Abstract
Marine plankton play a crucial yet understudied economic role, contributing both positively through ecosystem services (ES) and negatively through ecosystem disservices (EDS). This study employs a mixed-methods approach to address key research gaps in plankton valuation and plankton value-based policy optimization. We (i) conduct a semi-systematic review of marine plankton ES and EDS and their economic assessments, and (ii) develop an expert assessment tool to evaluate the expected effects of marine policy on plankton ES and EDS and apply it to a hypothetical fully protected marine area. Our review reveals that while plankton ES are increasingly recognized and classified, their economic valuation remains scarce due to their indirect and diffuse economic impacts. In contrast, plankton EDS, though not previously comprehensively classified as such, have been more extensively quantified, as their economic costs are typically localized and more directly attributable. The expert assessment, conducted with 19 marine scientists (including eight plankton specialists), highlighted the potential of marine protection to enhance research opportunities, support plankton’s role in marine food webs and fish larvae recruitment, and mitigate harmful algal blooms (HABs). However, experts also noted significant uncertainties, particularly due to the high spatial variability and mobility of plankton. Our findings underscore the need for comprehensive assessments of the cumulative social benefits of regulating ES beyond carbon sequestration to better capture plankton’s economic significance. Additionally, we advocate for more spatio-temporally flexible conservation approaches to account for plankton dynamism in marine policy and management.
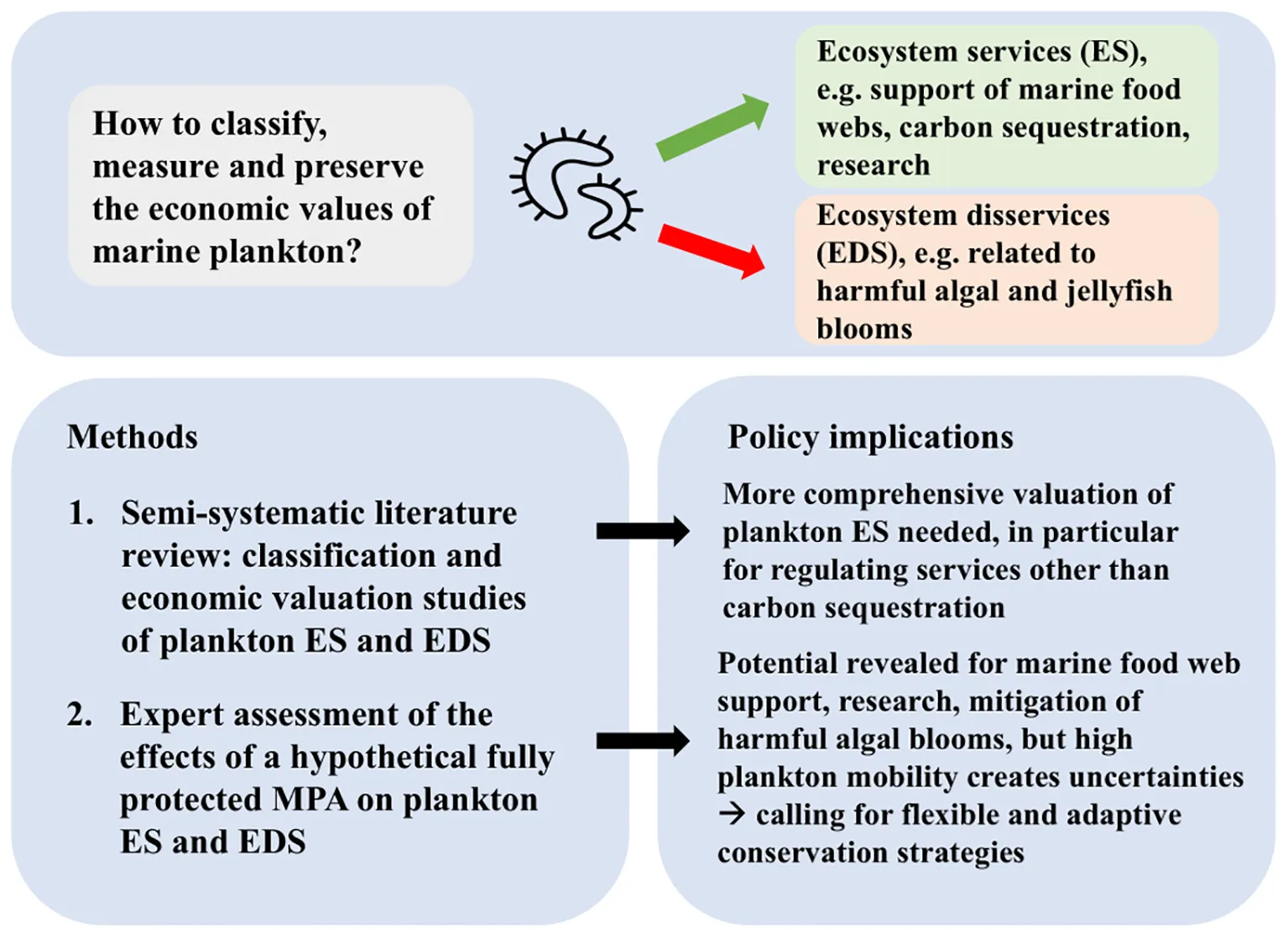
1 Introduction
Ecosystems create the necessary conditions for life on earth and provide a large number of benefits to humans, which for several decades have been conceptualized as Ecosystem Services (ES) to highlight their economic importance (Costanza and Daly, 1992; Costanza et al., 1997). On the other hand, some ecosystem activities have negative impacts on human wellbeing, and have therefore been referred to as Ecosystem Disservices (EDS) (Shackleton et al., 2016). Marine plankton, the innumerable organisms drifting in the ocean - including plants, animals, bacteria, viruses and fungi - are associated with both ecosystem services and disservices. These life forms play a crucial role in supporting marine food webs, regulating global carbon cycles, climate, and producing oxygen. However, they can also cause ecological disruptions, such as harmful algal blooms that deplete oxygen levels and release toxins, negatively impacting marine life and human activities.
The economic importance of marine plankton is increasingly being acknowledged, as shown by the recent surge in publications on plankton ecosystem services (B−Béres et al., 2023; Botterell et al., 2023; Calbet, 2024; Jean-Louis et al., 2025; Naselli-Flores and Padisák, 2023; Russo et al., 2025). The use of the ecosystem services classification underscores that, in addition to its vital supporting and regulating functions like primary production and carbon sequestration (Falkowski, 2012), plankton also offers provisioning services, such as food supplements, medical applications, and potential biofuel use (Naselli-Flores and Padisák, 2023), along with cultural services linked to its importance in scientific research and aesthetic values of certain species (Graham et al., 2014).
Planktonic activities are, however, not always beneficial for humans. Notably, blooms of toxin-releasing algae and jellyfish, often driven by anthropogenic stressors, exert negative impacts on health, economies and marine environments (Grattan et al., 2016; Landsberg, 2002; Nwankwegu et al., 2019; Sagarminaga et al., 2024). While a number of assessments of the economic damages caused by those planktonic activities exist (Carias et al., 2024; Kennerley et al., 2022; Kouakou and Poder, 2019; Osseni et al., 2021), they have not yet been classified in a comprehensive way as EDS. By contrast, plankton ecosystem services have recently been classified extensively following established guidelines (B−Béres et al., 2023; Botterell et al., 2023; Naselli-Flores and Padisák, 2023), but associations of beneficial planktonic ecosystem services with economic value indicators are comparatively scarce and the few existing studies focus on subsets of planktonic functions and value domains (Cavan et al., 2024; Jean-Louis et al., 2025).
The rise of ecosystem-based management entails a growing focus on ecosystem services for marine policy, including conservation (Luck et al., 2012; McDonald et al., 2020; Naselli-Flores and Padisák, 2023). Additionally, the recognition of the importance of marine plankton for human well-being has sparked proposals to prioritize plankton in marine policy-making (Tweddle et al., 2018). Yet, scientific investigation of how marine policies such as conservation measures impact plankton ecosystem services has not been undertaken. Open questions concern in particular whether and how marine policies can achieve synergies in addressing the provision of plankton ecosystem services and the mitigation of disservices, and whether, conversely, there are trade-offs to be considered (Schaafsma and Bartkowski, 2020).
Expert assessments have proven to be an insightful approach to evaluating policy outcomes on ES and EDS. A key advantage of these methods is that they enable the consideration of ES and EDS bundles deemed important, rather than focusing on a more easily quantifiable subset that may bias policy prioritization (Eastwood et al., 2016). Furthermore, they can provide rapid estimations for ES and EDS changes for which biophysical data approximation is complex and/or scarce (Pham et al., 2025), which is the case for plankton ES and EDS (Botterell et al., 2023). While expert assessments have been used for the evaluation of marine and coastal zone management regarding expected ES outcomes (Schernewski et al., 2018), they have not been used for the explicit assessment of plankton ES and EDS. In this article, we present an expert assessment tool for evaluating changes in plankton ES and EDS and demonstrate its application with 19 marine scientists to assess the expected impact of establishing a fully protected marine area in a European shelf sea.
Thus, building on the recognition of marine plankton’s significance for human well-being and the increasing use of ecosystem services in ecosystem-based management approaches to conservation, this article addresses a critical gap in current conservation strategies by providing (i) a semi-systematic review of marine planktonic ES and EDS and their respective economic measures, and (ii) an expert assessment of changes in ES and EDS provision assuming the implementation of a fully protected marine area according to IUCN guidelines (Day et al., 2019).
2 Methods
2.1 Classification and economic values of marine plankton ecosystem services and disservices
The Millennium Ecosystem Assessment defines ecosystem services in the following way: “Ecosystem services are the benefits people obtain from ecosystems. These include provisioning services such as food and water; regulating services such as flood and disease control; cultural services such as spiritual, recreational, and cultural benefits; and supporting services, such as nutrient cycling, that maintain the conditions for life on Earth” (MEA, 2005).
We base our classification of plankton ecosystem services on the MEA framework, primarily due to its simplicity, and its inclusion of supporting services, which are key components of the ecosystem services provided by plankton. Supporting services are typically treated as intermediates, contributing to a final service that can be valued economically (Morse-Jones et al., 2011). Regarding the services considered, we mainly rely on recently published reviews (B−Béres et al., 2023; Botterell et al., 2023; Naselli-Flores and Padisák, 2023), supplemented with literature from a review based on search terms listed in Supplementary Tables S1 and S2 of the Supplementary Material. We searched the Web of Science and Google Scholar for all possible combinations of plankton search terms and ecosystem service search terms, as well as plankton search terms and economic association search terms, and combinations of all three search term categories. Additionally, we considered relevant literature in the reference lists of the papers thus identified. For generalizability, we provide an alternative classification of ecosystem services following CICES (Common International Classification of Ecosystem Services) V5.1 (Haines-Young and Potschin-Young, 2018) in the Supplementary Material Table S3, which excludes the category of supporting services and instead considers them as underpinning provisioning services.
Ecosystem disservices have only recently been systematically categorized and incorporated into ecosystem service assessments (Campagne et al., 2018; Guo et al., 2022). Given that certain planktonic activities are associated with significant EDS that should be considered in marine policy, we deem it essential to include them in this review. There is not yet an agreed upon and standardized approach to ecosystem disservice classification (Anderson et al., 2025). Our classification follows the frameworks proposed by Shackleton et al. (2016) and Campagne et al. (2018).
The literature on plankton ES and EDS is highly heterogeneous and does not necessarily employ a common terminology, but is often focused on specific subgroups of plankton and specific ES. We therefore conducted a semi-systematic literature review (Botterell et al., 2023), combining different strategies. We selected search terms for plankton ecosystem services and disservices from previously published reviews and combined them with search terms for economic values (Supplementary Material S1 and S2). We included search terms for marine plankton and different subgroups such as jellyfish, copepods, diatoms and cyanobacteria that represent planktonic keystone groups. The literature search was conducted using Web of Science and Google Scholar for scientific literature, and Google for grey literature on economic values (see below).
The primary objective is to synthesize methods and outcomes of marine plankton valuation. This posed differentiated challenges to us for ES and EDS. Plankton ES valuation is almost completely absent from the literature, therefore we consulted both scientific publications as well as grey data and created associations between economic outcomes and plankton where possible, in a similar manner to Campagne et al. (2015)’s approach to the valuation of Posidonia oceanica ecosystem services valuation in the Mediterranean. Contrastingly, for some plankton EDS, valuation studies are quite abundant and have been subject to reviews. We do not give a full account of them here, but provide an overview of the different methods and outcomes and refer to the respective reviews for details.
Although the distinction is somewhat coarse (Russo et al., 2025), we distinguish between phyto- and zooplankton in this article. We make this choice because other recently published reviews employ these categories (Botterell et al., 2023; Naselli-Flores and Padisák, 2023), and because they are the most popular distinction used in policy documents, such as the European Union Marine Strategy Framework Directive.
2.2 Expert assessment of changes in ecosystem service and disservice provision assuming the implementation of a fully protected marine area
The current interest in marine plankton ES and EDS demonstrates their relevance for marine policy-making. Tweddle et al. (2018) argue that phytoplankton parameters should be explicitly considered within conservation objectives, due to their importance in supporting ecosystem functions and ES, but caution that a challenge to this approach is the difficulty in assessing phytoplankton ES such as primary production based on available data (which is improving with more advanced methods for including subsurface information to complement satellite data, but remains costly). Russo et al. (2025) underscore the strong connection between environmental conditions, plankton biodiversity and ES, however changes in plankton ES in response to changes in environmental conditions are very difficult to pinpoint because of the high number of factors that contribute to them in an interacting manner (Tagliabue et al., 2021).
The expert assessment aimed to evaluate the potential of area-based marine protection regarding the provision of plankton ES and EDS, thereby proposing an additional method for the literature branch dealing with the plankton ecosystem services case for marine protection (Jean-Louis et al., 2025). The method is useful to synthesize different experts’ knowledge and personal experiences into likely potentials for ES and EDS outcomes. We designed the methodology of the expert assessment based on previous studies of expert evaluation of ES/EDS potential of different ecosystems and/or policy options, notably Schernewski et al. (2018) and Campagne et al. (2018).
The research project BIOcean5D, in scope of which the work on this article was conducted, focuses on European marine life and unites many internationally renowned marine scientists and notably plankton experts. Therefore, we chose to create a marine protection scenario set in the BIOcean5D study region (Figure 1) and to be evaluated by BIOcean5D experts. Since the experts are based in different European countries and some work on different marine areas, while others do not conduct area-specific research, we decided to base the scenario in a representative region of the study site without a more specific localization.
Figure 1
BIOcean5D study region and location where the expert assessment was carried out during the 2025 general assembly (Barcelona).
While there is important heterogeneity regarding plankton ES depending on local conditions (Russo et al., 2025), we were interested in the general potential of marine protection for ES/EDS outcomes and refined the scenario by providing a broad description of environmental conditions and economic activities in a representative area (see below). The ES/EDS indicators were based on the literature review that constitutes the first part of this article. Under consideration of their relevance for our specific area of interest, we developed 19 quantitative ES and 12 EDS indicators (Tables 1 and 2). According to the framework by Haines-Young and Potschin (2010), a cascading relationship exists between the two ends of a “production chain” of ecosystem services: the ecological structures and processes, or supply, on the one end and the final benefits to humans, or demand, on the other. Certain plankton ES, such as provisioning services depending on extraction, require active demand, which may be confined to certain geographical areas. In such cases, we defined the quantitative indicators as the potential for the provision of the service. The scenario and indicators were developed with counseling by one of the experts.
Table 1
| MEA ecosystem service category | Description | Indicator | Label | References |
|---|---|---|---|---|
| Provisioning services | ||||
| Food, other materials | Zooplankton harvest | Potential quantity of zooplankton harvested for human consumption or other uses | P1 | Raposo et al. (2022); Edelist et al. (2021) |
| Phytoplankton harvest | Potential quantity of phytoplankton harvested for human consumption or other uses | P2 | Gantar and Svircev (2008) | |
| Regulating services | ||||
| Air quality maintenance | Photosynthesis | Quantity of oxygen produced by phytoplankton | R1 | Falkowski (2012) |
| Climate regulation | Phytoplankton as part of the biological carbon pump | Quantity of CO2 acquired by phytoplankton | R2 | Turner (2015); Cavan et al. (2024) |
| Zooplankton as part of the biological carbon pump | Quantity of CO2 sequestered through zooplankton fecal pellets | R3 | Turner (2015); Cavan et al. (2024) | |
| Quantity of CO2 sequestered through dead zooplankton | R4 | Turner (2015); Cavan et al. (2024) | ||
| Quantity of carbon sequestered through the lipid pump | R5 | Turner (2015); Cavan et al. (2024) | ||
| Water purification and waste treatment | Zooplankton regulation of waste | Quantity of N&P reduction by jellyfish and copepods | R6 | Prakash et al. (2022) |
| Biological control | Zooplankton role in biological control | Quantity of harmful species populations predated by zooplankton | R7 | Von Rückert and Giani (2008) |
| Phytoplankton role in biological control | Quantity of harmful phytoplankton outcompeted by non-harmful populations | R8 | Naselli-Flores and Barone (2011) | |
| Cultural Services | ||||
| Education and science | Zooplankton and phytoplankton research | Number of scientific studies based on plankton | C1 | Yuan et al. (2015); Bezares Calderon et al. (2024) |
| Zooplankton and phytoplankton as biomonitors | Number of ecosystem health assessments based on plankton | C2 | Boldrocchi et al. (2023) | |
| Recreation and tourism | Bioluminescent plankton as tourist attraction | Number of tourists engaging with bioluminescent plankton | C3 | Haddock et al. (2010) |
| Supporting Services | ||||
| Food source for higher trophic levels | Zooplankton as a food source for other species | Quantity of zooplankton that serves as food for pelagic organisms | S1 | Turner (2004); Hofmann and Murphy (2004) |
| Quantity of zooplankton that serves as food for benthic organisms | S2 | Wotton and Malmqvist (2001) | ||
| Phytoplankton as a food source for other species | Quantity of phytoplankton that serves as food for pelagic organisms | S3 | Falkowski (2012) | |
| Quantity of phytoplankton that serves as food for benthic organisms | S4 | Falkowski (2012) | ||
| Larvae recruitment to fisheries | Importance of zooplankton for larval recruitment to fisheries | Number of fish larvae that develop into adult fish | S5 | Lomartire et al. (2021) |
| Hosts and refugia for other animals | Importance of zooplankton as a host or refugia | Number of juvenile animals that find refuge in jellyfish | S6 | Doyle et al. (2014) |
List of plankton ecosystem services (MEA classification), associated quantitative indicators used in the expert assessment, labels used for the analysis and references that served as basis for the indicators.
