 Kaltham A. Ismail
Kaltham A. Ismail Mohammed Salim
Mohammed Salim- 1College of Marine Sciences and Aquatic Biology, University of Khorfakkan, Sharjah, United Arab Emirates
- 2Department of Meteorology, University of Reading, Reading, United Kingdom
Phytoplankton productivity in the Arabian Gulf is strongly influenced by regional oceanographic processes, yet the specific role of upwelling remains unclear. This study addresses this gap by analyzing seasonal and lagged correlations between Chlorophyll-a (Chl-a) concentrations and Vertical velocity (Wt) during the summer monsoon period (June–September 2018). The results show that in the Iranian regions (I & II), a slight delay in phytoplankton response to upwelling was observed in early summer, with the highest correlation at lag 1 in June (r = 0.80). In contrast, Region III, located in the northwestern Arabian Sea adjacent to the Omani coast, exhibited an immediate response throughout the season, with peak correlations at lag 0 (r = 0.72 in June and 0.67 in July). Stratification and horizontal currents played distinct roles in modulating Chl-a distributions: in Iran, stratification was positively correlated with Chl-a throughout the season (r = 0.37–0.42), while currents showed a consistently negative correlation (r = -0.57 to -0.40). In region III, stratification was negatively correlated with Chl-a in June (r = -0.54), becoming positive in later months (up to r = 0.39), and currents were positively correlated throughout (r = 0.15–0.42). These regional contrasts emphasize the importance of local oceanographic context, with upwelling, stratification, and horizontal transport exerting distinct controls on phytoplankton dynamics in the Arabian Gulf and northwestern Arabian Sea.
1 Introduction
Phytoplankton primary productivity constitutes the foundation of marine ecosystems, sustaining global biogeochemical cycles and the marine food web by driving the carbon cycle and influencing trophic interactions (Behrenfeld and Falkowski, 1997; Falkowski, 1994; Field et al., 1998). Chlorophyll-a (Chl-a), the primary photosynthetic pigment in phytoplankton, is widely used as a proxy for phytoplankton biomass and serves as an essential indicator of marine ecosystem responses to both physical and biological drivers (Lehman, 1981; Vörös and Padisák, 1991). Among these drivers, upwelling plays a critical role by delivering nutrient-rich deep water to the surface ocean, thereby elevating productivity in regions where nutrient availability often limits phytoplankton growth (Arístegui et al., 2004; Brandt et al., 2023).
Upwelling facilitates the vertical transport of cold, nutrient-rich waters from deeper ocean layers to the euphotic zone, which enhances phytoplankton photosynthesis and drives primary production (Carr, 2002; Chavez and Messié, 2009; Messié et al., 2009). Eastern Boundary Upwelling Ecosystems (EBUEs), including the Canary, California, Humboldt, and Benguela Current systems, demonstrate the global significance of upwelling processes (Carr and Kearns, 2003; Chavez and Messié, 2009). Although they cover only about 2% of the ocean surface, these regions contribute nearly 20% of global marine fish catches, highlighting their exceptional biological productivity and economic significance (Fréon et al., 2009). These systems emphasize the importance of wind-driven upwelling in sustaining fisheries and supporting ecological balance (Arístegui et al., 2004). In contrast to the extensively studied EBUEs, the Arabian Sea, recognized as one of the most productive regions globally, relies on seasonal upwelling driven by the southwest monsoon to sustain its high productivity (Yi et al., 2021). This process not only influences nutrient dynamics but also modulates sea surface temperatures (SSTs), with implications for regional air-sea interactions and contributing to broader climatic feedback (Pareja-Roman et al., 2024).
While upwelling systems are globally significant, the Arabian Gulf presents a unique case of nutrient dynamics. As a semi-enclosed water body, the Gulf is characterized by high salinity (Swift and Bower, 2003; Vasou et al., 2020) and oligotrophic conditions, with surface waters often limited by low nitrate concentrations (Al-Yamani et al., 2006). In addition to anthropogenic sources such as treated wastewater and industrial discharges, riverine input also plays a notable role in the nutrient budget of the Gulf. The Shatt Al-Arab River, formed by the confluence of the Tigris and Euphrates, is a major source of freshwater and nutrients to the northwestern Gulf, supporting elevated phytoplankton biomass and influencing regional nutrient regimes, especially near its delta (Ghaemi et al., 2024; Saad, 1978). Similarly, the Mond River discharges into the Gulf in proximity to Region I, contributing to localized increases in nutrient concentrations and primary productivity, particularly during periods of high flow (Ghaemi et al., 2024). These riverine inputs, together with limited water exchange through the narrow Strait of Hormuz, shape the Gulf’s nutrient landscape and highlight its vulnerability to both natural variability and human-induced changes (Sheppard et al., 2010; Jawad, 2021).
In contrast, the northwestern Arabian Sea adjacent to the Omani coast, including the Sea of Oman, is subject to seasonal upwelling primarily driven by the southwest monsoon. This upwelling brings nutrient-rich waters to the surface, supporting higher levels of primary productivity compared to the Arabian Gulf (Goes et al., 2005; Kumar et al., 2001; Piontkovski et al., 2011).Despite its ecological significance, detailed studies on the mechanisms, variability, and ecological consequences of upwelling in the northwestern Arabian Sea, including the Sea of Oman and the Omani coastal region, are limited. Most available research has focused on broad patterns of productivity or has documented specific bloom events, while comprehensive assessments of the spatial and temporal dynamics of upwelling and its direct relationship with chlorophyll-a concentrations remain scarce (Banse and English, 2000; Wiggert et al., 2005). This lack of detailed investigation highlights a gap in our understanding of how upwelling influences phytoplankton dynamics in this region.
The present study addresses the gap in understanding the mechanisms regulating phytoplankton productivity in the northwestern Arabian Sea and the Arabian Gulf by examining how upwelling (as indicated by vertical velocity), stratification, and horizontal currents interact to shape phytoplankton dynamics. Horizontal currents may transfer nutrients and Chl-a across regions, potentially mitigating the direct impact of upwelling, whereas stratification might restrict nutrient mixing at times of large thermal gradients (Lévy et al., 2012; McGillicuddy et al., 2007). Specifically, this work aims to: (1) examine the seasonal trends in Chlorophyl-a (Chl-a) and vertical velocity (Wt) to understand the temporal dynamics of upwelling-driven productivity; (2) analyze the lagged relationships between Chl-a and Wt to uncover temporal delays in phytoplankton responses; and (3) evaluate the roles of stratification and horizontal currents in influencing Chl-a beyond upwelling. By providing a nuanced analysis of these interactions, this study contributes new insights into the physical and biological processes that regulate productivity in a region that remains relatively understudied compared to other major upwelling systems.
2 Data and methods
2.1 Study area
The domain selected for this study includes the Arabian Gulf and the northwestern Arabian Sea adjacent to the Omani coast (hereafter referred to as the northwestern Arabian Sea), encompassing the Sea of Oman (Figure 1). A bathymetric map (GEBCO Bathymetric Compilation Group, 2024)) of the Arabian Gulf and the northwestern Arabian Sea, shown in Figure 1, provides the geographical context for this study. The map is overlaid with the climatological summer (JJA) general circulation pattern simulated by the model (Al-Shehhi et al., 2021; Ismail, 2023; Salim et al., 2024; Subeesh et al., 2025). These regions are characterized by dynamic oceanographic processes, including seasonal upwelling (Ismail and Al Shehhi, 2022). These areas are significant for understanding the dynamics of Chl-a and Wt, which are key factors driving phytoplankton productivity and nutrient dynamics in the region.
