 Federica Cacciatore1*
Federica Cacciatore1* Michele Cornello1
Michele Cornello1 Valentina Bernarello1Federica Oselladore1Marta Novello2Alessandra Girolimetto2Massimo Zorzi2Daniele Bon2Fabio Strazzabosco2
Valentina Bernarello1Federica Oselladore1Marta Novello2Alessandra Girolimetto2Massimo Zorzi2Daniele Bon2Fabio Strazzabosco2 Andrea Bonometto1
Andrea Bonometto1 Rossella Boscolo Brusà1
Rossella Boscolo Brusà1- 1Italian Institute for Environmental Protection and Research (ISPRA), Chioggia-Venice, Italy
- 2Environmental Prevention and Protection Agency of Veneto Region (ARPAV), Padova, Italy
Macrozoobenthos are invertebrates playing a fundamental role in the ecosystem processes of several environments, such as lagoons. They contribute to nutrient cycles, metabolization of pollutants, sediment oxygenation, as well as filtering phytoplankton, and are a food source for other organisms at higher level of the food web. The distribution of macrozoobenthos depends on both abiotic and biotic factors, such as salinity, depth, hydrodynamics, sediment size and composition, as well as inter- and intraspecific competition and predation. Macrozoobenthos communities are commonly used as bioindicators to assess the quality of coastal and transitional marine ecosystems. In fact, they represent one of the biological quality elements for evaluating the ecological quality status of transitional and coastal waters under the Water Framework Directive. The aim of this study is to investigate changes in the macrozoobenthos community and their relationship with environmental parameters in the Venice Lagoon, one of the largest and most important transitional water bodies in the Mediterranean Sea. To this end, ecological indices were applied to 268 samples and 252 taxa, collected from 2011 to 2022, with macrozoobenthos sampled once every three years during the spring-summer period. Trends in environmental conditions of the water coloumn (sampled seasonally each year) and sediments (sampled concurrently with macrozoobenthos) were also analyzed, highlighting a significant increase/decrease in some parameters, which were then related to dissimilarities in macrozoobenthos abundances. The results show that the macrozoobenthos community in the Venice lagoon has changed over time, with evidence of biodiversity loss and differences between polyhaline and euhaline areas, and between different hydrodynamic zones. The changes in macrozoobenthos are also significantly related to environmental parameters such as salinity, temperature, nutrients, organic carbon, silicate, and grain size.
Introduction
Soft bottom macrozoobenthos consists of benthic invertebrates generally defined as organisms retained by a 0.5 mm sieve size. Benthic assemblages play a fundamental role in the ecosystem processes of several environments, such as lagoons. Indeed, they contribute to nutrient cycles, metabolism of pollutants, sediment oxygenation, as well as the filtration of phytoplankton. Moreover, they are a food source for other organisms at higher level of the food web (Pearson and Rosenberg, 1978; Dauer et al., 2000).
Benthic assemblages are predominantly sedentary infaunal species, reflecting and responding to the environmental conditions in which they live (López-López and Sedeño-Díaz, 2015; Borja et al., 2000). The distribution of macrozoobenthos depends on both abiotic factors, such as salinity, depth, hydrodynamics, sediment size and composition, and biotic factors, such as inter- and intraspecific competition and predation (Pearson and Rosenberg, 1978; Dauer et al., 2000; Magni et al., 2009). Most species have long life spans and belong to a wide range of taxonomic, functional and trophic groups. Therefore, they can integrate disturbance events over time but also are sensitive to several different sources of disturbance, including groups with different tolerance levels (Pearson and Rosenberg, 1978; Dauer et al., 2000; Ponti et al., 2009). Therefore, macrozoobenthos is often used to predict imbalance in physical, chemical and biological conditions as well as the impact of anthropogenic activities and it is widely used worldwide as a bioindicator to assess the quality of marine, coastal and transitional ecosystems (see e.g.: Mistri et al., 2000; Mobashi et al., 2016; Li et al., 2017; Bangun et al., 2018; D’Alessandro et al., 2018; Dapeng et al., 2019). Indeed, benthic macroinvertebrates are among the biological quality elements to assess ecological quality in transitional and coastal waters according to the Water Framework Directive, in terms of their diversity, abundance and the presence of disturbance-sensitive taxa (European Parliament and Council, 2000).
Transitional aquatic environments are coastal areas where continental freshwater meets marine saltwater, characterized by a high variability in the chemical-physical parameters of the water, including temperature, salinity, and oxygen (Tagliapietra et al., 2009). As they are shallow surface waters, temperature and salinity are more variable compared to adjacent marine or freshwater systems. Additionally, they are zones of high biological production, as the nutrient enrichment that distinguishes them makes them optimal areas for reproduction, nesting, and wintering of various bird and fish species, some of which are of conservation and European community interest (Whitefield and Eliott, 2005). Sometimes, however, specific conditions of nutrient enrichment in the water column, as well as the reduction of water exchange with the open sea and the sedimentary nature of the seabed, can lead to common hypoxic and/or anoxic conditions (Cognetti et al., 2002; Knutsen et al., 2024).
All transitional environments are subject to natural formation and disappearance over the course of several millennia (Cognetti et al., 2002; Narayana and Priju, 2006). The formation and subsequent evolution of coastal lagoons and estuaries are due to the interaction of multiple factors, including fluvial sediment deposits, marine currents, wave dynamics, wind forces, coastal soil subsidence, and sea level fluctuation (Narayana and Priju, 2006; Barausse et al., 2015; Molinaroli et al., 2009; Pérez-Ruzafa et al., 2011; Alademomi et al., 2020). However, various anthropogenic factors can accelerate the natural processes of morphological variation. These include seabed erosion caused by fishing gear, the creation of artificial canals or sandbars, wave action from boats, subsidence induced by extractive activities at sea, and abrupt sea level changes due to climate change (Molinaroli et al., 2009: Barausse et al., 2015). Nutrient enrichment, pollution, increased turbidity, fishing pressure, seabed erosion, and, in general, all sudden environmental parameter changes, caused by anthropogenic factors, impact biological communities, which may not have sufficient time to adapt and respond promptly to the environmental pressures they encounter, resulting in significant ecosystem-level consequences (Molinaroli et al., 2009; Newton et al., 2014; Barausse et al., 2015; Facca et al., 2020; European Environment Agency, 2021).
Since 2011, a monitoring program in accordance with the WFD has been active in the Venice Lagoon. This program includes macrozoobenthos and sediment surveys conducted at 77 stations once every three years, and water parameter measurements taken seasonally at 30 sites each year across both natural and heavily modified water bodies. After more than a decade, the accumulated dataset has enabled the investigation of spatial and temporal patterns and dynamics in the lagoon.
This study aims to examine changes in the macrozoobenthos community in the Venice Lagoon from 2011 to 2022 and to assess the relationship between biodiversity dynamics and environmental parameters within the context of climate change. While several studies have explored benthic responses to environmental and climatic pressures in other coastal lagoons (e.g., Anthony et al., 2009; Alademomi et al., 2020; Boutoumit et al., 2021), and others have addressed specific ecological aspects of macrozoobenthos in the Venice Lagoon (e.g., Sfriso et al., 2001; Oselladore et al., 2022), or the broader impacts of climate change on the lagoon’s biodiversity (Lacoste et al., 2023), to the best of our knowledge, this study represents the first multi-station, long-term investigation specifically focused on macrozoobenthic biodiversity in the Venice Lagoon in relation to climate-driven environmental changes. By integrating biological and physico-chemical data over a 12-year period, this study provides a novel contribution to understanding the ecological dynamics of transitional ecosystems under increasing climate pressure.
Materials and methods
Study area
The Venice lagoon (Figure 1) is a delicate and fragile transitional environment in the northwestern part of the Adriatic Sea (Italy). It is one of the largest (about 550 km²) and most important transitional waters in the Mediterranean Sea. It is a shallow body of water of approximately 1.3 m in depth, separated from the sea by 3 wide inlets and with 14 freshwater inputs. It has been continually threatened by human activities, particularly over the last century, with significant man-made transformations (Sorokin et al., 2002; Guerzoni and Tagliapietra, 2006; Solidoro et al., 2010; Feola et al., 2024).
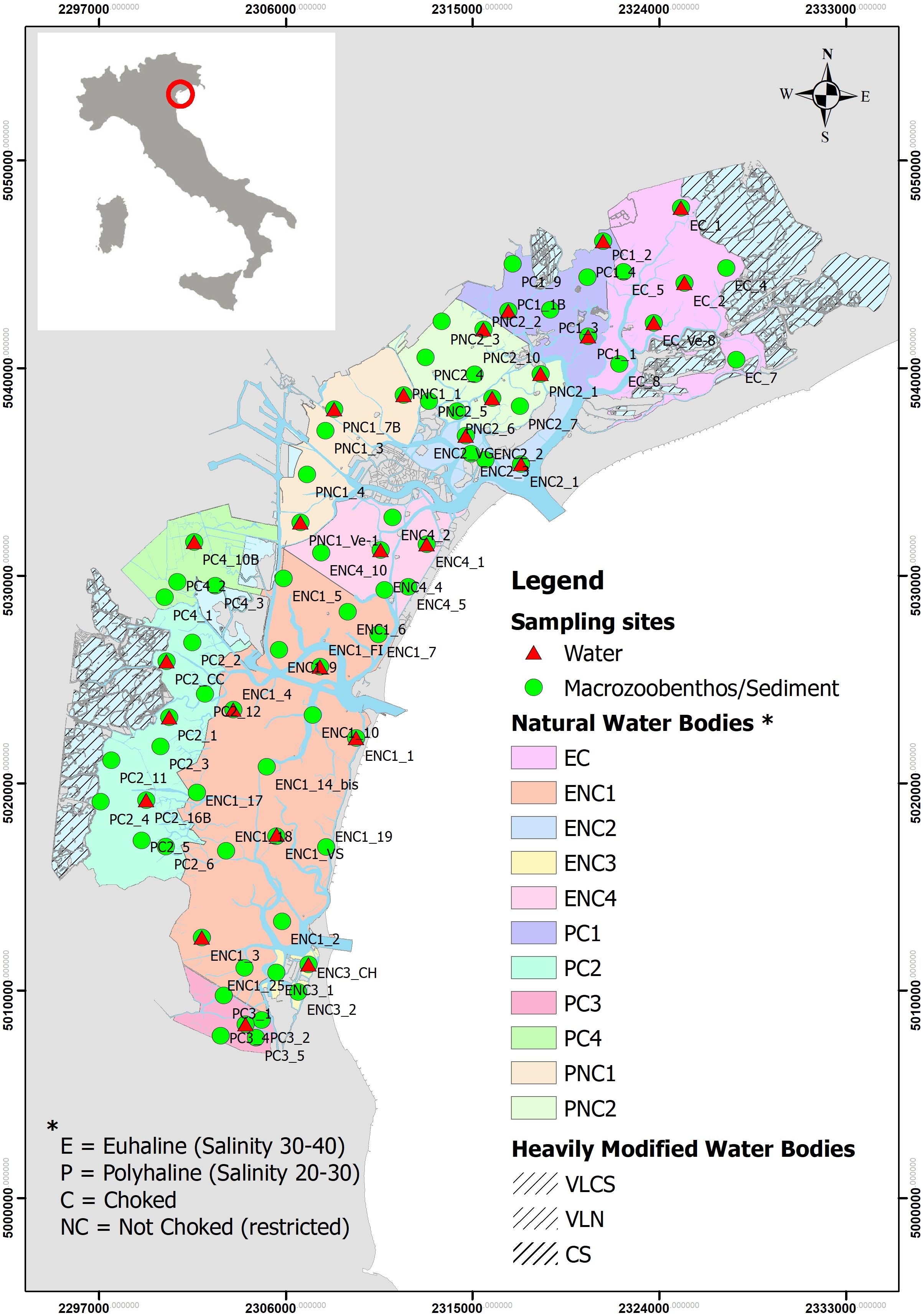
Figure 1. Map of the Venice Lagoon (Italy) with stations of macrozoobenthos, sediments and waters. Water body identification is also given.
According to the WFD, the Venice Lagoon is a large coastal microtidal lagoon, divided into 11 natural water bodies (WBs), belonging to 4 different types according to salinity (P =Polyaline with salinity range 20 - 30; E = Euhaline with salinity range 30-40) and by confinement degree (C= Choked; NC= Non-Choked) and 3 heavily modified WBs. For the present study, only natural WBs were considered.
Macrozoobenthos sampling and laboratory processing
Field surveys were conducted by boat in the spring (May/June) 2012, 2014, 2019 and 2022 in the Venice Lagoon at 75 stations (Figure 1). Sediment samples were collected using an Ekman-Birge grab with a surface area of 15 x 15 cm (225 cm²), capable of penetrating the sediment vertically up to approximately 20 cm. At each station, three separate sediment samples were collected through independent deployments of the grab, resulting in a total sampling surface area of 0.0675 m². The sampling procedure adopted in this study strictly follows the official protocols established by ISPRA (2010), developed within the framework of the European Water Framework Directive (European Parliament and Council, 2000). These protocols are uniformly applied by all Regional Environmental Agencies in Italy to ensure consistency and comparability of benthic data across transitional water bodies.
The three sediment aliquots were separately sieved with a 1 mm mesh sieve. Although macrofauna is generally defined as organisms retained by a 0.5 mm sieve, a 1 mm mesh was used as it offers a practical balance between data representativeness and processing effort, without significantly affecting community parameters (APAT-SIBM-ICRAM, 2003).
The material retained by the sieve was transferred to an anesthetic solution to prevent the organisms from contracting, which would make the identification process difficult. The samples were then fixed in 70% ethyl alcohol for later sorting and identification of the fauna in the laboratories. Although the samples were processed individually, the results were later combined during the data analysis phase.
Macroinvertebrates were initially divided into major taxonomic groups (mollusks, crustaceans, polychaetes, etc.) and then subjected to refined classification. The data were ordered and conformed according to the taxonomic nomenclature considered in the Checklist of Italian Marine Fauna, published by the Italian Society of Marine Biology (Relini, 2008, 2010). In the case of species not included in the Italian checklist due to changes in taxonomic nomenclature, reference was made to the World Register of Marine Species (WORMS, 2025; http://www.marinespecies.org/).
Once the complete list of macrozoobenthos species was obtained, further comparison was made with the check list of macrozoobenthos species for the Adriatic and Mediterranean Seas edited and periodically updated by the Italian Society of Marine Biology (SIBM) to identify species not yet recorded.
All datasets are available at ARPAV Website as reported on Data Availability Section.
Environmental parameters
Water was sampled seasonally at 28 stations (Figure 1) each year from 2011 to 2022. Sediment samples were collected at the same time and stations as in the macrozoobenthos surveys (Figure 1).
All datasets are available on the ARPAV Website as reported in the Data Availability Section.
Data processing and statistical analyses
In the first monitoring cycle, 85 stations were sampled for macrozoobenthos assessment in natural water bodies, later optimized to 75, according to the approach by Cacciatore et al. (2019). Briefly, the method presents a geostatistical approach to optimize the selection of monitoring stations for environmental studies. Although the article primarily addresses macrophytes, the same methodology was applied to macrozoobenthos in the Venice Lagoon. This approach ensures that the selected stations provide comprehensive and representative data while minimizing biases. This adjustment was made without compromising the comparability of data across cycles and remained consistent for all subsequent surveys.
The dataset obtained, based on data published by ARPAV (see Data Availability SEction), was organized into a single abundance matrix of taxonomic units. The stations considered for each campaign were 68 out of the 75 sampled (85 in 2011). Indeed, some stations were relocated for logistical reasons (construction of new lagoon morphological structures, occupation of areas for fishing, clam cultivation etc.). Therefore, the 68 stations refer to those that have remained unchanged in terms of geographical location., and data for all 4 sampling campaigns are available. This abundance matrix consists of 272 samples and 363 taxa. The number of taxa was then reduced to 252 by deleting all those species and taxonomic groups whose sum of individuals was less than or equal to 2, across the 4 samplings campaigns.
For each station, the following descriptors and univariate indices were calculated: total abundance (N), richness (number of taxa), Margalef’s richness index (Margalef, 1958), Shannon’s diversity index (Shannon and Weaver, 1949), and Pielou’s evenness index (Pielou, 1966).
Furthermore, the new reduced matrix was subjected to univariate analysis to calculate diversity indices (DIVERSE) after square root transformation (to reduce the influence of highly dominant taxa and to stabilize variance across samples) and calculation of the Bray-Curtis similarity index and multivariate analysis (Cluster Analysis (CLUSTER), Multidimensional Scaling (MDS), Analysis of Similarity (ANOSIM), Similarity Percentage (SIMPER), and Permutational Analysis of Variance (PERMANOVA)) using the software package Primer 6 & Permanova+ β3. The reduced similarity matrix and the complete similarity matrix for each sampling year were tested using Primer RELATE analysis to test the hypothesis of no relationship between multivariate patterns. Matrices were found to be strongly correlated (Rho values between 0.999 and 1.000 with p ≤.01).
M-AMBI (Borja et al., 2004; Muxika et al., 2007) was applied at all the sampling stations (85 for the first survey, then 75) to assess the ecological quality status according to the requirements of the WFD. Boundaries and reference conditions to apply M-AMBI are water body type dependent, and those defined in the Italian regulations (see D.Lgs. 152/2006 ss. mm.ii.) intercalibrated by the Med-GIG (Mediterranean Geographical Intercalibration group) in European Member States. To compute the M-AMBI value for each WB, the arithmetic means of the EQR values of the stations belonging to the same WB was calculated. This mean was then referenced against the same reference conditions used for each station, as outlined in the aforementioned national normative guidelines (see D.Lgs. 152/2006 ss. mm.ii.).
