 Kaitlyn A. O’Brien1*†
Kaitlyn A. O’Brien1*† John K. Carlson2†
John K. Carlson2† Enric Cortés2†
Enric Cortés2† William B. Driggers III3
William B. Driggers III3 Bryan S. Frazier4†
Bryan S. Frazier4† Robert J. Latour1†
Robert J. Latour1†- 1Virginia Institute of Marine Science, William & Mary, Gloucester Point, VA, United States
- 2Panama City Laboratory, National Marine Fisheries Service, Southeast Fisheries Science Center, Panama City, FL, United States
- 3Mississippi Laboratories, National Marine Fisheries Service, Southeast Fisheries Science Center, Pascagoula, MS, United States
- 4South Carolina Department of Natural Resources, Marine Resources Research Institute, Charleston, SC, United States
Introduction: Methods combining data from spatially limited, independently conducted surveys indicate a preliminary recovery for coastal shark species along the Atlantic. However, anthropogenic climate change is expected to shift distributions and alter migration timing for these highly migratory species, potentially affecting survey catchability and interpretation of abundance indices.
Methods: Vector autoregressive spatiotemporal (VAST) models were applied to data from six fishery-independent surveys of six coastal shark stocks to generate area-weighted indices of abundance. Area-weighted indices, trends in density over space and time, and analysis of density anomalies were used to evaluate changes in a stock’s spatial distribution across the U.S. southeast Atlantic. In addition to VAST, generalized linear mixed models were used to generate indices of abundance for each survey, which served as inputs to two previously implemented reconciliation methods in coastal shark stock assessments: dynamic factor analysis (DFA) and Bayesian hierarchical analysis (Conn).
Results: The index standardization methods, particularly VAST and Conn, largely agreed with one another and appeared robust to spatial patterns. Only two of the six shark stocks showed increasing trends by the end of the time series, with indices for multiple species plateauing or declining. Positive trends in density and increased variability in density anomalies in the VAST models across the northern extent of the surveyed spatial domain suggests a potential northward expansion or a timing discrepancy between migration onset and sampling efforts for multiple species.
Discussion: Overall, the VAST models provided evidence of spatial changes that could impact each survey’s catchability, thus complicating the interpretation of abundance trends. These findings underscore the importance of accounting for spatiotemporal dynamics in future stock assessments and fisheries management strategies.
1 Introduction
Abundance of coastal shark populations in the western North Atlantic Ocean declined dramatically from the mid-1970s through the 1990s due to overexploitation (Baum et al., 2003; Burgess et al., 2005; Baum and Blanchard, 2010; SEDAR, 2017, 2020). Despite evidence documenting a preliminary recovery of several coastal shark species inhabiting the southeast coast of the U.S. and Gulf of Mexico (GOM; Carlson et al., 2012; Peterson et al., 2017a), low intrinsic population growth rates and high susceptibility to overfishing underscore the need for continued monitoring and assessment (Musick, 1999; Dulvy and Forrest, 2010; Cortés et al., 2012). Uncertainty associated with the degree of unregulated or unreported fishing activity further emphasizes the importance of robust monitoring efforts (Peterson et al., 2022). Coastal sharks fulfill an important role in ecosystem stability and changes in their abundance or distribution, including climate change, can have far-reaching impacts on economically high-value fisheries and overall ecosystem health (Stevens et al., 2000; Ferretti et al., 2010; Britten et al., 2014).
An index of relative abundance (hereafter index) is a primary input for most stock assessment methods. Use of an index as a population indicator requires assuming it is proportional to total abundance, such that changes in the index can be used to infer changes in stock size (Cortés et al., 2015; Hoyle et al., 2024). Ideally, an index is developed from a fishery-independent survey that consistently samples the entire spatial range of the target species (Hilborn and Walters, 1992; Stevens et al., 2000). However, monitoring species within the large coastal shark (LCS) and small coastal shark (SCS) management complexes (NMFS, 1993) is particularly challenging because these species have extensive home ranges that extend across domestic and/or international management boundaries (Cortés et al., 2015; Calich et al., 2018; Kohler and Turner, 2019), complex spatiotemporal migration patterns (Hammerschlag et al., 2012, 2022; Papastamatiou et al., 2013), and overall low economic value. These factors collectively lead to limited resources for survey programs and biological sampling regimes (Stevens et al., 2000; Pilling et al., 2008; Ellis et al., 2009), which creates data constraints and reliance on information from several independent and spatially fragmented surveys to estimate trends of abundance. Given underlying complexities in coastal shark habitat utilization and movement, these spatially restricted surveys may not individually provide an index that accurately represents temporal trends in stock abundance (Maunder et al., 2006; Conn, 2010; Francis, 2011; SEDAR, 2017).
Changes in environmental conditions can disrupt coastal ecosystems, potentially triggering cascading effects on shark populations and their associated food webs (Ferretti et al., 2010). Oceanic conditions, habitat availability, prey abundance, and prey distribution will be affected by anthropogenic climate change, and the resultant effects are anticipated to impact coastal sharks (Perry et al., 2005). Sea level rise, ocean acidification, and deoxygenation may directly impact the physiological processes of sharks, including reproduction and metabolic rates (Rosa et al., 2017; Crear et al., 2019; Diaz-Carballido et al., 2022). Additionally, changes in ocean currents, primary productivity, and abiotic conditions can affect the availability of suitable habitats for coastal sharks during different life stages, such as nurseries for juveniles or feeding grounds for adults (Dulvy et al., 2014; Birkmanis et al., 2020; Niella et al., 2020; Osgood et al., 2021). Distributions and migration patterns of coastal sharks are also anticipated to be impacted by climate change (Chin et al., 2010; Hare et al., 2016; Hammerschlag et al., 2022; Quinlan, 2023; Manz et al., 2025) since changing sea temperatures and other abiotic factors can alter the distribution and abundance of prey, and by extension, the associated predatory fishes (Heithaus et al., 2008; Goodman et al., 2022). Understanding and predicting the cumulative effects of anthropogenic climate change on coastal shark populations can aid the development of effective management and conservation strategies amidst ongoing environmental change.
Previous studies of coastal sharks in the northwest Atlantic have utilized two different approaches to provide overall abundance trends based on multiple standardized indices: dynamic factor analysis (DFA) and a Bayesian hierarchical method (Conn). Both methods require individual time series of indices as inputs, but each handles reconciliation of the abundance indices differently. DFA is a multivariate time series dimension reduction technique (Zuur et al., 2003a, 2003b) that explains the common dynamics of a large number of time series with a small number of latent factors (Peterson et al., 2017a, 2021a, 2021b). However, application of DFA requires assigning implicit weights to each of the time series which can be challenging when the spatial extent of sampling by the constituent surveys differs substantially (Peterson et al., 2021b; Grüss et al., 2023a). The Conn method (Conn, 2010) is a hierarchical analysis that assumes each index is subject to sampling and process error. Sampling error (e.g., coefficient of variation; CV) is assumed to be estimated as part of the analysis that generates the indices, but process error, which describes the degree to which an index measures ‘artifacts’ above and below the relative abundance of the population, is accounted for in the hierarchical analysis. This method separates sampling and process errors for each time series, models the overall trend for all indices, and is assumed to remain robust to differences in gear selectivity across surveys and trends in spatial mixing (Conn, 2010). Additionally, uncertainty is generally large as the estimation procedure could overestimate process error and be overly conservative when estimating changes in abundance on the scale of the population.
Spatiotemporal models are an emerging class of statistical models that can be used to analyze survey data from multiple sources and provide estimates of population density over space and time, including derived quantities such as area-weighted indices of abundance (Thorson, 2019). Advantages of spatiotemporal models are refined estimates of precision (Cao et al., 2017; Thorson and Barnett, 2017), better characterization of how environmental variables shape spatial distributions (Thorson et al., 2020; Hansell and McManus, 2025), enhanced understanding of spatial and spatiotemporal distributions and potential distribution shifts (Thorson et al., 2016; Perretti and Thorson, 2019; O’Leary et al., 2020), and improved characterization of uncertainty (Thorson et al., 2015; Grüss et al., 2023a, 2023b). However, spatiotemporal models are computationally demanding and may require more data than are readily available for data-limited species such as coastal sharks.
Growing concerns regarding expected changes in survey catchability resulting from shifting distributions have generated interest in applying spatiotemporal models to coastal shark survey data. Thus, this study explored the potential use of spatiotemporal index standardization methods for coastal sharks. The specific objectives were to 1) develop spatiotemporal models for six western North Atlantic shark stocks by integrating data from six fishery-independent survey programs, 2) investigate potential shifts in species distributions along the U.S. Atlantic coast, and 3) compare the area-weighted indices of abundance from the spatiotemporal model to indices estimated from other reconciliation approaches (DFA and Conn). This study is intended to provide information for future stock assessments and management regulations for these largely data-limited species.
2 Materials and methods
2.1 Data sources
Catch and effort data were compiled from six fishery-independent shark surveys, including four bottom longline surveys (the Virginia Shark Monitoring and Assessment Program, VASMAP; the Southeast Fisheries Science Center Bottom Longline Survey, SEFSC; the South Carolina Coastal Longline Survey, SC; and the Georgia Red Drum Longline Survey, GA), one bottom gillnet survey (the Gulf of Mexico Shark Pupping and Nursery Gillnet Survey, GULFSPAN), and one bottom trawl survey (the SouthEast Area Monitoring and Assessment Program-South Atlantic Coastal Trawl Survey, SEAMAP; Supplementary Table S1, Figure 1; SEAMAP-SA Data Management Work Group, 2012). Data from six shark species were analyzed, including two SCS (blacknose shark, Carcharhinus acronotus, Atlantic stock only (A.); finetooth shark, C. isodon) and four LCS (bull shark, C. leucas; sandbar shark, C. plumbeus; spinner shark, C. brevipinna; tiger shark, Galeocerdo cuvier). Species were chosen based on differences in data availability, representation across surveys, variation in life history traits, and consultations with shark assessment scientists and experts in the field to ensure a diverse dataset for analysis. Further, the four LCS species were included as they were anticipated to be assessed in the coming years, providing an opportunity to compare results with future stock assessments (E. Cortés; NOAA Fisheries, personal communication).
