Abstract
The decline in common squid (Todarodes pacificus) stocks, combined with increasing environmental variability, underscores the need for science-based approaches to support adaptive fisheries management in Korea. This study aimed to standardize the catch per unit effort (CPUE) of common squid in Korean waters, specifically in the offshore jigging fishery, by incorporating spatiotemporal and sea surface temperature (SST) factors, and to assess future shifts in thermally suitable fishing grounds under the shared socioeconomic pathway 3-7.0 (SSP3-7.0) climate change scenario. A generalized additive model (GAM) revealed a significant nonlinear relationship between CPUE and SST, with squid abundance concentrated within the 13–23 °C range. The peak response was observed around 21 °C, which was identified as the species’ optimal temperature. We applied this preferred thermal range to projected SST data for 2050 and 2100 under the SSP3-7.0 scenario to map the spatial extent of thermally suitable habitats. Results indicated a contraction of suitable fishing areas during the main fishing season (September–October), while suitable temperatures reemerged in November, suggesting a potential delay in fishing ground formation. These findings highlight the importance of flexible fishery scheduling and management criteria in response to climate-driven changes in habitat suitability. This study provides foundational insights for spatially explicit resource management and the development of climate-adaptive fishery strategies.
1 Introduction
Climate change has a profound impact on the marine environment, and the sixth assessment report (AR6) by the Intergovernmental Panel on Climate Change (IPCC) projects that ocean warming will continue throughout the 21st century under the Shared Socioeconomic Pathways (SSP) scenarios (IPCC, 2023). In particular, rising sea temperatures are a major driver of ecological responses in marine organisms (Brander, 2007; Pörtner and Peck, 2010), altering population maintenance and reproductive strategies, and ultimately leading to shifts in habitats or distribution ranges (Cheung et al., 2009; Perry et al., 2010). These responses vary by species, depending on physiological tolerance and behavioral traits, with species more sensitive to environmental change exhibiting a greater likelihood of distributional shifts (Poloczanska et al., 2013; Brander et al., 2017). In fact, changes in the distribution of various commercially important species have been observed in response to ocean warming, affecting fishing activities and resource utilization patterns (Perry et al., 2005; Cruz et al., 2024). The IPCC anticipates regional variations in catch potential (IPCC, 2023), underscoring the importance of scientific forecasting and adaptive strategies (FAO, 2021; Bueno-Pardo et al., 2021).
The Korean coastal and offshore waters, particularly the eastern waters of Korea, represent a transitional zone where the subtropical Tsushima warm current and the subarctic Liman cold current converge, resulting in pronounced ecological responses to sea temperature changes (Tian et al., 2008). Within these environmental conditions, common squid (Todarodes pacificus) is recognized as a representative climatesensitive species (Pecl and Jackson, 2008; Sakurai et al., 2000; Song, 2018). As a short-lived, fast-growing, and highly migratory species, T. pacificus exhibits substantial shifts in its distribution range and fishing ground centers in response to changes in ocean conditions. In particular, in the eastern waters of Korea, the species responds sensitively to variations in water temperature and the position of thermal fronts, resulting in distinct seasonal and interannual differences in the spatial distribution of fishing grounds (Cho et al., 2004; Choi et al., 2008; Kim et al., 2014).
Common squid is classified into autumn, winter, and summer spawning cohorts based on spawning season, among which the autumn spawning cohort constitutes the primary population caught in Korean waters. This cohort predominantly migrates to the eastern waters of Korea, where it is mainly harvested from September to December by multiple fisheries, including offshore jigging, large trawl, and eastern sea trawl (National Institute of Fisheries Science, 2021; Statistics Korea, 2025). Based on the average catch over the past five years (2019–2023), the common squid ranked third among all species, underscoring its status as a commercially important resource (Statistics Korea, 2025). Catches increased rapidly during the 1990s, peaking at 250,000 tons in 1996. However, they gradually declined thereafter, followed by a sharp drop in 2015, reaching approximately 23,000 tons in 2023 (Statistics Korea, 2025).
The decline in T. pacificus catches has been attributed to a complex interplay of factors, including changes in the fishing environment, technological advancements, overfishing, and broader shifts in the marine ecosystem (Kim et al., 2011; Jo et al., 2019). In particular, increased fishing effort due to the development of automatic jigging machines and advanced navigation systems (Oh et al., 2018), cooperative operations between offshore jigging and trawl fisheries (Choi et al., 2008), and large-scale illegal and excessive fishing by Chinese vessels (Lee et al., 2017) have all contributed to the acceleration of resource depletion. Moreover, marine environmental changes driven by climate change are believed to influence both the distribution and population dynamics of the species, further contributing to the decline in catches (Sakurai et al., 2002; Rosa et al., 2011; Kim et al., 2018). Accordingly, there is a growing need for research on distribution forecasting and systematic population assessments that incorporate the impacts of climate-driven marine environmental changes, in order to improve the management of T. pacificus resources.
Catch per unit effort (CPUE) is a key indicator of the relative abundance of a resource and is often derived from commercial fishing data due to the difficulty of obtaining long-term scientific data (Bishop et al., 2004; Maunder and Punt, 2004). However, CPUE is affected by various factors—including oceanographic conditions, fishing gear, and fishing time—necessitating standardization through statistical adjustments for environmental and operational variables (Hinton and Maunder, 2004). In this process, it is important to account for spatial variation across fishing grounds by stratifying the survey area into discrete units and incorporating them as categorical variables (Hsu et al., 2022; Shi et al., 2023). Generalized additive models (GAMs) are particularly effective for incorporating multiple factors, as they flexibly capture nonlinear relationships between environmental variables and spatial structure (Ducharme-Barth et al., 2022).Due to these characteristics, GAMs have been widely applied to fishery survey data to investigate how environmental factors influence the distribution patterns of marine species (Winter et al., 2007; Murase et al., 2009).
In recent years, there has been growing interest in quantitatively predicting climate-driven shifts in fishing grounds. In particular, studies have increasingly utilized IPCC climate change scenarios (e.g., Representative Concentration Pathways (RCPs) and SSPs) to forecast the future distribution of marine species under projected changes in sea temperature (Lima et al., 2022; Ben Lamine et al., 2022; Lewis et al., 2023; Xiong et al., 2024). By integrating ecological responses of marine species with climate projections, this approach enables the prediction of spatial shifts in potential fishing grounds and provides a scientific basis for the development of resource management strategies. While climate scenario-based vulnerability assessments have been conducted in Korea’s fisheries sector (Kim et al., 2023, 2024b), studies that specifically predict spatial shifts in species distributions or fishing ground centers remain limited. Among available climate pathways, the SSP3-7.0 scenario was selected because the sea surface temperature (SST) trends from the CNRM-ESM2-1-based Regional Ocean Modeling System (ROMS) model closely matched observed temperature variations from Korea’s KODC dataset for the 2015–2024 period, making it the most appropriate scenario for hindcasting and near-future projection (Kim et al., 2024a).
In Korea, several studies have examined the relationship between the biological characteristics of T. pacificus and the marine environment (Kim et al., 2011), investigated the influence of climate indices on spawning ground conditions (Kim et al., 2018), and conducted stock assessments using Bayesian state-space production models based on catch data (An et al., 2021; Gim et al., 2022). However, these studies have primarily addressed catch variability and environmental factors separately, or have only partially accounted for the spatial characteristics of CPUE. In particular, to our knowledge, there has been no prior research that integrates standardized CPUE with climate scenario-based SST projections to predict spatial shifts in fishing ground distribution or to estimate the extent of suitable fishing areas for T. pacificus in Korean waters.
We hypothesize that the abundance of T. pacificus (as reflected in CPUE) is influenced by spatiotemporal variability and sea surface temperature, and that the species exhibits a preferred thermal range, with suitable habitats expected to shift geographically under future climate scenarios. This hypothesis is consistent with previous findings for T. pacificus and other squid species in various regions, which have reported SST and season dependent changes in distribution. The novelty of this study lies in applying this established ecological framework specifically to the Korean offshore jigging fishery and in explicitly integrating the preferred thermal range, estimated from standardized, fishery-dependent CPUE, with scenario-based SST projections from a high-resolution regional ocean model for Korean waters.
In this study, a GAM was applied to standardize the CPUE of the offshore jigging fishery for common squid by incorporating operational and SST-related covariates. We also estimated the species’ preferred thermal range from GAM analysis, which was then used to identify suitable fishing grounds under future SST conditions projected by the SSP3-7.0 climate scenario. Our study has three main objectives: (1) To identify the effects of spatiotemporal and SST variability on the CPUE of T. pacificus; (2) To estimate the species’ preferred temperature range based on GAM analysis; (3) To project spatial shifts in suitable fishing grounds under the SSP3-7.0 climate scenario for 2050 and 2100. The results of this study aim to provide a scientific basis for adaptive fishery management of T. pacificus, particularly by informing decisions related to fishing season adjustments, spatial management, and future stock assessments in Korean waters.
2 Materials and methods
The overall analytical process in this study consists of five sequential stages: (1) data collection and preprocessing, (2) spatial clustering, (3) CPUE standardization using GAM, (4) thermal preference estimation, and (5) future habitat projection under the SSP3-7.0 climate scenario. These components are detailed in the following subsections (Figure 1).
Figure 1
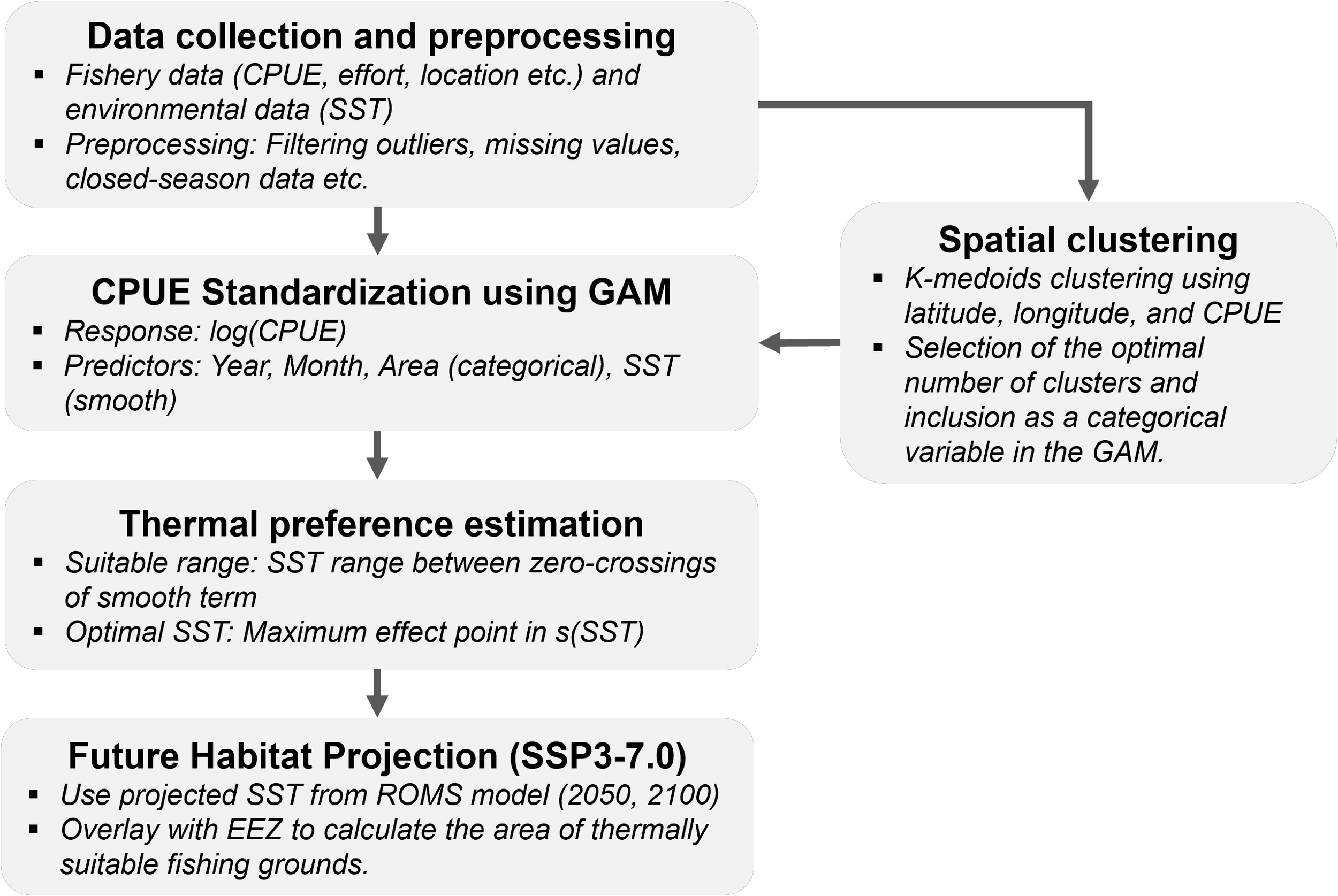
Flow chart of this study.