The Seasonal Kendall test, implemented via the kendall package in R, was applied to detect monotonic trends in the quarterly time series of water parameters from 2011 to 2022. This test extends the Mann-Kendall approach by accounting for intra-annual (seasonal) variability, evaluating trends within each season separately and then aggregating the results. Its use is particularly appropriate for datasets with regular seasonal sampling, as it improves the robustness and interpretability of trend detection in the presence of recurring seasonal patterns (Hirsch et al., 1982; Meals et al., 2011).
For the comparative analyses between macrozoobenthos abundances and water physico-chemical parameters, data from the 4 monitoring cycles of the 28 stations of physico-chemical quality elements were considered, as well as the corresponding stations of the EQB macrozoobenthos distributed in natural water bodies. To perform the appropriate comparisons, a three-year average value of the seasonal water samplings of each monitoring cycle was calculated for a total of 112 data points (28 stations for 4 cycles).
For comparative analyses between sediment parameters and macrozoobenthos, the 68 stations maintained over the years were considered. Because the set of sediment parameters analysed varied across the four sampling campaigns, and to ensure consistency in the statistical treatment of the data, comparisons between sediment characteristics and macrozoobenthic communities were conducted separately for each campaign.
Before performing the analyses, similarity matrices (Bray-Curtis’s similarity) of the appropriately transformed macrozoobenthos data (square root transformation) were calculated, while the abiotic parameter data were normalized and logarithmically transformed when necessary. To minimize autocorrelation and multicollinearity among environmental variables, a variable selection procedure was applied: for granulometric data, only pelite (%) was retained as a representative fraction, while other highly correlated size classes were excluded; for chemical parameters, we retained individual elemental concentrations and excluded summed variables (e.g., TOC) to avoid redundancy. These steps ensured the robustness of the multivariate analyses and reduced the risk of overfitting due to interdependent variables.
The BEST function of BIOENV (Primer v.6) was then applied, which selects environmental variables by maximizing a rank correlation between the respective similarity matrices. Correlations were considered significant for p < 0.01. To better visualize the correlation results, biplot graphs from the Principal Component Analysis (PCA) of water and sediment parameters (R software) and MDS graphs (Primer v.6) of the macrozoobenthos similarity matrices were created.
Pearson’s correlation coefficient was tested (after verifying the assumptions of linearity and normal distribution) for each monitoring cycle (2011, 2014, 2019 and 2022) among sediment parameters consistently analysed across all surveys (pelite, inorganic nitrogen and organic carbon) and the abundances and Shannon’s diversity index values (R software).
Results
Biodiversity and ecological status of macrozoobenthos
The total abundance and richness decreased especially in the last year (2022), as well as Shannon’s diversity and Margalef’s richness indices, whilst Pielou’s evenness index increased over time (Figures 2a-e). The distribution of taxa abundances (Figure 2f) showed the disappearance of some taxa over time, such as Clitellata, Hexapoda, Bryozoa, Copepoda, Nematoda and Pycnogonida, although very few organisms were found to belong to these last four taxa. Other taxa were halved or nearly disappeared such as Crustacea, Polychaeta, Bivalvia, Cnidaria, Echinodermata, Tunicata, Nemertea. The only taxon which increased was Phoronida, whilst Gastropoda, Decapoda and Polyplacophora (except in 2014) remained fairly stable across the surveys.
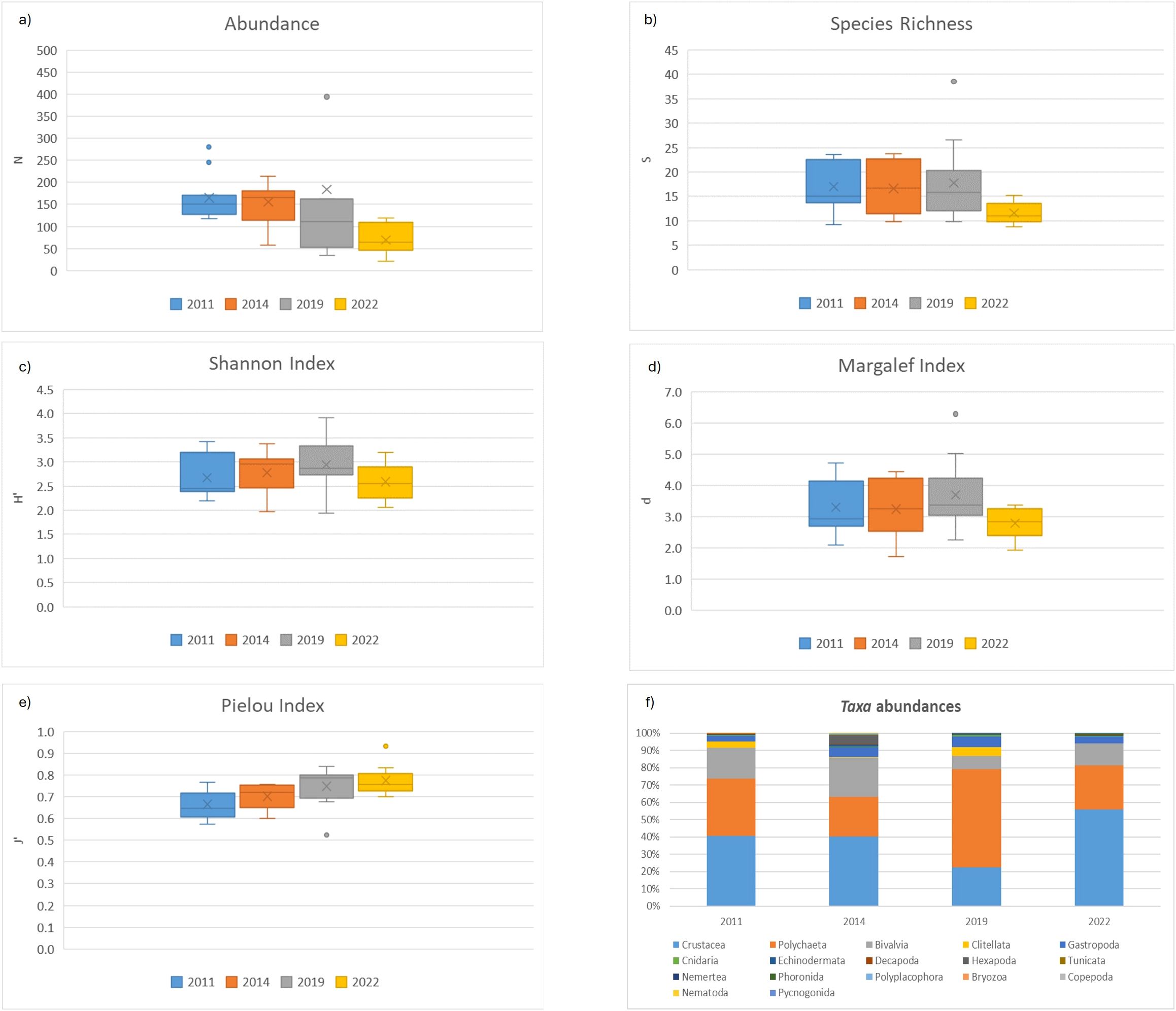
Figure 2. Box and whiskers of (a) abundance, (b) species richness (c) Shannon’s diversity index, (d) Margalef’s richness index, (e) Pielou’s evenness index and (f) taxa abundances, in macrozoobenthos sampled from 2011 to 2022 in the Venice Lagoon. (x = mean, – = median, box limits are 25° and 75° percentiles, whiskers are max and min).
The complete list of macrozoobenthic species recorded during the study is provided in Supplementary Table S1. Species highlighted in blue are those not listed in the current SIBM checklist of macrozoobenthic species of the Adriatic and Mediterranean seas. The CLUSTER analysis showed that, at a similarity level of 20%, eight significant groups with more than four elements can be identified. These groups were defined by the year of samplings and hydrological condition (choked/not choked) of water bodies (WB). In the first group, samples from 2011 predominated; in the second group, there were mainly choked (C) samples, especially from 2011 and 2014; in the third group, samples from 2014 and 2019 were predominant; in the fourth group, samples from non-choked (NC) stations prevailed; the fifth group consisted almost entirely of NC samples from 2011 and 2014; the sixth group also consisted of mainly NC samples, especially from 2022; the seventh group mostly included samples from 2019 and some from 2022; finally, the eighth group was entirely composed of samples from 2019 (mostly NC). Overall, it can be said that samples from 2019 and 2022 differed more from those of 2011 and 2014, which formed less homogeneous groups, and that many samples from 2022 tended to form small groups (with fewer than five elements), indicating a strong heterogeneity within the year.
The MDS graph (stress value of 0.28) depicts a differentiation both by confinement (Figure 3) and by salinity (Figure 4). ANOSIM results (Table 1) showed that “years” was among the factors that maximize intra-group similarity and minimize that from inter-group, followed by “salinity” and then “confinement”. The pairwise test confirmed that all years were significantly different (p = 0.01%) with lower R values between 2011 and 2014 and higher between 2011 and 2019. SIMPER analysis also confirmed lower similarity in each year (13-21%) and higher between years (mean dissimilarity values >83%): the lowest dissimilarity was between 2011 vs 2014 (83%), whilst the highest dissimilarity was between 2019 vs 2022 (90%). These findings mostly depend on high heterogeneity of low frequency species among groups, as well as the variation on abundances of the same species with relative higher abundance and common at all groups. The most common species were the bivalve Abra segmentum (Récluz, 1843) and the amphipod Gammarus insensibilis (Stock, 1966).
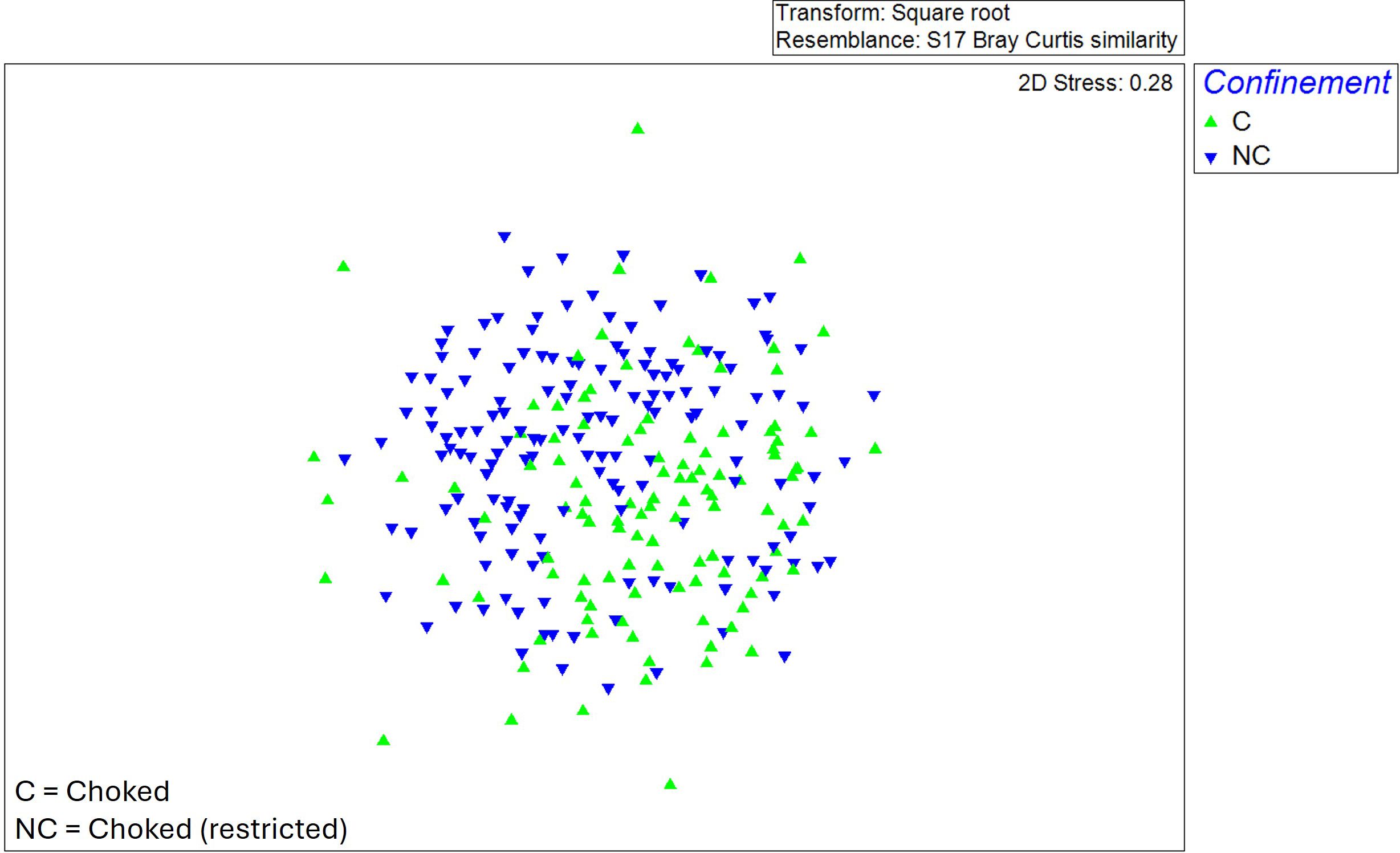
Figure 3. MDS (Multidimensional Scaling) plot with differentiation of samples by confinement (choked/unchoked) of stations.
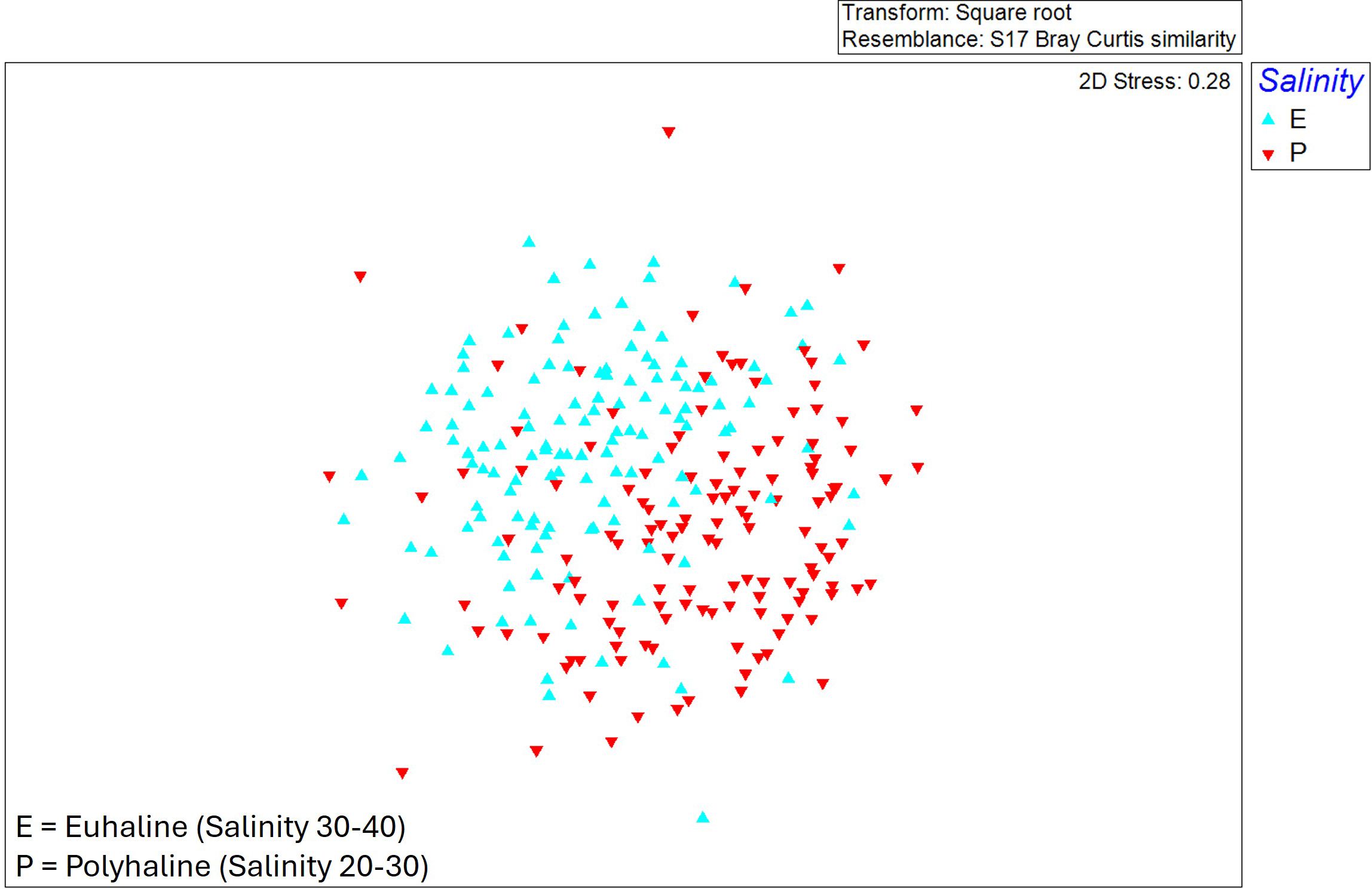
Figure 4. MDS (Multidimensional Scaling) plot with differentiation of samples by salinity (Euhaline/polyhaline) of stations.