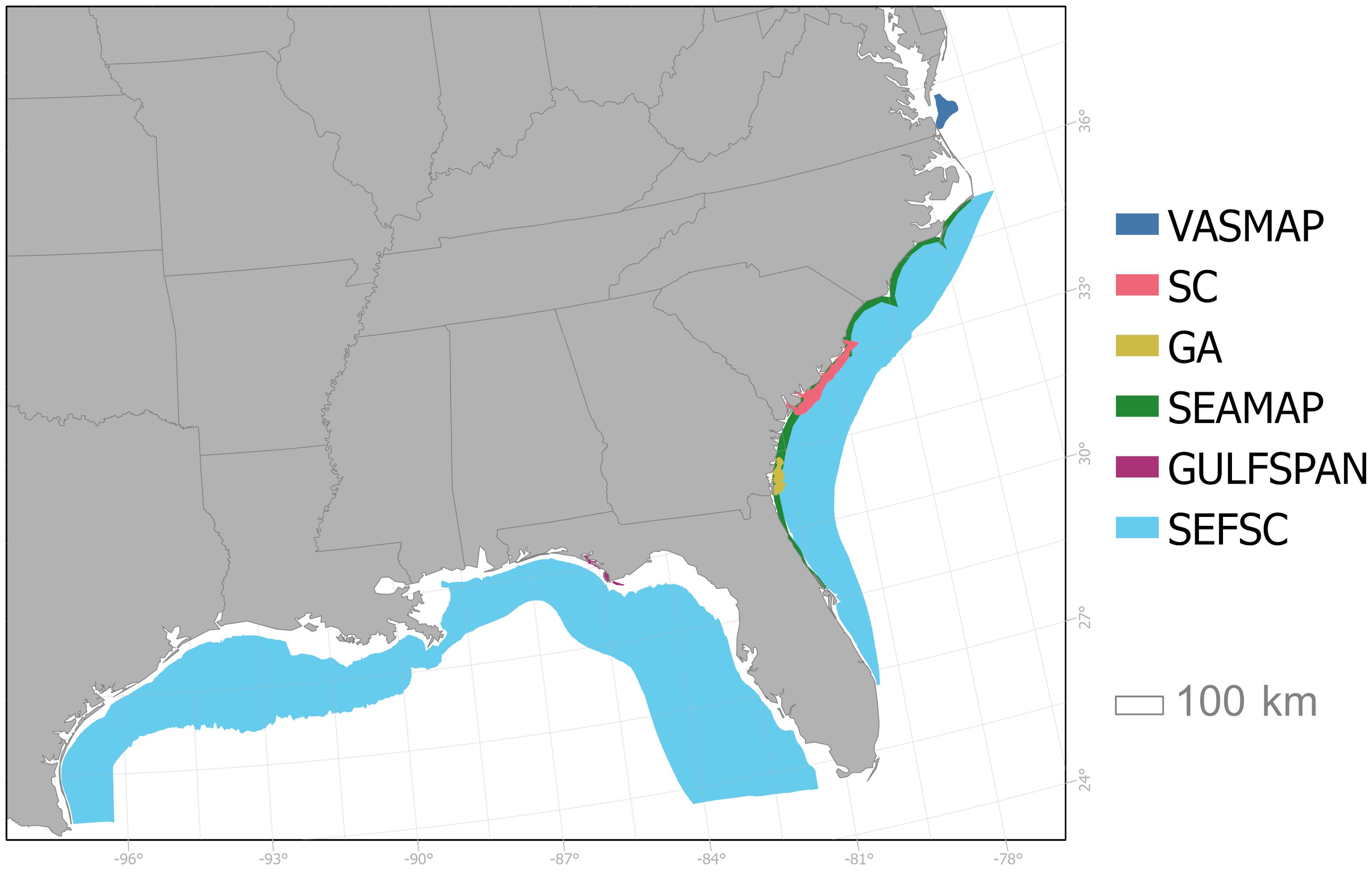
Figure 1. Map of the six fishery-independent surveys used in this study with shaded areas indicating the spatial extent of each survey. Abbreviations include: the Virginia Shark Monitoring and Assessment Program (VASMAP; dark blue), the South Carolina Coastal Longline Survey (SC; pink), the Georgia Red Drum Longline Survey (GA; yellow), the SouthEast Area Monitoring and Assessment Program-South Atlantic Coastal Trawl Survey (SEAMAP; green), the Gulf of Mexico Shark Pupping and Nursery Gillnet Survey (GULFSPAN; purple), the Southeast Fisheries Science Center Bottom Longline Survey (SEFSC; light blue).
A rubric based on previous Highly Migratory Species (HMS) stock assessments (ICCAT, 2012) was applied to evaluate the utility of survey programs for each species; considerations included geographic and temporal coverage, sampling design, and an overall proportion positive (sampling events where at least one target animal was captured) of at least 0.05 (Table 1). Neonates were removed from the survey data based on length-based criteria from the literature to maintain consistency with current stock assessments, which—when data permit—typically model neonates separately as indicators of recruitment (Supplementary Table S2). While sex and size data were available across all surveys included in this study, the analyses were not stratified by these variables due to data limitations and convergence constraints associated with finer-scale model resolution. Sample depth was treated as a continuous variable while year, month/season, and station/area were treated as categorical variables, and levels of those variables were excluded from analyses if the species of interest was not present during at least one sampling event.
Caution should be exercised when applying DFA to one or more time series with missing years as performance of the methods can decline (Peterson et al., 2021a, 2021b). For the six fishery-independent shark surveys, only one program sampled coastal sharks through the 1970s and early 1980s (VASMAP). Therefore, data from 1989–2021 were analyzed in this study to limit the number of missing years for constituent surveys and achieve a more balanced contribution of all data sources to the resulting indices. Coastal shark stocks were depleted by the early 1990s (SEDAR, 2006; Carlson et al., 2012; Peterson et al., 2017a) and federal management was in its infancy (NMFS, 1993). Therefore, defining a starting year in 1989 still enabled analysis of potential population recoveries following management.
2.2 Spatiotemporal analysis
2.2.1 Model structure
For the spatiotemporal modeling approach, the Vector Autoregressive Spatiotemporal modeling platform (VAST, Thorson and Barnett, 2017; Thorson, 2019) was utilized through the R package VAST (Thorson, 2024). The model structure consists of two processes to support delta or zero-inflated models that can predict variation in density across multiple locations and time intervals. Given the high frequency of zero observations in the survey datasets and to remain consistent with previous methodology, a zero-inflated negative binomial distribution (ZINB) was chosen for the response variable, catch (C). Sampling unoccupied habitat will always generate a catch of zero (i.e., true zero) and sampling occupied habitat may also generate a catch of zero if animals are not captured (i.e., false zero; Martin et al., 2005). Therefore, the ZINB probability mass function for is given by:
where is the probability of a false zero and is the negative binomial probability mass function with variance specified as a quadratic function of the mean (Equation 1).
Both the zero-inflation (, binomial) and count processes (, negative binomial) were modeled using VAST:
where the respective linear predictors were comprised of year intercepts , spatial variation (), spatiotemporal variation (), and catchability covariates , where is the location of sample , is the year of sample , and and are the effects of catchability covariate . The and are the link-transformed predictors for the zero-inflation (binomial) and count component (negative binomial), where is the area sampled or active area for sample treated as an offset for positive catches. Initial model runs included spatial and spatiotemporal variation in the zero-inflation component, but they were unstable due to a lack of sufficient information. Therefore, models were simplified to estimate only a single year intercept in the zero-inflation part which was a necessary departure from recommended model building practices (Equations 2, 3; Thorson and Barnett, 2017; Thorson, 2019).
The count component was structured to estimate temporal variation for each year and spatial and spatiotemporal effects. Both spatial and spatiotemporal effects were modeled as Gaussian Markov random fields, which describe the random variation in population density over latitude and longitude (or northings and eastings) with spatial covariance defined as a Matérn process (Thorson et al., 2015). Random fields were assumed to be stationary to enable analysis of geometric anisotropy, with the number of random fields dependent upon the user-specified spatial domain and the number of specified knots. Knots were defined at equally spaced locations within the spatial domain (i.e., 2D grid) due to the spatially unbalanced data from the different surveys (Thorson, 2019). Given differences in the surveys considered, encounter rates, and sample sizes per species, the number of knots was evaluated for each species individually (Table 2).

Table 2. The selected vector autoregressive spatiotemporal (VAST) models and dynamic factor analysis (DFA) model, fitted to survey-specific indices of abundance derived from generalized linear mixed models (GLMMs), for small coastal (top) and large coastal (bottom) shark species.
2.2.2 Covariates and derived quantities
Catchability covariates, or variables that could plausibly affect catch rates but not reflect variation in population density, were considered in both linear predictors. To acknowledge catchability differences among sampling programs, survey was included as a catchability effect in both linear predictors of the model. If convergence issues occurred, survey was retained in only one linear predictor. The survey with the largest spatial scale was selected as the reference data set for each target species prior to model fitting such that a fishing power ratio relative to the reference data set was estimated for the other surveys. Other catchability variables considered included the depth of sampling gear (treated as either a linear or nonlinear effect depending upon the species) and month of sampling.
Parameter estimation was performed in Template Model Builder (Kristensen et al., 2023), with model convergence checked by ensuring the absolute value of the log-likelihood final gradient at the maximum likelihood estimate was less than 0.0001 for all parameters, and that the Hessian of the likelihood function was positive definite. Model diagnostics were evaluated using the R package DHARMa (Hartig and Lohse, 2022), and model selection was based on Akaike’s Information Criterion (AIC; Akaike, 1973). A final model for each species was used to calculate area-weighted indices:
where is the area associated with knot that is within stratum , is the number of knots, is the predicted density at knot at time when , and is the abundance index at time for stratum (Equation 4). Standard errors for the index were calculated by TMB using the delta-method variance approximation.
To evaluate potential temporal patterns in species density along the Atlantic coast, the study area was divided into three sections: South (s< 30.5°N), Middle (30.5°N ≤ s< 33.5°N), and North (s ≥ 33.5°N). Area-weighted density indices for each section were plotted and compared to assess changes over time. Additionally, location-specific density trends through time were analyzed using quantile regression (R package quantreg; Koenker et al., 2024) and spatial and regional density anomalies – defined as where indicates the starting year for a particular species – were visualized graphically.
2.2.3 Area sampled estimation
Spatiotemporal models, like VAST, generally require an estimation of area sampled for each observation to estimate an area weighted index. For the SEAMAP bottom trawl survey, net dimensions and tow location information was used to estimate area swept. However, estimation of the area sampled by passive gears such as gillnets and longlines is more challenging because information on the physical properties of sampling sites is needed (Løkkeborg et al., 2010) but often not readily available. Therefore, area sampled for the passive gear surveys were approximated by incorporating the target species average swimming speed:
where indexes the sample, denotes the target species, is the estimated area sampled, is the length of gear (km), is the average swimming speed (km/hr) based on the scientific literature (Supplementary Table S2), is the amount of time (hrs) that the gear deployed, and is the average time of capture for species according to the hook timer data from the VASMAP survey (Equation 5; see Peterson et al., 2017b for details).
2.3 Dynamic factor and Bayesian hierarchical analysis
2.3.1 Indices of abundance
As mentioned previously, both DFA and Conn require standardized indices of abundance for each survey as inputs. For that reason, traditional generalized linear mixed models (GLMMs; Bolker et al., 2009) were used to standardize the species-specific catch data for each survey and provide estimated annual indices. There was a high frequency of zero observations for all focal species, which was expected given low overall abundance associated with predatory species relative to organisms at lower trophic levels. The number of sharks captured per sampling event was defined as the response variable with effort modeled as an intercept offset defined at the natural log of 100 hook-hours, net area-hours, and area swept for longline, gillnet, and trawl gears, respectively. Various discrete count distributions, including zero-inflated and zero-altered (also known as hurdle) parameterizations, were considered for the target species.
The generalized additive models for location, scale, and shape regression framework (R package gamlss; Stasinopoulos et al., 2023) was used to fit the GLMMs. Correlation and collinearity of variables were assessed using scatter plot matrices (SPLOMs) and variance inflation factors (VIFs) with highly correlated variables (≳ 0.7) or those with large VIFs (> 5) not mutually included in model parameterization (Zuur et al., 2009). The covariates examined varied by survey; year was included in all models, while station/area, month/season, and depth were only included in a subset of models. Model selection was based on AIC corrected for small sample size (AICc) and 10-fold cross-validation considering mean square error, root mean square error, and mean absolute error. Model validation was achieved through visual examination of diagnostic plots (QQ-plots and residuals), overdispersion was assessed by checking that the estimated dispersion ratio was close to 1.0, and selected distributions were verified through simulation analysis to ensure they could support the observed frequency of zeros. Final models were used to generate predicted indices of relative abundance and uncertainty estimates were generated from 1000 nonparametric bootstrapped samples (Efron and Tibshirani, 1993).
