 Leonardo Romero-Martínez
Leonardo Romero-Martínez Amalia Prieto-Pérez-Juez
Amalia Prieto-Pérez-Juez Enrique Nebot
Enrique Nebot Javier Moreno-Andrés
Javier Moreno-Andrés- Department of Environmental Technologies, Faculty of Marine and Environmental Sciences, INMAR-Marine Research Institute, University of Cadiz, Puerto Real, Cadiz, Spain
Introduction: The Vibrio genus includes aquatic bacteria from freshwater, estuarine, and marine environments, some of which pose significant risks to public and environmental health, highlighting the need for effective disinfection strategies. This study investigated the inactivation kinetics, reactivation potential, and energy efficiency of UV-LEDs emitting at 265 nm and 275 nm for controlling Vibrio alginolyticus and Vibrio parahaemolyticus, two emerging marine pathogens.
Methods: Bacteria were exposed to UV irradiation under different post-treatment conditions (0 d, 1 d-dark, 1 d-light), and both inactivation rate constants (k, mJ·cm-2) and electrical energy per n-log reduction (EEL,n) were obtained and analyzed.
Results and discussion: Results showed that V. parahaemolyticus was more UV-sensitive than V. alginolyticus, with inactivation rate constants of 1.22 and 1.60 cm2·mJ-1 at 265 and 275 nm, respectively, compared to 0.82 and 0.69 cm2·mJ-1 for V. alginolyticus. No significant differences were observed between the two wavelengths (265 nm and 275 nm) in terms of inactivation rate constants. As a result, achieving a 2-log reduction required lower fluences for V. parahaemolyticus (3.68–2.89 mJ·cm-2) than for V. alginolyticus (5.53–6.85 mJ·cm-2). Post-treatment incubation caused a decline in k-values one day after UV exposure, particularly under light conditions, with reductions of up to 59.9% at 275 nm and 48.8% at 265 nm. Although the decrease in k-values under dark conditions was less pronounced (average 27.9%), it remains noteworthy when compared with other fecal bacterial indicators. Electrical energy analyses revealed lower EEL,1 values at 275 nm, indicating greater energy efficiency, with values ranging from 0.012 to 0.050 kWh·m-3 for both Vibrio species. These results support the need for wavelength-specific optimization to balance disinfection efficacy and energy efficiency in marine water treatment and seafood safety applications.
1 Introduction
The Vibrio genus comprises diverse Gram-negative bacteria that inhabit a broad range of aquatic environments, including freshwater, estuarine, and marine ecosystems (Almagro-Moreno et al., 2023; Seymour and McLellan, 2025). While many Vibrio species are harmless members of the natural microbiota, others pose significant risks to public health, aquaculture, and marine ecosystems. Notably, Vibrio cholerae is well known for its ability to survive and proliferate in freshwater, unlike most other species in the genus, which are primarily adapted to brackish or saline conditions (Grimes, 2020; Brumfield et al., 2023).
Climate-driven warming and extreme weather, together with anthropogenic alterations, such as urbanized coastlines and chemically modified estuarine environments, are increasing the vulnerability of coastal ecosystems, promoting the spread of Vibrio spp. and other non-indigenous species with broad environmental tolerance (González-Ortegón and Moreno-Andrés, 2021; Seymour and McLellan, 2025). These factors contribute to higher bacterial densities in marine environments, increasing the likelihood of human exposure through direct contact with contaminated water or the consumption of contaminated seafood (Baker-Austin et al., 2013, Baker-Austin et al., 2018; Trinanes and Martinez-Urtaza, 2021). Additionally, aquaculture facilities located in estuarine areas are particularly vulnerable to Vibrio-related outbreaks, which can lead to significant economic losses (Sony et al., 2021; Yang et al., 2021; Lo et al., 2023; Koutsoumanis et al., 2024; Siboni et al., 2024). Furthermore, the global dispersal of marine species through non-natural means, such as ballast water discharge, may enhance the risk of Vibrio infections by introducing new pathogenic strains into previously unaffected regions (Ng et al., 2018; Georgiades et al., 2021; Lv et al., 2023). Given these threats, effective mitigation strategies are urgently needed to control Vibrio contamination in seafood and marine environments, as recently emphasized by the European Food Safety Agency (EFSA) (Koutsoumanis et al., 2024).
Among Vibrio species, Vibrio cholerae (non-O1/non-O139), Vibrio parahaemolyticus, Vibrio vulnificus, and Vibrio alginolyticus are particularly significant due to their pathogenicity, ubiquity, and sensitivity to warming marine conditions (Baker-Austin et al., 2017; Gyraitė et al., 2024), However, V. parahaemolyticus and V. alginolyticus stand out as emerging pathogens due to their increasing incidence in human infections and seafood contamination. Both species thrive in warm, brackish waters, with their prevalence expected to increase due to climatic factors (de Souza Valente and Wan, 2021; Brumfield et al., 2023; Baker-Austin et al., 2024; Koutsoumanis et al., 2024). Additionally, they are widely used as indicator bacteria for evaluating disinfection methods in marine and seafood-processing environments (Wang et al., 2021b; Kuroyanagi et al., 2022; Hamza and Zinjarde, 2023; Ma et al., 2023; Moreno-Andrés et al., 2023; Lim and Kang, 2024).
Control of these emerging pathogens requires effective treatment strategies. While chemical treatments like ozonation or chlorination can control biological quality (Moreno-Andrés et al., 2020; Pumkaew et al., 2021; Cho et al., 2024), they raise concerns about toxic substance management, disinfection by-products, and biofilm formation with bacterial adaptation (Mougin et al., 2023). UV treatment has become a preferred method for disinfecting intake water in aquaculture facilities and shellfish depuration systems (Koutsoumanis et al., 2024). It is also the main option for ballast water treatment systems (Hess-Erga et al., 2019), providing effective pathogen inactivation.
The transition from traditional mercury-based UV lamps to UV-LED technology is essential due to the environmental and regulatory constraints imposed by the Minamata Convention, which aims to phase out mercury-containing products (UNEP, 2019). UV-LEDs offer mercury-free operation, design flexibility, and long lifespan, and though current efficiencies at lower wavelengths are limited, improvements are expected in the coming years (Martín-Sómer et al., 2023). Additionally, UV-LEDs allow for precise wavelength selection, which is crucial given that different UV-C wavelengths may exhibit varying disinfection mechanisms and efficiencies (Martín-Sómer et al., 2023; Sun et al., 2023).
The effectiveness of UV-LEDs in microbial disinfection depends on the spectral sensitivity of each target organism, emphasizing the need for precise dose–response characterization (Sun et al., 2023). While DNA exhibits peak UV absorption at approximately 260 nm, variations in nucleic acid composition and cellular structure among microorganisms can influence the optimal wavelength for inactivation (Li et al., 2019; Martín-Sómer et al., 2023; Sun et al., 2023). For instance, 265 nm UV induces more DNA damage than 285 nm, particularly in Gram-negative bacteria, which are more susceptible due to their thinner peptidoglycan layer (Sun et al., 2023).
On the other hand, a key challenge in UV disinfection is the potential for microbial regrowth by organisms that were not inactivated by the treatment, as well as organisms that recovered their ability to reproduce due to photo-reactivation or dark repair mechanisms, particularly when sublethal UV doses are applied. This concern is particularly relevant for marine bacteria, which remain in their natural environment post-disinfection, unlike fecal bacteria that are typically introduced from external sources. Research suggests that longer UV-C wavelengths, such as 280 nm, while less germicidal than the 260–265 nm range, may be more effective in suppressing photoreactivation and dark repair (Li et al., 2017; Nyangaresi et al., 2018), highlighting the importance of wavelength selection for sustained microbial inactivation. Due to photo-reactivation and dark-repair mechanisms, post-treatment conditions should be considered when evaluating treatment efficacy, particularly in marine ecosystems where these processes remain less studied.
This study aims to evaluate and define the dose–response curves of two emerging marine pathogens, Vibrio alginolyticus and Vibrio parahaemolyticus, under UV-LED exposure at 265 nm and 275 nm. Additionally, it seeks to assess their potential for post-treatment reactivation under light and dark conditions, providing insights into the efficacy of UV-LED disinfection at tailored wavelengths. By analyzing interspecies differences, this research contributes to the optimization of UV-based disinfection strategies for improved microbial control in marine environments.
2 Material and methods
2.1 Organisms and microbial procedures
2.1.1 Bacterial strains and preparation of the challenge water
The organisms used in this study were the bacteria Vibrio alginolyticus (ATCC 17749/CECT 521T) and Vibrio parahaemolyticus (ATCC 17802/CECT 511T). The bacteria strains were acquired to the Spanish Type Culture Collection (CECT, University of Valencia, Spain) in lyophilized format. Lyophilized organisms were recovered in Marine Broth (2216 Difco™) at 30°C for 24 h and subsequently sub-cultured for another 24 h. The resulting culture was aliquoted, mixed with a 50:50 glycerol-water solution, and stored in cryovials at -30°C for long-term preservation.
To prepare the working strain, the contents of a cryovial were resuspended in 50 mL of Marine Broth and incubated at 30°C for 24 h. The culture was then transferred into 50 mL of fresh Marine Broth and incubated for an additional 24 h. These conditions were selected based on the recommendations of the Spanish Type Culture Collection (CECT) for V. alginolyticus and V. parahaemolyticus. Similar incubation protocols have also been used in previous disinfection studies involving marine bacteria, including the specific strains employed in this work, i.e., ATCC 17749 and ATCC 17802 (Wu et al., 2011; Lomelí-Ortega and Martínez-Díaz, 2014; Moreno-Andrés et al., 2018, 2020; Zhang et al., 2023). After incubation, 45 mL of the sub-culture were divided into sterile vials and centrifuged at 3000 rpm for 10 min. The supernatant was discarded, and 5 mL of artificial seawater (prepared with 35 g L-1 of Instant Ocean® sea salt in distilled water) were added to each vial. The bacterial pellets were resuspended in 50 mL of artificial seawater in a sterile flask, yielding the final bacterial inoculum.