Table 2
| Ecological process | Dimension of well-being affected | Indicator | Label | References | |
|---|---|---|---|---|---|
| Harmful algal blooms | Economic dimension | Share of harmed animals in aquaculture | HAB1 | Trottet et al. (2022) | |
| Share of harmed animals in fisheries | HAB2 | Karlson et al. (2021) | |||
| Reduction in tourism potential due to harmful algal blooms | HAB3 | Alvarez et al. (2024) | |||
| Health dimension | Number of people harmed through direct contact | HAB4 | Van Dolah et al. (2001) | ||
| Number of people harmed through toxic food intake | HAB5 | Van Dolah et al. (2001) | |||
| Ecological dimension | Number of marine organisms harmed | HAB6 | Karlson et al. (2021) | ||
| Jellyfish blooms | Economic dimension | Share of harmed animals in aquaculture | JB1 | Bosch-Belmar et al. (2021) | |
| Share of harmed animals in fisheries | JB2 | Bosch-Belmar et al. (2021) | |||
| Reduction in tourism potential due to jellyfish algal blooms | JB3 | Kennerley et al. (2022) | |||
| Health dimension | Share of people stung | JB4 | Lakkis et al. (2015) | ||
| Ecological dimension | Number of toxic algal blooms facilitated by jellyfish predation on grazing zooplankton | JB5 | Stoltenberg et al. (2021) | ||
| Albedo effects | Phytoplankton reduces ocean surface albedo | Changes in ocean surface layer temperature due to phytoplankton | A1 | Frouin and Iacobellis (2002) | |
List of plankton ecosystem disservices, associated quantitative indicators used in the expert assessment, labels used for the analysis and references that served as basis for the indicators.
The scenario description specified that the evaluation should consider a representative, currently unprotected sea shelf area within the BIOcean5D study region, which due to its ecological or biological characteristics, could be subject to protection. It was also supposed that economic activities, such as commercial fishing, aquaculture, tourism, transportation, recreation and scientific research occur in a way typical for the region. Table 3 summarizes general conditions of European sea shelf waters regarding plankton ecological conditions, anthropogenic pressures and socioeconomic context. Plankton primary productivity is generally moderate to high in coastal areas, with signs of stress due to eutrophication and warming. Habitat degradation is prevalent in seagrass beds and benthic communities and fish biomass is moderate to declining. Anthropogenic pressures include fishing, aquaculture, coastal development, pollution from agricultural runoff and maritime transport and localized tourism impacts. Coastal economies are generally mixed-use, with fisheries, aquaculture, tourism and port-related activities being the main sources of ocean-related income.
Table 3
| Baseline Category | General description | Exemplifications |
|---|---|---|
| Ecological Baseline | Moderate to high levels of plankton productivity in coastal and shelf areas, with signs of stress due to eutrophication and warming trends; habitat degradation in seagrass beds and benthic communities; moderate to declining fish biomass. | Plankton productivity and eutrophication: Coastal and shelf areas exhibit moderate to high levels of plankton productivity. However, this productivity is increasingly stressed by eutrophication and warming trends. Elevated nutrient inputs from anthropogenic sources lead to eutrophication and hypoxic conditions that disrupt benthic communities, reduce fish biomass and promote phytoplankton blooms that can disrupt ecosystem balance (Cermeño et al., 2008; Malone and Newton, 2020). Simultaneously, rising sea surface temperatures affect plankton community composition and reduce overall productivity (OSPAR, 2023). |
| Habitat degradation in seagrass beds and benthic ecosystems: Seagrass meadows are declining globally due to eutrophication, warming waters, and physical disturbances, which threaten their role in supporting marine biodiversity (Waycott et al., 2009). Together with benthic habitats play a crucial role in supporting plankton ecosystems, however due the degradation of these habitats and factors like coastal development and pollution, can alter nutrient cycling and light availability, impacting phytoplankton growth and diversity (Duffy, 2006). | ||
| Moderate to declining fish biomass: Overfishing and habitat degradation have led to declining fish biomass in many coastal regions, affecting the sustainability of fisheries and the health of marine ecosystems (Palomares et al., 2020). Changes in plankton communities, particularly reductions in phytoplankton biomass and shifts toward less nutritious species, have cascading effects on higher trophic levels. Such alterations can lead to declines in fish biomass, affecting fisheries and overall marine biodiversity (OSPAR, 2017). | ||
| Anthropogenic Pressures | Presence of artisanal and industrial fishing, aquaculture, coastal urbanization, moderate pollution loads (mainly from agricultural runoff and maritime transport), and localized tourism impacts, recreation, scientific research and other anthropic activities occur. | Artisanal and industrial fishing pressures, including aquaculture: Both artisanal and industrial fishing contribute to the depletion of fish stocks and can negatively impact marine ecosystems through bycatch and habitat destruction (Coll et al., 2010). Additionally, overfishing alters food web structures, potentially leading to changes in plankton predator populations and nutrient cycling, which in turn influence plankton community composition considering the top-down effect that fishing activities can have on planktonic communities (Reid, 2000). |
| Coastal urbanization, pollution and other anthropic activities: Urban development along coastlines increases pollution from sewage, agricultural runoff, and maritime activities, leading to the degradation of coastal habitats (Barragán and De Andrés, 2015). Climate change and elevated microplastic concentrations have been shown to exert the most significant negative effects on copepods and krill, with cascading consequences for ecosystem services such as climate regulation, water quality, material provisioning, scientific research, and recreational activities (Botterell et al., 2023) | ||
| Tourism and recreation impacts: Tourism in coastal areas can lead to habitat destruction, pollution, and increased pressure on marine resources, necessitating sustainable management practices (Mejjad et al., 2022). Increased nutrient loads from tourism infrastructure can exacerbate eutrophication, leading to algal blooms and altered plankton dynamics (Li et al., 2025), causing in turn negative impacts at the socioeconomic level due to the increase in harmful algal blooms (Alvarez et al., 2024). | ||
| Socioeconomic Context | Mixed-use coastal economies, with fisheries, aquaculture, tourism, and port-related activities being the main sources of income; moderate population density with varying dependence on marine resources across subregions. | Mixed-use coastal economies: Coastal economies often rely on a mix of fisheries, aquaculture, tourism, and port-related activities, making them vulnerable to environmental changes and resource depletion (European Commission. Directorate General for Maritime Affairs and Fisheries. & European Commission. Joint Research Centre, 2021). |
| Population and dependence on marine resources: The livelihoods of coastal communities are closely tied to marine resources, with varying degrees of dependence influenced by population density and economic activities (Selig et al., 2019). Communities relying heavily on fisheries and aquaculture are particularly vulnerable to changes in plankton ecosystems, which can affect food security and livelihoods (Chapman et al., 2020). |
Baseline of ecological conditions, anthropogenic pressures and socioeconomic context for the representative European sea shelf area considered in the expert assessment.
The change scenario to be evaluated was the introduction of a marine protected area (MPA) of average size. Most of the European sea shelf MPAs are smaller than 150 km² (Aminian-Biquet et al., 2024). The scenario specifies that the MPA would be fully protected following IUCN definition of category Ia: “Strictly protected areas set aside to protect biodiversity and also possibly geological/geomorphological features, where human visitation, use and impacts are strictly controlled and/or limited to ensure protection of the conservation values. Such protected areas can serve as indispensable reference areas for scientific research and monitoring.” (Day et al., 2019), meaning that the allowed activities are limited to sustainable tourism and scientific research. We chose this policy option to be evaluated because MPAs increasingly have multiple objectives, including sustaining and/or increasing the provision of ecosystem services (Arkema et al., 2024). We do not intend to suggest that full protection is the option that necessarily maximizes (plankton) ES, but instead aim to create a scenario that is both easily understandable and includes a significant contrast between the two comparison points. Rather than limiting the expert assessment tool to this case, we hope that it can be used to assess other scenarios, for instance to site MPAs and evaluate different management measures.
It is assumed in the scenario that the MPA would be well-managed and therefore ensure efficient protection, and that the evaluation period is long enough so that changes could be detected, which is generally assumed to be approximately 5 years after implementation (Brun et al., 2024; Aurellado et al., 2021; Lenihan et al., 2021; Marriott et al., 2021; Abesamis et al., 2014). However, land-based protection is not included in the scenario and thus typical pressures originating from terrestrial activities would still be present.
The expert assessment took place during a workshop at the BIOcean5D general assembly in February of 2025. Two study authors delivered a PowerPoint-supported oral presentation, followed by a Q&A and short deliberation among scientists to ensure common understanding of the task and indicators. Subsequently, the experts received a link to a Qualtrics-based online survey, where the scenario was once again briefly outlined and they were asked to individually provide their evaluations of expected changes in the ES/EDS indicators following the introduction of the MPA.
We distinguish between two groups of experts, based on their own declarations: those that are explicitly plankton experts because a significant part of their work deals with plankton, and other marine scientists. Additionally, we asked the respondents for details about their scientific background. The expertise of the self-identified plankton experts spanned the following domains: plankton ecology, including phytoplankton and zooplankton dynamics, HABs, and marine eukaryotic biodiversity; genomic approaches, including metagenomics and environmental genomics, with bioinformatics applied to the study of cyanobacteria, picophytoplankton, and microalgae; biogeography and system-level analysis, encompassing phytoplankton and fish; and numerical modeling, focused on simulating plankton dynamics. The areas of expertise of the other marine scientists were closely related, included marine biology and microbial ecology, with a focus on marine microorganisms, diatom-bacteria interactions, and microbial diversity in deep-sea sediments; environmental chemistry, particularly in relation to organic matter dynamics; ecosystem and food web modeling, with applications to plankton diversity and phytoplankton biogeochemical roles.
Additional information about the participants and their scientific backgrounds can be found in the Supplementary Material (S3).
A total of 19 scientists (8 plankton experts) responded to the survey. Campagne et al. (2017)’s investigation of panel sizes for ecosystem service expert capacity matrices suggests that ideally, a panel should include at least 10 experts.
For the assessment, the experts compared the present state (unprotected sea shelf area in the target region) to the hypothetical scenario (the same area as a fully protected MPA). The expected changes were measured on a 7-point scale ranging from significant decrease (-3), moderate decrease, slight decrease, no change (0), to slight increase, moderate increase and significant increase (+3). Participants were explicitly asked to indicate 0 if they expected no change, and to leave the score blank if they did not feel like they had sufficient knowledge to answer. The scale was complemented with relative quantitative indications, where -3 (+3) was taken to be a >10% decrease (increase), -2 (+2) a 5-10% decrease (increase) and -1 (+1) a ~5% decrease (increase). Thus, all values are estimated relative differences between the status quo and the hypothetical scenario.
In addition to the expected changes, experts were asked to indicate how confident they were in their answers, measured on a scale from 1 (low confidence) to 5 (high confidence), following the recommendation of Campagne et al. (2017). They could also provide comments and explanations of their scores where they deemed it necessary.
3 Results
3.1 Classification and economic values of marine plankton ES and EDS
Figure 2 provides a textual, Figure 3a visual overview of our findings from the literature review. Plankton ES are categorized following the MEA framework, distinguishing between zoo- and phytoplankton. Plankton EDS are categorized following Shackleton et al. (2016) and Campagne et al. (2018). We indicate with symbols whether and how economic associations have been established for the respective ES and EDS. The literature review is detailed below.
Figure 2
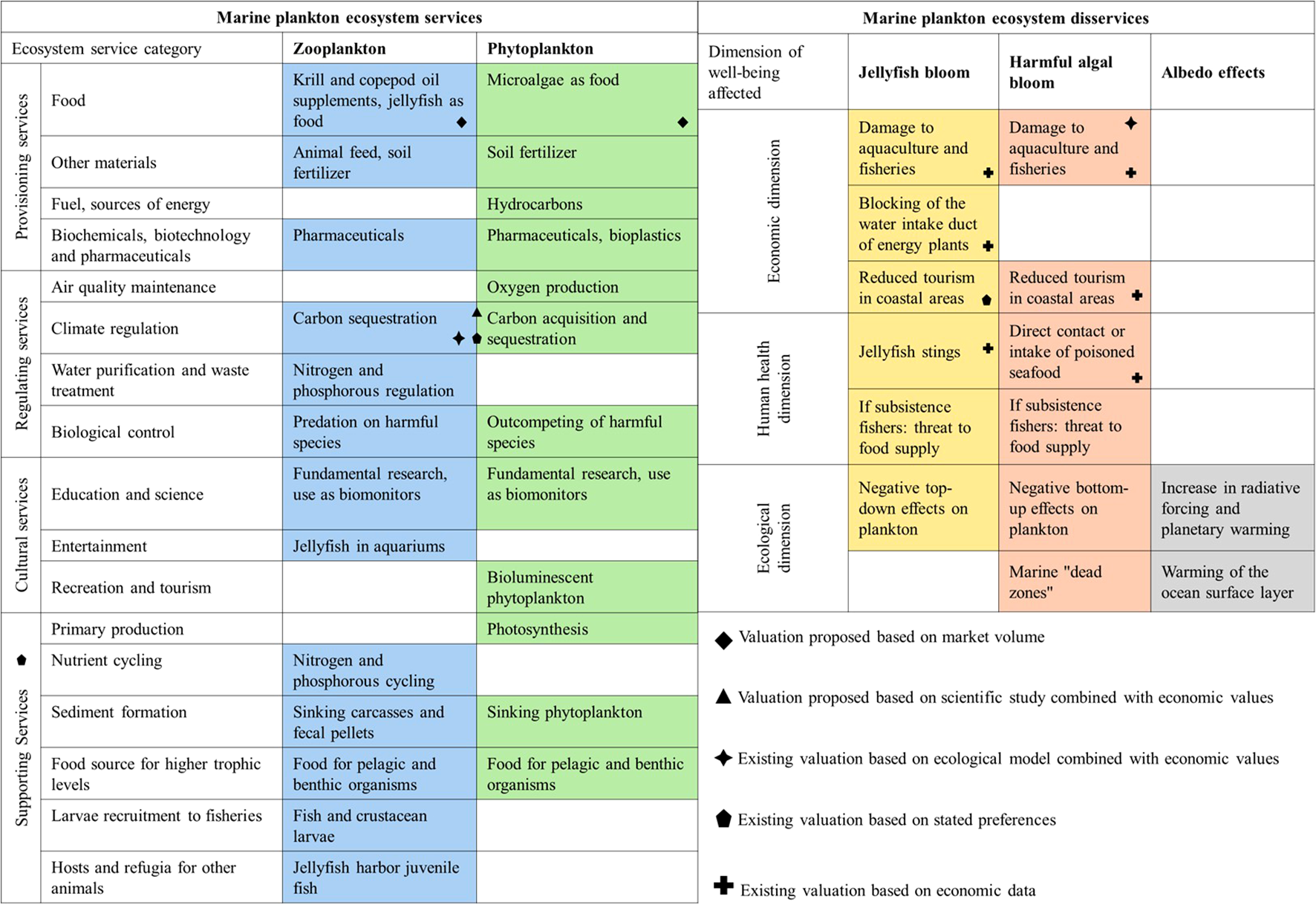
Plankton ES and EDS identified in the literature. Symbols indicate those for which valuation studies have been conducted (existing valuation) and for which proxies are proposed in this article (valuation proposed).
Figure 3
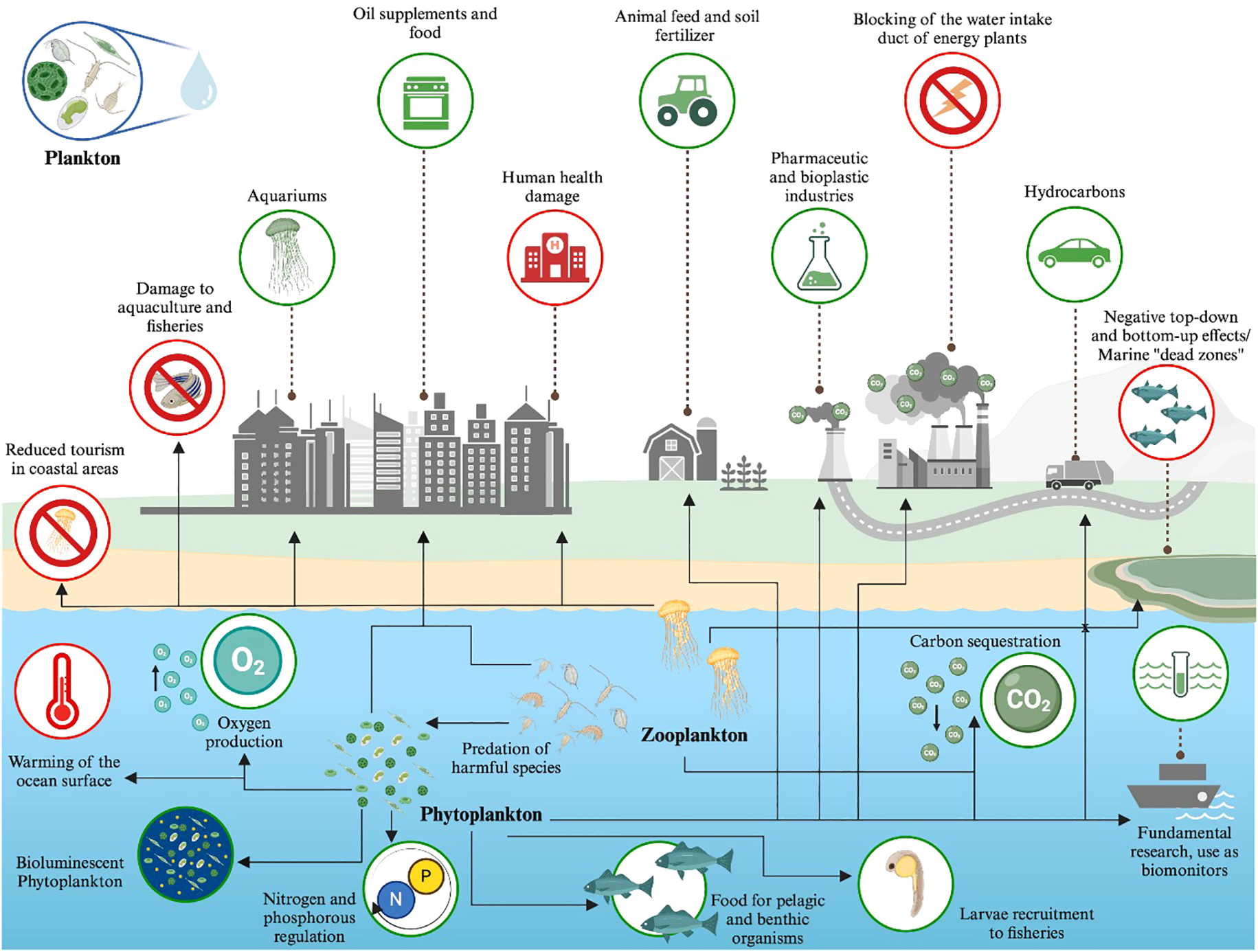
Visual representation of marine plankton ES (green) and EDS (red). Created in https://BioRender.com.