2.1 Data
In Korea, the offshore jigging fishery for common squid is primarily conducted in the eastern waters of Korea. Following the Korea–Russia fisheries cooperation, fishing activities by Korean offshore jigging vessels have also become active within the Russian exclusive economic zone (EEZ), particularly from July to October since 2001. Accordingly, this study incorporates both the eastern part of the Korean EEZ and Russian waters as the analysis area to reflect the operational characteristics of the fishery.
Catch data for common squid were obtained from a sampling survey conducted by the National Institute of Fisheries Science (NIFS). The dataset includes monthly catch records of offshore jigging in 0.5° × 0.5° grid cells, covering the period from January 1994 to December 2024. The data consist of fishing date (year and month), fishing location (latitude and longitude), fishing effort (number of jigging machines), and catch (kg). CPUE was calculated as the total catch divided by the number of jigging machines for each cell of the grid (Equation 1):
where Ci is the catch (kg) and Ei is the number of jigging machines in grid cell i.
To ensure the reliability of the dataset for common squid, several preprocessing steps were conducted. First, since small sample sizes can compromise the robustness of statistical analyses (Riley et al., 2019), grid cells with fewer than 30 CPUE observations were excluded. Second, records identified as outliers through boxplot analysis, zero CPUE values, and entries with missing data were removed. Third, to account for potential bias introduced by regulatory restrictions, fishing activities conducted during the closed season in April were excluded as part of seasonal adjustment. Following this procedure, a total of 85 grid cells were selected, and 8,664 observations were used in the analysis. Based on the preprocessed dataset, the response variable for CPUE standardization was defined as CPUE (catch per unit effort), where effort was measured by the number of jigging machines operating per observation, and calculated monthly for each grid cell. The explanatory variables used in the GAM included three spatiotemporal factors—Year, Month, and Area (derived through spatial clustering)—and one environmental factor, SST, which was treated as a continuous variable (Table 1).
Table 1
| Factor | Type | Explanatory variable | Description |
|---|---|---|---|
| Spatiotemporal factors | Categorical | Year | 21 years from 1994 to 2024) |
| Categorical | Month | 11 months, excluding the closed season (April) | |
| Categorical | Area | Classified into two spatial clusters | |
| Environmental factors | Continuous | s(SST) | Sea surface temperature (1994–2024), extracted from ROMS-based ocean climate model Kim et al. (2024a) |
Explanatory variables used in the GAM-based CPUE standardization model for common squid Todarodes pacificus..
2.2 Spatial clustering
Based on the spatial analysis conducted in this study, the study area was divided into two distinct subareas. When fishing locations are non-randomly selected, CPUE data collected from preferred fishing grounds may exert a disproportionate influence on the estimation of fish abundance (Conn et al., 2017; Hsu et al., 2022). To address this issue, a spatial clustering approach based on the k-medoids algorithm, as proposed by (Ono et al., 2015), was applied. The resulting stratified areas were used as a categorical explanatory variable in the CPUE standardization model. For the clustering process, latitude, longitude, and CPUE data of T. pacificus were used as input variables. The k-medoids algorithm assigns cluster centers as medoids—actual data points that minimize the average dissimilarity within each cluster. Euclidean distance was used to measure spatial dissimilarity between grids, and the center of each cluster was defined as the data point with the smallest average dissimilarity to all others in the same cluster.
To determine the optimal number of spatial clusters, the silhouette score was calculated, which evaluates both within-cluster cohesion and between-cluster separation (Rousseeuw, 1987). Clustering analyses were conducted for k values ranging from 2 to 5, and the silhouette score was computed for each configuration to identify the one with the highest score. The silhouette score ranges from –1 to 1, with higher values indicating better-defined and more cohesive clustering.
2.3 Sea surface temperature
Sea temperature is a key environmental factor that strongly influences the spawning and distribution of marine species (Kim et al., 2019). Common squid is primarily distributed within the upper water column, inhabiting depths of up to approximately 50 meters (Jo et al., 2019). Acoustic survey data and catch-depth analyses further indicate a pronounced tendency for the species to concentrate in surface layers shallower than 18 meters (Lee et al., 2009). Moreover, elevated sea surface temperatures are generally associated with increased catch rates, whereas lower temperatures are linked to reduced catches (Sakurai et al., 2000), indicating that common squid is particularly sensitive to variations in surface temperature. Accordingly, SST at a depth of 5 meters was selected as the representative environmental variable, as it best corresponds to the species’ primary habitat depth. However, due to limitations in the availability of consistent, long-term datasets, other environmental drivers such as salinity, zooplankton abundance, and ocean currents were not included in the analysis.
Historical and projected SST used in the analysis were derived from regional ocean climate model outputs developed by (Kim et al., 2024a), covering the period from 1994 to 2100. The regional ocean climate model was developed based on the ROMS and was configured to simulate ocean circulation in the waters surrounding the Korean Peninsula. Boundary conditions for ROMS were derived from global climate model outputs from Coupled Model Intercomparison Project Phase 6 (CMIP6), as included in the IPCC AR6 (IPCC, 2023), allowing for high-resolution downscaling of large-scale climate projections. The CMIP6 model employed was CNRM-ESM2-1, developed by the Centre National de Recherches Met´ eorologiques´ (CNRM), which has demonstrated high performance in reproducing oceanic conditions in the Northwest Pacific region. For the historical period (1993–2014), the model replicated seasonal and interannual variability in the marine environment through historical simulations. From 2015 to 2100, the model produced three-dimensional oceanographic projections including daily mean temperature, salinity, and current velocity under four SSPs: SSP1-2.6, SSP2-4.5, SSP3-7.0, and SSP5-8.5.
Comparison between the ROMS model outputs based on CNRM-ESM2–1 and observational data from the Korea Oceanographic Data Center (KODC) of the National Institute of Fisheries Science revealed that the SSP3-7.0 scenario most closely captured the observed trends in sea temperature during the period from 2015 to 2024. Accordingly, sea temperature projections under the SSP3-7.0 scenario were used to represent historical temperature variability for the same period.
2.4 CPUE standardization and optimal temperature estimation using GAM
In this study, a GAM was applied to identify the key factors influencing variations in the CPUE of common squid and to perform CPUE standardization. The response variable was the log-transformed CPUE, and the model incorporated both spatiotemporal and SST variables. Year, Month, and Area were included as categorical variables and encoded as factors, while SST was treated as a continuous variable and modeled with a smooth term, to account for nonlinear effects (Table 1). The GAM was selected for its flexibility in handling both linear and nonlinear relationships among explanatory variables. In particular, its ability to incorporate smooth functions allowed us to capture the potentially nonlinear effect of SST on CPUE, while also controlling for categorical spatiotemporal factors. The GAM applied in this study can be expressed in the following general form (Equation 2):
Here, s(·) represents a smooth function applied to continuous predictors to model potential nonlinear effects. The residual error term ϵiis assumed to follow a normal distribution. To model the nonlinear effect of SST, it was included as a smoothing term s(SST). The model was fitted using the mgcv package in the R statistical software, and model residuals and smoothing parameter adequacy were evaluated using the gam.check function. Also, To enhance model selection transparency, five GAM candidates were evaluated using different combinations of the variables listed in Table 1. Each model was compared using Akaike Information Criterion (AIC), Bayesian Information Criterion (BIC), and mean square error (MSE). In addition, a 10 fold cross-validation was conducted for all models, and the cross-validated mean square error (CV-MSE) was calculated to assess predictive robustness.
To identify the temperature thresholds at which the influence of SST on CPUE transitions from positive to negative (or vice versa), the zero-crossing points of the smoothing function where the estimated smooth term changes sign were extracted. The first and last zero-crossing points were interpreted as the lower and upper limits of the suitable thermal range, respectively. In addition, the SST value corresponding to the maximum of the smoothing function was identified as the species’ optimal temperature. The derived suitable thermal range and optimal value were subsequently used as criteria for projecting climatically suitable fishing habitats under future scenarios.
2.5 Climate change scenario and future habitat prediction
Future oceanographic projections were based on SST outputs for the years 2050 and 2100 under the SSP3-7.0 scenario, derived from a ROMS-based regional climate model developed by (Kim et al., 2024a). SSP3-7.0 represents an “inequality” scenario characterized by persistent socioeconomic disparities and limited climate mitigation efforts, reflecting a medium- to high-emissions trajectory (Korea Meteorological Administration, 2019). Using the optimal temperature and suitable thermal range identified through the GAM analysis, the spatial extent of climatically suitable fishing habitats for common squid in Korean waters was projected under the SSP3-7.0 scenario for mid-century (2050) and end-century (2100) conditions.
Subsequently, by overlaying the projected thermally suitable habitats with the fishable area in the eastern part of the Korean EEZ (Figure 2), this study estimated potential shifts in the coastal jigging fishery’s fishing grounds in the eastern waters of Korea during the current peak fishing season (September to December) for the years 2050 and 2100. Figure 2 delineates the spatial boundary of the fishable area within the eastern Korean EEZ, which served as the geographic reference for quantifying thermally suitable fishing habitats under projected SST scenarios.
