 Eric E. Grossman
Eric E. Grossman Ferdinand K. J. Oberle
Ferdinand K. J. Oberle Curt D. Storlazzi
Curt D. Storlazzi- U.S. Geological Survey, Santa Cruz, CA, United States
Coral reef mortality around the world is accelerating due to human activities and rising sea temperatures that cause bleaching, which is expected to become more frequent. Our ability to predict which corals will be most resilient, however, remains limited due to insufficient information characterizing nearshore temperature and habitat conditions. In this study, we examine how submarine groundwater discharge (SGD) reduces nearshore water temperatures and exposure of corals to heat stress, complementing the understanding that SGD can adversely affect coral when it contains elevated nutrient concentrations. Data from fixed nearshore sensors and vertical depth profiles along ~100 km of the western shoreline of the Island of Hawai’i from 2003 to 2014 demonstrate that submarine groundwater discharge (SGD) can reduce nearshore water temperatures by 1 °C–5°C and create estuarine-like conditions with salinities as low as 20 PSU, where the prevalent coral species, Pocillopora meandrina, Porites lobata, and Montipora capitata, thrive. Time-series temperature records reveal that exposure to high ambient ocean temperatures, which are known to initiate bleaching events, are reduced up to 5%–46% of the time. Coral health surveys indicated coral bleaching in response to moderately high annual temperatures in 2010 and 2011, with more colonies affected farther from cold, SGD-fed waters. Synthesis of these results, along with coral response data following the more extreme marine heat wave of 2014–2015, demonstrates lower coral loss and greater coral recovery near groundwater seeps, particularly those with higher flux and influence on reducing nearshore water temperatures. Our results demonstrate that SGD may therefore provide a beneficial ecosystem service and enhance coral reef resilience, particularly where human-related nutrient additions to groundwater can be mitigated. The implications of our findings are relevant across tropical coasts where groundwater inputs can be substantial, such as the Caribbean and Indo-Pacific, and contribute to improving our understanding of coral sensitivity to gradients in temperature and nutrient stress. Improved management of groundwater resources could thus be vital to local–regional strategies for mitigating future heat stress.
Highlights
● Thermal modulation: Submarine groundwater discharge (SGD) injects cooler (up to 5°C) bottom water temperatures along a coral reef-lined, tropical coast.
● Mitigation of thermal stress: Tidal pumping of SGD substantially reduced coral exposure (up to 46%) to high summer nearshore temperatures that can initiate coral bleaching and mortality.
● Implications for coastal resource management: Improved understanding and management of shallow, coastal groundwater as a vector for cooler bottom water and, conversely, land-derived nutrients or contaminants, are critical to evaluating net effects on coral reef health.
1 Introduction
Nearshore coral reefs rank among the most diverse and productive ecosystems on Earth, supporting more than 25% of all marine species (Connell, 1978). Despite their ecological importance, global coral cover has declined by over 50% in recent decades (Hughes et al., 2018; Jackson et al., 2014). Coral reefs provide ecosystem services valued at ~10 trillion US dollars per year in the form of fisheries production, recreational opportunities, and coastal shoreline protection (Costanza et al., 2014), and an estimated 0.5–1.0 billion people worldwide depend on coral reefs (Anthony et al., 2017). Whereas localized impacts to coral reefs from pollution and overfishing are increasing, the rise in ocean temperatures beyond corals’ upper thermal thresholds (Kleypas et al., 1999) is the principal global driver of coral loss (Hughes et al., 2018; Bruno et al., 2019). Acute and prolonged thermal stress causes the breakdown of the coral–algal symbiosis, thereby reducing the corals’ food supply—known as bleaching—that can result in coral mortality (Glynn, 1996). Four declared global coral reef bleaching events have occurred during 1997–1998, 2009–2010, 2014–2017, and most recently 2023–2024 (Souter et al., 2021; Reimer et al., 2024; Goreau and Hayes, 2024). Thermal stress is anticipated to increase in the coming decades (Van Hooidonk et al., 2013, 2016) and to place ~90% of the world’s reefs at risk of ecological collapse (Frieler et al., 2013).
Water temperature across coral reef systems can be highly variable at spatial scales of millimeters to kilometers due to the complex interaction of atmospheric, terrestrial, and coastal/marine processes with reef topography and depth, which can all affect mixing and the extent of thermal stress on corals (Jokiel and Brown, 2004; Oberle et al., 2022; Muñiz-Castillo et al., 2024). Temperature and heat exposure driven by residence time of waters also affect many non-coral constituents of reefs that indirectly and cumulatively can affect coral health and mortality. For example, harmful algal blooms, cyanobacterial outbreaks, and grazers that jeopardize coral tissue likely precondition coral susceptibility to stress, but their relative role along with temperature stress in coral mortality is poorly understood (Couch et al., 2014; Klein et al., 2024). Coral bleaching due to prolonged heat stress is commonly characterized by “Degree Heating Weeks” (DHW), a metric quantifying the accumulation of heat stress over a 12-week period (Liu et al., 2003). Heat stress for the widely used DHW metric is defined as sea-surface temperatures >1°C above the expected long-term average for a given location and time of year and is generally measured by satellite. It therefore portrays the temperature in the uppermost few millimeters of the sea surface, correcting for depth where data exist, and a DHW of 4 °C-weeks or higher signifies a high risk of significant coral bleaching.
Submarine groundwater discharge (SGD), defined as groundwater that seeps through the beach face and/or seafloor and driven by terrestrial and marine forcings, can be a prominent source of cooler, less salty water to the nearshore (Moore, 2010). SGD can create estuarine-like conditions in otherwise marine environments, and it influences the thermal, salinity, and nutrient budgets along tropical coral reef-lined coasts (Kikuchi, 1976). Corals typically thrive in oligotrophic, saline waters, and whereas corals can benefit from slight nutrient additions sourced from SGD (Kim et al., 2011; Silbiger et al., 2020), elevated nutrient concentrations in SGD adversely affect coral health (Luijendijk et al., 2020; Aguiar et al., 2023). Nutrient excesses or imbalances in the nearshore can result in shifts in predator–prey relationships, competition, population booms of grazers, bioeroders, or invasive species, which can adversely affect corals (Fabricius, 2005; Klein et al., 2024). SGD can naturally contain high nutrient concentrations, but land use activities commonly elevate nutrients in SGD (Knee et al., 2010; Young et al., 2015; Dulai et al., 2023). Many studies of recent and Holocene coral reef development show that corals have thrived in areas exposed to sustained SGD for thousands of years prior to poor land-use activities (Ribaud-Laurenti et al., 2001; Grossman and Fletcher, 2004; Fletcher et al., 2008).
The extent of influence of groundwater and changes in groundwater quality and flux to the nearshore of the State of Hawaii due to climate and land-use change has been a focus of many studies. Thermal infrared imagery from fixed-wing airplanes along the coast spanning the 1990s to 2008 demonstrated that cooler, SGD-derived water plumes were persistent over that timespan (Johnson et al., 2008; Peterson et al., 2009; Asner et al., 2024). Detailed radon, radium, nutrient, and other tracer studies, including salinity, indicated the flux of SGD through these seeps ranged from 500 to 12,000 m3/day (Knee et al., 2010; Johnson et al., 2008; Peterson et al., 2009), comparable with SGD along many other coasts (Garrison et al., 2003; Street et al., 2008; Dimova et al., 2012) and equivalent in size to small to moderate streams. Comparisons between studies and repeat analyses at individual sites suggest that SGD fluxes may vary through time in response to land use (Knee et al., 2010), prolonging the drought-driven decrease in baseflow (Oki, 2004). Although data quantifying the temperature of groundwater entering the nearshore are scarce, monitoring at Kaloko-Honokōhau near Kailua-Kona on the Island of Hawai’i demonstrated that groundwater is substantially colder than marine waters, uniformly fluctuating by less than 0.5°C around a mean of ~19°C (United States Geological Survey, 2024). Groundwater modeling indicated that groundwater well development since 1975 had the effect of decreasing the flux of SGD along the Kaloko-Honokōhau and Kailua-Kona coastal area by 50% (Oki, 1999). This could reduce the potential beneficial effects of SGD to cool the nearshore and, at the same time, intensify the negative impacts of nutrients by concentrating their delivery to coastal waters. Although groundwater fluxes along the western shoreline of the Island of Hawai’i have been well characterized, the extent to which changes in these fluxes through time affect coral health and resilience is still unclear.
Until about 1997, coral health in the State of Hawaii was not a concern and bleaching was relatively infrequent and localized (Rodgers et al., 2016). Hawaiian corals were thought to be protected from extreme thermal stress, in part, because of the island chain’s high latitude (Jokiel and Coles, 1990). This century, however, rising sea-surface temperatures, growing accounts of worldwide coral mortality, and degradation observed across the main Hawaiian Islands have led to increased concern over coral health, particularly because Hawaiian corals have a lower temperature bleaching threshold (between 26 °C and 27°C) than corals at lower latitudes (Jokiel and Coles, 1990; Jokiel and Brown, 2004). Regional assessments of nearshore coral health have found Hawaiian corals increasingly displaying a variety of compromised health indicators related to terrestrial runoff and land use (Couch et al., 2014; Knee et al., 2010; Street et al., 2008), which are modulated and/or exacerbated by changes in water runoff, temperature, and quality associated with SGD (Prouty et al., 2017; Swarzenski et al., 2017; Grossman et al., 2010; 2021; Gove et al., 2023; Johnson et al., 2008; Peterson et al., 2009). The role and extent that SGD mitigates or aggravates temperature stress and any additional interacting stressors to coral health remain poorly known. For example, although the prevalence of various diseases was observed to increase with temperature and was found up to 2.0 times higher when mean water temperatures were ~1°C higher (Couch et al., 2014), the degree to which SGD contributed to bleaching or interacting stressors affecting disease remains uncertain. The 2014–2015 extreme marine heat wave (MHW) led to high spatial variability in bleaching and subsequent recovery (Gove et al., 2023), indicating that conditions modulating temperature stress are nuanced and complex.
The extent to which SGD influences nearshore temperatures and thermal stress experienced by corals is poorly understood due to the challenges of measuring and modeling water temperatures at fine scales in the nearshore environment and the complexity of factors contributing to coral health. Remote sensing approaches, such as thermal infrared imaging, can distinguish the presence and extent of SGD plumes emanating from the shore; however, they can only resolve the uppermost few millimeters of the thermal structure without additional modeling or time-series analysis (Johnson et al., 2008; Peterson et al., 2009; Oberle et al., 2022). In-situ measurements of water temperature and salinity are fundamental for validating inferences from remote sensing and for quantifying conditions affecting coral growth, although these data are often limited due to logistical challenges, cost and effort required, and sensor damage (Hendee et al., 2020). Whereas the global extent to which SGD affects coral reefs is uncertain, more than ~56% of the world’s delivery of fresh SGD to the coast occurs in the tropics and high-relief, tectonically active margins like the Caribbean and Indo-Pacific (Zhou et al., 2019) where corals prevail and serve as important sources of coral genetics (Veron et al., 2009). Here, we report on a comprehensive suite of water property measurements collected along and across nearshore instrument arrays that show the extent and frequency to which SGD reduces ambient marine water temperatures and the associated decrease in coral exposure to heat stress known to cause bleaching. Improved understanding of SGD temperature, salinity, and nutrient interactions across complex nearshore ecosystems can therefore help assess coral resilience and fate given climate and land use projections (Klein et al., 2024) and the extent to which subtle gradations in temperature and nutrient affects coral health.
2 Materials and methods
2.1 Study area
This study examined nearshore water properties along the western shoreline of the Island of Hawai’i, USA (Figure 1), where coral reef systems in the main Hawaiian Islands are threatened by growing urban development (Friedlander et al., 2018) and SGD creates a mosaic of variable habitat with respect to effects on temperature and nutrients (Grossman et al., 2021; Johnson et al., 2008; Knee et al., 2010). This region is characterized by recent to late Pleistocene volcanics (Sherrod et al., 2021), low to moderate wave exposure, and a microtidal regime (Storlazzi and Presto, 2005; Grossman et al., 2010). The coast is arid, receiving between 250 and 500 mm of rainfall each year, and surface water runoff is largely non-existent except during intense rainfall events (Giambelluca et al., 2013). Estimates of sustainable groundwater yield from mapped aquifers vary by an order of magnitude and are higher in the south (Department of Land and Natural Resources, Commission on Water Resource Management, 2022). Thermal infrared imaging and radiochemistry have revealed numerous groundwater seeps alongshore (Johnson et al., 2008; Peterson et al., 2009) with estimated fluxes of SGD to the nearshore ranging 500–12,000 m3/s (Peterson et al., 2009; Knee et al., 2010) of relatively old-age water sourced from recharge at high elevation on the volcanic slopes (Fackrell et al., 2020). Marine electrical-resistivity surveys indicate discharge of fresh groundwater along the submerged flanks of the island through complex subterranean aquifers and fractures (Attias et al., 2020) similar to the many modest sized (e.g., 2–5-m-diameter) lava tubes found below mean sea level along the western coast of the Island of Hawai’i (e.g., at Kaloko-Honokōhau Harbor, Kaloko, and Kealakekua Bays).