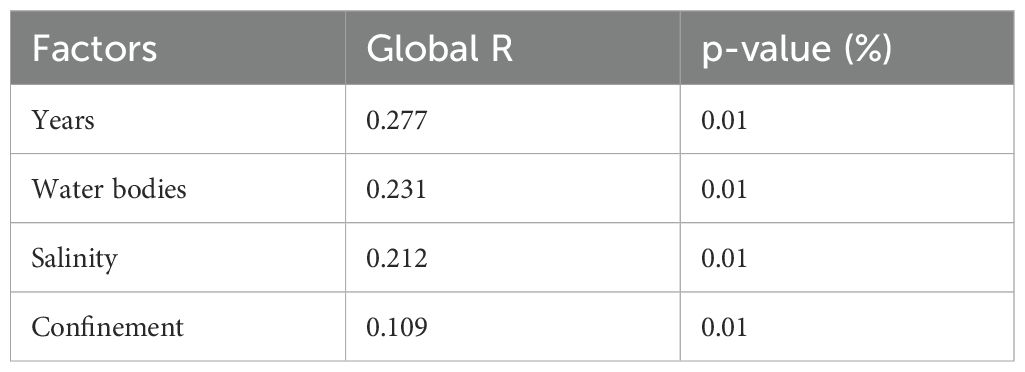
Table 1. Results of the analysis of similarity (ANOSIM) between groups identified by each factor: years (4 groups), water bodies (11 groups), salinity (2 groups), confinement (2 groups).
The application of two-way crossed PERMANOVA (Table 2) among groups determined by the factors “water bodies” (fixed, 11 levels) and “years” (fixed, 4 levels) allowed for the verification of whether these groups were significantly different from each other and if there was an interaction between the two factors. Both factors were found to be significant, as it was the interaction. Higher pseudo-F also indicated that the temporal factor is the most determining factor.
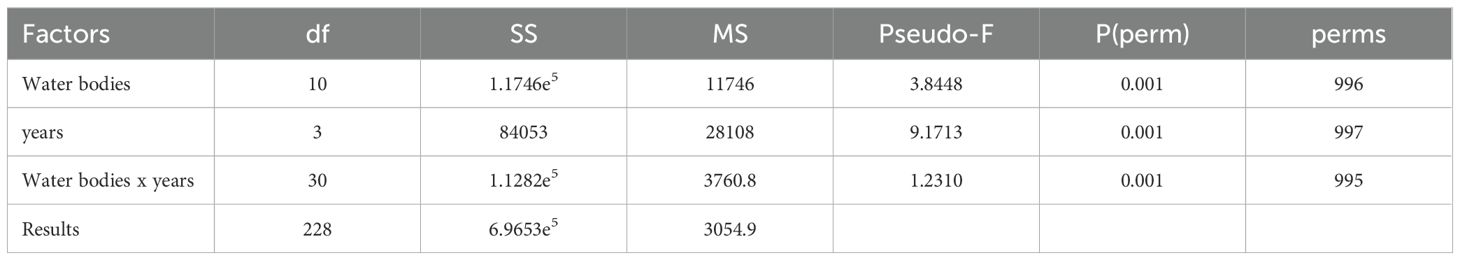
Table 2. Results of the Permutational Analysis of Variance (PERMANOVA) between groups identified by each factor: water bodies (11 groups) and years (4 groups).
Figure 5 depicts a decrease of ANOSIM’s R values in salinity-based differentiation, relatively stable values for water body differentiation after 2014, and an alternation of significance and non-significance results when seeing the confinement-related differentiation.
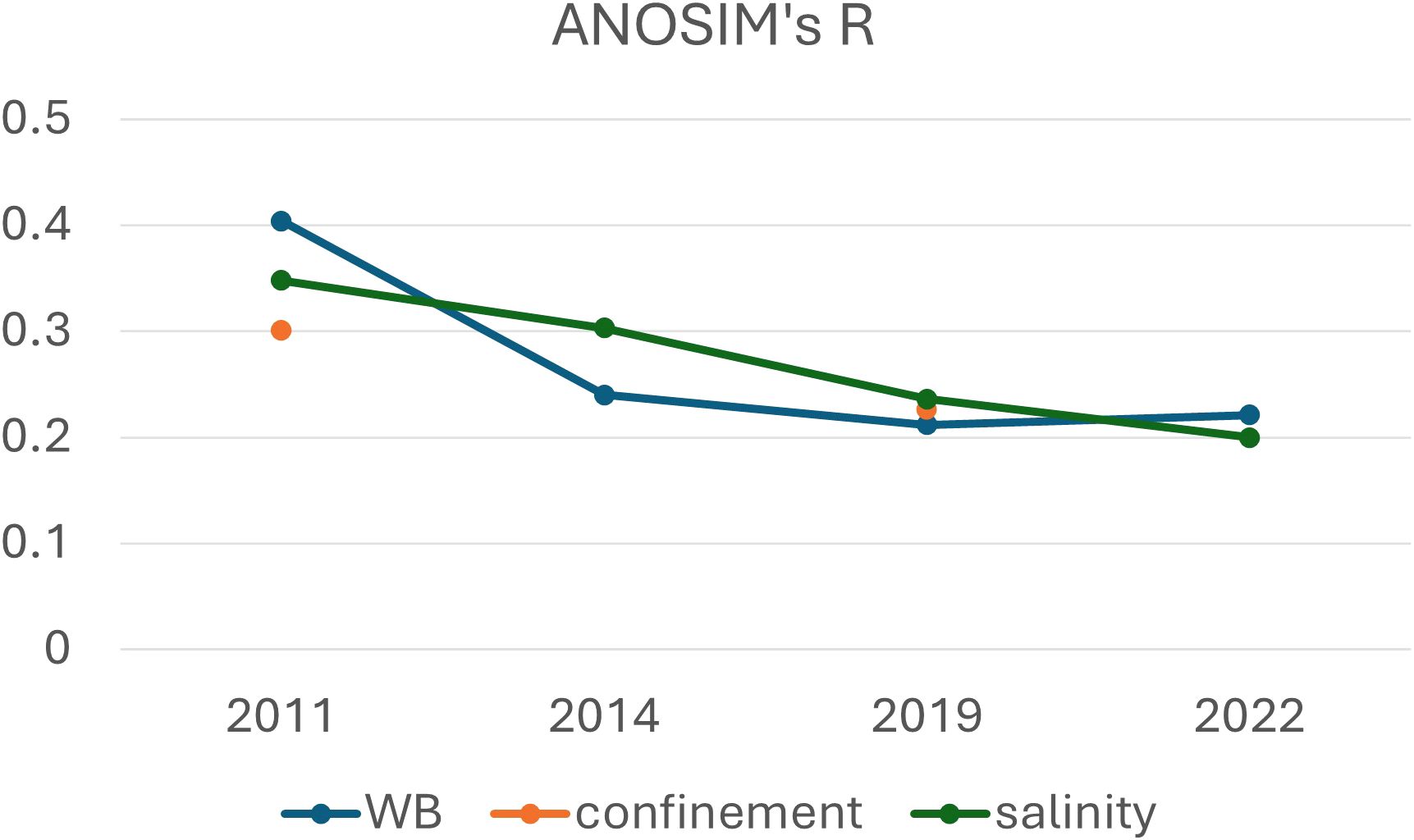
Figure 5. R trends of ANOSIM (Analysis of Similarity) resulted significantly (P =0.01). No data means not significant.
Ecological quality status of macrozoobenthos assessed by M-AMBI is reported in Figure 6. Ecological quality ratio (EQR) values ranged between 0.29 to 0.97 and covered all quality status from Bad to High. In the first monitoring cycle (2011), the water bodies’ EQR scores ranged between 0.54 and 0.74, with 2 WBs in Good ecological status, 1 in Poor, and the others in Moderate status. In the second cycle (2014), scores ranged between 0.57 and 0.81; only one WB was recorded in Good status, 1 in Poor, and the remaining 9 in Moderate condition. In the third cycle (2019), all WB were in Moderate condition except for 3, which were in Good condition; EQR values ranged between 0.57 and 0.82. Finally, in the fourth monitoring cycle (2022), no water bodies were in Good status, 4 were in Poor, and the remaining 7 were in Moderate status, with EQR values ranging between 0.54 and 0.70. Compared to 2011, in 2022 there was a deterioration in 5 water bodies: ENC1 and PC2, which were in Good status in 2011, were in Moderate status in 2022; ENC2, ENC4, and PC4, which were in Moderate status in 2011, were in Poor status in 2022. Compared to 2014, in 2022 the water bodies EC, ENC2, and PC4 shifted from Moderate to Poor status, and only one water body (PC3) shifted from Good to Moderate status; the other water bodies remained unchanged. Finally, compared to 2019, in 2022 the ecological quality deterioration affected 6 water bodies: EC, ENC4, and PC4 shifted from Moderate to Poor status; PC2 and PNC2 shifted from Good to Moderate status, while ENC2 shifted from Good to Poor status.
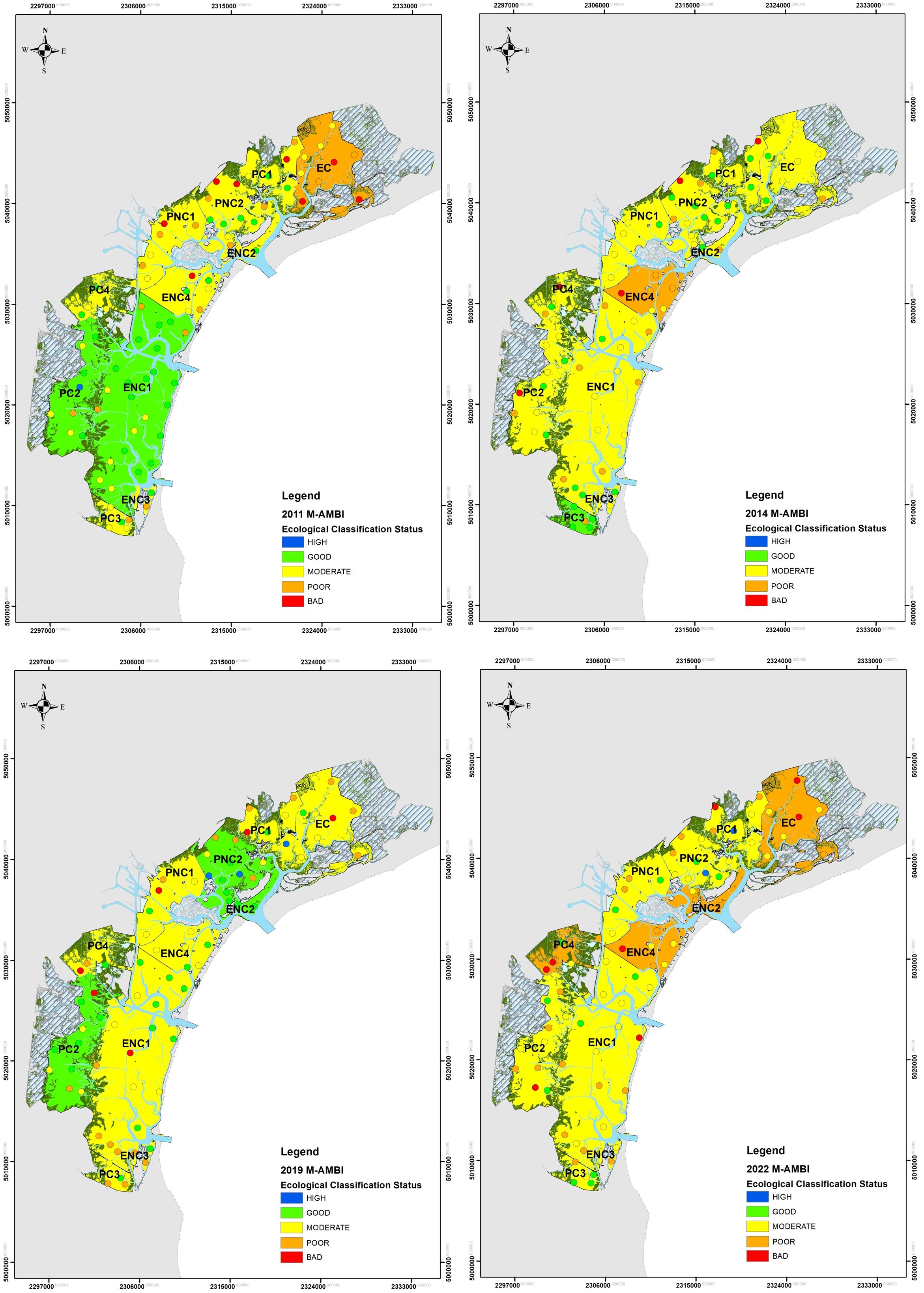
Figure 6. Maps of the ecological quality status of macrozoobenthos assessed by M-AMBI in stations and water bodies of the Venice Lagoon from 2011 to 2022.
Figure 7 shows that, in the latest monitoring cycle, the average M-AMBI values for all natural water bodies in the Venice Lagoon reflect the lowest levels of species richness and diversity, indicating a general decline in macrozoobenthic biodiversity. Conversely, the first three monitoring cycles generally show better quality, with higher diversity values in the second and third cycles, and higher richness in the first cycle.,
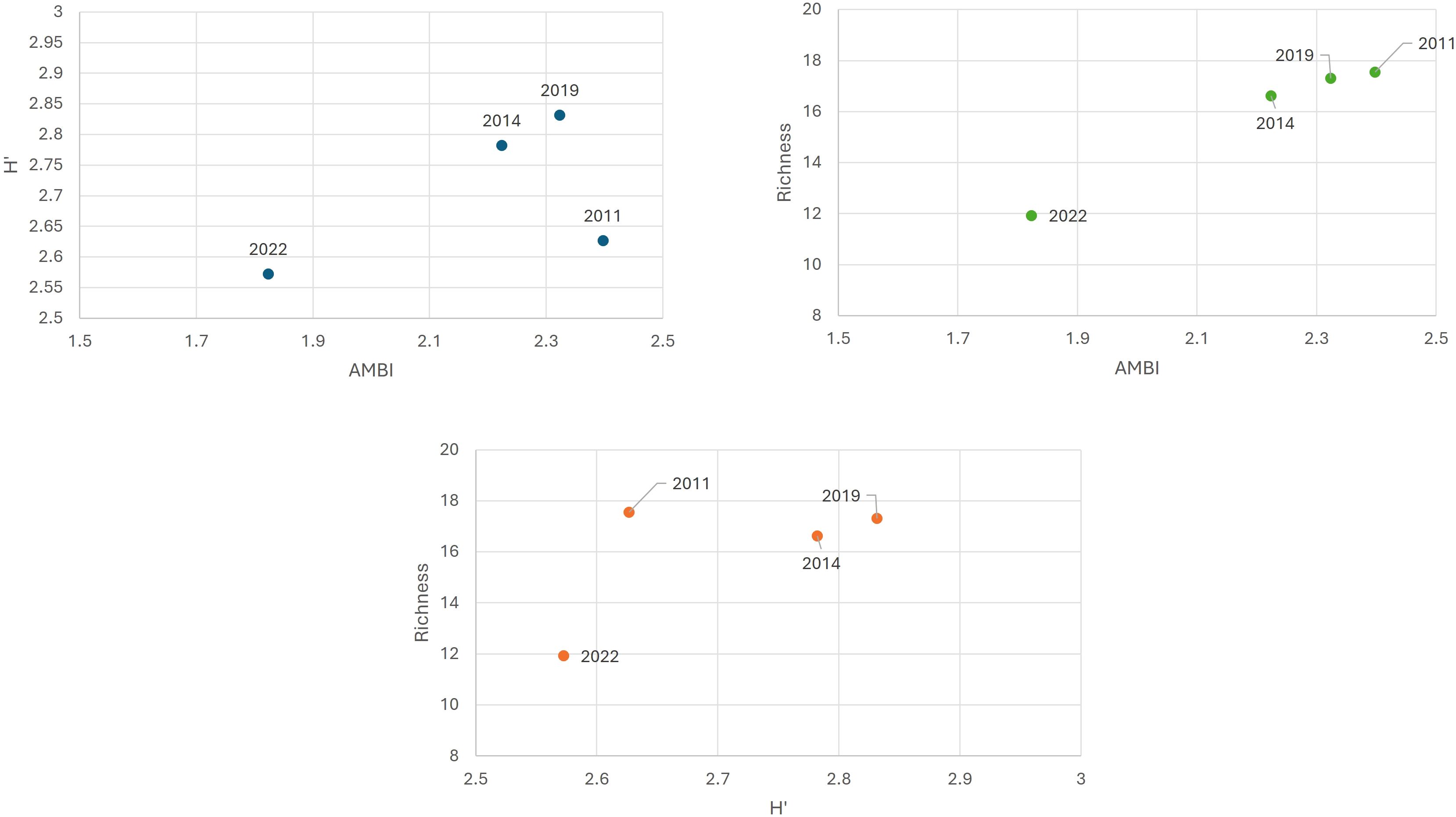
Figure 7. Relationship between the parameters of the M-AMBI index: Shannon Diversity Index (H’), number of taxa (Richness), AMBI for the Venice Lagoon for each monitoring cycle (2011, 2014, 2019, 2022).