3.1.1 Ecosystem services
Despite the recent emergence of literature on plankton ecosystem services, explicitly linking them with economic values is essentially a new practice. The only three economic valuation studies explicitly addressing plankton ecosystem services are by Berzaghi et al. (2025); Cavan et al. (2024) and Jean-Louis et al. (2025). The ecosystem service valued in the former two is carbon sequestration, considering the entire biological carbon pump (Berzaghi et al., 2025) and krill carbon sequestration in the Southern Ocean (Cavan et al., 2024), respectively. In the study by Jean-Louis et al. (2025), several attributes related to plankton biodiversity are valued, which, following our classification, contain both the provision of ES and the mitigation of EDS. Different ES categorizations were used in previously published reviews. Regarding phytoplankton, Naselli-Flores and Padisak (2023) and B-Béres et al. (2023) use the MEA framework. For zooplankton ES, Botterell et al. (2023) employed the CICES (Commin International Classification of Ecosystem Services) framework, with the addition of supporting services. We use the MEA framework because of its simplicity and the prospective inclusion of supporting services.
3.1.1.1 Provisioning services
Marine zooplankton have been associated with an array of provisioning services. Krill and copepods are used to make oil supplements that are rich in omega-3 fatty acids, and also contain proteins, vitamins, minerals and the antioxidant astaxanthin (Kwantes and Grundmann, 2015; Štepán et al., 2021). They are thus used for human consumption as well as animal feed. Gelatinous zooplankton, i.e. jellyfish, are also consumed as food, especially in Asia (Raposo et al., 2022), while other possible markets, notably in Europe, are being explored (Edelist et al., 2021). Jellyfish are further used as animal feed, soil fertilizer, and for pharmaceutical and biotechnological applications, notably due to their collagen, fatty acids and venom (Duarte et al., 2022).
Jellyfish are harvested through fishing, but jellyfish aquacultures also exist, mainly in China (Duarte et al., 2022). The global production of jellyfish in 2021 amounted to 208,000 t (FAO, 2024) with a “stable value of around USD 2500/ton” (Raposo et al., 2022, p. 3), and thus a total value of about USD 520 million.
Krill and copepods are fished, but not produced in aquaculture (Abate et al., 2015). The global market volume of krill oil in 2021 was estimated at USD 824.17 million (Grand View Research, 2021).
Phytoplankton, such as microalgae and cyanobacteria, are also consumed as food in several non-Western countries (Gantar and Svircev, 2008). They are further used as animal feed, soil fertilizer and their pigments are extracted as natural coloring agents (Abdulrahman et al., 2023; Pagels et al., 2021). Certain phytoplankton species possess the potential to produce hydrocarbons which represent a renewable energy source (Djoru and Gimin, 2020). Microalgae and cyanobacteria are used for the production of bioplastics (López-Pacheco et al., 2022) and, due to their capacity to produce toxins, for pharmaceutical applications. Microalgae are typically cultivated in hatcheries. Additionally, phytoplankton have been shown to biodegrade plastics, thereby contributing to waste treatment and the improvement of ocean health (Schenone et al., 2025). The global microalgae market was estimated at USD 3.4 billion in 2020 (Loke Show, 2022).
3.1.1.2 Regulating services
Both marine phyto- and zooplankton contribute to the biological carbon pump, i.e. the ocean’s biologically driven sequestration of carbon. Phytoplankton perform photosynthesis, thereby acquiring CO2 (Falkowski, 2012).
Subsequently, they are either consumed by other organisms, or they die and sink to the ocean floor. Zooplankton further contribute to carbon sequestration through the sinking of fecal pellets and carcasses. Additionally, vertically migrating zooplankton enhance carbon storage through their movement (Turner, 2015). The physical movement of water masses, particularly vertical mixing, also adds to the sequestration of carbon (Claustré et al., 2021).
The carbon sequestration potential of open-ocean pelagic ecosystems, and notably plankton, is vastly under-investigated. Cavan et al. (2024) conducted an estimation of the amount and economic value of carbon sequestered by antarctic krill. Using krill density based on samples, a fecal pellet egestion rate and an ocean circulation model, they conclude that krill sequester 20MtC per year, which is a similar amount to the key coastal “blue carbon” habitat represented by mangroves. Multiplication with a social cost of CO2 (SCCO2) range between USD 51 (SCCO2 in the US at the time of the study) - 640 (global average at the time of the study) per tCO2 leads to an estimated economic value range of USD 3.7 - 46.1 billion per year.
Nowicki et al. (2022) assimilated satellite and in situ ocean biogeochemical observations to model the entire biological pump and quantify global and regional carbon export and sequestration. They used the Carbon, Acidification, Fisheries, Ecosystems (CAFE) model for their estimation of net primary production. The total amount of carbon exported via the biological pump is estimated at 10.2 Pg per year. The study authors do not provide monetary estimates, but if we use Cavan et al. (2024)’s approach and SCCO2 range, we obtain the following results: with a conversion factor of 3.67 (EPA, 2024), the biological pump export is 37.43 Pg CO2 and the total economic value USD 1.91 - 23.99 trillion per year. More recently, Berzaghi et al. (2025) also modelled global carbon export attributable to the biological pump, using the NEMO-PISCES-APECOSM model which explicitly includes fish and zooplankton vertical migration, and obtained an estimate of 2.81 Pg of carbon per year. They applied a CO2 offsetting price of USD 90 and subsequently reported a yearly total value of USD 928 billion for this plankton ES.
The deliberative choice experiment by Jean-Louis et al. (2025) included an attribute on marginal increases in carbon sequestration in a proposed MPA. On average, the workshop participants from 5 European countries were willing to pay about 0.30€ (USD 0.32) for a 1% increase in carbon sequestration by plankton.
Additionally, zooplankton can contribute to water purification and the regulation of waste, by reducing nitrogen and phosphorus concentrations (Prakash et al., 2022). Zooplankton and phytoplankton can fulfil functions of biological control, through predation (Von Rückert and Giani, 2008) and competition (Naselli-Flores and Barone, 2011) of harmful species.
3.1.1.3 Cultural services
Marine plankton provide ecosystem services related to education and science, notably by functioning as bioindicators for the ecological status of marine environments (Boldrocchi et al., 2023), as well as serving as a foundation for fundamental research (Yuan et al., 2015; Bezares Calderon et al., 2024). Recent European marine research and citizen science projects with a strong focus on plankton, such as BIOcean5D, Plankt’Eco and Plankton Planet, have received substantive research funds, demonstrating marine plankton’s high scientific relevance.
Regarding the cultural service of entertainment, jellyfish are displayed in aquariums, providing an aesthetic and educational experience to visitors (Hayward, 2012). Additionally, bioluminescent phytoplankton has become a tourist attraction (Haddock et al., 2009).
3.1.1.4 Supporting services
Plankton are among the main providers of oceanic supporting services. Phytoplankton-based primary production generates vital biomass and oxygen (Falkowski, 2012). Both zoo- and phytoplankton are important for the cycling of nutrients, notably nitrogen and phosphorus (Arrigo, 2005; Karakuş et al., 2022). Both provide food sources for higher trophic levels (Turner, 2004) and contribute to sediment formation by sinking to the ocean floor (Bostrom et al., 1974). Fish and crustaceans have an early developmental stage within the zooplankton (Lomartire et al., 2021), and thus the recruitment of zooplankton larvae is a supporting service for fisheries. Jellyfish, moreover, act as refugia for juvenile fish which they shelter under their bells (Doyle et al., 2014).
In the deliberative choice experiment by Jean-Louis et al. (2025), the attribute “stable plankton composition” is described as essential for enabling/supporting all plankton ecosystem services. On average, participants were willing to pay at least 50€ (USD 54) to increase plankton composition stability, making it the highest-valued attribute.
3.1.2 Ecosystem disservices
Besides the numerous benefits that marine plankton provide for humans, planktonic activities are also associated with negative impacts on well-being. In particular, blooms of jellyfish and toxic algae have been characterized with regard to the damages they cause to economies, human health and marine environments (Bosch-Belmar et al., 2021; Carias et al., 2024).
Jellyfish blooms have been characterized by Botterell et al. (2023) and Graham et al. (2014) along the lines of ecosystem services categories, i.e. as negative effects on provisioning, regulating, cultural and supporting ecosystem services. However, as Shackleton et al. (2016) point out, a characterization of EDS as the absence or diminishment of ES may be insufficient for a weighing of policy options, especially because not all dimensions of EDS may be covered. For instance, classifying effects of jellyfish stings as a diminishment in a cultural service seems too reductive, as the effect is not limited to the elimination of a pleasant experience, but creates an additional negative impact that may cause serious impairments or even death. Therefore, we propose a classification of plankton EDS, following the framework by Shackleton et al. (2016) with modifications inspired by Campagne et al. (2018), which distinguishes between three dimensions of well-being affected: economic, health-related and ecological.
3.1.2.1 Jellyfish blooms - economic dimension
Important economic harms of jellyfish blooms affect the fisheries sector and are linked to damaged catch, i.e. the negative effects of predation and toxins on commercially valuable fish, and to damaged or clogged fishing nets (Bosch-Belmar et al., 2021). Detailed information on the extent of these economic impacts on the fishing sector is rare and global estimates are not currently available. Several local estimations using different quantification methods have been undertaken and shall be briefly reviewed here.
The most comprehensive study focusing on the economic impact of jellyfish blooms on fisheries was published by Palmieri et al. (2014) and focused on the Italian northern Adriatic Sea. The estimation was based on interviews conducted with fishers in 2012 regarding their economic losses which were then extrapolated to the regional level. Economic losses due to reductions in fish catches were estimated at USD 8.2 million per year and fuel costs related to the displacement of fishing activities were estimated at USD 460,000. The Northern Adriatic fleet makes up 30% of the Italian fleet in terms of size (European Commission, 2022). Fishing volume in the Italian Adriatic Sea decreased from 88,376 tons in 2012 to 59,773 tons in 2023 (Eurostat, 2025) in line with the objectives of the EU Common fisheries policy (CFP), therefore the nominal losses linked to jellyfish may have decreased as well.
Another study based on self-reported costs to fishers was conducted by Uye (2011). It focused on damages caused by the giant Nemopilema jellyfish to set-nets around the Japanese coast. During bloom periods, clogging can lead to breaking nets, fishing is suspended and fishermen may be laid off. An important bloom happened in 2005, which caused economic damages worth USD 380 million to fisheries according to reports made of fishers to the Japan Sea National Fisheries Research Institute. In 2009, despite an important bloom, economic losses were reduced to USD 125 million thanks to an improved warning system (Lucas et al., 2014).
Graham et al. (2003) reported an expert assessment conducted by a member of the Mississippi Department of Marine Resources regarding the losses to the US shrimp fishery in the Gulf of Mexico in the year 2000 as a result of the invasive rhizostome Phyllorhiza punctata. They estimated the direct losses associated with the clogging of shrimp nets to be about USD 10 million, while indirect effects related to predation on eggs and larvae could not be estimated due to a lack of detailed information.
Regarding aquaculture, jellyfish impacts have mainly been linked to increases in fish mortality and gill damage (Bosch-Belmar et al., 2021). Their full economic extent is difficult to estimate, because jellyfish involvement in gill pathologies is currently unquantifiable (Clinton et al., 2021). Bosch-Belmar et al. (2021) provide the first compilation of economic estimations of jellyfish interactions on aquaculture, based on “grey” literature. According to this data, economic losses due to a Pelagia noctiluca bloom killed 100,000 salmon in Ireland in 2013, leading to an economic loss of at least USD 1.2 million. The same species caused a loss of USD 1.9 million in Scotland in 2014, when 300,000 salmon died. In 2018, a bloom of Aurelia led to the death of an unspecified number of salmon in Tasmania, causing damages estimated at USD 7.1 million.
Another economic aspect of wellbeing impacted by jellyfish blooms is the obstruction of cooling water intake for nuclear- and coal-fired power stations (Lucas et al., 2014). Following increases in the volume of marine biota, for example due to blooms, the intake of water can be significantly reduced. Power stations then run at reduced efficiency or they may even temporarily shut down as a precautionary measure to prevent overheating of the reactors. The only available economic assessment we found was conducted in south-east India in 1995 and 1996 (Masilamoni et al., 2000). It was found that large numbers of jellyfish appeared on the water intake screens of the Madras Atomic Power Station in the summer, leading to head loss worth about USD 2,000 per day. Additionally, the plant was forced to temporarily shut down because of jellyfish blocking the cooling water intake, causing losses of USD 100,000 per day.
Jellyfish blooms also negatively impact the highly lucrative tourism sector. Due to the risk of stings and aesthetic concerns, tourists tend to avoid jellyfish bloom-prone areas. Stated preference studies were a preferred way to value this EDS. A study eliciting hypothetical beach trip frequency conducted in Israel in 2013 (Ghermandi et al., 2015) found that a jellyfish bloom would reduce the number of beach trips between 3% and 10.5%, which corresponds to a monetary loss in the range between 8.9 million ILS (USD 2.37 million, converted to 2013 USD and adjusted for purchasing power parity) and 31.1 million ILS (USD 8.29 million). A choice experiment conducted in Spain in 2011 (Nunes et al., 2015), estimated the potential welfare gains related to a reduction in jellyfish bloom risk. It was reported that aggregated wellbeing gains associated with a reduction of jellyfish blooms in Catalonia would be around 422.60 million Euro annually (USD 503.1 million). In 2018, another choice experiment study was conducted among international tourists in the island of Mallorca (Spain) (Ruiz-Frau, 2023), which observed that tourists were on average willing to pay 33.3 Euro (USD 37.84) to avoid high jellyfish presence. No population-level extrapolation was made in the study, but if we consider that 11.7 million international tourists visited Mallorca in 2018 (https://roadgenius.com/statistics/tourism/spain/mallorca/), then the total willingness-to-pay (WTP) may be up to 389.61 million Euro (USD 442.74 million).
In the choice experiment study by Jean-Louis et al. (2025), the risk for jellyfish blooms and for harmful algal blooms was combined into a single dichotomous attribute. Across all study locations, bloom avoidance received the lowest WTP of the attributes, between 12€ and 17€ per year (USD 13-18). However, its perceived value is likely conflated with that of the supporting service “stable plankton composition”, as discussed above.
3.1.2.2 Jellyfish blooms - human health dimension
Some species of jellyfish possess stinging cells which produce harmful toxic effects on humans. Depending on the type of venom, reactions include skin irritation, pain, nausea, or even cardiac and respiratory arrest which can be fatal (Lucas et al., 2014). Jellyfish sting-related deaths are quite rare, but the most frequent occurrences seem to be in Malaysia and the Philippines, where about 20 to 50 people die each year as a result of jellyfish stings (Fenner et al., 2010). Non-fatal stings are especially frequent in Australia, where about 10,000 stings each summer are attributed to Physalia physalis alone (Lucas et al., 2014). We found one study that specifically investigated jellyfish sting-induced health costs. Based on hospital admission data from the Salento area (Italy), a study conducted between 2007 and 2011 estimated that about 40,000 jellyfish stings occurred on Italian coasts during the study period, causing 2 million Euros per year in cost to the Italian healthcare system (De Donno et al., 2014).
Finally, human health is affected when subsistence fisheries, i.e. fishers and their families who consume the fish they catch themselves, rather than selling them, are threatened by the massive presence of jellyfish. A recent study ascertained that 52.8 million people engaged in subsistence fishing at some point during the year in 2016 (Virdin et al., 2023). Jellyfish blooms threaten subsistence fisher’s vital catches through the direct effects on fishing described above or through trophic cascades detailed in the next paragraph (Lloret et al., 2018).
3.1.2.3 Jellyfish blooms - ecological dimension
Jellyfish, as alluded to above, fulfil important functions in marine trophic webs. They are notably predators of zooplankton and can influence the plankton ecosystem structure through trophic cascades (Wright et al., 2021). With grazing zooplankton diminished by jellyfish, increases in phytoplankton can be observed. They also contribute to the biogeochemical cycle through excretion and sinking carcasses (ibid.). During jellyfish blooms, the high biomass thus achieved make them a significant vector for the export of nutrients to the benthos. Because of their bloom-and-bust dynamic, jellyfish represent unpredictable and ephemeral biomass (Fernandez-Alias et al., 2024).