The challenge water was prepared by adding 1 mL of the bacterial inoculum into 1 L of artificial seawater. The bacteria concentration in the challenge water ranged between 9.35·106 and 2.83·107 CFU mL-1.
2.1.2 Determination of the bacterial concentration
The UV treatment efficacy was determined by enumerating the culturable bacteria concentration in the unirradiated controls and the treated samples. Ten-fold dilutions were made from every sample, and 1 mL of each dilution tier was filtered through gridded membranes of 0.45 µm (Pall Corporation) according to the membrane filtration method. The membranes with the retained bacteria were plated in Petri dishes with Thiosulfate Citrate Bile Salts Sucrose (TCBS Agar, Scharlau) and incubated at 30°C for 24 h. Although the possibility of inducing viable but non-culturable (VBNC) states during UV disinfection cannot be ruled out (Wang et al., 2021a), this limitation is softened by the use of spiked ATCC strains under controlled laboratory conditions. In fact, previous studies have shown that plate count methods can reliably reflect bacterial survival in similar experimental setups (Wennberg et al., 2013). Nevertheless, in future applications involving environmental isolates or complex microbial communities, the use of complementary culture-independent methods is recommended to account for sublethally injured or VBNC populations.
After the incubation, the colonies were enumerated. For a proper accuracy of the calculations, the plates with a CFU number between 15 and 150 were considered as valid for calculation; less than 15 CFU were considered as non-representative and more than 150 CFU does not allow the colonies to growth properly and hampers the correct enumeration. The culturable bacteria concentration in the samples was calculated as the quotient of the CFU and the filtered volume multiplied by the corresponding dilution factor.
2.2 UV reactors and dose calculation
Two UV devices were used in the experiments (Photolab LED265-0.1er/cb and Photolab LED275-0.1er/cb, APRIA Systems S.L., Spain). Each reactor was equipped with a LED (100 mW), emitting at λmax = 265 nm or λmax = 275 nm, allowing the evaluation of microbial inactivation and reactivation at two representative UV-C wavelengths, while ensuring consistency with previous studies and current technological constraints.
The UV dose was determined as the product of the mean intensity (Im) and the exposure time (t). The Im was calculated according to the protocol by Bolton and Linden (2003) (Bolton and Linden, 2003). The irradiance at the center of the sample surface was determined by a radiometer (HD 2102.1, Delta OHM) equipped with a UV-CB probe (Delta OHM LP471). The water transmittance (Tw) at either 265 or 275 nm was measured right before the UV irradiation, providing values between 70.3% and 75.0%, and used to calculate the Im value in each experiment. The relatively low transmittance values can be attributed to the formation of micro-precipitates after autoclaving and the high bacterial density. Once determined the Im (0.809-0.843 mW·cm-2), the exposure time required to apply UV doses between 4 and 18 mJ·cm-2 were calculated (Supplementary Table S1). The range of UV doses was selected to determine the inactivation kinetics according to preliminary studies.
2.3 Experimental approach
The challenge water containing either V. alginolyticus or V. parahaemolyticus was irradiated with different UV doses emitting at λmax = 265 nm or λmax = 275 nm and subjected to different post-treatment illumination regimes (Figure 1).
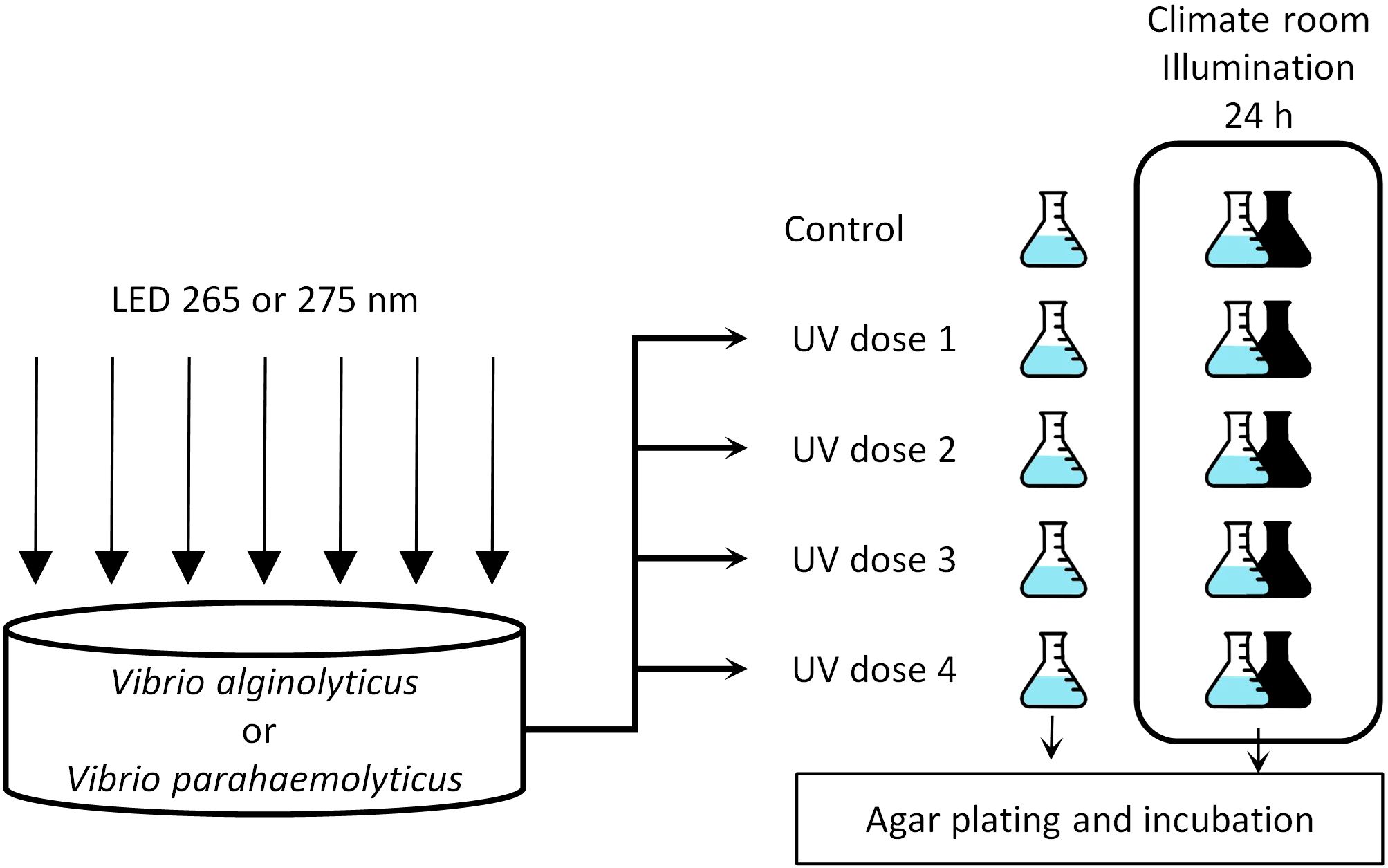
Figure 1. Experimental setup for assessing V. alginolyticus and V. parahaemolyticus inactivation under two different UV-LED wavelengths (λmax = 265 nm; λmax = 275 nm), as well as evaluating photoreactivation and dark repair potential 24 hours post UV exposure.
20 mL of the challenge water was placed into a glass Petri dish with 49 mm of internal diameter. The sample volume was irradiated for the time required to apply the target UV dose; then the sample volume was transferred into a 50 mL sterile borosilicate Erlenmeyer flask and subjected to the microbiological determination procedure (section 2.1.2). The irradiation procedure was repeated, and the treated sample was transferred into another sterile Erlenmeyer and placed into a climate room at 24°C and illumination at 37.3 μmol photons m-2 s-1, with mixed cold white 6,500K and warm white 2,700K light. For dark repair assessment, the same UV dose was applied for a third time, with the resulting sample transferred into another Erlenmeyer flask, but covered with aluminum foil to prevent bacteria exposure to the environmental light and placed in the climate room. Flasks were covered but allowing the air exchange and incubated in static condition. The same procedure was repeated for the three remaining target UV doses and for non-irradiated control. The incubated samples associated with photo-reactivation and dark repair experimentation were subjected to the bacterial determination procedure after 24 h of incubation (1d-light or 1d-dark, respectively) as the repair processes have placed between 1 and 12 h after the UV irradiation (Oguma et al., 2001; Nebot et al., 2007; Quek and Hu, 2008). Two samples were collected per UV dose, and the experiment was repeated twice for each organism, thus obtaining four samples for every combination of organism, UV dose, wavelength and post-treatment condition.
2.4 Data analysis and statistics
For each experimental series, the survival (S) of every sample was calculated as the quotient between the concentration of culturable bacteria in the treated sample (N) and their corresponding non-irradiated and non-incubated control (N0). Therefore, S includes the effect by the UV irradiation as well as the possible growth or mortality occurring throughout to the 24 h storage.