2.3.2 Dynamic factor analysis
DFAs were fitted using the state-space multivariate autoregressive modeling R package MARSS (Holmes et al., 2023). The general form of a DFA model is:
where Equation 6a and Equation 6b represent the observation and process components at time , respectively. The vector is comprised of the abundance time series, is the vector of common trends , is the matrix of factor loadings of each time series on the common trend(s), the matrix contains the estimated coefficients for the covariates , and and are variance-covariance matrices associated with observation error vector and process error vector , respectively. The matrix was set equal to identity matrix while four parameterizations were investigated for matrix : diagonal with equal variance and zero covariance, diagonal with unequal variance and zero covariance, non-diagonal with equal variance and equal covariance, and unconstrained (Holmes et al., 2021). Statistical significance of covariates was inferred when 0 was not included in the 95% confidence intervals (CIs).
Application of DFAs in this study followed the approach of Peterson et al. (2017a) where the four covariance structures were first evaluated in the absence of covariates followed by the introduction of covariates to the model with the selected covariance structure. The analysis was restricted to estimate a single latent trend (m = 1). The covariates considered were large scale climate indices – the North Atlantic Oscillation (NAO) index, the Atlantic Multidecadal Oscillation (AMO) index, and the Gulf Stream (GSI) index – all of which are correlated with highly suitable habitat for multiple coastal shark species (O’Brien et al., 2024; Beltz, 2024; Supplementary Figure S1). Graphical analysis, AICc, and fit ratio (Zuur et al., 2003b) were used to discern the most parsimonious model at each step of the analysis.
Time series are typically standardized (z-scored) prior to performing a DFA (Zuur et al., 2003a, 2003b) resulting in the estimated common trend(s) being in log-space on a unitless scale that spans positive and negative values. However, when the time series are indices of relative abundance, back-transformation from log space can distort the relative scale of the indices, thereby compromising interpretation of abundance trends. To address this issue, a rescaling technique was applied that preserves the error structure and relative scale of the survey indices, remains consistent with the requirements of a DFA (z-score), and allows for successful back-transformation of the resulting DFA index (see Peterson et al., 2021b for details).
2.3.3 Bayesian hierarchical analysis
The general form of the Conn model is:
where is index in year (scaled by its mean), reflects the changes in abundance at population scale at time , is the scaling coefficient for index (related to ), and and are the process and sampling standard deviations (Conn, 2010) (Equation 7). Parameters were estimated within a Bayesian framework, following prior distribution specifications and Markov Chain Monte Carlo configurations outlined by Conn (2010). Models were fitted using the R2jags package to interface the R programming environment with the JAGS software.
3 Results
3.1 Data sources
The number of informative surveys varied by species, ranging from one for bull sharks to five for blacknose (A.) sharks (Table 1). Data from SEFSC and SC were used in the analysis of five of six species; only the finetooth shark analysis excluded SEFSC data, while the tiger shark analysis did not include SC data. The SEAMAP dataset provided data solely for blacknose (A.) sharks. Data from VASMAP were used in the analysis of all LCS species (except for bull sharks, which were analyzed only in the GOM) and one SCS species—the blacknose (A.) shark—while GA data were used only for blacknose (A.) and sandbar sharks. Despite data filtering, a high frequency of zero observations remained for all species, ranging from 81.5% for sandbar sharks to 93.2% for spinner sharks (Table 1).
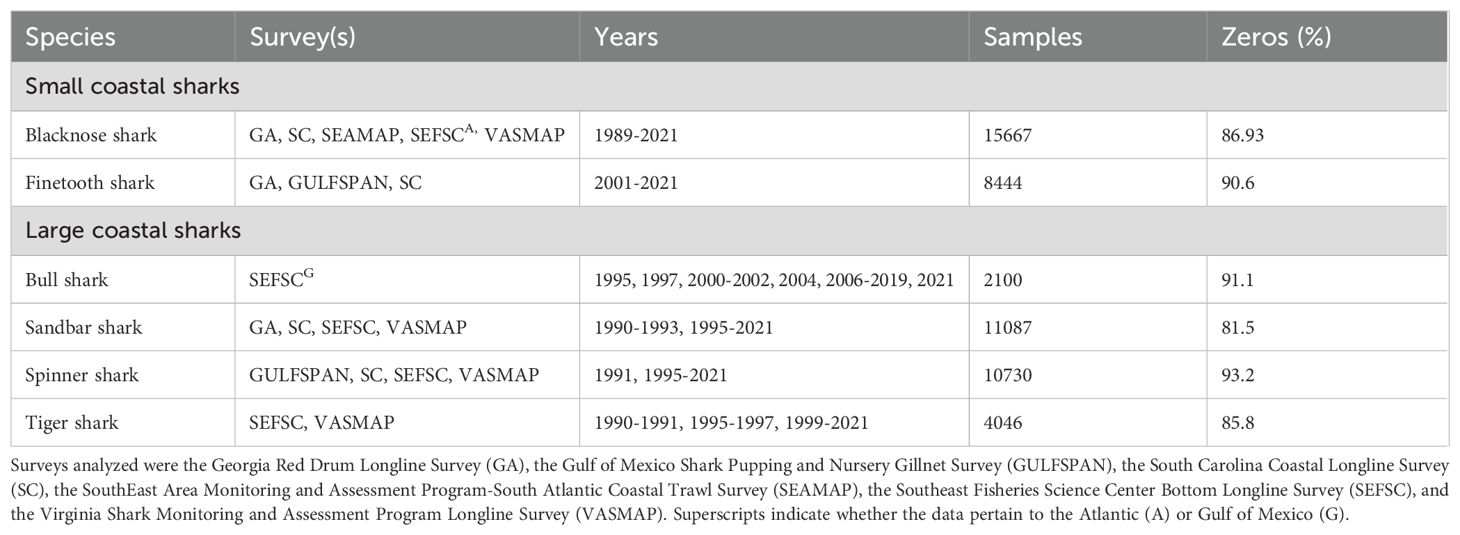
Table 1. Surveys, years examined, total samples, and frequency of samples not capturing the species of interest (Zeros %) for species in the small coastal (top) and large coastal shark (bottom) complexes.
3.2 Spatiotemporal analysis
VAST models successfully converged for all species expect finetooth sharks (Table 2). Inclusion of spatial and spatiotemporal random effects for the count model led to convergence issues for finetooth sharks, likely because the surveys with adequate catch information were spatially small and distinct. Similarly, models for bull sharks that included spatiotemporal effects in the count component also failed to converge; however, parameterizations with a spatial random field held constant over time were successful. Depth emerged as the most consistently supported catchability effect in the zero-inflated component, often modeled as a nonlinear effect (Table 2). For the most empirically supported models, depth, survey, and month were frequently included as catchability covariates in the count component, again with depth often represented as a nonlinear effect. Additionally, further evaluation of the estimated area sampled for passive gear was shown to have a scaling effect. While this factor should be further considered for stock assessment applications, the method performed well for comparative purposes in this study.
Areas of highest predicted density varied across species. The blacknose (A.) shark was the only SCS analyzed using VAST, and predicted densities were highest in shallower waters close to shore, particularly at the openings of estuaries and bays, and lowest farther offshore in deeper waters (the blacknose (A.) shark was selected as a representative species in Figure 2; see Supplementary Materials – Supplementary Figures S2–S5 for all other species). These predictions generally differed from those of the LCS, where higher densities were often farther offshore. However, there was notable variability within the LCS complex. In the GOM, predicted densities for bull sharks (only GOM was considered due to extremely low catches in the Atlantic) increased in the Northern Gulf of Mexico Hypoxic Zone (NGMHZ) over time, and the highest predicted density for spinner shark was almost exclusively inside the NGMHZ (Supplementary Figures S2, S3). Conversely, densities were consistently low for sandbar and tiger sharks inside the NGMHZ (Supplementary Figures S4, S5). Offshore waters along the coasts of Georgia, South Carolina, and southern North Carolina consistently showed high predicted densities for both sandbar and tiger sharks, but density patterns for those species in waters off the coast of Virginia were variable across years.

Figure 2. Spatial plots of log (density) for the Atlantic stock of blacknose shark, 1989-2021, from the VAST model. Warm tones represent areas of high log(density) while cool tones indicate areas of low log (density).
The area-weighted indices for all species were generally low in the 1990s and early 2000s, but trends differed toward the end of the time series showing three general patterns: a gradual increase, relative consistency, and an increase followed by a decline (Figure 3). Relative abundance trends for blacknose (A.) and sandbar sharks consistently increased through the end of the time series (Figures 3A, F). Area-weighted index values increased from 1991 to 2001 for bull and spinner sharks, but overall, the index values remained close to the mean with minimal directional changes thereafter (Figures 3E, H). Like other species, tiger shark indices increased after a period of low abundance in the 1990s, peaked around 2013, but then showed a subsequent decline (Figure 3J). Overall, estimated uncertainty associated with the indices was high, as evidenced by wide 95% confidence intervals (CIs).
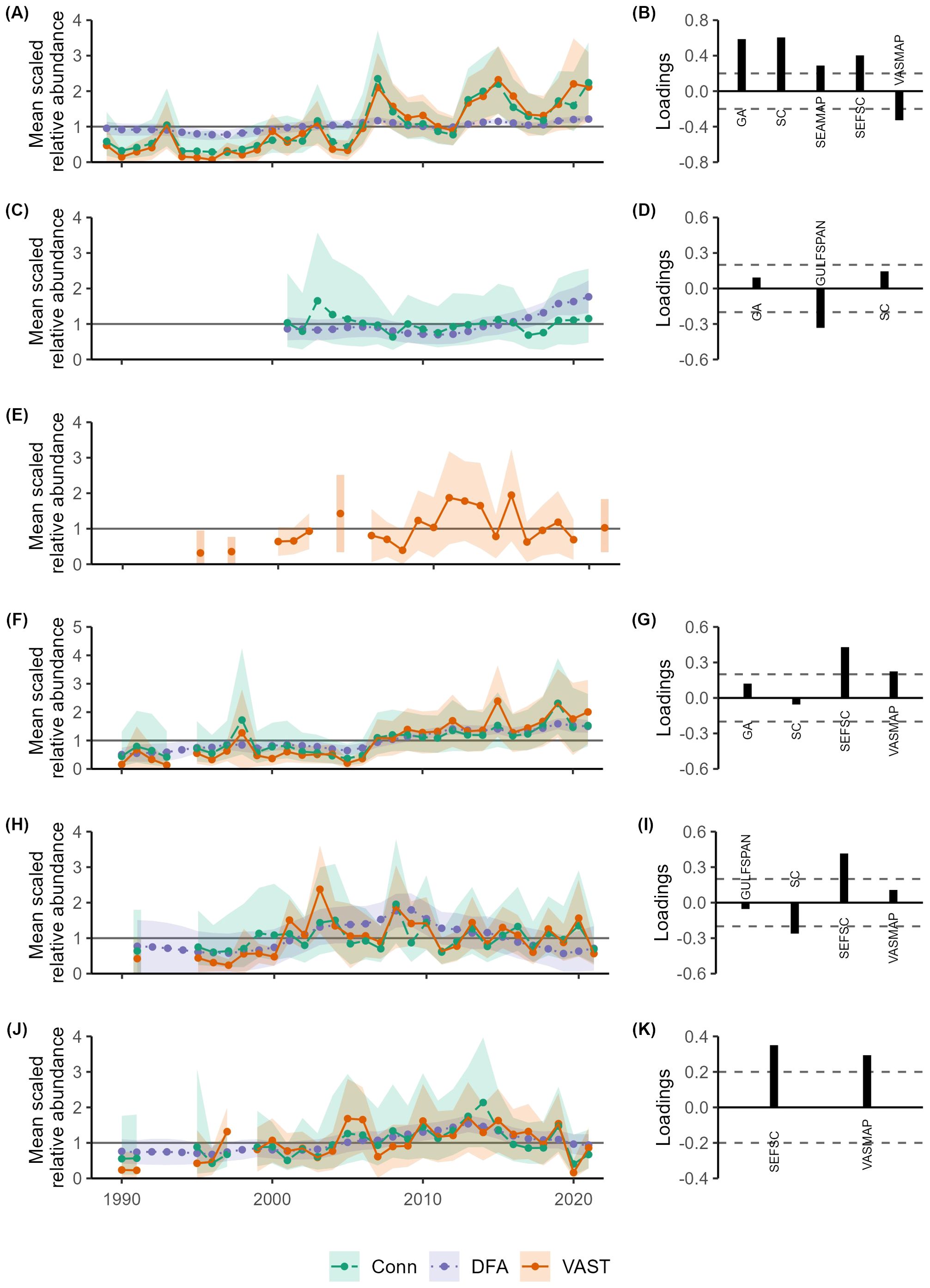
Figure 3. Mean scaled indices of relative abundance estimated using dynamic factor analysis (DFA; purple) and Bayesian hierarchical analysis (Conn; green), and mean scaled area-weighted indices of relative abundance from vector autoregressive spatiotemporal (VAST; orange) models for the Atlantic stock of blacknose shark (A), finetooth shark (C), bull shark (E), sandbar shark (F), spinner shark (H), and tiger shark (J). Trends are represented by dotted, dashed, and solid lines with uncertainty denoted by the shaded regions (95% confidence intervals for DFA and VAST and 95% credible intervals for Conn). Factor loadings for selected DFA models for the Atlantic stock of blacknose shark (B), finetooth shark (D), sandbar shark (G), spinner shark (I), and tiger shark (K) are displayed to the right with dashed lines indicating 0.2. Factor loadings greater than 0.2 correspond to indices that had a relatively strong influence on the resulting common trend, and negative factor loadings denote indices that follow an opposite trend relative to the DFA common trend. Abbreviations include: the Georgia Red Drum Longline Survey (GA), the Gulf of Mexico Shark Pupping and Nursery Gillnet Survey (GULFSPAN), the South Carolina Coastal Longline Survey (SC), the SouthEast Area Monitoring and Assessment Program-South Atlantic Coastal Trawl Survey (SEAMAP), the Southeast Fisheries Science Center Bottom Longline Survey (SEFSC), the Virginia Shark Monitoring and Assessment Program (VASMAP).
