 João P. Sousa1*
João P. Sousa1* Luísa Marques1
Luísa Marques1 Daniela P. Rodrigues1
Daniela P. Rodrigues1 Felisa Rey2,3
Felisa Rey2,3 Manuel Albuquerque1
Manuel Albuquerque1 Maria Rosário M. Domingues2,3
Maria Rosário M. Domingues2,3 Rafael C. Duarte1
Rafael C. Duarte1 Ricardo Calado1*
Ricardo Calado1*- 1ECOMARE & CESAM, Centre for Environmental and Marine Studies and Department of Biology, University of Aveiro, Aveiro, Portugal
- 2CESAM, Centre for Environmental and Marine Studies and Department of Chemistry, University of Aveiro, Aveiro, Portugal
- 3Mass Spectrometry Centre and LAQV-REQUIMTE, Department of Chemistry, University of Aveiro, Aveiro, Portugal
The accelerated growth of marine aquaculture targeting species that require formulated aquafeeds creates a pressing need to find new sustainable ingredients and additives that can secure a premium farming performance, as well as the nutritional value of farmed species. Marine gammarid amphipods can be an excellent source of essential fatty acids (FA), namely, long-chain n-3 polyunsaturated FA (LC-PUFA). However, their potential contribution to aquaculture feeds remains largely unexplored. The present work evaluated the effect of five mono-specific macroalgal diets (Ulva sp., Fucus sp., Laminaria sp., Gracilaria sp., and Chondrus sp.) on the FA composition of the offspring of Gammarus locusta fed on these macroalgae and compared it with the offspring of wild conspecifics. Results revealed that offspring from wild G. locusta showed the highest amount of PUFA (4.95 ± 0.31 μg mg-1 DW), as well as essential FA, namely eicosapentaenoic acid (EPA, 20:5n-3) (2.49 ± 0.21 μg mg-1 DW) and docosahexaenoic acid (DHA, 22:6n-3) (0.87 ± 0.16 μg mg-1 DW). The treatment with the highest amount of PUFA and EPA was the one using Fucus sp. as a mono-specific macroalgal diet (4.46 ± 0.31 μg mg-1 DW and 2.42 ± 0.24 μg mg-1 DW, respectively). The best levels of DHA in offspring were recorded for specimens feeding on Chondrus sp. (0.56 ± 0.10 μg mg-1 DW). Significant differences in the abundance of stearic acid (18:0) and DHA were observed between the offspring of adults fed algal-based diets and those from the wild. These findings suggest that G. locusta selectively conserves and directly or indirectly biosynthesises essential FA for growth and reproduction, particularly LC-PUFA. Overall, G. locusta can perform a trophic upgrade of its diet, making this gammarid amphipod a promising additive for the formulation of high-quality aquafeeds supplying LC-PUFA.
1 Introduction
Over the past two decades, significant advancements in aquaculture nutrition have been driven by a deeper understanding of the nutrient requirements of farmed organisms, feed efficiency, and sustainable supply of aquafeeds (Naylor et al., 2021; Glencross et al., 2023; FAO, 2024). Due to a lack of sustainable sources, the pressure on high-quality aquafeed ingredients, particularly fish meal (FM) and fish oil (FO), grows proportionally to the demand for sustainable aquaculture products. These conventional aquafeed components contain essential amino acids and long-chain polyunsaturated fatty acids (LC-PUFA), which play key-roles in the development, reproduction, immune system and stress response of multiple marine species targeted for aquaculture (Ribes-Navarro et al., 2022; Glencross et al., 2024; FAO, 2024). Consequently, while FM and FO are some of the most balanced and nutritious ingredients for the formulation of aquafeeds, the increase in marine aquaculture is challenging the sustainability of their use, as these are derived from finite marine resources (Naylor et al., 2021; FAO, 2024). For this reason, the search for nutritionally rich and environmentally friendly alternatives to FM and FO is a priority in marine aquaculture systems (Sadegh Alavi-Yeganeh et al., 2008; Baeza-Rojano et al., 2013; Jiménez-Prada et al., 2020; Rodrigues et al., 2022).
Several alternatives have emerged to relieve pressure on traditional sources of marine ingredients, such as the introduction of plant and algal-based oils (e.g., soy meal, agricultural waste and microalgae) and animal-based and single-cell solutions (e.g., insect meal, yeast and bacteria-based feeds, respectively), which have been moderately successful in replacing FM and FO (Cottrell et al., 2020). It is worth highlighting, that replacing commonly used marine-based ingredients by land-based ones when formulating aquafeeds for marine organisms is often associated with a reduction in the nutritional value of the final product; this caveat is mostly due to a decrease, or even a lack, of essential amino acids and essential n-3 LC-PUFA, namely eicosapentaenoic acid (EPA, 20:5n-3) and docosahexaenoic acid (DHA, 22:6n-3) (Álvarez et al., 2020; Napier and Betancor, 2023).
Marine gammarid amphipods have been highlighted as promising candidates to diversify the supply of FM and FO for the formulation of aquafeeds due to their high protein (40-45% in dried Gammarus), essential amino acids and LC-PUFA content, as well as their natural high abundance and larger body size (> 1 mm) when compared to other aquatic micro-invertebrates (Baeza-Rojano et al., 2014; Harlıoğlu and Farhadi, 2018; Jiménez-Prada et al., 2018). Besides being highly abundant in aquatic environments, these organisms are able to grow under a large range of environmental conditions, making them suitable for large-scale production (Costa and Costa, 2000; Alberts-Hubatsch et al., 2019). Additionally, gammarids can be farmed using agricultural by-products and aquaculture waste streams as feed, closing the nutrient cycle and reducing waste, thus enhancing sustainability and fostering a circular bioeconomy (Alberts-Hubatsch et al., 2019; Jiménez-Prada et al., 2020; Ribes-Navarro et al., 2022). It has already been documented that extracts of amphipods from genus Gammarus can enhance the immune response, growth, and resistance to stress in fish production, making them potentially valuable additives to improve farmed fish performance (Harlıoğlu and Farhadi, 2018). Feed represents a significant expense in aquaculture, so low-cost and nutrient-rich alternatives, such as marine gammarid amphipods, can contribute to reducing the reliance on increasingly expensive FM and FO, while also improving aquaculture profitability (Naylor et al., 2021; FAO, 2024).
To unravel the full potential of marine gammarid amphipods as ingredients or additives for the formulation of aquafeeds, it is urgent to foster basic research studies to optimise their cultivation techniques and evaluate their potential applications (Harlıoğlu and Farhadi, 2018). Recent works on several temperate gammarid species, such as Gammarus locusta and Echinogammarus marinus, more recently renamed as Marinogammarus marinus (Horton, 2023), have provided valuable insights into their nutritional profile and growth performance under specific diets, such as agricultural sidestreams, which lack essential LC-PUFA, and aquaculture waste. Nonetheless, those gammarids present high levels of PUFA and LC-PUFA and show no radical changes in growth and survival (Alberts-Hubatsch et al., 2019; Jiménez-Prada et al., 2020; Ribes-Navarro et al., 2022).
The objective of the present study was to evaluate the impact of mono-specific macroalgal diets supplied to G. locusta broodstock, (Figure 1 – left) on the fatty acid (FA) composition of their offspring (Figure 1 – right) and compare it to the offspring obtained from wild conspecifics. The macroalgae Ulva sp., Fucus sp. and Gracilaria sp. were chosen because they are a natural source of food and shelter for this gammarid species (Costa and Costa, 2000), while Laminaria sp. and Chondrus sp. were also tested as diets due to their high abundance in the sampling area of the amphipod species being addressed in the present study. By feeding G. locusta broodstock with mono-specific macroalgal diets that display very low absolute levels of EPA and lack DHA, we aimed to: 1) test the ability of this marine gammarid amphipod species to perform a trophic upgrade by producing offspring featuring these biomolecules in their biomass; and 2) contrast the FA profile of farmed offspring with that of offspring spawned by conspecifics collected from the wild.

Figure 1. Left: A breeding pair of Gammarus locusta (male is the larger specimen on top). Right: Gammarus locusta newborn.
2 Materials and methods
2.1 Sampling and laboratory procedures
Adults of G. locusta were sampled from Mira channel in the coastal lagoon Ria de Aveiro (Portugal, 40°37’15.4”N, 8°44’23.6” W), at 2.5 km from the lagoon’s inlet and with a salinity of 35 (Quintino et al., 2009). Specimens of G. locusta were collected from the wild in June 2023 at low tide from the macroalgae Ulva sp., Fucus sp., and Gracilaria sp. by gently shaking algal fronds inside a 20-L bucket filled with seawater from that same location to release attached amphipods. Subsequently, all specimens were sieved through a 1-mm net and transferred into another 20-L bucket also containing water from the sampling location and transported to the laboratory. At the laboratory, females of G. locusta in pre-copula were sorted and paired with a conspecific male, until forming a total of 25 breeding pairs. All breeding pairs were fed ad libitum on freshly collected Ulva sp. while being individually acclimated for seven days in custom-built floating PVC chambers (63 mm in diameter x 100 mm high, with their bottom being covered with a 1-mm mesh) randomly distributed through four 50-L tanks (350 mm long x 400 mm wide x 500 mm high) with recirculating natural seawater at a temperature of 20°C and a photoperiod of 12 h light/12 h dark provided by a pair of 58 W white fluorescent lamps per each two tanks (see Figure 2). The life support system of these four 50-L tanks was equipped with a sump, housing a Deltec 1000ix internal protein skimmer, a TMC UV-steriliser P1 of 55 W, a Hailea HC-500A chiller, which was connected to an EHEIM universal 1200 water pump placed inside the sump to maintain the temperature stable (20 ± 1 °C), and an EHEIM universal 2400 pump returning the water to the tanks at a flow of 2400 L h-1. Salinity was kept stable at 35 through a Deltec Aquastat 1001 osmoregulator connected to an EHEIM 1000 pump placed inside a reservoir filled with freshwater purified by reverse osmosis to compensate for water evaporation (Silva et al., 2021).

Figure 2. (A) Lateral view of floating PVC chambers employed to house the breeding pairs of Gammarus locusta (left) and detachable chamber to collect their newly hatched offspring (right); (B) Lateral view of the assembled PVC chamber; (C) Bottom view of the PVC chamber with 1-mm net at the bottom; (D) Bottom view of the detachable chamber to collect newly hatched offspring with the 150-µm net at the bottom; and (E) Experimental life support system employed featuring the four 50-L tanks used to house the floating PVC chambers stocked with the breeding pairs of G. locusta.