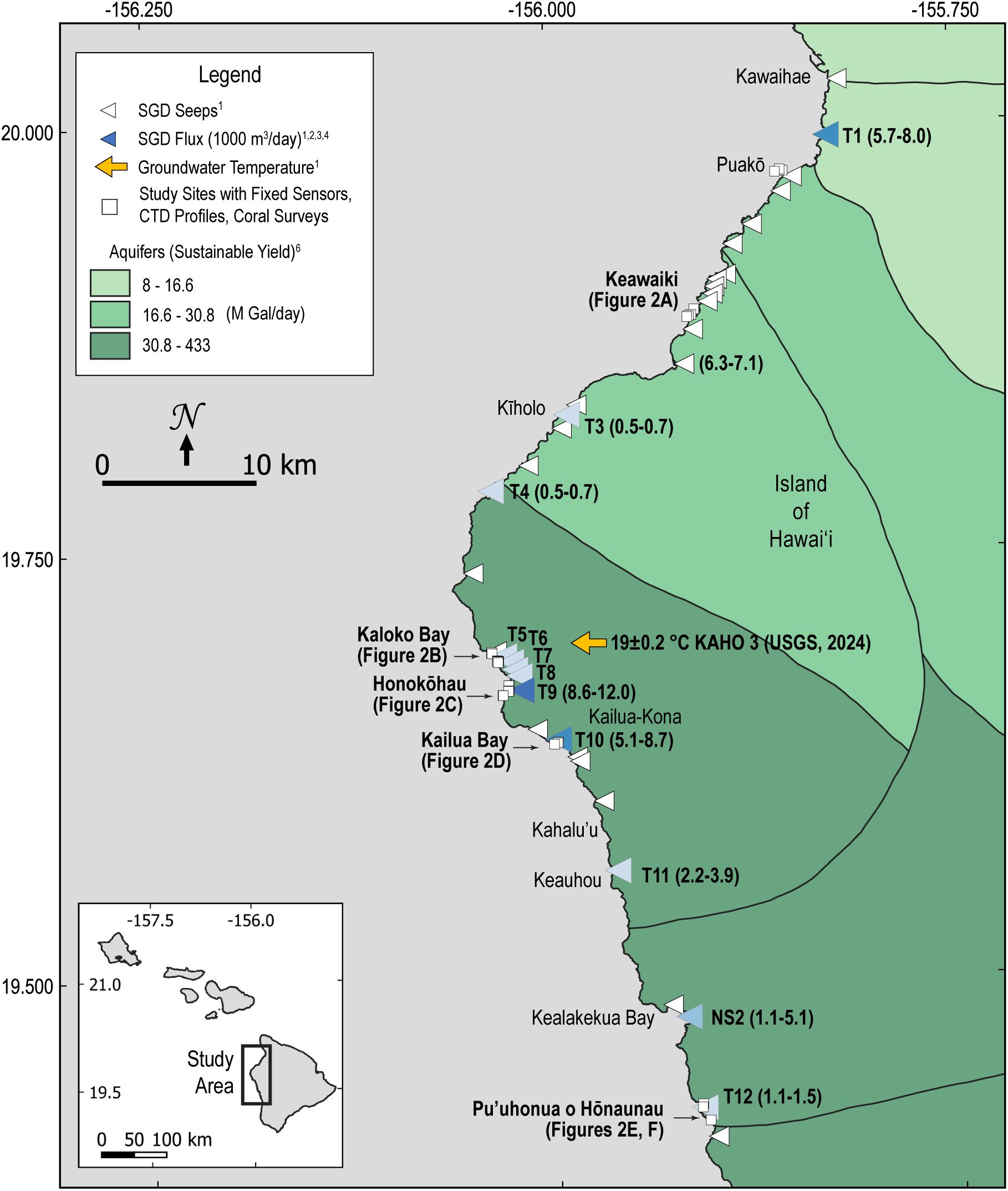
Figure 1. Map showing locations of mapped groundwater seeps (white triangles), estimated fluxes of submarine groundwater discharge (blue triangles), groundwater temperature (orange arrow), and study sites (white squares) with reference to detailed sample maps in Figure 2 highlighted in bold text. 1Johnson et al., 2008,2Peterson et al., 2009, 3Knee et al., 2010, 4Grossman et al., 2010, 5United States Geological Survey, 2024, 6Department of Land and Natural Resources, Commission on Water Resource Management, 2022.
Paired water property measurements and benthic surveys of coral health metrics were made at 31 stations in 12 study areas along ~100 km of the western shoreline of the Island of Hawai’i between Kawaihae and Pu’uhonua o Hōnaunau (Figure 2) over the period 2004-2014. Time series of water temperature, salinity, and water level were made at fixed locations, and discrete vertical profiles and surface tows were conducted to characterize variability more expansively over a range of seasonal, tidal, and weather conditions. Benthic surveys were conducted by SCUBA along predefined, randomly placed transects located within ~10 m of the fixed oceanographic sensors. Together, these data make it possible to evaluate the extent and frequency to which SGD affects nearshore temperatures and water quality important to coral health.
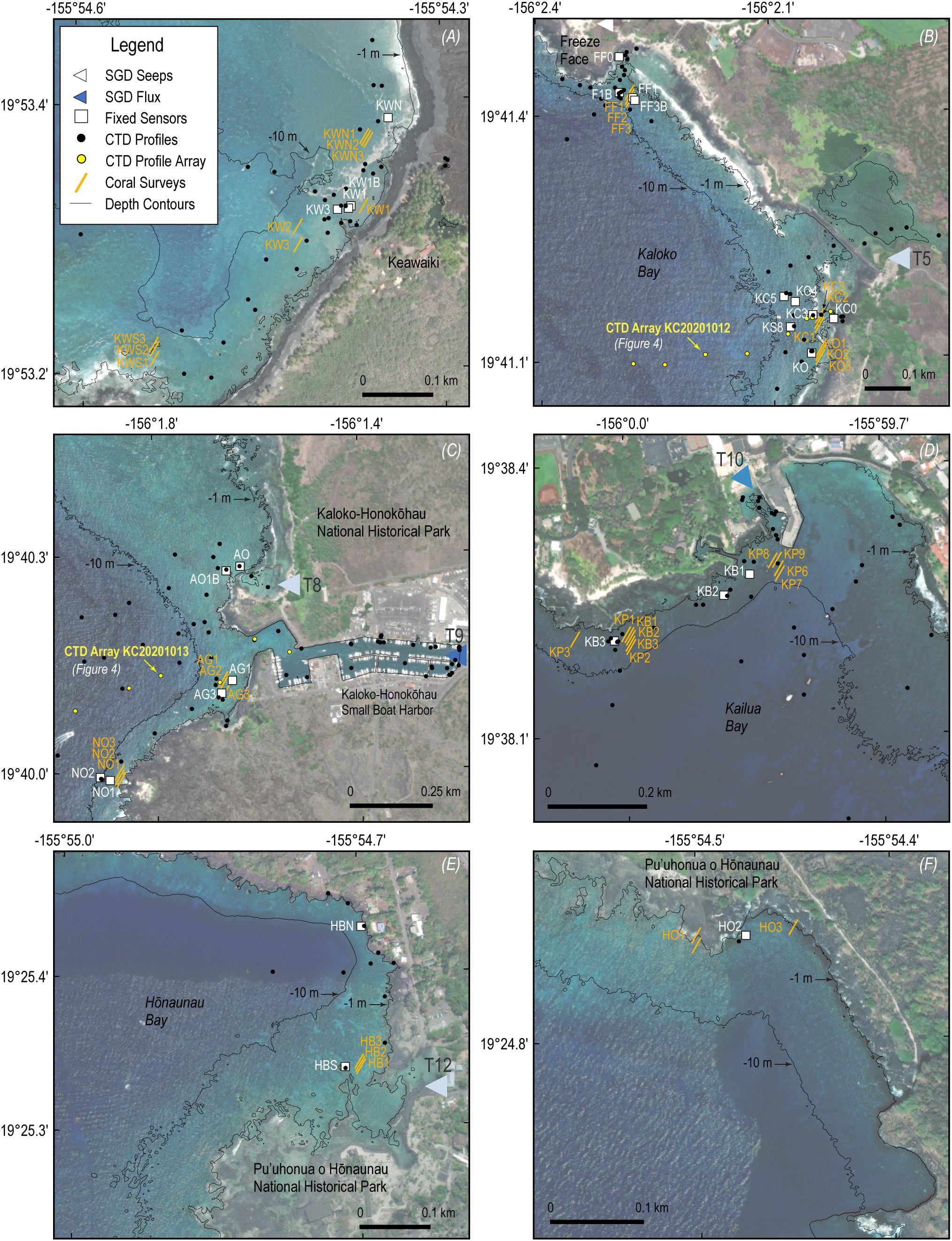
Figure 2. Maps of sampling locations including fixed sensors (white squares), CTD profiles (black circles), featured CTD profile arrays (yellow circles), and benthic surveys of coral cover and bleaching (gold lines) at Keawaiki (A), Kaloko Bay (B), Kaloko- Honokōhau Harbor entrance (C), Kailua Bay (D), Hōnaunau Bay (E), and outer Hōnaunau (F).
2.2 Fixed sensor deployments of water properties
Hobo TempV2 thermistors were attached to the benthos on stable substrates (dead coral or basalt) within 0.5 m of live coral—typically 3–5 cm above the seafloor. Within each study area, one station also included a Van Essen CTD-Diver that measured conductivity, temperature, and depth (CTD) to gather time-series fluctuations in salinity and water level. Sensors recorded data at 10-min intervals from June 2010 to December 2013 and were serviced every 3–4 months to correct sensor drift, remove biofouling, and replace batteries. Isolated data loss occurred at a few stations due to sensor damage associated with large wave events or biofouling, but sufficient repetition resulted in near continuous data at each study site to characterize tidal, daily, seasonal, and interannual variability in water properties.
2.3 Spatial measurements of salinity–temperature–depth profiles
Discrete salinity–temperature–depth profile measurements were collected using a Sontek CastAway profiler in gridded alongshore and across-shore arrays at study sites at 5 Hz from ~10-cm below the surface to the seafloor to characterize spatial variability around the fixed deployment sites. The location of the start and end of each cast was recorded with GPS, and the profile data were processed to characterize down- versus up-casts after averaging values into 0.25-m depth bins. Spatial surface tow data were gathered with a YSI 6920V2 multiparameter probe at ~0.5-m depth to resolve spatial gradients in surface water temperature, salinity, pH, dissolved oxygen, and turbidity. Measurements and GPS positions were both recorded at 1 Hz. Under collection speeds of ~2 m/s, these data recorded water properties every 4–5 m along-track. Tow and profile data from study areas were collected within 1-2 h to provide a snapshot of conditions during specific tidal, diurnal, wind, and seasonal states.
2.4 Benthic surveys of coral health
Coral health data were gathered by line-transect mapping methods using SCUBA between 2010 and 2014 following (Rogers and Miller, 2013; Couch et al., 2014). Three randomly placed 10-m transects were installed in each study area with pins fixed to the seafloor at the start and end of the survey line and located with GPS. Surveys were conducted along a survey tape temporarily placed between the pins in June 2010 for baseline conditions and repeated in October 2010 and November 2011 following peak annual late summer-early autumn peak temperatures. Photoquadrats corrected for white balance at each site were collected every 0.5 m along the survey tape set 0.6 m above the bottom on each side of the transect for a 2-m swath of continuous imagery. Divers also recorded in situ coral cover, species, number of colonies, other substrates and presence, and counts and size of lesions associated with coral bleaching along with other adverse health indicators described in Couch et al. (2014). These data were recorded across a 1 m × 10 m swath centered along the tape following Couch et al. (2014) and included the surface area and severity of bleaching (partial or total) where it occurred. A minimum lesion size of 10 cm was used to map and count coral bleaching or other tissue impairment. Additional details of the benthic surveys are provided by Grossman et al. (2019; 2021).
2.5 Data processing and analyses
All sensors were calibrated and checked for consistency both individually and across instruments. Uncertainty in temperature following factory specifications and repeat ice bath tests was ±0.1–0.2°C for the Hobo Temp V2 sensors and ±0.1°C for Van Essen CTD-Divers used for the fixed deployments. Temperatures were accurate to within ±0.5°C for depth profiles measured by the Sontek CastAway profiler and ±0.15°C for tows using the YSI 6920V2 multiparameter probes. Fixed sensor, depth profile, and tow salinities were deemed accurate to ±0.1 PSU determined after a two-point calibration. Water depths of fixed sensors were resolved to within ±0.5 cm, and those for casts were ±1.0 cm. Fixed deployment data were interpolated onto a common time series across all stations which occasionally required filling 1-3 h of missing data by linear interpolation. Common statistics, spectral power analysis, and station differences from area mean and maximum statistics were computed using MathWorks MATLAB at each time step. Depth profile and tow data were similarly compiled, and quality was assessed in MATLAB occasionally treating missing or spurious data (e.g., out of water) by filling with NaN (not a number) or interpolating between quality-controlled, environmentally consistent values. Spatial patterns were analyzed through standard gridding, kriging, and mapping approaches using MATLAB and geographic information systems including QGIS. Exceedances were determined by differencing observed temperature and published temperature thresholds of heat stress to corals (Jokiel and Coles, 1990; Coles et al., 2018) at each time step and computing the time above each threshold as a fraction of total time sampled.