Spatial differences over time were evident for all species where spatiotemporal random effects were successfully included in the VAST models (Figures 4; Supplementary Figures S6–S10). The quantile regressions revealed largely significant positive slopes for blacknose (A.) sharks, with the largest estimated values off the coasts of northern Florida, Georgia, South Carolina, and southern North Carolina (Figure 4A). Additionally, density anomalies were consistently positive across the Atlantic, indicating increased density compared to the starting year, and more variable over time, particularly in the Middle region (Supplementary Figures S6A, S7). This area also showed the most substantial increase in density (Figure 5A). For sandbar sharks, areas along coastal Georgia and offshore Florida exhibited consistently positive significant temporal trends (Figure 4B), while areas along North Carolina and Virginia were largely stable with few anomalies (Supplementary Figures S6B, S8). Area-weighted abundance increased more in the Middle and South compared to the North (Figure 5B). Estimated density trends for spinner sharks showed increasingly positive slopes at both the southern and northern extents of the surveyed area (Figure 4C). Notably, largely positive density anomalies were estimated off the coasts of Virginia and North Carolina starting around 2001 (Supplementary Figure S9), while those in the South showed increasing variability (Supplementary Figure S6C). Area-weighted indices were quite variable across all three regions (Figure 5C). For tiger sharks, density trends were steeply positive in the Middle region (Figure 4D), while density anomalies exhibited greater variability in both the Middle and North regions (Supplementary Figures S6D, S10). Area-weighted indices were also larger in the Middle and North compared to the South (Figure 5D).
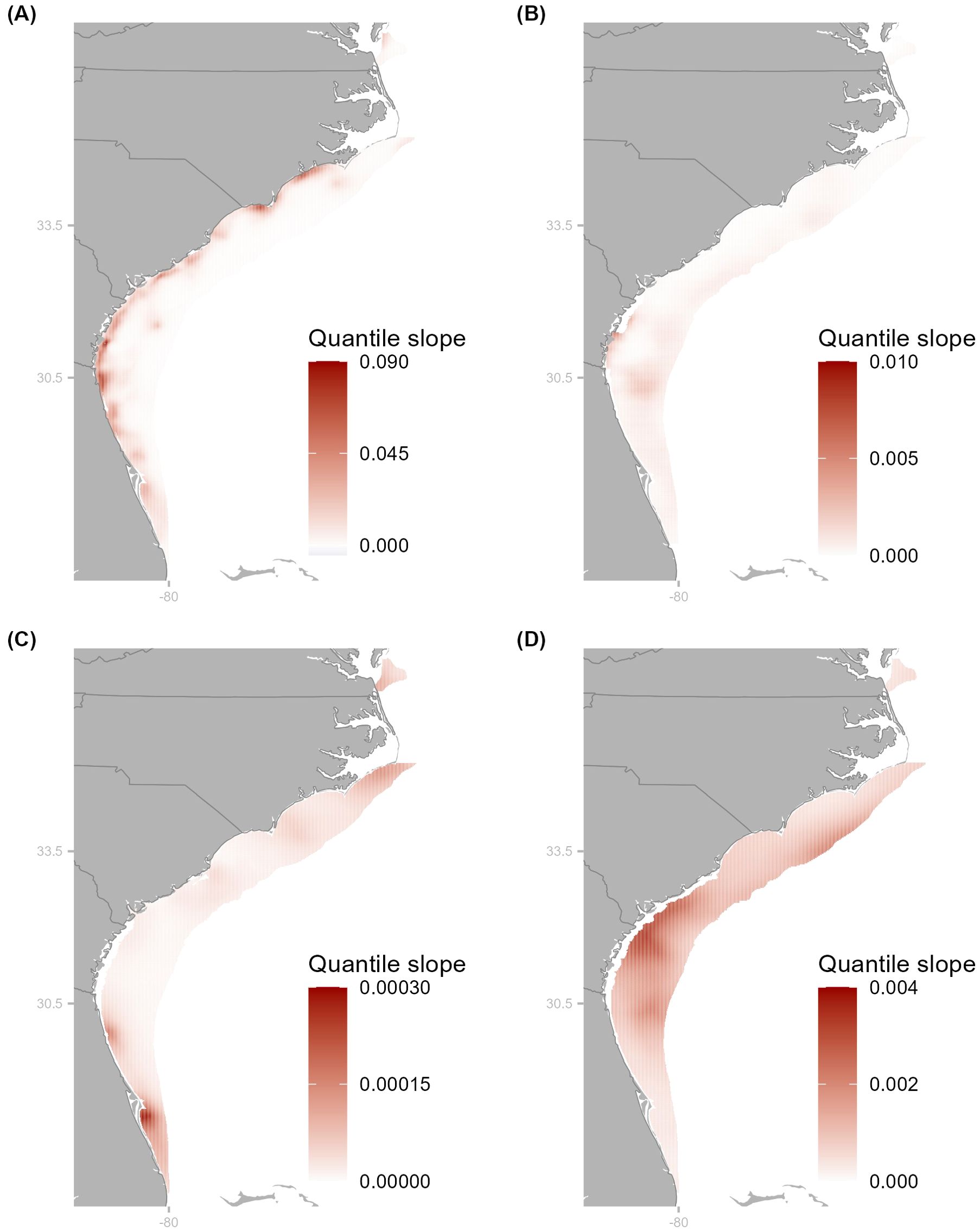
Figure 4. Maps of significant slopes from quantile regression analyses for Atlantic stock of blacknose (A), sandbar (B), spinner (C), and tiger (D) sharks from respective VAST models. Cooler (warmer) colors indicate a negative (positive) slope and the hue denotes the magnitude of the slope with larger values denoted by darker hues. Latitude markers denote where the three sections (South, Middle, and North) of the Atlantic were separated for spatiotemporal analysis.
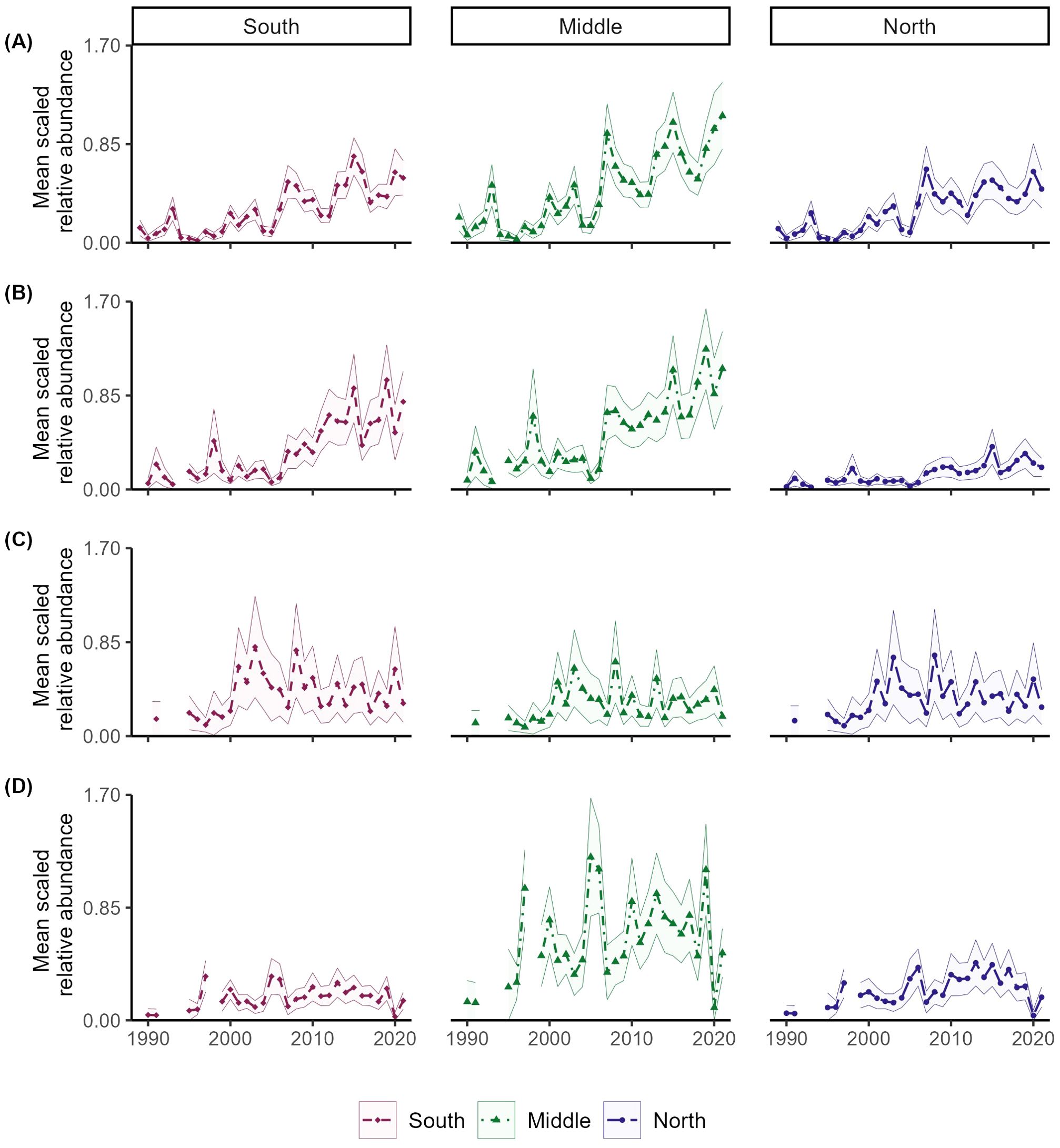
Figure 5. Mean scaled area-weighted indices with 95% confidence intervals for four coastal shark species over three different regions of the Atlantic: South (), Middle (), and North (). Species examined include Atlantic stock of blacknose (A), sandbar (B), spinner (C), and tiger (D) sharks.