Water and sediment parameters
Table 3 reports the results of the application of the Seasonal Kendall’s test in the trend analysis of water parameters sampled seasonally each year from 2011 to 2022. Figure 8 depicts trends of those parameters that were found to be significant in the Seasonal Kendall’s test at the lagoon scale. Nitrogen compounds showed that throughout the lagoon there was a general decreasing trend in concentrations over the years, significant for all analyzed forms except for nitrates. The trend analysis of phosphorus compounds was limited to total dissolved phosphorus (TDP), as orthophosphate (P-PO4) was often present in concentrations below the limit of quantification (LOQ) of analytical instruments. Like nitrogen compounds, there was a significant decrease in TDP concentration throughout the lagoon over the years. Considering the carbon compounds, i.e. total organic carbon (TOC), particulate organic carbon (POC), and dissolved organic carbon (DOC), the Seasonal Kendall’s test yielded significant results only for the latter parameter, despite all three compounds showing a general decreasing trend over the years. Both chlorophyll a and dissolved oxygen (O2) values showed a decreasing trend over the analyzed period, with significance observed only for dissolved oxygen.
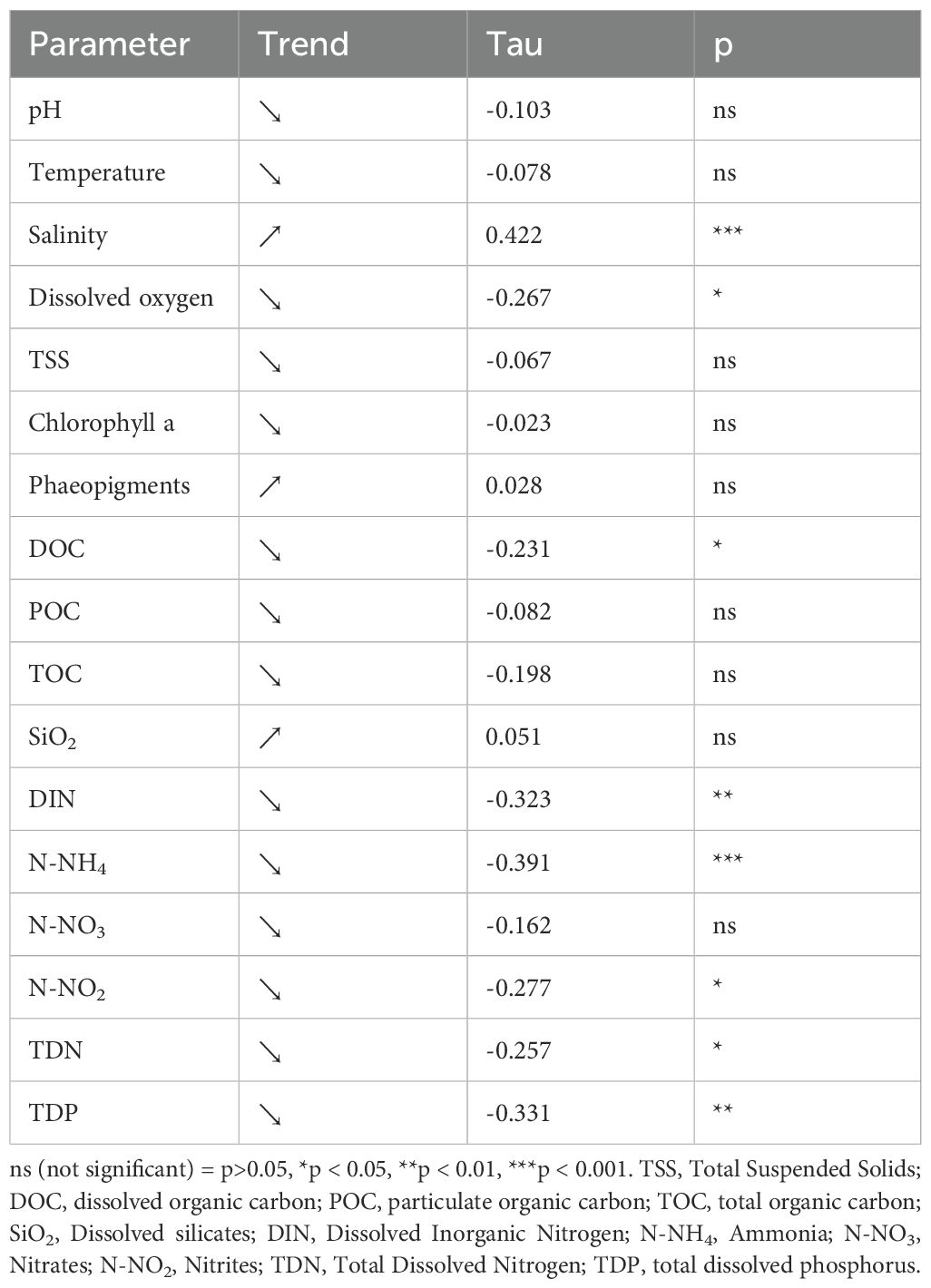
Table 3. Seasonal Kendall test’s results of water parameters sampled seasonally from 2011 to 2022.
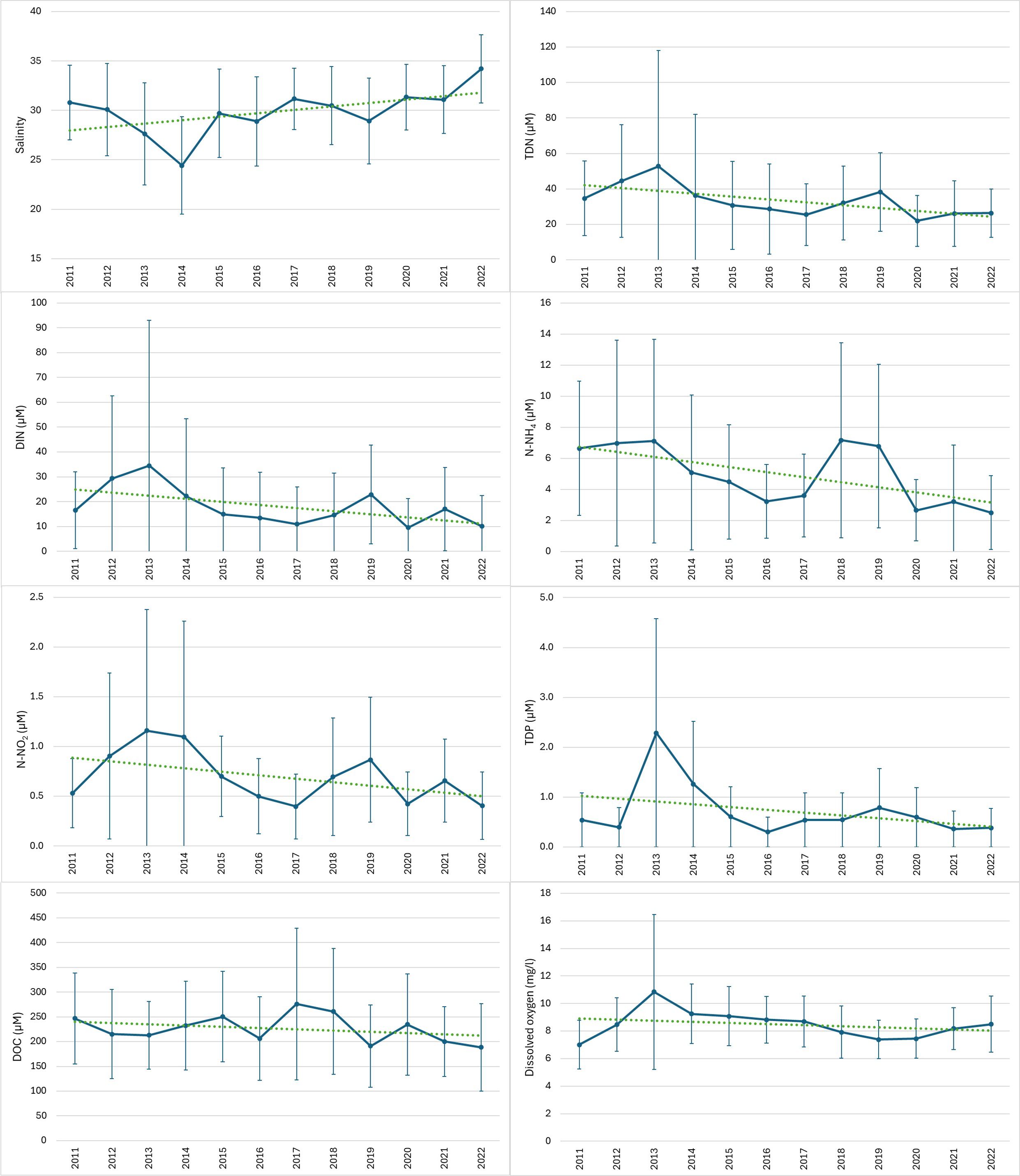
Figure 8. Trends of concentrations of parameters (salinity, TDN, DIN, N-NH4, N-NO2, TDP, DOC, dissolved Oxygen) from 2011 to 2022, found to be significant in the Seasonal Kendall’s test at the lagoon scale. Bars indicate standard deviations.
Table 4 reported the results of all sediment parameters analyzed in the Venice Lagoon from 2011 to 2022. Figure 9 depicts the results of the three sediment parameters analyzed across the four monitoring cycles (i.e.: pelite, total nitrogen and total organic carbon).
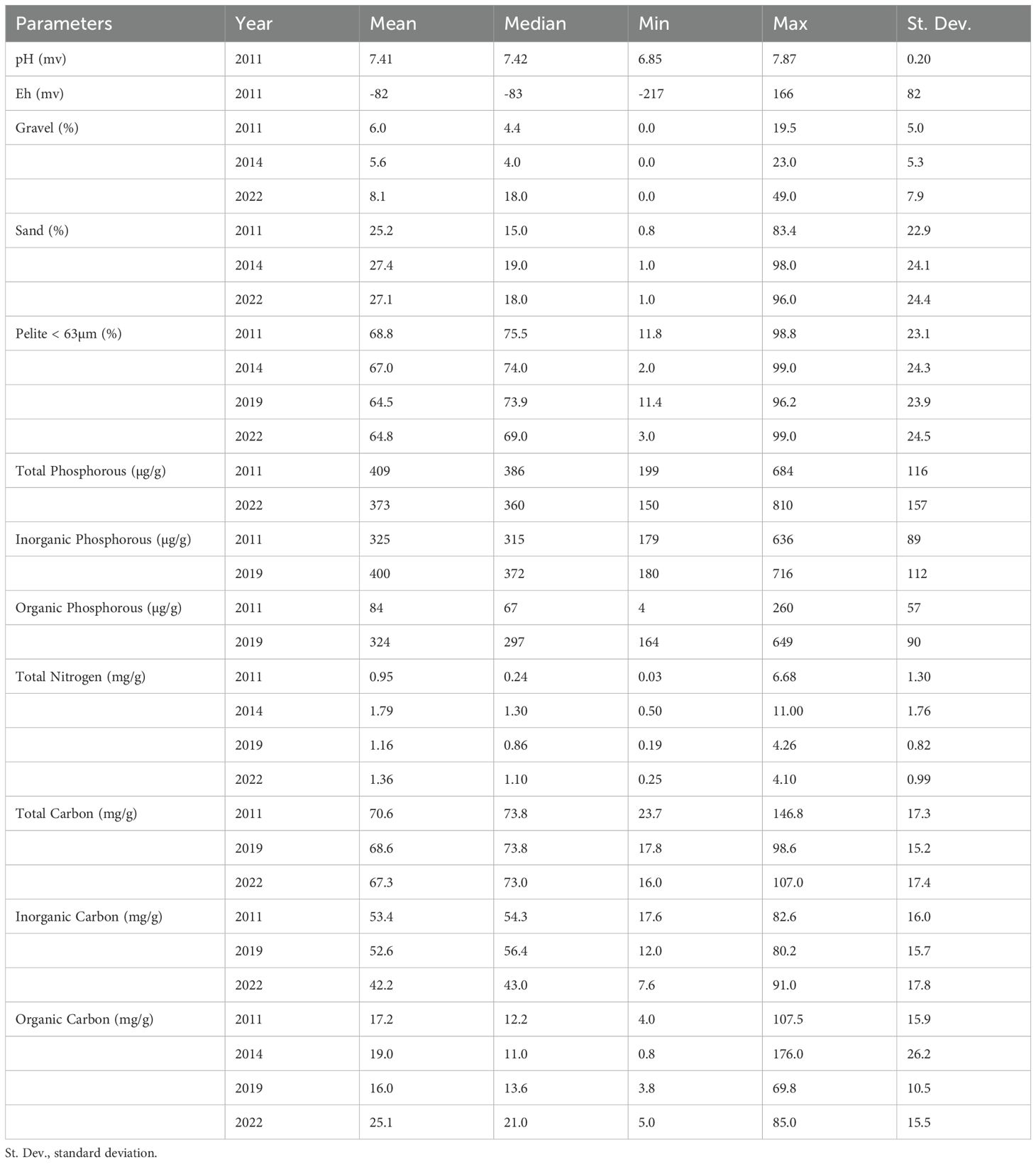
Table 4. Sediment parameters resulted in the Venice Lagoon from 2011 to 2022.
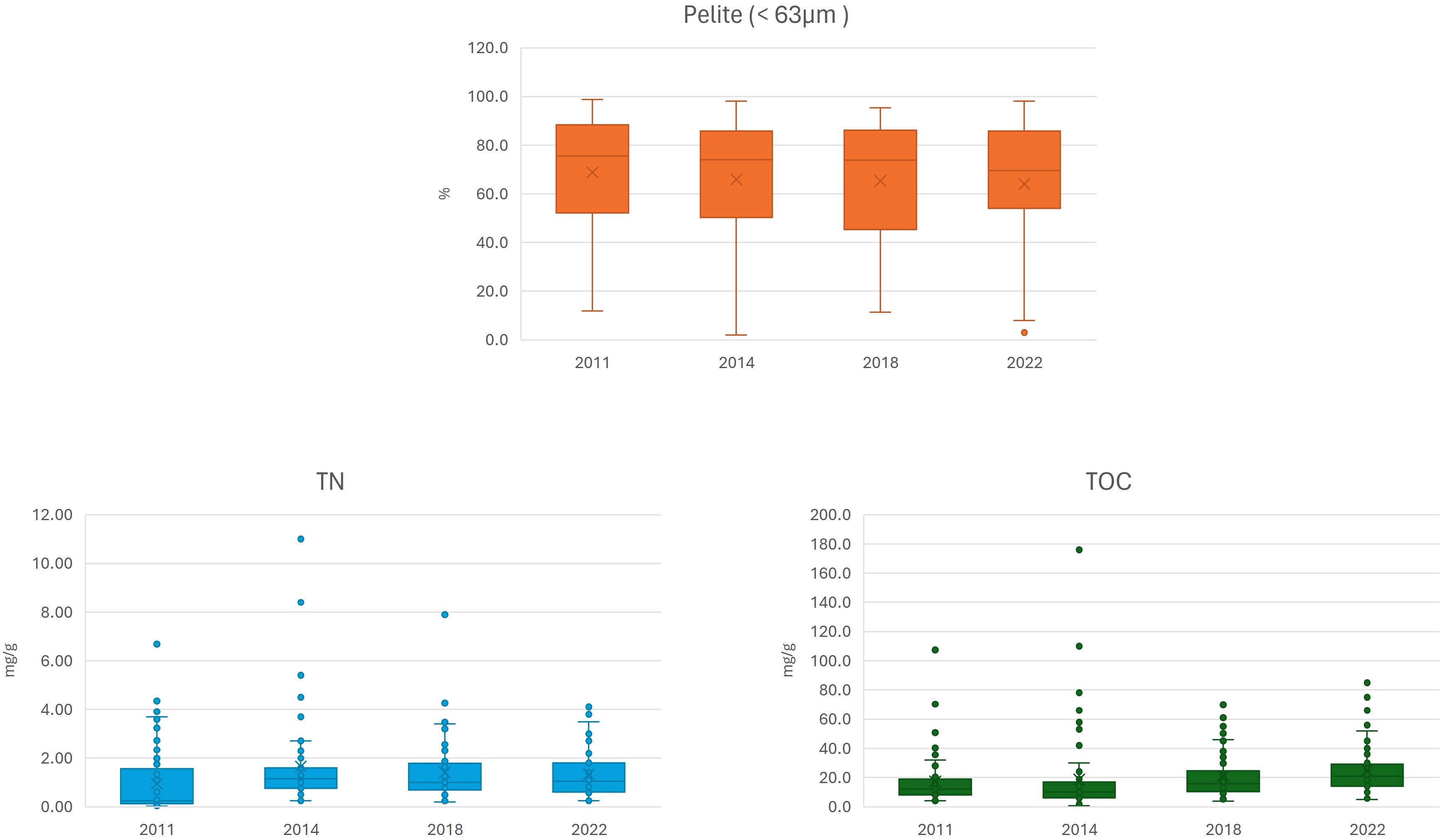
Figure 9. Pelite, total nitrogen (TN) and total organic carbon (TOC) concentrations of sediments in the Venice Lagoon across the four surveys from 2011 to 2022.