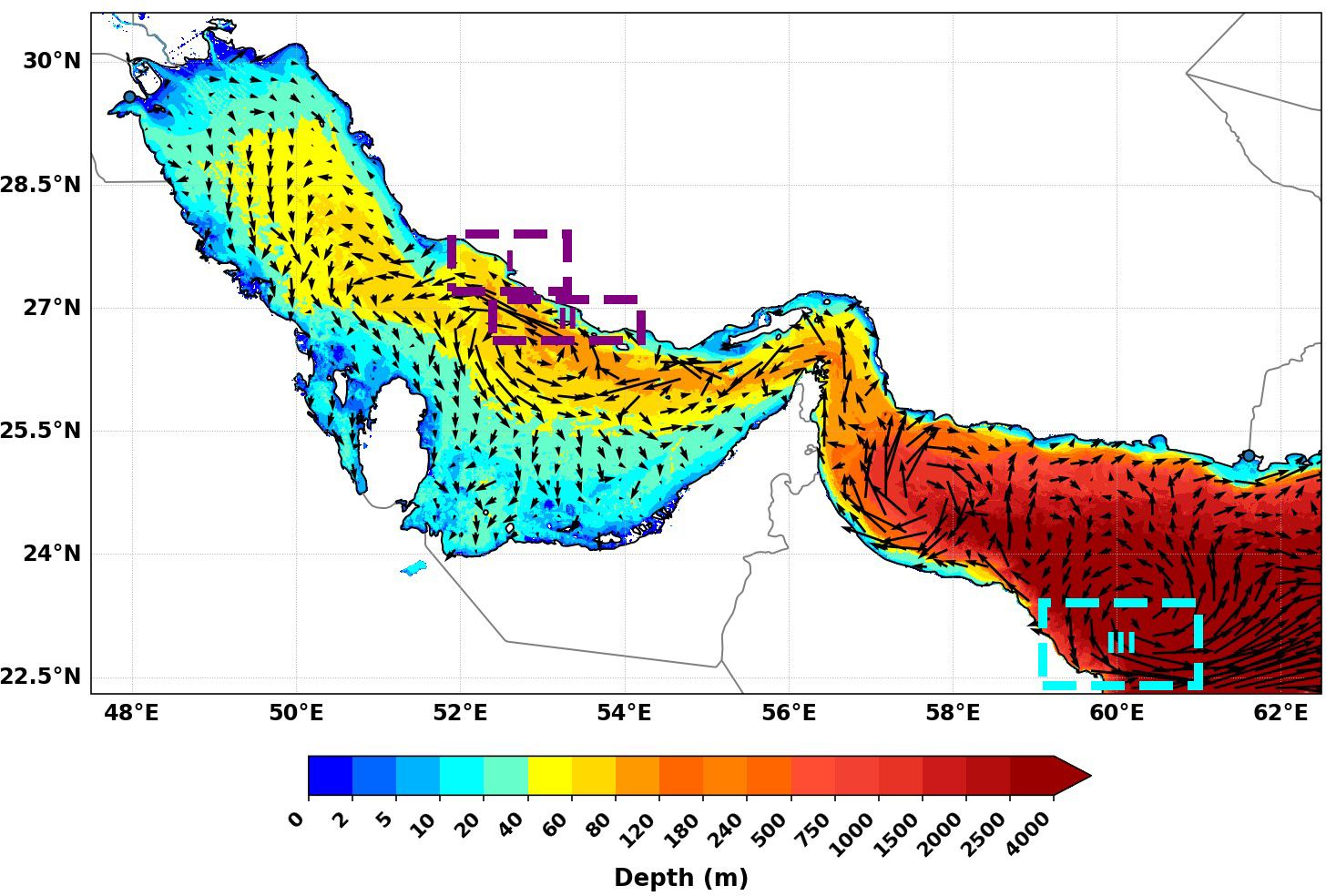
Figure 1. Bathymetric map (GEBCO-2024) of the Arabian Gulf and northwestern Arabian Sea, highlighting depth variations and primary upwelling zones (Iranian Region: I, II, and Northwestern Arabian Sea Region: III) marked by boxes with upward arrows. The bathymetry is overlaid with the climatological summer (JJA) general circulation pattern simulated by the model.
The study domain can be divided into two primary sub-regions:
Arabian Gulf: A semi-enclosed body of water with a highly variable nutrient regime (Al-Yamani and Naqvi, 2019), influenced by both natural factors (e.g., upwelling) and anthropogenic impacts (e.g., nutrient runoff from industrial areas) (Devlin et al., 2015; Ismail and Al Shehhi, 2022). The Gulf is connected to the Sea of Oman through the narrow Strait of Hormuz, which limits its exchange with the open Arabian Sea. This restricted exchange makes the Arabian Gulf particularly sensitive to both natural and human-induced changes in water quality and nutrient levels.
Northwestern Arabian Sea Adjacent to the Omani Coast (including the Sea of Oman): This region, encompassing the Omani coastal waters and the Sea of Oman, represents the northern part of the Arabian Sea and is characterized by more consistent upwelling and nutrient input (Bali et al., 2019; Jickells et al., 2005; Kumar and Narvekar, 2005; Lee et al., 2000). This provides a contrasting environment to the Arabian Gulf, where upwelling events are more variable. The northwestern Arabian Sea and Sea of Oman play a crucial role in nutrient transport and phytoplankton productivity (Ismail and Al Shehhi, 2022), making this area important for understanding marine ecosystem dynamics in the region.
2.2 Datasets
To investigate phytoplankton responses to upwelling in the Arabian Gulf and the northwestern Arabian Sea, this study utilized the following datasets to analyze the relationship between Chl-a concentrations and Wt: Chl-a: Monthly climatology Chl-a data were obtained from the NASA Ocean Color project, specifically the MODIS aqua (year 2018) (NASA Goddard Space Flight Center, Ocean Biology Processing Group, https://oceandata.sci.gsfc.nasa.gov/). MODIS chlorophyl anomaly calculated between 2018 Chl-a and MODIS climatology obtained from GMIS-MODIS-AQUA Monthly climatology data (Melin, 2013).These data provide monthly mean chlorophyll concentrations (µg/L) and were used to assess seasonal patterns in Chl-a and their correspondence with upwelling signals inferred from Wt.
2.2.1 Total vertical velocity
The Wt, used as a proxy for upwelling intensity, was derived from the ERA-Interim reanalysis dataset provided by the European Centre for Medium-Range Weather Forecasts (ECMWF) (https://www.ecmwf.int/en/forecasts/datasets/reanalysis-datasets/era-interim). The dataset includes 10 m zonal and meridional wind components (u10 and v10) that were used to estimate total vertical velocity (coastal and curl driven upwelling) via the Ekman transport method as described in (Ismail and Al Shehhi, 2022). The wind components have a spatial resolution of 0.75° x 0.75°, and the vertical velocities were derived from the curl of the wind field (Ismail and Al Shehhi, 2022).
2.2.2 Additional supporting data
Simulated Ocean Current Data: Monthly zonal (U) and meridional (V) ocean current components for the year 2012 were obtained from model-generated NetCDF datasets produced by the MIT General Circulation Model (MITgcm). These simulations have been validated and used in several regional studies in the Sea of Oman and the Arabian Gulf (Al-Shehhi et al., 2021; Ismail, 2023; Salim et al., 2024; Subeesh et al., 2025). The climatological or seasonal circulation patterns were well validated and closely aligned with those reported in several previous studies (e.g., Reynolds, 1993, Kampf and Sadrinasab, 2005). Regardless of the forcing year, all studies consistently revealed a similar general circulation pattern. Detailed model configurations and validation procedures are described in Salim et al. (2024); Subeesh et al. (2025), and Al-Shehhi et al. (2021).
The dataset provides ocean current velocity fields at multiple depths (up to 83 levels) and has a spatial resolution of 2.5 km for longitude and latitude. The U and V ocean current components were analyzed to assess their potential impact on Chl-a concentrations, in conjunction with Wt, which serves as a proxy for upwelling intensity. Ocean currents may affect nutrient transport pathways, phytoplankton distribution, and primary productivity, contributing to spatial variability in Chl-a concentrations across the Arabian Gulf and the Sea of Oman (Jordi et al., 2009).
Sea Surface Temperature (SST): Monthly SST data were obtained from the same MIT general circulation model outputs for the same period (year 2012) and were used to calculate the stratification index. The SST data have a spatial resolution of 2.5 km and include a depth range down to 83 levels. These model outputs, validated through comparison with observational data (Al-Shehhi et al., 2021; Salim et al., 2024), were used to assess the thermal stratification and upwelling influence in the Arabian Gulf and the northwestern Arabian Sea.
The stratification index was computed as follows:
Where: SSTsurface: Refers to the Sea Surface Temperature at the shallowest available layer in the model data, typically around 5 meters, as specified in the dataset.
SSTbottom: Refers to the Sea Surface Temperature at a deeper layer, which is 50 meters.
This index provides a normalized measure of thermal stratification, which can inhibit vertical nutrient mixing and thereby influence phytoplankton growth dynamics.
2.3 Methodology
2.3.1 Correlation analysis of Chl-a and upwelling (total vertical velocity)
We performed a seasonal correlation analysis to examine the relationship between Chl-a and upwelling intensity (Wt) in the Arabian Gulf and northwestern Arabian Sea. The Pearson correlation coefficient was used to quantify the strength of the linear relationship between monthly average values of Chl-a and Wt. Seasonal variations in upwelling intensity and phytoplankton biomass were assessed using the monthly SST stratification index and Chl-a data.
2.3.2 Lagged correlation analysis
To further investigate the timing of the relationship between upwelling and Chl-a concentration, we performed lagged correlation analysis. This approach accounts for the biological response time of phytoplankton to upwelling-induced nutrient supply, which may not be instantaneous due to ecological and biogeochemical processes (Ma et al., 2021).
2.3.2.1 Time series preparation
Monthly time series of upwelling index (Wt) and Chl-a concentration were extracted for each sub-region (Iran I & II and northwestern Arabian Sea III) within the study area.
2.3.2.2 Lag implementation
The Chl-a time series was systematically shifted relative to the upwelling time series by lag periods ranging from -2 months to +2 months (τ = -2, -1, 0, + 1, + 2 months). Negative lags indicate Chl-a leading upwelling, while positive lags indicate Chl-a lagging behind upwelling (Bograd et al., 2009; Jacox et al., 2015).
2.3.2.3 Correlation calculation
For each lag period τ, the Pearson correlation coefficient was computed between the upwelling index at time t and Chl-a concentration at time t + τ:
2.3.2.4 Optimal lag identification
The lag period producing the maximum absolute correlation coefficient was identified as the optimal lag, representing the characteristic response time of phytoplankton to upwelling forcing (Jacox et al., 2016).