The values of Log (S) were represented against the corresponding values of UV dose to obtain the inactivation curves. The inactivation curves were modeled according to the log-linear + tail inactivation model (Equation 1, in which S0 is the survival in absence of UV irradiation, Sres is the asymptotic value of survival at high UV doses, and k, cm2 mJ-1, is the inactivation kinetic constant) (Geeraerd et al., 2005).
Modeling was carried out using SigmaPlot 11.0, providing the values for the three model parameters, their standard errors and p-values, as well as the R2 for the correlation between estimated and experimental values (Supplementary Table S2). For a practical evaluation of the treatment efficacy, the inactivation model parameters were used to calculate the Dn, understood as the UV dose required to achieve “n” log-reductions of the culturable bacteria concentration. Dn was calculated for every organism, wavelength and post-treatment regime.
The inactivation curves data were also subjected to ANCOVA using Log (S) as dependent variable, the UV dose as covariate, and the bacterial species (V. alginolyticus or V. parahaemolyticus), the wavelength (265 or 275 nm) and the post-treatment regime (0 d, 1 d - light, or 1 d - dark) as qualitative factors, to determine their potential impact on the treatment efficacy. p-values were adjusted according to the Bonferroni procedure. The interaction terms (Organism x UV dose, Wavelength x UV dose, and Post-treatment x UV dose) were not significant (p > 0.05) and thus discarded from the analysis.
Energy efficiency assessments have been widely applied to evaluate the performance of various UV disinfection systems, including those utilizing UV-LEDs and low-pressure mercury lamps (Beck et al., 2017; Martín-Sómer et al., 2023; Sun et al., 2023). In this study, the Electrical Energy per Log Reduction (EEL,n) was calculated (Equation 2), which incorporates parameters from the experimental set-up and inactivation results (Beck et al., 2017; Sun et al., 2023). The EEL,n values were obtained using the following equation:
The parameters used in the EELn calculations include A, which represents the irradiated surface area (cm2), and Dn, the UV fluence (mJ·cm-2) required for achieving an n-log reduction. The sample volume (V) is measured in mL, while C corresponds to the wall-plug efficiency of the UV-LED, a value provided by the manufacturer. Additionally, the water factor (WF) is considered, following the methodology described by Bolton and Linden (2003).
3 Results and discussion
3.1 Dose-response profiles for inactivation
The inactivation curves exhibited similar patterns for both V. alginolyticus and V. parahaemolyticus across UV exposures at 265 nm and 275 nm, as well as for measurements taken immediately after UV irradiation (0 d) and those taken after 24 h of incubation under dark (1 d - dark) or light (1 d - light) conditions. As shown in Figure 2, the curves initially display a linear decrease in Log (S) with increasing UV dose, followed by a stage where increasing the dose no longer enhances inactivation, a phenomenon known as tailing (Cerf, 1977).
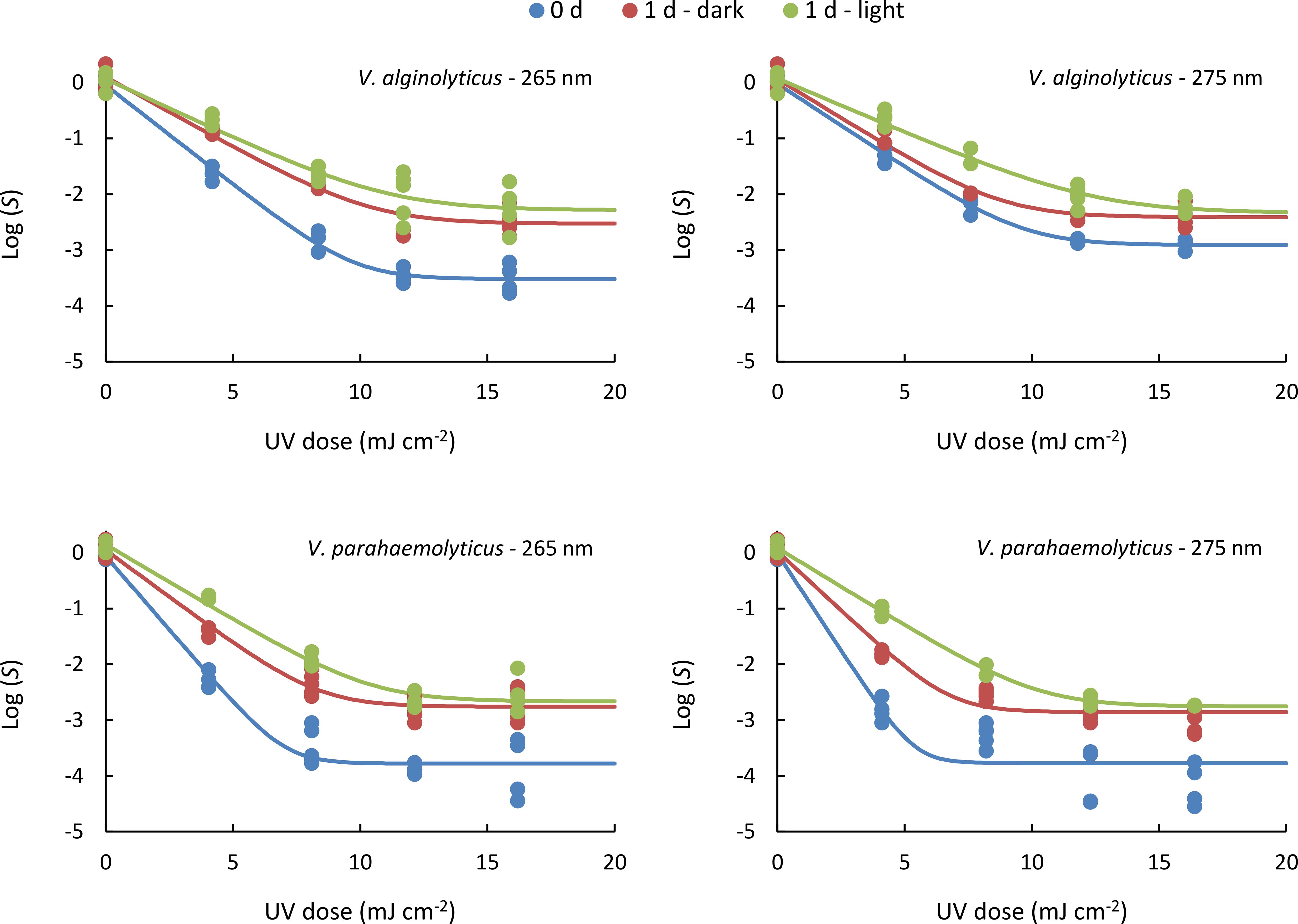
Figure 2. Inactivation curves obtained for both organisms treated at either 265 or 275 nm, under different post-treatment conditions: determined right after the UV irradiation (0 d) or after 24 h under dark (1 d - dark) or light (1 d - light) incubation. Lines represent the log-linear + tail model fitted to the experimental data. Data represent individual values obtained from analyses.
In all cases, inactivation was highest immediately after UV irradiation, followed by 1d - dark samples, and lowest in 1d-light samples. Thus, the data obtained reveal that the inactivation level is reduced one day after the UV irradiation (Figure 2). Bacterial regrowth can occur through reactivation from a viable but non-culturable state, repair of photo-induced DNA damage, and reproduction of bacteria surviving disinfection (Wang et al., 2021a). The values of Log (S) in absence of UV irradiation (Log (S0)) are non significant (Supplementary Table S2), indicating the absence of reproduction of survivor bacteria, since the water matrix was deprived of nutrients. For this reason, the greater concentration of culturable bacteria one day after the UV irradiation can be attributed to the UV-damage repairing processes (Sinha and Häder, 2002).
Similarly, tailing phenomena is observed in all cases (Figure 2). The tailing effect in inactivation processes has been commonly observed in bacteria (Rattanakul and Oguma, 2018; MacIsaac et al., 2024), including Vibrio species (Hamamoto et al., 2010; Romero-Martínez et al., 2022; Kyriazi et al., 2023). Traditionally, this behavior has been linked to the presence of a treatment-resistant microbial subpopulation (Nyangaresi et al., 2018), microbial aggregation (Vitzilaiou et al., 2021), or the protective role of the surrounding matrix, which can shield a fraction of the population from direct exposure (Azimi et al., 2017). Additionally, engineering and hydrodynamic factors, such as intensity-exposure reciprocity and reactor design, may influence tailing behavior (Oguma et al., 2013; Guerra-Rodríguez et al., 2022; Romero-Martínez et al., 2022; Blanchon et al., 2025). Recent research suggests an alternative explanation, proposing that stochastic variations in gene expression may contribute to the tailing effect (Oppezzo et al., 2024). Accordingly, the tailing phenomena is influenced by both biological factors and operational conditions, making it unpredictable without real-world testing in actual reactors and challenge water.
3.2 Inactivation kinetics
The inactivation curves observed (Figure 2) followed a log-linear + tail kinetics model (Equation 1), with all kinetic parameters detailed in Supplementary Table S2. The R2 for the correlation between estimated and experimental values was > 0.9 in all cases. The values of Log (S0) were determined as non-significant (p > 0.05). This indicates that bacterial concentration remained unchanged during light or dark incubation compared to the levels measured immediately after UV irradiation, indicating that differences in UV-treated samples are due to dark repair and photo-reactivation. The values of Log (Sres) were significant (p < 0.001) in all cases (Figure 3A; Supplementary Table S2). Log (Sres) values indicate that the maximum level of inactivation was greater in samples analyzed right after the UV irradiation, whereas the maximum level of inactivation reachable was reduced in the samples incubated for one day. The k values were significant (p < 0.001) in all cases (Figure 3B; Supplementary Table S2), with the highest values observed immediately after UV irradiation, followed by samples kept in the dark for one day, and the lowest k-values in those incubated under light for one day. This confirms that reactivation occurs in both Vibrio species and under both UV-C wavelengths.