3.1.2.4 Harmful algal blooms - economic dimension
Harmful algal blooms (HABs) are associated with some of the most feared plankton ecosystem disservices, which are closely linked to anthropogenic pressures such as overfishing and eutrophication (Berdalet et al., 2016). The fisheries sector is, in turn, economically susceptible to harmful algal blooms. Shellfish, and also finfish, are negatively affected by HABs, leading to temporary closing of fisheries, declines in consumer demand and reduced fish landings (Sanseverino et al., 2016). The full extent of economic damages to fisheries related to HABs is difficult to quantify because of a lack of detection or reporting as well as intricate relationships between HABs and their toxicity for fish. For example, blooms of the microalgae Heterosigma akashiwo may affect fish through the production of reactive oxygen species, excessive mucus production that impedes oxygen exchange and/or gill tissue damage, among others (Brown et al., 2020). Despite the persisting uncertainties, several studies and reviews have been conducted at different scales.
The most recent and comprehensive literature review on the valuation of HAB events was compiled by Carias et al. (2024). Several studies reviewed therein focus explicitly on losses to fisheries. A notable study was conducted by Park et al. (2013) and examined the period between 1981 and 2012. By means of multiplying average fishing revenue and the duration of HAB related closures, the authors estimate that losses of USD 5.4 million - USD 60 million were incurred by the Korean fishing sector. Mardones et al. (2020) also applied a price times quantity approach to estimate HAB-related damages to the shellfish industry in Chile between 2014 and 2018 and reached a cumulative estimate of USD 6.9 million. Another study conducted in Scotland focused on shellfish production between 2009 and 2018 (Martino et al., 2020). Unlike the price*quantity approach, the authors conducted multivariate time-series analysis using panel data from the Scottish Shellfish Production Survey to model variation in shellfish production including, besides HAB abundance, capital, labor and climate variables. The results suggest that HAB-induced economic losses of GBP 1.37 million per year in 2015 currency (USD 2.08 million) over a national annual industry turnover of GBP 12 million (USD 18.18 million) were generated.
Another study focused on salmon production in British Columbia between 2009 and 2012 (Haigh and Esenkulova, 2014). Based on survey data collected from aquaculture management, HAB-related losses were estimated to be CAD 16.135 million (USD 13.17 million, PPP-adjusted average 2009-2012) for the whole period.
Like jellyfish blooms, HAB events are economically detrimental for the tourism sector. A recent study analyzed revenues from tourism in Florida (USA) during the 2018 Karenia bloom (Alvarez et al., 2024). Tourism data was compared to counterfactual data in the assumed absence of HAB. The total loss to the tourism sector in Florida in 2018 is estimated to be USD 2.7 billion. A previously conducted study using the same 2018 data set as well as data from 2005 and 2006 focused on southwest Florida (Bechard, 2020) and applied a difference-in-difference approach to compare tourism revenues in HAB-affected counties to unaffected ones. Relative estimated sales losses are 5–7 percent in the lodging sector and 1.2-2.5 percent in the restaurant sector.
3.1.2.5 Harmful algal blooms - human health dimension
A primary source of health-related EDS caused by HABs is the consumption of affected shellfish and finfish. Different types of poisoning, depending on the type of toxin-releasing microalgae and the consumed species, exist (Grattan et al., 2016). Direct impacts result from skin contact with contaminated water and/or inhaling biotoxins in the form of aerosols (Berdalet et al., 2016). The main categories of resulting illnesses are respiratory and digestive. Determining the incidence of HAB-related diseases is very difficult due to lack of reporting and/or misdiagnosing (ibid.). In the United States, 95 cases of HAB-related illnesses were reported in 2022 (CDC, 2024), though no distinction was made between freshwater- and saltwater-related cases.
Kouakou and Poder (2019) conducted a systematic review on global public health costs caused by HAB events. Costs were calculated per reported case and converted into 2016 USD. Costs for respiratory illnesses were up to USD 14,600, while costs for digestive illnesses reached up to USD 12,605.
As with jellyfish, HAB events can threaten the livelihoods of subsistence fishers. In the United States, Indigenous communities are especially vulnerable to this threat because of their comparatively higher reliance on marine resources for subsistence (Kourantidou et al., 2022).
3.1.2.6 Harmful algal blooms - ecological dimension
Like humans, marine life is typically negatively affected by HABs either through direct exposure to the biotoxins, or through transfer through the food chain. As discussed above, HABs can be harmful for fish in a number of ways. They can lead to mass kills and diseases, which can spread also to birds that feed on impacted fish (Landsberg, 2002).
In some cases, phytoplankton blooms can fuel hypoxia (oxygen depletion), leading to marine “dead zones”, when fish die or migrate to other, oxygen-richer areas (Altieri and Diaz, 2019).
3.1.2.7 Albedo effects - ecological dimension
Albedo is the fraction of sunlight that is diffusely reflected by a body. Earth surface albedo ranges from 0.8 for freshly fallen snow (high albedo) to 0.04 for charcoal (low albedo) and plays a significant role in climate science (Stephens et al., 2015). Within the Gaia hypothesis conceptual framework, it was proposed that phytoplankton-produced sulphur acts as cloud condensation nuclei, leading to higher cloud albedo (Charlson et al., 1987). While this hypothesis has been revised and is now considered too simplistic (Green and Hatton, 2014), it contributed to opening a field of research on interrelationships of marine and atmospheric biogeochemistry. Marine phytoplankton has been suggested to reduce ocean surface albedo, thereby considerably contributing to atmospheric warming (Frouin and Iacobellis, 2002). Furthermore, albedo reductions due to phytoplankton have been linked with sea-surface temperature rises and evaporation, leading to atmospheric humidity increases, and thereby also the earth’s greenhouse effect (Patara et al., 2012). The albedo-reducing effect of phytoplankton is notably involved in a feedback loop where warming snow and ice environments are increasingly inhabited by pigmented microalgae, which in turn enhances surface melt (Chevrollier et al., 2023). Thus, we consider that in today’s rapidly warming climate, phytoplankton-induced albedo reductions may be considered an ecosystem disservice.
3.2 Expert assessment of changes in ecosystem service and disservice provision assuming the implementation of a fully protected marine area
The expected changes in plankton ES and EDS following the hypothetical implementation of a fully protected marine area of the 19 experts who participated in the assessment are summarized in Table 4. Plankton experts on average gave more estimations (non-blanks) than other surveyed marine scientists (22 and 17 out of 31 indicators, respectively). There is some heterogeneity with regard to the directions of the expected changes. For provisioning services, i.e. the potential extractive use of plankton for consumption or other purposes, most respondents indicated a slight to moderate positive expected change, however a minority expected slight to strong decreases.
Table 4
| ES/EDS label | Expert no. | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| P1 | -1 | 1 | 1 | -2 | 1 | 2 | 0 | 1 | 0 | 2 | 0 | -1 | -2 | 1 | 0 | |||||
| P2 | -3 | 0 | 1 | -1 | 1 | 2 | 0 | 1 | -1 | 2 | 0 | 2 | 0 | -1 | 0 | 2 | 0 | |||
| R1 | -2 | 1 | 1 | 2 | 1 | 2 | 0 | 0 | -1 | 2 | 1 | 1 | 1 | -2 | 1 | 0 | 3 | 0 | ||
| R2 | -1 | 1 | 1 | 2 | 2 | 2 | 0 | 0 | 1 | 2 | 1 | 1 | -1 | 1 | 0 | 3 | 0 | |||
| R3 | 1 | 1 | 2 | 0 | 2 | 1 | 1 | 1 | -1 | 0 | 2 | -2 | ||||||||
| R4 | -1 | 1 | 1 | 2 | 0 | 2 | 1 | 1 | 1 | 1 | -1 | 0 | 1 | -2 | ||||||
| R5 | -2 | 2 | 1 | 2 | 0 | 1 | 1 | 0 | -2 | |||||||||||
| R6 | 0 | 2 | 2 | 1 | 2 | -1 | 1 | 1 | 0 | -1 | 0 | |||||||||
| R7 | 2 | 1 | 2 | 1 | 1 | 3 | 1 | 0 | 0 | -1 | 0 | |||||||||
| R8 | 1 | 2 | 2 | 1 | 3 | 0 | 0 | 0 | 0 | 0 | 2 | -3 | ||||||||
| C1 | 3 | 3 | 2 | 2 | 3 | 3 | 1 | 2 | 1 | 3 | 2 | 2 | 3 | 3 | 3 | |||||
| C2 | 3 | 2 | 1 | 1 | 2 | 1 | 3 | 2 | 1 | 3 | 1 | 0 | 3 | 3 | ||||||
| C3 | 0 | 2 | 1 | 1 | 0 | 0 | 0 | 2 | 2 | 1 | 0 | 2 | 0 | |||||||
| S1 | -1 | 1 | 2 | 2 | 3 | 1 | 3 | 1 | 1 | 2 | 2 | 3 | ||||||||
| S2 | -1 | 1 | 2 | 2 | 3 | 2 | 1 | 1 | 2 | 2 | 3 | |||||||||
| S3 | -2 | 1 | 2 | 2 | 0 | 2 | 0 | 0 | 3 | 1 | 1 | -2 | 2 | 2 | 0 | |||||
| S4 | -2 | 1 | 2 | 2 | 1 | 2 | 1 | 0 | 3 | 1 | 1 | -2 | 2 | 2 | 0 | |||||
| S5 | -2 | 3 | 1 | 2 | 1 | 3 | 3 | 3 | 1 | 3 | 2 | 0 | ||||||||
| S6 | -1 | 3 | 1 | 2 | 0 | 2 | 3 | 0 | -1 | 0 | 0 | |||||||||
| HAB1 | -2 | -3 | -1 | -2 | -3 | -1 | 0 | 0 | -1 | -2 | -1 | -2 | ||||||||
| HAB2 | -2 | -1 | -1 | -2 | -3 | 0 | 0 | -1 | -2 | -1 | -2 | |||||||||
| HAB3 | -1 | -3 | -2 | -2 | -3 | -3 | 0 | -2 | -1 | -2 | ||||||||||
| HAB4 | -1 | -1 | -1 | -2 | -2 | -1 | -1 | 1 | -2 | -1 | ||||||||||
| HAB5 | -2 | 0 | -2 | -2 | -1 | -1 | 0 | 2 | 0 | -1 | ||||||||||
| HAB6 | -3 | -2 | -2 | -2 | -3 | -1 | -1 | -1 | -2 | -2 | ||||||||||
| JB1 | -3 | 0 | -1 | -2 | -1 | 0 | -2 | -2 | ||||||||||||
| JB2 | 0 | -1 | -2 | -1 | 0 | 0 | -2 | |||||||||||||
| JB3 | -2 | -3 | 0 | -1 | 0 | 0 | 0 | -2 | ||||||||||||
| JB4 | -3 | 0 | -1 | -1 | 0 | 1 | 0 | -2 | ||||||||||||
| JB5 | -3 | 0 | -2 | -3 | 0 | 0 | ||||||||||||||
| A1 | 0 | -2 | 0 | 0 | 0 | -2 | 0 | 0 | -2 | |||||||||||
Expert-assessed expected changes in ES/EDS provision following a hypothetical increase in marine protection from none to full protection. Plankton experts are highlighted in grey.
This observation mirrors a brief discussion of the items during the workshop, where it was discussed that a positive effect was conditional on the expectation that there would be an export of exploitable planktonic matter to the exterior of the hypothetically strongly protected area. Additionally, there may have been differences regarding the expectation of actual plankton harvesting activities in Europe, which we tried to circumvent somewhat with the indicators referring to “potential” (details in the methods section), but may not have eliminated all differences in interpretations.
Differences in marine scientists’ expectations regarding the direction of the effect on ES that are not contingent upon active demand, i.e. regulating and supporting services, persist, but are less frequent. Cultural services were unanimously evaluated as being positively impacted or unaffected. Regarding EDS, the plankton experts unanimously and other marine scientists almost unanimously expected decreases following increased protection, or no effects.
In order to obtain a more succinct overview of the assessment results, we provide descriptive statistics in Table 5. Employing the approach proposed by Schernewski et al. (2018), we use the median as an indicator of central tendency and calculate relevance and important scores. Relevance reflects the percentage of experts who considered the respective ES/EDS indicator as potentially affected by the scenario, which in our case meant a score that was not zero (no change expected) or blank (no knowledge). The importance score was then calculated as the absolute value of the product of the median and the relevance, divided by 10 to have a more manageable scale. Additionally, we report minimum and maximum scores.
Table 5
| ES/EDS label | Exp. change median | Relevance | Importance | Min, max |
|---|---|---|---|---|
| P1 | 0 | 57.89 | 0.00 | [-2,2] |
| P2 | 0 | 57.89 | 0.00 | [-3,2] |
| R1 | 1 | 73.68 | 7.37 | [-2,3] |
| R2 | 1 | 68.42 | 6.84 | [-1,3] |
| R3 | 1 | 52.63 | 5.26 | [-2,2] |
| R4 | 1 | 63.16 | 6.32 | [-2,2] |
| R5 | 1 | 36.84 | 3.68 | [-2,2] |
| R6 | 1 | 42.11 | 4.21 | [-1,2] |
| R7 | 1 | 42.11 | 4.21 | [-1,3] |
| R8 | 0.5 | 36.84 | 1.84 | [-3,3] |
| C1 | 3 | 78.95 | 23.68 | [1,3] |
| C2 | 2 | 68.42 | 13.68 | [0,3] |
| C3 | 1 | 36.84 | 3.68 | [0,2] |
| S1 | 2 | 63.16 | 12.63 | [-1,3] |
| S2 | 2 | 57.89 | 11.58 | [-1,3] |
| S3 | 1 | 57.89 | 5.79 | [-2,3] |
| S4 | 1 | 68.42 | 6.84 | [-2,3] |
| S5 | 2 | 57.89 | 11.58 | [-2,3] |
| S6 | 0 | 36.84 | 0.00 | [-1,3] |
| HAB1 | -1.5 | 52.63 | 7.89 | [-3,0] |
| HAB2 | -1 | 47.37 | 4.74 | [-3,0] |
| HAB3 | -2 | 47.37 | 9.47 | [-3,0] |
| HAB4 | -1 | 52.63 | 5.26 | [-2,1] |
| HAB5 | -1 | 36.84 | 3.68 | [-2,2] |
| HAB6 | -2 | 52.63 | 10.53 | [-3,-1] |
| JB1 | -1.5 | 31.58 | 4.74 | [-3,0] |
| JB2 | -1 | 21.05 | 2.11 | [-2,0] |
| JB3 | -0.5 | 21.05 | 1.05 | [-3,0] |
| JB4 | -0.5 | 26.32 | 1.32 | [-3,-1] |
| JB5 | -1 | 15.79 | 1.58 | [-3,0] |
| A1 | 0 | 15.79 | 0.00 | [-2,0] |
Median scores of all scientists for ES and EDS changes, relevance (% of respondents who indicated neither 0 nor blank for the respective indicator) and importance (|score|*relevance/10), and min/max of the change score.
The highest importance scores are obtained for cultural ES related to research (C1) and plankton biomonitoring (C2). Other ES that received high ratings are the quantity of zooplankton that serves as food for pelagic (S1) and benthic (S2) organisms as well as the number of planktonic fish larvae that develop into adult fish (S5). A moderate importance was assigned to the regulating ES of phytoplankton oxygen production (R1). Regarding EDS, all HAB-related EDS received high to moderate importance scores, with the share of harmed animals in aquacultures (HAB1) and the number of people harmed through toxic food intake (HAB6) being the most, and the number of people harmed through direct contact (HAB4) the least important. We report the same descriptive statistics when considering only plankton experts (Supplementary Material, Supplementary Table S4) and we observe the same tendencies as for all surveyed scientists, with the exception of plankton experts assigning considerably higher importance scores and expected positive changes in regulating services related to water purification and biological control. Specifically, these include the reduction of nitrogen and phosphorus by copepods and jellyfish (R6), the predation of harmful species populations by zooplankton (R7), and the competitive exclusion of harmful phytoplankton by non-harmful populations (R8).
A visual representation of expert-assessed potential of a fully protected MPA for the different categories of ES and EDS is given by Figure 4. Plankton experts showed slightly higher expectations for increases in the provision of regulating services and reductions of HAB and jellyfish-related disservices than other surveyed marine scientists. Notably, plankton experts who indicated that their work was directly related to HABs attributed the highest potential of marine protection to the reduction of HAB-related disservices. As reported above, the highest potential overall was reported for cultural and supporting ES (+) and for HAB-related EDS (-).
Figure 4
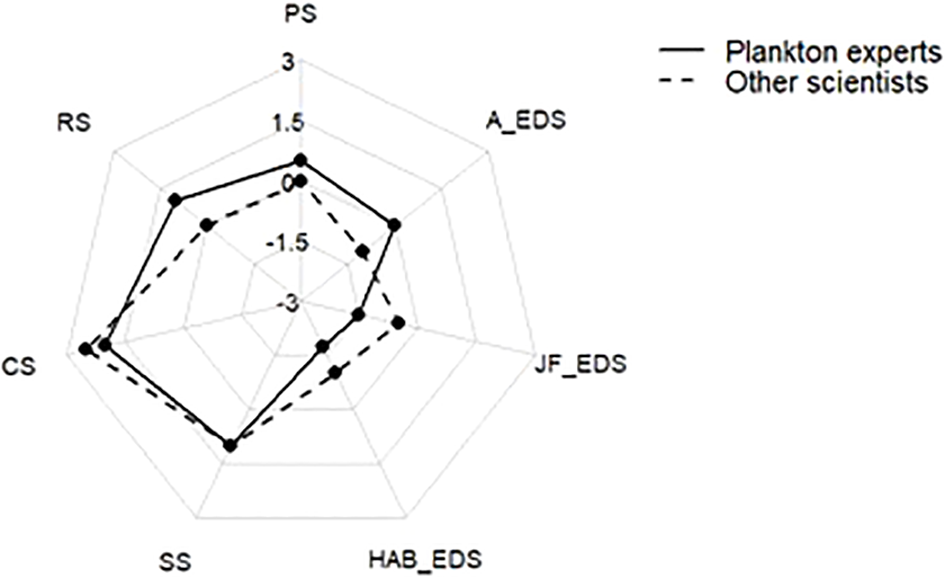
Potential of increased marine protection to contribute to plankton ES and EDS categories as expected by plankton experts and other marine scientists. Dots represent medians of the pooled ES/EDS scores of the respective categories (PS, provisioning services; RS, regulating services; CS, cultural services; SS, supporting services; HAB_EDS, HAB-related EDS; JF_EDS, jellyfish-related EDS; A_EDS, Albedo-related EDS).