Fresh biomass of the following five different species of macroalgae was sampled from three different sites within the Mira channel: the green macroalga Ulva sp. (popularly known as sea lettuce), the brown macroalgae Fucus sp. (popularly known as bladderwrack) and Laminaria sp. (popularly known as kelp), and the red macroalgae Gracilaria sp. (popularly known as ogonori) and Chondrus sp. (popularly known as Irish moss) (Figure 3).

Figure 3. Macroalgae sampled at Ria of Aveiro (Portugal) and used as monospecific diets to feed the breeding pairs of the marine amphipod Gammarus locusta: (A) Ulva sp. (green macroalga); (B) Fucus sp. (brown macroalga); (C) Laminaria sp. (brown macroalga); (D) Gracilaria sp. (red macroalga); and (E) Chondrus sp. (red macroalga).
After sampling, all macroalgae were transported to the laboratory in buckets filled with local seawater to be used as mono-specific diets for G. locusta. In the laboratory, all macroalgae were rinsed and hand-washed with seawater to remove any potential source of contamination during the experiment, such as small invertebrates and epiphytes. All collected macroalgae were kept in tanks under the same conditions as the amphipods (see above). Five pieces of at least 5 g each (fresh weight) were collected from each macroalgal biomass, rinsed with freshwater purified by reverse osmosis and the remaining epiphytes were manually removed, being subsequently frozen at -20°C and freeze-dried (Labogene CoolSafe 55-9L PRO) for further analyses, as recommended by Jiménez-Prada et al. (2020).
Each of the twenty-five breeding pairs of G. locusta was fed with one of the five mono-specific macroalgal diets after 24 h of starvation to purge their digestive tract (which could be confirmed visually through the absence of faecal material in their translucent dorsal intestine tube) and ensure that the breeding pair would derive any of the nutrients used to mature their gonads from the mono-specific macroalgal diets being tested. Additionally, 20 ovigerous females from the wild brooding embryos about to hatch were isolated and starved in the chambers detailed above until their newly hatched offspring was released from the female’s marsupium. Five (n = 5) of these newly hatched wild broods were collected and rinsed with water purified by reverse osmosis, gently dried with absorbent paper, counted, frozen at -20 °C, freeze-dried and stored at -20 °C until further analysis (Rollin et al., 2017).
2.2 Experimental setup
The twenty-five breeding pairs of G. locusta were separately allocated to a floating chamber, with these being randomly distributed throughout the four 50-L tanks detailed above and a total of five breeding pairs being allocated to each of the five mono-specific macroalgal diets. After seven days of acclimation (see section 2.1), each breeding pair was supplied daily with approximately 0.2 g (determined after preliminary feeding trials performed in the laboratory) of one of the five fresh mono-specific macroalgal diets tested. All breeding pairs of G. locusta were fed ad libitum and exclusively on their specific macroalgal diet, with any uneaten food being removed daily to safeguard water quality. Breeding pairs were fed with one of the five mono-specific macroalgal diets described above to obtain five broods of newly hatched offspring per mono-specific macroalgal diet (n = 5 per each mono-specific macroalgal diet), with each brood being considered as an independent replicate for experimental purposes. The newly hatched offspring of G. locusta breeding pairs was collected every day from the detachable chambers. The broods considered for biochemical analyses either originated from the second or third brood produced in the laboratory under the mono-specific macroalgal diet being tested, depending on whether the female was either non- or ovigerous since the start of the experiment, respectively. This procedure aimed to safeguard that the FA profiles displayed by their newly hatched offspring mostly reflected the influence of the mono-specific macroalgal diets being tested and were less likely to be biased by maternal reserves derived from their natural diet in the wild. The five broods of newly hatched offspring per mono-specific macroalgal diet selected for biochemical analysis were first rinsed with water purified by reverse osmosis, gently dried with absorbent paper, counted, frozen at -20 °C, freeze-dried and stored at -20 °C until further analyses (Rollin et al., 2017).
2.3 Biochemical analyses
2.3.1 Lipid extraction
2.3.1.1 Macroalgae
Freeze-dried samples of all macroalgae tested in the present work, with a mean dry weight (DW) of 252 ± 2.8 mg (mean ± standard deviation), were macerated in a mortar using a glass rod. Lipid extraction was performed using a modified protocol from the Bligh and Dyer (1959) method (Rey et al., 2023). A volume of 2500 μL of methanol (MeOH), (HPLC grade, Fisher Scientific, UK) and 1250 μL of dichloromethane (CH2Cl2) (HPLC grade, Fisher Scientific, UK) was added to each sample and then homogenised (Multi Reax Vortex Shaker, Heidolph Instruments, Germany) for 2 min, sonicated for 1 min and incubated on ice in an orbital shaker at 150 rpm (Stuart SSL2 Reciprocating Shaker, Cole Parmer, USA) for 2 h. After the addition of 1250 μL of CH2Cl2, samples were homogenised for 2 min and then centrifuged at 3000 rpm for 10 min at 4 °C (Centurion Scientific Pro Analytical C4000R with a BRK5324 rotor, Stoughton, UK). The organic phase was collected in a new tube, and the biomass residue was re-extracted by adding 2500 μL of MeOH and 2500 μL of CH2Cl2, followed by vortex and centrifugation. A volume of 2250 μL of Mili Q water (Synergysup®, Millipore Corporation, Billerica, MA, USA) was added to the organic phase, promoting phase separation. The organic phase was collected, following homogenisation and new centrifugation for 10 min at 3000 rpm at 4 °C. The aqueous phase was re-extracted with 2000 μL of CH2Cl2 and centrifuged. The combination of both organic phases was dried under a nitrogen stream, and total lipid content was determined by gravimetry.
2.3.1.2 Newly hatched offspring of Gammarus locusta
Freeze-dried samples of newly hatched offspring of G. locusta with a mean DW of 0.92 ± 0.1 mg (mean ± standard deviation) were transferred to glass tubes and macerated with a glass rod before adding 500 μL of MeOH and 250 μL of CH2Cl2. The resulting mixture was homogenised for 2 min and incubated on ice for 30 min. Posteriorly, 250 μL of dichloromethane were added and homogenised for 2 min prior to centrifugation at 3000 rpm for 10 min at 4 °C. The organic phase containing the lipid extract was transferred to a new tube and dried under a nitrogen stream before methylation (Rey et al., 2022).
2.3.2 Fatty acid analysis
Fatty acid methyl esters (FAME) were obtained from alkaline transmethylation. A lipid extract fraction corresponding to 30 μg of lipids from macroalgae or total lipid from G. locusta newly hatched offspring was transferred to glass tubes previously washed with n-hexane (99%) (Maia et al., 2025). After drying under a nitrogen stream, 1 mL of internal standard C19:0 (1.01 μg mL-1, CAS number 1731-94-8, Merck) in n-hexane (99%) was added to the tubes containing the lipid extracts, followed by the addition of 200 μL of a methanolic potassium hydroxide (KOH) solution (2 M). After sample homogenisation, 2 mL of a saturated sodium chloride solution (NaCl, 10 mg mL-1) was added, and the samples were centrifuged for 5 min at 2000 rpm. Then, 600 μL of organic phase containing the FAME were collected. In offspring samples, cholesterol from the organic phase was removed before gas chromatography mass spectrometry (GC-MS) analysis, according to the Christie (2023) protocol (Maia et al., 2025). Glass tips were filled with 0.1 g of silica and cotton at the bottom. The columns were activated with 5 mL of n-hexane and the 600 μL of the organic phase containing FAME were added to the column, followed by 3 mL of a hexane:diethyl ether solution (95:5, v/v). FAMEs without cholesterol were collected in a glass tube and dried under a nitrogen flow. Subsequently, FAMEs from both macroalgae and G. locusta offspring were dissolved in 100 μL of n-hexane (99%), from which 2 μL of the solution were injected in the GC-MS equipment (Agilent Technologies 5977 B GC/MSD, Santa Clara, CA, USA) with a DB-FFAP column (123-3232, J and W Scientific, Folsom CA, USA), presenting the following specifications: 30 m in length, an internal diameter of 320 μm and a film thickness of 0.25 μm. The equipment was connected to a Mass Selective Detector operating with an electron impact mode at 70 eV and a scanning mass range of m/z 50-550 (1-second cycle in full scan mode). The oven temperature was programmed from an initial temperature of 58 °C for 2 min, followed by a linear increase to 160 °C at 25 °C min-1, a linear increase of 2 °C min-1 to 210 °C, then 20 °C min-1 until reaching 225 °C which was maintained for 15 min. The injector was set at 220 °C and the detector at 230 °C. Helium was used as the carrier gas (constant flow 1.4 mL min-1). The data acquisition software employed was the GCMS 5977B/Enhanced MassHunter, with FA peaks being identified using Agilent MassHunter Qualitative Analysis 10.0 software through retention time values and comparison of MS spectra of each FAME with the 37-Component FAME Mix certified standard (Supelco 37 Component FAME Mix, ref. 47,885–1, Sigma-Aldrich) and confirmed by comparison with NIST chemical database library and the spectral library “The AOCS Lipid Library” (Christie, 2023).
2.4 Statistical analyses
A Principal Coordinate Analysis (PCoA) was performed separately for the macroalgae and the newly hatched offspring of G. locusta datasets, using the abundance of the most abundant FAs (expressed as µg of FA per mg of macroalgae or amphipod dry weight, DW), specifically those representing more than 1% of total FA abundance, while excluding branched and epoxy FAs. Prior to analysis, data were log (x+1)-transformed to reduce the impact of dominant FAs and prevent overemphasis on their contribution. The transformed data were then converted into a resemblance matrix using the Euclidean distance similarity coefficient. Using this resemblance matrix, we performed a one-way Permutational Multivariate Analysis of Variance (PERMANOVA; Anderson, 2001) with 9,999 permutations to compare FA composition across groups. We conducted separate comparisons for macroalgae (with five algal species as a fixed factor) and newly hatched offspring of G. locusta (from breeding pairs fed with the five mono-specific macroalgal diets and from ovigerous females collected from the wild as a fixed factor). Whenever significant effects were detected, pairwise comparisons were performed to identify differences between levels of significant factors. Additionally, the Similarity Percentage (SIMPER) method was also employed to determine which FAs contributed most to the observed differences between macroalgal species and broods of newly hatched offspring of G. locusta. All multivariate analyses were conducted using R v.4.4.3 (R Core Team, 2025). Specifically, PCoA was performed through the function ‘pcoa’ of the “ape” package (Paradis and Schliep, 2019), PERMANOVA with pairwise comparisons using the functions ‘adonis2’ and ‘pairwise.adonis2’ of the packages “vegan” and “pairwiseAdonis”, respectively, and SIMPER using the function ‘simper’ of the “vegan” package (Arbizu, 2017; Oksanen et al., 2025).