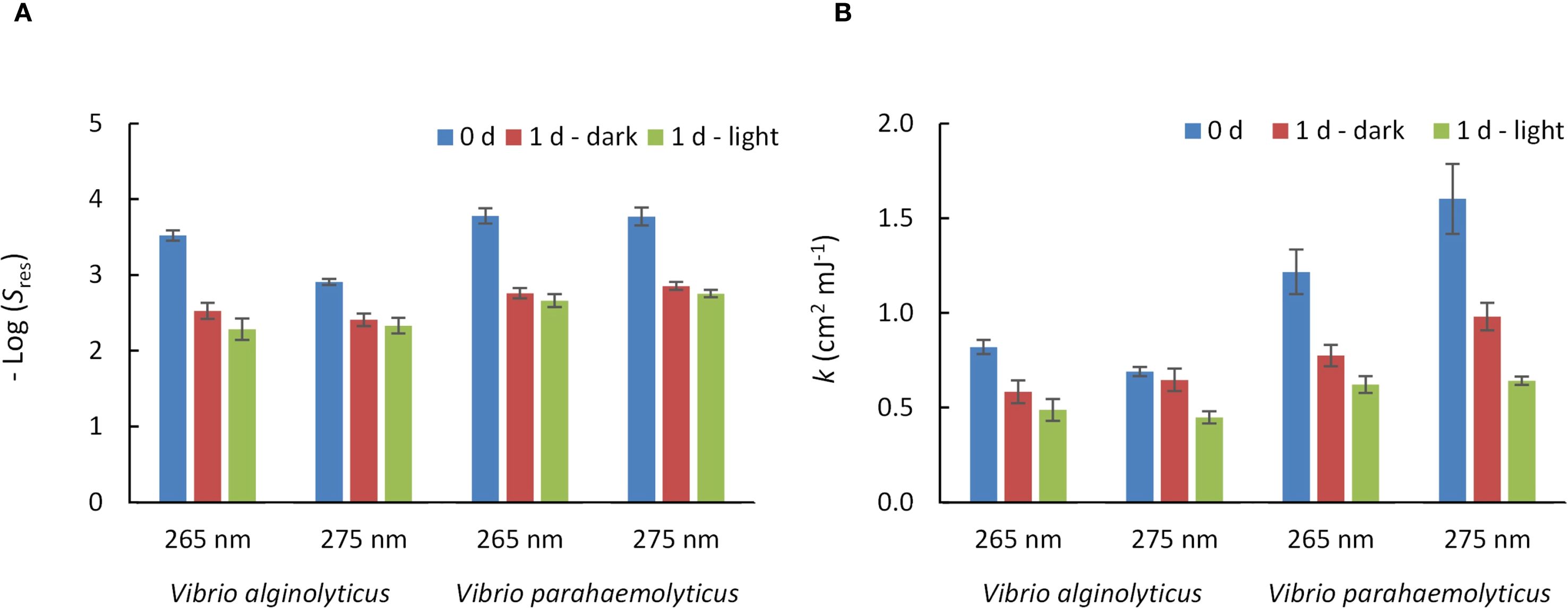
Figure 3. Inactivation kinetic parameters for V. alginolyticus and V. parahaemolyticus under UV exposure at 265 nm or 275 nm and different post-treatment incubation conditions. (A) Sres: asymptotic survival at high UV doses. (B) k: inactivation constant (cm2·mJ-1). Numerical values, standard errors and p-values of the kinetic parameters, and R2 values can be found in the Supplementary Table S2 of the Supplementary Material. Error bars represent the standard error of the model parameters.
To determine the significance of the studied factors involved in the UV inactivation (UV dose, target organism, treatment wavelength, and reactivation processes) an ANCOVA was applied to the inactivation curves data (Supplementary Table S3). The ANCOVA indicated significant (p< 0.001) effect of the UV dose on Log (S); significant (p< 0.001) differences between both species, with greater effect on V. parahaemolyticus with respect to V. alginolyticus (Figure 4A), non-significant (p = 0.521) differences between both emission wavelengths (Figure 4B), and significant differences (p< 0.001) between the three post-treatment conditions (Figure 4C).
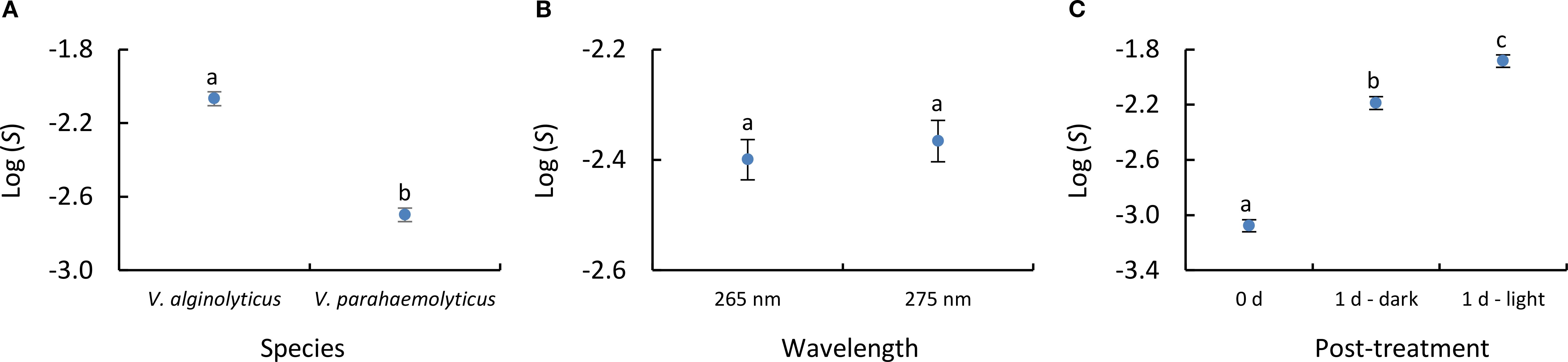
Figure 4. ANCOVA results obtained using Log (S) as dependent variable, UV dose as covariate, and (A) species, (B) emission wavelength and (C) post-treatment conditions as qualitative factors. Error bars represent the 95% confidence interval. Lowercase letters indicate the homogenous groups reported in the analysis.
The results revealed differences in UV sensitivity between the two Vibrio species. V. parahaemolyticus showed higher sensitivity, with greater inactivation rate constants at both 265 nm and 275 nm, compared to V. alginolyticus. Post-treatment incubation led to a notable reduction in inactivation rate constants, particularly under illuminated conditions. For V. parahaemolyticus, k-values decreased by 36.3% (1d-dark) and 48.8% (1d-light) at 265 nm, and by 38.8% and 59.9% at 275 nm, respectively. V. alginolyticus showed a less pronounced reduction of k-values, with decreases of up to 40.4% at 265 nm and 35.1% at 275 nm under light exposure. These results confirm that incubation under light conditions promotes a greater reduction in disinfection efficacy than incubation in darkness, highlighting the importance of controlling post-treatment exposure to light.
The differential sensitivity of V. alginolyticus and V. parahaemolyticus to UV-C radiation is likely influenced by physiological and genetic factors, as both strains were subjected to identical experimental conditions, meaning that observed differences arise from intrinsic strain characteristics. As Gram-negative bacteria, both species are expected to show moderate-to-high sensitivity to UV-C; however, variations in UV resistance may be attributed to the presence and expression of the RecA gene and other genetic factors, including oxidative stress defense mechanisms (Bauermeister et al., 2009; Pousty et al., 2021; Martín-Sómer et al., 2023). Another key factor influencing UV sensitivity is the role of outer membrane proteins (OMPs), which serve as the first structural barrier against UV-C radiation. Their composition and modifications can significantly impact bacterial resistance and adaptation to stress. Abdallah et al., 2012 suggest species-specific responses to UV stress based on OMP analysis in V. alginolyticus and V. parahaemolyticus, while Romero-Martínez et al., 2023 also observed differences in UV resistance among various E. coli strains. In fact, we observed intraspecific variability in the UV-sensitivity on bacteria, which, in some cases exceeds the interspecific differences (Romero-Martínez et al., 2023). Even more, the physiological status of the target bacteria has also an influence on the UV treatment outcome (Keller and Maxcy, 1984; Vitzilaiou et al., 2021). These findings imply that each species could employs distinct response mechanisms that contribute to their relative UV-C sensitivity.
While this study provides valuable insights into the behavior of two representative Vibrio species, it is important to acknowledge that only ATCC reference strains were used. Given the genetic diversity and environmental adaptability of Vibrio spp., wild-type isolates may exhibit different behavior (Quek and Hu, 2008; Jütte et al., 2023). Future studies should consider a broader panel of environmental strains to capture this variability and strengthen the applicability of UV-LED disinfection strategies under real-world conditions. For these reasons, the variability on bacteria species, strains and physiological statuses should have into account for the validation and application of the UV treatment on natural water.
On the other hand, reactivation became evident when comparing non-incubated samples with those incubated for 24 hours (Figures 2, 3). In addition, significant differences were also observed between 1d - dark and 1d - light samples (Figure 4). This fact indicates that both dark-repair and photo-reactivation mechanisms cause significant DNA-repairing effect, prevailing photo-reactivation over dark repair processes.