The proportions of confidence scores for each ES/EDS are summarized in Figure 5. Following the recommendation by Campagne et al. (2017), we use the confidence scores as supporting information, rather than to compute final scores. Confidence scores for ES were not significantly different from those for EDS (Mann-Whitney U test, p=0.38), and respondents were overall moderately confident in their answers, with a median score of 3 out of 5. However, considering the ES and EDS with the highest importance scores described above, relatively low median confidence scores of 2 were assigned to phytoplankton oxygen production (R1), nitrogen and phosphorus reduction (R6), the share of harmed animals in aquacultures (HAB1), and the number of people harmed through toxic food intake (HAB6). We discuss implications of low confidence scores in the Discussion section.
Figure 5
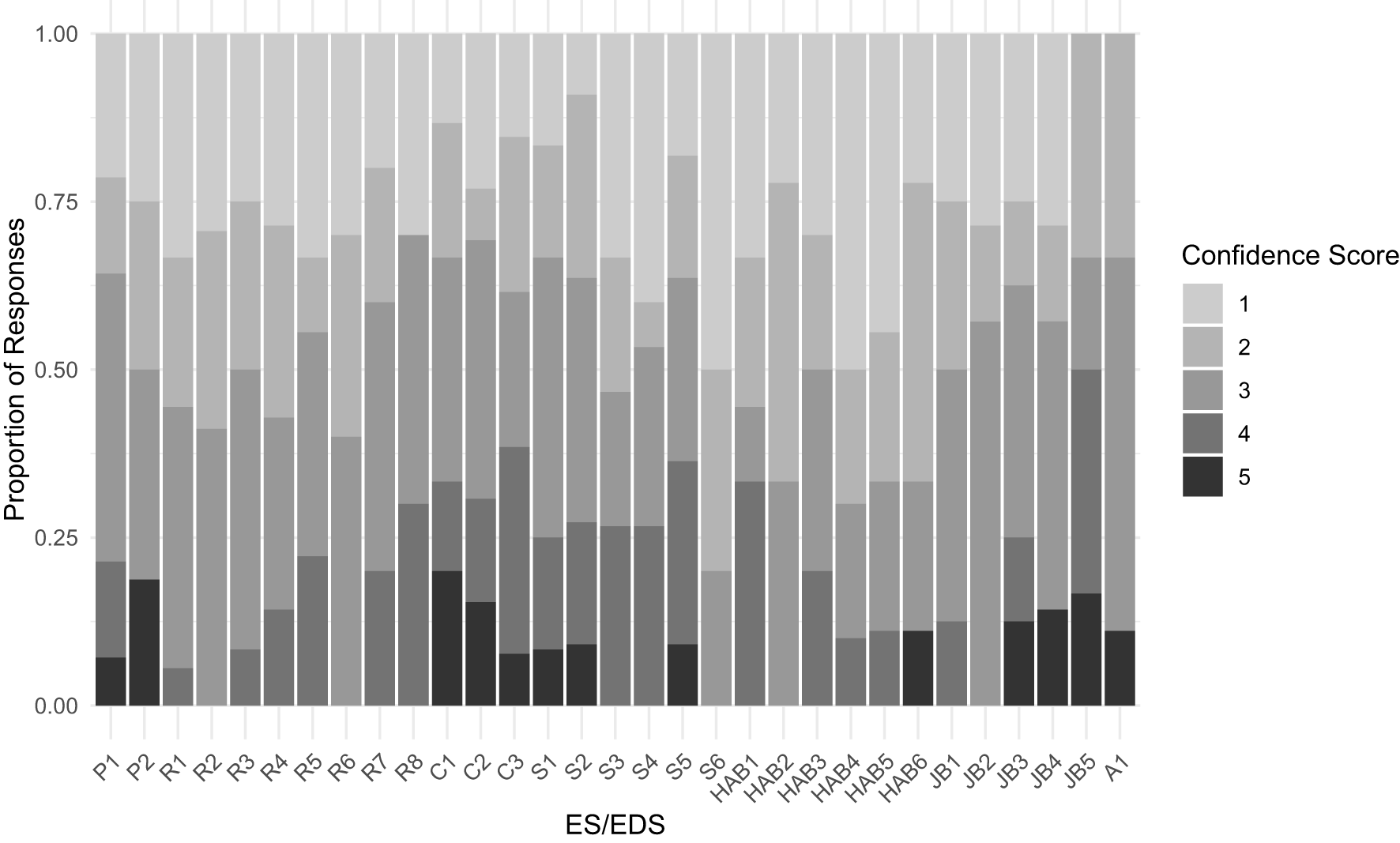
Proportions of confidence scores for each ES/EDS.
Explanatory comments added to the scores by the experts were helpful in interpreting the results, particularly in shedding light on differences between responses and identifying knowledge gaps. On average, plankton experts provided 11 explanatory comments, while other marine scientists added 8. Some overarching themes are discernible, which we outline here.
Many comments highlighted sources of uncertainty - attributed to distinct factors. Several scientists mentioned the importance of overall ocean biogeochemistry for plankton activity, which for the most part is independent of marine protection. Additionally, many factors are specific to localized conditions, such as concrete fishing activities and the presence of plankton predators. Besides differentiated local conditions, there is also scientific uncertainty regarding many issues. For example, a scientist pointed out a lack of scientific knowledge regarding the relationship between fish composition and plankton size and composition.
A major challenge in making estimates was the highly dynamic nature of plankton. Since area-based protection measures may not (or at least not in a predictable way) affect mobile organisms, this introduces additional uncertainty. Jellyfish, in particular, which are a long-lived subgroup of plankton, exhibit high mobility throughout their life cycle and are therefore potentially less affected by area-based measures. As one scientist noted, jellyfish born in winter in Tunisia can be observed along France’s Mediterranean coast in summer. A significant obstacle that persists in predicting jellyfish movement lies in accurately assessing their motion relative to the water velocity (Diamant et al., 2023). This may help to explain why jellyfish-related ES and EDS were assigned relatively low relevance scores.
Another challenge for providing estimates was the relative contribution of “active beneficiaries”. For extractive provisioning services as well as tourism, it is presupposed that there is some demand for those services. Even though we tried to somewhat circumvent this issue by labelling the quantitative indicators using “potential”, it is difficult to factor out the socio-economic dependencies. Several scientists voiced their doubts about demand for plankton extraction, especially in Europe. Another “active beneficiary” class of ES are those related to research. Here, the opposite seemed to be the case – many scientists deemed that there would be a mechanistically induced positive influence on plankton-related studies and plankton-based ecosystem health assessments, as MPAs are scientifically monitored and often serve as ecological baselines. This likely helps to explain why C1 and C2 received the highest estimated positive changes, as well as highest relevance and high confidence ratings. Nevertheless, one scientist noted that studies and ecosystem health assessments may not be focused on plankton, but on other metrics, for example related more to fish populations.
A second theme we identified concerns the observed differences in expected effects on regulating services, especially on the biological carbon pump. Most scientists expected null or slightly positive effects and comments were related to expected increases in primary production which would quasi-automatically lead to more carbon acquisition and sequestration. However, three scientists expected negative effects on at least one of ES related to the role of zooplankton in the biological carbon pump. One of them commented that they expected a larger population of plankton predators which would increase the share of carbon transferred to higher trophic levels instead of sequestered by plankton.
Finally, the interactions between protection outcomes and the different scales of protection were highlighted in the comments. Coastal macroalgae, which, as opposed to plankton, exist in geographically fixed areas, were mentioned as effective protection against HABs. On the other hand, a commenter remarked that the success of marine-based protection in coastal waters also depended on land-based protection and that, without a reduction in agriculture-related nutrient runoff, they would not expect noticeable effects on HAB-related negative outcomes.
4 Discussion
Marine plankton encompasses a vast diversity of species and ecological functions, and only recently have they been collectively considered in terms of their economic significance. As ecosystem services (ES), ecosystem disservices (EDS), and their economic values gain importance in marine policy-making, it is essential to incorporate plankton into these discussions.
Despite the growing acknowledgement of the economic importance of plankton, our review has shown that economic valuation of plankton ES in the scientific literature remains extremely scarce. Extractive uses of planktonic organisms, including jellyfish, krill, calanus and microalgae, generate revenues of several billion USD annually. As these markets are still emerging and remain underexplored in economic terms, existing literature focuses primarily on development pathways and potential rather than formal valuation studies. Regarding microalgae, Rahman (2020) points out that reliable data and statistics of market opportunities is very limited, making it difficult to assess their actual potential. The large-scale economic viability of microalgae-based lipids and hydrocarbons depends on the development of lower-cost, energy-efficient production and processing methods (Nethravathy et al., 2019; Ruiz et al., 2016; Rumin et al., 2020). For jellyfish, market potential depends critically on consumer demand. While the demand in China is well-established, market opportunities in Western countries are only beginning to be explored (Edelist et al., 2021). Shifts in consumer demand and cultural spillovers are increasingly shaping market dynamics (Bargain, 2024; Cao et al., 2024), so the untapped potential could be considerable.
The one ecosystem service (ES) that has received explicit valuation in the scientific literature is carbon sequestration. Cavan et al. (2024) estimated that carbon sequestered by Antarctic krill translates into an avoided social cost of carbon of several billion USD annually. Broader attempts to quantify the value of the biological carbon pump have produced highly variable results, with estimates ranging from under one trillion (Berzaghi et al., 2025) to 24 trillion USD per year (estimation by Nowicki et al., 2022, combined with the carbon prices used by Cavan et al., 2024). These discrepancies stem from three factors that drive estimate variability. Intrinsic factors are driven by ecosystem processes (Bisson et al., 2018) and include temporal and spatial variability in nutrient supply, phytoplankton growth, predator behavior and other ecological interactions as well as physical factors such as upwelling processes, which influence sequestration efficiency (DeVries et al., 2012). The second source of estimate variability is extrinsic and concerns sampling, data processing and modelling variations in biogeochemical and ecosystem models. Carbon export can, for instance, be measured using sediment traps that collect sinking particles (Ducklow et al., 2001). More recently, advances in satellite remote sensing have enabled estimates of carbon export from ocean surface data (Jönsson et al., 2023). However, these newer methods face specific challenges, particularly when it comes to inferring export beyond the euphotic zone (Siegel et al., 2023). The third source of variability in the estimation of the value of plankton carbon sequestration comes from differences in carbon prices. The social cost of carbon, reported in this article, is an approach that is based on the calculation of the economic damages associated with emitting one additional ton of CO2 into the atmosphere. To this end, Integrated Assessment Models (IAMs) are used to link economic activity to climate change and they exist in a large variety of specifications, ranging from general equilibrium models to simulations, focusing on the world as a whole or on individual countries or regions (Stern and Stiglitz, 2021). Another critical factor of variability within estimations of the social cost of carbon is the applied discount factor, i.e. the weighting of future costs and benefits relative to present ones. A considerable body of scholarly work has been dedicated to the debate on what constitutes an appropriate discount factor, founded on both epistemic and ethical grounds (Stern et al., 2006; Nordhaus, 2007). The aforementioned variations lead to estimated SSCO2 that range from USD–13.36/tCO2 to USD 2386.91/tCO2 (Wang et al., 2019). Besides this huge range within one conceptual approach, there are other carbon pricing strategies that also lead to varying estimates. Instead of quantifying cumulated costs of carbon emissions to societies, the marginal abatement cost approach focuses on CO2 reduction costs to emitters. Traditional economic theory proposes that the optimal level of abatement occurs where marginal abatement cost is equal to it marginal benefit, i.e. avoided SCCO2 (Clarkson and Deyes, 2002). Marginal abatement costs are sector-specific – an average value of 429 EUR/t CO2 (~ USD 485) has recently been estimated for the hard-to-abate European chemical sector considering the period 2015-2020 (Rekker et al., 2023). For the Italian energy sector, it was estimated that CO2 abatement was associated with costs of 165 EUR/t (~USD 230) when the abatement was realized by shifting energy production to wind power, and of 1000 EUR/t (~USD 1395) when it was realized by shifting to solar power, considering the period 2008-2011. Thus, while the immense value of the plankton-powered biological carbon pump is increasingly acknowledged, significant uncertainty remains regarding its economic magnitude.
Jean-Louis et al. (2025) found that surveyed individuals also stated significant WTP for planktonic carbon sequestration. Many plankton ES, especially those that can be classified as supporting and regulating services, have important but highly diffuse impacts on economies, and thus their economic use values are difficult to quantify. The finding by Jean-Louis et al. (2025) that stated preferences for a bundle of supporting services - subsumed under the attribute of ‘stable plankton composition’ - were the most highly valued suggests that people recognize and appreciate these values. Although supporting services are generally excluded from economic valuation to avoid double counting, regulating services like carbon sequestration are sometimes assigned a price, as seen with the social cost of carbon. When considering the full scope of the biological carbon pump and the social cost of carbon, this results in economic values of an almost unimaginable scale. Yet, the valuation of regulating services, as compared to provisioning and cultural services, poses specific challenges to evaluators. One needs to assess the capacity, demand and pressure for each regulating ES of interest (Sutherland et al., 2018), which can often fail due to a lack of quantifiable metrics, limiting in turn the inclusion of regulating ES in accounting frameworks and economic valuation. This difficulty is evident in our review of plankton ES valuation, where we found that only the regulating service of carbon sequestration has been assessed. Ecological-economic approaches have also been suggested to address the social costs of environmental issues such as eutrophication (Romstad, 2014), and in certain areas, nutrient trading schemes exist for nitrogen and phosphorus (Eger et al., 2023). Those emerging markets, combined with assessments of plankton’s nutrient cycling capacities, could be used in future research to economically value marine plankton’s water purification services, in order to more comprehensively account for the economic benefits that plankton provides. However, not only valuing, but also translating the economic valuation of plankton into actionable policies remains a challenge. The complexities of plankton dynamics, including its high spatial variability and mobility, complicate the design of policy instruments such as payments for ecosystem services (PES) or inclusion in blue carbon credit schemes. Addressing these gaps requires interdisciplinary collaboration between oceanographers, economists, and policymakers to develop robust methodologies for assessing and managing plankton ES at regional and global scales.
While plankton ES mostly contribute to human well-being in a general, often passive and/or unknown way, plankton EDS typically affect specific groups of people, such as fishers, beach goers or seafood consumers, in a more specific and discernible way. Thus, compared to ES, plankton EDS have been considerably more often the object of valuation, with a variety of applied methods. The economic dimensions of HAB and jellyfish bloom effects have been evaluated by means of survey data of affected industry (Haigh and Esenkulova, 2014; Palmieri et al., 2014), time-series economic models (Martino et al., 2020; Park et al., 2013), and stated-preference studies (Ghermandi et al., 2015; Ruiz-Frau, 2023). Their health dimensions have been economically valued based on public health costs (De Donno et al., 2014; Kouakou and Poder, 2019). Economic damages of several billion USD in total were reported in the studies we reviewed.
Notwithstanding the remaining knowledge gaps about the economic outcomes of plankton activities, our review has demonstrated its considerable economic significance. Thus, the question of how societies can foster its services and mitigate its disservices is of interest for economic values-based policy-making (Naselli-Flores and Padisak, 2023). An obvious theme of evaluation is area-based marine protection as a key tool for conservation and increasingly also considered for sustaining ecosystem services (Boulton et al., 2016; Leenhardt et al., 2015; Marcos et al., 2021; McDonald et al., 2020). In an ecosystem-based approach to MPA design, taking into account plankton indicators is crucial, as they form the basis of marine life (Benedetti et al., 2019). We suggest that including plankton ES and EDS indicators can be especially useful in view of a social-ecological systems approach, as they highlight the interconnectedness of the systems and add a new dimension to the more long-standing economics-based considerations represented by the fishing and tourism industries.
We developed an expert assessment tool for the evaluation of policies regarding their impact on plankton ES and EDS. Using this tool, 19 marine scientists - including 8 explicitly specialized in plankton - evaluated the anticipated effects of establishing a fully protected marine area in a previously unprotected European sea shelf zone. Besides cultural services related to research and monitoring, the experts saw an important potential of area-based marine protection to increase supporting services related to sustaining the marine food web and larvae recruitment for fish. Additionally, they expected important reductions in economic, ecological and health-related negative impacts of harmful algal blooms. Plankton specialists additionally assigned considerable potential to the services of water purification and biological control. However, some of the aforementioned ES and EDS received low median confidence scores, indicating that despite their possibly important potential for improvements, there is currently not enough scientific evidence to make reliable predictions – particular in relation to water purification and the mitigation of HAB-related EDS. Those results should therefore be interpreted with particular caution. Given the limited number of evaluations of plankton indicators in protected areas (Benedetti et al., 2019), it is not surprising that expert confidence remains relatively low in our assessment. We acknowledge this limitation and emphasize that the only durable way to increase confidence is through further research that systematically incorporates plankton indicators into protected area evaluations.
We note that experts whose work focuses explicitly on plankton provided slightly higher estimations for the potential of protection to increase ES and decrease EDS. In particular, HAB specialists had high expectations for area-based protection to reduce HAB-related disservices. This observation corroborates previous findings on the relationship between the level of expertise on ecosystem services and expectations regarding protection measures to preserve them (Peng et al., 2020; Prodanova and Varadzhakova, 2022). These findings suggest that domain-specific knowledge may shape perceptions, underscoring the importance of expert diversity in environmental decision-making processes. While the central tendencies do not hint to trade-offs between different ES/EDS, the directions of the expected changes were in most cases not unanimous, which indicates scientific uncertainty which at the very least does not exclude trade-offs.