For this analysis, monthly Chl-a concentrations and Wt were extracted for the period April to September for the year (2018) in the study. This period was selected because it corresponds to the primary upwelling and phytoplankton growing season in the region, during which both physical and biological signals are most pronounced. The analysis was restricted to this six-month window to focus on the period of greatest ecological relevance for upwelling-driven phytoplankton dynamics. As only six data points from a single year were available for the lagged correlation analysis, the results should be interpreted as exploratory and descriptive. The short time series does not allow for robust statistical significance testing or generalization beyond the 2018 upwelling season. Future work including multi-year or full-annual-cycle data is recommended to strengthen the analysis.
2.3.3 Spatial analysis
The spatial distribution of Chl-a and upwelling was examined by analyzing the geographical extent of high and low upwelling regions within the study domain. We computed the spatial correlations between Chl-a and Wt in selected upwelling hotspots (Ismail and Al Shehhi, 2022) along the Iranian and northwestern Arabian Sea coasts (I, II, III), Figure 1.
2.3.4 Statistical significance
Statistical significance of the correlation analyses between Chl-a and Wt was determined using p-values derived from the correlation coefficients. A threshold of p-value< 0.05 was considered statistically significant (Zar, 2014) for the relationship between Chl-a and vertical velocity.
2.4 Limitations and data comparability
This study integrates remotely sensed surface Chl-a data, reanalysis-derived Wt, and model-simulated oceanographic parameters to investigate upwelling-driven phytoplankton dynamics in the Arabian Gulf and northwestern Arabian Sea. While this multi-source approach offers a comprehensive perspective, several limitations and considerations should be acknowledged regarding data comparability, selection, and interpretation.
2.4.1 Comparability of data sources
Vertical velocity used here as a proxy for upwelling intensity, was calculated from ERA-Interim reanalysis wind data using the Ekman transport method. While ERA-Interim provides consistent and widely used gridded atmospheric data, it may not capture all fine-scale or transient wind patterns, especially in coastal or semi-enclosed regions. The stratification index and ocean current fields were obtained from the MITgcm, a high-resolution ocean circulation model previously validated for this region. However, as with all models, uncertainties remain due to parameterizations and boundary conditions.
Chlorophyll-a concentrations were derived from MODIS satellite observations, which represent surface-layer phytoplankton biomass. Satellite Chl-a estimates are subject to potential errors from atmospheric correction, water turbidity, and colored dissolved organic matter, particularly in optically complex coastal environments like the Arabian Gulf. Furthermore, satellite sensors primarily detect surface signals and may not fully capture subsurface phytoplankton maxima or vertical variability.
2.4.2 Justification for data selection and time frames
The datasets and time frames were selected based on their regional validation, spatial and temporal resolution, and relevance to the study objectives. The period from April to September 2018 was chosen to focus on the main upwelling and phytoplankton growth season, as documented in a previous study (Ismail and Al Shehhi, 2022). Monthly averaging of data was employed to minimize the influence of short-term variability and to align the temporal resolution across all datasets.
2.4.3 Limitations and implications
While the use of a six-month, single-year dataset limits the ability to generalize results across years or different regional settings, the findings provide valuable insights into seasonal-scale phytoplankton-upwelling interactions in the Arabian Gulf and northwestern Arabian Sea. These results should be viewed as a baseline contribution that emphasizes the importance of further multi-year studies for robust temporal generalization.
2.4.4 Recommendations
Future studies should incorporate multi-year and higher-frequency datasets, as well as in situ measurements of Chl-a and nutrients, to validate and extend the findings presented here. Such efforts would help resolve vertical and temporal variability more accurately and support more robust statistical analyses.
3 Results and discussions
3.1 Seasonal trends in chlorophyll-a and vertical velocity
Understanding the seasonal dynamics of phytoplankton productivity requires examining how surface Chl-a responds to physical processes such as upwelling. In this study, we focus on the summer monsoon period (June–September), when upwelling is expected to intensify due to prevailing southwesterly winds (Kumar et al., 2001; Wiggert et al., 2005). The Arabian Gulf and the northwestern Arabian Sea represent two adjacent yet hydrographically distinct systems. The Arabian Gulf is semi-enclosed, shallow, and strongly stratified during summer (Al Azhar et al., 2016), whereas the northwestern Arabian Sea is deeper, open to the Arabian Sea, and more directly influenced by monsoon-driven upwelling (Piontkovski et al., 2011). These regional contrasts provide an ideal framework to investigate how phytoplankton respond to variability in vertical nutrient flux.
Figure 2 displays average Chl-a concentrations from June to September (for the upwelling season), overlaid with vertical velocity vectors to indicate the main upwelling zones and areas of high phytoplankton biomass. The spatial alignment of elevated Chl-a with strong vertical velocities suggests a linkage between upwelling intensity and surface phytoplankton biomass. This analysis examines three key upwelling zones: the Iranian upwelling regions (Regions I and II) and the northwestern Arabian Sea upwelling region (Region III), as delineated in Figure 1. These subregions were selected to evaluate spatial differences in the coupling between vertical nutrient supply and biological response, thereby offering insight into the drivers of primary production in each zone.
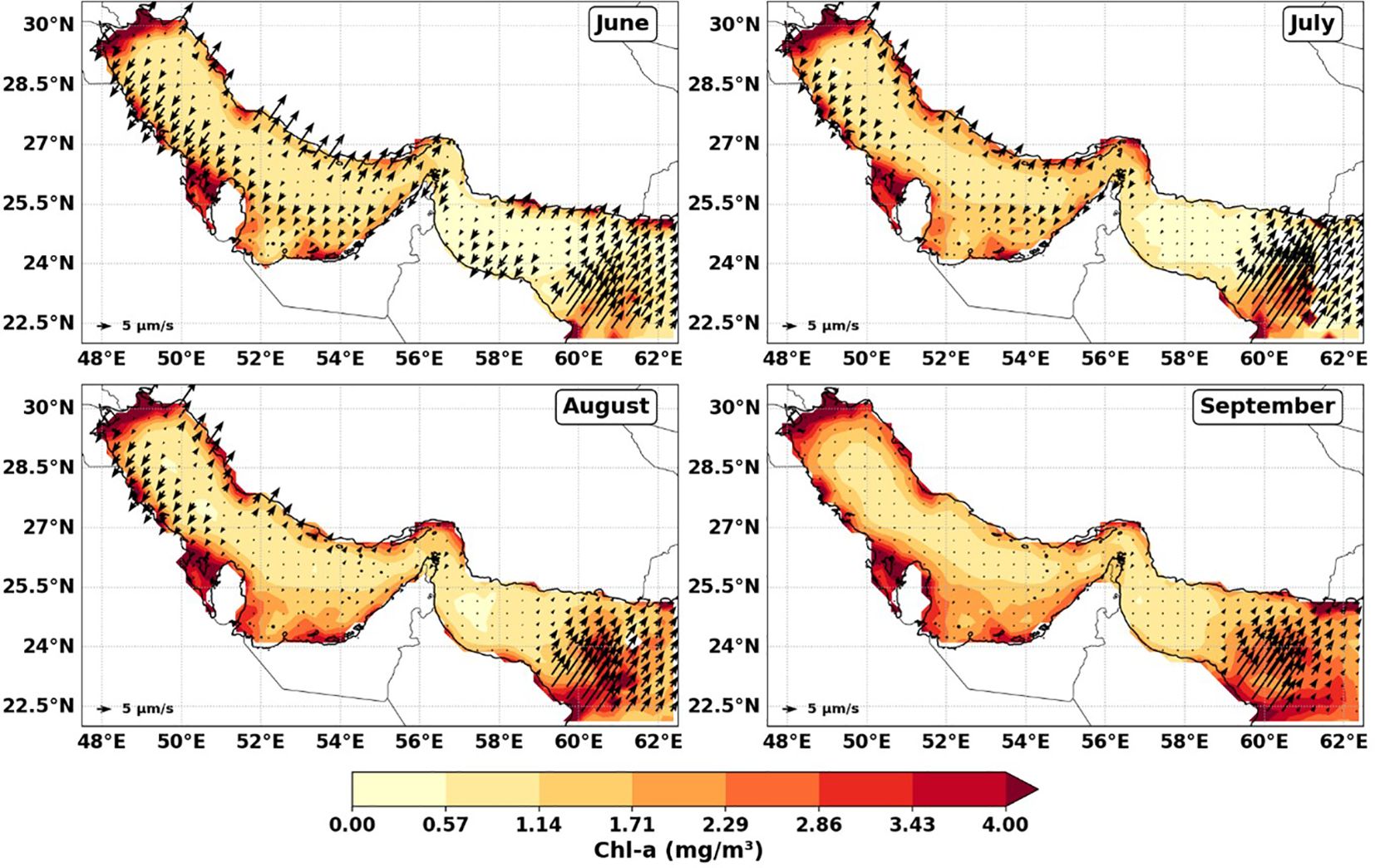
Figure 2. Monthly spatial distribution of chlorophyll-a concentration (mg m-3) for June, July, August, and September, overlaid with vectors representing upwelling index (vertical velocity, m s-1) as a proxy for upwelling intensity. Arrows indicate areas of enhanced vertical water movement and associated phytoplankton accumulation.