Microorganisms recover from UV-induced DNA damage through photoreactivation and dark repair (Sinha and Häder, 2002). Photoreactivation, driven by photolyase and UVA/visible light, repairs cyclobutane pyrimidine dimers (CPDs) in DNA. Dark repair, in contrast, replaces damaged nucleotides without light, using nucleotide excision repair. However, dark repair is less efficient, as light enhances the repair process (Li et al., 2017; Nyangaresi et al., 2018; Xiao et al., 2018; Martín-Sómer et al., 2023; Zhang et al., 2023). This is supported by our results, which show a significant difference between samples incubated for 1 day in the dark compared to those incubated under light (Figure 4). Specifically, the kinetic rate constant was reduced by 15.5-19.5% at 265 nm and 30.8-34.7% at 275 nm (depending on Vibrio species) when comparing 1d-dark samples to 1d-light samples (Figure 3). These trends in reactivation align with findings from other studies specifically focused on Vibrio species. Research on Vibrio cholerae has shown that photoreactivation is significantly more efficient than dark repair (Das et al., 1981), while Vibrio natriegens has been reported to possess both photo-reactivation and dark-repair mechanisms (Hoff et al., 2020).
Finally, although dark repair in V. alginolyticus and V. parahaemolyticus had a smaller impact than photo-repair, it still showed a significant effect when compared to 0 d samples (Figure 4). This is particularly relevant for marine bacteria, as previous studies using fecal indicators reported non-significant reductions in k-values after 1 day of dark incubation (Romero-Martínez et al., 2023). In fact, repair percentages were also calculated based on bacterial concentrations and UV dose, following the approach by Lindenauer and Darby, 1994, yielding values between 0.096–8.98% at 265 nm and 0.088–7.32% at 275 nm, depending on the bacterial species and the UV dose applied (Supplementary Figure S1). Similar trends have been reported in disinfection studies, where marine bacteria like Aeromonas salmonicida show significantly higher dark repair percentages than common fecal indicators such as E. coli (Zhang et al., 2023). This highlights the greater resilience of marine bacteria, which can subsequently result in regrowth episodes after UV treatment.
These results suggest that current UV disinfection standards (based on non-native fecal bacteria) may not fully represent the risks in marine environments. Although these indicators, such Enterococcus faecalis, are more UV-resistant (Malayeri et al., 2016), their limited survival in saline waters reduces their relevance in coastal or aquaculture settings (Winfield and Groisman, 2003; Tiwari et al., 2019). In contrast, marine pathogens such as V. alginolyticus and V. parahaemolyticus can grow to high levels in seawater and recover after treatment through photo-repair or dark repair mechanisms. This may compromise the effectiveness of disinfection. Therefore, it is recommended that marine-relevant bacteria be included in performance validations of UV systems, especially for applications like aquaculture or ballast water management. Incorporating native marine indicators could lead to more accurate risk assessments and more robust regulatory frameworks.
3.3 Operational and practical considerations: UV-doses and energy efficiency
The values of Dn provide an approach of the inactivation efficacy that is more direct and intuitive in comparison with the inactivation kinetics parameters (Hijnen et al., 2006; Malayeri et al., 2016). In all cases, 1-log (90%) and 2-log (99%) reductions were achieved, allowing consistent calculation of D1 and D2 values (Table 1). At 0 d, D1 ranged from 1.40 to 3.24 mJ·cm-2, and D2 from 2.89 to 6.85 mJ·cm-2. These values are consistent with those reported for other Vibrio species, such as V. cholerae and V. anguillarum (Hijnen et al., 2006; Malayeri et al., 2016), and support the classification of Vibrio spp. as UV-sensitive organisms due to the relatively low doses required for effective inactivation. Although the inactivation efficacy of V. alginolyticus at 275 nm was slightly lower than that reported by Romero-Martínez et al., 2022 or Moreno-Andrés et al., 2023, this may be explained by the higher irradiance used in the present study. These differences suggest that intensity–time reciprocity and wavelength dependence can influence disinfection outcomes, highlighting the need for further research on these interactions across different microbial targets.
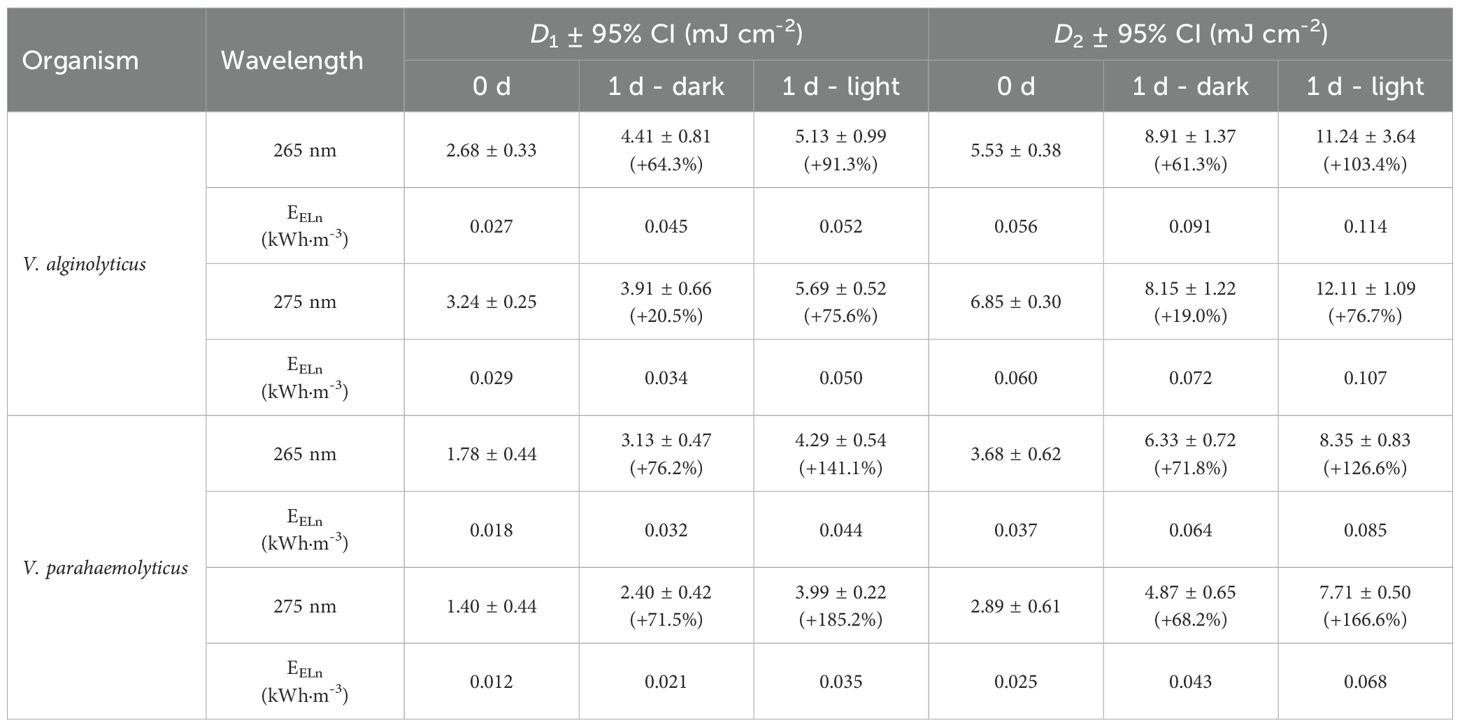
Table 1. Values of D1 and D2 (UV dose required to achieve 1 or 2 log-reductions, mJ·cm-2) and electrical energy per 1 or 2-log reduction (EEL,1; EEL,2; kWh·m-3) for the different target organisms, emission wavelengths and post-treatment regimes. It is also specified the increasing percentage of the UV dose requirement due to the incubation under dark or light conditions.
As highlighted in previous sections, reactivation processes can substantially increase the UV dose required to achieve effective disinfection. In the case of V. alginolyticus, reactivation may lead to an increase in UV dose of up to 91.3% for D1 and 103.4% for D2, while for V. parahaemolyticus, the required increase can reach 185.2% for D1 and 166.6% for D2. These results emphasize the critical impact of post-treatment conditions on disinfection efficacy. Therefore, final use conditions (whether the treated water is to be stored, discharged, or reused) should be carefully considered, particularly in terms of exposure to light or darkness.
At this point, it is important to note that the UV-LED disinfection performance evaluated in this study was based on controlled laboratory assays using synthetic saline water. In natural marine environments, variable concentrations of dissolved organic matter (DOM), suspended solids, inorganic ions, and native microorganisms can significantly influence disinfection outcomes (Gandhi and Prakash, 2023). These constituents may attenuate UV radiation by absorption and scattering, reducing the effective fluence and increasing the dose required for equivalent inactivation. DOM, in particular, can act both as a UV absorber and as a photosensitizer depending on its composition and the wavelength used. While the photosensitization effect of DOM can promote bacterial inactivation via singlet oxygen or triplet state reactions leading to membrane and DNA damage, it may also shield bacteria from direct UV exposure (Serna-Galvis et al., 2018; Gandhi and Prakash, 2023). Moreover, UV disinfection may alter microbial community composition in treated marine waters (e.g., ballast water, aquaculture effluents) by promoting selective survival or recolonization (Hess-Erga et al., 2019). In this study, water transmittance was incorporated into UV-Dose calculations according to standard protocols (Bolton and Linden, 2003); however, future work should include matrix-specific assessments to enhance the applicability of these findings under real-world marine conditions.