Furthermore, a separate one-way Analysis of Variance (ANOVA) was employed to assess significant differences in the abundance of FA classes, namely saturated fatty acids (SFA), monounsaturated fatty acids (MUFA), and polyunsaturated fatty acids (PUFA), as well as FA ratios (n-3/n-6 and PUFA/SFA) among the five macroalgal species. Similarly, we used separate one-way ANOVAs to compare the individual abundances of the most abundant FAs, FA classes, and FA ratios (including the EPA/DHA ratio) in the newly hatched offspring of G. locusta, as well as the number of neonates produced per brood and their DW, in relation to parental diet (wild or fed with the five mono-specific macroalgal diets). For all models, we assessed the normality of residuals using q-q plots and tested the homogeneity of variances with Bartlett’s test, using the “performance” package in R (Lüdecke et al., 2021). Log transformation was applied where necessary, including for SFA, MUFA, and PUFA/SFA in macroalgae, as well as for DW, certain individual FAs (16:0, 18:0, and 20:4 n-6), and SFA in newly hatched offspring of G. locusta. In cases of significant effects, we performed Tukey’s post-hoc tests to identify differences between factor levels using the “emmeans” package in R (Lenth, 2017). All univariate statistical analyses were conducted using R v.4.4.3 (R Core Team, 2025).
3 Results
3.1 Gammarus locusta offspring
The average number of newly hatched amphipods released per brood and the DW of the whole brood differed significantly between treatments (F5,24 = 14.89, p < 0.001 and F5,24 = 3.91, p = 0.01) (Supplementary Figure 1). In the wild, adult G. locusta (i.e., not exposed to the experimental diets) produced a significantly higher number of offspring per brood than conspecifics fed on mono-specific macroalgal diets, except for those supplied with Ulva, which featured a similar production of offspring per brood (mean ± SE: 87 ± 4 and 81 ± 3 for offspring per brood from wild and Ulva, respectively). In contrast, conspecifics fed with Chondrus produced fewer neonates per brood than those in the other mono-specific macroalgal diets, except for those fed with Gracilaria, which exhibited a comparable number (46 ± 2 offspring per brood from Chondrus and 59 ± 3 offspring per brood from Gracilaria). The DW of the whole brood did not vary considerably among treatments, except for amphipods fed with Chondrus (1.13 ± 0.04 mg), which displayed significantly lower values than those fed with Laminaria (2.01 ± 0.32 mg) and those from the wild (1.82 ± 0.17 mg).
3.2 Fatty acid analyses
3.2.1 Macroalgae
A total of 19 FAs were identified in Ulva, 17 in Fucus, 15 in Laminaria, 12 in Gracilaria, and 16 in Chondrus, with the number of carbons ranging from 14 to 24 (Supplementary Table 1). As expected, the overall FA profile of macroalgae significantly differed among species (PERMANOVA, F4,20 = 24.37, p = 0.001), with all species differing from each other in the pairwise comparisons and clearly segregating in the PCoA plot (Figure 4). The most abundant FA in Ulva, Laminaria, Gracilaria, and Chondrus was palmitic acid (16:0) (mean ± SD: 0.41 ± 0.03, 1.74 ± 1.56, 0.87 ± 0.19, 0.17 ± 0.06 µg mg−1 DW, respectively), while oleic acid (18:1n-9) was the most abundant FA in Fucus (3.28 ± 0.15 µg mg−1 DW) (Table 1). Although SFA were the most representative FA class in Ulva, Laminaria and Gracilaria, they were significantly more abundant in the brown macroalgae Laminaria and Fucus (F4,20 = 38.05, p < 0.001, Supplementary Figure 2). Similarly, MUFA and PUFA were significantly more abundant in Fucus, followed by Laminaria, with the first being the most common FA class found in Fucus and the second in Chondrus (Supplementary Figure 2). The ratio between n-3 and n-6 PUFAs significantly differed among macroalgae (F4,20 = 13.14, p < 0.001), being lower for Fucus (0.53 ± 0.03) and Laminaria (1.27 ± 0.90) and higher for Ulva (2.87 ± 0.45) (Table 1). On the contrary, the ratio between PUFA and SFA did not vary significantly among the macroalgal species tested (F4,20 = 0.99, p = 0.434).
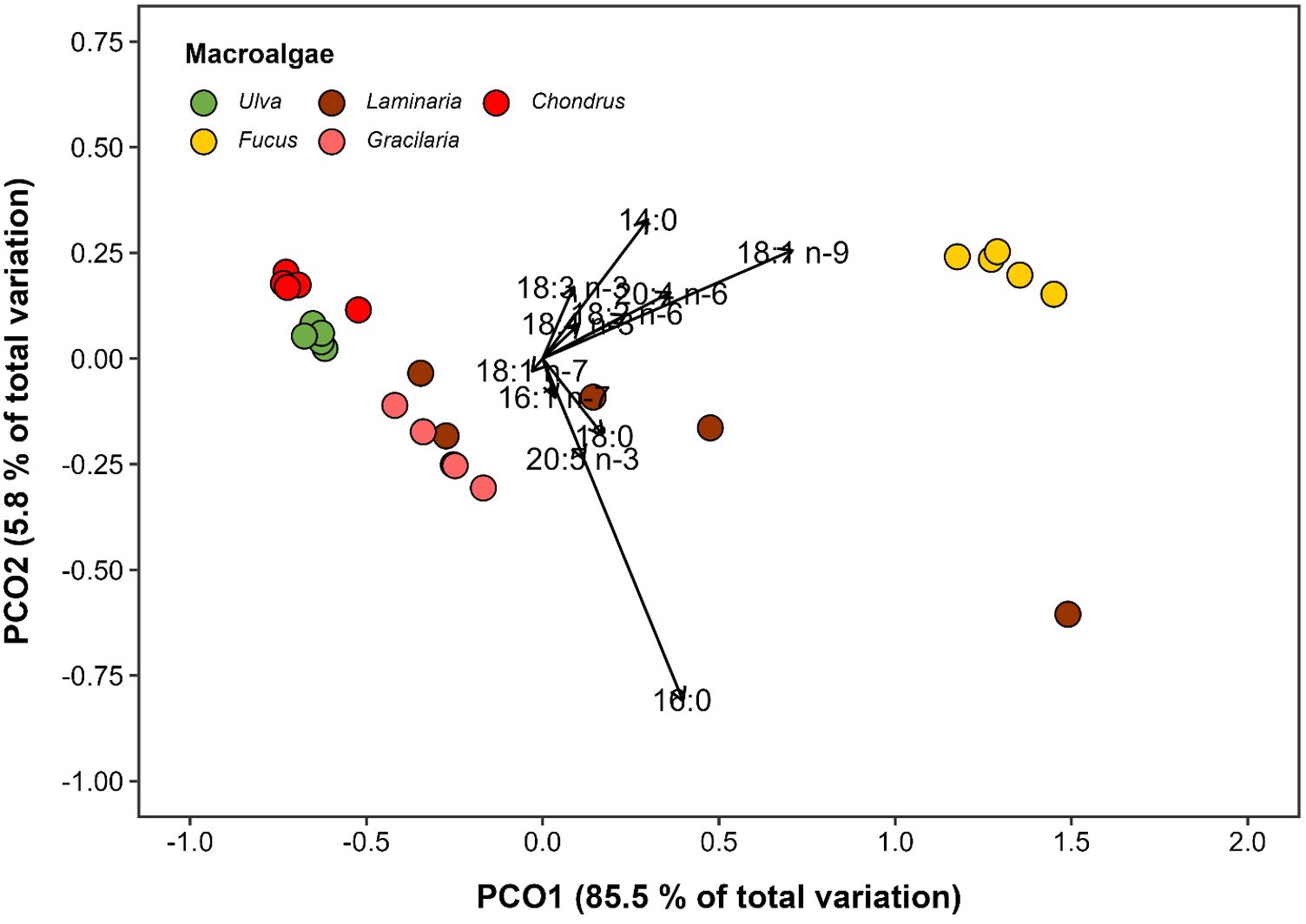
Figure 4. Principal Coordinate Analysis (PCoA) illustrating the variation in the fatty acid composition of the five macroalgal species (Ulva sp., Fucus sp., Laminaria sp., Gracilaria sp. and Chondrus sp.) used as mono-specific diets to Gammarus locusta broodstocks. Analysis was restricted to fatty acids in the samples accounting for more than 1% of total fatty acid abundance.