Benthic photoquadrats were analyzed for counts and percentages of live coral cover, live coral cover by species, bleached coral, algae cover, and other substrate details using the National Coral Reef Initiative CPCe software (Kohler and Gill, 2006). Measurements were made at 45 randomly placed locations in each photo, which was shown to be sufficient to quantify total coral cover with low standard error (Marrack et al., 2014). CPCe data and coral health metrics were assembled for each transect and general statistics were derived by area. All water property data and metadata are provided in Grossman and Marrack (2019), and all benthic survey data are provided in Grossman et al. (2019; 2021).
Data characterizing coral response to the 2014–2015 MHW described in Gove et al. (2023) were analyzed to assess potential relationships with mapped groundwater seeps and fluxes (Figure 1) and temperature patterns (this study, Grossman and Marrack, 2019; Grossman et al., 2021). Percent coral loss was computed for each site as the difference in cover between 2016 and 2014 or 2015 where 2014 data was not available divided by the initial cover value. Percent coral recovery was defined the change from the minimum found after the 2014–2015 MHW (e.g., computed as loss) and the maximum surveyed through 2018 before the compounding effects of the 2019 MHW divided by the initial minimum cover value.
3 Results
3.1 Nearshore temperature and salinity
Measurements from seabed-mounted sensors in Kaloko Bay over three summers between June 2010 and September 2012 demonstrated seasonal variations in nearshore temperature that ranged between ~24°C in winter to >27°C in late summer/early fall (Figure 3). Comparable ~1 °C–2°C temperature oscillations also occur on daily and semidiurnal timescales. Semidiurnal temperature oscillations were marked by salinity variations of 1–4 PSU with minimum temperatures and salinity occurring daily at or immediately after low tide (Figures 3B–D). Greater variability with lower temperature and salinity was observed during spring tides. Temperatures in Kaloko Bay were 1 °C–5°C lower, with larger temperature excursions closer to identified SGD seeps through time.
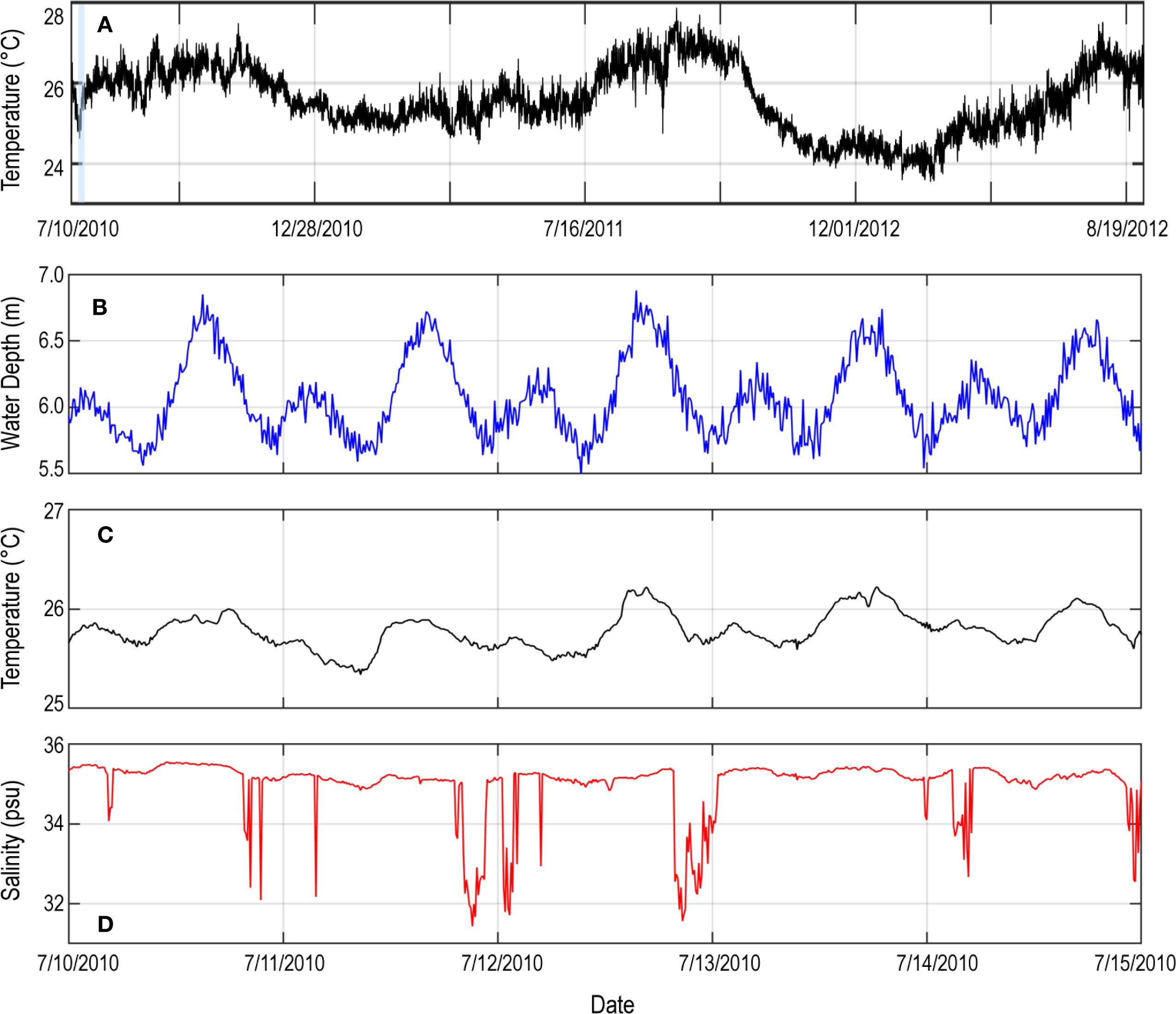
Figure 3. Time-series plots of nearshore water temperature for summers 2010, 2011, and 2012 (A) and corresponding water depth (B), temperature (C), and salinity (D) for the period 10 Jul 2010 through 15 Jul 2010 (blue bar in (A)) at site KO located at -4 m (MSL) in Porites lobata coral habitat, showing decreases in temperature reaching 0.75°C and salinity of ~4 PSU in response to daily ebb tides.
Depth profiles of temperature and salinity demonstrated similar patterns of generally stratified, colder, lower salinity waters closer to shore and closer to SGD seeps (Figure 4). Some areas had warmer and more saline waters offshore or in the middle of the water column associated with a marine source (Figure 4F). Depth profile and surface tow surveys collected within 1- to 2-h periods corroborated fixed-sensor results that lower temperature and salinity surface plumes extended farther offshore at and shortly following low tides than during high tide when nearshore waters were stratified. The lower temperature and salinity surface plumes were observed multiple times over each of the seasons surveyed and during low to moderate (<5 m/s) winds, demonstrating persistence despite mixing.
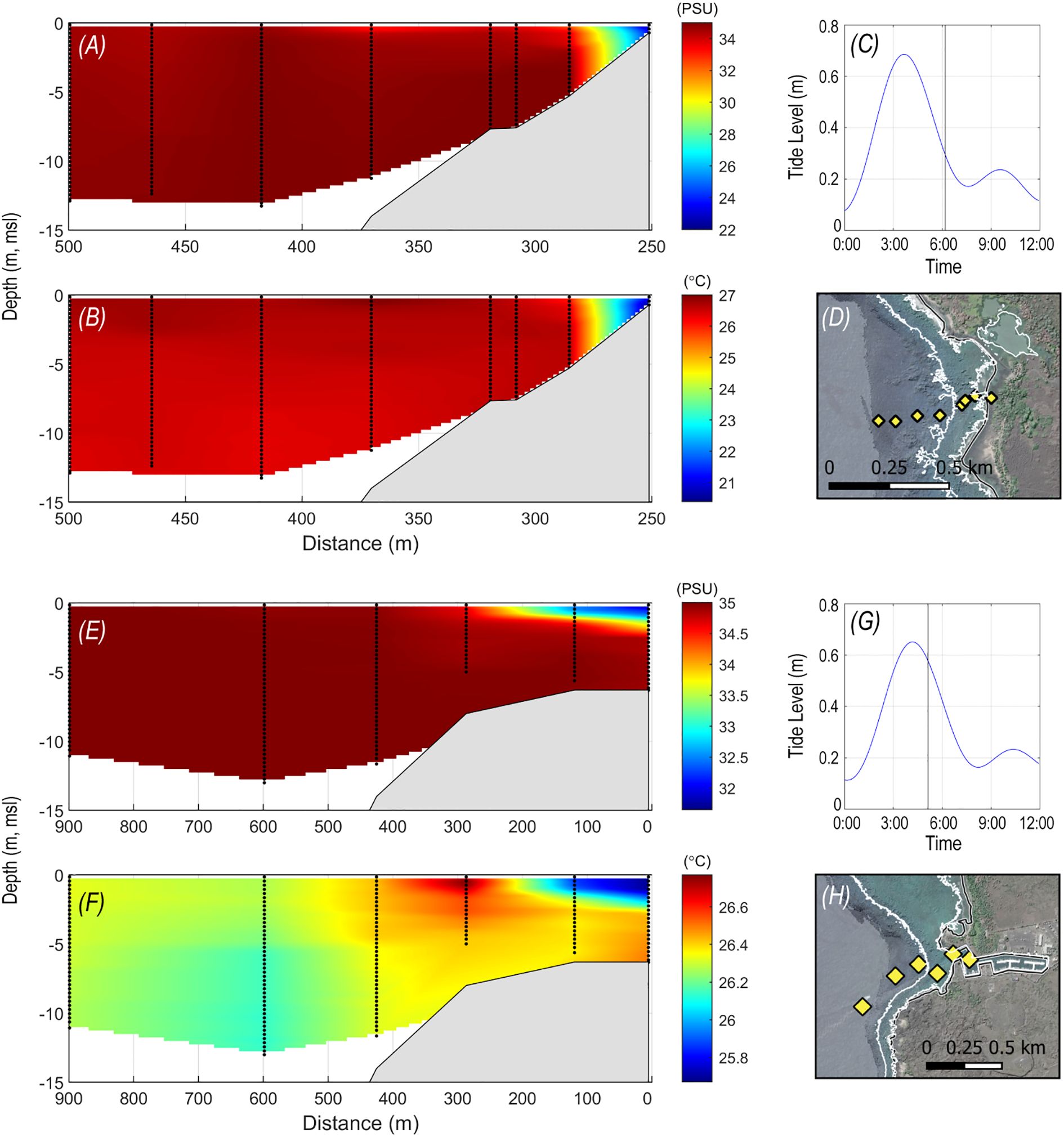
Figure 4. Plots of salinity (A) and temperature (B) depth profiles during ebb of 12 October 2010 (C) extending offshore of Kaloko Bay (D) and salinity (E) and temperature (F) depth profiles during early ebb on 13 October 2010 (G) offshore of Kaloko-Honokōhau Harbor (H) showing water that bathes extensive areas of coral reef in 3–5-m depth with lower salinity and 5°C–6°C lower temperatures than offshore marine water. Note varying color bar scale in (A, B) and (E, F).
3.2 Influence of submarine groundwater discharge
Measurements from fixed sensors, CTD casts, and tows indicate that SGD reduced water temperatures and salinity of ambient marine water at all the sites studied. A general decrease in temperature and salinity was observed with proximity to shore (Figure 5A). Surface salinities were systematically lower during the wetter winter and spring months relative to drier summers and late fall periods. CTD profiles reveal that SGD effects extend to ~7–8 m depth (Figure 4B), which can occur as far as 500 m offshore based on survey transects (Figures 1B). Across these areas, SGD reduces nearshore temperatures by 1–5°C and salinities by 5–18 PSU (Figure 5B) with reduced temperatures of 1°C or more extending 200–300 m offshore. CTD profile data demonstrated that water of similar density and salinity can vary up to 4 °C–6°C in temperature (Figure 5C). Whereas much of this temperature depression was associated with stratified surface conditions, repeated surveys indicated a persistent pattern of reduced water temperatures toward the shoreline at all depths relative to offshore marine water temperatures (Figure 4).