The electrical energy consumption per 1 or 2 log reduction (EEL,1; EEL,2) was compared for V. alginolyticus and V. parahaemolyticus at 265 nm and 275 nm, considering different post-irradiation conditions: 0 d, 1 d-dark, and 1 d-light (Table 1). Among the tested conditions, 275 nm generally required lower energy consumption for inactivation compared to 265 nm, particularly for V. parahaemolyticus. For instance, for D1, V. parahaemolyticus exhibited the lowest EEL,1 at 275 nm with 0.012 kWh·m-3, whereas V. alginolyticus required 0.029 kWh·m-3 under the same conditions, with similar trend observed for D2. EEL,n values increased under post-irradiation conditions, particularly after 1d-light incubation. For 99% inactivation, EEL,2 increases ranged from 37.9–41.8% (265 nm) and 15.9–40.6% (275 nm) under 1d-dark, and 50.8–55.9% (265 nm) and 43.4–62.5% (275 nm) under 1d-light, depending on the bacterial species.
Compared to previous studies, our results show similar EEL,n trends for UV-LEDs at 260–280 nm. Beck et al., 2017 reported that 280 nm LEDs required less EEL,n than 260 nm LEDs for E. coli inactivation (a typical fecal indicator, Gram-negative bacteria, as Vibrio), a pattern also observed by Rattanakul and Oguma, 2018, or Sun et al., 2023 across multiple bacterial species. In our study, 275 nm was more energy-efficient than 265 nm, despite requiring slightly higher UV doses, emphasizing that wavelength selection should consider both inactivation efficiency and electrical energy consumption to optimize UV-LED disinfection systems.
4 Conclusions
This study evaluated the effectiveness of UV-LEDs emitting at 265 nm and 275 nm for the inactivation of two emerging Vibrio pathogens, Vibrio alginolyticus and Vibrio parahaemolyticus. The inactivation kinetics, reactivation potential (photo repair and dark repair), and electrical energy consumption were analyzed to determine the most efficient disinfection conditions.
Both species were efficiently inactivated at relatively low UV doses, with inactivation rate constants (k) ranging from 0.69 to 1.60 cm2·mJ-1 depending on the species and wavelength used. No significant differences were observed in inactivation rate constants between 265 nm and 275 nm. This confirms the feasibility of UV-LEDs for Vibrio disinfection in marine environments. The inactivation constant of V. alginolyticus was 32.8% lower than that of V. parahaemolyticus at 265 nm, and 56.9% lower at 275 nm, indicating greater UV-resistance for V. alginolyticus.
Reactivation was observed, with post-treatment exposure to light having a greater impact than incubation in darkness. Compared to 0 d (immediate post-UV inactivation), the k-values decreased by up to 38.8% at 275 nm under dark conditions, while photoreactivation led to reductions of up to 59.9%. Interestingly, the dark repair percentages observed for V. alginolyticus and V. parahaemolyticus were considerably higher than those typically reported for fecal bacteria.
Electrical energy consumption per log reduction (EEL,1) was lower at 275 nm, with values of 0.012–0.035 kWh/m³ for V. parahaemolyticus and 0.029–0.050 kWh·m-3 for V. alginolyticus, compared to higher EEL,1 values at 265 nm. This suggests that 275 nm LEDs are a more energy-efficient choice.
These results demonstrate that UV-LEDs are effective for Vibrio inactivation, but wavelength selection should balance germicidal performance, energy consumption, and post-treatment bacterial recovery to optimize disinfection strategies. The findings also have relevant implications for scaling up UV-LED systems. Although no significant differences in inactivation were found between 265 nm and 275 nm, the higher energy efficiency of 275 nm supports its selection for practical applications. Importantly, the observed dark repair and photo-reactivation in V. alginolyticus and V. parahaemolyticus must be considered in system design to ensure microbiological safety. Transitioning from batch to flow-through reactors (necessary for full-scale implementation) may alter inactivation kinetics due to changes in UV intensity and exposure time; thus, time–dose reciprocity should be further investigated. Additionally, real water matrix effects (e.g., DOM) and strain-specific UV resistance must be addressed to ensure consistent performance in seawater treatment.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors upon request.
Ethics statement
The manuscript presents research on animals that do not require ethical approval for their study.
Author contributions
LR-M: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Writing – original draft, Writing – review & editing. AP-P-J: Data curation, Formal analysis, Investigation, Methodology, Writing – review & editing. EN: Conceptualization, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Writing – review & editing. JM-A: Conceptualization, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Visualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This work is part of the project MOBILED: PCM_00015 co-funded by the Regional Ministry of University, Research, and Innovation of the Government of Andalusia and by the European Union NextGenerationEU/PRTR. C.17, I1. Authors also wants to acknowledge the financial support of the project ECOTRANSEAS (TED2021-130994B-C31) funded by MCIN/AEI/10.13039/501100011033 and by the European Union NextGenerationEU/PRTR. The author L. Romero-Martínez is hired in the frame of the Programme Qualifica 2021 (Ref: QUAL21_019_UCA, Junta de Andalucía), funded by the “Plan Andaluz de Investigación, Desarrollo e Innovación (PAIDI 2020)” of the Regional Government of Andalusia.
Acknowledgments
Amalia Prieto developed his Final Degree Project within the framework of this study; she acknowledges “Plan Propio – UCA 2023-2024” for the Grant INICIA-INV. Ref.: 2023-012/PU/PPINICIA-INV/BN. The authors thank the "Plan Propio de apoyo y estímulo a la Investigación y Transferencia 2025-2027" for funding the Open Access fees (Ref.: PB2025-062).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that Generative AI was used in the creation of this manuscript. Generative AI tools were used solely for language editing and grammar refinement during the preparation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2025.1608367/full#supplementary-material
References
Abdallah F., Ben, Lagha R., Ellafi A., Namane A., Rousselle J. C., Lenormand P., et al. (2012). Identification of Outer Membrane Proteins Altered in Response to UVC-Radiation in Vibrio parahaemolyticus and Vibrio alginolyticus. Indian J. Microbiol. 52, 660–665. doi: 10.1007/s12088-012-0299-2
Almagro-Moreno S., Martinez-Urtaza J., and Pukatzki S. (2023). Vibrio infections and the twenty-first century. Adv. Exp. Med. Biol. 1404, 1–16. doi: 10.1007/978-3-031-22997-8_1
Azimi Y., Liu Y., Tan T. C., Allen D. G., and Farnood R. R. (2017). The tail of two models: Impact of circularity and biomass non-homogeneity on UV disinfection of wastewater flocs. Water Res. 126, 70–78. doi: 10.1016/j.watres.2017.09.011
Baker-Austin C., Lake I., Archer E., Hartnell R., Trinanes J., and Martinez-Urtaza J. (2024). Stemming the rising tide of Vibrio disease. Lancet Planet Heal. 8, e515–e520. doi: 10.1016/S2542-5196(24)00124-4
Baker-Austin C., Oliver J. D., Alam M., Ali A., Waldor M. K., Qadri F., et al. (2018). Vibrio spp. infections. Nat. Rev. Dis. Prim. 41, 1–19. doi: 10.1038/s41572-018-0005-8
Baker-Austin C., Trinanes J., Gonzalez-Escalona N., and Martinez-Urtaza J. (2017). Non-cholera vibrios: the microbial barometer of climate change. Trends Microbiol. 25, 76–84. doi: 10.1016/j.tim.2016.09.008
Baker-Austin C., Trinanes J. A., Taylor N. G. H., Hartnell R., Siitonen A., and Martinez-Urtaza J. (2013). Emerging Vibrio risk at high latitudes in response to ocean warming. Nat. Clim. Change 3, 73–77. doi: 10.1038/nclimate1628
Bauermeister A., Bentchikou E., Moeller R., and Rettberg P. (2009). Roles of PprA, IrrE, and RecA in the resistance of Deinococcus radiodurans to germicidal and environmentally relevant UV radiation. Arch. Microbiol. 191, 913–918. doi: 10.1007/s00203-009-0522-7