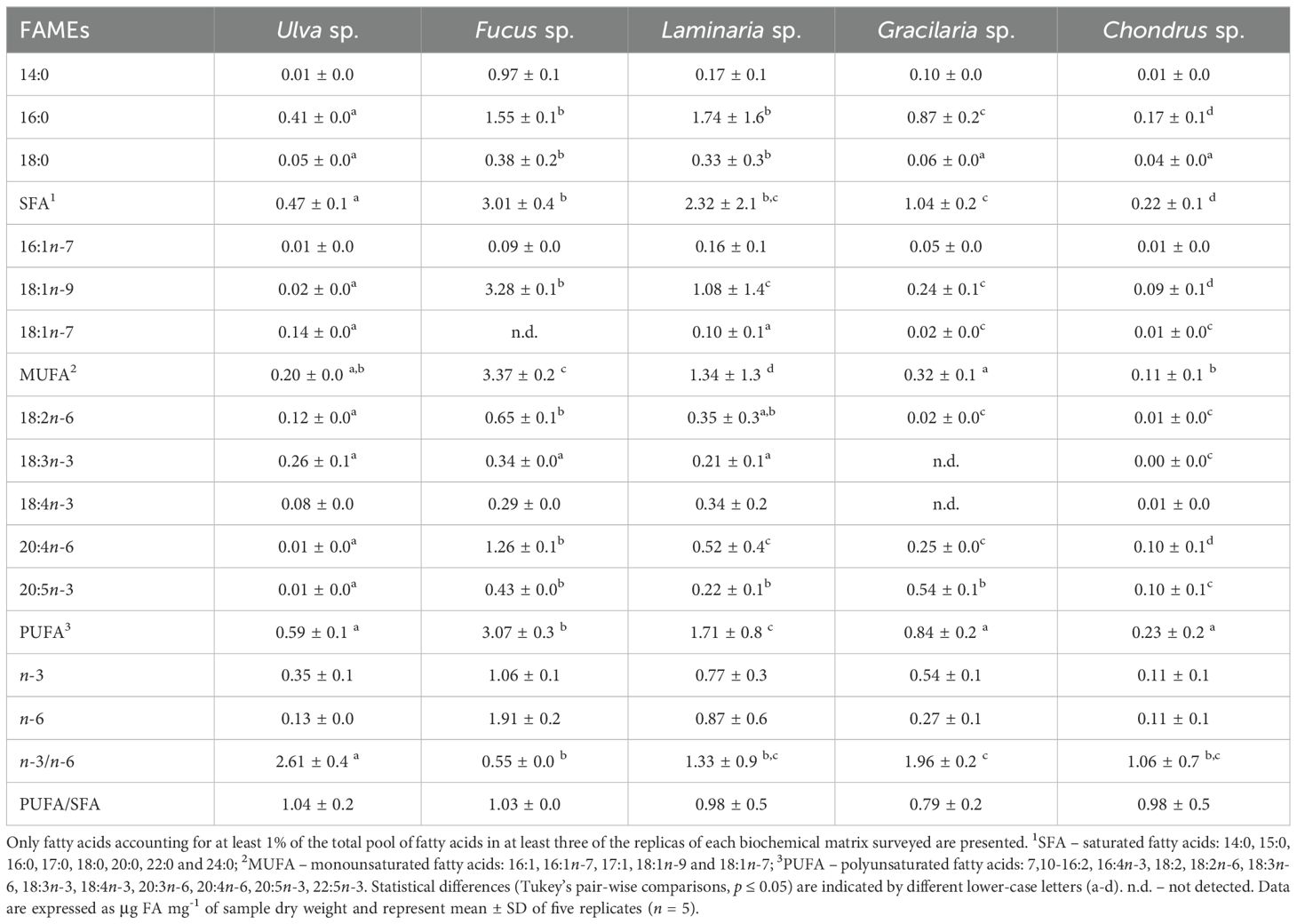
Table 1. Most abundant fatty acids (FAs) (i.e. those representing more than 1% of total FA abundance) identified in the five macroalgae species used as mono-specific diets in the feeding of the amphipod Gammarus locusta (Ulva sp., Fucus sp., Laminaria sp., Gracilaria sp., and Chondrus sp.).
3.2.2 Gammarus locusta offspring
A total of 22 FAs were identified in G. locusta’s newly hatched offspring, with the number of carbons ranging from 14 to 22 (Supplementary Table 2). The overall FA profile of G. locusta offspring significantly differed among the feeding treatments supplied to breeding pairs (PERMANOVA, F5,24 = 5.26, p = 0.001), with a clear segregation between the wild-derived offspring and that originated from macroalgal-fed adults (Figure 5). Such segregation was supported by pairwise comparisons, which indicated significant differences between all mono-specific algal diet treatments and wild-derived offspring (all p values < 0.01), as well as between offspring of G. locusta breeding pairs supplied with Ulva and Fucus (p = 0.019). Regardless of the feeding treatments supplied, the most abundant FA in the offspring of adults fed with a mono-specific macroalgal diet were, in decreasing order: palmitic acid (16:0; mean ± SD: from 2.11 ± 0.51 to 2.59 ± 0.08 μg FA mg-1 DW), EPA (20:5n-3; from 1.93 ± 0.72 to 2.42 ± 0.24 μg FA.mg-1 DW) and the oleic acid (18:1n-9; from 1.09 ± 0.38 to 1.66 ± 0.15 μg FA mg-1 DW) (Table 2). On the contrary, G. locusta offspring from the wild featured, in decreasing order of abundance, the following FAs: stearic acid (18:0; 2.88 ± 0.71 μg FA mg-1 DW), palmitic acid (16:0; 2.75 ± 0.76 μg FA mg-1 DW) and EPA (20:5n-3; 2.49 ± 0.21 μg FA mg-1 DW) (Table 2). According to the SIMPER analysis, stearic acid (18:0) was the fatty acid that most contributed to the dissimilarities recorded between the offspring of G. locusta from the wild and those amphipods fed with a mono-specific macroalgal diet and those exposed to the mono-specific algal diet treatments, accounting for more than 60% of the dissimilarities for all comparisons. It is also worth referring that oleic acid (18:1n-9) was the FA that contributed the most (30.75%) for the dissimilarities recorded on the FA profile of offspring from breeding pairs fed Ulva or Fucus (Supplementary Table 3).
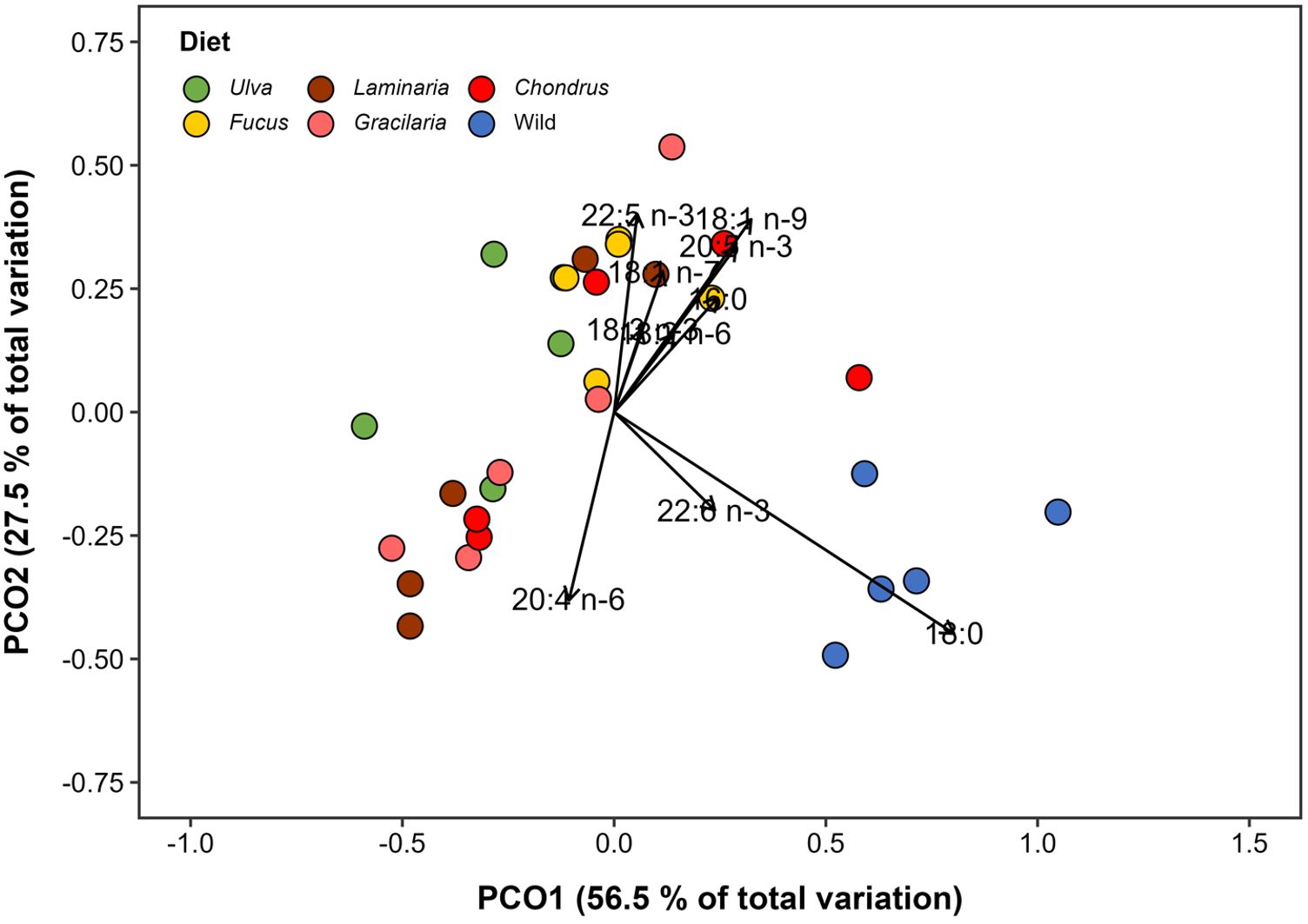
Figure 5. Principal Coordinate Analysis (PCoA) illustrating the variation in the fatty acid composition of Gammarus locusta offspring from breeding pairs fed with a mono-specific macroalgal diet (Ulva sp., Fucus sp., Laminaria sp., Gracilaria sp. and Chondrus sp.) or from specimens sourced from the wild. Analysis was restricted to fatty acids in the samples accounting for more than 1% of total fatty acid abundance.