One possible trade-off is the impact of MPAs on trophic cascades within the marine food web (Dimitriadis et al., 2021). By reducing fishing pressure and enhancing predator populations, MPAs may indirectly affect plankton community structure through altered grazing pressure. For example, an increase in filter-feeding fish populations could suppress certain planktonic species, potentially reducing primary production or disrupting existing biogeochemical cycles. Conversely, protecting large predators that control jellyfish populations might mitigate jellyfish blooms, an EDS that negatively impacts fisheries and tourism. Given these potential cascading effects, it is essential that future research evaluates the indirect ecological consequences of MPAs on planktonic ecosystems to inform holistic, multi-trophic conservation planning.
The use of expert consultation for ES/EDS assessments is a well-established practice (Campagne et al., 2017; 2018; Pham et al., 2025; Schernewski et al., 2018; Zaucha and Kreiner, 2021), appreciated for its efficiency, accessibility, and adaptability. Jacobs et al. (2015) suggest that such assessments can enhance collaboration between natural and social sciences, experts, stakeholders, and decision-makers. Expert assessments can be used throughout the entire policy cycle, e.g. for setting agendas, formulating policies, implementation and evaluation (Edelmann and Albrecht, 2023). An assessment of plankton ES and EDS, such as the one conducted in this study for a representative protected area, can help inform conservation agendas by highlighting these often overlooked values as policy priorities. It may help to designate areas for protection or compare different conservation approaches (some examples for alternatives to static area-based approaches are provided below). The assessment could furthermore be used to integrate ES/EDS information into plankton-based indicators for the evaluation of specific MPAs (c.f. Bryndum-Buchholz et al., 2023; Vassallo et al., 2021).
It should, however, be noted that the small sizes of expert panels may affect the universality of the results. Campagne et al. (2017) investigated the effect of expert panel size on ecosystem service capacity scores and found that standard errors decreased significantly for panels with more than 10 experts, but that the mean scores did not stabilize even with a comparatively large expert panel size of 30. Thus, we acknowledge that while our panel has a satisfactory size for an expert assessment, it remains a small absolute panel, which carries the risk of subjectivity in the assessments. Another limitation of our assessment is the lack of focus on a specific marine area, which likely introduces confounding effects. Nonetheless, we believe that the assessment is useful in drawing attention to the some well-established relationships between the reduction of acute anthropic pressures and valued plankton activities, as well as to scientific uncertainties.
One important source of uncertainty regarding protection outcomes on plankton and its services and disservices, as reported by the experts, is one of its most defining characteristics – its highly dynamic, drifting nature. Furthermore, the focus on area-based protection implies that only acute stressors like fishing, transport and other marine-based anthropic activities are removed, while chronic stressors such as climate change, ocean acidification and pollution originating from land, which significantly influence plankton (Botterell et al., 2023), are still present. However, such localized protection may have limited long-term impacts on population stability. This underscores the need for integrated conservation strategies that go beyond static MPAs.
While MPAs serve as key conservation tools, their static nature presents challenges for protecting highly dynamic and mobile organisms such as plankton. As plankton populations shift due to ocean currents, seasonal variations, and climate-driven changes, fixed MPA boundaries may fail to enhance their full ecological and economic value (Esteban-Cantillo et al., 2024; Game et al., 2009; United Nations Global Compact, 2024). The idea of dynamic protected areas has emerged in the scientific literature in the early 2000s (Agardy et al., 2003). The main idea is to take into account the natural dynamism of the ocean in area-based management tools by integrating data on target species or ecosystems and to adapt the spatial extent and location of protection measures according to their shifting distributions. Planktonic ecosystems, characterized by their high mobility and temporal variability, exemplify the need for such adaptive management but also pose significant data and monitoring challenges. However, recent technological advances are making this approach increasingly feasible. Digital tools are already contributing to marine sustainability in various domains, from decarbonizing the emission-intensive shipping sector (Xu et al., 2025; Xiao et al., 2025; Zou et al., 2025) to optimizing fishing efforts and reducing bycatch through predictive habitat modeling (Hazen et al., 2018). One of the most promising innovations is dynamic ocean management (DOM), a flexible, data-driven strategy that tailors conservation measures in response to real-time environmental conditions (McDonnell et al., 2024). For instance, Australia’s Great Barrier Reef Marine Park Authority employs adaptive zoning strategies that respond to ecosystem conditions in near real-time (Vella et al., 2024). The European Union’s Maritime Spatial Planning Directive has also facilitated adaptive fisheries management, integrating ecosystem-based conservation priorities into regulatory frameworks. At an international scale, the recently adopted Agreement on the Conservation and Sustainable Use of Marine Biological Diversity of Areas Beyond National Jurisdiction (BBNJ Agreement) has come to complement the United Nations Law of the Sea (UNCLOS). While the BBNJ Agreement does not explicitly mention dynamic protection measures, it gives way to protected areas with fluid boundaries, as long as the boundaries are defined at any given point in time (Klerk et al., 2024; Maxwell et al., 2020). Additionally, dynamic elements in the scope of the Agreement, such as the Scientific and Technical Body and the periodic monitoring and review of area-based management tools could facilitate the implementation of dynamic MPAs (ibid.). Applying these principles to plankton conservation could involve the use of AI-driven satellite monitoring to track plankton blooms, identify high-value carbon sequestration zones, and implement spatio-temporally flexible restrictions on industrial activities. Such adaptive conservation frameworks could enhance the effectiveness of marine policies in sustaining plankton ecosystem services while mitigating ecosystem disservices, such as harmful algal blooms.
Furthermore, complementary policies - such as strengthened land-based pollution controls and stricter emissions regulations are also essential to mitigate climate-driven and anthropogenic stressors that impact plankton communities globally. Future research should investigate whether adaptive and integrative conservation strategies can more effectively manage plankton ES and EDS compared to static MPAs. This could be assessed through comprehensive evaluations, incorporating expert assessment tools like the one presented in this article, alongside biophysical analyses and economic valuation of plankton indicators.
Statements
Data availability statement
The raw expert scoring data, codebook and R code for the analyses conducted can be retrieved here: https://osf.io/sptcr/.
Author contributions
EW: Conceptualization, Methodology, Writing – original draft, Writing – review & editing. OE-C: Investigation, Visualization, Writing – original draft, Writing – review & editing. SB-G: Supervision, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. EW, OE-C and SB-G acknowledge the BIOcean5D (marine biodiversity assessment and prediction across spatial, temporal and human scales, https://biocean5d.org/) project funded by the European Union’s under grant agreement No. 101059915.
Acknowledgments
We gratefully acknowledge the participation of the 19 BIOcean5D marine scientists who participated in the expert assessment.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Correction note
A correction has been made to this article. Details can be found at: 10.3389/fmars.2025.1715640.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Author disclaimer
Views and opinions expressed are however those of the authors only and do not necessarily reflect those of the European Union. Neither the European Union nor the granting authority can be held responsible for them.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2025.1607996/full#supplementary-material
References
1
Abate T. G. Nielsen R. Nielsen M. Drillet G. Jepsen P. M. Hansen B. W. (2015). Economic feasibility of copepod production for commercial use : Result from a prototype production facility. Aquaculture436, 72−79. doi: 10.1016/j.aquaculture.2014.10.012
2
Abdulrahman M. S. Harikrishnan S. Suvathika B. Sarath A. Arun Suthish U. Mjm Basha S. et al . (2023). Development of phytoplankton extract and their potential role on vegetable plant growth. Biochem. Cell. Arch.23, 789–794. doi: 10.51470/bca.2023.23.2.789
3
Abesamis R. A. Green A. L. Russ G. R. Jadloc C. R. L. (2014). The intrinsic vulnerability to fishing of coral reef fishes and their differential recovery in fishery closures. Rev. Fish. Biol. Fisheries24, 1033–1063. doi: 10.1007/s11160-014-9362-x
4
Agardy T. Bridgewater P. Crosby M. P. Day J. Dayton P. K. Kenchington R. et al . (2003). Dangerous targets? Unresolved issues and ideological clashes around marine protected areas. Aquat. conservation: Mar. Freshw. Ecosyst.13, 353–367. doi: 10.1002/aqc.583
5
Altieri A. H. Diaz R. J. (2019). “ Dead Zones : Oxygen Depletion in Coastal Ecosystems,” in World Seas : An Environmental Evaluation (Amsterdam: Elsevier), 453−473. doi: 10.1016/B978-0-12-805052-1.00021-8
6
Alvarez S. Brown C. E. Garcia Diaz M. O’Leary H. Solís D. (2024). Non-linear impacts of harmful algae blooms on the coastal tourism economy. J. Environ. Manage.351, 119811. doi: 10.1016/j.jenvman.2023.119811
7
Aminian-Biquet J. Gorjanc S. Sletten J. Vincent T. Laznya A. Vaidianu N. et al . (2024). Over 80% of the European Union’s marine protected area only marginally regulates human activities. One Earth7, 1614−1629. doi: 10.1016/j.oneear.2024.07.010
8
Anderson C. C. Metzemacher A. Adem Esmail B. (2025). Boxes of rain: A systematic review on the classes and frameworks of ecosystem disservices. Ambio, 1–19. doi: 10.1007/s13280-025-02157-1
9
Arkema K. K. Field L. Nelson L. K. Ban N. C. Gunn C. Lester S. E. (2024). Advancing the design and management of marine protected areas by quantifying the benefits of coastal ecosystems for communities. One Earth7, 989−1006. doi: 10.1016/j.oneear.2024.04.019
10
Arrigo K. R. (2005). Marine microorganisms and global nutrient cycles. Nature437, 349−355. doi: 10.1038/nature04159
11
Aurellado M. E. B. Ticzon V. S. Nañola C. L. Cabansag J. B. P. Bacabac M. M. A. Sorgon K. E. S. et al . (2021). Effectiveness of Philippine nationally managed marine reserves in improving biomass and trophic structure of coral reef fish communities. Coast. Manage.49, 293–312. doi: 10.1080/08920753.2021.1899944
12
Bargain O. (2024). Globalization and cultural spillover in trade: evidence from the Japanese food culture. Rev. World Economics160, 55–73. doi: 10.1007/s10290-023-00498-1
13
Barragán J. M. De Andrés M. (2015). Analysis and trends of the world’s coastal cities and agglomerations. Ocean Coast. Manage.114, 11–20. doi: 10.1016/j.ocecoaman.2015.06.004
14
B-Béres V. Stenger-Kovács C. Buczkó K. Padisák J. Selmeczy G. B. Lengyel E. et al . (2023). Ecosystem services provided by freshwater and marine diatoms. Hydrobiologia850, 2707−2733. doi: 10.1007/s10750-022-04984-9
15
Bechard A. (2020). Harmful algal blooms and tourism : the economic impact to counties in Southwest Florida. Rev. Regional Stud.50, 170–188. doi: 10.52324/001c.12705
16
Benedetti F. Jalabert L. Sourisseau M. Becker B. Cailliau C. Desnos C. et al . (2019). The seasonal and inter-annual fluctuations of plankton abundance and community structure in a north Atlantic marine protected area. Front. Mar. Sci.6. doi: 10.3389/fmars.2019.00214
17
Berdalet E. Fleming L. E. Gowen R. Davidson K. Hess P. Backer L. C. et al . (2016). Marine harmful algal blooms, human health and wellbeing : Challenges and opportunities in the 21st century. J. Mar. Biol. Assoc. United Kingdom96, 61−91. doi: 10.1017/S0025315415001733
18
Berzaghi F. Pinti J. Aumont O. Maury O. Cosimano T. Wisz M. S. (2025). Global distribution, quantification, and valuation of the biological carbon pump. Nat. Climate Change15, 385–392. doi: 10.1038/s41558-025-02295-0
19
Bezares Calderón L. A. Shahidi R. Jékely G. (2024). Mechanism of barotaxis in marine zooplankton. eLife13, RP94306. doi: 10.7554/eLife.94306
20
Bisson K. M. Siegel D. A. DeVries T. Cael B. B. Buesseler K. O. (2018). How data set characteristics influence ocean carbon export models. Global Biogeochemical Cycles32, 1312–1328. doi: 10.1029/2018GB005934
21
Boldrocchi G. Villa B. Monticelli D. Spanu D. Magni G. Pachner J. et al . (2023). Zooplankton as an indicator of the status of contamination of the Mediterranean Sea and temporal trends. Mar. pollut. Bull.197, 115732. doi: 10.1016/j.marpolbul.2023.115732
22
Bosch-Belmar M. Milisenda G. Basso L. Doyle T. K. Leone A. Piraino S. (2021). Jellyfish impacts on marine aquaculture and fisheries. Rev. Fisheries Sci. Aquaculture29, 242−259. doi: 10.1080/23308249.2020.1806201
23
Bostrom K. Joensuu O. Brohm I. (1974). Plankton : Its chemical composition and its significance as a source of pelagic sediments. Chem. Geol.14, 255−271. doi: 10.1016/0009-2541(74)90063-1
24
Botterell Z. L. R. Lindeque P. K. Thompson R. C. Beaumont N. J. (2023). An assessment of the ecosystem services of marine zooplankton and the key threats to their provision. Ecosystem Serv.63, 101542. doi: 10.1016/j.ecoser.2023.101542
25
Boulton A. J. Ekebom J. Gíslason G. M. (2016). Integrating ecosystem services into conservation strategies for freshwater and marine habitats: a review. Aquat. Conservation: Mar. Freshw. Ecosyst.26, 963–985. doi: 10.1002/aqc.2703
26
Brown A. R. Lilley M. Shutler J. Lowe C. Artioli Y. Torres R. et al . (2020). Assessing risks and mitigating impacts of harmful algal blooms on mariculture and marine fisheries. Rev. Aquaculture12, 1663−1688. doi: 10.1111/raq.12403
27
Brun V. De Ville d’Avray L. T. Ma. M. Dolorosa R. G. Mecha N. J. M. F. Sorgon K. E. S. et al . (2024). Baseline assessment and early effects of a network of marine protected areas. Conserv. Sci. Pract.6, e13121. doi: 10.1111/csp2.13121
28
Bryndum-Buchholz A. Blanchard J. L. Coll M. Pontavice H. D. Everett J. D. Guiet J. et al . (2023). Applying ensemble ecosystem model projections to future-proof marine conservation planning in the Northwest Atlantic Ocean. Facets8, 1–16. doi: 10.1139/facets-2023-0024
29
Calbet A. (2024). Plankton in a Changing World : The Impact of Global Change on Marine Ecosystems (Switzerland: Springer Nature). doi: 10.1007/978-3-031-76121-8
30
Campagne C. S. Roche P. Gosselin F. Tschanz L. Tatoni T. (2017). Expert-based ecosystem services capacity matrices : Dealing with scoring variability. Ecol. Indic.79, 63−72. doi: 10.1016/j.ecolind.2017.03.043
31
Campagne C. S. Roche P. K. Salles J.-M. (2018). Looking into Pandora’s Box : Ecosystem disservices assessment and correlations with ecosystem services. Ecosystem Serv.30, 126−136. doi: 10.1016/j.ecoser.2018.02.005
32
Campagne C. S. Salles J.-M. Boissery P. Deter J. (2015). The seagrass Posidonia oceanica : Ecosystem services identification and economic evaluation of goods and benefits. Mar. pollut. Bull.97, 391−400. doi: 10.1016/j.marpolbul.2015.05.061
33
Cao M. Jin Y. Hu Y. Huang X. Wu J. (2024). A decision framework for Chinese-style cruise ship design based on informativeness weight method and group consensus reaching model. Advanced Eng. Inf.62, 102698. doi: 10.1016/j.aei.2024.102698
34
Carias J. Vásquez-Lavín F. Barrientos M. Ponce Oliva R. D. Gelcich S. (2024). Economic valuation of Harmful Algal Blooms (HAB) : Methodological challenges, policy implications, and an empirical application. J. Environ. Manage.365, 121566. doi: 10.1016/j.jenvman.2024.121566
35
Cavan E. L. Mackay N. Hill S. L. Atkinson A. Belcher A. Visser A. (2024). Antarctic krill sequester similar amounts of carbon to key coastal blue carbon habitats. Nat. Commun.15, 7842. doi: 10.1038/s41467-024-52135-6
36
Centers for Disease Control and Prevention (CDC) (2024). Summary Report – One Health Harmful Algal Bloom System (OHHABS), United States 2022 (Atlanta, Georgia: U.S. Department of Health and Human Services, CDC).
37
Cermeño P. Marañín E. Harbour D. Figueiras F. G. Crespo B. G. Huete-Ortega M. et al . (2008). Resource levels, allometric scaling of population abundance, and marine phytoplankton diversity. Limnol. Oceanogr.53, 312–318. doi: 10.4319/lo.2008.53.1.0312
38
Chapman E. J. Byron C. J. Lasley-Rasher R. Lipsky C. Stevens J. R. Peters R. (2020). Effects of climate change on coastal ecosystem food webs: Implications for aquaculture. Mar. Environ. Res.162, 105103. doi: 10.1016/j.marenvres.2020.105103
39
Charlson R. J. Lovelock J. E. Andreae M. O. Warren S. G. (1987). Oceanic phytoplankton, atmospheric sulphur, cloud albedo and climate. Nature326, 655−661. doi: 10.1038/326655a0
40
Chevrollier L. A. Cook J. M. Halbach L. Jakobsen H. Benning L. G. Anesio A. M. et al . (2023). Light absorption and albedo reduction by pigmented microalgae on snow and ice. J. Glaciol.69, 333–341. doi: 10.1017/jog.2022.64
41
Clarkson R. Deyes K. (2002). Estimating the social cost of carbon emissions (London: HM Treasury).