Beck S. E., Ryu H., Boczek L. A., Cashdollar J. L., Jeanis K. M., Rosenblum J. S., et al. (2017). Evaluating UV-C LED disinfection performance and investigating potential dual-wavelength synergy. Water Res. 109, 207–216. doi: 10.1016/J.WATRES.2016.11.024
Blanchon C., Toulza E., Calvayrac C., Stavrakakis C., Eichendorff S., and Plantard G. (2025). Trials and in silico simulations to predict daily photo-oxidative capacity under solar conditions: A case study on the inactivation of an oyster pathogenic bacteria, Vibrio harveyi. J. Water Process Eng. 70, 107016. doi: 10.1016/J.JWPE.2025.107016
Bolton J. R. and Linden K. G. (2003). Standardization of methods for fluence (UV dose) determination in bench-scale UV experiments. J. Environ. Eng. 129, 209–215. doi: 10.1061/(ASCE)0733-9372(2003)129:3(209
Brumfield K. D., Chen A. J., Gangwar M., Usmani M., Hasan N. A., Jutla A. S., et al. (2023). Environmental Factors Influencing Occurrence of Vibrio parahaemolyticus and Vibrio vulnificus. Appl. Environ. Microbiol. 89, e00307-23. doi: 10.1128/AEM.00307-23/SUPPL_FILE/AEM.00307-23-S0001.PDF
Cerf O. (1977). Tailing of survival curves of bacterial spores. J. Appl. Bacteriol. 42, 1–19. doi: 10.1111/j.1365-2672.1977.tb00665.x
Cho J., Kim T., Cha D., Lee J. C., and Lee C. (2024). Inactivation of Edwardsiella tarda and Vibrio harveyi by Chlorination in Seawater. ACS ES T Eng. 4, 1158–1166. doi: 10.1021/ACSESTENGG.3C00584/ASSET/IMAGES/LARGE/EE3C00584_0006.JPEG
Das G., Sil K., and Das J. (1981). Repair of ultraviolet-light-induced DNA damage in Vibrio cholerae. BBA Sect. Nucleic Acids Protein Synth. 655, 413–420. doi: 10.1016/0005-2787(81)90053-8
de Souza Valente C. and Wan A. H. L. (2021). Vibrio and major commercially important vibriosis diseases in decapod crustaceans. J. Invertebr. Pathol. 181, 107527. doi: 10.1016/j.jip.2020.107527
Gandhi J. and Prakash H. (2023). Photo-disinfection processes for bacterial inactivation and underlying principles for water constituents’ impact: A review. Chem. Eng. J. Adv. 14, 100482. doi: 10.1016/J.CEJA.2023.100482
Geeraerd A. H., Valdramidis V. P., and Van Impe J. F. (2005). GInaFiT, a freeware tool to assess non-log-linear microbial survivor curves. Int. J. Food Microbiol. 102, 95–105. doi: 10.1016/j.ijfoodmicro.2004.11.038
Georgiades E., Scianni C., Davidson I., Tamburri M. N., First M. R., Ruiz G., et al. (2021). The role of vessel biofouling in the translocation of marine pathogens: management considerations and challenges. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.660125
González-Ortegón E. and Moreno-Andrés J. (2021). Anthropogenic modifications to estuaries facilitate the invasion of non-native species. Processes 9, 740. doi: 10.3390/pr9050740
Grimes D. J. (2020). The vibrios: scavengers, symbionts, and pathogens from the sea. Microb. Ecol. 80, 501–506. doi: 10.1007/s00248-020-01524-7
Guerra-Rodríguez S., Rodríguez E., Moreno-Andrés J., and Rodríguez-Chueca J. (2022). Effect of the water matrix and reactor configuration on Enterococcus sp. inactivation by UV-A activated PMS or H2O2. J. Water Process Eng. 47, 102740. doi: 10.1016/J.JWPE.2022.102740
Gyraitė G., Kataržytė M., Bučas M., Kalvaitienė G., Kube S., Herlemann D. P. R., et al. (2024). Epidemiological and environmental investigation of the ‘big four’ Vibrio species 1994 to 2021: a Baltic Sea retrospective study. Eurosurveillance 29, 2400075. doi: 10.2807/1560-7917.ES.2024.29.32.2400075
Hamamoto A., Bandou C., Nakano M., Mawatari K., Lian X., Yamato M., et al. (2010). Differences in stress response after UVC or UVA irradiation in Vibrio parahaemolyticus. Environ. Microbiol. Rep. 2, 660–666. doi: 10.1111/j.1758-2229.2010.00154.x
Hamza F. and Zinjarde S. (2023). Use of marine microorganisms in designing anti-infective strategies for sustainable aquaculture production. J. Appl. Microbiol. 134, 1–21. doi: 10.1093/JAMBIO/LXAD128
Hess-Erga O.-K., Moreno-Andrés J., Enger Ø., and Vadstein O. (2019). Microorganisms in ballast water: Disinfection, community dynamics, and implications for management. Sci. Total Environ. 657, 704–716. doi: 10.1016/J.SCITOTENV.2018.12.004
Hijnen W. A. M., Beerendonk E. F., and Medema G. J. (2006). Inactivation credit of UV radiation for viruses, bacteria and protozoan (oo)cysts in water: A review. Water Res. 40, 3–22. doi: 10.1016/j.watres.2005.10.030
Hoff J., Daniel B., Stukenberg D., Thuronyi B. W., Waldminghaus T., and Fritz G. (2020). Vibrio natriegens: an ultrafast-growing marine bacterium as emerging synthetic biology chassis. Environ. Microbiol. 22, 4394–4408. doi: 10.1111/1462-2920.15128
Jütte M., Abdighahroudi M. S., Waldminghaus T., Lackner S., and V Lutze H. (2023). Bacterial inactivation processes in water disinfection – mechanistic aspects of primary and secondary oxidants – A critical review. Water Res. 231, 119626. doi: 10.1016/J.WATRES.2023.119626
Keller L. C. and Maxcy R. B. (1984). Effect of physiological age on radiation resistance of some bacteria that are highly radiation resistant. Appl. Environ. Microbiol. 47, 915–918. doi: 10.1128/aem.47.5.915-918.1984
Koutsoumanis K., Allende A., Alvarez-Ordóñez A., Bolton D., Bover-Cid S., Chemaly M., et al. (2024). Public health aspects of Vibrio spp. related to the consumption of seafood in the EU. EFSA J. 22, e8896. doi: 10.2903/J.EFSA.2024.8896
Kuroyanagi Y., Tsuchiya J., Jiang C., Mino S., Kasai H., Motooka D., et al. (2022). Light response of Vibrio parahaemolyticus. Front. Mar. Sci. 9. doi: 10.3389/FMARS.2022.1037594/BIBTEX
Kyriazi A., Gounaki I., Mantzavinos D., and Venieri D. (2023). The intriguing role of the aqueous matrix in advanced water disinfection: can a harmonic relationship be achieved? J. Chem. Technol. Biotechnol. 99, 2113–2118 doi: 10.1002/JCTB.7340
Li X., Cai M., Wang L., Niu F., Yang D., and Zhang G. (2019). Evaluation survey of microbial disinfection methods in UV-LED water treatment systems. Sci. Total Environ. 659, 1415–1427. doi: 10.1016/J.SCITOTENV.2018.12.344
Li G. Q., Wang W. L., Huo Z. Y., Lu Y., and Hu H. Y. (2017). Comparison of UV-LED and low pressure UV for water disinfection: Photoreactivation and dark repair of Escherichia coli. Water Res. 126, 134–143. doi: 10.1016/j.watres.2017.09.030
Lim J. H. and Kang J. W. (2024). Assessing biofilm formation and resistance of vibrio parahaemolyticus on UV-aged microplastics in aquatic environments. Water Res. 254, 121379. doi: 10.1016/J.WATRES.2024.121379
Lindenauer K. G. and Darby J. L. (1994). Ultraviolet disinfection of wastewater: Effect of dose on subsequent photoreactivation. Water Res. 28, 805–817. doi: 10.1016/0043-1354(94)90087-6
Lo L. S. H., Liu X., Liu H., Shao M., Qian P. Y., and Cheng J. (2023). Aquaculture bacterial pathogen database: Pathogen monitoring and screening in coastal waters using environmental DNA. Water Res. X 20, 100194. doi: 10.1016/J.WROA.2023.100194
Lomelí-Ortega C. O. and Martínez-Díaz S. F. (2014). Phage therapy against Vibrio parahaemolyticus infection in the whiteleg shrimp (Litopenaeus vannamei) larvae. Aquaculture 434, 208–211. doi: 10.1016/j.aquaculture.2014.08.018
Lv B., Zhu G., Tian W., Guo C., Lu X., Han Y., et al. (2023). The prevalence of potential pathogens in ballast water and sediments of oceangoing vessels and implications for management. Environ. Res. 218, 114990. doi: 10.1016/J.ENVRES.2022.114990
Ma J., Meng L., Wang S., Li J., and Mao X. (2023). Inactivation of Vibrio parahaemolyticus and retardation of quality loss in oyster (Crassostrea gigas) by ultrasound processing during storage. Food Res. Int. 168, 112722. doi: 10.1016/J.FOODRES.2023.112722
MacIsaac S. A., Reid B., Ontiveros C., Linden K. G., Stoddart A. K., and Gagnon G. A. (2024). UV LED wastewater disinfection: The future is upon us. Water Res. X 24, 100236. doi: 10.1016/J.WROA.2024.100236
Malayeri A. H., Mohseni M., Cairns B., and Bolton J. R. (2016). Fluence (UV dose) required to achieve incremental log inactivation of bacteria, protozoa, viruses and algae. IUVA News 18, 4–6.