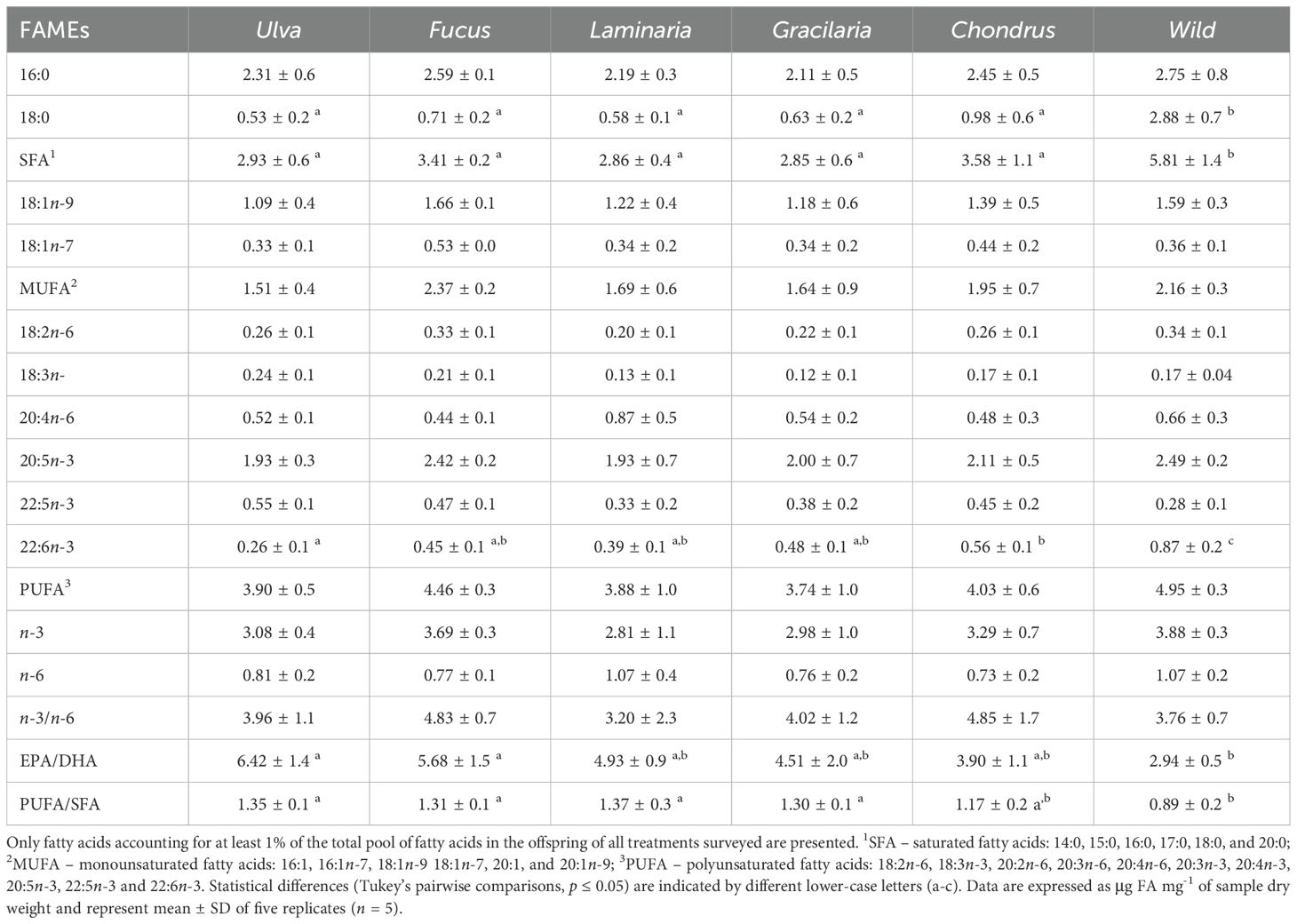
Table 2. Most abundant fatty acids (FAs) (i.e. those representing more than 1% of total FA abundance) identified in Gammarus locusta offspring originated from adult broodstock fed with five different macroalgal diets (Ulva sp., Fucus sp., Laminaria sp., Gracilaria sp., and Chondrus sp.), and Gammarus locusta originated from broodstock retrieved from the wild.
Concerning the univariate comparisons, there were significant differences in the abundance of stearic acid (18:0; F5,24 = 15.69, p < 0.001) between the different treatments, with offspring from the wild showing an abundance four times greater on average of this FA than those originated from breeding pairs fed with a mono-specific macroalgal diet (Table 2). Moreover, the FA profile of wild offspring displayed a notable contribution of both EPA and DHA, although only DHA levels (0.87 ± 0.16 μg mg-1 DW) were significantly higher (F5,24 = 13.64, p < 0.001) when compared to offspring from breeding pairs fed with a mono-specific macroalgal diet (from 0.26 ± 0.09 to 0.56 ± 0.10 μg mg-1 DW) (Table 2). As such, the EPA/DHA ratio significantly differed between treatments (F5,24 = 5.24, p = 0.006), being lower for offspring from the wild (2.94 ± 0.54) when compared to offspring from breeding pairs fed Ulva (6.42 ± 1.44) or Fucus (5.68 ± 1.48) (Table 2).
PUFA was the most abundant FA class in offspring from breeding pairs fed with a mono-specific macroalgal diet (ranging from 3.74 ± 1.02 to 4.46 ± 0.31 μg mg-1 DW). On the contrary, SFA was the most abundant FA class in the offspring of G. locusta from the wild (5.81 ± 1.4 μg mg-1 DW) and ranged between 2.85 ± 0.58 and 3.58 ± 1.12 μg mg-1 DW in offspring from breeding pairs fed the mono-specific macroalgal diets (Figure 6). Regardless of the treatment, MUFA was the least abundant FA class (ranging from 1.51 ± 0.44 to 2.37 ± 0.19 μg mg-1 DW), with offspring from breeding pairs fed on Fucus exhibiting the highest levels of MUFA, mostly due to the high abundance of oleic acid (18:1n-9) in their FA profile (Table 2). However, although the overall abundance of MUFA (F5,24 = 1.73, p = 0.165) and PUFA (F5,24 = 2.26, p = 0.081) were similar among treatments, the levels of SFA were significantly higher in wild offspring compared to conspecifics hatched from breeding pairs fed the mono-specific macroalgal diets (F5,24 = 8.61, p < 0.001) (Figure 6). Consequently, the PUFA/SFA ratio for wild offspring was significantly lower than that of the other experimental groups (F5,24 = 4.48, p = 0.005), which exhibited ratios above 1 (due to a higher abundance of PUFA over SFA) (Table 2). Finally, regardless of the treatment, G. locusta offspring always featured a greater abundance of n-3 rather than n-6 FAs, with the n-3/n-6 ratio being similar among the offspring of all experimental groups (F5,24 = 1.03, p = 0.421) and all of them exhibiting values above 1 (from 3.20 ± 2.26 to 4.85 ± 1.75) (Table 2).
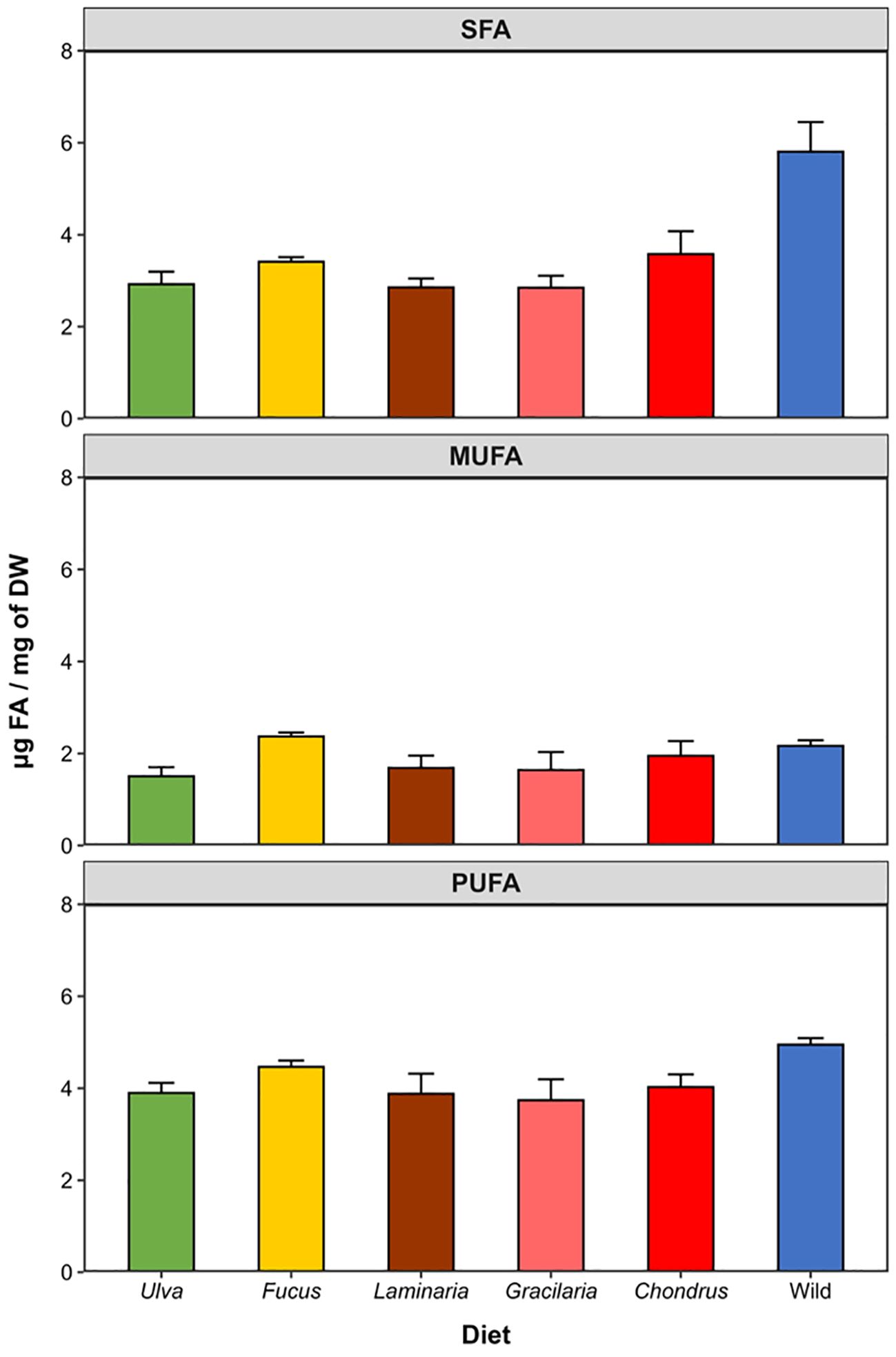
Figure 6. Abundance of fatty acid classes in the offspring of Gammarus locusta from the wild and breeding pairs fed with five different macroalgal diets (Ulva sp., Fucus sp., Laminaria sp., Gracilaria sp., and Chondrus sp.) or in G. locusta offspring originated from broodstock retrieved from the wild. Data are expressed as μg mg-1 DW and represent the mean ± SE of 5 replicates (n = 5). SFA – saturated fatty acids; MUFA – monounsaturated fatty acids; PUFA – polyunsaturated fatty acids.
The absolute differences recorded between the mean values of the most abundant FA identified in G. locusta offspring from breeding pairs fed mono-specific macroalgal diets, when contrasted with those of offspring from conspecifics from the wild, are displayed in Figure 7. In general, offspring derived from breeding pairs fed mono-specific macroalgal diets displayed lower average abundances for most of the FAs when compared to offspring from the wild, as indicated by the negative values in the plot (Figure 7). However, a few exceptions occurred for MUFA (18:1n-7 and 18:1n-9), with some levels being higher for Fucus and Chondrus treatments, and PUFA, with the levels of 18:3n-3 being slightly higher for Ulva and Laminaria, and the level of 20:4n-6 for offspring from breeding pairs fed with Fucus. Furthermore, 22:5n-3 was the single PUFA whose levels were consistently higher for offspring originating from breeding pairs fed a mono-specific macroalgal diet when compared with offspring from the wild (Figure 7).