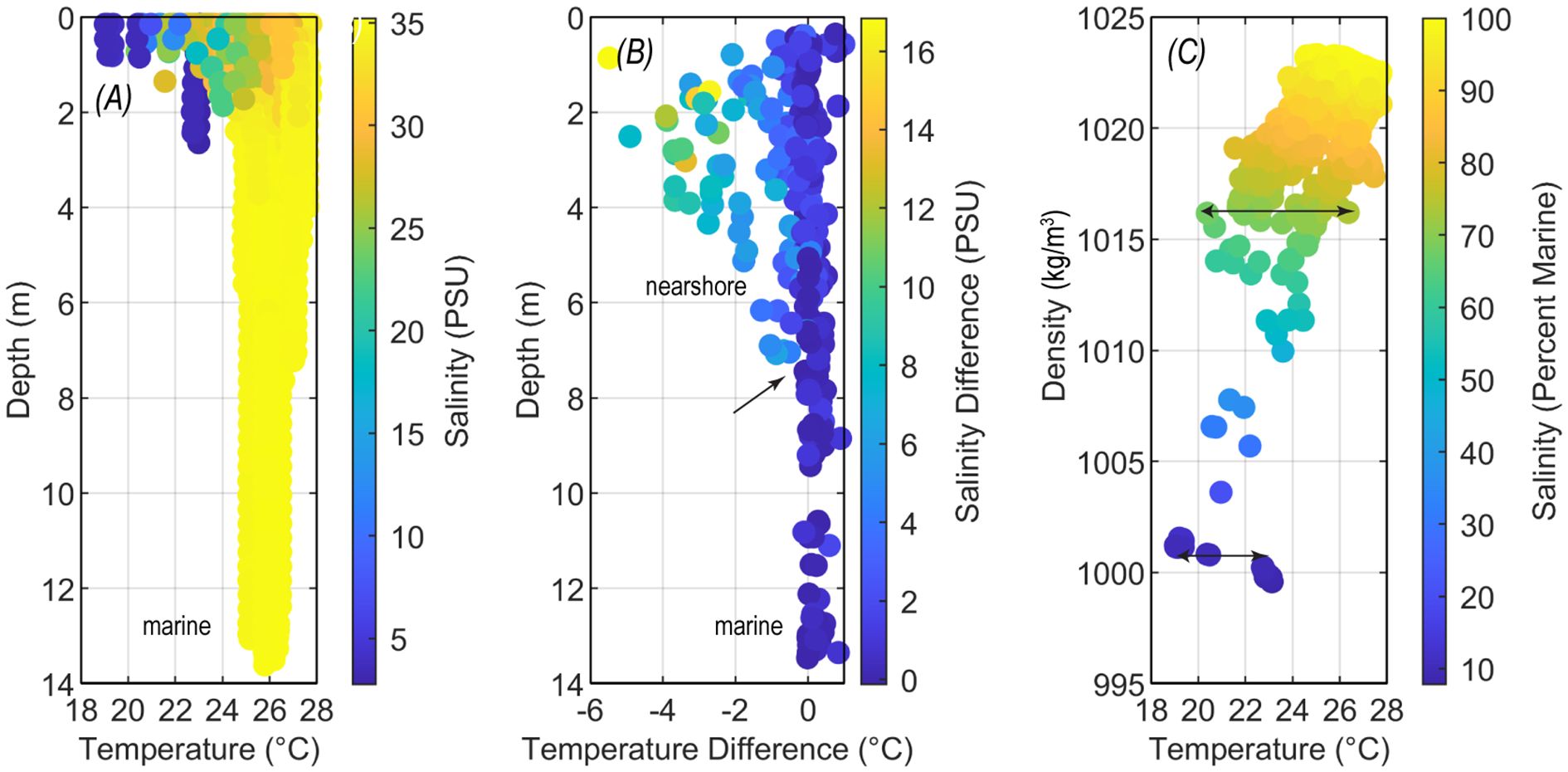
Figure 5. Plots of CTD profiles across the western coast of the Island of Hawai’i between 2010 and 2013 showing lower water temperatures near the surface and with lower salinity (A), the difference between top and bottom water reflecting stratified low temperature and salinity to ~7-m depth [arrow in (B)], and variability in temperature for discrete density and salinity that ranges 4°C–6°C [arrows in (C)].
Time-series measurements across Kaloko Bay spanning depths of 0.3 to 8.0 m and distances up to 300 m from mapped groundwater seeps indicate the extent to which SGD reduces temperatures at each location. The difference in temperature at stations KC3 (100 m from seep) and KO (250 m from seep) from the corresponding instantaneous area mean and maximum demonstrate that site-specific conditions largely structured by SGD and its mixing reduce ambient marine water temperatures across tidal to seasonal timescales (Figure 6). Area mean temperatures during the study were reduced by 0.25 °C–1.00°C (black line), and maximum area temperatures were depressed by up to 1.50°C (orange line) and characteristic of all Kaloko Bay sites. Slightly greater temperature depression was observed at stations closer to shore, although variations in overall range at stations such as KC4 and KS8 indicate the importance of circulation and potentially other processes (localized heating) in modulating the effects of SGD on nearshore water temperatures.
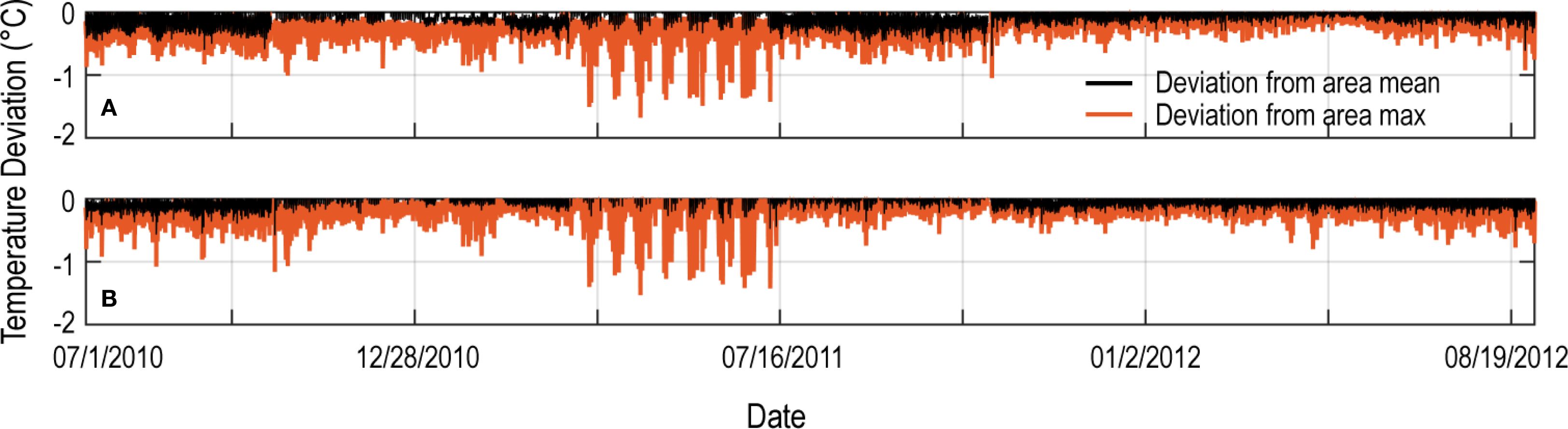
Figure 6. Time-series plot showing the extent that water temperatures at site KC3 [(A) water depth 4 m, 100 m from groundwater seep] and KO [(B) water depth 4 m, 250 m from groundwater seep] were depressed below the mean (black) and maximum (orange) marine water temperatures between 1 July 2010 and 1 September 2012 observed in Kaloko Bay (locations shown in Figure 2B).
3.3 Temperature threshold exceedances
Exceedances of 26.0 °C, 26.5 °C, and 27.0°C temperature thresholds computed as a percentage of the time recorded at each fixed sensor indicate that nearshore temperatures were frequently reduced (Figure 7). Exceedance of 26°C for example in Kaloko Bay ranged between 49% offshore (site KS8) to as little as ~3% close to shore (site KC0) over the 3.5-year study. Exceedances of 26.5°C and 27.0°C occurred 2%–27% and ~1%–7% of the time, respectively. The difference in exceedances between offshore sites and sites closer to shore (Figure 7) is principally driven by increasing exposure to SGD close to shore. Furthermore, SGD compensated for the additional heating expected from solar insolation in shallower nearshore waters and, in particular, across areas of uniquely black (low albedo) volcanic substrate susceptible to high heat absorption. Even so, across the 500-m reef, marine water temperatures of 26°C were reduced by SGD ~46% of the time relative to offshore sites; temperatures of 26.5 °C and 27.0°C were depressed by SGD ~25% and ~5% of the time, respectively (Figure 7).
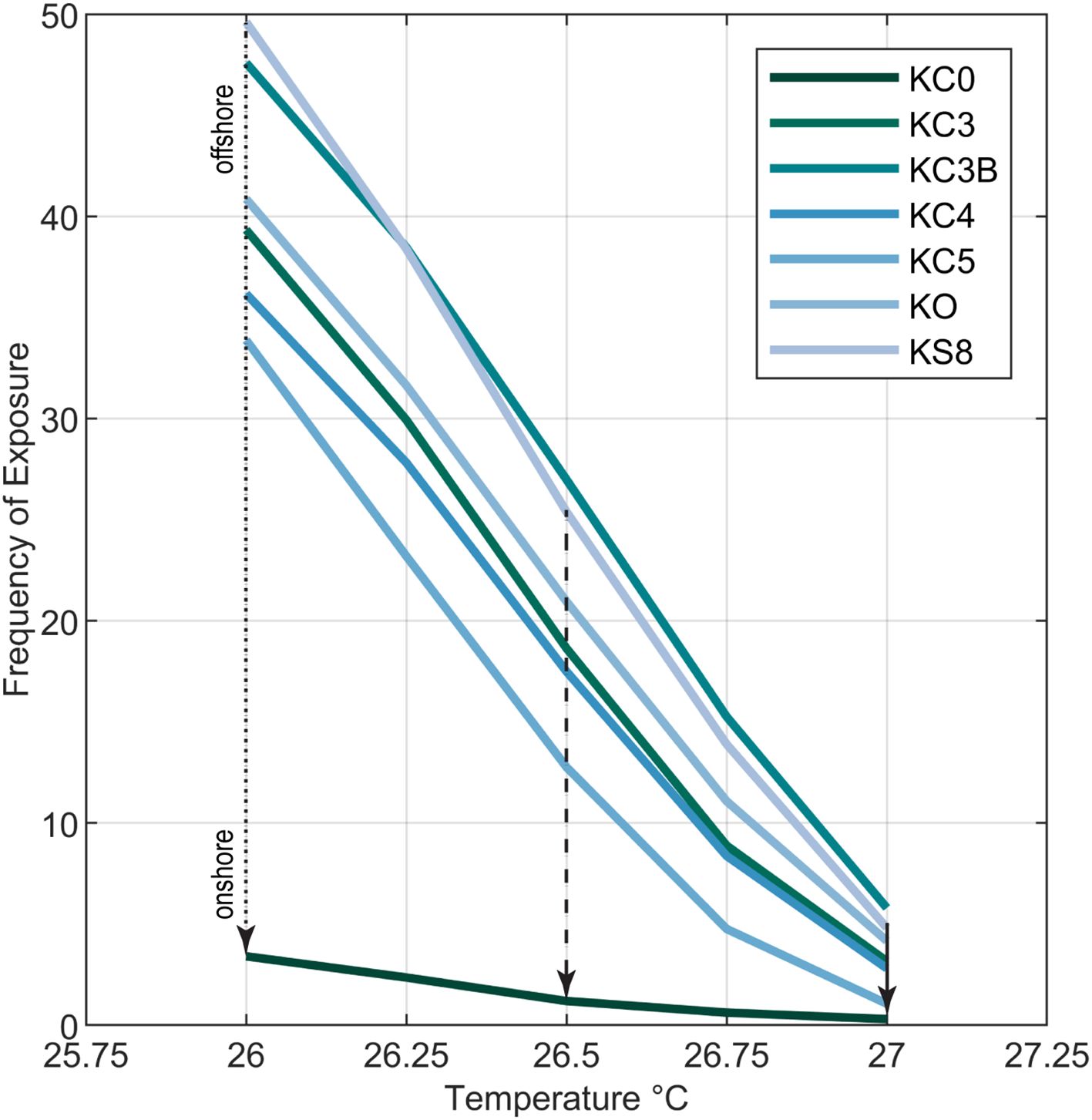
Figure 7. Plot of the frequency that water temperatures at sites in Kaloko Bay exceeded thresholds that induce thermal stress (dotted, dashed, and solid arrows for 26.0°C, 26.5°C, and 27.0°C, respectively) showing that exposure is steadily reduced by SGD closer to shore (between sites) and up to 47% of the time relative to offshore (dotted arrow).
3.4 Coral bleaching and health
Coral bleaching was observed at eight of our study areas along the western shoreline of the Island of Hawai’i in response to the moderate heat stress measured in 2010 and 2011 (Figure 8). The number of bleached coral colonies in any one area ranged from 1 to 15, with individual survey transects generally having one or two colonies observed. The maximum number of colonies bleached was 12 on one triplicate transect at outer Hōnaunau (HO). In 2010, bleaching was comparable among areas with variable SGD influence like Keawaiki (KW) relative to Keawaiki North (KWN) or Keawaiki South (KWS) and inner Kaloko (KC) relative to outer Kaloko (KO). Bleaching on individual transects in each area of KW, FF, KC, AG, and KP was higher on transects farther offshore or farther from SGD seeps (e.g., KW2 vs. KW1, FF2 vs. FF1, KC2 vs. KC1, AG3 vs. AG1, and KP7 vs. KP8, Figure 2). In 2011, a greater number of bleached coral colonies were found in areas offshore or farther from SGD seeps, particularly outer Kaloko (KO) relative to inner Kaloko (KC) and especially outer Hōnaunau (HO) relative to inner Hōnaunau Bay (HB).We note that some sites were not surveyed in 2011 and results from steeply sloping areas like Freeze Face (FF) and Noio Point (NO) may be less comparable with other areas because available substrate to survey only existed at greater depth. Even so, these results indicate that bleaching was lower closer to SGD seeps and inside small embayments than offshore or at greater distance from SGD seeps.