42
Claustré H. Legendre L. Boyd P. W. Levy M. (2021). The oceans’ biological carbon pumps: framework for a research observational community approach. Front. Mar. Sci.8, 780052. doi: 10.3389/fmars.2021.780052
43
Clinton M. Ferrier D. E. K. Martin S. A. M. Brierley A. S. (2021). Impacts of jellyfish on marine cage aquaculture : An overview of existing knowledge and the challenges to finfish health. ICES J. Mar. Sci.78, 1557−1573. doi: 10.1093/icesjms/fsaa254
44
Coll M. Piroddi C. Steenbeek J. Kaschner K. Ben Rais Lasram F. Aguzzi J. et al . (2010). The biodiversity of the Mediterranean Sea: estimates, patterns, and threats. PloS One5, e11842. doi: 10.1371/journal.pone.0011842
45
Costanza R. d’Arge R. De Groot R. Farber S. Grasso M. Hannon B. et al . (1997). The value of the world’s ecosystem services and natural capital. Nature387, 253–260. doi: 10.1038/387253a0
46
Costanza R. Daly H. E. (1992). Natural capital and sustainable development. Conserv. Biol.6, 37−46. doi: 10.1046/j.1523-1739.1992.610037.x
47
Day J. Dudley N. Hockings M. Holmes G. Laffoley D. Stolton S. et al . (2019). Guidelines for applying the IUCN protected area management categories to marine protected areas. 2nd ed. (Gland, Switzerland: IUCN).
48
De Donno A. Idolo A. Bagordo F. Grassi T. Leomanni A. Serio F. et al . (2014). Impact of stinging jellyfish proliferations along south Italian coasts : human health hazards, treatment and social costs. Int. J. Environ. Res. Public Health11, 2488−2503. doi: 10.3390/ijerph110302488
49
DeVries T. Primeau F. Deutsch C. (2012). The sequestration efficiency of the biological pump. Geophysical Res. Lett.39, L13601. doi: 10.1029/2012GL051963
50
Diamant R. Alexandri T. Barak N. Lotan T. (2023). A remote sensing approach for exploring the dynamics of jellyfish, relative to the water current. Sci. Rep.13, 14769. doi: 10.1038/s41598-023-41655-8
51
Dimitriadis C. Fournari-Konstantinidou I. Sourbès L. Koutsoubas D. Katsanevakis S. (2021). Long term interactions of native and invasive species in a marine protected area suggest complex cascading effects challenging conservation outcomes. Diversity13, 71. doi: 10.3390/d13020071
52
Djoru M. R. Gimin R. (2020). “ Phytoplankton (microalgae) as an alternative of renewable energy sources,” in IOP Conference Series: Materials Science and Engineering, Vol. 823. (Bristol, UK: IOP Publishing). doi: 10.1088/1757-899X/823/1/012019
53
Doyle T. K. Hays G. C. Harrod C. Houghton J. D. R. (2014). “ Ecological and Societal Benefits of Jellyfish,” in Jellyfish Blooms. Eds. PittK. A.LucasC. H., (New York, NY: Springer) 105−127.
54
Duarte I. M. Marques S. C. Leandro S. M. Calado R. (2022). An overview of jellyfish aquaculture : For food, feed, pharma and fun. Rev. Aquaculture14, 265−287. doi: 10.1111/raq.12597
55
Ducklow H. W. Steinberg D. K. Buesseler K. O. (2001). Upper ocean carbon export and the biological pump. Oceanography14, 50–58. doi: 10.5670/oceanog.2001.06
56
Duffy J. (2006). Biodiversity and the functioning of seagrass ecosystems. Mar. Ecol. Prog. Ser.311, 233–250. doi: 10.3354/meps311233
57
Eastwood A. Brooker R. Irvine R. J. Artz R. R. E. Norton L. R. Bullock J. M. et al . (2016). Does nature conservation enhance ecosystem services delivery? Ecosystem Serv.17, 152−162. doi: 10.1016/j.ecoser.2015.12.001
58
Edelist D. Angel D. L. Canning-Clode J. Gueroun S. K. M. Aberle N. Javidpour J. et al . (2021). Jellyfishing in Europe : current status, knowledge gaps, and future directions towards a sustainable practice. Sustainability13, 12445. doi: 10.3390/su132212445
59
Edelmann N. Albrecht V. (2023). The Policy Cycle: a framework for knowledge management of practitioners’ expertise and role in participatory processes. Front. Political Sci.5, 1223013. doi: 10.3389/fpos.2023.1223013
60
Eger A. M. Marzinelli E. M. Beas-Luna R. Blain C. O. Blamey L. K. Byrnes J. E. et al . (2023). The value of ecosystem services in global marine kelp forests. Nat. Commun.14, 1894. doi: 10.1038/s41467-023-37385-0
61
EPA (U.S. Environmental Protection Agency) (2024). Greenhouse gas equivalencies calculator. Available online at: https://www.epa.gov/energy/greenhouse-gas-equivalencies-calculator (Accessed February 14, 2025).
62
Esteban-Cantillo O. J. Abreu A. Bourgeois-Gironde S. Wanek E. Gurchani U. Eveillard D. et al . (2024). Dynamic Oceans, Dynamic Solutions (Paris, France: Policy Brief).
63
European Commission (2022). Annual report on the efforts made by Italy in 2022 to reach a sustainable balance between fishing capacity and fishing opportunities (in accordance with Article 22 of Regulation (EU) No 1380/2013) (Brussels, Belgium: European Union).
64
European Commission Directorate General for Maritime Affairs and Fisheries European Commission. Joint Research Centre (2021). The EU blue economy report 2021 ( Publications Office). Available online at: https://data.europa.eu/doi/10.2771/8217 (Accessed January 28, 2025).
65
Eurostat (2025). Fish catches in the Northeast Atlantic (Custom extraction 15172260). Available online at: https://ec.europa.eu/eurostat/databrowser/view/fish_ca_atl37:custom_15172260/default/table?lang=en (Accessed February 14, 2025).
66
Falkowski P. (2012). Ocean Science: The power of plankton. Nature483, S17–S20. doi: 10.1038/483S17a
67
FAO (2024). Fishery and Aquaculture Statistics – Yearbook 2021. (Rome, Italy: Food and Agriculture Organization of the United Nations). doi: 10.4060/cc9523en
68
Fenner P. J. Lippmann J. Gershwin L. (2010). Fatal and nonfatal severe jellyfish stings in Thai waters. J. Travel Med.17, 133−138. doi: 10.1111/j.1708-8305.2009.00390.x
69
Fernández-Alías A. Marcos C. Pérez-Ruzafa A. (2024). The unpredictability of scyphozoan jellyfish blooms. Front. Mar. Sci.11. doi: 10.3389/fmars.2024.1349956
70
Frouin R. Iacobellis S. F. (2002). Influence of phytoplankton on the global radiation budget. J. Geophysical Research: Atmospheres107, 4377. doi: 10.1029/2001JD000562
71
Game E. T. Bode M. McDonald-Madden E. Grantham H. S. Possingham H. P. (2009). Dynamic marine protected areas can improve the resilience of coral reef systems. Ecol. Lett.12, 1336–1346. doi: 10.1111/j.1461-0248.2009.01384.x
72
Gantar M. Svirčev Z. (2008). MICROALGAE AND CYANOBACTERIA : FOOD FOR THOUGHT. J. Phycol.44, 260−268. doi: 10.1111/j.1529-8817.2008.00469.x
73
Ghermandi A. Galil B. Gowdy J. Nunes P. A. L. D. (2015). Jellyfish outbreak impacts on recreation in the Mediterranean Sea : Welfare estimates from a socioeconomic pilot survey in Israel. Ecosystem Serv.11, 140−147. doi: 10.1016/j.ecoser.2014.12.004
74
Graham W. M. Gelcich S. Robinson K. L. Duarte C. M. Brotz L. Purcell J. E. et al . (2014). Linking human well-being and jellyfish : Ecosystem services, impacts, and societal responses. Front. Ecol. Environ.12, 515−523. doi: 10.1890/130298
75
Graham W. M. Martin D. L. Felder D. L. Asper V. L. Perry H. M. (2003). “ Ecological and economic implications of a tropical jellyfish invader in the Gulf of Mexico,” in Marine Bioinvasions : Patterns, Processes and Perspectives. Ed. PedersonJ. ( Springer, Netherlands), 53−69. doi: 10.1007/978-94-010-0169-4_6
76
Grand View Research (2021). Krill oil market size, share & trends analysis report [Market research report]. Available online at: https://www.grandviewresearch.com/industry-analysis/krill-oil-market (Accessed January 22, 2025).
77
Grattan L. M. Holobaugh S. Morris J. G. (2016). Harmful algal blooms and public health. Harmful Algae57, 2−8. doi: 10.1016/j.hal.2016.05.003
78
Green T. K. Hatton A. D. (2014). “ The Claw Hypothesis : A New Perspective on the Role of Biogenic Sulphur in the Regulation of Global Climate,” in Oceanography and Marine Biology. Eds. HughesR. N.HughesD. J.SmithI. P. (Boca Raton, FL: CRC Press), 315−336. doi: 10.1201/b17143-7
79
Guo R.-Z. Song Y.-B. Dong M. (2022). Progress and prospects of ecosystem disservices : an updated literature review. Sustainability14, 10396. doi: 10.3390/su141610396
80
Haddock S. H. D. Moline M. A. Case J. F. (2009). Bioluminescence in the Sea. (Cambridge, MA: Harvard University Press).
81
Haddock S. H. Moline M. A. Case J. F. (2010). Bioluminescence in the Sea. Annu. Rev. Mar. Sci.2, 443–493. doi: 10.1146/annurev-marine-120308-081028
82
Haigh N. Esenkulova S. (2014). Economic losses to the British Columbia salmon aquaculture industry due to harmful algal blooms 2009-2012. PICES Sci. Rep.47), 2.
83
Haines-Young R. Potschin M. (2010). “ The links between biodiversity, ecosystem services and human well-being,” in Ecosystem Ecology, Eds. RaffaelliD. G.FridC. L. J. (Cambridge: Cambridge University Press), 110−139. doi: 10.1017/CBO9780511750458.007
84
Haines-Young R. Potschin-Young M. (2018). Common International Classification of Ecosystem Services (CICES) V5.1 and guidance on the application of the revised structure ( European Environment Agency). Available online at: https://cices.eu (Accessed February 12, 2025).
85
Hayward E. (2012). Sensational jellyfish : aquarium affects and the matter of immersion. Differences23, 161−196. doi: 10.1215/10407391-1892925
86
Hazen E. L. Scales K. L. Maxwell S. M. Briscoe D. K. Welch H. Bograd S. J. et al . (2018). A dynamic ocean management tool to reduce bycatch and support sustainable fisheries. Sci. Adv.4, eaar3001. doi: 10.1126/sciadv.aar3001
87
Hofmann E. E. Murphy E. J. (2004). Advection, krill, and Antarctic marine ecosystems. Antarctic Sci.16, 487499. doi: 10.1017/S0954102004002275
88
Jacobs S. Burkhard B. Van Daele T. Staes J. Schneiders A. (2015). ‘The Matrix Reloaded’ : A review of expert knowledge use for mapping ecosystem services. Ecol. Model.295, 21−30. doi: 10.1016/j.ecolmodel.2014.08.024
89
Jean-Louis G. Massenberg J. Bartkowski B. (2025). Data on Europe-wide public preferences for plankton-based ecosystem services and marine biodiversity from a series of Deliberative Monetary Valuation workshops. Data Brief forthcoming60, 111488. doi: 10.1016/j.dib.2025.111488
90
Jönsson B. F. Kulk G. Sathyendranath S. (2023). Review of algorithms estimating export production from satellite derived properties. Front. Mar. Sci.10, 1149938. doi: 10.3389/fmars.2023.1149938
91
Karakuş O. Völker C. Iversen M. Hagen W. Hauck J. (2022). The role of zooplankton grazing and nutrient recycling for global ocean biogeochemistry and phytoplankton phenology. J. Geophysical Research: Biogeosciences127, e2022JG006798. doi: 10.1029/2022JG006798
92
Karlson B. Andersen P. Arneborg L. Cembella A. Eikrem W. John U. et al . (2021). Harmful algal blooms and their effects in coastal seas of Northern Europe. Harmful Algae102, 101989. doi: 10.1016/j.hal.2021.101989
93
Kennerley A. Wood L. E. Luisetti T. Ferrini S. Lorenzoni I. (2022). Economic impacts of jellyfish blooms on coastal recreation in a UK coastal town and potential management options. Ocean Coast. Manage.227, 106284. doi: 10.1016/j.ocecoaman.2022.106284
94
Klerk B. E. Heinrich K. Primicerio R. (2024). Beyond equilibrium thinking: dynamic area-based management tools in a changing ocean. Front. Mar. Sci.11, 1418435. doi: 10.3389/fmars.2024.1418435
95
Kouakou C. R. C. Poder T. G. (2019). Economic impact of harmful algal blooms on human health : A systematic review. J. Water Health17, 499−516. doi: 10.2166/wh.2019.064
96
Kourantidou M. Jin D. Schumacker E. J. (2022). Socioeconomic disruptions of harmful algal blooms in indigenous communities : The case of Quinault Indian nation. Harmful Algae118, 102316. doi: 10.1016/j.hal.2022.102316
97
Kwantes J. M. Grundmann O. (2015). A brief review of krill oil history, research, and the commercial market. J. Dietary Suppl.12, 23−35. doi: 10.3109/19390211.2014.902000
98
Lakkis N. A. Maalouf G. J. Mahmassani D. M. (2015). Jellyfish stings : A practical approach. Wilderness Environ. Med.26, 422−429. doi: 10.1016/j.wem.2015.01.003
99
Landsberg J. H. (2002). The effects of harmful algal blooms on aquatic organisms. Rev. Fisheries Sci.10, 113−390. doi: 10.1080/20026491051695
100
Leenhardt P. Low N. Pascal N. Micheli F. Claudet J. (2015). “ The Role of Marine Protected Areas in Providing Ecosystem Services,” in Aquatic Functional Biodiversity (Amsterdam: Elsevier). doi: 10.1016/B978-0-12-417015-5.00009-8
101
Lenihan H. S. Gallagher J. P. Peters J. R. Stier A. C. Hofmeister J. K. K. Reed D. C. (2021). Evidence that spillover from Marine Protected Areas benefits the spiny lobster (Panulirus interruptus) fishery in southern California. Sci. Rep.11, 2663. doi: 10.1038/s41598-021-82371-5
102
Li Y. Cheng D. Adam N. A. Mirzalevens S. Zhang G. (2025). Exploring the nexus between coastal tourism growth and eutrophication: Challenges for environmental management. Mar. pollut. Bull.216, 117922. doi: 10.1016/j.marpolbul.2025.117922
103
Lloret J. Cowx I. G. Cabral H. Castro M. Font T. Gonçalves J. M. S. et al . (2018). Small-scale coastal fisheries in European Seas are not what they were : Ecological, social and economic changes. Mar. Policy98, 176−186. doi: 10.1016/j.marpol.2016.11.007
104
Loke Show P. (2022). Global market and economic analysis of microalgae technology : Status and perspectives. Bioresource Technol.357, 127329. doi: 10.1016/j.biortech.2022.127329
105
Lomartire S. Marques J. C. Gonçalves A. M. M. (2021). The key role of zooplankton in ecosystem services : A perspective of interaction between zooplankton and fish recruitment. Ecol. Indic.129, 107867. doi: 10.1016/j.ecolind.2021.107867
106
López-Pacheco I. Y. Rodas-Zuluaga L. I. Cuellar-Bermudez S. P. Hidalgo-Vázquez E. Molina-Vazquez A. Araújo R. G. et al . (2022). Revalorization of microalgae biomass for synergistic interaction and sustainable applications : bioplastic generation. Mar. Drugs20, 601. doi: 10.3390/md20100601
107
Lucas C. H. Gelcich S. Uye S.-I. (2014). “ Living with Jellyfish : Management and Adaptation Strategies,” in Jellyfish Blooms. Eds. PittK. A.LucasC. H. ( Springer, Netherlands), 129−150. doi: 10.1007/978-94-007-7015-7_6
108
Luck G. W. Chan K. M. Klien C. J. (2012). Identifying spatial priorities for protecting ecosystem services. F1000Research1, 17. doi: 10.12688/f1000research.1-17.v1
109
Malone T. C. Newton A. (2020). The globalization of cultural eutrophication in the coastal ocean: causes and consequences. Front. Mar. Sci.7. doi: 10.3389/fmars.2020.00670
110
Marcos C. Díaz D. Fietz K. Forcada A. Ford A. García-Charton J. A. et al . (2021). Reviewing the ecosystem services, societal goods, and benefits of marine protected areas. Front. Mar. Sci.8, 613819. doi: 10.3389/fmars.2021.613819
111
Mardones J. I. Holland D. S. Anderson L. Le Bihan V. Gianella F. Clément A. et al . (2020). “ Estimating and mitigating the economic costs of harmful algal blooms on commercial and recreational shellfish harvesters,” in PICES Scientific Report No. 59. Global HAB: Evaluating, reducing, and mitigating the cost of harmful algal blooms: A compendium of case studies. (Victoria, BC, Canada: North Pacific Marine Science Organization - PICES), 66–83.
112
Marriott S. E. Cox C. Amolo R. C. Apistar D. Mancao R. H. De Mutsert K. (2021). Implications of community-based management of marine reserves in the Philippines for reef fish communities and biodiversity. Front. Mar. Sci.8. doi: 10.3389/fmars.2021.731675
113
Martino S. Gianella F. Davidson K. (2020). An approach for evaluating the economic impacts of harmful algal blooms : The effects of blooms of toxic Dinophysis spp. on the productivity of Scottish shellfish farms. Harmful Algae99, 101912. doi: 10.1016/j.hal.2020.101912
114
Masilamoni J. G. Jesudoss K. S. Nandakumar K. Satpathy K. K. Nair K. V. K. Azariah J. (2000). Jellyfish ingress: a threat to the smooth operation of coastal power plants. Curr. Sci.79, 567–569.
115
Maxwell S. M. Gjerde K. M. Conners M. G. Crowder L. B. (2020). Mobile protected areas for biodiversity on the high seas. Science367, 252–254. doi: 10.1126/science.aaz9327
116
McDonald H. Hoffman H. Ressurreição A. Röschel L. Gerdes H. Lago M. et al . (2020). “ Ecosystem-based management for more effective and equitable marine protected areas: A case study on the Faial-Pico channel marine protected area, Azores,” in Ecosystem-based management, ecosystem services and aquatic biodiversity: Theory, tools and applications. (Cham, Switzerland: Springer), 499–515.