Environmental parameters and macrozoobenthos relations
The results of the comparison between the chemical-physical parameters of water and the macrozoobenthos are reported in Figure 10. The PCA score plot (Figure 10a) allows for the characterization of water samples according to the first and second components, which together explain 61.5% of the total variance. The first component includes the variables salinity, TDN, TDP, TSS, POC, and Chl a, while pH, dissolved oxygen, and temperature belong to the second component. DOC and silicates are in an intermediate position.
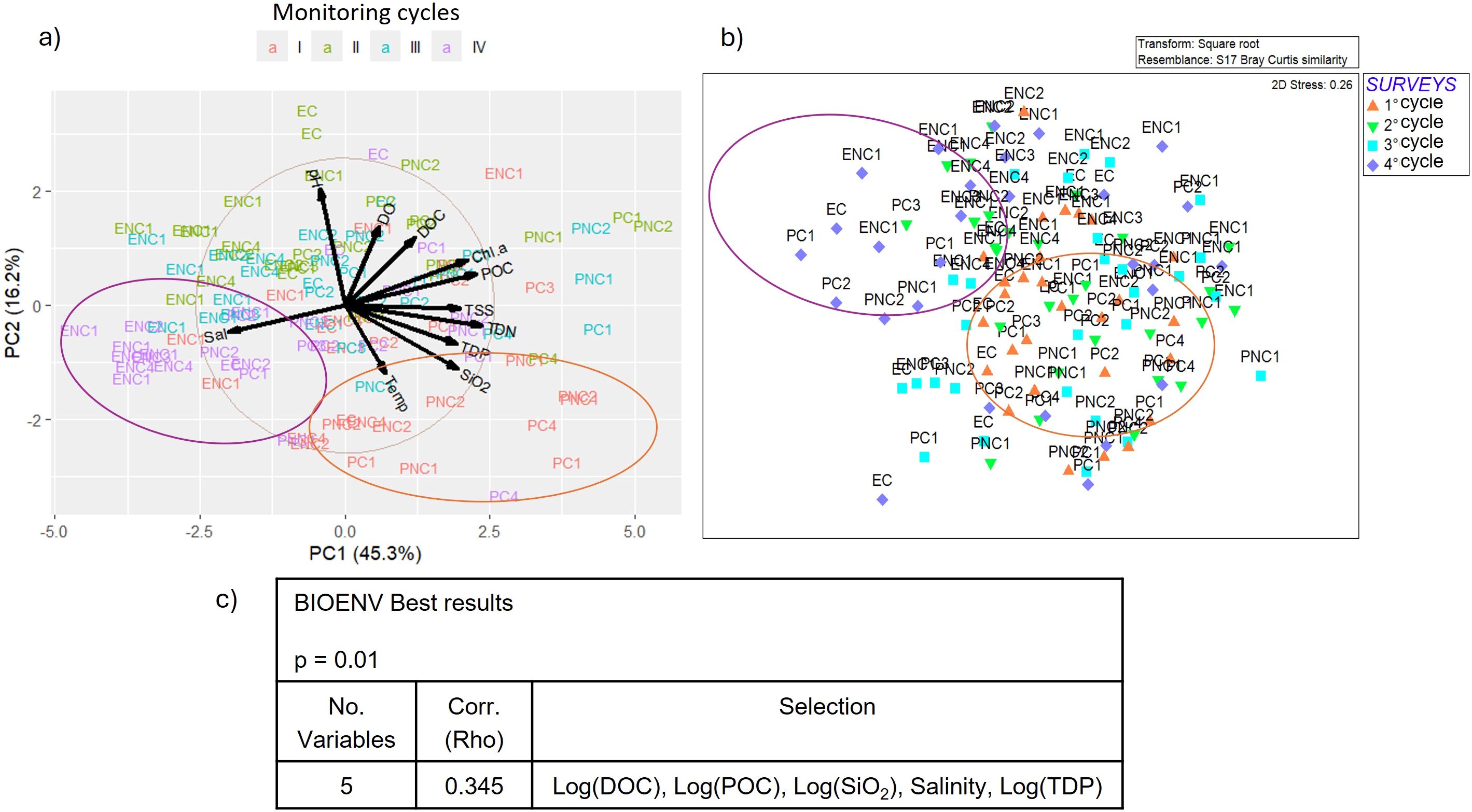
Figure 10. (a) Biplot of PCA (Principal Component Analysis) applied to water parameters, (b) MDS (Multidimensional Scaling) plot of macrozoobenthos similarity matrix and (c) BEST (BIOENV) results showing which environmental variables best explain the macrozoobenthic assemblages in the Venice Lagoon from 2011 to 2022. (DOC=Dissolved Organic Carbon, POC = Particulate Organic Carbon, SiO2 = Silicates, TDP = Total Dissolved Phosphorous).
By comparing the PCA results with the MDS plot (Figure 10b), two groups of cases can be identified, corresponding, respectively, to the first and last monitoring cycles. The BIOENV analysis (Figure 10c) showed that DOC, POC, silicates, salinity, and TDP are the environmental variables most significatively correlated (Rho=0.345, p=0.01%) between the two similarity matrices.
The results of the comparison between the sediment parameters and the macrozoobenthos for each monitoring cycle are reported in Figures 11-14. The PCA results of the abiotic parameters (Figures 11a-14a) are mostly similar, despite modifications to the analytical panel over the years, consistent with the largely conservative environmental matrix. The factor weight plots for all four cycles show that sediment samples mainly differed in grain size and organic content.
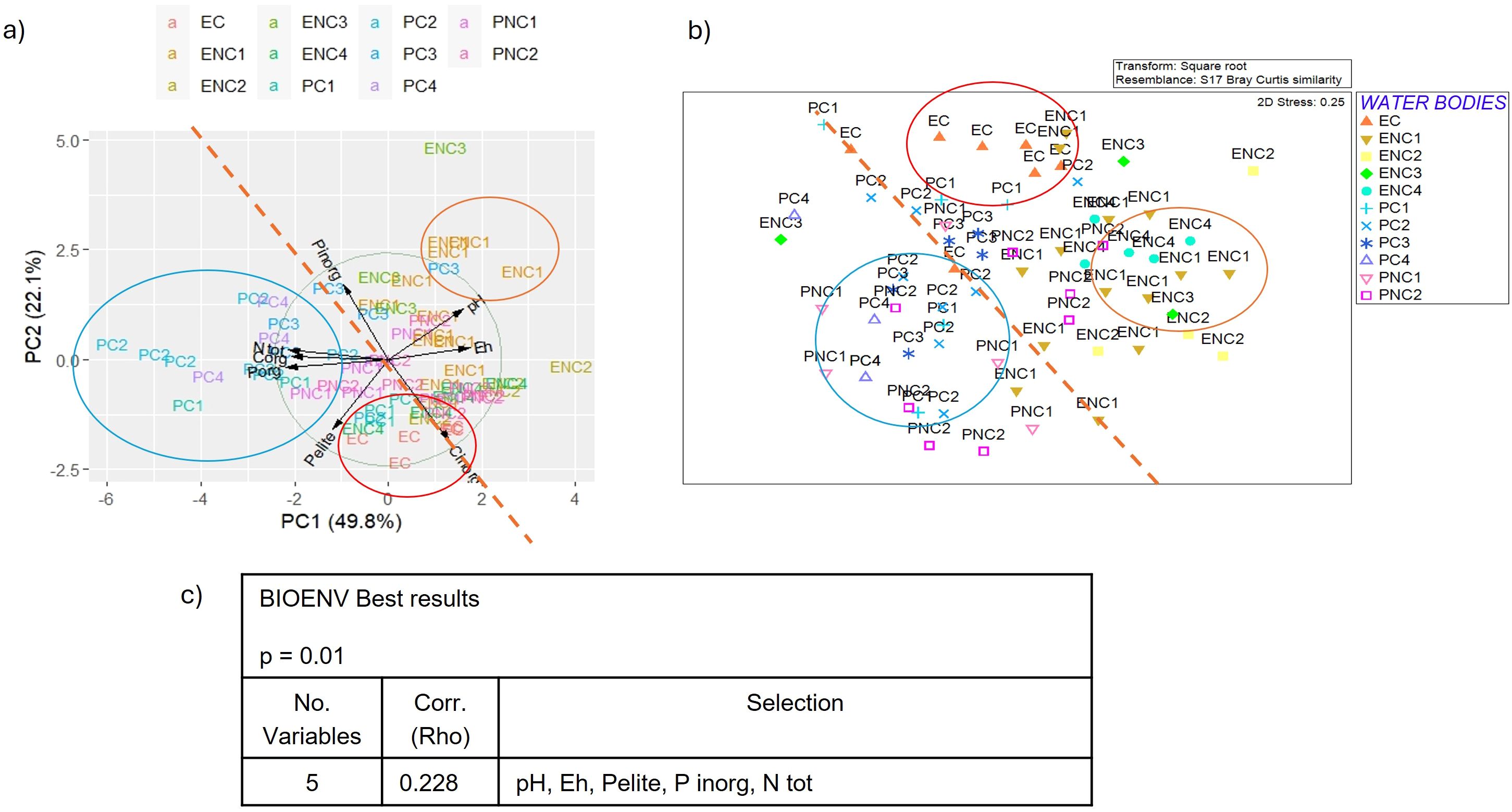
Figure 11. (a) Biplot of PCA (Principal Component Analysis) applied to sediment parameters, (b) MDS (Multidimensional Scaling) plot of macrozoobenthos similarity matrix and (c) BEST (BIOENV) results showing which environmental variables best explain the macrozoobenthic assemblages in the Venice Lagoon in the first monitoring cycle (2011). (P inorg = inorganic Phosphorous, N tot = Total Nitrogen).
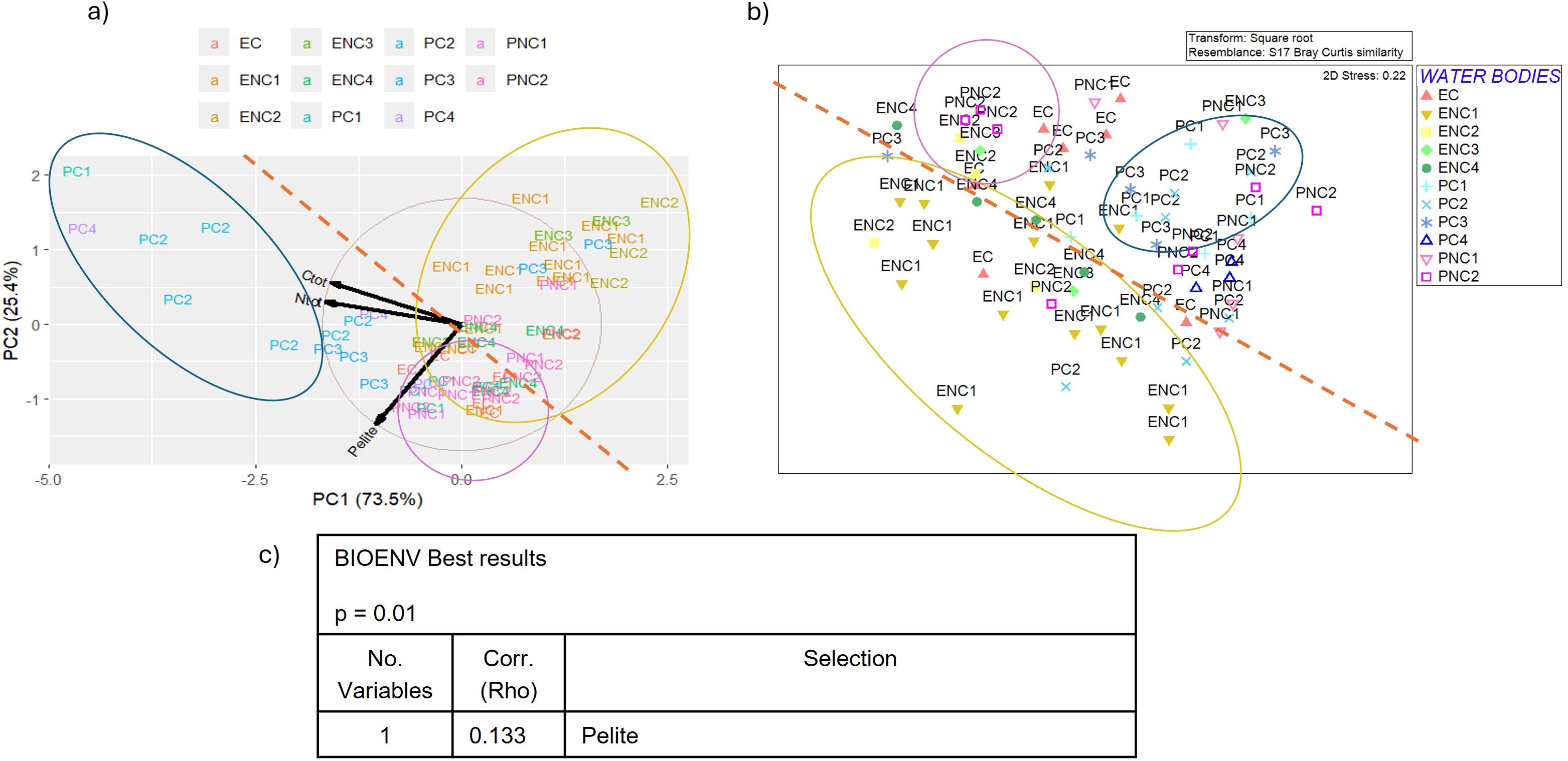
Figure 12. (a) Biplot of PCA (Principal Component Analysis) applied to sediment parameters, (b) MDS (Multidimensional Scaling) plot of macrozoobenthos similarity matrix and (c) BEST (BIOENV) results showing which environmental variables best explain the macrozoobenthic assemblages in the Venice Lagoon in the second monitoring cycle (2014).
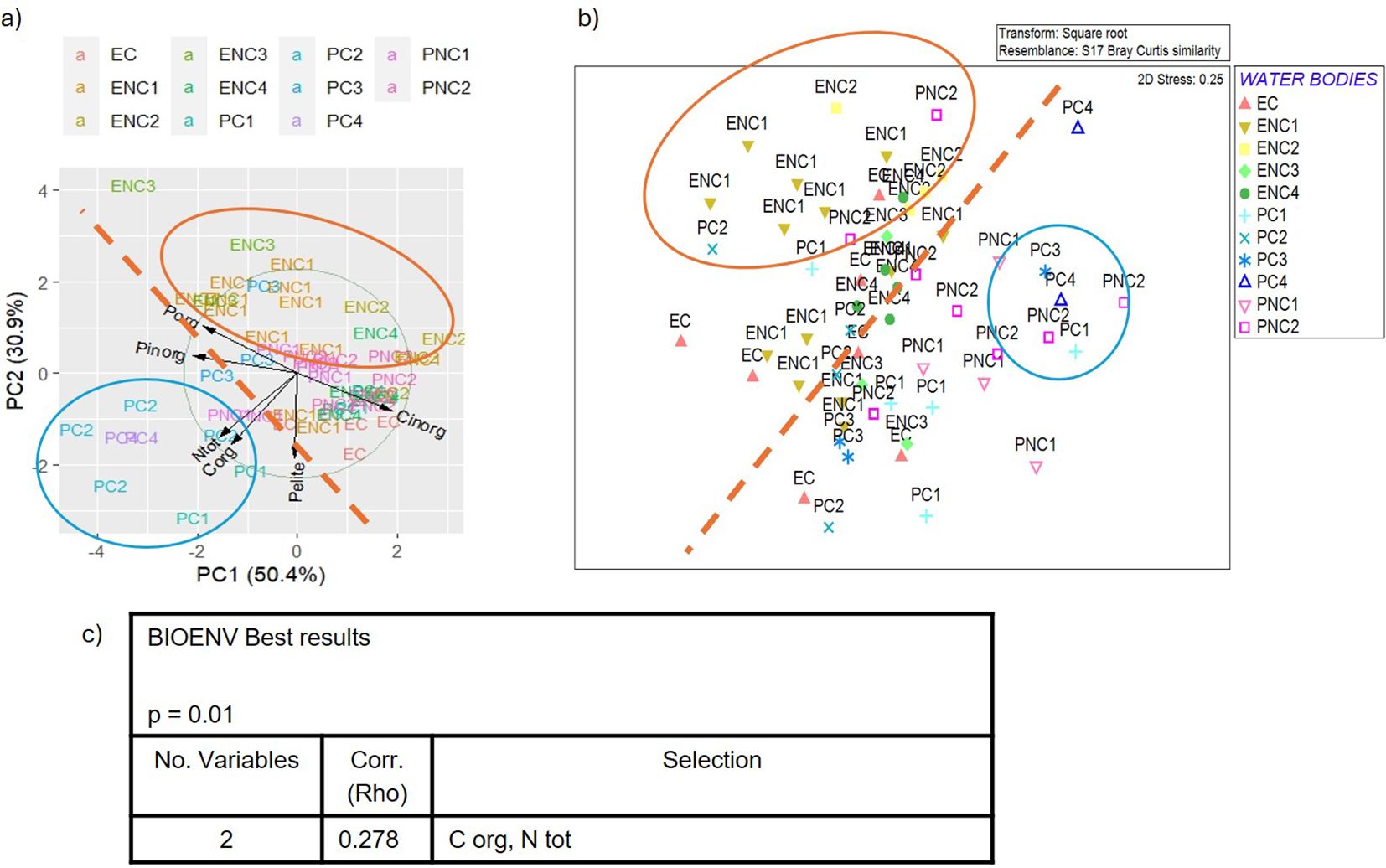
Figure 13. (a) Biplot of PCA (Principal Component Analysis) applied to sediment parameters, (b) MDS (Multidimensional Scaling) plot of macrozoobenthos similarity matrix and (c) BEST (BIOENV) results showing which environmental variables best explain the macrozoobenthic assemblages in the Venice Lagoon in the third monitoring cycle (2019). (C org = total organic Carbon, P inorg = inorganic Phosphorous, N tot = total Nitrogen).