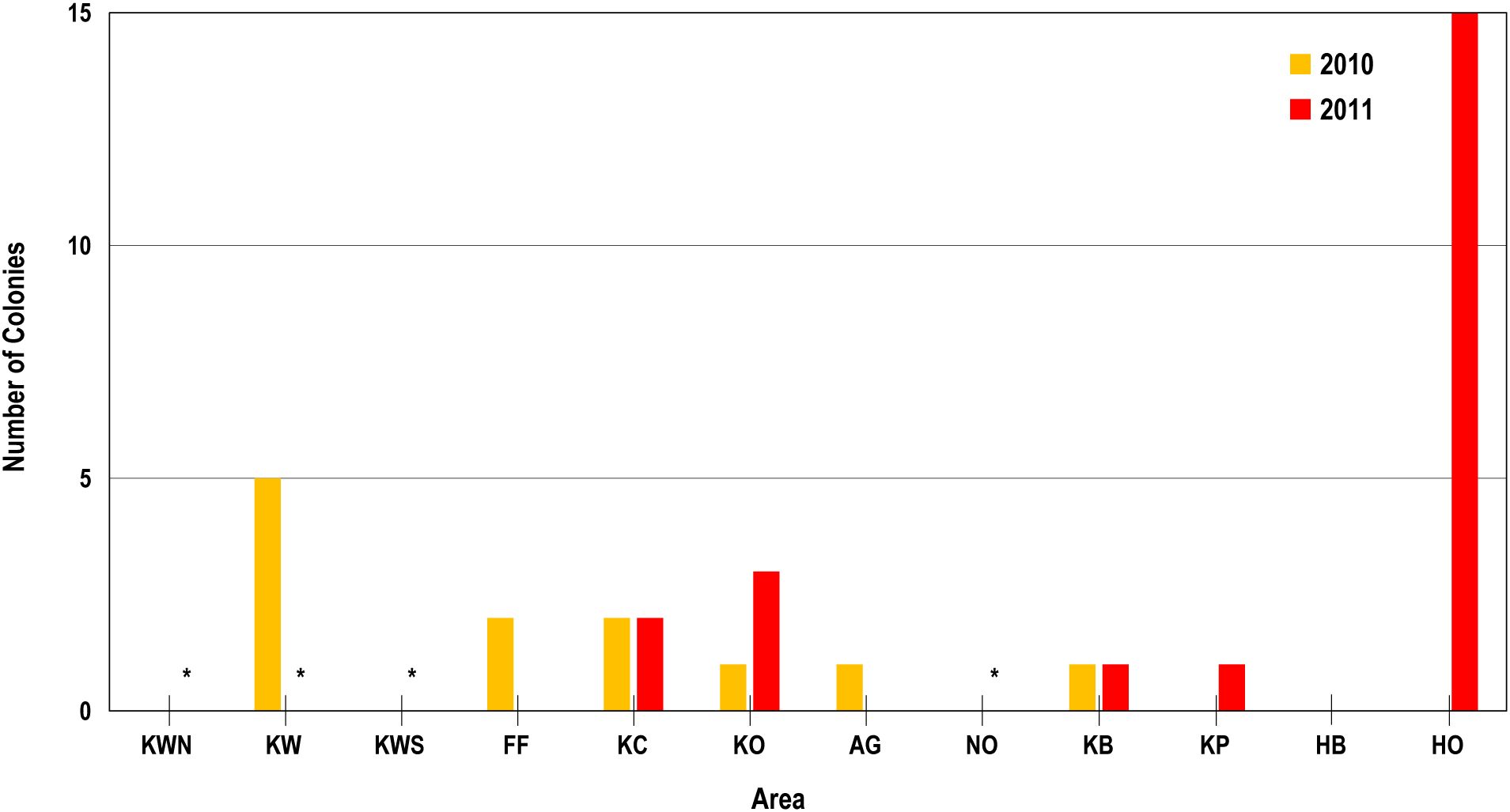
Figure 8. Bar plot of number of bleached coral colonies in each area in October 2010 (orange) and November 2011 (red) showing generally less bleaching at sites KC, KP, and HB that were closer to cold SGD seeps relative to site offshore KO, KB, and HB, respectively. Asterisks indicate site not surveyed in 2011.
Coral loss following the 2014–2015 MHW and subsequent recovery through 2018 demonstrated notable relationships to mapped groundwater seeps and colder nearshore temperatures where flux and influence of SGD have been observed to be high (Figure 9). More than 25% of the sites with the lowest coral loss occurred offshore of the densely mapped seeps north of the Kailua-Kona Airport, including several sites like R-19, R-20, and R-6 at Keawaiki, where we measured temperature depression by SGD (Figure 9A), as well as R-23, R-24 (near Kīholo) and R-11 (Puakō) known for high influence of SGD (Dimova et al., 2012; Aguiar et al., 2023). Coral loss was also lower at sites Fixed-2 offshore of Freeze Face, Fixed-5 immediately offshore of Kaloko, and Site 11 offshore of the Kaloko-Honokōhau Harbor, although nearby sites also had higher loss (Figure 9B). Coral recovery was generally higher along shorelines with presumed high SGD flux like Kawaihae (sites R-03, R05) and/or high measured fluxes like R-08, R-10 near T1, and R-24 at Kīholo (Figure 9A), Fixed-4 near T5, R-37, Fixed-11 and Fixed-12 offshore of T9 (Figure 9B).
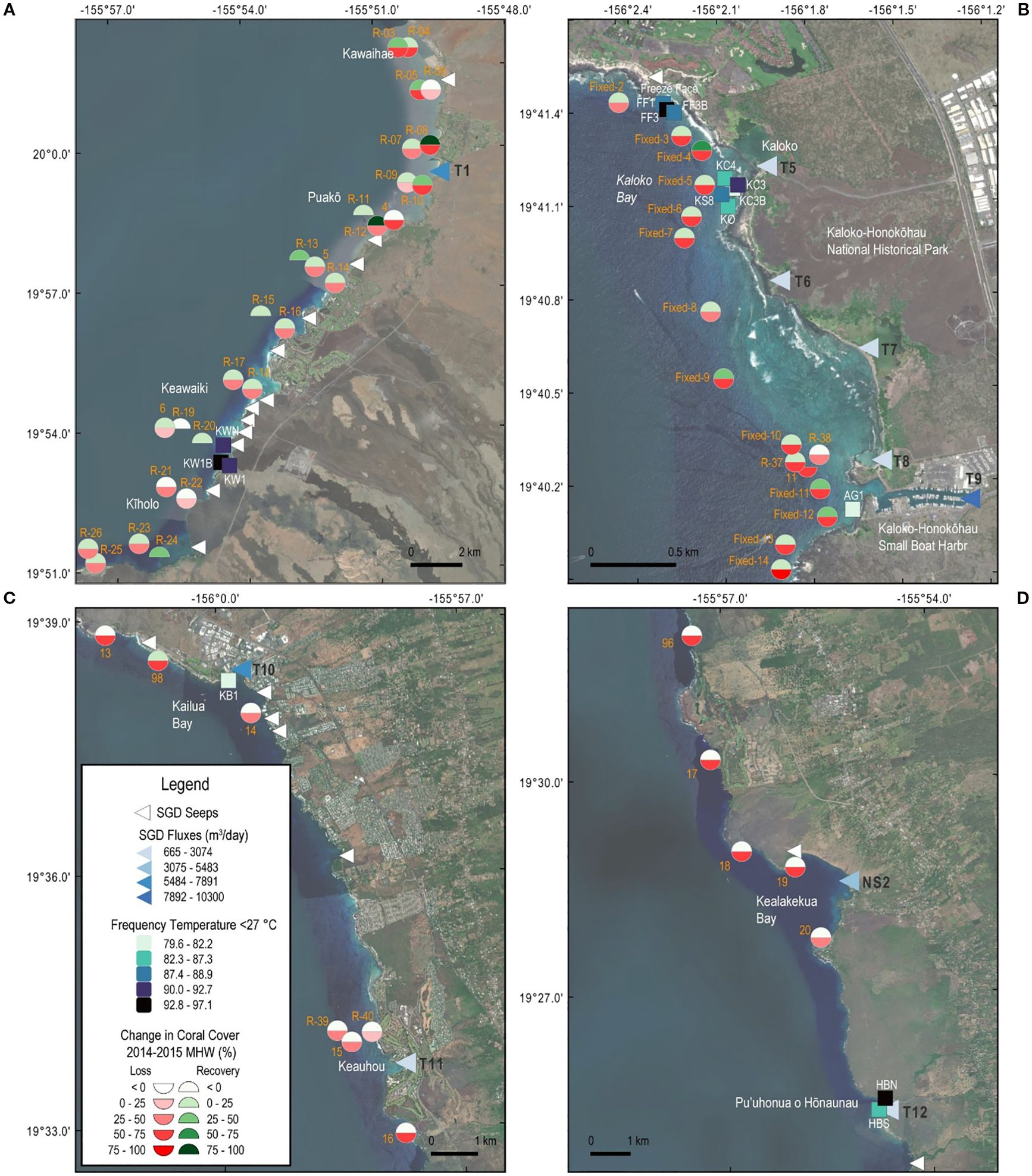
Figure 9. Maps showing generally lower coral bleaching and loss (red semi-circles) and higher recovery (green semi-circles) following the 2014–2015 marine heat wave (after Gove et al., 2023) close to SGD seeps, especially seeps with higher flux (darker blue triangles), and more frequent temperatures below 27 °C driven by SGD (darker blue squares) including R-06 (Kawaihae), R-11 and R-12 (Puakō), R-20, 6, R-22 (Keawaiki), and R-23 and R-24 (Kīholo) (A), Fixed-3 (Freeze Face), Fixed-4 and Fixed-5 (Kaloko), and R-38, Fixed-11, Fixed-12 (Kaloko-Honokōhau Harbor entrance) (B), 98 and 14 (Kailua) and R-40 (Keauhou) (C), and 20 (Kealakekua Bay) (D).
At most sites where multiple coral change measurements were available, results demonstrated high variability across fine spatial scales. In general, coral loss was lower at sites located closer to shore compared with paired sites farther offshore. For example, nearshore sites showed less coral loss than offshore counterparts at R-06 vs. R-05 in Kawaihae, R-14 vs. 5 south of Puakō, R-18 vs. R-17 and R-22 vs. R-21 surrounding Keawaiki, and R-23 and R-25 vs. R-26 near Kīholo (Figure 9A), R-38 vs. R-37 and Fixed-10 vs. Fixed-11 just north of the Kaloko-Honokōhau Harbor entrance (Figure 9B), R-40 vs. R-39 and 15 offshore of Keauhou (Figure 9C), and 20 vs. 18 and 19 offshore of Kealakekua Bay (Figure 9D). Similarly, coral recovery was often greater at nearshore sites compared with adjacent offshore sites. This pattern was observed at R-08 vs R-07, R-10 vs. R-09 surrounding groundwater seep T1 north of Puakō, R-12 vs. R-11 in Puakō, R-16 vs. R-15 south of Puakō, R-18 vs. R-17, R-20 vs. R-19 surrounding Keawaiki, R-24 vs. R-23 in Kīholo (Figure 9A), and Fixed-4 vs. Fixed-5 and Fixed-11 and Fixed-12 relative to Fixed-10, R-37. Fixed-13 and Fixed-14, located closer to the SGD plumes emanating from Kaloko Fishpond and Kaloko-Honokōhau Small Boat Harbor, respectively (Figure 9B).
4 Discussion
4.1 Variability in nearshore temperature depression by submarine groundwater discharge
Our findings add substantial information quantifying the temporal variability and spatial scales at which SGD influences nearshore water temperatures, and particularly the frequency with which it reduces temperatures that can affect coral heat stress along ~100 km of the western shoreline of the Island of Hawai’i. The seasonal and semidiurnal patterns displayed at Kaloko (Figures 3, 6) were also observed at the 11 other areas studied along the western shoreline of the Island of Hawai’i reported by Grossman and Marrack (2019) and Grossman et al. (2021) associated with SGD influence on the nearshore (Johnson et al., 2008; Peterson et al., 2009; Grossman et al., 2010). This study demonstrates that uniformly cold (~19°C) groundwater (United States Geological Survey, 2024) seeping through the beach face and seafloor as SGD consistently reduces nearshore temperatures by 1°C–5°C comparable with previous studies (Johnson et al., 2008; Peterson et al., 2009; Grossman et al., 2010; 2021; Swarzenski et al., 2017). The extent and persistence of SGD influence across the nearshore Hawaiian coast results in discrete areas where the mean and maximum ambient marine temperatures are reduced up to 1°C–2°C (Figure 6) over timescales of days to years and are consistent with prior studies (Presto et al., 2007; Grossman et al., 2010; 2021). Although daily patterns of stratified plumes were observed in the present study, they were generally less spatially extensive and less pervasive along the seafloor than reported in the past (Presto et al., 2007; Grossman et al., 2010). Colder, stratified surface waters fed by SGD were found to reestablish within hours to days following mixing events from winds and waves and to reduce temperatures at depth during mixing (Grossman et al., 2010; Storlazzi and Presto, 2005), complementing the role of diurnal tidal pumping in SGD cooling nearshore waters.