Figure 2
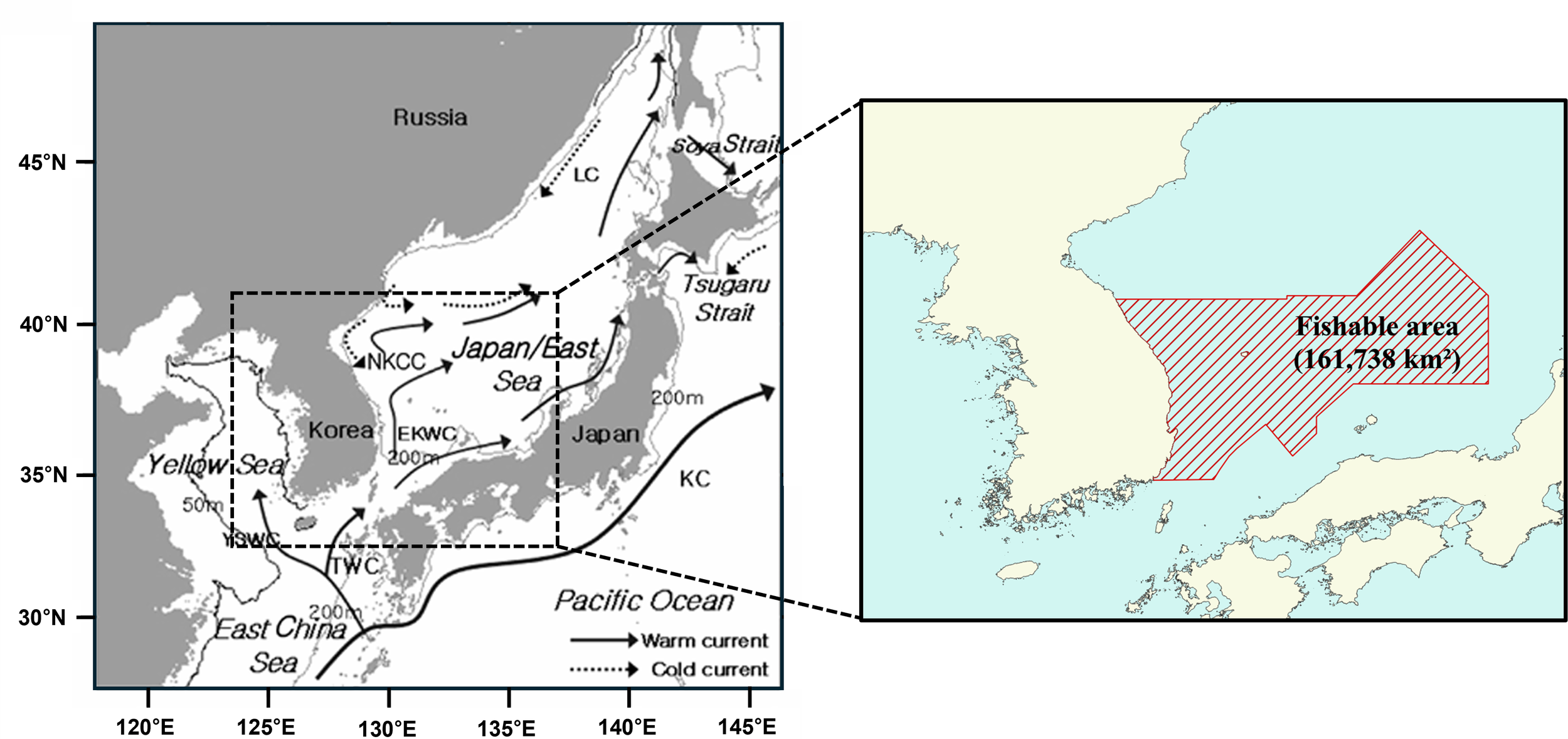
Geography and current system around the Korean Peninsula (left; modified from McFarlane et al. (2009)) and the fishable area in the eastern Korean exclusive economic zone (EEZ) designated as the analysis domain in this study (right).
3 Results
3.1 Clustering and validation of analytical data
Based on the silhouette analysis (Figure 3; Table 2), fishing locations were classified into two spatial clusters (k = 2), which yielded the highest silhouette score among the tested values. These clusters were then used as the categorical variable Area in the GAM model (Figure 4). As the assumptions of normality and homogeneity of variance were violated, a Kruskal–Wallis test was conducted instead of ANOVA. The results revealed a statistically significant difference in nominal CPUE between the two subareas (p < 0.0001), supporting that the CPUE-based spatial classification effectively captured underlying spatial heterogeneity in squid abundance.
Figure 3
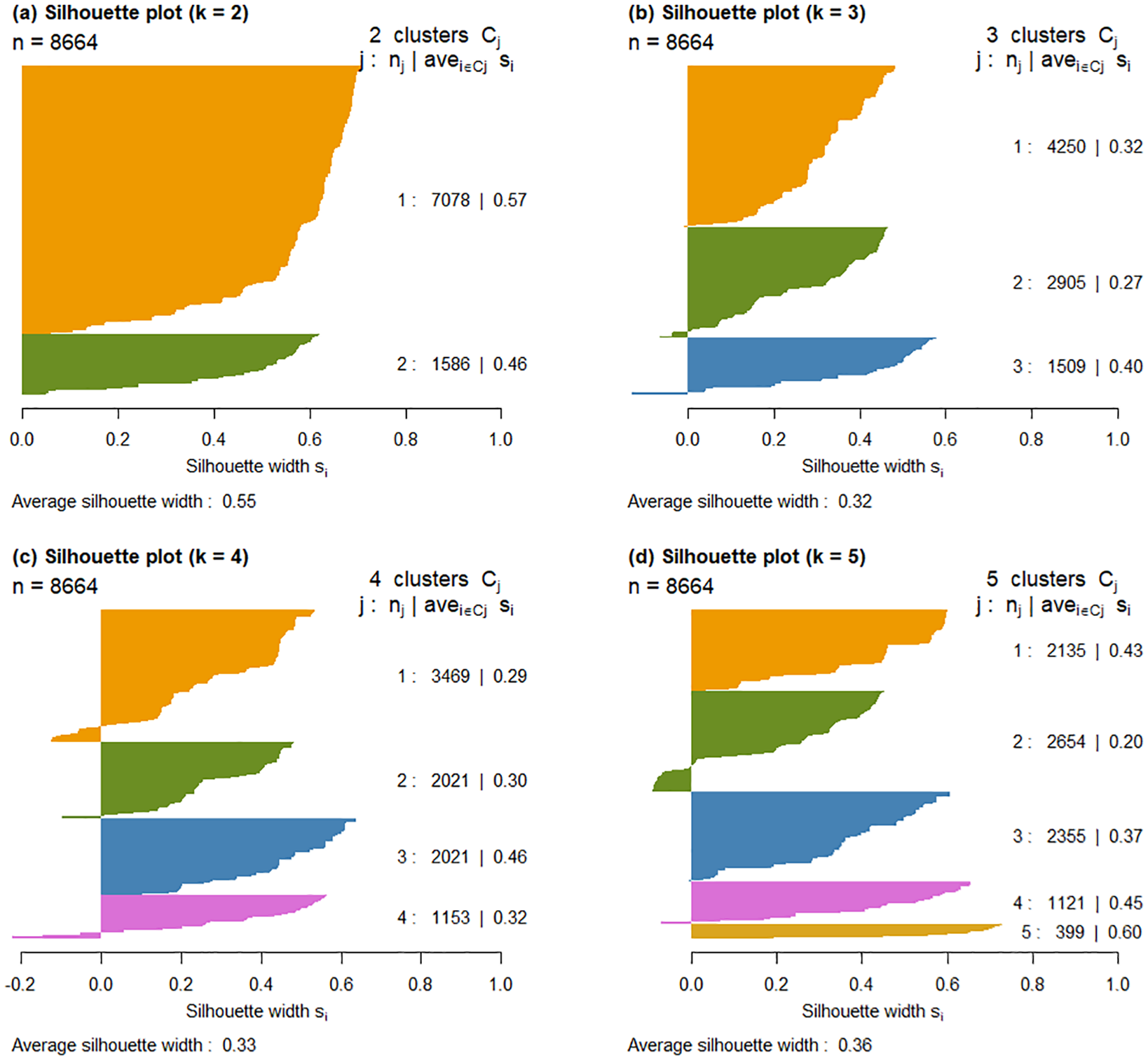
Silhouette score plots for spatial clustering of the jigging fishery’s fishing grounds for common squid Todarodes pacificus using k-medoids clustering: (a) 2 clusters, (b) 3 clusters, (c) 4 clusters, (d) 5 clusters.
Table 2
| The number of cluster (k) | k=2 | k=3 | k=4 | k=5 |
|---|---|---|---|---|
| Silhouette score | 0.552 | 0.317 | 0.333 | 0.355 |
Silhouette scores for k-medoids clustering with 2 to 5 clusters applied to offshore jigging data for common squid.
Figure 4
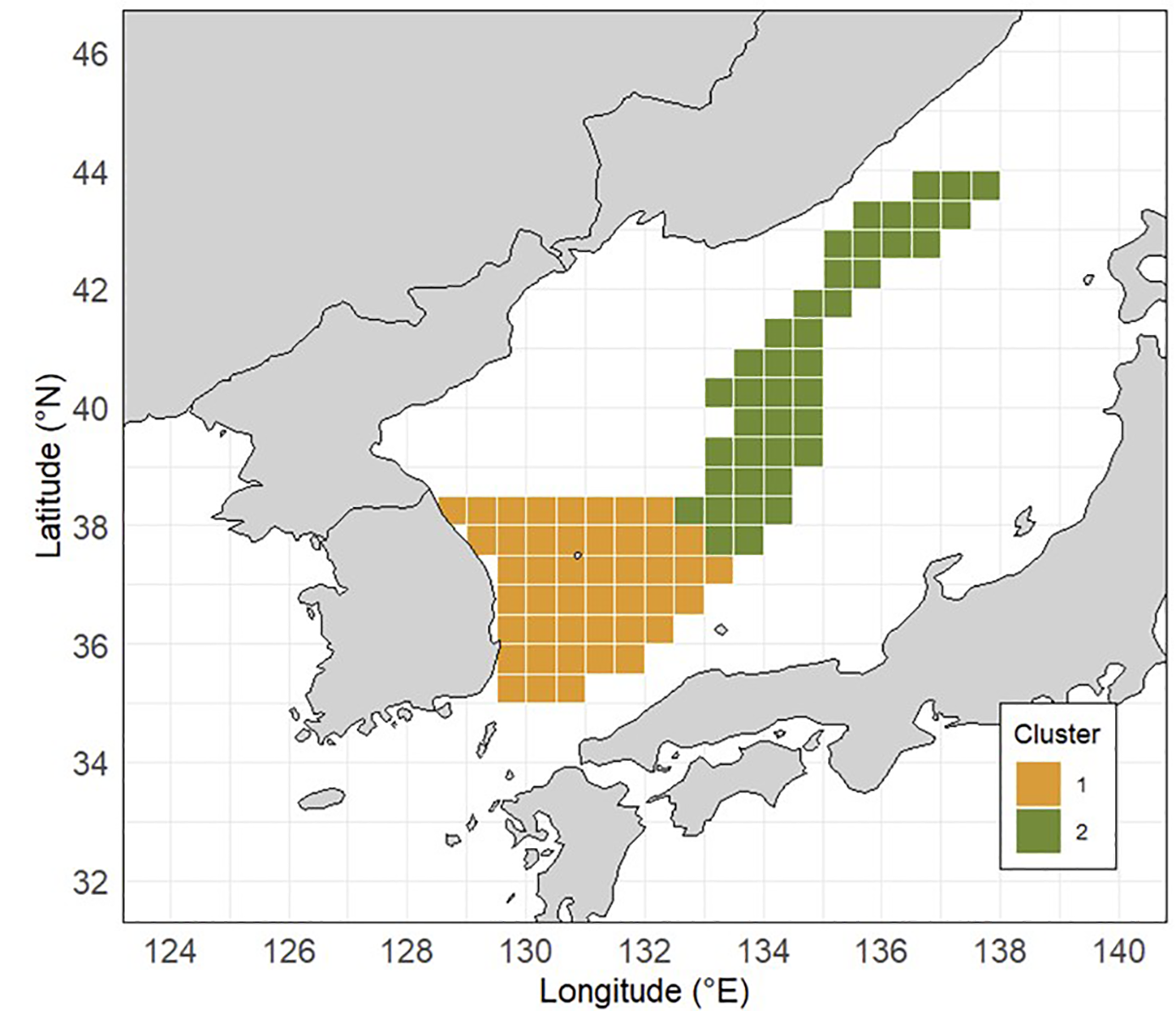
Spatial stratification of the jigging fishery’s fishing grounds for common squid Todarodes pacificus based on k-medoids clustering using catch location data from 1994 to 2021. The fishing area was classified into two clusters (Area 1 and Area 2).