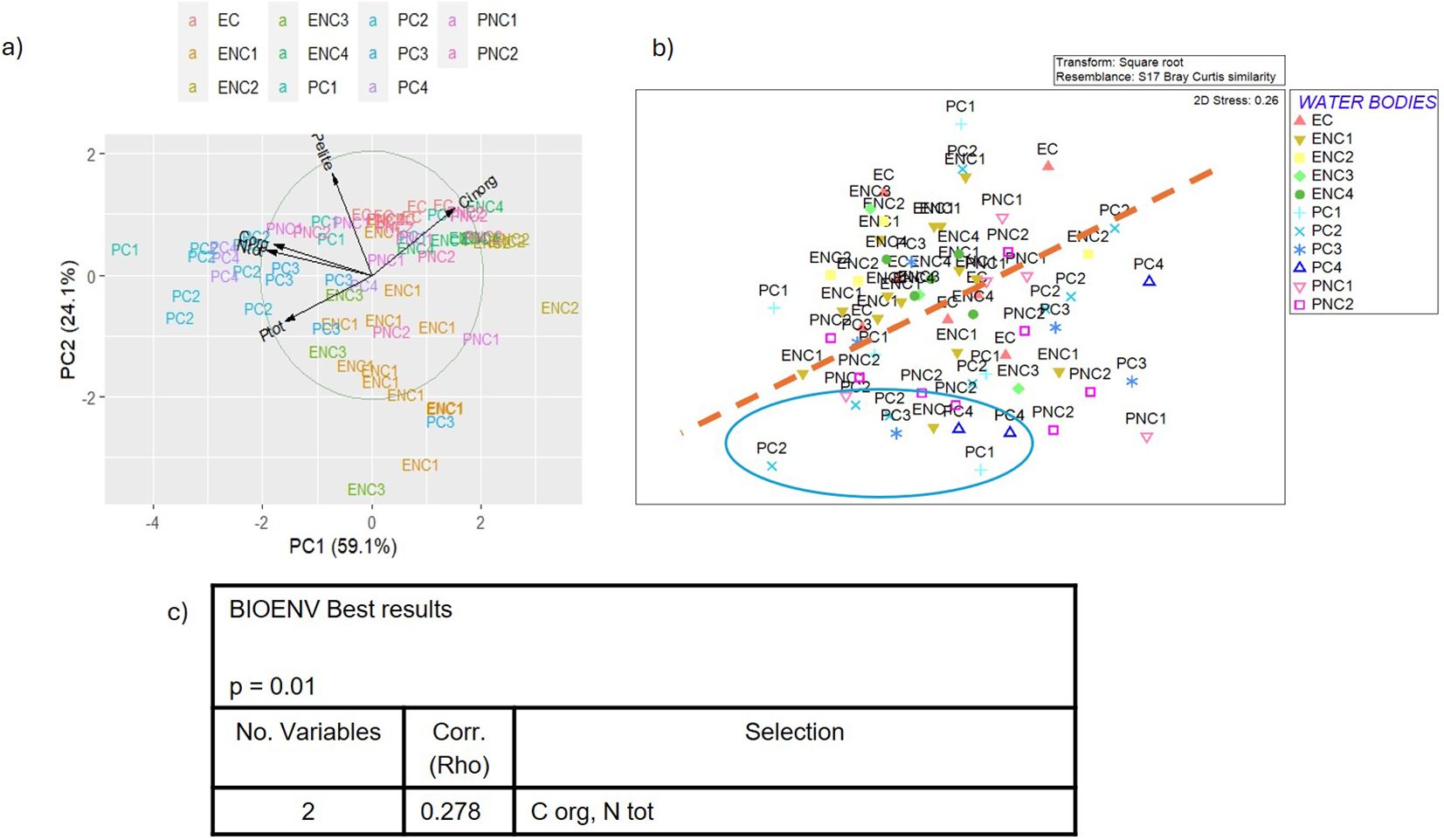
Figure 14. (a) Biplot of PCA (Principal Component Analysis) applied to sediment parameters, (b) MDS (Multidimensional Scaling) plot of macrozoobenthos similarity matrix and (c) BEST (BIOENV) results showing which environmental variables best explain the macrozoobenthic assemblages in the Venice Lagoon in the fourth monitoring cycle (2022). (C org = total organic Carbon, N tot = total Nitrogen).
In the first cycle, 2011 (Figure 11a), the first component (explaining 49.8% of the total variance) includes total nitrogen, total organic carbon and phosphorus, Eh and pH, while pelite, inorganic carbon and phosphorus are positioned between the two main components. In the second cycle, 2014 (Figure 12a), only three parameters were usable for PCA, after removing autocorrelations, with total carbon and nitrogen belonging to the first component (73.5% of the total variance), and pelite to the second (25.4% of the total variance). In the third cycle, 2019 (Figure 13a) the first component (50.4% of the total variance) includes inorganic phosphorus and carbon, while pelite belongs to the second component (30.9% of the total variance). Finally, in the fourth cycle, 2022 (Figure 14a) the PCA plot is very similar to that of the first cycle, with the first component (59.1% of the total variance) characterized by organic carbon, total nitrogen and phosphorus, while pelite belongs to the second component (24.1%).
Comparing the PCA and MDS plots for each cycle (Figures 11b-14b), similar groupings are evident over the years, with choked samples on one side and non-choked on the other. Notably, most samples from choked polyhaline WBs form a distinct group. In the first cycle, three groups are identified: the group of choked polyhaline WB samples, the group of samples from the only choked euhaline WB in the lagoon, and the group of non-choked euhaline samples. In the second cycle, the groups are again formed by choked polyhaline, non-choked euhaline samples, and a heterogeneous group of non-choked polyhaline and euhaline samples. In the third cycle, besides the choked polyhaline group, a group mainly represented by ENC1 and ENC2 samples is identified. Finally, in the last cycle, the only recognizable group in both data distributions is represented by choked polyhaline samples. BIOENV results of each cycle are reported in Figures 11c-14c.
Pearson’s correlation analyses among sediments parameters and macrozoobenthos characteristics revealed significant inverse correlation between Shannon’s index and organic carbon contents in 2014 (R = -0.6, p < 0.05), 2019 (R = -0.53, p < 0.05) and 2022 (R = -0.70, p< 0.05).
Discussion
The analysis of the biodiversity of the macrozoobenthic community in the Venice Lagoon, based on data collected from 2011 to 2022, during ecological classification monitoring according to WFD, revealed a temporal decrease in taxa and number of organisms. Spatially, polyhaline water bodies consistently exhibited lower richness than euhaline ones, while choked water bodies had fewer species compared to those non-choked. This spatial heterogeneity is quite common in benthic assemblage in relation to different abiotic environmental characteristics, such as hydrology and sediment texture (Kneib, 1984; Pearson and Rosenberg, 1978; Dauer et al., 2000). However, 2022 generally recorded the lowest number of species, while the species richness of previous years remained relatively similar; in terms of abundance, there was a continuous year-on-year decline from 2011 to 2022. Moreover, the analysis of similarity (ANOSIM’s R) of inter- and intra-groups confirmed a consistent decrease in salinity-based differentiation, while there was a stabilization in the differentiation between water bodies, and an alternation between significance and non-significance in confinement-related differentiation. These findings indicate a trend towards homogenization, particularly concerning confinement. This is consistent with numerical simulations that predicted, in a global warming perspective, a general reduction in variability of physical characteristics in the Mediterranean lagoons, with a tendency towards marinization (Ferrarin et al., 2014). The sharp decline in species richness and abundance observed in 2022 coincides with an extreme climatic event. According to ARPAV (2023), 2022 was the warmest year on record in the Veneto region, with an average air temperature anomaly of +1.2 °C compared to the 1993–2021 reference period. The year was also characterized by recurrent heatwaves and exceptionally low precipitation, leading to widespread drought. As reported by ISPRA (2023), these conditions resulted in the highest annual average salinity values recorded in the Venice Lagoon over the past decade. Such combined thermal and hydrological stressors likely contributed to the observed ecological shifts in benthic communities.
The macrozoobenthic community in Venice Lagoon presented a faunal composition that is characteristic of lagoon environments with the dominance of Crustacea, Polychaeta and Bivalvia. Similar results were obtained for other lagoons in the Northern and Southern Adriatic Sea and Sardinia Island (Sfriso et al., 2001; Marzano et al., 2003; Munari et al., 2010; Oselladore et al., 2022). However relative abundances changed over time, as well as some taxa declined sharply or nearly disappeared.
A general decline in species richness was observed when comparing the earlier sampling years (2011 and 2014) with the more recent ones (2019 and 2022), with 2022 showing the lowest overall diversity. This trend was consistent across the most represented taxonomic groups, including polychaetes, amphipods, and bivalves. Polychaetes showed a partial recovery in 2019, mainly due to the proliferation of spionids, particularly Streblospio shrubsolii. This species is known for its opportunistic behaviour, short lifespan, and rapid growth, and is often considered an indicator of environmental stress (Borja et al., 2000). Chironomids were scarcely present in 2011 and 2019 and completely absent in 2022, with a notable peak only in 2014. Decapods maintained relatively stable numbers across the years, except for a marked increase in 2014, although their overall abundance remained low. Malacostracan crustaceans showed a decline from a peak in 2011 to a minimum in 2019, followed by a slight recovery in 2022. This pattern was largely driven by amphipods, with the 2022 increase attributed to a high number of tanaidaceans. Echinoderms were most abundant in 2014, with abundances three times higher than in 2011 and 2019, and sharply declined in 2022. Their distribution is typically associated with areas under strong marine influence. Bivalves were abundant in 2011 and 2014 but their numbers halved in 2019 and 2022. Gastropods increased in 2014 and 2019 but dropped significantly in 2022. Polychaetes declined in 2014 and collapsed in 2022, with sedentary species being the most represented throughout the study period.
Ferrarin et al. (2012), considering a long period with several hydromorphological changes and related variations in Venice Lagoon, hypothesized a scenario for the benthic community predicting a progressive decrease in the number of species. This scenario included a general increase in polychaetes and a reduction in bivalves, attributed to the hydrological impacts of human interventions, which were expected to result in conditions more similar to the inner areas of the lagoon. Our findings, although covering a shorter period and fewer changes than those simulated by Ferrarin et al. (2012), confirm the reduction in Bivalvia, as well as in Crustacea and Polychaeta.
In the last two sampling campaigns, eight species were recorded that were not listed in the checklist of macrozoobenthic species of the Adriatic and Mediterranean Seas of the Italian Society of Marine Biology (SIBM). Of these, six belong to the Crustacea group and are relatively small and morphologically similar, which increases the likelihood of misidentification (Del Pasqua et al., 2022). For some of the recently detected taxa, such as certain Tanaid species, taxonomic uncertainty remains due to their cryptogenic nature, and ongoing studies are focusing on their genetic characterization and distribution (Faasse, 2013; Del Pasqua et al., 2022).
It is important to note that the Venice Lagoon is widely recognized as a hotspot for non-indigenous species (NIS) introductions in the Mediterranean Sea, primarily due to intense anthropogenic pressures such as maritime traffic, aquaculture, and canalization (Occhipinti-Ambrogi, 2000; Occhipinti-Ambrogi and Savini, 2003; Marchini et al., 2015a, b). This context underscores the importance of continuous monitoring and taxonomic refinement to detect and track emerging NIS and assess their potential ecological impacts.
Biological invasions are among the greatest threats to native biodiversity, and recently, Venice Lagoon has been identified as an ideal habitat for the spread of Mnemiopsis leidyi A. Agassiz, 1865 (Piccardi et al., 2025). This species is an invasive zooplanktivorous ctenophore that preys on various organisms, affecting the biodiversity of zooplankton (Schroeder et al., 2023) and impacting fishing activities (Piccardi et al., 2025). The abundance of the comb jelly has significantly increased since 2014 (Piccardi et al., 2025), making it plausible that the explosion of this species has also had direct consequences on the biodiversity of the macrozoobenthos in Venice Lagoon. Indeed, most of the taxa that disappeared in our findings after 2014, mostly crustaceans, gastropods, and bivalves, have zooplanktonic larval stages.
Regarding another well-known alien species recently invading the Northern Adriatic lagoons, Callinectes sapidus, its proliferation began in 2023 and primarily impacted the Po Delta River lagoons, where clam farming is the predominant activity (Chiesa et al., 2025). The present study reports data up to 2022. However, it is crucial to remain vigilant for potential consequences of this voracious species, which has caused severe impacts on native ecological communities (Onofri et al., 2008; Marchessaux et al., 2023). Indeed, it prefers benthic infauna, particularly bivalve mollusks, but it also consumes polychaetes, amphipods, and other benthic prey (Mansour and Lipcius, 1991; Mansour, 1992).
The ecological quality of the macrozoobenthos in the Venice Lagoon, as indicated by the M-AMBI index, showed variable classifications across space and time, predominantly reflecting a Moderate status according to the WFD definition. Across the entire lagoon, considering the average values of all natural water bodies, the ratio of the M-AMBI index parameters (Shannon’s diversity index, number of species and AMBI) indicated that, in the last monitoring cycle, species richness and diversity reached their lowest values, confirming a general decline in the biodiversity of the macrozoobenthos in Venice Lagoon. The M-AMBI seems to behave predictably in the presence or absence of pressure, in both coastal and transitional water bodies, such as different hydro-morphological pressures, including dredging or canalization, with high AMBI values and low diversity and richness levels especially in presence of organic matter, nutrient and pollutant discharges, which can result ultimately in oxygen depletion, eutrophication, etc (Borja et al., 2008). Venice lagoon is a dynamic and unique ecosystem, experiencing several anthropogenic activities which are modifying its hydrodynamic regimes (Ferrarin et al., 2012), as well as causing hypoxia or anoxia events (Brigolin et al., 2021), or pollutant and nutrient discharges (Bellucci et al., 2002; Maggi et al., 2012; Guolo et al., 2025). However, unlike other studies that analysed M-AMBI or AMBI responses to adverse impacts (Muxika et al., 2005; Borja et al., 2008), in Venice Lagoon diversity and richness declined, whilst AMBI improved with values dropping from 2.2-2.4 in the first surveys to 1.8 in 2022. These ranges of AMBI values are reported to indicate a slightly disturbed benthic community in Good status (Muxika et al., 2005; Borja et al., 2008).
To evaluate changes in environmental conditions in Venice Lagoon, trends in water parameters and assessments of sediment characteristics were performed. The results indicated significant decreasing temporal trends from 2011 to 2022 for dissolved oxygen, dissolved organic carbon (DOC), ammonium (NH4), nitrite (NO2), total dissolved nitrogen (TDN), and total dissolved phosphorus (TDP). Conversely, a significant increasing trend was observed for salinity. The nutrient reduction is consistent with Acri et al. (2020) who observed similar trends from 1998 to 2017 in Venice Lagoon. Nutrient inputs in Venice Lagoon are typically associated with riverine inflows (Zirino et al., 2016) and the trophic conditions of this environment have been generally improving in the recent years (Acri et al., 2020; Ponis et al., 2024). Therefore, higher salinity can be linked to greater marine influence, as evidenced by the rising average sea level in the Venice Lagoon (Crosato, 2024), one of the impacts of climate change. Indeed, some predictive studies for 2050 indicated that the decrease in river runoff, coupled with increased evaporation, will lead to higher salinity and temperature in the Adriatic basin, even though site-specific phenomena may cause deviations from this trend (Verri et al., 2024; da Costa et al., 2024).
Rising temperature is one of the primary indicators of global warming, and in the Venice Lagoon, an increase in water temperature was reported from 1998 to 2017 due to climate trends (Acri et al., 2020). The trend test of the present studydid not yield significant results, but data were obtained from quarterly campaigns from 2011 to 2022. Although the sampling frequency of water parameters is designed to provide the necessary information for water quality management according to the WFD, an analysis of data from continuous monitoring and longer time series could corroborate the results of the trend test of the present study, as well as better meet climate change assessment objectives.
In addition, recent studies and institutional reports have highlighted the occurrence of extreme climatic events that may have influenced the ecological dynamics of the lagoon. For instance, data from the National Climate Change Adaptation Platform (ISPRA, 2023) indicates that all lagoons in the Northern Adriatic Sea, including the Venice Lagoon, have experienced a general increase in both temperature and salinity over the past decade. Specifically, during the period 2011–2023, polyhaline and euhaline water bodies in the Venice Lagoon exhibited a positive trend in temperature, albeit not statistically significant, and a statistically significant increase in salinity. These findings are consistent with broader regional climate trends. As reported above, according to ARPAV (2023), the year 2022 was the warmest on record in the Veneto region, with an average air temperature anomaly of +1.2 °C compared to the 1993–2021 reference period. Notably, the average of daily maximum and minimum temperatures reached +1.2 °C and +3.2 °C, respectively. In addition to recurrent heatwaves, 2022 was characterized by exceptionally low precipitation, leading to widespread and prolonged drought conditions. This extreme event prompted the declaration of a national state of emergency by the Italian government. As reported by ISPRA (2023), the resulting hydrological stress contributed to the highest annual average salinity values recorded in the Venice Lagoon over the past decade.