We suspect that the extent of nearshore cooling by SGD is higher where fluxes are higher (Figure 1), such as offshore of Kealakekua Bay and Kaloko-Honokōhau Small Boat Harbor, where fluxes can reach 5,100 and ~120,000 m3/day, two orders of magnitude larger than at Kaloko Bay (Johnson et al., 2008; Peterson et al., 2009; Knee et al., 2010), respectively. Similar SGD fluxes and effects to nearshore plumes and temperature have been described along other coral reef-lined coasts (Garrison et al., 2003; Street et al., 2008; Knee et al., 2010; Dimova et al., 2012; Swarzenski et al., 2017; Oberle et al., 2022) and coastal environments (Taniguchi et al., 2019; Alorda-Kleinglass et al., 2021), indicating that SGD is likely important, not only globally for structuring nearshore habitats, but for nuanced temperature effects, including cold-water refugia. Although the many groundwater plumes observed along the western shoreline of the Island of Hawai’i have been observed for decades, the variability in their temperature buffering potential is uncertain and likely varies in response to changes in SGD flux, wave energy, and circulation. We note that the extent of SGD plumes observed in this study were less extensive than documented in several past studies (Johnson et al., 2008; Grossman et al., 2010; Knee et al., 2010). A concern exists that the SGD plumes, associated colder water refugia, and estuarine habitats they shape could be decreasing in extent and influence with statewide drought (Oki, 2004) and land use activities affecting groundwater supply and quality (Oki, 1999; Knee et al., 2010; Street et al., 2008; Grossman et al., 2021).
4.2 Thermal refugia and stress mitigation potential for corals
The reduction in ambient seawater temperatures up to 5°C and of more than 1°C for 5%–47% of the 4-year time period 2010–2014 across the nearshore of Kaloko Bay (Figure 7) demonstrates the importance of SGD in creating colder nearshore waters around coral reefs. Similar spatial patterns and gradients in temperatures with distance offshore and depth were found at other study areas along the western shoreline of the Island of Hawai’i (Grossman et al., 2010; 2021). The near-continuous colder nearshore temperatures year-round and specifically during summer peak warm conditions (Figure 2A) found at most, if not all, study sites (Grossman et al., 2021; 2010) demonstrate that SGD reduces near-bed water temperatures and thus exposure of corals to thermal stress (Figure 2C). SGD may be playing a greater role generally by additionally cooling shallower areas closer to shore that experience greater warming from solar insolation, thus compensating for extra heating of the water column in Hawaii and along many tropical reef-lined islands where low-albedo volcanic substrates absorb and re-radiate heat.
Nearshore temperatures observed during our 3.5-year study were stressful, inducing bleaching (Figure 8) that was moderate relative to the extreme marine heat waves of 2014–2015 (Gove et al., 2023; Perelman et al., 2024) and 2019 (Asner et al., 2022). Even so, nearshore sites with lower SGD-fed temperatures such as inner Kaloko (Figures 6, 7) and Hōnaunau Bays (Grossman and Marrack, 2019) experienced less coral bleaching during our study than sites offshore or with greater marine influence (Figure 9). Although coral bleaching and mortality were substantially more widespread during the 2014–2015 and 2019 marine heatwaves than during our study, they were generally lower at sites closer to the numerous groundwater seeps on the western part of the Island of Hawai’i (Figure 1) and associated colder SGD-driven nearshore temperatures such as Kaloko (Figure 6) and Keawaiki and inner Hōnaunau Bay (Figure 9). Greater coral recovery following the 2014–2015 MHW was generally observed at sites located closer to shore, near groundwater seeps, and in areas of high SGD flux associated with lower nearshore temperatures—for example, in Kaloko (Figure 7), Kawaihae, Puakō, Keawaiki, Kīholo, Kailua, and Hōnaunau Bays (Figure 9). These patterns indicate that, in many cases—particularly when SGD is low in nutrients and contaminants—groundwater discharge can be beneficial to coral health.
These results indicate that coral bleaching, mortality, and recovery are sensitive to fine-scale and nuanced differences in habitat conditions structured by SGD among these nearshore sites. Whereas both temperature reduction and nutrient concentrations are suspected to be greater closer to the source of SGD, lower coral mortality and higher recovery at many sites influenced by SGD indicate that either lower temperatures helped reduce heat stress and bleaching, lower temperatures compensated for nutrient stress, or adverse effects from SGD, such as elevated nutrients, were lower than previously characterized. Also, the extent to which SGD fluxes and resulting temperature depression varied during these extreme events from past estimates is uncertain and any reductions would have contributed to the extensive coral mortality documented for the 2014–2015 and 2019 marine heatwaves. Our findings also suggest that more comprehensive quantification of tidally driven SGD cooling and role in heat stress relief that can last several hours each day is important to improving understanding coral resilience (mortality and recovery), which may not be fully captured by daily average temperature metrics (Perelman et al., 2024; Asner et al., 2022).
4.3 Implications of SGD and ecological tradeoffs for coral resilience
The fine-scale spatial and temporal variability in habitat shaped by SGD may be especially important during and following prolonged heat stress events across reefs like those examined in this study, where fast-growing, pioneer coral species serve as sources of larval replenishment and genetic diversity for reef recovery following disturbance (Dollar, 1982). The degree to which these cooler conditions interact with other stressors, either exacerbating or compensating for them, such as nutrient enrichment and contaminant effects from terrestrial runoff, is not fully understood. Additionally, it remains unclear to what extent stressors that are sensitive to temperature or interact with SGD precondition coral susceptibility to bleaching and mortality (Couch et al., 2014; Grossman et al., 2021). For example, subtle and sublethal stress from grazing, lesions, growth anomalies, and algal overgrowths that have been correlated with SGD (Walsh et al., 2013; Grossman et al., 2021) and that can be modulated by temperature, salinity, and nutrients influenced by SGD (Couch et al., 2014; Klein et al., 2024) are not yet predicted sufficiently to guide coral reef resilience management. Also, recent in-situ experiments show that SGD exposure can reshape growth and community metabolism on reefs, highlighting tradeoffs alongside potential thermal benefits (Barnas et al., 2025). Therefore, improved understanding of the diverse mechanisms that SGD directly or indirectly affects coral health is important to resilience planning for coral reefs.
An additional consequence of the pronounced spatial variability in SGD documented here is the potential for microhabitat niche partitioning among reef organisms. Variations in thermal and salinity regimes across relatively short spatial scales may foster differential survival and recovery (Burn et al., 2023), enabling certain coral species and associated biota to exploit localized refugia more effectively during thermal stress events. The shallow nearshore zone studied here is increasingly affected by human activities and terrestrial runoff that modulate the flux and quality of water delivered to the coast, leading to fine-scale gradients in habitat disturbance. Future investigations could aim to elucidate how these fine‐scale gradients mediate community composition and enhance overall reef resilience in the face of rising ocean temperatures.
The tradeoffs of SGD to corals and coastal ecosystems, including possible benefits of cold-water refugia and associated reduced heat exposure relative to possible adverse impacts (e.g., elevated nutrients), will determine the net response of affected coral communities. Where elevated nutrients occur, such as noted by Oberle et al. (2019), improved understanding (Street et al., 2008; Knee et al., 2010; Hunt, 2015) is vital to managing coral reef resilience. The fact that coral reefs thrived in similar settings for thousands of years prior to urban development of the last 150 years (Ribaud-Laurenti et al., 2001; Grossman and Fletcher, 2004; Fletcher et al., 2008) demonstrates that SGD likely provided net ecosystem benefits during past marine heat waves or anomalous warm events (McKenzie et al., 2019) prior to concentrating excess nutrients and contaminants from human activities (Street et al., 2008; Knee et al., 2010; Hunt, 2015). Furthermore, improved understanding of nearshore water residence time can be used to help evaluate tradeoffs between the potential temperature benefits of colder SGD versus adverse impacts of contaminants in SGD and indirect environmental stressors including dissolved oxygen concentrations, invasive species, and grazers. Extended residence times can lead to accumulation of sediment, nutrients, pollutants, and thermal stress (Storlazzi et al., 2018) that can, in turn, reduce oxygen levels, exacerbating physiological stress to corals (Oberle et al., 2019), giving a competitive edge to invasives and grazers over corals (Couch et al., 2014).
4.4 Management and research implications
Quantifying the relative benefits and impacts of SGD as well as the cost associated with improved groundwater management is also critical to inform planning for anticipated changes to coral reef and coastal ecosystems sensitive to land-use and climate change. Rising sea surface temperatures, combined with ongoing drought and groundwater extraction for urban development, are expected to alter both the availability of SGD and its capacity to sustain “estuarine systems” that span wetlands, springs, ponds, and coastal waters, along with the ecosystem services these environments provide to people. These estuarine environments provide functioning habitat for threatened and endangered species including the Hawaiian coot and stilt, endemic shrimp, Hawaiian damselfly, rare water eel, and diverse insects and other food prey (MacKenzie and Bruland, 2012; Sakihara et al., 2015; Santos et al., 2021) that provide for commercial and subsistence fisheries and Hawaiian indigenous culture. Our results have broad implications for the potential role of SGD on coral health in light of increasing global coral mortality in response to rising ocean temperatures (Kleypas et al., 1999; Hughes et al., 2018) and nutrient runoff from poor land use (D’Angelo and Wiedenmann, 2014; Hughes et al., 2018). This benefit may be particularly important as ~56% of the world’s fresh SGD occurs in the tropics and high-relief, tectonically active margins like the Caribbean and Indo-West Pacific (Zhou et al., 2019), the latter of which is also a critical area for the coral gene pool that replenishes corals broadly following disturbance (Veron et al., 2009). Our results suggest that maintaining sufficient SGD flux to coastal waters could serve as a management strategy to support coral reef resilience, particularly in areas where runoff is decreasing due to drought or land use. This approach would be conceptually similar to the “in-stream flow requirements” established in river management to ensure adequate water for fish populations (Bradford and Heinonen, 2008).
Based on our in-situ temperature records and mapped transects showing that SGD-driven cooling varies over hours to seasons and across tens to hundreds of meters, paired abiotic–biotic studies along disturbance gradients could be used to pinpoint where and when SGD creates cooler-water refugia and to quantify how those refugia mediate physiological stress responses. For management, our in-situ observations are complemented by thermal remote sensing that maps SGD features across reefscapes—from island-wide coverage such as the western part of the Island of Hawai’i to coral-head–scale imaging in American Samoa—yielding scalable, actionable targets for site selection, monitoring, and protection (Oberle et al., 2022; Asner et al., 2024). Given our tidal-scale cooling and evidence that winds and waves restructure SGD plumes, future research and management monitoring could complement standard long-duration logging (≤1 Hz; 5–10-min archives) with targeted high-frequency thermistors at SGD hotspots to capture any short-lived, wave- and current-mixed cool excursions that our sensors likely undersampled. Quantifying nutrient concentrations and the resulting ecological effects are equally challenging (Camacho-Cruz et al., 2022; Street et al., 2008; Knee et al., 2010; D’Angelo and Wiedenmann, 2014). Integrated and long-term research and monitoring studies can help to define thresholds and variability in drivers of disturbance, as well as to track and validate the success of thresholds and metrics employed in management measures.
5 Conclusions
This study demonstrates that submarine groundwater discharge can create localized areas of cooler water that may serve as refugia for corals during periods of heat stress and recovery. Along the western coast of the Island of Hawai’i, we found that reefs with groundwater input experienced 1 °C–2°C lower temperatures and reduced bleaching compared with nearby reefs without such input. Over 3 years during peak summer temperatures when we observed coral bleaching, the exposure of coral to temperatures above 26 °C–27°C was reduced 5%-46% of the time in areas with low to moderate flux of cooler groundwater. Moreover, lower coral loss and greater coral recovery following the extreme 2014–2015 MHW were more commonly found near mapped groundwater seeps and especially those with high flux and a strong influence in reducing nearshore temperatures. These findings demonstrate a natural mechanism of resilience that could help certain coral populations withstand the impacts of ocean warming. Protecting and managing these cool-water refugia—for example, by safeguarding groundwater water quality and ensuring sufficient flows similar to the “in-stream flow requirements” established for fish—could become a vital component of coral reef conservation strategies in areas impacted by groundwater removal and/or increasing drought. Improved understanding and characterization of SGD zones, related cold-water refugia, and associated reef community structure could be used to further quantifying the role of water management for diversity, marine protected areas, and reef sensitivity to drought and variability in runoff. A systematic census of community structure and diversity across gradients in SGD influence can therefore be an important tool for guiding marine spatial planning and coral reef conservation.