To assess multicollinearity among the explanatory variables, variance inflation factors (VIFs) were calculated. All VIF values were below 3 (1.010–1.286), well below the commonly used threshold of 10, indicating that multicollinearity was not a concern in the model.
3.2 CPUE standardization and effect of sea surface temperature
Model selection results comparing five GAM configurations are presented in Table 3. Among the five tested models, Model 1, which incorporated all covariates, yielded the lowest AIC and BIC values (73,234 and 73,589, respectively), as well as the smallest MSE (271.30) and CV-MSE (305.36). Based on these results, Model 1 was selected as the final model structure for CPUE standardization in this study.
Table 3
| Model | GAM formula | AIC | BIC | MSE | CV- MSE |
|---|---|---|---|---|---|
| Model 1 | ln(CPUE) ˜ Year + Month + Area + s(SST) | 73,234 | 73,589 | 271 | 305 |
| Model 2 | ln(CPUE) ˜ Year + Month + s(SST) | 74,428 | 74,778 | 311 | 337 |
| Model 3 | ln(CPUE) ˜ Year + s(SST) | 76,179 | 76,463 | 382 | 390 |
| Model 4 | ln(CPUE) ˜ Year + Month + Area | 73,485 | 73,789 | 280 | 313 |
| Model 5 | ln(CPUE) ˜ Year + Month | 74,653 | 74,950 | 320 | 345 |
Formulas and evaluation results of five generalized additive models (GAMs) used for standardizing CPUE, including Akaike Information Criterion (AIC), Bayesian Information Criterion (BIC), mean square error (MSE), and cross-validated mean square error (CV-MSE) from 10-fold cross-validation.
The GAM explained approximately 45.4% of the deviance in CPUE. SST showed a significant nonlinear effect on CPUE variation (p < 0.001), with an effective degrees of freedom (edf) of 6.58, indicating a complex, nonlinear relationship rather than a simple linear trend. All parametric terms in the model were statistically significant (mostly at p < 0.001). Model diagnostics indicated that the smoother was appropriately specified without overfitting or underfitting. The GAM diagnostic check using gam.check confirmed that the smoother basis dimension was suitable (k-index = 0.98, p = 0.07).
Visualization of the relationship between SST and CPUE showed that the smooth term s(SST) remained positive between approximately 13 °C and 23 °C, indicating a favorable effect of temperature on CPUE within this range (Figure 5). The effect peaked near 21 °C, beyond which CPUE declined sharply. Based on this pattern, the suitable thermal range for common squid in the coastal jigging fishery was estimated to be approximately 13–23 °C, with an optimal temperature of 21 °C. The 95% confidence interval for this optimum temperature was estimated to range from 18.1 °C to 22.0 °C.
Figure 5
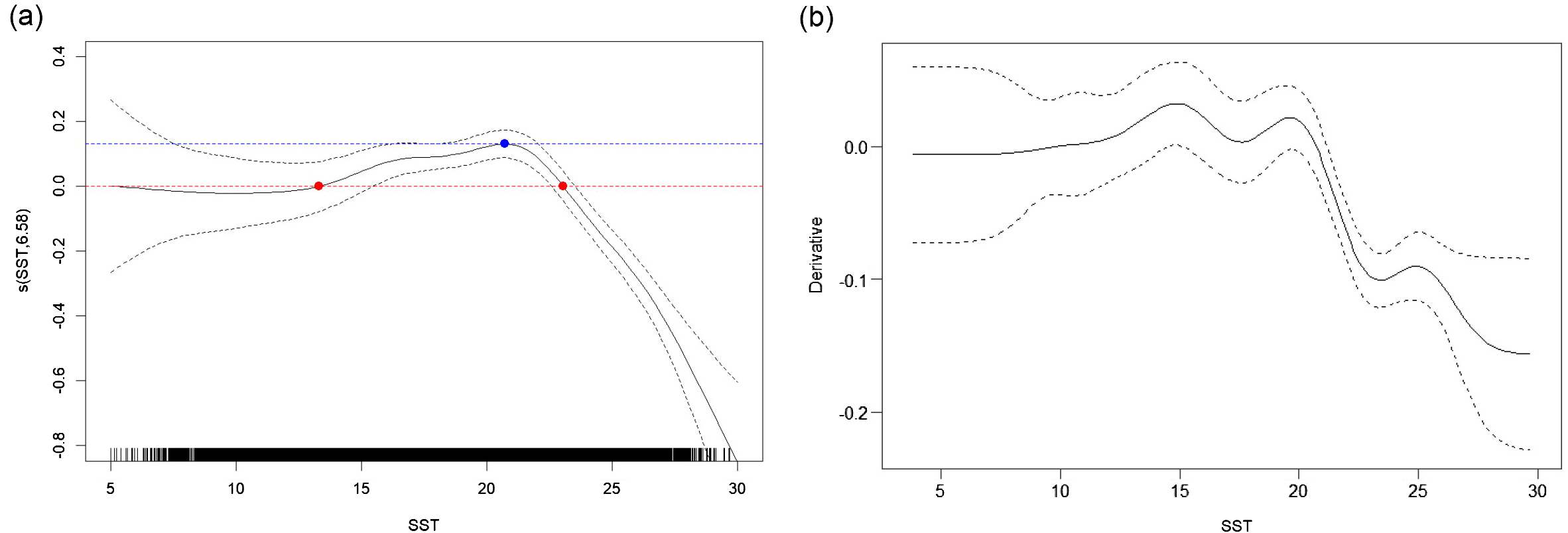
(a) Estimated smooth effect of sea surface temperature (SST) on the standardized catch per unit effort (CPUE) of the common squid using a generalized additive model (GAM). The solid black line represents the fitted smooth function with 95% confidence intervals (dashed lines). The red points indicate the SST values (13 °C and 23 °C) where the smooth effect crosses zero. The blue point indicates the SST (21 °C) at which the smooth effect reaches its maximum. (b) First derivative of the smooth SST effect, showing the rate of change in s(SST). The solid line represents the estimated derivative, and the dashed lines show the 95% confidence interval.
The first derivative of the smooth term s(SST) revealed the rate of change in the temperature effect on standardized CPUE (Figure 5b). The derivative was significantly positive between approximately 13–20 °C, indicating a strong increasing response of CPUE with rising SST within this range. Around 20–23 °C, the rate of increase slowed, reaching a plateau near the optimal temperature. Beyond 23 °C, the derivative became significantly negative, suggesting a rapid decline in CPUE with further warming. These results quantitatively support the presence of a thermal preference window for T. pacificus, bounded by a lower threshold (13 °C) and an upper limit (23 °C), with steep ecological transitions beyond this range.
To further evaluate the influence of SST, temporal trends in nominal CPUE (black dots) and standardized CPUE (red line) were compared (Figure 6). The standardized CPUE exhibited a gradual decline after the early 2000s, with reduced interannual variability compared to nominal CPUE. Notably, while nominal CPUE fluctuated irregularly between 2008 and 2016, the standardized CPUE showed a more consistent downward trend. This pattern suggests that SST may have played a significant role in explaining the observed variations.
Figure 6
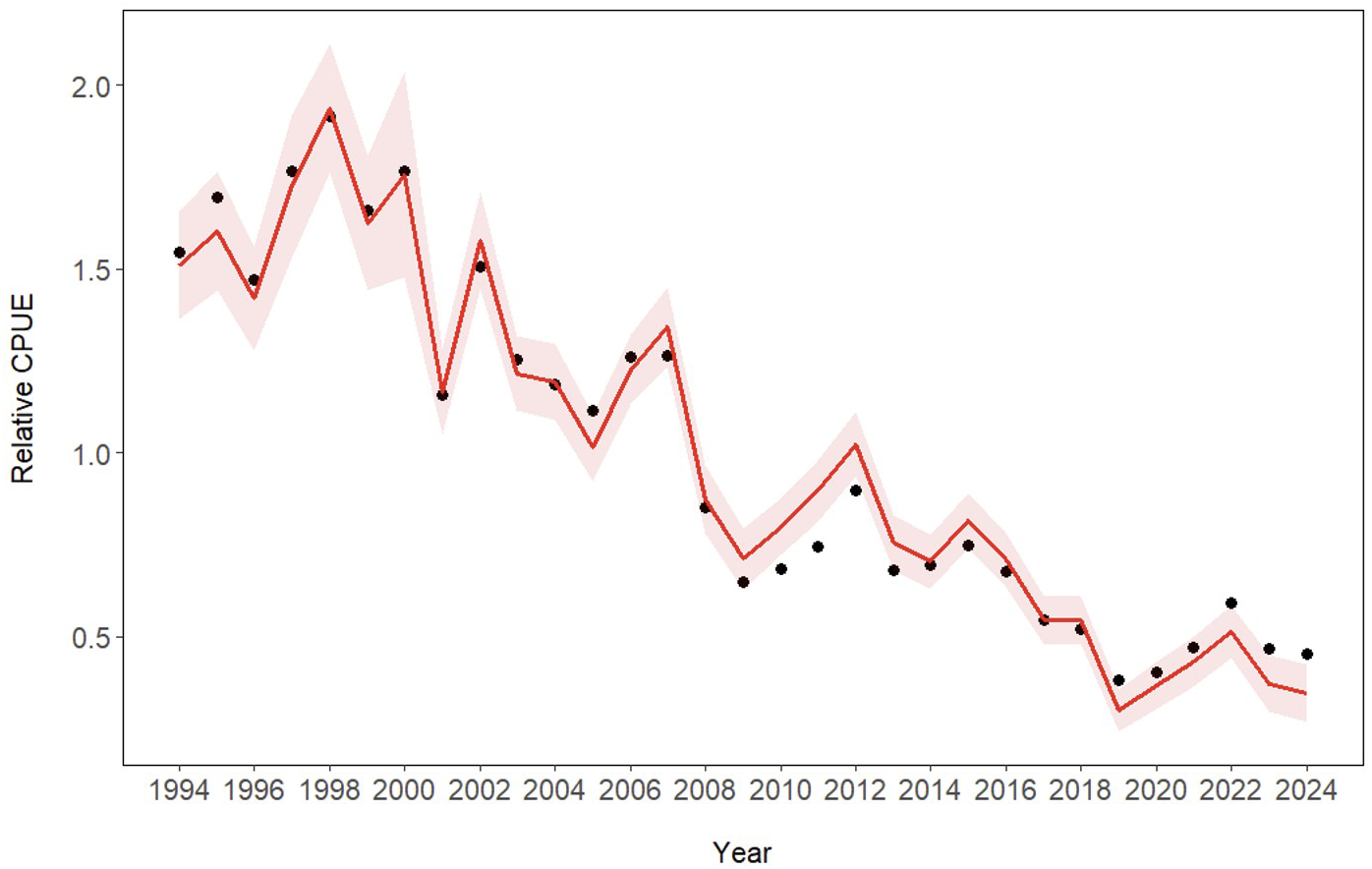
Annual trends in nominal and standardized CPUE for of the common squid in the Korean coastal jigging fishery from 1994 to 2024. Black dots represent nominal CPUE, while the red line and shaded area indicate the standardized CPUE and its confidence interval estimated using a GAM.