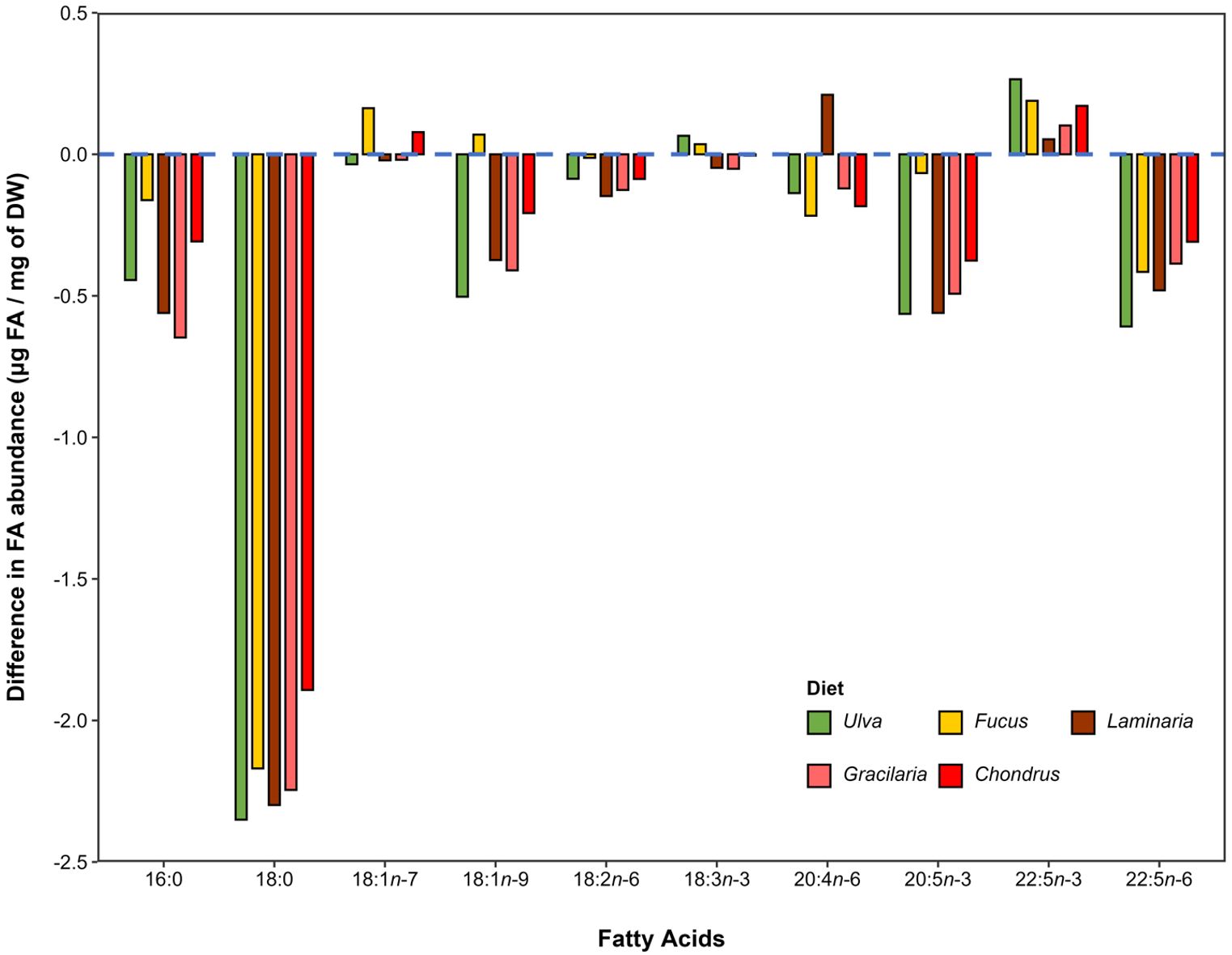
Figure 7. Absolute differences in the most abundant fatty acids (i.e. those accounting for more than 1% of their relative abundance calculated for total fatty acid abundance in the samples) between Gammarus locusta offspring from breeding pairs fed with mono-specific macroalgal diets (Ulva sp., Fucus sp., Laminaria sp., Gracilaria sp. and Chondrus sp.) and offspring from conspecifics from the wild. Bars represent the variation in the mean absolute abundance of the fatty acid (expressed as µg mg−1 DW) relative to offspring of conspecifics from the wild. Negative values indicate lower levels in macroalgae-fed treatments when compared to offspring of conspecifics from the wild, while positive values indicate higher levels. The dashed blue line set at 0 represents the baseline level of fatty acids for the offspring of G. locusta from the wild.
4 Discussion
Recently, gammarids have gained attention for their promising nutritional profile as a new ingredient/supplement for aquafeeds (Baeza-Rojano et al., 2013, 2014; Jiménez-Prada et al., 2018; Ribes-Navarro et al., 2022). Their ease of cultivation under different feeding substrates, including agricultural and aquaculture side-streams, further highlights their potential for the sustainable production of formulated feeds (Alberts-Hubatsch et al., 2019; Jiménez-Prada et al., 2020; Ribes-Navarro et al., 2022). However, few studies have examined the influence of gammarid diets on their FA composition, therefore limiting our understanding of how it can shape their nutritional profile (Alberts-Hubatsch et al., 2019; Ribes-Navarro et al., 2022). Additionally, to the author’s knowledge, no study has ever addressed the potential effects of diets on subsequent generations of these crustaceans. To address this gap, the present study determined the FA composition of G. locusta offspring from breeding pairs supplied mono-specific macroalgal diets and compared it to wild conspecifics, aiming to assess the impact of parental diets on the nutritional FA profile of their offspring.
Breeding pairs from the wild exhibited larger and heavier broods than conspecifics supplied with mono-specific macroalgal diets. Our findings align with those from previous studies that suggest that natural diets provide greater nutritional value to marine amphipods than mono-specific diets (Ribes-Navarro et al., 2022). Potential causes for varying brood sizes may include the limitation and/or exhaustion of some essential nutrients (Mccabe and Dunn, 1994), reinforcing previous research indicating that amphipods benefit from enhanced reproductive metabolism when provided with nutritionally rich and diverse feeding substrates (Clarke et al., 1985).
Concerning the macroalgae provided as diets, their FA relative content in the total FA composition was, in general, well-aligned with previous studies. Nevertheless, some apparent differences may be pointed out, as the values of EPA found in the red macroalgae employed in the present study (G. gracilis and C. crispus) were higher than those previously reported in the literature (Melo et al., 2015; Da Costa et al., 2017). Ulva sp. and the two brown macroalgae (Fucus sp. and Laminaria sp.) also displayed a similar composition when compared to other research (Schmid et al., 2016; Lorenzo et al., 2017; Marques et al., 2021), except for the higher values recorded in the present study for linolenic acid (18:3n-3) and oleic acid (18:1n-9), respectively.
Conversely to what has been described for G. locusta cultured with macroalgae and analysed at sexual maturity (Alberts-Hubatsch et al., 2019; Ribes-Navarro et al., 2022), offspring from breeding pairs fed the mono-specific macroalgal diets showed a similar FA composition between them, regardless of broodstock diet, with the sole exception being for Ulva and Fucus. The sole comparison that can be performed with our study in terms of newly hatched offspring is that with Gammarus oceanicus, with offspring hatching in the spring or in the winter (Clarke et al., 1985). In our study, newly hatched G. locusta from breeding pairs fed a mono-specific macroalgal diet displayed higher values of linolenic acid (18:3n-3), arachidonic acid (20:4n-6) and EPA, but not for DHA, which was found in higher concentration only in newly hatched conspecifics from the wild when compared to winter-born G. oceanicus. These results suggest that, despite diet being an important driver in the modulation of the FA composition of G. locusta (Ribes-Navarro et al., 2022), this species appears to display a selective retention and utilisation of some dietary FA, particularly LC-PUFA, which are known to be essential for their growth and reproduction (Clarke et al., 1985). This is most relevant for DHA, as this FA is paramount for nervous system function and plays a key role in the development of marine organisms (Tocher et al., 2019), with its incorporation influencing the cell membrane’s fluidity and permeability (Stillwell and Wassall, 2003). Indeed, as DHA and EPA play an important role in crustacean development and moulting, their requirements are most likely higher during early life stages, as these are known to moult more frequently to allow somatic growth (Wang et al., 2021). In a study with a freshwater gammarid species, Pallaseopsis quadrispinosa, Taipale et al. (2021) showed that, when starved for 12 days, EPA and DHA appeared to be preferentially retained, most likely to maintain cell membrane functions. Therefore, P. quadrispinosa do seem to be able to preserve physiologically essential FAs when nutritionally limited by their diet (Taipale et al., 2021). As such, it would not be surprising if G. locusta displayed a similar adaptation to safeguard survival. Under nutrient limitation, breeding pairs likely conserved or biosynthesised a baseline level of essential LC-PUFAs (Alberts-Hubatsch et al., 2019; Taipale et al., 2021; Ribes-Navarro et al., 2022), such as DHA, which was transmitted to the offspring, despite its absence from the parental diets (Melo et al., 2015; Schmid et al., 2016; Da Costa et al., 2017; Lorenzo et al., 2017; Marques et al., 2021). This may explain the lack of significant differences in the FA profiles of G. locusta offspring across macroalgal treatments. Parental nutrition is known to be a potential driver for the occurrence of carry-over (or latent) effects, which can shape offspring performance post-hatching (Calado and Leal, 2015). Briefly, carry-over effects can be defined as traits that have their origin during the development history of an organism (e.g., during embryonic development in G. locusta, as it displays no larval stages) that commonly stay latent for a given period and subsequently re-emerge at juvenile and/or adult stages (Pechenik, 2006). Maternal investment is particularly important in species featuring direct development (as displayed by G. locusta), as the energetic burden of this reproductive strategy is commonly higher than when species hatch as a less developed larval stage. Moreover, when breeding pairs of a species that exhibits direct development are exposed to nutritional constraints (such as the mono-specific macroalgal diets tested in our study), it may not be possible for them to successfully allocate all the nutrients required to ensure an optimal growth performance and survival of their offspring. As such, future studies should document the post-hatching performance of G. locusta offspring hatching from breeding pairs fed on mono-specific macroalgal diets, as it would not be surprising to detect the occurrence of carry-over effects. Moreover, given the short time required to produce sexually mature G. locusta in the laboratory from newly hatched specimens (two to three months), the potential existence of transgenerational effects should also be investigated, as such traits remain poorly elucidated (Calado and Leal, 2015).