3.1.1 Iranian region (I and II)
In the Iranian coastal upwelling zones (Regions I and II), Chl-a concentrations exhibit a clear seasonal pattern, closely linked to upwelling dynamics during the summer monsoon. Chl-a increases from 1.23 mg m-3 in June to a peak of 1.68 mg m-3 in July, followed by a gradual decline to 1.45 mg m-3 in August and 1.37 mg m-3 in September (Figure 3, left y-axis). Simultaneously, Wt exhibits a decreasing trend, indicating that upwelling intensity weakens over time. Wt declines from 6.34 × 10-6 m s-1 in June to 3.46 × 10-6 m s-1 in July, followed by 3.29 × 10-6 m s-1 in August, reaching its lowest value of 1.50 × 10-6 m s-1 in September (Figure 3, right y-axis) suggesting that the strongest upwelling occurs in early summer (June–July), when rising water masses enhance nutrient supply to the euphotic zone, promoting phytoplankton growth as reflected by the peak in Chl-a concentrations (Banse and English, 2000; Wiggert et al., 2005). The subsequent decline in vertical velocity from July onward suggests a weakening upwelling signal, but Chl-a remains relatively high throughout the summer, indicating a sustained phytoplankton response. This persistence may be attributed to several factors, including nutrient retention within the system, continued stratification dynamics, and biological processes such as grazing pressure and changes in community composition (Behrenfeld and Falkowski, 1997; Landry et al., 2024; Teixeira et al., 2014). For example, shifts in phytoplankton community structure toward larger or less palatable species, or the dominance of taxa with lower grazing susceptibility, can reduce the efficiency of grazer control and allow Chl-a to remain elevated (Cloern, 1996; Teixeira et al., 2014). Conversely, if grazer populations (such as zooplankton) do not increase proportionally with phytoplankton, or if there is a mismatch in timing, phytoplankton biomass can persist at high levels despite ongoing grazing (Calbet and Landry, 2004; Sommer et al., 2012). Thus, it is the interplay between phytoplankton community composition and grazer dynamics that influences the persistence of high Chl-a concentrations during periods of reduced upwelling (Landry et al., 2024). These findings are consistent with other upwelling systems, such as the California Current and the Peru Current, where phytoplankton productivity remains elevated even after upwelling intensity diminishes, due to delayed biological responses and nutrient recycling (Checkley and Barth, 2009; Espinoza-Morriberón et al., 2017; Igarza et al., 2019). In particular, (Jacox et al., 2016) reported that upwelling alone does not always dictate biological productivity, as pre-existing oceanic conditions, nutrient retention, and local hydrography can mediate the phytoplankton response to upwelling variability.
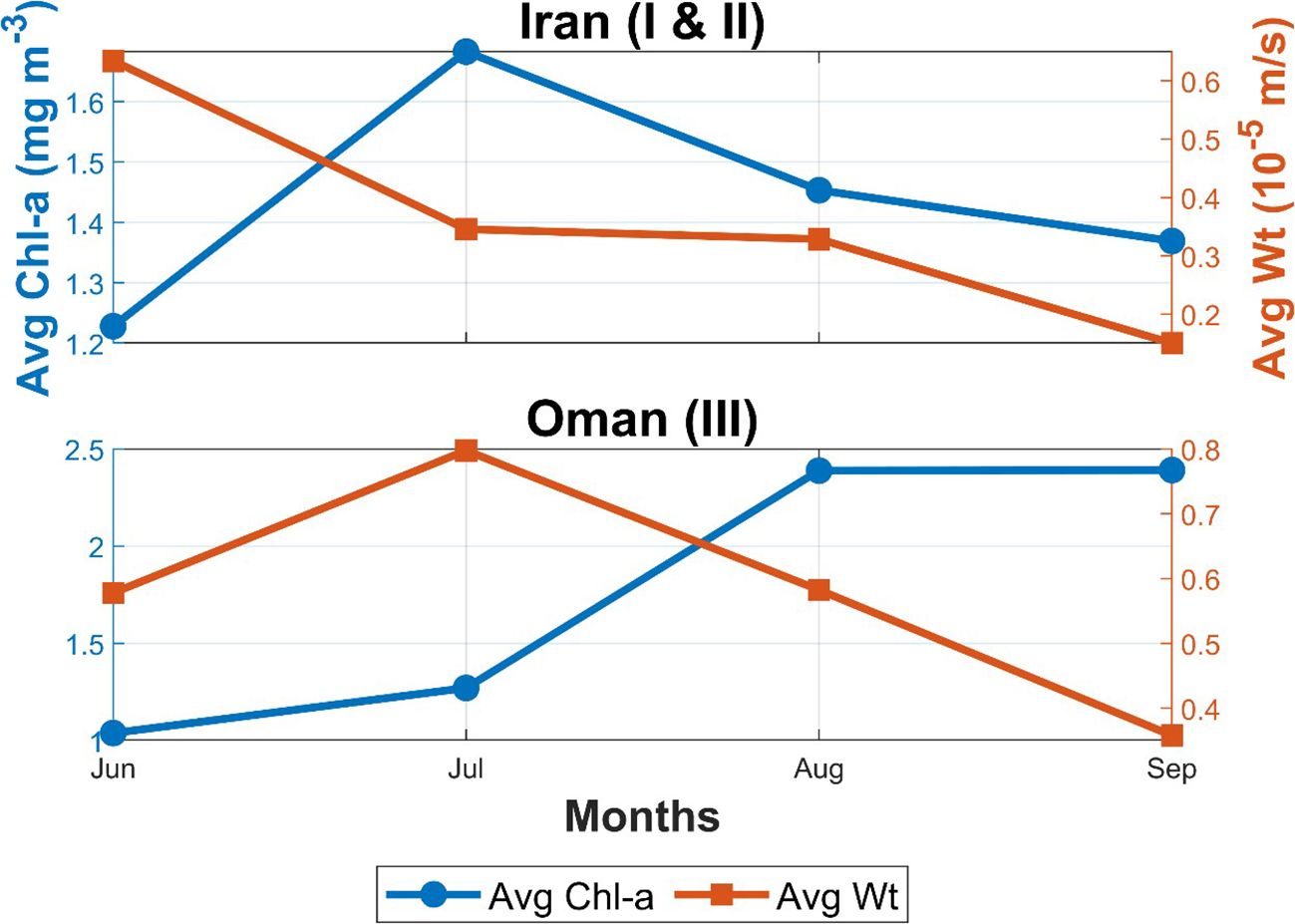
Figure 3. Monthly averages of chlorophyll-a (Chl-a, mg m-3) and vertical velocity (Wt, 10-5 m s-1) for the Iranian (Regions I & II) and northwestern Arabian Sea (Region III) coasts from June to September.
3.1.2 Northwestern Arabian Sea region (III)
In the northwestern Arabian Sea (Region III), Chl-a concentrations follow a clear seasonal pattern, increasing from 1.04 mg m-3 in June to 2.39 mg m-3 in August, then stabilizing at 2.39 mg m-3 in September. This trend closely corresponds with the seasonal evolution of Wt, which peaks in July at 7.97 × 10-6 m s-1, before declining to 5.82 × 10-6 m s-1 in August and 3.58 × 10-6 m s-1 in September. The rapid rise in Chl-a response to the intensifying Wt during early to mid-summer suggests a direct link between upwelling-driven nutrient flux and surface phytoplankton growth. This pattern is similar to other coastal upwelling systems, where rapid nutrient input fuels phytoplankton blooms almost immediately during strong upwelling periods (Chavez and Messié, 2009; Broullón et al., 2023). The alignment between the peak in Wt and the highest Chl-a values in July highlights the role of upwelling in supporting productivity in northwestern Arabian Sea waters. This pattern is consistent with earlier studies that have documented the significant impact of monsoon-driven upwelling on seasonal phytoplankton dynamics in the region (Wiggert et al., 2005; Al-Azri et al., 2010).
Despite the decline in Wt after July, Chl-a remains high in August and September, suggesting that nutrient recycling or retention may help sustain phytoplankton growth beyond the peak upwelling period. This pattern has been observed in other major upwelling systems, such as the Benguela Current, where efficient nutrient utilization allows phytoplankton blooms to persist even after upwelling slows down (Hutchings et al., 2009; Monteiro et al., 2011). Additionally, the persistence of Chl-a despite weakening upwelling intensity could also be influenced by stratification dynamics, which have been shown to regulate phytoplankton growth in other eastern boundary upwelling systems (Messié and Chavez, 2015).
Furthermore, biological processes, as discussed above, likely contribute to sustaining productivity after the decline in upwelling intensity (Landry et al., 2024; Teixeira et al., 2014). Studies in the California Current have demonstrated that phytoplankton blooms can persist even during weakened upwelling phases due to organic matter recycling and continued nitrate availability in subsurface layers (Jacox et al., 2016). A similar mechanism may be at play in northwestern Arabian Sea upwelling region, supporting a prolonged phytoplankton response beyond the peak upwelling season.