3.3 Dynamic factor and Bayesian hierarchical analyses
3.3.1 Indices of abundance
GLMMs parameterized with the negative binomial and zero-inflated Sichel distributions received the most empirical support across species and surveys, followed by the Waring and beta negative binomial distributions (Supplementary Table S3). All models provided acceptable fits to the raw survey data as determined by graphical residual analysis, cross validation, variance inflation factors, and dispersion analysis. Station/area and month were the covariates most frequently included in the supported GLMMs for each species, followed by depth (Supplementary Table S1). For all species, individual survey indices displayed clear data conflict (Supplementary Figure S11), making it difficult to visually interpret broad-scale patterns in species relative abundance.
3.3.2 Dynamic factor analysis
DFA models were successfully fitted to all species (except the bull shark since only one survey was informative), and at least one survey loaded significantly to the common trend (Table 2, Supplementary Table S4; Figures 3B, D, G, I, K). A diagonal covariance structure was empirically supported for all species, which was consistent with previous work that reported no covariance between survey indices (Peterson et al., 2017a).
Common trends estimated for the five species can be divided into two groups. Blacknose (A.), finetooth, and sandbar sharks all displayed low relative abundance in the 1990s and early 2000s and a modest recovery through the terminal year of 2021, though rates of recovery were species-specific (Figures 3A, C, F). In contrast, the common trends for spinner and tiger sharks showed low relative abundance from the beginning of the time series through the early 2000s followed by a modest recovery, but declined after approximately 2009 and 2014, respectively (Figures 3H, J). Except for spinner sharks, trends of indices from the DFA generally agreed with those from VAST, although uncertainty associated with the DFA indices was lower (Figure 3).
The inclusion of covariates was supported in DFA models for all species except sandbar shark. Results indicated statistically significant linkages, at 85% CI, between AMO and GSI for the SCS and LCS species, respectively. Specific to the SCS, the AMO had a significant positive effect on the VASMAP index for blacknose sharks (A.), the GA index for finetooth sharks, and the GULFSPAN index for finetooth sharks (Supplementary Table S4). The GSI had a significant negative effect on the GULFSPAN index and a positive effect on the VASMAP index for spinner sharks, and a positive effect on the VASMAP index for tiger sharks (Supplementary Table S4).
3.3.3 Bayesian hierarchical analysis
Estimated indices from the Conn method were most similar to those predicted using VAST, though the associated uncertainties were slightly larger (Figure 3). Estimated Conn and DFA indices differed for spinner and finetooth sharks, with the DFA indices decreasing or increasing for the two species while the Conn indices showed consistency over time (Figures 3C, H).
Index values were low for all species in the 1990s through the early 2000s but differed near the end of the time series. Both blacknose and sandbar sharks displayed a largely positive trend beginning around 2007 (Figures 3A, F). Conn indices for finetooth and spinner sharks had no discernable trend and some of the largest credible intervals (Figures 3C, H). Tiger shark relative abundance increased until 2014, after which the index declined (Figure 3J). There was little variation in the process error estimates across surveys and species apart from the comparably large value associated with GA index for finetooth shark (Supplementary Figure S12).
4 Discussion
This study is the first to utilize spatiotemporal models to standardize indices for assessed and unassessed coastal shark populations and provide insights into spatial distribution patterns in the U.S. southeast Atlantic and GOM. Analysis of the density trends and anomalies suggested that species’ availability to the surveys may be changing, potentially due to range expansions, distribution shifts, or changes in migration timing. Environmental factors, such as climate change (Chin et al., 2010; Hare et al., 2016) and multidecadal variability (Peterson et al., 2017a; O’Brien et al., 2024), may be impacting coastal shark distributions in similar ways to those documented for teleost fishes (Nye et al., 2009, 2014; Kleisner et al., 2017; Bowers and Kajiura, 2023). Potential changes in catchability within surveys could be changing and should be further investigated and accounted for in future stock assessments used to inform management. The estimated area-weighted indices were comparable to those produced by previous standardization methods (DFA and Conn) and, at minimum, offer an alternative option for sensitivity analyses for future stock assessments.
Density maps for all species modeled using VAST generally aligned with previously published geographic distributions, but the model-based predictions highlighted several areas of interest. Higher bull shark densities are expected in shallower coastal waters near the mouths of estuaries (Drymon et al., 2014; Calich et al., 2018; TinHan and Wells, 2021). However, the presence of higher predicted densities offshore in the GOM may suggest sexual segregation, similar to patterns observed off the coast of Australia (Werry and Clua, 2013), especially given bull sharks’ tolerance to hypoxic conditions (Heithaus et al., 2009). Unfortunately, this could not be further explored due to data limitations.
For blacknose (A.) sharks, results corroborated previous findings that the species inhabited nearshore shallow waters (Ulrich et al., 2007; Castro, 2000), but also revealed areas of higher density in deeper offshore waters off Georgia and South Carolina, which visually align with offshore structures (Crimian and Conley, 2019). In contrast, consistently low densities of sandbar sharks were predicted in these same areas, underscoring the complexity of marine ecosystems and the importance of considering species-specific habitat requirements.
The expansion of hypoxic waters, along with the increasing frequency and intensity of temporal hypoxia, could impact shark abundance and diversity in coastal and shelf environments (Waller et al., 2024). Dissolved oxygen concentrations have influenced multiple SCS species, and changes in the shape and size of the NGMHZ could also influence the distribution and survey availability of LCS species. Despite the estimated overall increase in abundance over time, predicted sandbar shark density remained low in the NGMHZ, likely due to this species’ intolerance of hypoxic conditions (Crear et al., 2019; Latour et al., 2022). In contrast, areas of higher predicted spinner shark density were almost exclusively located within the NGMHZ, potentially attributable to the exploitation of prey aggregations at the edges of hypoxic zones or near the water surface (Craig, 2012; Pickens et al., 2022). More research is necessary to understand how hypoxia may impact other coastal shark species, such as tiger shark, although predicted densities within the NGMHZ were consistently low over time.
Historically, the blacknose (A.) shark inhabited warm nearshore waters, with a northern range limit extending to North Carolina (Ulrich et al., 2007; Castro, 2000). However, results of this study provide indications of a northward range expansion along the Atlantic coast, as evidenced by significant positive trends in spatial density in coastal North Carolina and Virginia waters, increased variation in density anomalies in the Middle and North regions, and a greater increase in abundance in the Middle region. Increased density trends in Northern latitudes coincide with a dramatic shift in catch recorded on the northernmost survey (VASMAP), from just five animals between 1990–2014 to 124 sampled from 2015-2023. This poleward range expansion may be driven by rising water temperatures linked to climate change (O’Brien et al., 2024) or shifts in prey species distributions (Nye et al., 2009; Morley et al., 2018). Although incorporating prey species and habitat effects into a multivariate spatiotemporal model could provide valuable insights into the specific factors influencing this northward migration, such an analysis was beyond the scope of this study. Documenting these range expansions help facilitate modeling of climate change impacts and marine food webs, which is important for resource managers as it allows for the development of targeted conservation measures to ensure sustainable management.
The estimated positive trends of spinner shark density in both the North and South, along with increasing density anomalies over the western North Atlantic Ocean suggests either a range expansion or timing disconnect between the onset of summer migration and sampling activities by the various survey programs. The VAST and Conn models may have captured these dynamics more effectively than the DFA, as indicated by the consistent trends in relative abundance produced by these analyses. Although the Conn model is not spatially explicit, it has been described as robust to assumption violations such as trends in spatial mixing proportions (Conn, 2010). The distribution of spinner sharks extends north of the surveyed area, and the incomplete survey spatial coverage of their full range likely influenced the interpretation of all indices. Spinner shark abundance north of the VASMAP sampling domain has been high during summer months over recent years (T. Curtis; NOAA Fisheries, personal communication). Including data from sampling earlier in the year or from areas near the northern extent of the focal species’ ranges would help clarify whether the discrepancies between indices are due to changes in survey catchabilities or a genuine decline in abundance.
The tiger shark decline in abundance towards the end of the time series was unexpected since it contradicts previous work indicating early signs of population recovery (Peterson et al., 2017a). Recent studies have suggested that climate change and multidecadal variability may be shifting the timing of summer and overwintering migrations for tiger sharks (Hammerschlag et al., 2022; O’Brien et al., 2024). The positive trends in density estimated for the Middle and North regions, along with increased variability in density anomalies along the western North Atlantic could indicate an earlier onset of migration. This potential timing discrepancy could significantly affect the availability of tiger sharks to the SEFSC survey, which annually samples in the Atlantic from July-August. Such misalignment could lead to potential misinterpretations of population trends that are ultimately used to support management decisions. Lastly, tiger shark populations may be declining given increased mortality on neonate and juveniles due to increased predation rates from recovering LCS species (e.g., sandbar sharks; W. Driggers; NOAA Fisheries, personal communication). Expanding monitoring efforts to earlier times in the year or incorporating additional data sources (e.g., fisheries-dependent surveys) could aid in interpreting abundance trends for tiger shark.
Increased indices through the end of the time series across all standardization methods support previous findings that the sandbar shark population is still recovering (Carlson et al., 2012; Peterson et al., 2017a, 2022). However, while density trends were consistent in the North, they showed an increase in the Middle and South regions. Additionally, area-weighted indices were lower in the North compared to the Middle and South, suggesting a potential southward shift in the population. Although there is less highly suitable habitat for large sandbar sharks in the northern extent of their range during positive NAO phases (Peterson et al., 2017a; O’Brien et al., 2024), other potential explanations for the southward shift could be changes in the size and age composition and/or migration timing. The size and age composition has likely shifted to smaller and younger individuals following the period of overexploitation, and these sizes and ages, particularly males, tend to inhabit in the warmer waters off the east coast of Florida, Georgia, and South Carolina during the summer and fall months, rather than migrating north like gravid mature females (Grubbs, 2010; Collatos et al., 2020; Baremore and Hale, 2012). Sandbar sharks also migrate farther north than the VASMAP survey, where abundance has been high during summer months over recent years (B. Frazier; SCDNR, personal communication). Additionally, migration timing for sandbar sharks may also be changing, which could affect the catches of gravid females during their northward migration to pupping grounds, though this hypothesis has not been comprehensively investigated. Future application of multivariate spatiotemporal modeling techniques to distinct age and size classes would aid exploration of distributional patterns through ontogeny.
Multidecadal climate variability may intensify or counteract shifts in the distribution of coastal shark populations (Peterson et al., 2017a; O’Brien et al., 2024). The AMO was a significant covariate in the DFA analyses of both blacknose (A.) and finetooth sharks. A warm AMO phase, which is associated with increased rainfall in Florida and heightened hurricane activity in the Atlantic, was correlated with above average blacknose (A.) shark abundance off the Virginia coast, likely due to increased water temperatures along the southeastern U.S. coast. Increased abundance of finetooth sharks in the nursery and nearshore areas sampled by the GA and GULFSPAN surveys was also associated with a warm AMO phase and could indicate higher habitat utilization (Carlson et al., 2003; Carpenter, 2017). Gene flow in finetooth sharks, as well as other SCS, is limited around the southern tip of Florida due to relatively localized movement patterns, and the GOM and southeast Atlantic represent two separate stocks (Portnoy et al., 2016; Kohler and Turner, 2019; Vinyard et al., 2019). Positive GSI values, which indicate a northern shift in the Gulf Stream, were associated with increased abundance of spinner and tiger sharks off the Virginia coast, potentially due to increased water temperature or abundance of tropical and sub-tropical prey species. Conversely, recruitment and habitat utilization may decline for spinner sharks in the northeastern GOM during positive GSI as indicated by the decreased indices for GULFSPAN. NAO was previously associated with a decreased abundance of sandbar sharks off the coast of Virginia (Peterson et al., 2017a) but was not found to be significant in this study. This discrepancy could be attributed to the different starting years and increased sample size, environmental changes, shifts in population demographics, or variations in migration timing.