The findings of our present study appear to support the hypothesis that females may rely on endogenous biosynthetic pathways to maintain a stable FA provisioning to their offspring (Alberts-Hubatsch et al., 2019; Ribes-Navarro et al., 2022), thus ensuring that the energetic demands of reproduction are met either through food intake or the catabolism of energy reserves (Clarke et al., 1985). The potential capacity of gammarids for trophic upgrading has already been put forward by several authors (Jiménez-Prada et al., 2018; Alberts-Hubatsch et al., 2019; Jiménez-Prada et al., 2020; Ribes-Navarro et al., 2022), suggesting that these crustaceans do feature the ability to bioconvert shorter-chain and more saturated FA present in macroalgae into LC-PUFA (Ribes-Navarro et al., 2022). The gene expression and the closely linked function of two different types of enzymes, such as desaturases and elongases, are particularly important for the biosynthesis of LC-PUFA (Monroig et al., 2022; Monroig and Kabeya, 2018; Ribes-Navarro et al., 2022). Nevertheless, the presence and activity of front-end desaturases in marine gammarid amphipods remains unclear (Ribes-Navarro et al., 2021, 2023), with the metabolic pathway(s) that allow the biosynthesis of LC-PUFA yet to be clarified. A recent investigation on the brine shrimp Artemia franciscana addressed the presence of desaturases that operate only on C18 FAs, with limited ability to target LC-PUFA (Ramos-Llorens et al., 2023). Another study on three distinct elongases in the marine gammarid species Marinogammarus marinus suggests that these may be able to produce LC-PUFA endogenously, although they also lack the front-end desaturases that produce key LC-PUFAs such as DHA (Ribes-Navarro et al., 2021). On the other hand, gammarids can also be associated with other organisms which feature essential enzymes that play a role in the biosynthesis of LC-PUFA, such as epibiont rotifers associating with freshwater Gammarus sp (Ribes-Navarro et al., 2023). Overall, the pathway that allows marine gammarids, such as M. marinus and G. locusta to perform a trophic upgrade of their diet by biosynthesising LC-PUFA remains elusive, as the presence of genes encoding front-end desaturases that allow the biosynthesis of certain LC-PUFA is yet to be recorded in their genome (Ribes-Navarro et al., 2021, 2023).
From a production point of view, the findings of the present study underscore the need to refine broodstock diets to optimise the reproductive yield of gammarid amphipods, as indicated by the low number of specimens per brood and low DW obtained from breeding pairs supplied mono-specific macroalgal diets. Nonetheless, it is worth highlighting that the FA composition of gammarid amphipods is nutritionally adequate for several aquaculture applications (Baeza-Rojano et al., 2013, 2014; Jiménez-Prada et al., 2018, 2020; Alberts-Hubatsch et al., 2019), even under nutritionally limited conditions such as the use of mono-specific macroalgal diets or when fed with non-marine side streams (Alberts-Hubatsch et al., 2019; Ribes-Navarro et al., 2022). Future research should explore the potential effects of mono-specific diets over consecutive generations of G. locusta, investigating the potential existence of nutritional programming, as well as evaluating the use of non-marine diets, such as agricultural co-products/side streams, to further enhance the circularity of their large-scale production.
In conclusion, our study revealed that the offspring of G. locusta hatching from parental organisms fed on mono-specific macroalgal diets always displayed DHA on their fatty acid profiles, regardless of the absence of this n-3 LC-PUFA on parental diets. Moreover, the offspring from wild G. locusta always showed higher levels of PUFA, as well as DHA, opening a window of opportunity to improve parental diets based on macroalgae that can be used in the farming of marine gammarid amphipods. Further research is needed to fully elucidate the mechanisms underlying the trophic upgrade performed by marine gammarid amphipods when supplied with diets that feature little or even no LC-PUFA, namely EPA and DHA. Testing different mono-specific and multi-specific diets, from both marine and non-marine origins, while also investigating the potential existence of nutritional programming, will allow advancing the state of the art of marine gammarid amphipods’ large-scale production. Moreover, by complementing these actions with the selective breeding of specimens that allow to establish lineages that can either match or even over-perform conspecifics in the wild in terms of reproductive output and FA content, one can truly unleash the potential of marine gammarid amphipods for the formulation of high-quality aquaculture feeds.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The manuscript presents research on animals that do not require ethical approval for their study.
Author contributions
JS: Conceptualization, Data curation, Formal Analysis, Investigation, Methodology, Writing – original draft, Writing – review & editing. LM: Data curation, Formal Analysis, Supervision, Validation, Visualization, Writing – review & editing. DPR: Data curation, Formal Analysis, Supervision, Validation, Visualization, Writing – review & editing. FR: Software, Writing – review & editing, Data curation, Formal Analysis, Funding acquisition, Methodology, Supervision, Validation, Visualization. MA: Conceptualization, Investigation, Methodology, Writing – review & editing. RCD: Resources, Supervision, Validation, Visualization, Writing – review & editing. MRMD: Data curation, Formal Analysis, Software, Validation, Visualization, Writing – review & editing. RC: Conceptualization, Funding acquisition, Methodology, Project administration, Resources, Supervision, Validation, Visualization, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This study was performed under the scope of the project “PUFAPODS: Merging blue and green food systems - Using marine gammarid amphipods supplied with plant food processing side streams to produce n-3 LC-PUFA”, which is supported by FCT/MEC (https://doi.org/10.54499/2022.01620.PTDC) and project “BLUE BIOECONOMY PACT” (Project N°. C644915664-00000026), co-funded by the Next Generation EU European Fund, under the incentive line “Agendas for Business Innovation” within Component 5—Capitalization and Business Innovation of the Portuguese Recovery and Resilience Plan (RRP), specifically under the vertical WP7 – Fish (LowTrophAqua). We thank the University of Aveiro, Fundação para a Ciência e Tecnologia (FCT), and Ministério da Ciência Tecnologia e Ensino Superior (MCTES) for financial support to the research units UID Centro de Estudos do Ambiente e Mar (CESAM) + LA/P/0094/2020 and UID/50006 -Laboratório Associado para a Química Verde - Tecnologias e Processos Limpos. The authors acknowledge FCT/MCTES for individual funding in the scope of the Individual Call to Scientific Employment Stimulus (CEECIND/00580/2017 to Felisa Rey (https://doi.org/10.54499/CEECIND/00580/2017/CP1459/CT0005).
Acknowledgments
We acknowledge Maria Adelaide Salvador for the help with the lipid extractions and fatty acid analysis methodology. We thank Simão Maia for helping with the fatty acid analysis software and for his expertise on laboratory techniques, data organisation and presentation. The authors would like to thank two reviewers for their constructive criticism of the original version of our manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2025.1621216/full#supplementary-material.
References
Alberts-Hubatsch H., Slater M. J., and Beermann J. (2019). Effect of diet on growth, survival and fatty acid profile of marine amphipods: implications for utilisation as a feed ingredient for sustainable aquaculture. Aquaculture Environ. Interact. 11, 481–491. doi: 10.3354/AEI00329
Álvarez A., Fontanillas R., Hernández-Contreras A., and Hernández M. D. (2020). Partial replacement of fish oil with vegetal oils in commercial diets: The effect on the quality of gilthead seabream (Sparus aurata). Anim. Feed Sci. Technol. 265, 114504. doi: 10.1016/j.anifeedsci.2020.114504
Anderson M. J. (2001). A new method for non-parametric multivariate analysis of variance. Austral Ecol. 26, 32–46. doi: 10.1111/J.1442-9993.2001.01070.PP.X
Arbizu P. M. (2017). Pairwise Multilevel Comparison using Adonis. Available online at: https://Github.Com/Pmartinezarbizu/PairwiseAdonis (Accessed April 15, 2017).
Baeza-Rojano E., Domingues P., Guerra-García J. M., Capella S., Noreña-Barroso E., Caamal-Monsreal C., et al. (2013). Marine gammarids (crustacea: Amphipoda): A new live prey to culture Octopus maya hatchlings. Aquaculture Res. 44, 1602–1612. doi: 10.1111/j.1365-2109.2012.03169.x
Baeza-Rojano E., Hachero-Cruzado I., and Guerra-García J. M. (2014). Nutritional analysis of freshwater and marine amphipods from the Strait of Gibraltar and potential aquaculture applications. J. Sea Res. 85, 29–36. doi: 10.1016/j.seares.2013.09.007
Bligh E. G. and Dyer W. J. (1959). A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 37, 911–917. doi: 10.1139/o59-099
Calado R. and Leal M. C. (2015). Trophic ecology of benthic marine invertebrates with bi-phasic life cycles: what are we still missing? Adv. Mar. Biol. 71, 1–70. doi: 10.1016/bs.amb.2015.07.001
Christie W. W. (2023). The lipidweb. Available online at: https://www.lipidmaps.org/resources/lipidweb/index.php?page=ms/methesters/me-arch/index.htm (Accessed November 15, 2023).
Clarke A., Skadsheim A., and Holmes L. J. (1985). Lipid biochemistry and reproductive biology in two species of Gammaridae (Crustacea: Amphipoda). Mar. Biol. 88, 247–263. doi: 10.1007/BF00392587
Costa F. O. and Costa M. H. (2000). Review of the ecology of Gammarus locusta (L.). Polskie Archiwum Hydrobiologii 47, 541–559.
Cottrell R. S., Blanchard J. L., Halpern B. S., Metian M., and Froehlich H. E. (2020). Global adoption of novel aquaculture feeds could substantially reduce forage fish demand by 2030. Nat. Food 1, 301–308. doi: 10.1038/s43016-020-0078-x
Da Costa E., Melo T., Moreira A. S. P., Bernardo C., Helguero L., Ferreira I., et al. (2017). Valorization of lipids from Gracilaria sp. through lipidomics and decoding of antiproliferative and anti-inflammatory activity. Mar. Drugs 15, 62. doi: 10.3390/md15030062
FAO (2024). “The state of world fisheries and aquaculture 2024,” in The state of world fisheries and aquaculture (Rome: FAO). doi: 10.4060/cd0683en
Glencross B. D., Bachis E., Betancor M. B., Calder P., Liland N., Newton R., et al. (2024). Omega-3 futures in aquaculture: exploring the supply and demands for long-chain omega-3 essential fatty acids by aquaculture species. Review Fisheries Sci. Aquaculture. 33 (2), 167–216. doi: 10.1080/23308249.2024.2388563
Glencross B., Fracalossi D. M., Hua K., Izquierdo M., Mai K., Øverland M., et al. (2023). Harvesting the benefits of nutritional research to address global challenges in the 21st century. J. World Aquaculture Soc. 54, 343–363. doi: 10.1111/jwas.12948
Harlıoğlu M. M. and Farhadi A. (2018). Importance of gammarus in aquaculture. Aquaculture Int. 26, 1327–1338. doi: 10.1007/s10499-018-0287-6
Horton T. (2023). WoRMS - world register of marine species. Available online at: https://www.marinespecies.org/aphia.php?p=taxdetails&id=102303 (Accessed March 30, 2023).