This prolonged biological or phytoplankton response is further supported by spatial anomaly maps (Figure 4), which reveal localized, near-instantaneous positive Chl-a anomalies along the northwestern Arabian Sea coast (Sea of Oman) during July–August, indicative of a rapid response to strong upwelling. In contrast, the Iranian coast displays predominantly neutral to negative anomalies during early upwelling, with only modest positive anomalies emerging one to two months later. This spatial and temporal differentiation reflects a lagged biological response in the Iranian regions, likely arising from slower nutrient replenishment and local stratification constraints, and is consistent with established observations of delayed chlorophyll peaks following upwelling events in the northern Arabian Gulf.
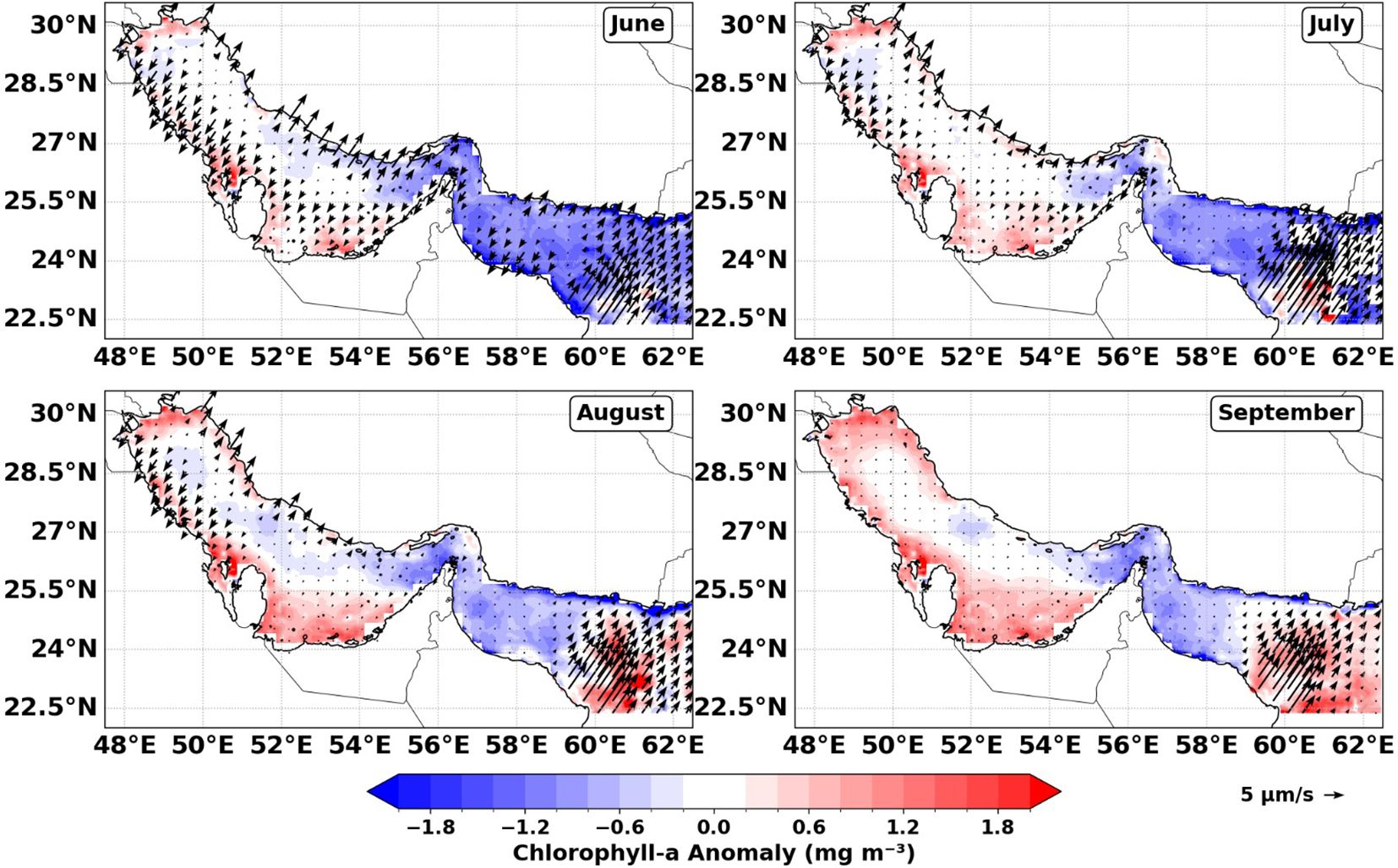
Figure 4. Monthly chlorophyll-a (Chl-a) anomaly map for 2018, computed relative to the MODIS climatological mean from Melin (2013). Positive anomalies (shades of red) represent regions where 2018 Chl-a concentrations exceeded the climatological average, while negative anomalies (shades of blue) indicate below-average values.
3.2 Correlation between chlorophyll-a and vertical velocity
To assess the physical–biological coupling across the study region, we examined the relationship between Chl-a concentrations and Wt in the Iranian (Regions I and II) and northwestern Arabian Sea (Region III) upwelling zones. These correlations provide insight into how seasonal upwelling, stratification, and horizontal transport influence surface phytoplankton productivity.
3.2.1 Iranian region (I & II)
In the Iranian upwelling zones, Chl-a exhibits a strong positive correlation with Wt during early summer, with coefficients of 0.76 in June and 0.83 in July at lag 0, declining to 0.50 in August and 0.48 in September (Figure 5). This pattern indicates that upwelling-driven nutrient supply strongly fuels phytoplankton growth, especially in early summer when upwelling peaks (Ismail and Al Shehhi, 2022; Ghaemi et al., 2024). As upwelling weakens in August and September, the correlation declines, implying that other processes such as remineralization, lateral transport, and biological interactions sustain phytoplankton biomass (Hutchings et al., 2009; Jacox et al., 2016; Kudela et al., 2008). These findings align with previous observations in the Arabian Gulf and Sea of Oman, where subsurface nutrient intrusions and organic matter remineralization contribute to elevated nitrate and silicate concentrations during late summer and early autumn (Ghaemi et al., 2021; Al-Azri et al., 2010) In addition to natural nutrient sources, anthropogenic nutrient inputs such as treated wastewater, industrial runoff, and coastal pollution may elevate surface nutrient concentrations and support phytoplankton growth even in the absence of vertical mixing (Al-Yamani et al., 2006; Devlin et al., 2015).
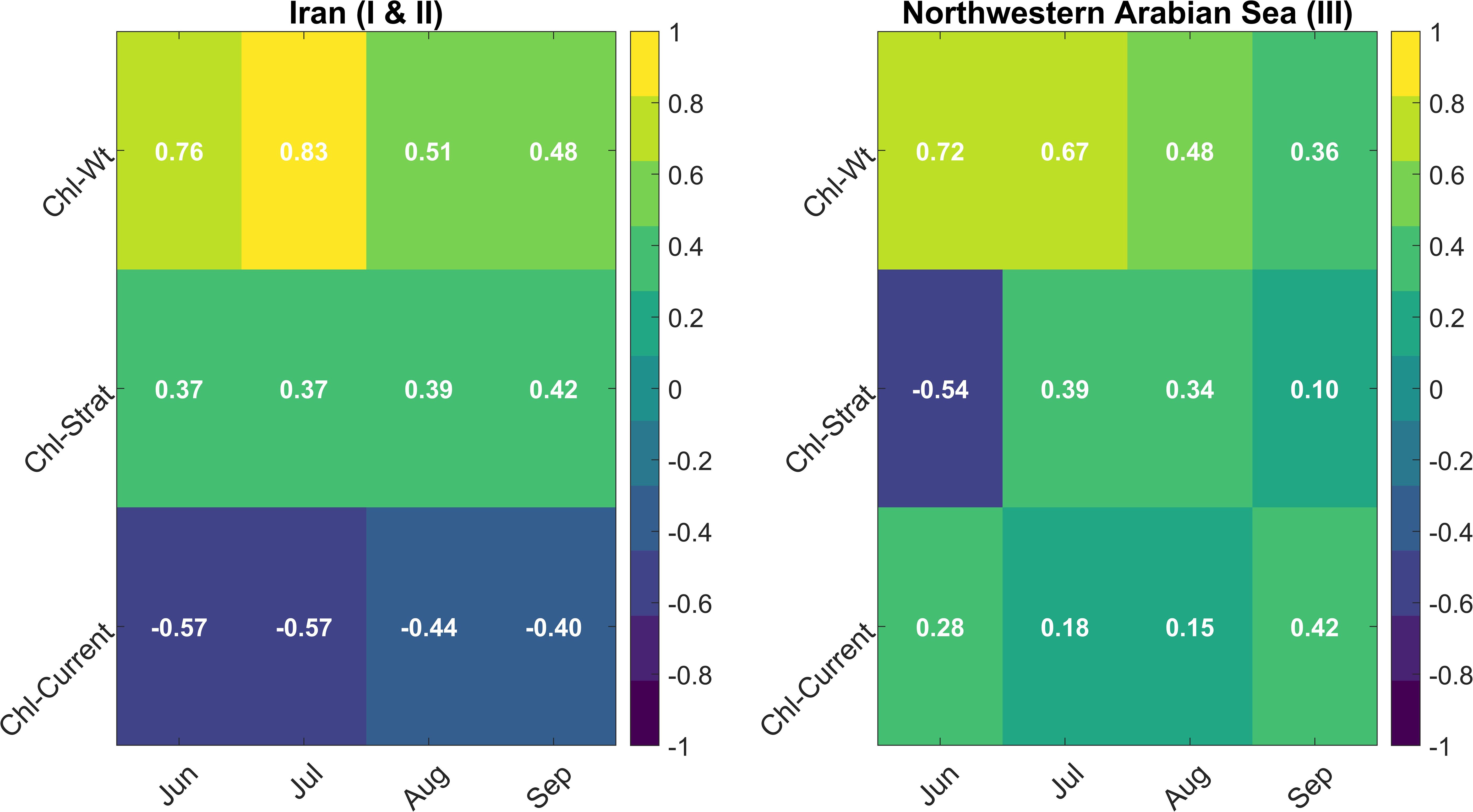
Figure 5. Monthly correlation coefficients between chlorophyll-a concentrations (Chl-a) and oceanographic variables (Vertical Velocity (Wt), Stratification index (Strat), and Ocean Currents (current)) for Iranian (I, II) and northwestern Arabian Sea (III) regions (Jun–Sep).