These anomalies, although not directly captured by the quarterly sampling design of this study, represent important indirect signals of climate change. The limited temporal resolution of biological and physico-chemical data may hinder the detection of short-term ecological responses to such events, including shifts in species composition or community structure. Therefore, integrating high-frequency environmental monitoring with biological observations could enhance the capacity to detect and attribute climate-driven changes in transitional ecosystems such as the Venice Lagoon.
Concerning sediments, the analytical panels varied over time, hindering a comprehensive temporal analysis. Nevertheless, it can be observed that the sediment matrix remained largely conservative over time, apart from organic carbon, which nearly doubled from 2011 to 2022, and organic phosphorus, which increased almost fourfold from 2011 to 2019. However, PCA results confirmed that choked areas were characterized by higher values of pelite and organic carbon compared to areas more influenced by marine waters and stronger hydrodynamics.
Analysing the biodiversity of the macrozoobenthos in relation to water parameters, revealed a significant correlation with salinity, silicates, organic carbon (DOC and POC), and total dissolved phosphorus (TDP). Salinity emerged as a key factor influencing the structure of the benthic assemblage in the studied lagoon. The observed variations in community composition and organism performance are consistent with previous findings that highlight the sensitivity of benthic taxa to salinity fluctuations, particularly in transitional waters (Gibson, 1982; Muresan et al., 2019; Semprucci et al., 2019; Boutoumit et al., 2021). In our case, increased salinity levels were associated with reduced species richness and shifts in dominant taxa, suggesting a stress response likely linked to osmotic regulation challenges. These results align with documented impacts of salinity on survival, growth, and metabolic activity in aquatic organisms (Folt and Chen, 1999; Velasco et al., 2019), supporting the interpretation that salinity acts as a structuring environmental variable in this ecosystem.
Spatial and temporal changes in food sources can also affect the biodiversity of macrozoobenthos, as the animals within this community exhibit diverse feeding habits (Doi et al., 2005; Park et al., 2017). These variations could explain the significant relationships between macrozoobenthos and both organic carbon and total dissolved phosphorus (TDP) highlighted in this study. Additionally, silicates are equally important, as their concentrations can reflect environmental variations that may impact the distribution of macrozoobenthos (Ittekkot et al, 2000; Faresi et al., 2012).
Regarding sediments, the parameters found significant in statistical analyses (granulometry, organic content and nutrients) are among those that typically influence the distribution of taxa in the macrozoobenthic community (Pearson and Rosenberg, 1978; Dauer et al., 2000; Sharani et al., 2018). Moreover, lower levels of diversity were significantly associated with higher organic carbon contents, especially in the last three surveys. The richness of benthic communities in transitional waters typically declines as the organic content in the sediments increases (Magni et al., 2009). The accumulation of organic carbon in sediments is typical of eutrophicated systems, which are likely to support a limited number of taxa (Pusceddu et al., 2007). Although trophic trends have improved recently, eutrophic conditions persist in the inner parts of the Venice Lagoon (Ponis et al., 2024). Therefore, it can still be considered as one of the causes of the lower macrozoobenthos biodiversity as observed in the present study, but particularly in the choked water bodies influenced by freshwater.
Lastly, it is well known that the Venice Lagoon has various point and diffuse sources of pollutants and contaminants, including industrial discharges from Porto Marghera, sewage outfalls from the cities of Venice and Chioggia, harbor activities, agricultural and urban runoff, and atmospheric deposition (Bellucci et al., 2002; Frignani et al., 2003; Maggi et al., 2012; Feltracco et al., 2022; Pizzini et al., 2024; Guolo et al., 2025). These activities contribute to a mixture of contaminants that can directly affect the macrozoobenthos community by exposing organisms to contaminated sediments. However, continuous and spatially extended monitoring of emerging and re-emerging contaminants could help to better assess the spatial and temporal dynamics of macrozoobenthos revealed in the present study.
Conclusions
The analysis of the macrozoobenthic community in the Venice Lagoon from 2011 to 2022 indicates a significant temporal decline in both taxa richness and abundance, with 2022 marking the lowest biodiversity levels recorded across the entire lagoon. This decline is consistent across different lagoonal typologies, with polyhaline and choked water bodies showing lower species richness compared to euhaline and non-choked areas.
Significant correlations were found between macrozoobenthic biodiversity and environmental variables such as salinity, organic carbon, silicates, and total dissolved phosphorus. Most of these parameters, except salinity, showed a decreasing trend from 2011 to 2022. The sediment matrix remains largely conservative, with granulometry, organic content, and nutrients acting as key factors influencing taxa distribution especially in spatial rather than temporal dimensions.
These ecological patterns coincide with broader environmental changes observed in the region. Recent institutional reports (ISPRA, 2023; ARPAV, 2023) highlight a decade-long trend of increasing temperature and salinity in the Northern Adriatic lagoons, including Venice. The year 2022 was characterized by extreme climatic conditions—record-breaking temperatures, recurrent heatwaves, and severe drought—which led to unprecedented salinity levels in the lagoon. These combined thermal and hydrological stressors likely contributed to the sharp ecological shifts observed in benthic communities.
Although the quarterly sampling design of water parameters limits the detection of short-term ecological responses, these anomalies represent important indirect signals of climate change. Integrating high-frequency environmental monitoring with biological observations could significantly enhance the capacity to detect and attribute climate-driven changes in transitional ecosystems such as the Venice Lagoon.
Finally, although the current monitoring strategy was not specifically designed to detect non-indigenous species, a comparison with the most recent national checklist of macrozoobenthic species for the Adriatic Sea revealed the presence of some taxa not previously recorded in the area. Most of these were small crustaceans, often difficult to identify morphologically, and are currently considered cryptogenic species whose origin remains uncertain due to insufficient taxonomic or biogeographic information. Their detection underscores the importance of integrating traditional morphological approaches with molecular tools to improve the resolution of biodiversity assessments, particularly in regions like the Venice Lagoon, which is recognized as a hotspot for species introductions.
The results of this study carry important implications for environmental policy and management. The ecological shifts observed, especially those recorded in 2022, underscore the vulnerability of transitional ecosystems to extreme climatic events. These findings suggest the need for adaptive management strategies that incorporate climate resilience into lagoon governance.
The study also revealed the need to complement routine monitoring programs, primarily designed for ecological status assessment under the Water Framework Directive, with targeted strategies aimed at early detection and tracking of non-indigenous species. This is especially relevant in the context of accelerating environmental change, where shifts in species composition may occur rapidly and involve cryptic or newly introduced taxa.The long-term and spatially extensive dataset collected in this study represents a valuable resource for environmental policy. Its integration of biological and physico-chemical data over more than a decade provides a solid basis for evaluating water quality trends and anticipating climate-driven ecological changes, supporting more informed and adaptive management strategies.
In summary this study reveals a concerning decline in biodiversity and underscore the importance of continuous monitoring and targeted management strategies to mitigate further ecological degradation in the Venice Lagoon.
Data availability statement
Publicly available datasets were analyzed in this study. This data can be found here: https://www.arpa.veneto.it/dati-ambientali/open-data/idrosfera/acque-di-transizione/acque-di-transizione-laguna-di-venezia-monitoraggio-ecologico.
Ethics statement
The manuscript presents research on animals that do not require ethical approval for their study.
Author contributions
FC: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Project administration, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. MC: Conceptualization, Data curation, Formal analysis, Investigation, Validation, Visualization, Writing – review & editing. VB: Data curation, Formal analysis, Investigation, Software, Visualization, Writing – review & editing. FO: Data curation, Formal analysis, Visualization, Writing – review & editing. MN: Data curation, Project administration, Validation, Writing – review & editing. AG: Data curation, Investigation, Resources, Writing – review & editing. MZ: Data curation, Investigation, Writing – review & editing. DB: Data curation, Investigation, Validation, Writing – review & editing. FS: Funding acquisition, Project administration, Supervision, Writing – review & editing. AB: Funding acquisition, Investigation, Project administration, Validation, Writing – review & editing. RBB: Conceptualization, Funding acquisition, Investigation, Project administration, Resources, Supervision, Validation, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by ARPAV in the framework of the Mo.V.Eco. V Project, funded by Veneto Region, according to WFD, under the contract between ISPRA and ARPAV of 10/11/2023.
Acknowledgments
The authors of ISPRA would like to thank Dr. Michela Del Pasqua for sharing her valuable experience and useful advice.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that Generative AI was used in the creation of this manuscript. To improve and check academic english language of the text.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2025.1603523/full#supplementary-material
Supplementary Table 1 | Faunal lists of macrozoobenthic species recorded during each sampling campaign (2011, 2014, 2019 and 2022). Species names highlighted in blue indicate taxa not included in the most recent Checklist of Macrozoobenthic Species of the Adriatic and Mediterranean Seas published by the Italian Society of Marine Biology (SIBM).
References
Acri F., Braga F., and Bernardi Aubry F. (2020). Long-term dynamics in nutrients, chlorophyll a and water quality parameters in the Lagoon of Venice. Scientia Marina 84, 1–11. doi: 10.3989/scimar.05022.30
Alademomi S. A., Manning A. J., Abbott V. J., and Whitehouse R. J. S. (2020). Morphodynamics in a Tropical Shallow Lagoon: Observation and Inferences of Change. Lagoon Environments Around the World - A Scientific Perspective. IntechOpen. doi: 10.5772/intechopen.90189
Anthony A., Atwood J., August P., Byron C., Cobb S., Foster C., et al. (2009). Coastal lagoons and climate change: ecological and social ramifications in US Atlantic and Gulf coast ecosystems. Ecol. Soc. 14, 8. doi: 10.5751/ES-02719-140108
APAT-SIBM-ICRAM (2003). Manuale di metodologie di campionamento e studio del benthos marino mediterraneo. Eds. Gambi M. C. and Dappiano. Società italiana di biologia marina.
ARPAV (2023). Available online at: https://www.arpa.veneto.it/arpavinforma/indicatoriambientali/indicatori_ambientali/clima-e-rischi-naturali/clima/precipitazione-annua/2022 (Accessed 16-May-2025).
Bangun A. P., Wahyuningsih H., and Muhtadi A. (2018). Impacts of macro- and microplastic on macrozoobenthos abundance in intertidal zone. IOP Conf. Ser.: Earth Environ. Sci. 122, 12102. doi: 10.1088/1755-1315/122/1/012102
Barausse A., Grechi L., Martinello N., Musner T., Smania D., Zangaglia A., et al. (2015). An integrated approach to prevent the erosion of salt marshes in the lagoon of Venice. EQA-International J. Environ. Qual. 18, 43–54. doi: 10.6092/issn.2281-4485/5799
Bellucci L. G., Frignani M., Paolucci D., and Ravanelli M. (2002). Distribution of heavy metals in sediments of the Venice Lagoon: the role of the industrial area. Sci. Total Environ. 295, 35–49. doi: 10.1016/s0048-9697(02)00040-2
Borja A., Franco J., and Pérez V. (2000). A marine biotic index to establish the ecological quality of soft-bottom benthos within European estuarine and Coastal environments. Mar. pollut. Bull. 40, 1100–1114. doi: 10.1016/S0025-326X(00)00061-8
Borja A., Franco F., Valencia V., Bald J., Muxika I., Belzunce M. J., et al. (2004). Implementation of theEuropean Water Framework Directive from the Basque country (northern Spain): a methodological approach. Mar. pollut. Bull. 48, 209–2018. doi: 10.1016/j.marpolbul.2003.12.001
Borja A., Muxika I., and Rodriguez J. G. (2008). Paradigmatic responses of marine benthic communities to different anthropogenic pressures, using M-AMBI, within the European Water Framework Directive Mar. Ecol. 30, 214–227. doi: 10.1111/j.1439-0485.2008.00272.x
Boutoumit S., Bououarour O., El Kamcha R., Pouzet P., Zourarah B., Benhoussa A., et al. (2021). Spatial patterns of macrozoobenthos assemblages in a sentinel coastal lagoon: biodiversity and environmental drivers. J. Mar. Sci. Eng. 9, 461. doi: 10.3390/jmse9050461
Brigolin D., Rabouille C., Demasy C., Bombled B., Monvoisin G., and Pastres R. (2021). Early Diagenesis in Sediments of the Venice Lagoon (Italy) and its relationship to hypoxia. Front. Mar. Sci. 7. doi: 10.3389/fmars.2020.575547
Cacciatore F., Bonometto A., Paganini E., Sfriso A., Novello M., Parati P., et al. (2019). Balance between the reliability of classification and sampling effort: A multi-approach for the water framework directive (WFD) ecological status applied to the venice lagoon (Italy). Water 11, 1572. doi: 10.3390/w11081572
Chiesa S., Petochi T., Boscolo Brusà R., Raicevich S., Cacciatore F., Franceschini G., et al. (2025). Impacts of the blue crab invasion on Manila clam aquaculture in Po Delta coastal lagoons (Northern Adriatic Sea, Italy), Estuarine. Coast. Shelf Sci. 312, 109037. doi: 10.1016/j.ecss.2024.109037
Crosato F. (2024). Livello medio del mare a Venezia. Dati e indicatori della Piattaforma Nazionale Adattamento Cambiamenti Climatici. Available on: Piattaforma Nazionale Adattamento Cambiamenti Climatici, https://climadat.isprambiente.it/dati-e-indicatori/indicatori-di-impatto-dei-cambiamenti-climatici/8981-2 (Accessed March 19, 2025).
D’Alessandro M., Esposito V., Porporato E. M. D., Berto D., Renzi M., Giacobbe S., et al. (2018). Relationships between plastic litter and chemical pollutants on benthic biodiversity. Environ. pollut. 242, 1546–1556. doi: 10.1016/j.envpol.2018.08.002
da Costa V. S., Alessandri J., Verri G., Mentachi L., Guerra R., and Pinardi N. (2024). Marine climate indicators in the Adriatic Sea. Front. Clim. 6. doi: 10.3389/fclim.2024.1449633
Dapeng N., Zhinan Z., and Xiaoshou L. (2019). Benthic ecological quality assessment of the Bohai Sea, China using marine biotic indices. Mar. pollut. Bull. 142, 457–464. doi: 10.1016/j.marpolbul.2019.03.055
Dauer D. M., Ranasinghe J. A., and Weisberg S. B. (2000). Relationships between benthic community condition, water quality, sediment quality, nutrient loads, and land use patterns in chesapeake bay. Estuaries 23, 80–96. doi: 10.2307/1353227
Del Pasqua M., Bernarello V., Esquete P., Cornello M., Cacciatore F., Oselladore F., et al. (2022). First Records of the Tanaid Species Zeuxo holdichi and Apseudopsis tridens (Crustacea: Peracarida) From the Venice Lagoon (Italy, Northern Adriatic Sea). Thalassas 38, 417–430. doi: 10.1234/jmb.2025.12345
Doi H., Matsumasa M., Toya T., Satoh N., Mizota C., Maki Y., et al. (2005). Spatial shifts in food sources for macrozoobenthos in an estuarine ecosystem: carbon and nitrogen stable isotope analyses. Estuar. Coast. Shelf Sci. 64, 316–322. doi: 10.1016/j.ecss.2005.02.028
European Environment Agency European Topic Centre on Biological Diversity—Report under the Article 17 of the Habitats Directive Period 2013–2018. Available online at: https://nature-art17.eionet.europa.eu/article17/ (Accessed 18 February 2025).
European Parliament and Council (2000). Directive 2000/60/EC of the European Parliament and of the Council establishing a framework for Community action in the field of water policy. Off. J. Eur. Communities 327, 1–73. Available at: http://eur-lex.europa.eu/LexUriServ/LexUriServ.do?uri=CELEX:32000L0060:en:NOT
Faasse M. (2013). Northward dispersal of the tanaid Zeuxo holdichi to The Netherlands (Crustacea: Tanaidae) 39, 95–98. Available at: https://natuurtijdschriften.nl/pub/1002120/NFM39_095-104_Faasse.pdf (Accessed July 07, 2025).
Facca C., Cavraro F., Franzoi P., and Malavasi S. (2020). Lagoon resident fish species of conservation interest according to the habitat directive (92/43/CEE): A review on their potential use as ecological indicator species. Water 12, 2059. doi: 10.3390/w12072059
Faresi L., Bettoso N., and Aleffi I. F. (2012). Vertical distribution of soft bottom macrozoobenthos in the gulf of trieste (Northern adriatic sea). Annales Ser. Hist. Naturalis 22, 123–135. Available at: https://zdjp.si/wp-content/uploads/2015/08/faresi-bettoso-aleffi.pdf (Accessed March 19, 2025).