117
McDonnell L. H. Hazen E. L. Mach K. J. (2024). Characteristics of animal movement and environmental data for dynamic ocean management: Insights and guidance. Conserv. Sci. Pract.6, e13268. doi: 10.1111/csp2.13268
118
Mejjad N. Rossi A. Pavel A. B. (2022). The coastal tourism industry in the Mediterranean: A critical review of the socio-economic and environmental pressures & impacts. Tourism Manage. Perspect.44, 101007. doi: 10.1016/j.tmp.2022.101007
119
Millennium Ecosystem Assessment (2005). Ecosystems and Human Well-being: Synthesis (Washington, DC: Island Press).
120
Morse-Jones S. Luisetti T. Turner R. K. Fisher B. (2011). Ecosystem valuation: some principles and a partial application. Environmetrics22, 675–685. doi: 10.1002/env.1073
121
Naselli-Flores L. Barone R. (2011). Invited review—Fight on plankton ! Or, phytoplankton shape and size as adaptive tools to get ahead in the struggle for life. Cryptogamie Algologie32, 157−204. doi: 10.7872/crya.v32.iss2.2011.157
122
Naselli-Flores L. Padisák J. (2023). Ecosystem services provided by marine and freshwater phytoplankton. Hydrobiologia850, 2691−2706. doi: 10.1007/s10750-022-04795-y
123
Nethravathy M. U. Mehar J. G. Mudliar S. N. Shekh A. Y. (2019). Recent advances in microalgal bioactives for food, feed, and healthcare products: commercial potential, market space, and sustainability. Compr. Rev. Food Sci. Food Saf.18, 1882–1897. doi: 10.1111/1541-4337.12500
124
Nordhaus W. D. (2007). A review of the Stern review on the economics of climate change. J. economic literature45, 686–702. doi: 10.1257/jel.45.3.686
125
Nowicki M. DeVries T. Siegel D. A. (2022). Quantifying the carbon export and sequestration pathways of the ocean’s biological carbon pump. Global Biogeochemical Cycles36, e2021GB007083. doi: 10.1029/2021GB007083
126
Nunes P. A. L. D. Loureiro M. L. Piñol L. Sastre S. Voltaire L. Canepa A. (2015). Analyzing beach recreationists’ Preferences for the reduction of jellyfish blooms : economic results from a stated-choice experiment in Catalonia, Spain. PloS One10, e0126681. doi: 10.1371/journal.pone.0126681
127
Nwankwegu A. S. Li Y. Huang Y. Wei J. Norgbey E. Sarpong L. et al . (2019). Harmful algal blooms under changing climate and constantly increasing anthropogenic actions : The review of management implications. 3 Biotech.9, 449. doi: 10.1007/s13205-019-1976-1
128
OSPAR (2017). Summary status of the changes in phytoplankton biomass and zooplankton abundance. Intermediate assessment 2017. Available online at: https://oap.ospar.org/en/ospar-assessments/intermediate-assessment-2017/biodiversity-status/habitats/plankton-biomass/ (Accessed January 22, 2025).
129
OSPAR (2023). “ Pelagic habitats thematic assessment,” in . In: OSPAR 2023: quality status report 2023 (London: OSPAR Commission). Available online at: https://oap.ospar.org/en/ospar-assessments/quality-status-reports/qsr-2023/thematic-assessments/pelagic-habitats/ (Accessed March 17, 2025).
130
Osseni A. F. Bareille F. Dupraz P. (2021). Hedonic valuation of harmful algal bloom pollution : Why econometrics matters? Land Use Policy107, 104283. doi: 10.1016/j.landusepol.2019.104283
131
Pagels F. Pereira R. N. Vicente A. A. Guedes A. C. (2021). Extraction of pigments from microalgae and cyanobacteria—A review on current methodologies. Appl. Sci.11, 5187. doi: 10.3390/app11115187
132
Palmieri M. G. Barausse A. Luisetti T. Turner K. (2014). Jellyfish blooms in the Northern Adriatic Sea : Fishermen’s perceptions and economic impacts on fisheries. Fisheries Res.155, 51−58. doi: 10.1016/j.fishres.2014.02.021
133
Palomares M. L. D. Froese R. Derrick B. Meeuwig J. J. Nöel S.-L. Tsui G. et al . (2020). Fishery biomass trends of exploited fish populations in marine ecoregions, climatic zones and ocean basins. Estuarine Coast. Shelf Sci.243, 106896. doi: 10.1016/j.ecss.2020.106896
134
Park J. Jeong H. J. Yoo Y. D. Yoon E. Y. (2013). Mixotrophic dinoflagellate red tides in Korean waters : Distribution and ecophysiology. Harmful Algae30, S28−S40. doi: 10.1016/j.hal.2013.10.004
135
Patara L. Vichi M. Masina S. Fogli P. G. Manzini E. (2012). Global response to solar radiation absorbed by phytoplankton in a coupled climate model. Climate Dynamics39, 1951−1968. doi: 10.1007/s00382-012-1300-9
136
Peng L. C. Lien W. Y. Lin Y. P. (2020). How experts’ Opinions and knowledge affect their willingness to pay for and ranking of hydrological ecosystem services. Sustainability12, 10055. doi: 10.3390/su122310055
137
Pham T. T. T. Armstrong C. W. Solvang H. K. Skern-Mauritzen M. (2025). Risk assessment to Barents Sea ecosystem services : An expert-based approach. Front. Mar. Sci.12. doi: 10.3389/fmars.2025.1493592
138
Prakash D. Kumar R. Rajan K. Patel A. Yadav D. K. Dhanker R. et al . (2022). Integrated application of macrophytes and zooplankton for wastewater treatment. Front. Environ. Sci.10. doi: 10.3389/fenvs.2022.941841
139
Prodanova H. Varadzhakova D. (2022). How individual scores affect the final expert-based assessments of ecosystem services: Range and mean scores analysis of natural heritage supply maps. Eur. J. Geogr.13, 074–097. doi: 10.48088/ejg.h.pro.13.4.074.097
140
Rahman K. M. (2020). “ Food and High Value Products from Microalgae: Market Opportunities and Challenges,” in Microalgae Biotechnology for Food, Health and High Value Products. Eds. AlamM.XuJ. L.WangZ. ( Springer, Singapore). doi: 10.1007/978-981-15-0169-2_1
141
Raposo A. Alasqah I. Alfheeaid H. Alsharari Z. Alturki H. Raheem D. (2022). Jellyfish as food : A narrative review. Foods11, 2773. doi: 10.3390/foods11182773
142
Reid P. (2000). Impacts of fisheries on plankton community structure. ICES J. Mar. Sci.57, 495–502. doi: 10.1006/jmsc.2000.0740
143
Rekker L. Kesina M. Mulder M. (2023). Carbon abatement in the European chemical industry: assessing the feasibility of abatement technologies by estimating firm-level marginal abatement costs. Energy Economics126, 106889. doi: 10.1016/j.eneco.2023.106889
144
Romstad E. (2014). “ The Economics of Eutrophication,” in Eutrophication : Causes, Consequences and Control. Eds. AnsariA. A.GillS. S. ( Springer, Netherlands), 45−53. doi: 10.1007/978-94-007-7814-6_4
145
Ruiz J. Olivieri G. De Vree J. Bosma R. Willems P. Reith J. H. et al . (2016). Towards industrial products from microalgae. Energy Environ. Sci.9, 3036–3043. doi: 10.1039/C6EE01493C
146
Ruiz-Frau A. (2023). Impacts of jellyfish presence on tourists’ holiday destination choices and their willingness to pay for mitigation measures. J. Environ. Plann. Manage.66, 2107−2125. doi: 10.1080/09640568.2022.2061926
147
Rumin J. Nicolau E. Gonçalves de Oliveira Junior R. Fuentes-Grünewald C. Picot L. (2020). Analysis of scientific research driving microalgae market opportunities in Europe. Mar. Drugs18, 264. doi: 10.3390/md18050264
148
Russo L. Bellardini D. Casotti R. Licandro P. Mazzocchi M. G. Murillas A. et al . (2025). The spatiotemporal variability of marine plankton ecosystem services at the regional scale: A combined approach using a systematic review and network analysis. Sustainability17, 1182. doi: 10.3390/su17031182
149
Sagarminaga Y. Piraino S. Lynam C. P. Leoni V. Nikolaou A. Jaspers C. et al . (2024). Management of jellyfish outbreaks to achieve good environmental status. Front. Ocean Sustainability2. doi: 10.3389/focsu.2024.1449190
150
Sanseverino I. Conduto D. Pozzoli L. Dobricic S. Lettieri T. (2016). Algal bloom and its economic impact (Luxembourg: Publications Office of the European Union).
151
Schaafsma M. Bartkowski B. (2020). “ Synergies and Trade-Offs Between Ecosystem Services,” in Life on Land. Eds. FilhoW.L.AzulA. M.BrandliL.ÖzuyarP. G.WallT. (Cham, Switzerland: Springer International Publishing), 1−11. doi: 10.1007/978-3-319-71065-5_117-1
152
Schenone L. Capitani L. Lora U. Setälä O. Kaartokallio H. Seppälä J. et al . (2025). Microbial plankton uptake enhances the degradation of a biodegradable microplastic. Environ. pollut.374, 126252. doi: 10.1016/j.envpol.2025.126252
153
Schernewski G. Inácio M. Nazemtseva Y. (2018). Expert based ecosystem service assessment in coastal and marine planning and management : A Baltic lagoon case study. Front. Environ. Sci.6. doi: 10.3389/fenvs.2018.00019
154
Selig E. R. Hole D. G. Allison E. H. Arkema K. K. McKinnon M. C. Chu J. et al . (2019). Mapping global human dependence on marine ecosystems. Conserv. Lett.12, e12617. doi: 10.1111/conl.12617
155
Shackleton C. M. Ruwanza S. Sinasson Sanni G. K. Bennett S. De Lacy P. Modipa R. et al . (2016). Unpacking pandora’s box : understanding and categorising ecosystem disservices for environmental management and human wellbeing. Ecosystems19, 587−600. doi: 10.1007/s10021-015-9952-z
156
Siegel D. A. DeVries T. Cetinić I. Bisson K. M. (2023). Quantifying the ocean’s biological pump and its carbon cycle impacts on global scales. Annu. Rev. Mar. Sci.15, 329–356. doi: 10.1146/annurev-marine-040722-115226
157
Štěpán M. Daďová K. Matouš M. Krauzová E. Sontáková L. Koc M. et al . (2021). Exercise training combined with calanus oil supplementation improves the central cardiodynamic function in older women. Nutrients14, 149. doi: 10.3390/nu14010149
158
Stephens G. L. O’Brien D. Webster P. J. Pilewski P. Kato S. Li J. (2015). The albedo of earth. Rev. Geophysics53, 141−163. doi: 10.1002/2014RG000449
159
Stern N. H. Peters S. Bakhshi V. Bowen A. Cameron C. Catovsky S. et al . (2006). Stern Review: The economics of climate change Vol. 30 (Cambridge: Cambridge University Press).
160
Stern N. Stiglitz J. E. (2021). The social cost of carbon, risk, distribution, market failures: An alternative approach Vol. 15 (Cambridge, MA, USA: National Bureau of Economic Research).
161
Stoltenberg I. Dierking J. Müller-Navarra D. C. Javidpour J. (2021). Review of jellyfish trophic interactions in the Baltic Sea. Mar. Biol. Res.17, 311326. doi: 10.1080/17451000.2021.1964532
162
Sutherland I. J. Villamagna A. M. Dallaire C. O. Bennett E. M. Chin A. T. M. Yeung A. C. Y. et al . (2018). Undervalued and under pressure : A plea for greater attention toward regulating ecosystem services. Ecol. Indic.94, 23−32. doi: 10.1016/j.ecolind.2017.06.047
163
Tagliabue A. Kwiatkowski L. Bopp L. Butenschön M. Cheung W. Lengaigne M. et al . (2021). Persistent uncertainties in ocean net primary production climate change projections at regional scales raise challenges for assessing impacts on ecosystem services. Front. Climate3. doi: 10.3389/fclim.2021.738224
164
Trottet A. George C. Drillet G. Lauro F. M. (2022). Aquaculture in coastal urbanized areas : A comparative review of the challenges posed by Harmful Algal Blooms. Crit. Rev. Environ. Sci. Technol.52, 28882929. doi: 10.1080/10643389.2021.1897372
165
Turner J. T. (2004). The importance of small planktonic copepods and their roles in pelagic marine food webs. Zoological Stud.43, 255–266.
166
Turner J. T. (2015). Zooplankton fecal pellets, marine snow, phytodetritus and the ocean’s biological pump. Prog. Oceanogr.130, 205−248. doi: 10.1016/j.pocean.2014.08.005
167
Tweddle J. F. Gubbins M. Scott B. E. (2018). Should phytoplankton be a key consideration for marine management? Mar. Policy97, 1−9. doi: 10.1016/j.marpol.2018.08.026
168
United Nations Global Compact (2024). The plankton manifesto. Available online at: https://ungc-communications-assets.s3.amazonaws.com/docs/publications/PlanktonManifesto_MG_DIGITAL-2.pdf (Accessed February 5, 2025).
169
Uye S. (2011). Human forcing of the copepod–fish–jellyfish triangular trophic relationship. Hydrobiologia666, 71−83. doi: 10.1007/s10750-010-0208-9
170
Van Dolah F. M. Roelke D. Greene R. M. (2001). Health and ecological impacts of harmful algal blooms : risk assessment needs. Hum. Ecol. Risk Assessment: Int. J.7, 13291345. doi: 10.1080/20018091095032
171
Vassallo P. Bellardini D. Castellano M. Dapueto G. Povero P. (2021). Structure and functionality of the mesozooplankton community in a coastal marine environment: Portofino marine protected area (Liguria). Diversity14, 19. doi: 10.3390/d14010019
172
Vella K. Dale A. Calibeo D. Limb M. Gooch M. Eberhard R. (2024). Monitoring the governance system underpinning the implementation and review of the Reef 2050 Plan : A preliminary benchmark (Brisbane, Australia: Queensland University of Technology and James Cook University). doi: 10.5204/rep.eprints.254729
173
Virdin J. Basurto X. Nico G. Harper S. Del Mar Mancha-Cisneros M. Vannuccini S. et al . (2023). Fishing for subsistence constitutes a livelihood safety net for populations dependent on aquatic foods around the world. Nat. Food4, 874−885. doi: 10.1038/s43016-023-00844-4
174
Von Rückert G. Giani A. (2008). Biological interactions in the plankton community of a tropical eutrophic reservoir : Is the phytoplankton controlled by zooplankton? J. Plankton Res.30, 1157−1168. doi: 10.1093/plankt/fbn065
175
Wang P. Deng X. Zhou H. Yu S. (2019). Estimates of the social cost of carbon: A review based on meta-analysis. J. cleaner production209, 1494–1507. doi: 10.1016/j.jclepro.2018.11.058
176
Waycott M. Duarte C. M. Carruthers T. J. B. Orth R. J. Dennison W. C. Olyarnik S. et al . (2009). Accelerating loss of seagrasses across the globe threatens coastal ecosystems. Proc. Natl. Acad. Sci.106, 12377–12381. doi: 10.1073/pnas.0905620106
177
Wotton R. S. Malmqvist B. (2001). Feces in aquatic ecosystems. BioScience51, 537. doi: 10.1641/0006-3568
178
Wright R. M. Le Quéré C. Buitenhuis E. Pitois S. Gibbons M. J. (2021). Role of jellyfish in the plankton ecosystem revealed using a global ocean biogeochemical model. Biogeosciences18, 1291−1320. doi: 10.5194/bg-18-1291-2021
179
Xiao G. Pan L. Lai F. (2025). Application, opportunities, and challenges of digital technologies in the decarbonizing shipping industry: a bibliometric analysis. Front. Mar. Sci.12, 1523267. doi: 10.3389/fmars.2025.1523267
180
Xu L. Wu J. Yan R. Chen J. (2025). Is international shipping in right direction towards carbon emissions control? Transport Policy166, 189–201. doi: 10.1016/j.tranpol.2025.03.009
181
Yuan J. Li M. Lin S. (2015). An improved DNA extraction method for efficient and quantitative recovery of phytoplankton diversity in natural assemblages. PloS One10, e0133060. doi: 10.1371/journal.pone.0133060
182
Zaucha J. Kreiner A. (2021). Engagement of stakeholders in the marine/maritime spatial planning process. Mar. Policy132, 103394. doi: 10.1016/j.marpol.2018.12.013
183
Zou Y. Xiao G. Li Q. Biancardo S. A. (2025). Intelligent maritime shipping: A bibliometric analysis of internet technologies and automated port infrastructure applications. J. Mar. Sci. Eng.13, 979. doi: 10.3390/jmse13050979
Summary
Keywords
plankton, ecosystem services, ecosystem disservices, expert assessment, economic value, semi-systematic review, marine policy, conservation approaches
Citation
Wanek E, Esteban-Cantillo OJ and Bourgeois-Gironde S (2025) Valuing marine plankton: a review of ecosystem services and disservices and an expert assessment of the potential of area-based protection. Front. Mar. Sci. 12:1607996. doi: 10.3389/fmars.2025.1607996
Received
08 April 2025
Accepted
24 June 2025
Published
16 July 2025
Corrected
24 October 2025
Volume
12 - 2025
Edited by
Lang Xu, Shanghai Maritime University, China
Reviewed by
Zachary Long, University of North Carolina Wilmington, United States; Guangnian Xiao, Shanghai Maritime University, China; Mingshuo Cao, Shanghai Maritime University, China
Updates
Copyright
© 2025 Wanek, Esteban-Cantillo and Bourgeois-Gironde.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Eva Wanek, eva.wanek@ens.psl.eu
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.