It is important to note that the SEFSC survey data had the most influence on the estimated abundance indices because of its comparably large spatial footprint. Therefore, VAST model results should be interpreted relative to the SEFSC survey characteristics. For instance, the survey does not sample in waters shallower than 9 m where important juvenile bull shark and adult finetooth shark habitat is located (Carlson et al., 2003; TinHan and Wells, 2021). The notably low tiger shark index value for 2020 is likely due to COVID-19 and the resulting significantly reduced sampling caused by the pandemic. Peak abundance estimates for spinner (2001 and 2003) and blacknose (A.) sharks (2007) correspond with years when the SEFSC did not sample the Atlantic, while abnormal indices for 2005 are likely due to Hurricane Katrina disrupting sampling schedules. Previous studies have highlighted the value of incorporating multiple data sources and different data types (e.g., encounter/non-encounter, fishery-dependent, etc.), to better capture true trends when unforeseen circumstances impact fishery-independent sampling (Thorson and Barnett, 2017; Grüss and Thorson, 2019; Grüss et al., 2023a). Despite these challenges, the analyses presented here are based on the best available fisheries-independent data.
While species-specific trends in abundance were generally consistent across methodologies (except for spinner and finetooth sharks), each index standardization method used in this study offers distinct advantages and disadvantages for stock assessment applications. A key advantage of the VAST model is that it simultaneously standardizes data and integrates information across multiple surveys within a unified spatiotemporal framework, eliminating the need to pre-standardize indices for distinct sampling programs prior to model fitting. The VAST model also incorporates spatial and spatiotemporal correlations, allowing for improved estimation of abundance and density surfaces that account for shifting distributions and variable survey effort (Grüss and Thorson, 2019; Grüss et al., 2023a). Additionally, the spatial output provided by VAST can aid in the interpretation of indices with respect to changes in survey catchability, which can bias stock assessments and negatively affect management advice (Peltonen et al., 1999). This is particularly valuable for mobile or wide-ranging species whose availability to surveys varies over time (Morgan et al., 2020; O’Brien et al., 2024). However, VAST and other spatiotemporal modeling frameworks (e.g., sdmTMB) require relatively rich datasets, are computationally intensive, and may not converge reliably for sparse or spatially fragmented datasets (i.e., finetooth shark), limiting their utility in some cases (Thorson, 2019; Anderson et al., 2024). Uncertainty associated with VAST was also greater, as indicated by larger 95% CIs. While this may reflect true ecological and sampling variability, greater uncertainty can ultimately influence stock assessment model output and total allowable catch (TAC) levels that incorporate buffers for scientific uncertainty (Maunder and Piner, 2015; Bi et al., 2022).
Although the Conn model is not spatially explicit, it is generally more robust to violations of assumptions such as constant spatial structure or stable catchability. Notably, Conn performed well in cases involving multiple spatially fragmented surveys (e.g., finetooth shark), where the VAST model failed to converge. However, 95% CIs for the Conn model were comparable to those estimated by VAST and notably greater than those estimated by DFA. The DFA method produced smoother relative abundance trajectories, yielded lower uncertainty bounds compared to the other approaches, and also successfully incorporated multiple spatially fragmented surveys, but may underestimate uncertainty when spatial or temporal variation in catchability are not explicitly modeled (Zuur et al., 2003a, 2003b, Peterson et al., 2021b). A key advantage of DFA is its ability to interpolate across occasional missing data, although this should be done sparingly and the results for years with minimal to no sampling – such as 2020 – should be interpreted with caution (Peterson et al., 2021a, 2021b).
Scientific uncertainty—whether arising from natural variability, observation error, or model structure—is an inherent aspect of stock assessment and should be explicitly considered in management advice. Identifying changes in catchability and survey efficiency is important for developing robust science-based fisheries management policies, and long-term fishery-independent survey programs remain the best available data source for most species, particularly given the known limitations of commercial catch data (Burgess et al., 2005). Although the Conn and VAST models yielded higher estimated uncertainties, they may be more robust than DFA to shifts in species distribution or migration timing that alter survey catchability (Peterson et al., 2021b). Ultimately, the choice of standardization method should reflect the species’ spatial ecology, the survey design and coverage, and the trade-offs between model complexity, robustness, and decision-making risk.
Climate change may impact catchability of fishery-independent surveys targeting coastal sharks by altering migration timing and shifts or expansions in distribution thus posing a significant challenge for their management. VAST spatiotemporal models have emerged as a promising tool for index standardization in fisheries management, demonstrating their potential for analyzing complex spatiotemporal patterns in coastal shark population abundance. However, VAST may not be suitable for all species or scenarios due to data constraints and model convergence issues. In such cases, index derived standardization methods, such as DFA and Conn, provide similar results and may be better suited for analyzing basin-wide abundance patterns.
Overall, the recovery of coastal shark populations may not be as optimistic as previously indicated, as only two of the six stocks examined showed increasing trends across all three index methods at the end of the time series. Spatial outputs from VAST provided evidence of distribution shifts among these stocks, but further investigation is needed to determine whether trends in indices reflect true changes in abundance or shifts in survey catchability due to spatial movements or migration timing. Despite these uncertainties, VAST models offer valuable insights that can support adaptive management strategies, such as spatial management measures, to effectively respond to the dynamic nature of coastal shark populations in a changing environment.
Data availability statement
The data analyzed in this study is subject to the following licenses/restrictions: Catch data from GA and SEAMAP-SA Coastal Trawl Survey are publicly accessible online with access information provided in the main text. Catch data from GULFSPAN, SC, SEFSC, and VASMAP are available from the respective authors upon reasonable request. Requests to access these datasets should be directed to: SEFSC - William Driggers, d2lsbGlhbS5kcmlnZ2Vyc0Bub2FhLmdvdg==; SC - Bryan Frazier, RnJhemllckJAZG5yLnNjLmdvdg==; VASMAP - Robert Latour, bGF0b3VyQHZpbXMuZWR1; GULFSPAN - John Carlson, am9obi5jYXJsc29uQG5vYWEuZ292.
Ethics statement
The animal study was approved by various groups for the different surveys considered, with each survey in this study complying with U.S. Animal Welfare Act laws, guidelines, and policies as approved by NOAA Fisheries. The SEFSC, SEAMAP-SA, and GULFSPAN surveys do not require ethical approval in accordance with institutional requirements because research was covered under federally authorized permits. Sharks tagged by VASMAP were approved by William & Mary Institutional Animal Care and Use Committee (IACUC). Sampling conducted in South Carolina complied with the SC Code of Law: Section 50-5-20. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
KO: Conceptualization, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Visualization, Writing – original draft, Writing – review & editing, Data curation. JC: Data curation, Writing – review & editing, Conceptualization, Investigation, Resources. EC: Conceptualization, Writing – review & editing, Methodology, Investigation. WD: Data curation, Writing – review & editing, Conceptualization, Investigation, Resources. BF: Data curation, Writing – review & editing, Conceptualization, Investigation, Resources. RL: Conceptualization, Data curation, Funding acquisition, Investigation, Supervision, Writing – review & editing, Methodology, Resources.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This research was made possible by the VIMS Commonwealth Coastal Research Fellowship, as well as a Virginia Sea Grant Professional Development Grant #V724500. We acknowledge the SEAMAP-SA Data Management Work Group for providing the GA and SEAMAP-SA Coastal Trawl Survey data. Recent funding for the VASMAP survey was provided by the Atlantic States Marine Fisheries Commission Agency Award #22-0901 and NOAA, US Department of Commerce Agency Award #NA22NMF4540361. Funding for SC has been provided by the SEAMAP-SA and the South Carolina Saltwater Recreational Fishing License Funds.
Acknowledgments
We would like to acknowledge the many unnamed staff, researchers, and volunteers associated with the various surveys for their efforts in collecting the data and maintaining these valuable sampling programs.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2025.1621720/full#supplementary-material
References
Akaike H. (1973). Maximum likelihood identification of Gaussian autoregressive moving average models. Biometrika 60, 255–265. doi: 10.1093/biomet/60.2.255
Anderson S. C., Ward E. J., English P. A., Barnett L. A. K., and Thorson J. T. (2024). sdmTMB: an R package for fast, flexible, and user-friendly generalized linear mixed effects models with spatial and spatiotemporal random fields 2022.03.24.485545. doi: 10.1101/2022.03.24.485545
Baremore I. E. and Hale L. F. (2012). Reproduction of the sandbar shark in the western north atlantic ocean and Gulf of Mexico. Mar. Coast. Fisheries 4, 560–572. doi: 10.1080/19425120.2012.700904
Baum J. K. and Blanchard W. (2010). Inferring shark population trends from generalized linear mixed models of pelagic longline catch and effort data. Fisheries Res. 102, 229–239. doi: 10.1016/j.fishres.2009.11.006
Baum J. K., Myers R. A., Kehler D. G., Worm B., Harley S. J., and Doherty P. A. (2003). Collapse and conservation of shark populations in the northwest atlantic. Science 299, 389–392. doi: 10.1126/science.1079777
Beltz B. (2024). NOAA-EDAB/ecodata. Available online at: https://github.com/NOAA-EDAB/ecodata (Accessed January 15, 2024).
Bi R., Collier C., Mann R., Mills K. E., Saba V., Wiedenmann J., et al. (2022). How consistent is the advice from stock assessments? Empirical estimates of inter-assessment bias and uncertainty for marine fish and invertebrate stocks. doi: 10.1111/faf.12714
Birkmanis C. A., Freer J. J., Simmons L. W., Partridge J. C., and Sequeira A. M. M. (2020). Future distribution of suitable habitat for pelagic sharks in Australia under climate change models. Front. Mar. Sci. 7. doi: 10.3389/fmars.2020.00570
Bolker B. M., Brooks M. E., Clark C. J., Geange S. W., Poulsen J. R., Stevens M. H. H., et al. (2009). Generalized linear mixed models: a practical guide for ecology and evolution. Trends Ecol. Evol. 24, 127–135. doi: 10.1016/j.tree.2008.10.008
Bowers M. and Kajiura S. (2023). A critical evaluation of adult blacktip shark, Carcharhinus limbatus, distribution off the United States East Coast. Environ. Biol. Fishes 106, 1–17. doi: 10.1007/s10641-023-01449-3
Britten G. L., Dowd M., Minto C., Ferretti F., Boero F., and Lotze H. K. (2014). Predator decline leads to decreased stability in a coastal fish community. Ecol. Lett. 17, 1518–1525. doi: 10.1111/ele.12354
Burgess G., Beerkircher L., Cailliet G., Carlson J., Cortés E., Goldman K., et al. (2005). Reply to “Robust estimates of decline for pelagic shark populations in the Northwest Atlantic and Gulf of Mexico.“. Fisheries 30, 30–31. doi: 10.1577/1548-8446(2005)30[19:ITCOSP]2.0.CO;2
Calich H., Estevanez M., and Hammerschlag N. (2018). Overlap between highly suitable habitats and longline gear management areas reveals vulnerable and protected regions for highly migratory sharks. Mar. Ecol. Prog. Ser. 602, 183–195. doi: 10.3354/meps12671
Cao J., Thorson J. T., Richards R. A., and Chen Y. (2017). Spatiotemporal index standardization improves the stock assessment of northern shrimp in the Gulf of Maine. Can. J. Fish. Aquat. Sci. 74, 1781–1793. doi: 10.1139/cjfas-2016-0137
Carlson J. K., Cortés E., and Bethea D. M. (2003). Life history and population dynamics of the finetooth shark (Carcharhinus isodon) in the northeastern Gulf of Mexico. Available online at: https://aquadocs.org/handle/1834/30976 (Accessed November 3, 2022).