Interestingly, the correlation between stratification and Chl-a in Iran is positive (0.37 to 0.42), contradicting the expected inverse relationship. Stratification typically suppresses nutrient mixing (Behrenfeld et al., 2006). However, in semi-enclosed basins like the Arabian Gulf, other processes can sustain productivity despite stratification. Potential explanations include lateral nutrient transport from the Sea of Oman (Al Azhar et al., 2016), nutrient retention and recycling within the mixed layer, maintaining high surface productivity despite weaker upwelling (Ismail and Al Shehhi, 2022), and the dominance of small phytoplankton (e.g., flagellates, cyanobacteria) that thrive in stratified, stable conditions, rather than diatoms which require strong mixing (Igarza et al., 2019). The negative correlation between Chl-a and currents (-0.57 to -0.40) suggests that horizontal advection disperses phytoplankton rather than enhancing growth. Similar patterns were observed in the California Current, where circulation spreads phytoplankton biomass without necessarily increasing local chlorophyll levels. Compared to northwestern Arabian Sea, upwelling in Iranian waters is weaker and delivers fewer nutrients, so currents tend to disperse phytoplankton rather than support their growth (Ismail and Al Shehhi, 2022; Lachkar et al., 2022). Similar patterns have been documented in other upwelling systems, such as the California and Peru Currents, where phytoplankton blooms often persist after peak upwelling due to nutrient recycling and delayed uptake (Checkley and Barth, 2009; Espinoza-Morriberón et al., 2017; Igarza et al., 2019).
3.2.2 Northwestern Arabian Sea region (III)
In the northwestern Arabian Sea (Region III), the pattern of correlation between Chl-a and Wt differs from the Iranian upwelling zones. The correlation is strongest in June (0.72), reflecting a rapid phytoplankton response to upwelling. This relationship weakens through the season, declining to 0.67 in July, 0.48 in August, and 0.35 in September, paralleling the seasonal decrease in upwelling intensity. This trend suggests that as upwelling weakens, additional factors such as nutrient variability and physical transport processes (e.g., eddies) increasingly influence phytoplankton growth (Al-Azri et al., 2010). Stratification in this region plays a more dynamic role than in Iran. A negative correlation between stratification and Chl-a in June (-0.54) indicates that strong early-season stratification suppresses nutrient mixing and limits phytoplankton production. As upwelling intensifies, this effect diminishes, and the influence of stratification on phytoplankton blooms weakens, consistent with previously reported seasonal dynamics (Lachkar et al., 2022; Wiggert et al., 2005). By September, the correlation between stratification and Chl-a is weakly positive (0.1), suggesting that processes such as horizontal transport, remineralization, and biological interactions become more important (Al-Azri et al., 2010). The late-summer shift from diatom-dominated blooms to dinoflagellates aligns with this pattern, as dinoflagellates are better adapted to lower-nutrient, stratified conditions (Piontkovski et al., 2011; Wiggert and Murtugudde, 2007).
Unlike the Iranian zones, currents in the northwestern Arabian Sea region show a positive correlation with Chl-a, particularly in September (0.42). This suggests that horizontal transport may enhance Chl-a concentrations by redistributing nutrients and phytoplankton, especially when retention times are favorable (Lachkar and Gruber, 2011). This contrasts with the Iranian region, where horizontal advection tends to disperse phytoplankton, a pattern also observed in regions with strong export processes, such as the central Arabian Sea (Bauer et al., 1991). The sustained upwelling along the Omani coast likely maintains nutrient availability even as surface waters are advected offshore (Ismail and Al Shehhi, 2022). Similar mechanisms have been reported in other upwelling systems, such as the Canary and Benguela Currents, where lateral transport supports persistent phytoplankton blooms (Chavez and Messié, 2009).
3.3 Lagged response of Chl-a to upwelling
To further investigate the timing of phytoplankton response to upwelling, lagged correlation analyses were performed between Chl-a concentrations and Wt, revealing distinct regional differences (Figure 6).
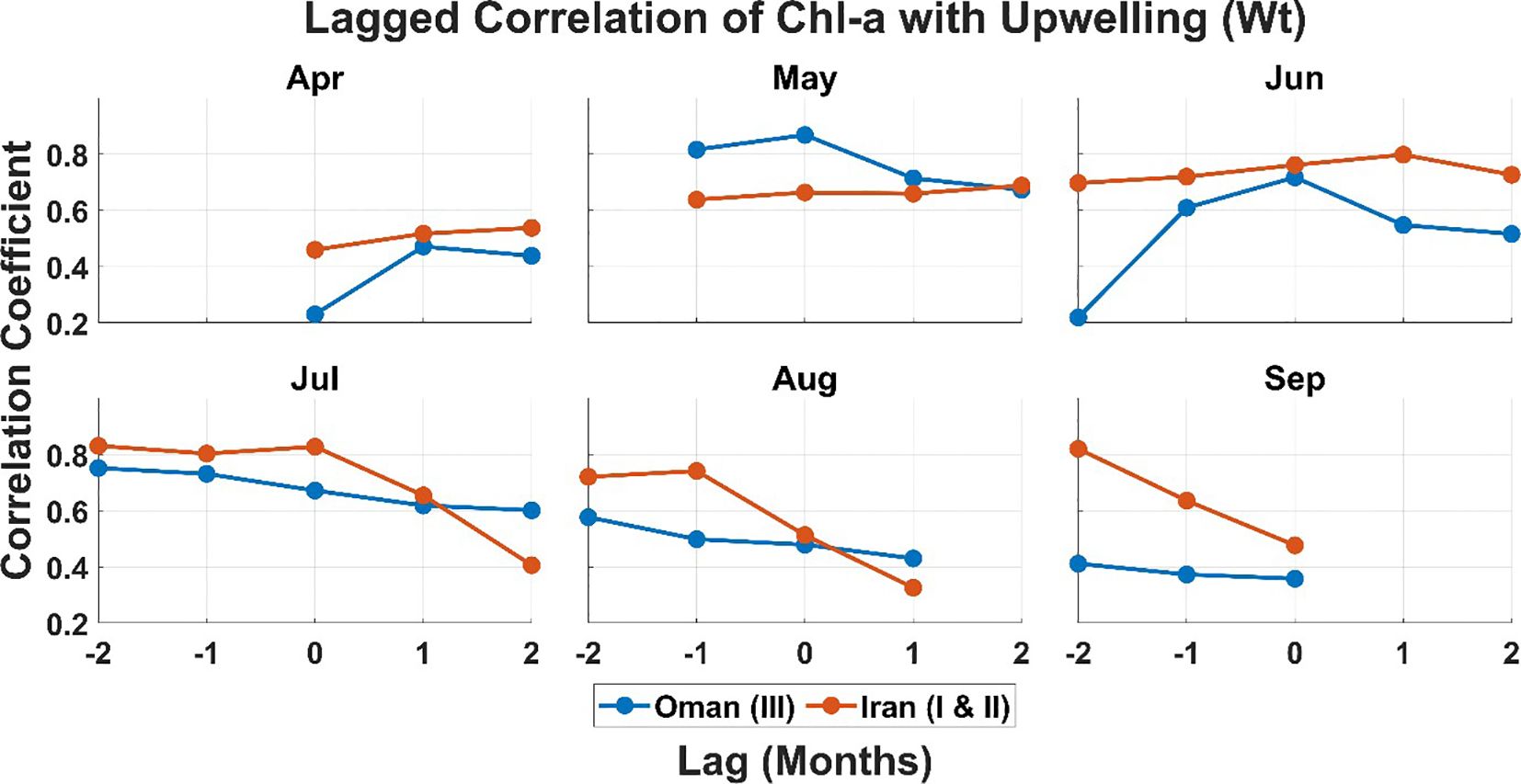
Figure 6. Monthly lagged correlations between chlorophyll-a and upwelling index (vertical velocity, Wt) from April to September for Iranian waters (Regions I & II; orange) and northwestern Arabian Sea waters (Region III; blue). The x-axis denotes time lag (months), with negative lags indicating upwelling leading phytoplankton response. Lines represent correlation coefficients for each month and region, highlighting seasonal and regional differences in upwelling–chlorophyll dynamics.