3.3 Projected suitable fishing grounds under climate change scenario
The projected spatial distributions of SST for the years 2024, 2050, and 2100 under the SSP3-7.0 scenario revealed shifts in the thermally suitable habitat range (13–23 °C) for common squid, particularly during the main fishing season from May to December (Figure 7). This period corresponds to ecologically important phases of the species’ northward migration and southward return, allowing for a quantitative assessment of seasonal and interannual changes in habitat suitability driven by SST variation.
Figure 7
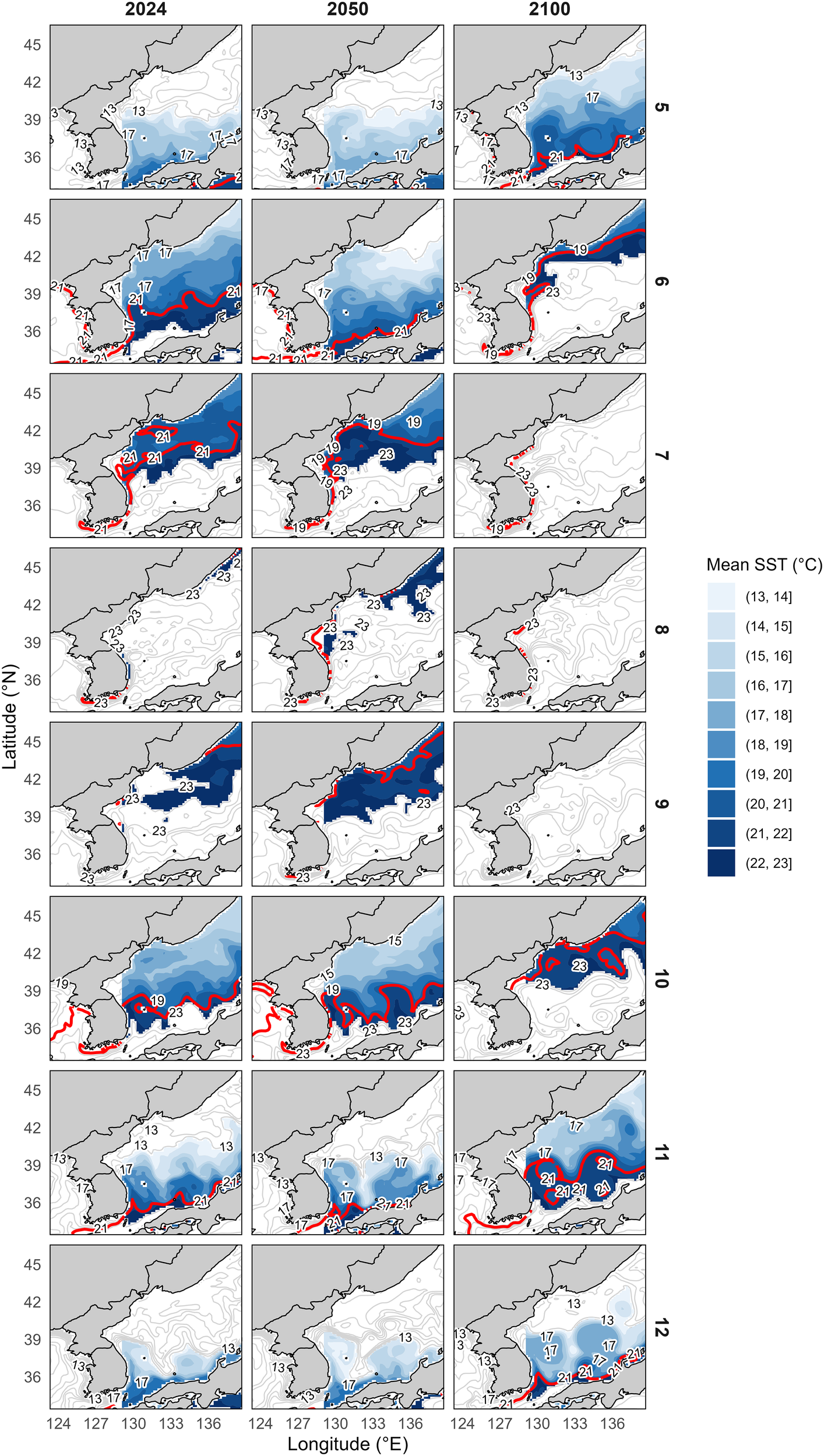
Projected monthly mean SST distributions from May to December in 2024, 2050, and 2100 under the SSP3-7.0 scenario. The suitable thermal range of the common squid Todarodes pacificus, 13–23 °C, is highlighted to identify potential fishing grounds. Black contour lines indicate SST intervals, and colored shading represents the SST range. The red contour line indicates the optimal temperature of 21 °C. Spatial patterns indicate a northward shift of thermally suitable habitats during May and June, and a notable reduction in suitable areas in the eastern part of the Korean EEZ during September and October—the peak fishing season—by 2100. Suitable conditions reappear in the south from November onward.
The spatiotemporal analysis indicated a gradual northward shift in the thermally suitable habitat range (13–23 °C) of common squid over time. In particular, the 21 °C isotherm—historically observed along Korea’s southern coast in May and June—was projected to move progressively toward the northern part of the eastern waters of Korea and Russian waters. Conversely, during September and October, traditional fishing grounds along the southern part of the eastern waters of Korea were projected to experience elevated temperatures exceeding 23 °C. Under the 2100 scenario, thermally suitable habitats in southern waters disappeared entirely during this period and reappeared in November and December.
In addition, the portion of the fishable area within the eastern part of the Korean EEZ (Figure 2) that falls within the suitable thermal range for common squid was quantitatively estimated (Table 4). According to the monthly projections under the SSP3-7.0 scenario, the thermally suitable area increased slightly from 2024 to 2050, but contracted substantially by 2100 (Figure 8). During the peak fishing season, the suitable area showed notable seasonal and temporal variation. In September, suitable habitat was already limited in 2024 and virtually disappeared by 2100. In contrast, October initially had a large extent of suitable area, but this declined sharply by 2100. Meanwhile, November and December exhibited relatively stable thermal conditions, with most of the fishable area projected to remain within the suitable range by 2100.
Table 4
| Month | 2024 | 2050 | 2100 |
|---|---|---|---|
| 9 | 8,906 (5.51) | 7,834 (4.84) | 5 (0.00) |
| 10 | 137,341 (84.92) | 149,827 (92.64) | 836 (0.52) |
| 11 | 161,557 (99.89) | 160,378 (99.16) | 156,022 (96.47) |
| 12 | 106,216 (65.67) | 123,109 (76.12) | 161,738 (100.00) |
| Total | 414,020 | 441,148 | 318,601 |
Projected area (km2) of suitable fishing grounds for common squid within the eastern part of the Korean exclusive economic zone (EEZ), based on the optimal thermal habitat range (13–23 °C) under the SSP3-7.0 scenario.
The numbers in parentheses represent the proportion (%) of each monthly value relative to the total fishing ground area (161,738 km2) in the eastern part of the Korean EEZ.
Figure 8
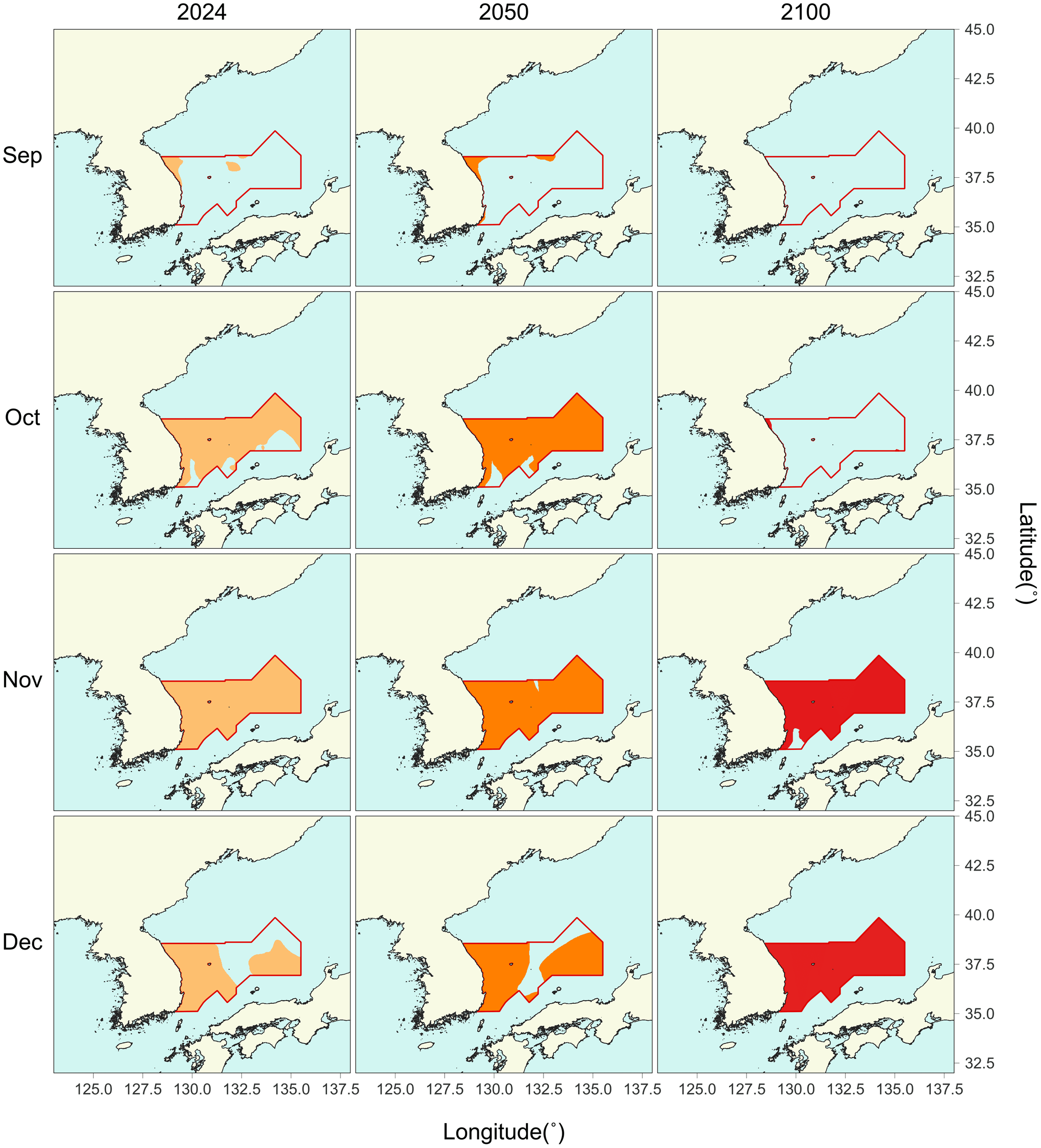
Monthly distribution of thermally suitable range (13–23 °C) for common squid Todarodes pacificus in September to December under three time slices (2024, 2050, and 2100) based on the SSP3-7.0 scenario, and the fishable area in the eastern Korean EEZ (red boundary).