Carlson J. K., Hale L. F., Morgan A., and Burgess G. H. (2012). Relative abundance and size of coastal sharks derived from commercial shark longline catch and effort data. J. Fish Biol. 80, 1749–1764. doi: 10.1111/j.1095-8649.2011.03193.x
Carpenter J. (2017). Survey Gear Comparisons and Shark Nursery Habitat Use in Southeast Georgia Estuaries. Available online at: https://digitalcommons.unf.edu/etd/731 (Accessed April 22, 2024).
Castro J. I. (2000). Biology of the Nurse Shark, Ginglymostoma cirratum, Off the Florida East Coast and the Bahama Islands. Environ. Biol. Fishes 58, 1–22. doi: 10.1023/A:1007698017645
Chin A., Kyne P. M., Walker T. I., and McAuley R. B. (2010). An integrated risk assessment for climate change: analysing the vulnerability of sharks and rays on Australia’s Great Barrier Reef. Global Change Biol. 16, 1936–1953. doi: 10.1111/j.1365-2486.2009.02128.x
Collatos C., Abel D. C., and Martin K. L. (2020). Seasonal occurrence, relative abundance, and migratory movements of juvenile sandbar sharks, Carcharhinus plumbeus, in Winyah Bay, South Carolina. Environ. Biol. Fish 103, 859–873. doi: 10.1007/s10641-020-00989-2
Conn P. B. (2010). Hierarchical analysis of multiple noisy abundance indices. Can. J. Fish. Aquat. Sci. 67, 108–120. doi: 10.1139/F09-175
Cortés E., Brooks E. N., and Gedamke T. (2012). “Population Dynamics, Demography, and Stock Assessments,” in Biology of Sharks and their Relatives, 453–477.
Cortés E., Brooks E. N., and Shertzer K. W. (2015). Risk assessment of cartilaginous fish populations. ICES J. Mar. Sci. 72, 1057–1068. doi: 10.1093/icesjms/fsu157
Craig K. (2012). Aggregation on the edge: Effects of hypoxia avoidance on the spatial distribution of brown shrimp and demersal fishes in the Northern Gulf of Mexico. Mar. Ecol. Prog. Ser. 445, 75–95. doi: 10.3354/meps09437
Crear D. P., Brill R. W., Bushnell P. G., Latour R. J., Schwieterman G. D., Steffen R. M., et al. (2019). The impacts of warming and hypoxia on the performance of an obligate ram ventilator. Conserv. Physiol. 7. doi: 10.1093/conphys/coz026
Crimian R. L. and Conley M. F. (2019). Coastal Georgia recreational use mapping project: summary report. Nat. Conservancy.
Diaz-Carballido P. L., Mendoza-González G., Yañez-Arenas C. A., and Chiappa-Carrara X. (2022). Evaluation of shifts in the potential future distributions of carcharhinid sharks under different climate change scenarios. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.745501
Drymon J. M., Ajemian M. J., and Powers S. P. (2014). Distribution and dynamic habitat use of young bull sharks carcharhinus leucas in a highly stratified northern gulf of Mexico estuary. PloS One 9, e97124. doi: 10.1371/journal.pone.0097124
Dulvy N. and Forrest R. (2010). “Life Histories, Population Dynamics, and Extinction Risks in Chondrichthyans,” in Sharks and their Relatives II: Biodiversity, Adaptive Physiology, and Conservation (Taylor and Francis Group), 639–679.
Dulvy N. K., Fowler S. L., Musick J. A., Davidson L. N., Fordham S. V., Francis M. P., et al. (2014). Extinction risk and conservation of the world’s sharks and rays. eLife 3. doi: 10.7554/eLife.00590
Efron B. and Tibshirani R. J. (1993). An Introduction to the Bootstrap (Boston, MA: Springer US). doi: 10.1007/978-1-4899-4541-9
Ellis J., Clarke M., Cortés E., Heessen H. J. L., Apostolaki P., Carlson J. K., et al. (2009). Management of elasmobranch fisheries in the North Atlantic. Adv. fisheries science: 50 years Beverton Holt. doi: 10.1002/9781444302653.ch9
Ferretti F., Worm B., Britten G. L., Heithaus M. R., and Lotze H. K. (2010). Patterns and ecosystem consequences of shark declines in the ocean. Ecol. Lett. 13, 1055–1071. doi: 10.1111/j.1461-0248.2010.01489.x
Francis R. I. C. C. (2011). Data weighting in statistical fisheries stock assessment models. Can. J. Fish. Aquat. Sci. 68, 1124–1138. doi: 10.1139/f2011-025
Goodman M. C., Carroll G., Brodie S., Grüss A., Thorson J. T., Kotwicki S., et al. (2022). Shifting fish distributions impact predation intensity in a sub-Arctic ecosystem. Ecography 2022, e06084. doi: 10.1111/ecog.06084
Grubbs R. D. (2010). “Ontogenetic Shifts in Movements and Habitat Use,” in Sharks and their relatives II: Biodiversity, adaptive physiology, and conservation, 319–350. doi: 10.1201/9781420080483-c7
Grüss A., Charsley A. R., Thorson J. T., Anderson O. F., O’Driscoll R. L., Wood B., et al. (2023a). Integrating survey and observer data improves the predictions of New Zealand spatio-temporal models. ICES J. Mar. Sci. 80, 1991–2007. doi: 10.1093/icesjms/fsad129
Grüss A. and Thorson J. T. (2019). Developing spatio-temporal models using multiple data types for evaluating population trends and habitat usage. ICES J. Mar. Sci. 76, 1748–1761. doi: 10.1093/icesjms/fsz075
Grüss A., Thorson J. T., Anderson O. F., O’Driscoll R. L., Heller-Shipley M., and Goodman S. (2023b). Spatially varying catchability for integrating research survey data with other data sources: case studies involving observer samples, industry-cooperative surveys, and predators as samplers. Can. J. Fish. Aquat. Sci. 80, 1595–1615. doi: 10.1139/cjfas-2023-0051
Hammerschlag N., Luo J., Irschick D. J., and Ault J. S. (2012). A Comparison of Spatial and Movement Patterns between Sympatric Predators: Bull Sharks (Carcharhinus leucas) and Atlantic Tarpon (Megalops atlanticus). PloS One 7, e45958. doi: 10.1371/journal.pone.0045958
Hammerschlag N., McDonnell L. H., Rider M. J., Street G. M., Hazen E. L., Natanson L. J., et al. (2022). Ocean warming alters the distributional range, migratory timing, and spatial protections of an apex predator, the tiger shark (Galeocerdo cuvier). Global Change Biol. 28, 1990–2005. doi: 10.1111/gcb.16045
Hansell A. C. and McManus M. C. (2025). Integrating fisheries independent surveys to account for the spatiotemporal dynamics of spiny dogfish (Squalus acanthias) in US waters of the northwest Atlantic. Fisheries Res. 281, 107173. doi: 10.1016/j.fishres.2024.107173
Hare J. A., Morrison W. E., Nelson M. W., Stachura M. M., Teeters E. J., Griffis R. B., et al. (2016). A vulnerability assessment of fish and invertebrates to climate change on the northeast U.S. Continental shelf. PloS One 11, e0146756. doi: 10.1371/journal.pone.0146756
Hartig F. and Lohse L. (2022). DHARMa: Residual Diagnostics for Hierarchical (Multi-Level / Mixed) Regression Models. Available online at: https://cran.r-project.org/web/packages/DHARMa/index.html (Accessed October 23, 2023).
Heithaus M. R., Delius B. K., Wirsing A. J., and Dunphy-Daly M. M. (2009). Physical factors influencing the distribution of a top predator in a subtropical oligotrophic estuary. Limnology Oceanography 54, 472–482. doi: 10.4319/lo.2009.54.2.0472
Heithaus M. R., Frid A., Wirsing A. J., and Worm B. (2008). Predicting ecological consequences of marine top predator declines. Trends Ecol. Evol. 23, 202–210. doi: 10.1016/j.tree.2008.01.003
Hilborn R. and Walters C. J. (Eds.) (1992). Quantitative Fisheries Stock Assessment: Choice, Dynamics and Uncertainty (New York, NY: Springer US). doi: 10.1007/978-1-4615-3598-0
Holmes E. E., Ward E. J., and Scheuerell M. D. (2021). Analysis of multivariate time series using the MARSS package. Version 3.11.4. Seattle, WA: NOAA Fisheries, Northwest Fisheries Science Center. doi: 10.5281/ZENODO.5781847
Holmes E. E., Ward E. J., Scheuerell M. D., and Wills K. (2023). MARSS: Multivariate Autoregressive State-Space Modeling. Available online at: https://cran.r-project.org/web/packages/MARSS/index.html (Accessed October 18, 2023).
Hoyle S. D., Campbell R. A., Ducharme-Barth N. D., Grüss A., Moore B. R., Thorson J. T., et al. (2024). Catch per unit effort modelling for stock assessment: A summary of good practices. Fisheries Res. 269, 106860. doi: 10.1016/j.fishres.2023.106860
ICCAT (2012). “Report of the 2012 ICCAT Working Group on Stock Assessment Methods Meeting (WGSAM),” in Meeting of the ICCAT Working Group on Stock Assessment(Madrid, Spain).
Kleisner K. M., Fogarty M. J., McGee S., Hare J. A., Moret S., Perretti C. T., et al. (2017). Marine species distribution shifts on the U.S. Northeast Continental Shelf under continued ocean warming. Prog. Oceanography 153, 24–36. doi: 10.1016/j.pocean.2017.04.001
Koenker R., Portnoy S., Tain Ng P., Melly B., Zeileis A., Grosjean P., et al. (2024). quantreg: Quantile Regression. Available online at: https://cran.r-project.org/web/packages/quantreg/index.html (Accessed July 13, 2024).
Kohler N. E. and Turner P. A. (2019). Distribution and movements of atlantic shark species: A 52-year retrospective atlas of mark and recapture data. Mar. Fisheries Rev. 81, 1–93. doi: 10.7755/MFR.81.2.1
Kristensen K., Bell B., Skaug H., Magnusson A., Berg C., Nielsen A., et al. (2023). TMB: Template Model Builder: A General Random Effect Tool Inspired by “ADMB.“. Available online at: https://cran.r-project.org/web/packages/TMB/index.html (Accessed October 23, 2023).
Latour R. J., Gartland J., and Peterson C. D. (2022). Ontogenetic niche structure and partitioning of immature sandbar sharks within the Chesapeake Bay nursery. Mar. Biol. 169, 76. doi: 10.1007/s00227-022-04066-3
Løkkeborg S., Fernö A., and Humborstad O.-B. (2010). “Fish Behavior in Relation to Longlines,” in Behavior of Marine Fishes (John Wiley & Sons, Ltd), 105–141. doi: 10.1002/9780813810966.ch5
Manz M. H., Shipley O. N., Cerrato R. M., Hueter R. E., Newton A. L., Tyminski J. P., et al. (2025). Predictions of southern migration timing in coastal sharks under future ocean warming. Conserv. Biol., e70080. doi: 10.1111/cobi.70080
Martin T. G., Wintle B. A., Rhodes J. R., Kuhnert P. M., Field S. A., Low-Choy S. J., et al. (2005). Zero tolerance ecology: improving ecological inference by modelling the source of zero observations. Ecol. Lett. 8, 1235–1246. doi: 10.1111/j.1461-0248.2005.00826.x
Maunder M. N. and Piner K. R. (2015). Contemporary fisheries stock assessment: many issues still remain. ICES J. Mar. Sci. 72, 7–18. doi: 10.1093/icesjms/fsu015
Maunder M. N., Sibert J. R., Fonteneau A., Hampton J., Kleiber P., and Harley S. J. (2006). Interpreting catch per unit effort data to assess the status of individual stocks and communities. ICES J. Mar. Sci. 63, 1373–1385. doi: 10.1016/j.icesjms.2006.05.008
Morgan A., Calich H., Sulikowski J., and Hammerschlag N. (2020). Evaluating spatial management options for tiger shark (Galeocerdo cuvier) conservation in US Atlantic Waters. ICES J. Mar. Sci. 77, 3095–3109. doi: 10.1093/icesjms/fsaa193
Morley J. W., Selden R. L., Latour R. J., Frölicher T. L., Seagraves R. J., and Pinsky M. L. (2018). Projecting shifts in thermal habitat for 686 species on the North American continental shelf. PloS One 13, e0196127. doi: 10.1371/journal.pone.0196127
Musick J. (1999). Life in the Slow Lane: Ecology and Conservation of Long-Lived Marine Animals (American Fisheries Society). doi: 10.47886/9781888569155
Niella Y., Smoothey A. F., Peddemors V., and Harcourt R. (2020). Predicting changes in distribution of a large coastal shark in the face of the strengthening East Australian Current. Mar. Ecol. Prog. Ser. 642, 163–177. doi: 10.3354/meps13322
NMFS (1993). Fishery Management Plan for Sharks of the Atlantic Ocean (Silver Springs: U.S. department of Commerce).