Jiménez-Prada P., Hachero-Cruzado I., Giráldez I., Fernández-Diaz C., Vilas C., Cañavate J. P., et al. (2018). Crustacean amphipods from marsh ponds: A nutritious feed resource with potential for application in Integrated Multi-Trophic Aquaculture. PeerJ 2018, 4194. doi: 10.7717/peerj.4194
Jiménez-Prada P., Hachero-Cruzado I., and Guerra-García J. M. (2020). Aquaculture waste as food for amphipods: the case of Gammarus insensibilis in marsh ponds from southern Spain. Aquaculture Int. 29, 139–153. doi: 10.1007/s10499-020-00615-z
Lenth R. V. (2017). emmeans: estimated marginal means, aka least-squares means. CRAN: Contributed Packages. doi: 10.32614/CRAN.PACKAGE.EMMEANS
Lorenzo J. M., Agregán R., Munekata P. E. S., Franco D., Carballo J., Şahin S., et al. (2017). Proximate composition and nutritional value of three macroalgae: Ascophyllum nodosum, Fucus vesiculosus and Bifurcaria bifurcata. Mar. Drugs 15, 360. doi: 10.3390/md15110360
Lüdecke D., Ben-Shachar M. S., Patil I., Waggoner P., and Makowski D. (2021). performance: an R package for assessment, comparison and testing of statistical models. J. Open Source Software 6, 3139. doi: 10.21105/JOSS.03139
Maia S., Rey F., Domingues M. R., Figueira J., Dupont S., Leandro S. M., et al. (2025). Fatty acid profile of newly extruded embryos of brown crab (Cancer pagurus) and potential value of their lipids for human consumption. Appl. Food Res. 5, 100773. doi: 10.1016/J.AFRES.2025.100773
Marques L., Domingues M. R., da Costa E., Abreu M. H., Lillebø A. I., and Calado R. (2021). Screening for health-promoting fatty acids in ascidians and seaweeds grown under the influence of fish farming activities. Mar. Drugs 19, 469. doi: 10.3390/md19080469
Mccabe J. and Dunn A. M. (1994). Is resource partitioning among offspring a response to brood sex ratio in an amphipod with environmental sex determination? Oikos 69, 203–206. doi: 10.2307/3546139
Melo T., Alves E., Azevedo V., Martins A. S., Neves B., Domingues P., et al. (2015). Lipidomics as a new approach for the bioprospecting of marine macroalgae - unraveling the polar lipid and fatty acid composition of Chondrus crispus. Algal Res. 8, 181–191. doi: 10.1016/j.algal.2015.02.016
Monroig Ó. and Kabeya N. (2018). Desaturases and elongases involved in polyunsaturated fatty acid biosynthesis in aquatic invertebrates: a comprehensive review. Fisheries Sci. 84, 911–928. doi: 10.1007/s12562-018-1254-x
Monroig, Shu-Chien A. C., Kabeya N., Tocher D. R., and Castro L. F. C. (2022). Desaturases and elongases involved in long-chain polyunsaturated fatty acid biosynthesis in aquatic animals: From genes to functions. Prog. Lipid Res. 86, 101157. doi: 10.1016/j.plipres.2022.101157
Napier J. A. and Betancor M. B. (2023). Engineering plant-based feedstocks for sustainable aquaculture. Curr. Opin. Plant Biol. 1, 102323. doi: 10.1016/j.pbi.2022.102323
Naylor R. L., Hardy R. W., Buschmann A. H., Bush S. R., Cao L., Klinger D. H., et al. (2021). A 20-year retrospective review of global aquaculture. Nature 591, 551–563. doi: 10.1038/s41586-021-03308-6
Oksanen J., Simpson G., Blanchet F., Kindt R., Legendre P., Minchin P., et al. (2025). “…,” in Package “vegan” Title community ecology package. Available online at: https://github.com/vegandevs/vegan (Accessed April 15, 2025).
Paradis E. and Schliep K. (2019). ape 5.0: an environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 35, 526–528. doi: 10.1093/bioinformatics/bty633
Pechenik J. A. (2006). Larval experience and latent effects - Metamorphosis is not a new beginning. Integr. Comp. Biol. 46, 323–333. doi: 10.1093/icb/icj028
Quintino V., Sangiorgio F., Ricardo F., Mamede R., Pires A., Freitas R., et al. (2009). In situ experimental study of reed leaf decomposition along a full salinity gradient. Estuarine Coast. Shelf Sci. 85, 497–506. doi: 10.1016/j.ecss.2009.09.016
Ramos-Llorens M., Ribes-Navarro A., Navarro J. C., Hontoria F., Kabeya N., and Monroig Ó. (2023). Can Artemia franciscana produce essential fatty acids? Unveiling the capacity of brine shrimp to biosynthesise long-chain polyunsaturated fatty acids. Aquaculture 563, 738869. doi: 10.1016/j.aquaculture.2022.738869
R Core Team (2025). A language and environment for statistical computing (Vienna, Austria: R Foundation for Statistical Computing). Available online at: https://www.r-project.org/.
Rey F., Cartaxana P., Cruz S., Melo T., and Domingues M. R. (2023). Revealing the polar lipidome, pigment profiles, and antioxidant activity of the giant unicellular green alga, Acetabularia acetabulum. J. Phycology 59, 1025–1040. doi: 10.1111/JPY.13367
Rey F., Greenacre M., Silva Neto G. M., Bueno-Pardo J., Domingues M. R., and Calado R. (2022). Fatty acid ratio analysis identifies changes in competent meroplanktonic larvae sampled over different supply events. Mar. Environ. Res. 173, 105517. doi: 10.1016/j.marenvres.2021.105517
Ribes-Navarro A., Alberts-Hubatsch H., Monroig Ó., Hontoria F., and Navarro J. C. (2022). Effects of diet and temperature on the fatty acid composition of the gammarid Gammarus locusta fed alternative terrestrial feeds. Front. Mar. Sci. 9. doi: 10.3389/FMARS.2022.931991/BIBTEX
Ribes-Navarro A., Kabeya N., Castro F. L. C., Gomes-Dos-Santos A., Fonseca M. M., Alberts-Hubatsch H., et al. (2023). Examination of gammarid transcriptomes reveals a widespread occurrence of key metabolic genes from epibiont bdelloid rotifers in freshwater species. Open Biol. 13, 230196. doi: 10.1098/rsob.230196
Ribes-Navarro A., Navarro J. C., Hontoria F., Kabeya N., Standal I. B., Evjemo J. O., et al. (2021). Biosynthesis of long-chain polyunsaturated fatty acids in marine gammarids: Molecular cloning and functional characterisation of three fatty acyl elongases. Mar. Drugs 19, 226. doi: 10.3390/MD19040226
Rodrigues D. P., Ameixa O. M. C. C., Vázquez J. A., and Calado R. (2022). Improving the lipid profile of Black Soldier Fly (Hermetia illucens) larvae for marine aquafeeds: Current state of knowledge. Sustainability (Switzerland) 14, 6472. doi: 10.3390/su14116472
Rollin M., Coulaud R., Danger M., Sohm B., Flayac J., Bec A., et al. (2017). Additive effect of calcium depletion and low resource quality on Gammarus fossarum (Crustacea, Amphipoda) life history traits. Environ. Sci. pollut. Res. 25, 11264–11280. doi: 10.1007/s11356-017-9390-x
Sadegh Alavi-Yeganeh M., Abedian Kenari A., and Rezaei M. (2008). Effect of Gammarus powder as a supplementary diet on growth and survival of rainbow trout larvae (Oncorhynchus mykiss). Pajouhesh Sazandegi 77, 113–123.
Schmid M., Guihéneuf F., and Stengel D. B. (2016). Evaluation of food grade solvents for lipid extraction and impact of storage temperature on fatty acid composition of edible seaweeds Laminaria digitata (Phaeophyceae) and Palmaria palmata (Rhodophyta). Food Chem. 208, 161–168. doi: 10.1016/j.foodchem.2016.03.123
Silva R. X. G., Cartaxana P., and Calado R. (2021). Prevalence and photobiology of photosynthetic dinoflagellate endosymbionts in the nudibranch Berghia stephanieae. Animals 11, 2200. doi: 10.3390/ani11082200
Stillwell W. and Wassall S. R. (2003). Docosahexaenoic acid: Membrane properties of a unique fatty acid. Chem. Phys. Lipids 126, 1–27. doi: 10.1016/S0009-3084(03)00101-4
Taipale S. J., Kers E., Peltomaa E., Loehr J., and Kainz M. J. (2021). Selective fatty acid retention and turnover in the freshwater amphipod Pallaseopsis quadrispinosa. Biomolecules 11, 478. doi: 10.3390/biom11030478
Tocher D. R., Betancor M. B., Sprague M., Olsen R. E., and Napier J. A. (2019). Omega-3 long-chain polyunsaturated fatty acids, EPA and DHA: Bridging the gap between supply and demand. Nutrients 11, 89. doi: 10.3390/nu11010089
Wang X., Jin M., Cheng X., Hu X., Zhao M., Yuan Y., et al. (2021). Dietary DHA/EPA ratio affects growth, tissue fatty acid profiles and expression of genes involved in lipid metabolism in mud crab Scylla paramamosain supplied with appropriate n-3 LC-PUFA at two lipid levels. Aquaculture 532, 736028. doi: 10.1016/j.aquaculture.2020.736028
Keywords: aquafeeds, fatty acids, GC-MS, LC-PUFA, marine gammarids, trophic upgrading
Citation: Sousa JP, Marques L, Rodrigues DP, Rey F, Albuquerque M, Domingues MRM, Duarte RC and Calado R (2025) Fatty acid profile of Gammarus locusta newly hatched offspring from the wild and from adults farmed on mono-specific macroalgal diets. Front. Mar. Sci. 12:1621216. doi: 10.3389/fmars.2025.1621216
Received: 30 April 2025; Accepted: 30 June 2025;
Published: 18 July 2025.
Edited by:
Kwaku Amoah, Guangdong Ocean University, ChinaReviewed by:
Amit Ranjan, Tamil Nadu Fisheries University, IndiaSahya Maulu, University of Plymouth, United Kingdom
Copyright © 2025 Sousa, Marques, Rodrigues, Rey, Albuquerque, Domingues, Duarte and Calado. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ricardo Calado, cmpjYWxhZG9AdWEucHQ=; João P. Sousa, anBzb3VzYTE3QHVhLnB0