , PMID:
69
Vidal P. d. C.J. González M. O. A. Vasconcelos R. Melo D. Ferreira P. Sampaio P. G. V. et al . (2022). Decommissioning of offshore oil and gas platforms: A systematic literature review of factors involved in the process. Ocean Eng.255, 111428. doi: 10.1016/j.oceaneng.2022.111428
Summary
Keywords
artificial reefs, fish, biodiversity, species richness, biomass, offshore platforms, Arabian Gulf, rigs-to-reefs
Citation
Sørensen JM, Torquato F, Faulkner L, Lowe CG, Carrara M, Godina A, Al-Jamali F, Ben-Hamadou R, Møller PR and Range P (2025) Species diversity, abundance, and biomass of fish at offshore platforms in the Arabian Gulf. Front. Mar. Sci. 12:1589927. doi: 10.3389/fmars.2025.1589927
Received
08 March 2025
Accepted
07 July 2025
Published
02 September 2025
Volume
12 - 2025
Edited by
Nadav Shashar, Ben-Gurion University of the Negev, Israel
Reviewed by
Frederico Pereira Brandini, University of São Paulo, Brazil
Amit Lerner, Hebrew University of Jerusalem, Israel
Updates
Copyright
© 2025 Sørensen, Torquato, Faulkner, Lowe, Carrara, Godina, Al-Jamali, Ben-Hamadou, Møller and Range.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Johan Mølgård Sørensen, johan.sorensen@snm.ku.dk; Pedro Range, prange@qu.edu.qa
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.