Martín-Sómer M., Pablos C., Adán C., van Grieken R., and Marugán J. (2023). A review on LED technology in water photodisinfection. Sci. Total Environ. 885, 163963. doi: 10.1016/J.SCITOTENV.2023.163963
Moreno-Andrés J., Acevedo-Merino A., and Nebot E. (2018). Study of marine bacteria inactivation by photochemical processes: disinfection kinetics and growth modeling after treatment. Environ. Sci. pollut. Res. 25, 27693–27703. doi: 10.1007/s11356-017-1185-6
Moreno-Andrés J., Morillo-Ponce J., Ibáñez-López M. E., Acevedo-Merino A., and García-Morales J. L. (2020). Disinfection enhancement of single ozonation by combination with peroxymonosulfate salt. J. Environ. Chem. Eng. 8, 104335. doi: 10.1016/j.jece.2020.104335
Moreno-Andrés J., Tierno-Galán M., Romero-Martínez L., Acevedo-Merino A., and Nebot E. (2023). Inactivation of the waterborne marine pathogen Vibrio alginolyticus by photo-chemical processes driven by UV-A, UV-B, or UV-C LED combined with H2O2 or HSO5–. Water Res. 232, 119686. doi: 10.1016/J.WATRES.2023.119686
Mougin J., Midelet G., Leterme S., Best G., Ells T., Joyce A., et al. (2023). Benzalkonium chloride disinfectant residues stimulate biofilm formation and increase survival of Vibrio bacterial pathogens. Front. Microbiol. 14. doi: 10.3389/fmicb.2023.1309032
Nebot E., Salcedo Dávila I., Andrade Balao J. A., and Quiroga Alonso J. M. (2007). Modelling of reactivation after UV disinfection: Effect of UV-C dose on subsequent photoreactivation and dark repair. Water Res. 41, 3141–3151. doi: 10.1016/j.watres.2007.04.008
Ng C., Goh S. G., Saeidi N., Gerhard W. A., Gunsch C. K., and Gin K. Y. H. (2018). Occurrence of Vibrio species, beta-lactam resistant Vibrio species, and indicator bacteria in ballast and port waters of a tropical harbor. Sci. Total Environ. 610–611, 651–656. doi: 10.1016/j.scitotenv.2017.08.099
Nyangaresi P. O., Qin Y., Chen G., Zhang B., Lu Y., and Shen L. (2018). Effects of single and combined UV-LEDs on inactivation and subsequent reactivation of E. coli in water disinfection. Water Res. 147, 331–341. doi: 10.1016/j.watres.2018.10.014
Oguma K., Katayama H., Mitani H., Morita S., Hirata T., and Ohgaki S. (2001). Determination of pyrimidine dimers in Escherichia coli and Cryptosporidium parvum during UV light inactivation, photoreactivation, and dark repair. Appl. Environ. Microbiol. 67, 4630–4637. doi: 10.1128/AEM.67.10.4630
Oguma K., Kita R., Sakai H., Murakami M., and Takizawa S. (2013). Application of UV light emitting diodes to batch and flow-through water disinfection systems. Desalination 328, 24–30. doi: 10.1016/J.DESAL.2013.08.014
Oppezzo O. J., Abrevaya X. C., and Giacobone A. F. F. (2024). An alternative interpretation for tailing in survival curves for bacteria exposed to germicidal radiation. Photochem. Photobiol. 100, 129–136. doi: 10.1111/PHP.13808
Pousty D., Hofmann R., Gerchman Y., and Mamane H. (2021). Wavelength-dependent time–dose reciprocity and stress mechanism for UV-LED disinfection of Escherichia coli. J. Photochem. Photobiol. B Biol. 217, 112129. doi: 10.1016/J.JPHOTOBIOL.2021.112129
Pumkaew M., Taweephitakthai T., Satanwat P., Yocawibun P., Chumtong P., Pungrasmi W., et al. (2021). Use of ozone for Vibrio parahaemolyticus inactivation alongside nitrification biofilter treatment in shrimp-rearing recirculating aquaculture system. J. Water Process Eng. 44, 102396. doi: 10.1016/J.JWPE.2021.102396
Quek P. H. and Hu J. (2008). Indicators for photoreactivation and dark repair studies following ultraviolet disinfection. J. Ind. Microbiol. Biotechnol. 35, 533–541. doi: 10.1007/s10295-008-0314-0
Rattanakul S. and Oguma K. (2018). Inactivation kinetics and efficiencies of UV-LEDs against Pseudomonas aeruginosa, Legionella pneumophila, and surrogate microorganisms. Water Res. 130, 31–37. doi: 10.1016/J.WATRES.2017.11.047
Romero-Martínez L., Duque-Sarango P., González-Martín C., Moreno-Andrés J., Acevedo-Merino A., and Nebot E. (2023). Inactivation efficacy and reactivation of fecal bacteria with a flow-through LED ultraviolet reactor: Intraspecific response prevails over interspecific differences. J. Water Process Eng. 52, 103497. doi: 10.1016/J.JWPE.2023.103497
Romero-Martínez L., Moreno-Andrés J., Acevedo-Merino A., and Nebot E. (2022). Development of a geometrical model for the determination of the average intensity in a flow-through UV-LED reactor and validation with biodosimetry and actinometry. J. Water Process Eng. 49, 103137. doi: 10.1016/J.JWPE.2022.103137
Serna-Galvis E. A., Troyon J. A., Giannakis S., Torres-Palma R. A., Minero C., Vione D., et al. (2018). Photoinduced disinfection in sunlit natural waters: Measurement of the second order inactivation rate constants between E. coli and photogenerated transient species. Water Res. 147, 242–253. doi: 10.1016/j.watres.2018.10.011
Seymour J. R. and McLellan S. L. (2025). Climate change will amplify the impacts of harmful microorganisms in aquatic ecosystems. Nat. Microbiol. 10, 615–626. doi: 10.1038/s41564-025-01948-2
Siboni N., King W. L., Williams N. L. R., Scanes E., Giardina M., Green T. J., et al. (2024). Increased abundance of potentially pathogenic Vibrio and a marine heatwave co-occur with a Pacific Oyster summer mortality event. Aquaculture 583, 740618. doi: 10.1016/J.AQUACULTURE.2024.740618
Sinha R. P. and Häder D. P. (2002). UV-induced DNA damage and repair: a review. Photochem. Photobiol. Sci. 1, 225–236. doi: 10.1039/B201230H
Sony M., Sumithra T. G., Anusree V. N., Amala P. V., Reshma K. J., Alex S., et al. (2021). Antimicrobial resistance and virulence characteristics of Vibrio vulnificus, Vibrio parahaemolyticus and Vibrio harveyi from natural disease outbreaks of marine/estuarine fishes. Aquaculture 539, 736608. doi: 10.1016/J.AQUACULTURE.2021.736608
Sun W., Jing Z., Zhao Z., Yin R., Santoro D., Mao T., et al. (2023). Dose-response behavior of pathogens and surrogate microorganisms across the ultraviolet-C spectrum: inactivation efficiencies, action spectra, and mechanisms. Environ. Sci. Technol. 57, 10891–10900. doi: 10.1021/acs.est.3c00518
Tiwari A., Kauppinen A., and Pitkänen T. (2019). Decay of Enterococcus faecalis, vibrio cholerae and MS2 coliphage in a laboratory mesocosm under brackish beach conditions. Front. Public Heal. 7. doi: 10.3389/fpubh.2019.00269
Trinanes J. and Martinez-Urtaza J. (2021). Future scenarios of risk of Vibrio infections in a warming planet: a global mapping study. Lancet Planet Heal. 5, e426–e435. doi: 10.1016/S2542-5196(21)00169-8
UNEP (2019). Minamata convention on mercury. Available online at: www.mercuryconvention.org (Accessed February 15, 2025).
Vitzilaiou E., Kuria A. M., Siegumfeldt H., Rasmussen M. A., and Knøchel S. (2021). The impact of bacterial cell aggregation on UV inactivation kinetics. Water Res. 204, 117593. doi: 10.1016/J.WATRES.2021.117593
Wang M., Ateia M., Awfa D., and Yoshimura C. (2021a). Regrowth of bacteria after light-based disinfection — What we know and where we go from here. Chemosphere 268, 128850. doi: 10.1016/J.CHEMOSPHERE.2020.128850
Wang W., Xie H., Li G., Li J., Wong P. K., and An T. (2021b). Visible light-induced marine bacterial inactivation in seawater by an in situ photo-fenton system without additional oxidants: implications for ballast water sterilization. ACS ES&T Water. 1, 1483–1494. doi: 10.1021/acsestwater.1c00048
Wennberg A. C., Tryland I., Østensvik Ø., Secic I., Monshaugen M., and Liltved H. (2013). Effect of water treatment on the growth potential of Vibrio cholerae and Vibrio parahaemolyticus in seawater. Mar. Environ. Res. 83, 10–15. doi: 10.1016/j.marenvres.2012.10.002
Winfield M. D. and Groisman E. A. (2003). Role of nonhost enviroments in the lifestyles of Salmonella and E. coli. Appl. Environ. Microbiol. 69, 3687–3694. doi: 10.1128/AEM.69.7.3687
Wu D., You H., Zhang R., Chen C., and Lee D. J. (2011). Ballast waters treatment using UV/Ag-TiO2+O3 advanced oxidation process with Escherichia coli and Vibrio alginolyticus as indicator microorganisms. Chem. Eng. J. 174, 714–718. doi: 10.1016/j.cej.2011.09.087
Xiao Y., Chu X. N., He M., Liu X. C., and Hu J. Y. (2018). Impact of UVA pre-radiation on UVC disinfection performance: Inactivation, repair and mechanism study. Water Res. 141, 279–288. doi: 10.1016/j.watres.2018.05.021
Yang B., Zhai S., Li X., Tian J., Li Q., Shan H., et al. (2021). Identification of Vibrio alginolyticus as a causative pathogen associated with mass summer mortality of the Pacific Oyster (Crassostrea gigas) in China. Aquaculture 535, 736363. doi: 10.1016/J.AQUACULTURE.2021.736363
Keywords: marine disinfection, emerging waterborne pathogens, bacterial repair, LED-based water treatment, species-specific inactivation, aquaculture biosecurity, marine pathogen control
Citation: Romero-Martínez L, Prieto-Pérez-Juez A, Nebot E and Moreno-Andrés J (2025) UV-LED disinfection of emerging Vibrio pathogens: inactivation kinetics and reactivation potential. Front. Mar. Sci. 12:1608367. doi: 10.3389/fmars.2025.1608367
Received: 09 April 2025; Accepted: 09 September 2025;
Published: 30 September 2025.
Edited by:
Brett Froelich, George Mason University, United StatesReviewed by:
Neeraj Kumar, National Institute of Abiotic Stress Management (ICAR), IndiaCharmaine Cheuk Man Yung, Hong Kong University of Science and Technology, Hong Kong SAR, China
Copyright © 2025 Romero-Martínez, Prieto-Pérez-Juez, Nebot and Moreno-Andrés. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Javier Moreno-Andrés, amF2aWVyLm1vcmVub0B1Y2EuZXM=