Feltracco M., Barbaro E., Morabito E., Zangrando R., Piazza R., Barbante C., et al. (2022). Assessing glyphosate in water, marine particulate matter, and sediments in the Lagoon of Venice. Environ. Sci. pollut. Res. 29, 16383–16391. doi: 10.1007/s11356-021-16957-x
Feola A., Bonometto A., Canesso D., Pedroncini A., Cacciatore F., Novello M., et al. (2024). A method to quantify the drainage basin contributions to transitional water bodies: numerical modeling applied to the case study of venice lagoon. Environments 11, 234. doi: 10.3390/environments11110234
Ferrarin C., Bajo M., Bellafiore D., Cucco A., De Pascalis F., Ghezzo M., et al. (2014). Toward homogenization of Mediterranean lagoons and their loss of hydrodiversity. Geophys. Res. Lett. 41, 5935–5941. doi: 10.1002/2014GL060843
Ferrarin C., Ghezzo M., Umgiesser G., Tagliapietra D., and Camatti E. (2012). Assessing hydrological effects of human interventions on coastal systems: numerical applications to the Venice Lagoon. Hydrol. Earth Syst. Sci. Discuss 9, 13839–13878. doi: 10.5194/hessd-9-13839-2012
Folt C. and Chen C. (1999). Synergism and antagonism among multiple stressors. Limnol. Oceanogr. 44, 864–877. doi: 10.4319/lo.1999.44.3
Frignani M., Bellucci L. G., Fagotto M., and Albertazzi S. (2003). Polycyclic aromatic hydrocarbons in sediments of the Venice Lagoon. Hydrobiologia. 494, 283–290. doi: 10.1023/A:1025451307225
Gibson R. N. (1982). Recent studies on the biology of intertidal fishes. Oceanogr Mar. Biol. Annu. Rev. 20, 363–414.
Guerzoni S. and Tagliapietra D. (2006). Atlas of the lagoon: Venice between land and sea, (Venezia, IT: Marsilio Editori), 242.
Guolo P. P., Calgaro L., Bonetto A., Xu Q., Lu J., and Marcomini A. (2025). Investigating contaminants of emerging concern (CECs) in the venice lagoon: A suspect screening approach for the analysis of water and sediment quality. doi: 10.2139/ssrn.5152600
Hirsch R. M., Slack J. R., and Smith R. A. (1982). Techniques of trend analysis for monthly water quality data. Water Resour. Res. 18, 107–121. doi: 10.1029/WR018i001p00107
ISPRA (2010). Protocolli per il Campionamento e la Determinazione Degli Elementi di Qualità Biologica e Fisico-Chimica Nell’ambito dei Programmi di Monitoraggio ex 2000/60/CE delle Acque di Transizione; El-Pr-TW-Protocolli Monitoraggio-03.06 (Roma, Italy: ISPRA).
ISPRA (2023). National climate change adaptation platform. Available online at: https://climadat.isprambiente.it/dati-e-indicatori/indicatori-di-impatto-dei-cambiamenti-climatici/ (Accessed 16-May-2025).
Ittekkot V., Humborg C., and Schäfer P. (2000). Hydrological Alterations and Marine Biogeochemistry: A Silicate Issue? Silicate retention in reservoirs behind dams affects ecosystem structure in coastal seas. BioScience 50, 776–782. doi: 10.1641/0006-3568(2000)050[0776:HAAMBA]2.0.CO;2
Kneib R. T.. (1984). Patterns of invertebrate distribution and abundance in the intertidal salt marsh: causes and questions. Estuaries 7, 392–412.
Knutsen Ø., Stefanakos C., Slagstad D., Ellingsen I., Zacharias I., Biliani I., et al. (2024). Studying the evolution of hypoxia/anoxia in Aitoliko lagoon, Greece, based on measured and modeled data. Front. Mar. Sci. 11. doi: 10.3389/fmars.2024.1299202
Lacoste É., Jones A., Callier M., Klein J., Lagarde F., Derolez V., et al. (2023). A review of knowledge on the impacts of multiple anthropogenic pressures on the soft-bottom benthic ecosystem in mediterranean coastal lagoons. Estuaries Coasts 46, 2190–2207. doi: 10.1007/s12237-023-01188-9
Li B., Li X., Bouma T. J., Soissons L. M., Cozzoli F., Wang Q., et al. (2017). Analysis of macrobenthic assemblages and ecological health of Yellow River Delta, China, using AMBI & M-AMBI assessment method. Mar. pollut. Bull. 119, 23–32. doi: 10.1016/j.marpolbul.2017.03.044
López-López E. and Sedeño-Díaz J. E. (2015). “Biological indicators of water quality: the role of fish and macroinvertebrates as indicators of water quality,” in Environmental indicators, vol. Chapter 37 . Eds. Armon R. H. and Hänninen O. Dordrecht: Springer pp. 643–661. doi: 10.1007/978-94-017-9499-2_37
Maggi C., Ausili A., Boscolo Brusà R., Cacciatore F., Bonometto A., Cornello M., et al. (2012). Sediment and biota in trend monitoring of contaminants in transitional waters. Trends Analytical Chem. 36, 82–61. doi: 10.1016/j.trac.2012.01.006
Magni P., Tagliapietra D., Lardicci C., Balthis L., Castelli A., Como S., et al. (2009). Animal-sediment relationships: evaluating the ‘Pearson-Rosenberg paradigm’ in Mediterranean coastal lagoons. Mar. pollut. Bull. 58, 478–486. doi: 10.1016/j.marpolbul.2008.12.009
Mansour R. A. (1992). Foraging ecology of the blue crab, Callinectes sapidus Rathbun, in lower Chesapeake Bay [dissertation] (Gloucester Point (VA: The College of William and Mary, Virginia Institute of Marine Science).
Mansour R. A. and Lipcius R. N. (1991). Density-dependent foraging and mutual interference in blue crabs preying upon infaunal clams. Mar. Ecol. Prog. Ser. 72, 239–246. doi: 10.3354/meps072239
Marchessaux G., Mangano M. C., Bizzarri S., M’Rabet C., Principato E., Lago N., et al. (2023). Invasive blue crabs and small-scale fisheries in the Mediterranean sea: Local ecological knowledge, impacts and future management. Mar. Policy 148, 105461. doi: 10.1016/j.marpol.2022.105461
Marchini A., Ferrario J., Sfriso A., and Occhipinti-Ambrogi A. (2015a). Current status and trends of biological invasions in the Lagoon of Venice, a hotspot of marine NIS introductions in the Mediterranean Sea. Biol. Invasions 17, 2943–2962. doi: 10.1007/s10530-015-0922-3
Marchini A., Galil B. S., and Occhipinti-Ambrogi A. (2015b). Recommendations on standardizing lists of marine alien species: Lessons from the Mediterranean Sea. Mar. pollut. Bull. 101, 267–273. doi: 10.1016/j.marpolbul.2015.09.054
Marzano C. N., Liaci L. S., Fianchini A., Gravina F., Mercurio M., and Corriero G. (2003). Distribution persistence and change in the macrobenthos of the lagoon of Lesina (Apulia, southern Adriatic Sea). Oceanol. Acta 26, 57–66. doi: 10.1016/S0399-1784(02)01229-X
Meals D. W., Spooner J., Dressing S. A., and Harcum J. B. (2011). Statistical analysis for monotonic trends. Tech Notes 6, November 2011 (Fairfax, VA: Developed for U.S. Environmental Protection Agency by Tetra Tech, Inc), 23.
Mistri M., Fano E. A., Rossi G., Caselli K., and Rossi R. (2000). Variability in macrobenthos communities in the valli di comacchio, northern Italy, a hypereutrophized lagoonal ecosystem. Estuarine Coast. Shelf Sci. 51, 599–611. doi: 10.1006/ecss.2000.0697
Molinaroli E., Guerzoni S., Sarretta A., Masiol M., and Pistolato M. (2009). Thirty-year changechanges, (1970 to 2000) in bathymetry and sediment texture recorded in the Lagoon of Venice sub-basins, Italy. Mar. Geology 258, 115–125. doi: 10.1016/j.margeo.2008.12.001
Mosbahi N., Pezy J.-P., Dauvin J.-C., and Neifar L. (2016). Spatial and temporal structures of the macrozoobenthos from the intertidal zone of the kneiss islands (Central mediterranean sea). Open J. Marine. Sci. 6, 223–237. doi: 10.4236/ojms.2016.62018
Munari C., Como S., Keppel E., Lardici C., Magni P., Mistri M., et al. (2010). Towards the management of transitional waters: assessing the potential of benthic tools. Biol. Marina Mediterranea 17 pp, 51–54.
Muresan A. N., Gaglio M., Aschonitis V., Nobili G., Castaldelli G., and Fano E. A. (2019). Structural and functional responses of macroinvertebrate communities in small wetlands of the Po delta with different and variable salinity levels. Estuarine Coast. Shelf Sci. 238, 106726. doi: 10.1016/j.ecss.2020.106726
Muxika I., Borja A., and Bald J. (2007). Using historical data, expert judgement and multivariate analysis in assessing reference conditions and benthic ecological status, according to the European Water Framework Directive. Mar. pollut. Bull. 55, 16–29. doi: 10.1016/j.marpolbul.2006.05.025
Muxika I., Borja A., and Bonne W. (2005). The suitability of the marine biotic index (AMBI) to new impact sources along European coasts. Ecol. Indic. 5, 19–31. doi: 10.1016/j.ecolind.2004.08.004
Narayana A. C. and Priju C. P. (2006). Evolution of the coastal landforms and environments of the central Kerala coast, India. J. Coast. Res. 22, 1237–1247.
Newton A., Icely J., Cristina S., Brito A., Cardoso A. C., Colijn F., et al. (2014). An overview of ecological status, vulnerability and future perspectives of European large shallow, semi-enclosed coastal systems, lagoons and transitional waters. Estuar. Coast. Shelf Sci. 140, 95–122. doi: 10.1016/j.ecss.2013.05.023p
Occhipinti-Ambrogi A. (2000). Biotic invasions in a mediterranean lagoon. Biol. Invasions 2, 165–176. doi: 10.1023/A:1010004926405
Occhipinti-Ambrogi A. and Savini D. (2003). Biological invasions as a component of global change in stressed marine ecosystems. Mar. pollut. Bull. 46, 542–551. doi: 10.1016/S0025-326X(02)00363-6
Onofri V., Dulčić J., Conides A., Matić-Skoko S., and Glamuzina B. (2008). The occurrence of the blue crab, Callinectes sapidus Rathbun 1896 (Decapoda, Brachyura, Portunidae) in the eastern Adriatic (Croatian coast). Crustaceana 81, 403–409. doi: 10.1163/156854008783797561
Oselladore F., Bernarello V., Cacciatore F., Cornello M., Boscolo Brusà R., Sfriso A., et al. (2022). Changes in macrozoobenthos community after aquatic plant restoration in the northern venice lagoon (IT). Int. J. Environ. Res. Public Health 19, 4838. doi: 10.3390/ijerph19084838
Park H. J., Kang H. Y., Park T. H., and Kang C.-K. (2017). Comparative trophic structures of macrobenthic food web in two macrotidal wetlands with and without a dike on the temperate coast of Korea as revealed by stable isotopes. Mar. Environ. Res. 131, 134–145. doi: 10.1016/j.marenvres.2017.09.018
Pearson T. H. and Rosenberg R. (1978). Macrobenthic succession in relation to organic enrichment and pollution of the marine environment. Oceanography Mar. Biology: Annu. Rev. 16, 229–311.
Pérez-Ruzafa A., Marcos C., and Pérez-Ruzafa I. M. (2011). Mediterranean coastal lagoons in an ecosystem and aquatic resources management context. Phys. Chem. Earth Parts A/B/C 36, 160–166. doi: 10.1016/j.pce.2010.04.013
Piccardi F., Poli F., Sguotti C., Tirelli V., Borme D., Mazzoldi C., et al. (2025). Assessing the impact of the invasive ctenophore Mnemiopsis leidyi on artisanal fisheries in the Venice Lagoon: an interdisciplinary approach. Hydrobiologia 852, 2387–2405. doi: 10.1007/s10750-024-05505-6
Pielou (1966). The measurement of diversity in different types of biological collections. J. Theor. Biol. 13, 131–144. doi: 10.1016/0022-5193(66)90013-0
Pizzini S., Giubilato E., Morabito E., Barbaro E., Bonetto A., Calgaro L., et al. (2024). Contaminants of emerging concern in water and sediment of the Venice Lagoon, Italy. Environ. Res. 249, 118401. doi: 10.1016/j.envres.2024.118401
Ponis E., Cacciatore F., Bernarello V., Boscolo Brusà R., Novello M., Sfriso A., et al. (2024). Assessment of the trophic status and trend using the transitional water eutrophication assessment method: A case study from venice lagoon. Environments 11, 251. doi: 10.3390/environments11110251
Ponti M., Vadrucci M. R., Orfanidis S., and Pinna M. (2009). Biotic indices for ecological status of transitional water ecosystems. Transit. Waters Bull. 3, 32–90. doi: 10.1285/i1825229Xv3n3p32
Pusceddu A., Gambi C., Manini E., and Danovaro R. (2007). Trophic state, ecosystem efficiency and biodiversity of transitional aquatic ecosystems: analysis of environmental quality based on different benthic indicators. Chem. Ecol. 23, 505–515. doi: 10.1080/02757540701760494
Relini G. (2008). “Checklist della flora e della fauna dei mari italiani,” Parte I. Biologia Marina Mediterranea 15 (Suppl. 1) - 2008 (ITA/ENG). pp. 438.
Relini G. (2010). “Checklist della flora e della fauna dei mari italiani/Checklist of the flora and fauna in italian seas”. na dei mari italiani / Checklist of the flora and fauna in italian seas". Parte II. Biologia Marina Mediterranea 17 (Suppl. 1) - 2010 (ITA/ENG). pp 385.
Schroeder A., Camatti E., Pansera M., Pallavicini A., et al. (2023). Feeding pressure on meroplankton by the invasive ctenophore Mnemiopsis leidyi. Biol. Invasions 25, 2007–2021.
Semprucci F., Gravina M., and Magni P. (2019). Meiofaunal dynamics and heterogeneity along salinity and trophic gradients in a Mediterranean transitional system. Water 11, 1488. doi: 10.3390/w11071488
Sfriso A., Birkemeyer T., and Ghetti P. F. (2001). Benthic macrofauna changes in areas of Venice lagoon populated by seagrasses or seaweeds. Mar. Environ. Res. 52, 323–349. doi: 10.1016/S0141-1136(01)00089-7
Shannon C. E. and Weaver W. (1949). The mathematical theory of communication Vol. 117 (Urbana: The University of Illinois Press).
Sharani J., Putro S. P., and Hidayat J. W. (2018). Correlation between macrobenthic structure (biotic) and water-sediment characteristics (abiotic) adjacent aquaculture areas at Tembelas Island, Indonesia. J. Physics: Conf. Ser. 1025, 12081. doi: 10.1088/1742-6596/1025/1/012081
Solidoro C., Bandelj V., Bernardi Aubry F., Camatti E., Ciavatta S., Cossarini G., et al. (2010). “Response of Venice Lagoon ecosystem to natural and anthropogenic pressures over the last 50 years,” in Coastal lagoons: critical habitats of environmental change. Eds. Kennish M. J. and Paerl H. W. (CRC Press, Boca Raton, FL, USA), 483–511.
Sorokin P. Y., Sorokin Y. I., Zakuskina O. Y., and Ravagnan C. P. (2002). On the changing ecology of Venice lagoon. Hydrobiologia 487, 1–18. doi: 10.1023/A:1022939426396
Tagliapietra D., Sigovini M., and Volpi Ghirardini A. (2009). A review of terms and definitions to categorise estuaries, lagoons and associated environments. Mar. Freshw. Res. 60, 497–509. doi: 10.1071/MF08088
Velasco J., Gutierrez-Canovas C., Botella-Cruz M., Sanchez-Fernandez D., Arribas P., Carbonell J. A., et al. (2019). Effects of salinity changes on aquatic organisms in a multiple stressor context. Phil.Trans. R. Soc B 374, 20180011. doi: 10.1098/rstb.2018.0011
Verri G., Furnari L., Gunduz M., Senatore A., Santos da Costa V., De Lorenzis A., et al. (2024). Climate projections of the Adriatic Sea: role of river release. Front. Clim. 6. doi: 10.3389/fclim.2024.1368413
Whitfield A. K. and Elliott M. (2005). Fishes as indicators of environmental and ecological changes within estuaries: A review of progress and some suggestions for the future. J. Fish Biol. 61A, 229–250. doi: 10.1111/j.1095-8649.2002.tb01773.x
WORMS, World Register of Marine Species. (2025). Ahyong S., Boyko C. B., Bernot J., Brandão S. N., Daly M., De Grave S.. doi:10.14284/170 Available from https://www.marinespecies.org at VLIZ (Accessed July 08, 2025).
Keywords: biodiversity, climate change, Directive 2000/60/EC, salinity, nutrients, M-AMBI, sediments, transitional waters
Citation: Cacciatore F, Cornello M, Bernarello V, Oselladore F, Novello M, Girolimetto A, Zorzi M, Bon D, Strazzabosco F, Bonometto A and Boscolo Brusà R (2025) Macrozoobenthos and environmental changing in the Venice Lagoon (Italy). Front. Mar. Sci. 12:1603523. doi: 10.3389/fmars.2025.1603523
Received: 31 March 2025; Accepted: 27 June 2025;
Published: 28 July 2025.
Edited by:
Cristina Munari, University of Ferrara, ItalyCopyright © 2025 Cacciatore, Cornello, Bernarello, Oselladore, Novello, Girolimetto, Zorzi, Bon, Strazzabosco, Bonometto and Boscolo Brusà. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Federica Cacciatore, ZmVkZXJpY2EuY2FjY2lhdG9yZUBpc3ByYW1iaWVudGUuaXQ=