3.3.1 Iranian region (I and II)
In the Iranian region, the correlation between Chl-a and Wt increases slightly from 0.76 at lag 0 to 0.80 at lag 1 in June and decreases from 0.83 at lag 0 to 0.65 at lag 1 in July. While this minor change suggests a possible delayed phytoplankton response to upwelling in June, the difference is small and not statistically significant due to the limited data. Thus, it should be interpreted as a potential trend rather than definitive evidence. Physical and biological processes such as nutrient remineralization, vertical mixing, and subsurface nutrient transport may contribute to this lag (Brock et al., 1991; Ershadifar et al., 2023; Lachkar et al., 2022). Similar lagged responses have been observed in other upwelling systems, including the Western Iberian Coast (Favareto et al., 2023) and the California and Peru Currents, where delays are attributed to nutrient cycling and grazing pressure (Chavez and Messié, 2009; Wilkerson et al., 2006).
Greater stratification along the Iranian coast, compared to Oman, may limit upward diffusion of subsurface nutrients, causing remineralized nutrients to accumulate at depth and become available only during subsequent upwelling events (Alosairi et al., 2011; Al-Yamani and Naqvi, 2019; Kämpf and Sadrinasab, 2005). Subsurface inflows from the Sea of Oman may also sustain nutrient levels in Iranian waters beyond the peak upwelling period (Al-Azri et al., 2010; Al-Yamani and Naqvi, 2019; Ghaemi et al., 2024). Zooplankton grazing and community structure may further influence bloom persistence, as lower grazing pressure could allow phytoplankton biomass to remain elevated for longer durations (Roman et al., 2000).
3.3.2 Northwestern Arabian Sea region (III)
In contrast, the northwestern Arabian Sea upwelling region displays an immediate phytoplankton response to upwelling, with the strongest correlation between Chl-a and Wt at lag 0 (0.72) in June and weaker correlations at negative lags (Figures 5, 6). This rapid response is consistent with the immediate injection of nutrients into the euphotic zone following upwelling events (Zhong et al., 2019). Similar rapid responses have been documented in other systems, such as the Mediterranean (Fuchs et al., 2023) and the California and Peru Currents (Chavez and Messié, 2009). The most intense upwelling along the northwestern Arabian Sea and along Omani coasts occurs in June and July, coinciding with the southwest monsoon and enhanced Ekman transport, which delivers cold, nutrient-rich waters to the surface (Al-Azri et al., 2010; Watanabe et al., 2017; Wiggert et al., 2005). This supports high chlorophyll levels and local fisheries productivity (Goes et al., 2005).
While our lag analysis provides insight into seasonal-scale phytoplankton responses, it is based on monthly data and may not capture finer-scale (e.g., weekly or daily) dynamics. Therefore, findings regarding the timing of delayed responses should be interpreted as broad trends rather than precise estimates.
4 Conclusions
This study explored the influence of upwelling on phytoplankton productivity in the Arabian Gulf and Sea of Oman, focusing on regional differences between the Iranian (Regions I and II) and northwestern Arabian Sea (Region III) coastal systems. Results revealed that upwelling plays a central but regionally variable role in regulating surface Chl-a concentrations: along the northwestern Arabian Sea coast, strong vertical nutrient transport during peak upwelling months (June–July) supports higher Chl-a levels and a rapid phytoplankton response, while on the Iranian shelf, a slightly delayed response is observed, with productivity peaking later in the season likely sustained by lateral nutrient transport, remineralization, and persistent stratification. Stratification showed a positive correlation with Chl-a in the Iranian region, suggesting that surface nutrient retention can support ongoing growth despite weaker mixing, whereas in the northwestern Arabian Sea, the limiting effect of early stratification diminishes as upwelling intensifies. Currents are negatively correlated with Chl-a in Iran, indicating a dispersive role, while in the northwestern Arabian Sea, the correlation becomes slightly positive by late summer, suggesting horizontal transport may help retain or redistribute phytoplankton biomass. Collectively, these findings indicate the importance of local oceanographic context in shaping phytoplankton responses to physical drivers and highlight the need for region-specific management and monitoring strategies in semi-enclosed and upwelling-influenced marine systems.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
KI: Conceptualization, Data curation, Formal Analysis, Investigation, Methodology, Validation, Writing – original draft, Writing – review & editing. MS: Data curation, Methodology, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. The authors would like to thank Univeristy of Khorfakkan for covering the Article Processing Charges (APC) for this publication.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that Generative AI was used in the creation of this manuscript. The authors used ChatGPT (OpenAI) to improve the language, clarity, and structure of the manuscript text. All content was verified and edited by the authors to ensure scientific accuracy and integrity.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Al Azhar M., Temimi M., Zhao J., and Ghedira H. (2016). Modeling of circulation in the Arabian Gulf and the Sea of Oman: Skill assessment and seasonal thermohaline structure. J. Geophysical Research: Oceans 121, 1700–1720. doi: 10.1002/2015JC011038
Al-Azri A. R., Piontkovski S. A., Al-Hashmi K. A., Goes J. I., and do Gomes H. R. (2010). Chlorophyll a as a measure of seasonal coupling between phytoplankton and the monsoon periods in the Gulf of Oman. Aquat. Ecol. 44, 449–461. doi: 10.1007/s10452-009-9303-2
Alosairi Y., Imberger J., and Falconer R. A. (2011). Mixing and flushing in the Persian gulf (Arabian gulf). J. Geophysical Research: Oceans 116 (C3), 1–14. doi: 10.1029/2010JC006769
Al-Shehhi M. R., Song H., Scott J., and Marshall J. (2021). Water mass transformation and overturning circulation in the Arabian gulf. J. Phys. Oceanography 51, 3513–3527. doi: 10.1175/JPO-D-20-0249.1
Al-Yamani F. and Naqvi S. W. A. (2019). Chemical oceanography of the Arabian gulf. Deep-Sea Res. Part II: Topical Stud. Oceanography 161, 72–80. doi: 10.1016/j.dsr2.2018.10.003
Al-Yamani F., Subba Rao D. V., Mharzi A., Ismail W., and Al-Rifaie K. (2006). Primary production off Kuwait, an arid zone Environment, Arabian Gulf. Int. J. Oceans Oceanography 1, 67–85.
Arístegui J., Álvarez-Salgado X., Barton E., Figueiras F., Hernández-León S., Roy C., et al. (2004). “Chapter 23 : oceanography and fisheries of the Canary current/Iberian region of the eastern North Atlantic (18a,E),” in The Sea, vol. 14 . Eds. Robinson A. R. and Brink K. H. (Cambridge, UK: Harvard University Press), 877–897.
Bali K., Mishra A. K., Singh S., Chandra S., and Lehahn Y. (2019). Impact of dust storm on phytoplankton bloom over the Arabian Sea: a case study during March 2012. Environ. Sci. pollut. Res. 26, 11940–11950. doi: 10.1007/s11356-019-04602-7
Banse K. and English D. C. (2000). Geographical differences in seasonality of CZCS-derived phytoplankton pigment in the Arabian Sea for 1978-1986. Deep-Sea Res. II 47, 1623–1677. doi: 10.1016/S0967-0645(99)00157-5
Bauer S., Hitchcock G. L., and Olson D. B. (1991). Influence of monsoonally-forced Ekman dynamics upon surface layer depth and plankton biomass distribution in the Arabian Sea. Deep Sea Res. Part A. Oceanographic Res. Papers 38, 531–553. doi: 10.1016/0198-0149(91)90062-K
Behrenfeld M. J. and Falkowski P. G. (1997). Photosynthetic rates derived from satellite-based chlorophyll concentration. Limnology Oceanography 42, 1–20. doi: 10.4319/lo.1997.42.1.0001
Behrenfeld M. J., O’Malley R. T., Siegel D. A., McClain C. R., Sarmiento J. L., Feldman G. C., et al. (2006). Climate-driven trends in contemporary ocean productivity. Nature 444, 752–755. doi: 10.1038/nature05317
Bograd S. J., Schroeder I., Sarkar N., Qiu X., Sydeman W. J., and Schwing F. B. (2009). Phenology of coastal upwelling in the California Current. Geophysical Res. Lett. 36, L01602. doi: 10.1029/2008GL035933
Brandt P., Alory G., Awo F. M., Dengler M., Djakouré S., Anicet R., et al. (2023). Physical processes and biological productivity in the upwelling regions of the tropical Atlantic. Ocean Sci. 19, 581–601. doi: 10.5194/os-19-581-2023
Brock J. C., Mcclain C. R., Luther M. E., Hay W. W., Brock J. C., Mcclain C. R., et al. (1991). The phytoplankton bloom in the northwestern Arabian Sea during the southwest monsoon of 1979. J. Geophysical Res. 83, 623–643. Available online at: https://digitalcommons.usf.edu/msc_facpub/83 (Accessed April 20, 2025).