4 Discussion
This study quantified future shifts in the fishing ground distribution of common squid in the eastern waters of Korea under the SSP3-7.0 climate change scenario, by standardizing CPUE data from the coastal jigging fishery and incorporating spatiotemporal and oceanographic variables. The GAM revealed a significant nonlinear relationship between CPUE and SST, with positive effects observed within the 13–23 °C range. The highest CPUE was detected near 21 °C, which was identified as the optimal temperature, while 13–23 °C was defined as the species’ suitable thermal range. Compared to the broader thermal tolerance range of 5–27 °C reported by (Roper et al., 1984), the refined range identified in this study reflects temperature conditions more closely associated with active fishing grounds. These results provide ecologically grounded temperature thresholds that can serve as a foundation for forecasting future habitat suitability under climate change.
Under the SSP3-7.0 scenario, projected SST distributions for 2050 and 2100 indicated a progressive northward shift and reduction in the spatial extent of areas meeting the suitable thermal range for common squid during its peak fishing season (September–October). Notably, by September–October 2100, suitable thermal habitats may disappear from the southern part of the eastern waters of Korea, suggesting potential constraints on the availability of traditional fishing grounds. In contrast, suitable conditions are projected to reemerge in the southern part of the eastern waters of Korea in November and remain available through December, implying a possible delay in the timing of fishing ground formation (Figure 8). These results suggest that climate change may not only alter the spatial distribution of squid fishing grounds but also shift the temporal window of fishing activities. As the fishable area is expected to contract or shift seasonally in response to ocean warming, adjustments to current fishing practices may be required. This highlights the need for adaptive strategies, including revisions to fishing seasons, adjustments in management benchmarks, and the development of predictive, climate-informed fisheries management.
While the thermal range identified in this study reflects the biological temperature preference of common squid, it is important to note that the CPUE data used were limited to areas where commercial fishing operations were conducted. As a result, the estimated thermal preference range may be influenced not only by the actual ecological distribution of the species, but also by fishing behavior and gear-specific selectivity. Such biases are inherent to fishery-dependent data, where observed catch rates can reflect where fishers choose to operate rather than where the species is most abundant. Although CPUE standardization reduces biases related to fishing time and location, it does not account for squid populations in unfished areas, which may limit the accuracy of stock status. To address this limitation, previous studies have suggested the integration of fishery-independent data sources, such as scientific surveys, as a complementary approach (Hoyle et al., 2024). Future studies should adopt an integrated approach that combines standardized CPUE data with biologically derived temperature preferences from the literature and fishery-independent data—such as acoustic monitoring, larval sampling, and seasonal trawl surveys—to improve the precision and ecological validity of habitat models under climate change.
Anthropogenic factors—including advances in fishing technology, changes in fisheries regulations, and illegal, unreported, and unregulated (IUU) fishing activities—can influence the spatiotemporal interpretation of CPUE (Cosgrove et al., 2014; Chiarini et al., 2022; Lee et al., 2017). These factors may alter fishing intensity and the spatial concentration of effort, thereby affecting not only the actual distribution of the resource but also the outcomes of CPUE standardization. If such influences are not adequately accounted for, the relationship between CPUE and SST may be over- or underestimated, leading to biased inferences about the environmental drivers of species distribution. While this study focused on spatiotemporal and SST factors, future CPUE standardization efforts should adopt a more integrated approach that also considers anthropogenic drivers. Doing so would improve the accuracy of identifying key environmental determinants of species distribution and contribute to the development of effective, climate-resilient fisheries management strategies.
Common squid are generally distributed at depths of 0–200 meters, with particularly high densities observed in waters shallower than 50 meters (Roper et al., 1984; Jo et al., 2019). In this study, only SST was used due to the limited availability of other oceanographic variables—such as salinity, chlorophyll, and primary productivity—at sufficient temporal (monthly) and spatial resolution over the full study period. While this may limit the vertical resolution of the modeled habitat, the use of SST is justified given the species’ strong presence in the upper water column and its behavioral aggregation near thermal fronts. Previous studies have shown that SST alone can effectively capture large-scale environmental influences on squid distribution and spawning ground shifts (Kuroda et al., 2020; Lai and Zhou, 2025). Therefore, SST at 5 m depth was used as a practical and interpretable proxy in the context of climate change forecasting.
Despite this limitation, incorporating more ecologically meaningful variables remains essential. Future research should include vertical oceanographic features, such as depth-specific temperature profiles and thermocline structure, which can be derived from ARGO float data or ocean reanalysis products. The larvae of common squid disperse into the eastern waters of Korea via the Tsushima Warm Current through the Korea Strait (Goto, 2002; Yamamoto et al., 2002), and the pathways and strength of this current system play a crucial role in determining spawning ground conditions and fishing ground formation (Cho et al., 2004; Choi et al., 2008). In addition to these physical factors—including current dynamics—the distribution and abundance of common squid are also influenced by biological and chemical drivers such as zooplankton availability and salinity (Hwang et al., 2012; Kang et al., 2002; Furukawa and Sakurai, 2008). Zooplankton serve as a key food source during the early life stages (Uchikawa and Kidokoro, 2014), and their distribution has been closely linked to fluctuations in squid abundance (Hwang et al., 2012). Although autumn-spawning cohorts are generally less exposed to low-salinity waters (Furukawa and Sakurai, 2008), potential changes in salinity under climate change must not be overlooked. Therefore, future modeling efforts should integrate various oceanographic variables—such as currents, salinity, and zooplankton—into nonlinear statistical frameworks to improve the accuracy of CPUE standardization and habitat prediction, thereby supporting habitat-based and adaptive fisheries management.
Declines in common squid stocks, coupled with climate-driven changes in oceanographic conditions, have led to a geographic expansion and shift of fishing grounds. During the 1980s and 1990s, advances in vessel performance and the adoption of automated jigging technologies enabled offshore jigging vessels to operate over increasingly distant areas (Oh et al., 2018). In particular, as catches in the eastern waters of Korea have sharply declined, the Yellow Sea has come to appear relatively more productive, even though actual catch levels in the region have not significantly increased (Statistics Korea, 2025). The Yellow Sea is known to harbor a mixture of autumn- and winter-spawning cohorts, highlighting the need to reexamine spatial distribution characteristics of squid populations and emphasizing the growing significance of this region in future stock assessments and predictive modeling.
Previous studies have often assumed, either implicitly or explicitly, that fishers are capable of autonomously adjusting their fishing grounds in response to climate-induced shifts in species distributions (Cheung et al., 2010; Erauskin-Extramiana et al., 2019; Tai et al., 2019). This assumption is supported by empirical evidence from Korea, where the primary fishing grounds of common squid in offshore jigging have shown a consistent northward shift over recent decades (Hwang et al., 2012). A recent study (Jo et al., 2025) analyzed the relationship between SST changes and fluctuations in squid abundance from the 1990s to 2023. The findings demonstrated that the warming-induced contraction of thermally suitable habitats, combined with increased fishing pressure, delayed the seasonal formation of fishing grounds and contributed to a long-term decline in standardized CPUE with a consistent trend shown in Figure 6. In addition to environmental stressors, anthropogenic pressures have likely further exacerbated the decline in squid abundance. These include intensified fishing effort from the adoption of advanced jigging technologies, cooperative fishing with trawlers, and illegal, unreported, and unregulated (IUU) fishing by Chinese vessels (Lee et al., 2017; Oh et al., 2018). Our projections under the SSP3-7.0 scenario similarly suggest that continued ocean warming may amplify these processes, resulting in reduced catch efficiency and greater variability and uncertainty in future catch levels.
Behavioral adaptation may represent a proactive response to changing ocean conditions, but it also imposes various socioeconomic burdens, including increased fuel costs, safety risks, and reduced operational efficiency. In particular, the northward shift of fishing grounds can lead to longer travel distances and higher fuel expenses for fishers, directly affecting the economic viability and sustainability of fishing operations. Rising fuel prices have been shown to significantly reduce fishing fleet profitability (Cheilari et al., 2013) and to induce spatial shifts in fishing activity, with fleets concentrating closer to shore under high fuel cost conditions (Yoon et al., 2024).
Moreover, the spatiotemporal shifts in suitable habitats may require fishing fleets to adjust the timing and location of their operations, which could lead to instability in market supply chains and raise concerns over food security. Therefore, effective management of expanding and shifting fishing grounds will require not only scientific analysis of spatial distributional changes but also the development of anticipatory and adaptive resource management policies such as flexible, season- or region-specific fishing permits and dynamic total allowable catch (TAC) allocations that are responsive to climate-driven habitat shifts.
The seasonal projections of thermally suitable habitats presented in this study can be used to develop risk maps for fisheries managers. These maps may help identify high-risk months or zones such as areas where suitable temperature conditions are expected to contract or disappear entirely (e.g., September–October by 2100). Such visual tools can support early warning systems and spatial planning. Our findings also have implications for designing adaptive TAC frameworks. For example, TACs could be seasonally or regionally reallocated based on projections of suitable habitat availability. Given that thermally suitable areas are projected to expand in November–December but decline in September–October, managers might consider shifting fishing quotas accordingly to enhance sustainability. The GAM-derived optimal SST and thermal range can be integrated into operational decision-support tools that utilize near-real-time SST data to predict the daily or weekly likelihood of suitable fishing conditions. Such tools could assist fishers in adjusting fishing locations and timing in response to short-term climate variability. These tools and frameworks, when coupled with predictive oceanographic models, may serve as a foundation for the implementation of climate-resilient, habitat-based management systems in Korea’s squid fisheries.
This study classified fishing grounds into two areas (Area 1 and Area 2) using spatial clustering based on fishing locations, and a statistically significant difference in CPUE was found between them. Area 2 includes part of the Russian EEZ, suggesting that common squid fishing grounds are spatially differentiated. The Russian EEZ has served as a strategic fishing area for Korean offshore jigging since the Korea–Russia fisheries agreement, with fishing activity in this region intensifying between July and October. This pattern likely reflects the combined effects of seasonal squid migration and institutional factors. Therefore, a comprehensive evaluation of fishing sustainability is needed, considering access fees, stock distribution, and catch performance. Adaptive strategies such as region-specific fishing plans and flexible timing of access should also be considered in response to seasonal shifts in fishing grounds. In particular, implementing dynamic seasonal closures based on habitat shifts and negotiating transboundary fishing agreements with neighboring countries may provide more effective and flexible management under changing environmental conditions. These policy instruments can help ensure continued access to productive grounds while supporting long-term resource sustainability amid climate-driven distribution changes. The findings of this study provide a valuable basis for the strategic use of foreign waters and the development of spatially explicit resource management. A more detailed assessment of fishing efficiency and long-term sustainability in the Russian EEZ is warranted.
Common squid stock has experienced a marked decline in recent years, prompting the Korean government to implement various management measures in response to the need for sustainable resource use. Currently, a seasonal fishing closure is enforced during April and May (limited to April for offshore jigging, coastal multi-fishery, and set nets), and a minimum legal mantle length of 15 cm has been established. In addition, since 2007, the TAC system has been introduced for four major fisheries—offshore jigging, Eastern Sea trawl, large otter trawl, and large powered purse seines—to regulate harvest levels (Korea Law Information Center, 2025). The species was also designated as a target of the national resource restoration program in 2018. However, these conventional management strategies have limited capacity to account for the impacts of climate-induced changes in oceanographic conditions, shifting fishing grounds, and altered species distributions. The thermally suitable range and climate-based habitat projections presented in this study can serve as foundational information to support more adaptive and forward-looking management. Future efforts should focus on establishing climate-resilient strategies that incorporate predictive tools, flexible fishing schedules, and revised resource management criteria. In particular, spatially dynamic seasonal closures, regionally reallocated total allowable catches (TACs), and habitat-based access regulations should be considered to more directly address catch declines and shifting squid distributions under climate change. Additionally, given the transboundary nature of squid distribution, future cooperative management frameworks—particularly with neighboring countries such as Russia and others—may be essential to ensure sustainable harvests under shifting oceanographic conditions.