It remains uncertain to what extent corals may respond to the depression in ocean temperatures from SGD during increasingly extreme heat stress, relative to the combined effects of other direct, indirect, lethal, sublethal, abiotic, and biotic stresses that are sensitive to variations in temperature, salinity, or the chemical composition of SGD. Many of these stressors to corals and coastal ecosystems are known to be related to the addition of excess nutrients and contaminants in groundwater and coastal aquifers that discharge to nearshore ecosystems. Therefore, improved understanding of the tradeoffs of reducing groundwater contamination while ensuring the benefits of SGD to coral health and the coastal ecosystem services on which people rely can help to enhance overall resilience. This will be especially consequential for maintaining ecosystems that support commercial and subsistence fisheries and coastal cultures in general, particularly in light of anticipated climate and land use change. These results have widespread implications for coral reef conservation and coastal groundwater management across the tropics where corals and more than half of the world’s fresh SGD occur. By modifying coastal thermal and chemical environments, SGD may be critical in the steep, tectonically active margins of the Caribbean and Indo-West Pacific—regions that serve as principal sources of coral genetic dispersal essential for reef recovery following increasingly frequent mass mortality events driven by rising sea-surface temperatures.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://doi.org/10.5066/F7154FJQ, U.S. Geological Survey data release https://doi.org/10.5066/F74X569K, U.S. Geological Survey data release.
Ethics statement
The manuscript presents research on animals that do not require ethical approval for their study.
Author contributions
EG: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. FO: Writing – review & editing. CS: Writing – review & editing.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This work was supported by the U.S. Geological Survey Coastal and Marine Hazards and Resources Program, the USGS Coral Reef Project, and the National Park Service Water Resources Program.
Acknowledgments
A special thanks is warranted for Dr. Sallie Beavers and Paula Cutillo of the National Park Service who provided logistical and programmatical support for the project; Sara Peck of the National Oceanic and Atmospheric Administration for community engagement; valiant field efforts of Lisa Marrack, Lindsey Kramer, Rebecca Most, and Josh Logan; and the guidance of Courtney Couch toward mapping coral health metrics. Ilsa Kuffner (USGS) contributed numerous excellent suggestions and a timely review of the manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Author disclaimer
Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the U.S. Government.
References
Aguiar D. K., Wiegner T. N., Colbert S. L., Burns J., Abaya L., Beets J., et al. (2023). Detection and impact of sewage pollution on south kohala’s coral reefs, Hawai‘i. Mar. pollut. Bull. 188, 114662. doi: 10.1016/j.marpolbul.2023.114662
Alorda-Kleinglass A., Ruiz-Mallén I., Diego-Feliu M., Rodellas V., Bruach-Menchén J. M., and Garcia-Orellana J. (2021). The social implications of submarine groundwater discharge from an ecosystem services perspective: A systematic review. Earth-Science Rev. 221, 103742. doi: 10.1016/j.earscirev.2021.103742
Anthony K., Bay L. K., Costanza R., Firn J., Gunn J., Harrison P., et al. (2017). New interventions are needed to save coral reefs. Nat. Ecol. Evol. 1, 1420–1422. doi: 10.1038/s41559-017-0313-5
Asner G. P., Vaughn N. R., and Heckler J. (2024). Operational mapping of submarine groundwater discharge into coral reefs: application to west Hawai‘i Island. Oceans 5, 547–595. doi: 10.3390/oceans5030031
Asner G. P., Vaughn N. R., Martin R. E., Foo S. A., Heckler J., Neilson B. J., et al. (2022). Mapped coral mortality and refugia in an archipelago-scale marine heat wave. Proc. Natl. Acad. Sci. 119, e2123331119. doi: 10.1073/pnas.2123331119
Attias E., Thomas D., Sherman D., Ismail K., and Constable S. (2020). Marine electrical imaging reveals novel freshwater transport mechanism in Hawai‘i. Sci Adv. 6, eabd4866. doi: 10.1126/sciadv.abd4866
Barnas D. M., Zeff M., and Silbiger N. J. (2025). Submarine groundwater discharge drives both direct and indirect effects on organismal and community metabolism on coral reefs. Proc. R. Soc. B: Biol. Sci. 292, 20241554. doi: 10.1098/rspb.2024.1554
Bradford M. J. and Heinonen J. S. (2008). Low Flows, Instream Flow Needs and Fish Ecology in Small Streams. Canadian Water Resources J. 33, 165–180. doi: 10.4296/cwrj3302165
Bruno J. F., Côté I. M., and Toth L. T. (2019). Climate change, coral loss, and the curious case of the parrotfish paradigm: why don’t marine protected areas improve reef resilience? Annu. Rev. Mar. Sci 11, 307–345. doi: 10.1146/annurev-marine-010318-095300
Burn D., Hoey A. S., Matthews S., Harrison H. B., and Pratchett M. S. (2023). Differential Bleaching Susceptibility among Coral Taxa and Colony Sizes, Relative to Bleaching Severity across Australia’s Great Barrier Reef and Coral Sea Marine Parks. Mar. pollut. Bull. 191, 114907. doi: 10.1016/j.marpolbul.2023.114907
Camacho-Cruz K., Rey-Villiers N., Ortiz-Hernández M. C., González-Jones P., Galán-Caamal R. D. J., Matus-Hernández M., et al. (2022). Changes in the enrichment of dissolved inorganic nutrients in the coastal waters of the mexican caribbean, influenced by submarine groundwater discharges 2016–2019. Mar. pollut. Bull. 185, 114308. doi: 10.1016/j.marpolbul.2022.114308
Coles S. L., Bahr K. D., Rodgers K. S., May S. L., McGowan A. E., Tsang A., et al. (2018). Evidence of acclimatization or adaptation in hawaiian corals to higher ocean temperatures. PeerJ 6, e5347. doi: 10.7717/peerj.5347
Connell J. H. (1978). Diversity in tropical rain forests and coral reefs: high diversity of trees and corals is maintained only in a nonequilibrium state. Science 199, 1302–1310. doi: 10.1126/science.199.4335.1302
Costanza R., De Groot R., Sutton P., Van Der Ploeg S., Anderson S. J., Kubiszewski I., et al. (2014). Changes in the global value of ecosystem services. Global Environ. Change 26, 152–158. doi: 10.1016/j.gloenvcha.2014.04.002
Couch C. S., Garriques J. D., Barnett C., et al. (2014). Spatial and temporal patterns of coral health and disease along leeward hawai’i island. Coral Reefs 33, 693–704. doi: 10.1007/s00338-014-1174-x
D’Angelo C. and Wiedenmann J. (2014). Impacts of nutrient enrichment on coral reefs: new perspectives and implications for coastal management and reef survival. Curr. Opin. Environ. Sustainability 7, 82–93. doi: 10.1016/j.cosust.2013.11.029
Department of Land and Natural Resources, Commission on Water Resource Management. (2022). Aquifers of Hawaii. Available online at: https://geoportal.hawaii.gov/ (Accessed July 15, 2025).
Dimova N. T., Swarzenski P. W., Dulaiova H., and Glenn C. R. (2012). Utilizing multichannel electrical resistivity methods to examine the dynamics of the fresh water–seawater interface in two hawaiian groundwater systems. J. Geophysical Research: Oceans 117, 2011JC007509. doi: 10.1029/2011JC007509
Dollar S. J. (1982). Wave stress and coral community structure in Hawaii. Coral Reefs 1, 71–81. doi: 10.1007/BF00301688
Dulai H., Smith C. M., Amato D. W., Gibson V., and Bremer L. L. (2023). Risk to native marine macroalgae from land-use and climate change-related modifications to groundwater discharge in Hawai'i. Limnology Oceanography Lett. 8, 141–535. doi: 10.1002/lol2.10232
Fabricius K. E. (2005). Effects of terrestrial runoff on the ecology of corals and coral reefs: review and synthesis. Mar. pollut. Bull. 50, 125–146. doi: 10.1016/j.marpolbul.2004.11.028
Fackrell J. K., Glenn C. R., Thomas D., Whittier R., and Popp B. N. (2020). Stable isotopes of precipitation and groundwater provide new insight into groundwater recharge and flow in a structurally complex hydrogeologic system: West Hawai‘i, USA. Hydrogeology J. 28, 1191–2075. doi: 10.1007/s10040-020-02143-9
Fletcher C. H., Bochicchio C., Conger C. L., Engels M. S., Feirstein E. J., Frazer N., et al. (2008). “Geology of Hawaii Reefs,” in Coral Reefs of the USA. Eds. Riegl B. M. and Dodge R. E. (Dordrecht, Netherlands: Springer), 435–487. doi: 10.1007/978-1-4020-6847-8_11
Friedlander A. M., Donovan M. K., Stamoulis K. A., Williams I. D., Brown E. K., Conklin E. J., et al. (2018). Human-induced gradients of reef fish declines in the hawaiian archipelago viewed through the lens of traditional management boundaries. Aquat. Conservation: Mar. Freshw. Ecosyst. 28, 146–157. doi: 10.1002/aqc.2832
Frieler K., Meinshausen M., Golly A., Mengel M., Lebek K., Donner S. D., et al. (2013). Limiting global warming to 2 °C is unlikely to save most coral reefs. Nat. Climate Change 3, 165–170. doi: 10.1038/nclimate1674
Garrison G. H., Glenn C. R., and McMurtry G. M. (2003). Measurement of submarine groundwater discharge in Kahana Bay, O’ahu, Hawai’i. Limnology Oceanography 48, 920–928. doi: 10.4319/lo.2003.48.2.0920
Giambelluca T. W., Chen Q., Frazier A. G., Price J. P., Chen Y.-L., Chu P.-S., et al. (2013). Online rainfall atlas of Hawai‘i. Bull. Am. Meteorological Soc. 94, 313–316. doi: 10.1175/BAMS-D-11-00228.1
Glynn P. W. (1996). Coral reef bleaching: facts, hypotheses and implications. Global Change Biol. 2, 495–509. doi: 10.1111/j.1365-2486.1996.tb00063.x
Goreau T. J.F. and Hayes R. L. (2024). 2023 record marine heat waves: coral reef bleaching hotSpot maps reveal global sea surface temperature extremes, coral mortality, and ocean circulation changes. Oxford Open Climate Change 4, kgae005. doi: 10.1093/oxfclm/kgae005
Gove J. M., Williams G. J., Lecky J., Brown E., Conklin E., Counsell C., et al. (2023). Coral reefs benefit from reduced land–sea impacts under ocean warming. Nature 621, 536–542. doi: 10.1038/s41586-023-06394-w
Grossman E. E. and Fletcher C. H. (2004). Holocene reef development where wave energy reduces accommodation space, kailua bay, windward Oahu, Hawaii, U.S.A. J. Sedimentary Res. 74, 49–635. doi: 10.1306/070203740049
Grossman E. E., Logan J. B., Presto K. M., and Storlazzi C. D. (2010). Submarine groundwater discharge and fate along the coast of Kaloko-Honokōhau National Historical Park, Hawaii: Part III, spatial and temporal properties of nearshore waters and coastal groundwater plumes, December 2003–April 2006: U.S. Geological Survey. doi: 10.3133/sir20105081
Grossman E. E. and Marrack E. C. (2019). Nearshore water properties and estuary conditions along the coral reef coastline of West Hawaii Island, (2010-2014). U.S. Geological Survey Data Release. doi: 10.5066/F7154FJQ
Grossman E. E., Marrack E. C., Kramer L. K., and Most R. (2019). Coral cover and health determined from seafloor photographs and diver observations, West Hawai’i 2010-2011. U.S. Geological Survey Data Release. doi: 10.5066/F74X569K
Grossman E. E., Marrack E. C., and vanArendonk N. R. (2021). Nearshore water quality and coral health indicators along the west coast of the Island of Hawai'i, 2010–2014: U.S. Geological Survey. doi: 10.3133/ofr20201128
Hendee J., Amornthammarong N., Gramer L., and Gomez A. (2020). A novel low-cost, high-precision sea temperature sensor for coral reef monitoring. Bull. Mar. Sci 96, 97–1105. doi: 10.5343/bms.2019.0050
Hughes T. P., Anderson K. D., Connolly S. R., Heron S. F., Kerry J. T., Lough J. M., et al. (2018). Spatial and temporal patterns of mass bleaching of corals in the anthropocene. Science 359, 80–83. doi: 10.1126/science.aan8048
Hunt C. D. (2015). Baseline water-quality sampling to infer nutrient and contaminant sources at Kaloko-Honokōhau National Historical Park, Island of Hawai‘i, 2009: U.S. Geological Survey. doi: 10.3133/sir20145158
Jackson J., Donovan M., Cramer K., and Lam V. (2014). Status and Trends of Caribbean Coral Reefs: 1970-2012. (Gland, Switzerland: Global Coral Reef Monitoring Network, IUCN). Available online at: https://iucn.org/sites/default/files/import/downloads/caribbean_coral_reefs:_status_report_1970_2012.pdf (Accessed September 19, 2024).