Nye J. A., Baker M. R., Bell R., Kenny A., Kilbourne K. H., Friedland K. D., et al. (2014). Ecosystem effects of the atlantic multidecadal oscillation. J. Mar. Syst. 133, 103–116. doi: 10.1016/j.jmarsys.2013.02.006
Nye J. A., Link J. S., Hare J. A., and Overholtz W. J. (2009). Changing spatial distribution of fish stocks in relation to climate and population size on the Northeast United States continental shelf. Mar. Ecol. Prog. Ser. 393, 111–129. doi: 10.3354/meps08220
O’Brien K. A., Cortés E., Driggers W. B. III, Frazier B. S., and Latour R. J. (2024). Niche structure and habitat shifts for coastal sharks of the US Southeast Atlantic and Gulf of Mexico. Fisheries Oceanography 33, e12676. doi: 10.1111/fog.12676
O’Leary C. A., Thorson J. T., Ianelli J. N., and Kotwicki S. (2020). Adapting to climate-driven distribution shifts using model-based indices and age composition from multiple surveys in the walleye pollock (Gadus chalcogrammus) stock assessment. Fisheries Oceanography 29, 541–557. doi: 10.1111/fog.12494
Osgood G. J., White E. R., and Baum J. K. (2021). Effects of climate-change-driven gradual and acute temperature changes on shark and ray species. J. Anim. Ecol. 90, 2547–2559. doi: 10.1111/1365-2656.13560
Papastamatiou Y. P., Meyer C. G., Carvalho F., Dale J. J., Hutchinson M. R., and Holland K. N. (2013). Telemetry and random-walk models reveal complex patterns of partial migration in a large marine predator. Ecology 94, 2595–2606. doi: 10.1890/12-2014.1
Peltonen H., Ruuhijärvi J., Malinen T., and Horppila J. (1999). Estimation of roach (Rutilus rutilus (L.)) and smelt (Osmerus eperlanus (L.)) stocks with virtual population analysis, hydroacoustics and gillnet CPUE. 44, 25–36.
Perretti C. T. and Thorson J. T. (2019). Spatio-temporal dynamics of summer flounder (Paralichthys dentatus) on the Northeast US shelf. Fisheries Res. 215, 62–68. doi: 10.1016/j.fishres.2019.03.006
Perry A. L., Low P. J., Ellis J. R., and Reynolds J. D. (2005). Climate change and distribution shifts in marine fishes. Science 308, 1912–1915. doi: 10.1126/science.1111322
Peterson C. D., Belcher C. N., Bethea D. M., Driggers W. B., Frazier B. S., and Latour R. J. (2017a). Preliminary recovery of coastal sharks in the south-east United States. Fish Fish 18, 845–859. doi: 10.1111/faf.12210
Peterson C. D., Courtney D. L., Cortés E., and Latour R. J. (2021a). Reconciling conflicting survey indices of abundance prior to stock assessment. ICES J. Mar. Sci. 78, 3101–3120. doi: 10.1093/icesjms/fsab179
Peterson C. D., Gartland J., and Latour R. J. (2017b). Novel use of hook timers to quantify changing catchability over soak time in longline surveys. Fisheries Res. 194, 99–111. doi: 10.1016/j.fishres.2017.05.010
Peterson C. D., Wilberg M. J., Cortés E., Courtney D. L., and Latour R. J. (2022). Effects of unregulated international fishing on recovery potential of the sandbar shark within the southeastern United States. Can. J. Fish. Aquat. Sci. 79, 1497–1513. doi: 10.1139/cjfas-2021-0345
Peterson C. D., Wilberg M. J., Cortés E., and Latour R. J. (2021b). Dynamic factor analysis to reconcile conflicting survey indices of abundance. ICES J. Mar. Science. doi: 10.1093/icesjms/fsab051
Pickens B. A., Taylor J. C., Campbell M. D., and Iii W. B. D. (2022). Offshore snapper and shark distributions are predicted by prey and area of nearby estuarine environments in the Gulf of Mexico, USA. Mar. Ecol. Prog. Ser. 682, 169–189. doi: 10.3354/meps13925
Pilling G., Apostolaki P., Failler P., Floros C., Large P., Morales-Nin B., et al. (2008). “Assessment and Management of Data-Poor Fisheries,” in Advances in Fisheries Science: 50 years on from Beverton and Holt, 280–305. doi: 10.1002/9781444302653.ch12
Portnoy D. S., Hollenbeck C. M., Bethea D. M., Frazier B. S., Gelsleichter J., and Gold J. R. (2016). Population structure, gene flow, and historical demography of a small coastal shark (Carcharhinus isodon) in US waters of the Western Atlantic Ocean. ICES J. Mar. Sci. 73, 2322–2332. doi: 10.1093/icesjms/fsw098
Quinlan J. A. (2023). Results from the gulf of Mexico climate vulnerability analysis for fishes and invertebrates. doi: 10.25923/5SVF-SE47
Rosa R., Rummer J. L., and Munday P. L. (2017). Biological responses of sharks to ocean acidification. Biol. Lett. 13, 20160796. doi: 10.1098/rsbl.2016.0796
SEDAR (2006). “Large Coastal Shark Complex, Blacktip and Sandbar Shark,” in Highly Migratory Species Management Division (Silver Spring, 1315 East West Highway).
Stasinopoulos M., Rigby R., Voudouris V., Akantziliotou C., Enea M., Kiose D., et al. (2023). gamlss: Generalised Additive Models for Location Scale and Shape. Available online at: https://cran.r-project.org/web/packages/gamlss/index.html (Accessed October 18, 2023).
Stevens J. D., Bonfil R., Dulvy N. K., and Walker P. A. (2000). The effects of fishing on sharks, rays, and chimaeras (chondrichthyans), and the implications for marine ecosystems. ICES J. Mar. Sci. 57, 476–494. doi: 10.1006/jmsc.2000.0724
Thorson J. T. (2019). Guidance for decisions using the Vector Autoregressive Spatio-Temporal (VAST) package in stock, ecosystem, habitat and climate assessments. Fisheries Res. 210, 143–161. doi: 10.1016/j.fishres.2018.10.013
Thorson J. (2024). James-Thorson-NOAA/VAST. Available online at: https://github.com/James-Thorson-NOAA/VAST (Accessed February 28, 2024).
Thorson J. T. and Barnett L. A. K. (2017). Comparing estimates of abundance trends and distribution shifts using single- and multispecies models of fishes and biogenic habitat. ICES J. Mar. Sci. 74, 1311–1321. doi: 10.1093/icesjms/fsw193
Thorson J. T., Maunder M. N., and Punt E. (2020). The development of spatio-temporal models of fishery catch-per-unit-effort data to derive indices of relative abundance. Fisheries Res. 230, 105611. doi: 10.1016/j.fishres.2020.105611
Thorson J. T., Pinsky M. L., and Ward E. J. (2016). Model-based inference for estimating shifts in species distribution, area occupied and centre of gravity. Methods Ecol. Evol. 7, 990–1002. doi: 10.1111/2041-210X.12567
Thorson J. T., Scheuerell M. D., Shelton A. O., See K. E., Skaug H. J., and Kristensen K. (2015). Spatial factor analysis: a new tool for estimating joint species distributions and correlations in species range. Methods Ecol. Evol. 6, 627–637. doi: 10.1111/2041-210X.12359
TinHan T. C. and Wells R. J. D. (2021). Spatial and ontogenetic patterns in the trophic ecology of juvenile bull sharks (Carcharhinus leucas) from the northwest gulf of Mexico. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.664316
Ulrich G., Jones C., Driggers W., Drymon M., Oakley D., and Riley C. (2007). Habitat utilization, relative abundance, and seasonality of sharks in the estuarine and nearshore waters of South Carolina. Am. Fisheries Soc. Symposium 50, 125–139.
Vinyard E. A., Frazier B. S., Drymon J. M., Gelsleichter J. J., and Bubley W. J. (2019). Age, growth, and maturation of the Finetooth Shark, Carcharhinus isodon, in the Western North Atlantic Ocean. Environ. Biol. Fish 102, 1499–1517. doi: 10.1007/s10641-019-00929-9
Waller M. J., Humphries N. E., Womersley F. C., Loveridge A., Jeffries A. L., Watanabe Y., et al. (2024). The vulnerability of sharks, skates, and rays to ocean deoxygenation: Physiological mechanisms, behavioral responses, and ecological impacts. J. Fish Biol. doi: 10.1111/jfb.15830
Werry J. M. and Clua E. (2013). Sex-based spatial segregation of adult bull sharks, Carcharhinus leucas, in the New Caledonian great lagoon. Aquat. Living Resour. 26, 281–288. doi: 10.1051/alr/2013063
Zuur A. F., Fryer R. J., Jolliffe I. T., Dekker R., and Beukema J. J. (2003a). Estimating common trends in multivariate time series using dynamic factor analysis. Environmetrics 14, 665–685. doi: 10.1002/env.611
Zuur A., Ieno E. N., Walker N., Saveliev A. A., and Smith G. M. (2009). Mixed Effects Models and Extensions in Ecology with R (New York, NY: Springer Science & Business Media).
Keywords: spatiotemporal models, dynamic factor analysis, Bayesian hierarchical, index standardization, stock assessment, data-limited
Citation: O’Brien KA, Carlson JK, Cortés E, Driggers WB III, Frazier BS and Latour RJ (2025) Evaluation of a spatiotemporal index standardization method for coastal shark species; implications for future stock assessments. Front. Mar. Sci. 12:1621720. doi: 10.3389/fmars.2025.1621720
Received: 01 May 2025; Accepted: 25 July 2025;
Published: 08 September 2025.
Edited by:
Wen-Pei Tsai, National Kaohsiung University of Science and Technology, TaiwanReviewed by:
José Lino Vieira De Oliveira Costa, University of Lisbon, PortugalAlexander Hansell, Northeast Fisheries Science Center (NOAA), United States
Copyright © 2025 O’Brien, Carlson, Cortés, Driggers, Frazier and Latour. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Kaitlyn A. O’Brien, a2FvYnJpZW5Admltcy5lZHU=
†ORCID: Kaitlyn A. O’Brien, orcid.org/0009-0003-2175-0636
John K. Carlson, orcid.org/0000-0001-8205-6558
Enric Cortés, orcid.org/0000-0001-6001-2482
Bryan S. Frazier, orcid.org/0000-0002-7718-9492
Robert J. Latour, orcid.org/0000-0002-8644-1733