5 Conclusion
This study standardized the catch per unit effort (CPUE) of common squid Todarodes pacificus in Korean waters using a generalized additive model (GAM) that incorporated spatiotemporal and sea surface temperature variables, and assessed future changes in thermally suitable fishing habitats under the shared socioeconomic pathway 3-7.0 (SSP3-7.0) climate change scenario. The model revealed a significant nonlinear relationship between CPUE and sea surface temperature (SST), with a suitable thermal range of 13–23 °C and an optimal temperature near 21 °C. Rather than forecasting future CPUE values, we applied these thermal thresholds to projected SST, which indicated a northward shift and seasonal delay in thermally suitable fishing grounds, especially under the 2100 scenario. These findings suggest that climate change is likely to affect not only the spatial distribution of squid fishing grounds, but also the timing of peak fishing seasons. As traditional fishing grounds in the eastern waters of Korea may become unsuitable during the autumn months, suitable habitats are projected to reemerge later in the season, indicating the need for flexible and adaptive management strategies. Although the analysis was limited to SST as the sole environmental factor and relied on fishery-dependent data, it provides a useful basis for identifying future thermally suitable habitats. Future studies that incorporate broader environmental variables and fishery-independent data could improve model accuracy and ecological relevance. These results may inform the development of operational habitat forecasting systems designed to enhance flexibility in future fishery management.
Statements
Data availability statement
The data analyzed in this study is subject to the following licenses/restrictions: The raw data supporting the conclusions of this article will be made available by the authors on request. Requests to access these datasets should be directed to kanghj87@outlook.kr.
Author contributions
M-JK: Methodology, Visualization, Writing – original draft, Writing – review & editing. CK: Writing – review & editing. HWK: Writing – review & editing. H-SJ: Writing – review & editing. HK: Conceptualization, Methodology, Visualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This research was funded by NIFS (National Institute of Fisheries Science) grant number R2025006.
Acknowledgments
We sincerely thank the reviewers for their insightful comments and constructive feedback, which greatly helped improve the quality of this article.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that Generative AI was used in the creation of this manuscript. English proofreading.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2025.1610859/full#supplementary-material
References
1
An D. Kim K. Kang H. Hyun S.-Y. (2021). A bayesian state-space production assessment model for common squid todarodes pacificus stock caught by multiple fisheries in korean waters. Korean J. Fisheries Aquat. Sci.54, 769–781. doi: 10.5657/KFAS.2021.0769
2
Ben Lamine E. Schickele A. Goberville E. Beaugrand G. Allemand D. Raybaud V. (2022). Expected contraction in the distribution ranges of demersal fish of high economic value in the mediterranean and european seas. Sci. Rep.12, 10150. doi: 10.1038/s41598-022-14151-8
3
Bishop J. Venables W. N. Wang Y.-G. (2004). Analysing commercial catch and effort data from a penaeid trawl fishery: A comparison of linear models, mixed models, and generalised estimating equations approaches. Fisheries Res.70, 179–193. doi: 10.1016/j.fishres.2004.08.003
4
Brander K. M. (2007). Global fish production and climate change. Proc. Natl. Acad. Sci.Hoboken, New Jersey, USA104, 19709–19714. doi: 10.1073/pnas.0702059104
5
Brander K. Cochrane K. Barange M. Soto D. (2017). “Climate change implications for fisheries and aquaculture,” in Climate Change Impacts on Fisheries and Aquaculture: A Global Analysis, vol. 1 . Eds. BarangeM.BahriT.BeveridgeM. C. M.CochraneK. L.Funge-SmithS.PoulainF. (Hoboken, New Jersey, USA: Wiley Online Library), 45–62.
6
Bueno-Pardo J. Nobre D. Monteiro J. N. Sousa P. M. Costa E. F. S. Baptista V. et al . (2021). Climate change vulnerability assessment of the main marine commercial fish and invertebrates of Portugal. Sci. Rep.11, 2958. doi: 10.1038/s41598-021-82595-5
7
Cheilari A. Guillen J. Damalas D. Barbas T. (2013). Effects of the fuel price crisis on the energy efficiency and the economic performance of the european union fishing fleets. Mar. Policy40, 18–24. doi: 10.1016/j.marpol.2012.12.006
8
Cheung W. W. L. Lam V. W. Y. Sarmiento J. L. Kearney K. Watson R. Zeller D. et al . (2009). Large-scale redistribution of maximum fisheries catch potential in the global ocean under climate change. Global Change Biol.16, 24–35. doi: 10.1111/j.1365-2486.2009.01995.x
9
Cheung W. W. L. Lam V. W. Y. Sarmiento J. L. Kearney K. Watson R. Zeller D. et al . (2010). Large-scale redistribution of maximum fisheries catch potential in the global ocean under climate change. Global Change Biol.16, 24–35. doi: 10.1111/j.1365-2486.2009.01995.x
10
Chiarini M. Guicciardi S. Angelini S. Tuck I. D. Grilli F. Penna P. et al . (2022). Accounting for environmental and fishery management factors when standardizing cpue data from a scientific survey: A case study for nephrops norvegicus in the pomo pits area (central adriatic sea). PLoS One17, e0270703. doi: 10.1371/journal.pone.0270703
11
Cho K.-D. Kim S.-W. Kang G.-H. Lee C.-I. Kim D.-S. Choi Y.-S. et al . (2004). Relationship between fishing condition of common squid and oceanic condition in the east sea. J. Korean Soc. Mar. Environ. Saf.10, 61–67.
12
Choi K. Lee C. I. Hwang K. Kim S.-W. Park J.-H. Gong Y. (2008). Distribution and migration of Japanese common squid, todarodes pacificus, in the southwestern part of the east (Japan) sea. Fisheries Res.91, 281–290. doi: 10.1016/j.fishres.2007.12.009
13
Conn P. B. Thorson J. T. Johnson D. S. (2017). Confronting preferential sampling when analysing population distributions: diagnosis and model-based triage. Methods Ecol. Evol.8, 1535–1546. doi: 10.1111/2041-210X.12803
14
Cosgrove R. Sheridan M. Minto C. Officer R. (2014). Application of finite mixture models to catch rate standardization better represents data distribution and fleet behavior. Fisheries Res.153, 83–88. doi: 10.1016/j.fishres.2014.01.005
15
Cruz L. Pennino M. Lopes P. (2024). Fisheries track the future redistribution of marine species. Nat. Climate Change14, 1093–1100. doi: 10.1038/s41558-024-02127-7
16
Ducharme-Barth N. D. Grüss A. Vincent M. T. Kiyofuji H. Aoki Y. Pilling G. et al . (2022). Impacts of fisheries-dependent spatial sampling patterns on catch-per-unit-effort standardization: A simulation study and fishery application. Fisheries Res.246, 106169. doi: 10.1016/j.fishres.2021.106169
17
Erauskin-Extramiana M. Arrizabalaga H. Hobday A. J. Cabré A. Ibaibarriaga L. Arregui I. et al . (2019). Large-scale distribution of tuna species in a warming ocean. Global Change Biol.25, 2043–2060. doi: 10.1111/gcb.14630
18
FAO (2021). The impact of covid-19 on fisheries and aquaculture food systems, possible responses: Information paper, november 2020. Rome, Italy: FAO. doi: 10.4060/cb2537en
19
Furukawa H. Sakurai Y. (2008). Effect of low salinity on the survival and development of Japanese common squid Todarodes pacificus hatchling. Fisheries Sci.74, 458. doi: 10.1111/j.1444-2906.2008.01546.x
20
Gim J. Hyun S.-Y. Yoon S. C. (2022). A state-space production assessment model with a joint prior based on population resilience: Illustration with the common squid todarodes pacificus stock. Korean J. Fisheries Aquat. Sci.55, 183–188. doi: 10.5657/KFAS.2022.0183
21
Goto T. (2002). Paralarval distribution of the ommastrephid squid todarodes pacificus during fall in the southern sea of Japan, and its implication for locating spawning grounds. Bull. Mar. Sci.71, 299–312.
22
Hinton M. G. Maunder M. N. (2004). Methods for standardizing cpue and how to select among them. Col. Vol. Sci. Pap. ICCAT56, 169–177.
23
Hoyle S. D. Campbell R. A. Ducharme-Barth N. D. Grüss A. Moore B. R. Thorson J. T. et al . (2024). Catch per unit effort modelling for stock assessment: A summary of good practices. Fisheries Res.269, 106860. doi: 10.1016/j.fishres.2023.106860
24
Hsu J. Chang Y.-J. Ducharme-Barth N. D. (2022). Evaluation of the influence of spatial treatments on catch-per-unit-effort standardization: A fishery application and simulation study of pacific saury in the northwestern pacific ocean. Fisheries Res.255, 106440. doi: 10.1016/j.fishres.2022.106440
25
Hwang K.-S. Kang S.-k. Oh T.-Y. Choi K. Lee D. W. (2012). Change in the fishing grounds and the relationship between the abundance of the common squid Todarodes pacificus and the distribution of zooplankton in the east sea. Korean J. Fisheries Aquat. Sci.45, 173–179. doi: 10.5657/KFAS.2012.0173
26
IPCC (2023). “Summary for policymakers,” in Climate change 2023: Synthesis report. contribution of working groups i, ii and iii to the sixth assessment report of the intergovernmental panel on climate change. Geneva, Switzerland: IPCC. doi: 10.59327/IPCC/AR6-9789291691647.001
27
Jo M. J. Kim M.-J. Kim H. W. Kang H. Kim C. S. (2025). Long-term changes in biological characteristics of Todarodes pacificus in response to environmental and stock variability in the korean jigging fishery. Korean J. Fisheries Aquat. Sci.