Johnson A. G., Glenn C. R., Burnett W. C., Peterson R. N., and Lucey P. G. (2008). Aerial infrared imaging reveals large nutrient-rich groundwater inputs to the ocean. Geophysical Res. Lett. 35, 2008GL034574. doi: 10.1029/2008GL034574
Jokiel P. L. and Brown E. K. (2004). Global warming, regional trends and inshore environmental conditions influence coral bleaching in Hawaii. Global Change Biol. 10, 1627–1415. doi: 10.1111/j.1365-2486.2004.00836.x
Jokiel P. L. and Coles S. L. (1990). Response of hawaiian and other indo-pacific reef corals to elevated temperature. Coral Reefs 8, 155–162. doi: 10.1007/BF00265006
Kikuchi W. K. (1976). Prehistoric hawaiian fishponds: indigenous aquaculture influenced the development of social stratification in Hawaii. Science 193, 295–299. doi: 10.1126/science.193.4250.295
Kim G., Kim J.-S., and Hwang D.-W. (2011). Submarine groundwater discharge from oceanic islands standing in oligotrophic oceans: implications for global biological production and organic carbon fluxes. Limnology Oceanography 56, 673–825. doi: 10.4319/lo.2011.56.2.0673
Klein S. G., Roch C., and Duarte C. M. (2024). Systematic review of the uncertainty of coral reef futures under climate change. Nat. Commun. 15, 22245. doi: 10.1038/s41467-024-46255-2
Kleypas J. A., Mcmanus J. W., and Meñez L. A.B. (1999). Environmental limits to coral reef development: where do we draw the line? Am. Zoologist 39, 146–595. doi: 10.1093/icb/39.1.146
Knee K. L., Street J. H., Grossman E. E., Boehm A. B., and Paytan A. (2010). Nutrient inputs to the coastal ocean from submarine groundwater discharge in a groundwater-dominated system: relation to land use (Kona Coast, Hawaii, U.S.A.). Limnology Oceanography 55, 1105–1225. doi: 10.4319/lo.2010.55.3.1105
Kohler K. E. and Gill S. M. (2006). Coral point count with excel extensions (CPCe): A visual basic program for the determination of coral and substrate coverage using random point count methodology. Comput. Geosciences 32, 1259–1695. doi: 10.1016/j.cageo.2005.11.009
Liu G., Strong A. E., and Skirving W. (2003). Remote sensing of sea surface temperatures during 2002 barrier reef coral bleaching. Eos Trans. Am. Geophysical Union 84, 137–415. doi: 10.1029/2003EO150001
Luijendijk E., Gleeson T., and Moosdorf N. (2020). Fresh groundwater discharge insignificant for the world’s oceans but important for coastal ecosystems. Nat. Commun. 11, 12605. doi: 10.1038/s41467-020-15064-8
MacKenzie R. A. and Bruland G. L. (2012). Nekton communities in hawaiian coastal wetlands: the distribution and abundance of introduced fish species. Estuaries Coasts 35, 212–265. doi: 10.1007/s12237-011-9427-1
Marrack L., Beavers S., Weijerman M., and Most R. (2014). Baseline Assessment of the Coral Reef Habitat in Kaloko-Honokōhau National Historical Park Adjacent to the Shores at Kohanaiki Development 2006-2007. Technical Report No. 190. (Honolulu, Hawai‘i: Pacific Cooperative Studies Unit, University of Hawai‘i), 58. Available online at: https://scholarspace.manoa.hawaii.edu/server/api/core/bitstreams/d0e26d82-3e35-4574-93b3-87a7fc0870a4/content (Accessed August 29, 2025).
McKenzie M. M., Giambelluca T. W., and Diaz H. F. (2019). Temperature trends in hawai'i: A century of change 1917–2016. Int. J. Climatology 39, 3987–40015. doi: 10.1002/joc.6053
Moore W. S. (2010). The effect of submarine groundwater discharge on the ocean. Annu. Rev. Mar. Sci 2, 59–88. doi: 10.1146/annurev-marine-120308-081019
Muñiz-Castillo A. I., Rivera-Sosa A., McField M., Chollett I., Eakin C. M., Enríquez S., et al. (2024). Underlying drivers of coral reef vulnerability to bleaching in the mesoamerican reef. Commun. Biol. 7, 1452. doi: 10.1038/s42003-024-07128-y
Oberle F. K.J., Prouty N. G., Swarzenski P. W., and Storlazzi C. D. (2022). High-Resolution observations of submarine groundwater discharge reveal the fine spatial and temporal scales of nutrient exposure on a coral reef: Faga’alu, AS. Coral Reefs 41, 849–545. doi: 10.1007/s00338-022-02245-8
Oberle F. K. J., Storlazzi C. D., Cheriton O. M., Takesue R. K., Hoover D. J., Logan J. B., et al. (2019). Physicochemical controls on zones of higher coral stress where black band disease occurs at mākua reef, Kaua‘i, Hawai‘i. Front. Mar. Sci 6. doi: 10.3389/fmars.2019.00552
Oki D. S. (1999). Ground-water resources in Kaloko-Honokōhau National Historical Park, Island of Hawai‘i, and numerical simulation of the effects of ground-water withdrawals: U.S. Geological Survey. doi: 10.3133/wri994070
Oki D. S. (2004). Trends in streamflow characteristics at long-term gaging stations, Hawaii: U.S. Geological Survey. doi: 10.3133/sir20045080
Perelman J. N., Tanaka K. R., Smith J. N., Barkley H. C., and Powell B. S. (2024). Subsurface temperature estimates from a regional ocean modelling system (ROMS) reanalysis provide accurate coral heat stress indices across the main hawaiian islands. Sci. Rep. 14, 66205. doi: 10.1038/s41598-024-56865-x
Peterson R. N., Burnett W. C., Glenn C. R., and Johnson A. G. (2009). Quantification of point-source groundwater discharges to the ocean from the shoreline of the big Island, Hawaii. Limnology Oceanography 54, 890–9045. doi: 10.4319/lo.2009.54.3.0890
Presto K. M., Storlazzi C. D., Logan J. B., and Grossman E. E. (2007). Submarine ground-water discharge and fate along the coast of Kaloko-Honokōhau National Historical Park, Hawaii, Part I: Time-series measurements of currents, waves, salinity, and temperature, November 2005–July 2006: U.S. Geological Survey. doi: 10.3133/ofr20071310
Prouty N. G., Swarzenski P. W., Fackrell J. K., Johannesson K., and Palmore C.D. (2017). Groundwater-derived nutrient and trace element transport to a nearshore kona coral ecosystem: experimental mixing model results. J. Hydrology: Regional Stud. 11, 166–177. doi: 10.1016/j.ejrh.2015.12.058
Reimer J. D., Peixoto R. S., Davies S. W., Traylor-Knowles N., Short M. L., Cabral-Tena R. A., et al. (2024). The fourth global coral bleaching event: where do we go from here? Coral Reefs 43, 1121–1125. doi: 10.1007/s00338-024-02504-w
Ribaud-Laurenti A., Hamelin B., Montaggioni L., and Cardinal D. (2001). Diagenesis and its impact on sr/ca ratio in holocene acropora corals. Int. J. Earth Sci. 90, 438–451. doi: 10.1007/s005310000168
Rodgers K. S., Jokiel P. L., and Hawaii Institute of Marine Biology (HIMB) (2016). Coral Reef Assessment and Monitoring Program (CRAMP) study sites: Main Hawaiian Islands. (Honolulu, HI: Distributed by the Pacific Islands Ocean Observing System (PacIOOS)). Available online at: http://pacioos.org/metadata/hi_himb_all_cramp_sites.html (Accessed October 7, 2019).
Rogers C. S. and Miller J. (2013). Coral diseases cause reef decline. Science 340, 1522–1522. doi: 10.1126/science.340.6140.1522-a
Sakihara T. S., Dudley B. D., MacKenzie R. A., and Beets J. P. (2015). Endemic grazers control benthic microalgal growth in a eutrophic tropical brackish ecosystem. Mar. Ecol. Prog. Ser. 519, 29–45. doi: 10.3354/meps11099
Santos I. R., Chen X., Lecher A. L., Sawyer A. H., Moosdorf N., Rodellas V., et al. (2021). Submarine groundwater discharge impacts on coastal nutrient biogeochemistry. Nat. Rev. Earth Environ. 2, 307–323. doi: 10.1038/s43017-021-00152-0
Sherrod D. R., Sinton J. M., Watkins S. E., and Brunt K. M. (2021). Geologic map of the State of Hawaii: U.S. Geological Survey. doi: 10.3133/sim3143
Silbiger N. J., Donahue M. J., and Lubarsky K. (2020). Submarine groundwater discharge alters coral reef ecosystem metabolism. Proc. R. Soc. B: Biol. Sci. 287, 20202743. doi: 10.1098/rspb.2020.2743
Souter D., Planes S., Wicquart J., Obura D., and Staub F. (2021). Status of Coral Reefs of the World: 2020 (Global Coral Reef Monitoring Network (GCRMN) and International Coral Reef Initiative (ICRI)). doi: 10.59387/WOTJ9184
Storlazzi C. D., Cheriton O. M., Messina A. M., and Biggs T. W. (2018). Meteorologic, oceanographic, and geomorphic controls on circulation and residence time in a coral reef-lined embayment: faga’alu bay, American Samoa. Coral Reefs 37, 457–469. doi: 10.1007/s00338-018-1671-4
Storlazzi C. D. and Presto K. M. (2005). Coastal circulation and water column properties along Kaloko-Honokōhau National Historical Park, Hawaii, Part I: Measurements of waves, currents, temperature, salinity, and turbidity, April–October 2004: U.S. Geological Survey. doi: 10.3133/ofr20051161
Street J. H., Knee K. L., Grossman E. E., and Paytan A. (2008). Submarine groundwater discharge and nutrient addition to the coastal zone and coral reefs of Leeward Hawai’i. Mar. Chem. 109, 355–376. doi: 10.1016/j.marchem.2007.08.009
Swarzenski P. W., Dulai H., Kroeger K. D., Smith C. G., Dimova N., Storlazzi C. D., et al. (2017). Observations of nearshore groundwater discharge: kahekili beach park submarine springs, Maui, Hawaii. J. Hydrology: Regional Stud. 11, 147–165. doi: 10.1016/j.ejrh.2015.12.056
Taniguchi M., Dulai H., Burnett K. M., Santos I. R., Sugimoto R., Stieglitz T., et al. (2019). Submarine groundwater discharge: updates on its measurement techniques, geophysical drivers, magnitudes, and effects. Front. Environ. Sci 7. doi: 10.3389/fenvs.2019.00141
United States Geological Survey (2024). USGS 194125156015501 8-4161–01 KAHO Well 3, HI, in USGS water data for the Nation: U.S. Geological Survey National Water Information System database. Available online at: https://waterdata.usgs.gov/monitoring-location/USGS-194125156015501 (Accessed July 26, 2024).
Van Hooidonk R., Maynard J. A., and Planes S. (2013). Temporary refugia for coral reefs in a warming world. Nat. Climate Change 3, 508–511. doi: 10.1038/nclimate1829
Van Hooidonk R., Maynard J., Tamelander J., Gove J., Ahmadia G., Raymundo L., et al. (2016). Local-scale projections of coral reef futures and implications of the paris agreement. Sci. Rep. 6, 39666. doi: 10.1038/srep39666
Veron J. E. N., Devantier L. M., Turak E., Green A. L., Kininmonth S., Stafford-Smith M., et al. (2009). Delineating the coral triangle. Galaxea J. Coral Reef Stud. 11, 91–100. doi: 10.3755/galaxea.11.91
Walsh W. J., Cotton S., Barnett C., Couch C., Preskitt L., Tissot B., et al. (2013). Long-Term Monitoring of Coral Reefs of the Main Hawaiian Islands. (Honolulu, HI: NOAA). Available online at: https://dlnr.hawaii.gov/dar/files/2014/04/NOAA_2013_WHi_-Mon_-Rep.pdf (Accessed July 15, 2015).
Young C., Tamborski J., and Bokuniewicz H. (2015). Embayment scale assessment of submarine groundwater discharge nutrient loading and associated land use. Estuarine Coast. Shelf Sci 158, 20–30. doi: 10.1016/j.ecss.2015.02.006
Keywords: coral, reef, groundwater, submarine groundwater discharge, cold-water refugia, thermal stress, resilience, Hawaii
Citation: Grossman EE, Oberle FKJ and Storlazzi CD (2025) Submarine groundwater discharge creates cold‐water refugia that can mitigate exposure of heat stress in nearshore corals. Front. Mar. Sci. 12:1621298. doi: 10.3389/fmars.2025.1621298
Received: 30 April 2025; Accepted: 12 September 2025;
Published: 07 October 2025.
Edited by:
Henrietta Dulai, University of Hawaii at Manoa, United StatesReviewed by:
Aarón Israel Muñiz-Castillo, Healthy Reefs Initiative, United StatesCourtney Saltonstall Couch, University of Hawaii at Mānoa, United States
Andrea Kealoha, University of Hawaii Maui College, United States
Néstor Rey Villiers, National Polytechnic Institute, Mexico
Copyright © 2025 Grossman, Oberle and Storlazzi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Eric E. Grossman, ZWdyb3NzbWFuQHVzZ3MuZ292
†ORCID: Eric E. Grossman, orcid.org/0000-0003-0269-6307
Ferdinand K. J. Oberle, orcid.org/0000-0001-8871-3619
Curt D. Storlazzi, orcid.org/0000-0001-8057-4490