28
Jo M. J. Kim J. J. Yang J. H. Kim C. S. Kang S. K. (2019). Changes in the ecological characteristics of todarodes pacificus associated with long-term catch variations in jigging fishery. Korean J. Fisheries Aquat. Sci.52, 685–695. doi: 10.5657/KFAS.2019.0685
29
Kang Y. S. Kim J. Y. Kim H. G. Park J. H. (2002). Long-term changes in zooplankton and its relationship with squid, Todarodes pacificus, catch in Japan/east sea. Fisheries Oceanography11, 337–346. doi: 10.1046/j.1365-2419.2002.00211.x
30
Kim M.-J. Han I.-S. Lee J.-S. Kim D.-H. (2024b). A climate change risk assessment in aquaculture in korea. Climatic Change177, 93. doi: 10.1007/s10584-024-03758-7
31
Kim M.-J. Hong J.-B. Han I.-S. Lee J.-S. Kim D.-H. (2023). Vulnerability assessment of korean fisheries to climate change. Mar. Policy155, 105735. doi: 10.1016/j.marpol.2023.105735
32
Kim Y.-h. Jung H. K. Lee C. I. (2018). Changes in the spawning ground environment of the common squid, todarodes pacificus due to climate change. Ocean Polar Res.40, 127–143. doi: 10.4217/OPR.2018.40.3.127
33
Kim J. J. Kim C.-H. Lee J.-S. Kim S. (2014). Seasonal characteristics of Todarodes pacificus paralarval distribution in the northern east China sea. Korean J. Fisheries Aquat. Sci.47, 59–61. doi: 10.5657/KFAS.2014.0059
34
Kim S. R. Kim J. J. Stockhausen W. T. Kim C.-S. Kang S. Cha H. K. et al . (2019). Characteristics of the eggs and larval distribution and transport process in the early life stage of the chub mackerel scomber japonicus near korean waters. Korean J. Fisheries Aquat. Sci.52, 666–684. doi: 10.5657/KFAS.2019.0666
35
Kim J.-J. Lee H.-H. Kim S. Park C. (2011). Distribution of larvae of the common squid todarodes pacificus in the northern east China sea. Korean J. Fisheries Aquat. Sci.44, 267–275. doi: 10.5657/KFAS.2011.0267
36
Kim C. Lee J.-S. Yang J.-Y. Han I.-S. (2024a). Dynamic downscaling for regional ocean climate modeling around the korean peninsula and its application in fisheries. Korean J. Fisheries Aquat. Sci.57, 177–185. doi: 10.5657/KFAS.2024.0177
37
Korea Law Information Center (2025). Enforcement decree of the fishery resources management act. Available online at: https://www.law.go.kr/LSW/main.html (Accessed 2025-01-20).
38
Korea Meteorological Administration (2019). Climate Information Portal. Available online at: http://www.climage.go.kr (Accessed 2025-01-20).
39
Kuroda H. Saito T. Kaga T. Takasuka A. Kamimura Y. Furuichi S. et al . (2020). Unconventional sea surface temperature regime around Japan in the 2000s–2010s: Potential influences on major fisheries resources. Front. Mar. Sci. Volume7. doi: 10.3389/fmars.2020.574904
40
Lai Q. Zhou W. (2025). Multiscale variation analysis of sea surface temperature in the fishing grounds of pelagic fisheries. Front. Mar. Sci. Volume12. doi: 10.3389/fmars.2025.1567030
41
Lee K.-H. Choi J.-H. Shin J.-K. Chang D.-S. Park S.-W. (2009). Acoustical backscattering strength characteristics and density estimates of Japanese common squid distributed in yellow sea. Bull. Korean Soc. Fisheries Technol.45, 157–163. doi: 10.3796/ksft.2009.45.3.157
42
Lee J.-S. Ryu J.-G. Kee H.-K. (2017). A study on the status of chinese fishing in the east sea off North Korea and directions for countermeasures. J. Fisheries Business Administration48, 61–74. doi: 10.12939/FBA.2017.48.3.061
43
Lewis S. A. Stortini C. H. Boyce D. G. Stanley R. R. (2023). Climate change, species thermal emergence, and conservation design: a case study in the canadian northwest atlantic. FACETS8, 1–16. doi: 10.1139/facets-2022-0191
44
Lima A. R. Baltazar-Soares M. Garrido S. Riveiro I. Carrera P. Piecho-Santos A. M. et al . (2022). Forecasting shifts in habitat suitability across the distribution range of a temperate small pelagic fish under different scenarios of climate change. Sci. Total Environ.804, 150167. doi: 10.1016/j.scitotenv.2021.150167
45
Maunder M. N. Punt A. E. (2004). Standardizing catch and effort data: a review of recent approaches. Fisheries Res.70, 141–159. doi: 10.1016/j.fishres.2004.08.002
46
McFarlane G. A. Zhang C. I. King J. R. Kim S. Beamish R. J. Oh J. H. (2009). Contrast in life histories of exploited fishes and ecosystem structures in coastal waters off west Canada and east korea. Ocean Sci. J.44, 43–60. doi: 10.1007/s12601-009-0006-3
47
Murase H. Nagashima H. Yonezaki S. Matsukura R. Kitakado T. (2009). Application of a generalized additive model (gam) to reveal relationships between environmental factors and distributions of pelagic fish and krill: a case study in sendai bay, Japan. ICES J. Mar. Sci.66, 1417–1424. doi: 10.1093/icesjms/fsp105
48
National Institute of Fisheries Science (2021). Ecology and Fishing Ground of Fisheries Resources in Korean Waters (Korean: Ye-moon Publishing Co).
49
Oh T.-Y. Seo Y.-I. Cha H.-K. Jo H.-S. An Y.-S. Lee Y.-W. (2018). Change of fishing power index by technological development in the offshore squid jigging fishery. J. Korean Soc. Fisheries Ocean Technol.54, 224–230. doi: 10.3796/KSFOT.2018.54.3.224
50
Ono K. Punt A. E. Hilborn R. (2015). Think outside the grids: An objective approach to define spatial strata for catch and effort analysis. Fisheries Res.170, 89–101. doi: 10.1016/j.fishres.2015.05.021
51
Pecl G. T. Jackson G. D. (2008). The potential impacts of climate change on inshore squid: biology, ecology and fisheries. Rev. Fish Biol. Fisheries18, 373–385. doi: 10.1007/s11160-007-9077-3
52
Perry R. I. Cury P. Brander K. Jennings S. Möllmann C. Planque B. (2010). Sensitivity of marine systems to climate and fishing: Concepts, issues and management responses. J. Mar. Syst.79, 427–435. doi: 10.1016/j.jmarsys.2008.12.017
53
Perry A. L. Low P. J. Ellis J. R. Reynolds J. D. (2005). Climate change and distribution shifts in marine fishes. Science308, 1912–1915. doi: 10.1126/science.1111322
54
Poloczanska E. S. Brown C. J. Sydeman W. J. Kiessling W. Schoeman D. S. Moore P. J. et al . (2013). Global imprint of climate change on marine life. Nat. Climate Change3, 919–925. doi: 10.1038/nclimate1958
55
Pörtner H. O. Peck M. A. (2010). Climate change effects on fishes and fisheries: towards a cause-andeffect understanding. J. Fish Biol.77, 1745–1779. doi: 10.1111/j.1095-8649.2010.02783
56
Riley R. D. Snell K. I. Ensor J. Burke D. L. Harrell F. E. Jr. Moons K. G. et al . (2019). Minimum sample size for developing a multivariable prediction model: Part i–continuous outcomes. Stat Med.38, 1262–1275. doi: 10.1002/sim.7993
57
Roper C. F. E. Sweeny M. J. Nauen C. E. (1984). “Family ommastrephidae,” in FAO Species Catalogue. Vol. 3. Cephalopods of the World. An annotated and illustrated catalogue of species of interest to fisheries. Eds. RoperC. F. E.SweenyM. J.NauenC. E. (FAO Fisheries Synopsis, Rome, Italy), 156–186.
58
Rosa A. L. Yamamoto J. Sakurai Y. (2011). Effects of environmental variability on the spawning areas, catch, and recruitment of the Japanese common squid, Todarodes pacificus (cephalopoda: Ommastrephidae), from the 1970s to the 2000s. ICES J. Mar. Sci.68, 1114–1121. doi: 10.1093/icesjms/fsr037
59
Rousseeuw P. J. (1987). Silhouettes: A graphical aid to the interpretation and validation of cluster analysis. J. Comput. Appl. Mathematics20, 53–65. doi: 10.1016/0377-0427(87)90125-7
60
Sakurai Y. Kiyofuji H. Saitoh S. Goto T. Hiyama Y. (2000). Changes in inferred spawning areas of todarodes pacificus (cephalopoda: Ommastrephidae) due to changing environmental conditions. ICES J. Mar. Sci.57, 24–30. doi: 10.1006/jmsc.2000.0667
61
Sakurai Y. Kiyofuji H. Saitoh S.-I. Yamamoto J. Goto T. Mori K. et al . (2002). Stock fluctuations of the Japanese common squid, Todarodes pacificus, related to recent climate changes. Fisheries Sci.68, 226–229. doi: 10.2331/fishsci.68.sup1_226
62
Shi Y. Zhang X. Yang S. Dai Y. Cui X. Wu Y. et al . (2023). Construction of cpue standardization model and its simulation testing for chub mackerel (Scomber japonicus) in the northwest Pacific Ocean. Ecol. Indic.155, 111022. doi: 10.1016/j.ecolind.2023.111022
63
Song H. (2018). Fluctuations of common squid todarodes pacificus catches in the northwestern pacific under changing climate and habitat temperature. Korean J. Fisheries Aquat. Sci.51, 338–343. doi: 10.5657/KFAS.2018.0338
64
Statistics Korea (2025). Fishery Production Statistics. Available online at: https://www.kosis.kr/index/index.do (Accessed 2025-01-18).
65
Tai T. C. Steiner N. S. Hoover C. Cheung W. W. L. Sumaila U. R. (2019). Evaluating present and future potential of arctic fisheries in Canada. Mar. Policy108, 103637. doi: 10.1016/j.marpol.2019.103637
66
Tian Y. Kidokoro H. Watanabe T. Iguchi N. (2008). The late 1980s regime shift in the ecosystem of tsushima warm current in the Japan/east sea: Evidence from historical data and possible mechanisms. Prog. Oceanography77, 127–145. doi: 10.1016/j.pocean.2008.03.007
67
Uchikawa K. Kidokoro H. (2014). Feeding habits of juvenile Japanese common squid todarodes pacificus: Relationship between dietary shift and allometric growth. Fisheries Res.152, 29–36. doi: 10.1016/j.fishres.2013.07.001
68
Winter A. Coyle K. Swartzman G. (2007). Variations in age-0 pollock distribution among eastern bering sea nursery areas: A comparative study through acoustic indices. Deep Sea Res. Part II54, 2869–2884. doi: 10.1016/j.dsr2.2007.08.009
69
Xiong P. Cai Y. Jiang P. Xu Y. Sun M. Fan J. et al . (2024). Impact of climate change on the distribution of trachurus japonicus in the northern south China sea. Ecol. Indic.160, 111758. doi: 10.1016/j.ecolind.2024.111758
70
Yamamoto J. Masuda S. Miyashita K. Uji R. Sakurai Y. (2002). Investigation on the early stages of the ommastrephid squid Todarodes pacificus near the oki islands (sea of Japan). Bull. Mar. Sci.71, 987–992.
71
Yoon S. C. Lee J. H. Kang H. (2024). A study on distribution and fluctuation of the fishing grounds of chionoecetes japonicus by the offshore trap fishery in the east sea. J. Korean Soc. Fish Ocean Technol.60, 340–347. doi: 10.3796/KSFOT.2024.60.4.340
Summary
Keywords
common squid, fishing ground forecasting, CPUE standardization, sea surface temperature, generalized additive model
Citation
Kim M-J, Kim C, Kim HW, Ji H-S and Kang H (2025) Forecasting the spatial variation of optimal sea surface temperature for common squid (Todarodes pacificus) in the Korean jigging fishery. Front. Mar. Sci. 12:1610859. doi: 10.3389/fmars.2025.1610859
Received
13 April 2025
Accepted
13 August 2025
Published
01 September 2025
Volume
12 - 2025
Edited by
Pranaya Kumar Parida, Central Inland Fisheries Research Institute (ICAR), India
Reviewed by
Leif Rasmuson, Oregon Department of Fish and Wildlife, United States
Gyanaranjan Dash, Central Marine Fisheries Research Institute (ICAR), India
Aratrika Ray, National Taiwan Ocean University, Taiwan
Updates
Copyright
© 2025 Kim, Kim, Kim, Ji and Kang.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Heejoong Kang, kanghj87@korea.kr
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.