Broullón E., Franks P. J. S., Fernández Castro B., Gilcoto M., Fuentes-Lema A., Pérez-Lorenzo M., et al. (2023). Rapid phytoplankton response to wind forcing influences productivity in upwelling bays. Limnology Oceanography Lett. 8, 529–537. doi: 10.1002/lol2.10309
Calbet A. and Landry M. R. (2004). Phytoplankton growth, microzooplankton grazing, and carbon cycling in marine systems. Limnology Oceanography 49, 51–57. doi: 10.4319/lo.2004.49.1.0051
Carr M.-E. (2002). Estimation of potential productivity in Eastern Boundary Currents using remote sensing. Deep-Sea Res. II 49, 59–80. doi: 10.1016/S0967-0645(01)00094-7
Carr M.-E. and Kearns E. J. (2003). Production regimes in four Eastern Boundary Current systems. Deep-Sea Res. Part II: Topical Stud. Oceanography 50, 3199–3221. doi: 10.1016/j.dsr2.2003.07.015
Chavez F. P. and Messié M. (2009). A comparison of eastern boundary upwelling ecosystems. Prog. Oceanography 83, 80–96. doi: 10.1016/j.pocean.2009.07.032
Checkley D. M. and Barth J. A. (2009). Patterns and processes in the California current system. Prog. Oceanography 83, 49–64. doi: 10.1016/j.pocean.2009.07.028
Cloern J. E. (1996). Phytoplankton bloom dynamics in coastal ecosystems: a review with some general lessons from sustained investigation of San Francisco Bay, California. Rev. Geophysics 34, 127–168. doi: 10.1029/96RG00986
Devlin M. J., Massoud M. S., Hamid S. A., Al-Zaidan A., Al-Sarawi H., Al-Enezi M., et al. (2015). Changes in the water quality conditions of Kuwait’s marine waters: Long term impacts of nutrient enrichment. Mar. pollut. Bull. 100, 607–620. doi: 10.1016/j.marpolbul.2015.10.022
Ershadifar H., Saleh A., Koochaknejad E., Kor K., and Ghazilou A. (2023). Spatial and seasonal distribution of particulate phosphorous and nitrogen in the Persian Gulf: Nitrogen enrichment ties to diazotroph bloom in stratified warm waters. Mar. Chem. 253, 104280. doi: 10.1016/j.marchem.2023.104280
Espinoza-Morriberón D., Echevin V., Colas F., Tam J., Ledesma J., Vásquez L., et al. (2017). Impacts of El Niño events on the Peruvian upwelling system productivity. J. Geophysical Research: Oceans 122, 5423–5444. doi: 10.1002/2016JC012439
Falkowski P. G. (1994). The role of phytoplankton photosynthesis in global biogeochemical cycles. Photosynthesis Res. 39, 235–258. doi: 10.1007/BF00014586
Favareto L., Brotas V., Rudorff N., Zacarias N., Tracana A., Lamas L., et al. (2023). Response of phytoplankton to coastal upwelling: The importance of temporal and spatial scales. Limnology Oceanography 68, 1376–1387. doi: 10.1002/lno.12353
Field C. B., Behrenfeld M. J., Randerson J. T., Field C. B., Behrenfeld M. J., and Randerson J. T. (1998). Primary production of the biosphere : integrating terrestrial and oceanic components. Am. Assoc. Advancement Sci. 281, 237–240. Available online at: http://www.jstor.org/stable/2896014 (Accessed May 10, 2025).
Fréon P., Barange M., and Arístegui J. (2009). Eastern boundary upwelling ecosystems: integrative and comparative approaches. Prog. Oceanography 83, 1–14. doi: 10.1016/j.pocean.2009.08.001
Fuchs R., Rossi V., Caille C., Bensoussan N., Pinazo C., Grosso O., et al. (2023). Intermittent upwelling events trigger delayed, major, and reproducible pico-nanophytoplankton responses in coastal oligotrophic waters. Geophysical Res. Lett. 50 (5), e2022GL102651. doi: 10.1029/2022GL102651
GEBCO Bathymetric Compilation Group (2024). The GEBCO_2024 Grid - a continuous terrain model of the global oceans and land (Liverpool, UK: NERC EDS British Oceanographic Data Centtre NOC). doi: 10.5285/1c44ce99-0a0d-5f4f-e063-7086abc0ea0f
Ghaemi M., Abtahi B., and Gholamipour S. (2021). Spatial distribution of nutrients and chlorophyll a across the Persian Gulf and the Gulf of Oman. Ocean Coast. Manage. 201, 105476. doi: 10.1016/j.ocecoaman.2020.105476
Ghaemi M., Hamzei S., Saleh A., and Gholamipour S. (2024). Nutrient regimes in a semi-enclosed marginal sea: The Persian Gulf. J. Operational Oceanography 17, 124–136. doi: 10.1080/1755876X.2024.2333596
Goes J. I., Thoppil P. G., Gomes H. D. R., and Fasullo J. T. (2005). Warming of the Eurasian landmass is making the Arabian sea more productive. Science 308, 545–547. doi: 10.1126/science.1106610
Hutchings L., van der Lingen C. D., Shannon L. J., Crawford R. J. M., Verheye H. M. S., Bartholomae C. H., et al. (2009). The Benguela Current: An ecosystem of four components. Prog. Oceanography 83, 15–32. doi: 10.1016/j.pocean.2009.07.046
Igarza M., Dittmar T., Graco M., and Niggemann J. (2019). Dissolved organic matter cycling in the coastal upwelling system off central Peru during an “El Niño” year. Front. Mar. Sci. 6. doi: 10.3389/fmars.2019.00198
Ismail K. A. (2023). Modeling Physical-Biogeochemical Interactions in the Marine System of the Arabian Gulf.
Ismail K. A. and Al Shehhi M. R. (2022). Upwelling and nutrient dynamics in the Arabian Gulf and sea of Oman. PloS One 17, e0276260. doi: 10.1371/journal.pone.0276260
Jacox M. G., Fiechter J., Moore A. M., and Edwards C. A. (2015). ENSO and the California current coastal upwelling response. J. Geophysical Research: Oceans 120, 1691–1702. doi: 10.1002/2014JC010650
Jacox M. G., Hazen E. L., Zaba K. D., Rudnick D. L., Edwards C. A., Moore A. M., et al. (2016). Impacts of the 2015–2016 El Niño on the California Current System: Early assessment and comparison to past events. Geophysical Res. Lett. 43, 7072–7080. doi: 10.1002/2016GL069716
Jawad L. A. (2021). The Arabian Seas: Biodiversity, Environmental Challenges and Conservation Measures Vol. 1–2 (Cham, Switzerland: Springer Nature Switzerland AG). doi: 10.1007/978-3-030-51506-5
Jickells T. D., An Z. S., Andersen K. K., Baker A. R., Bergametti G., Brooks N., et al. (2005). Global iron connections between desert dust, ocean biogeochemistry, and climate. Science 308, 67–71. doi: 10.1126/science.1105959
Jordi A., Basterretxea G., and Anglès S. (2009). Influence of ocean circulation on phytoplankton biomass distribution in the Balearic Sea: Study based on sea-viewing wide field-of-view sensor and altimetry satellite data. J. Geophysical Research: Oceans 114, C11005. doi: 10.1029/2009JC005301
Kämpf J. and Sadrinasab M. (2005). The circulation of the Persian Gulf: a numerical study. Ocean Sci. Discussions 2, 129–164. Available online at: www.ocean-science.net/osd/2/129/ (AccessedMay 10, 2025).
Kudela R. M., Banas N. S., Barth J. A., Frame E. R., Jay D. A., Largier J. L., et al. (2008). New Insights into the controls and Mechanisms of Plankton Productivity in coastal Upwelling Waters of the Northern california current system. Oceanography Soc. 21, 46–59. doi: 10.5670/oceanog.2008.04
Kumar S. P. and Narvekar J. (2005). Seasonal variability of the mixed layer in the central Arabian Sea and its implication on nutrients and primary productivity. Deep Sea Res. Part II: Topical Stud. Oceanogr 52 (14-15), 1848–1861.
Kumar S. P., Madhupratap M., Kumar M. D., Muraleedharan P. M., De Souza S. N., Gauns M., et al. (2001). High biological productivity in the central Arabian Sea during the summer monsoon driven by Ekman pumping and lateral advection. Curr. Sci. 81 (12), 1633–1638. Available online at: https://drs.nio.res.in/drs/handle/2264/275 (Accessed April 25, 2025).
Lachkar Z. and Gruber N. (2011). What controls biological production in coastal upwelling systems? Insights from a comparative modeling study. Biogeosciences 8, 2961–2976. doi: 10.5194/bg-8-2961-2011
Lachkar Z., Mehari M., Lévy M., Paparella F., and Burt J. A. (2022). Recent expansion and intensification of hypoxia in the Arabian Gulf and its drivers. Front. Mar. Sci. 9. doi: 10.3389/fmars.2022.891378
Landry M. R., Freibott A. L., Stukel M. R., Selph K. E., Allen A. E., and Rabines A. (2024). Phytoplankton growth and grazing dynamics during anomalous heat wave and suppressed upwelling conditions in the southern California Current. Deep-Sea Res. Part I: Oceanographic Res. Papers 210, 104353. doi: 10.1016/j.dsr.2024.104353
Lee C. M., Jones B. H., Brink K. H., and Fischer A. S. (2000). The upper-ocean response to monsoonal forcing in the Arabian Sea: seasonal and spatial variability. Deep-Sea Res. II 47, 1177–1226. doi: 10.1016/S0967-0645(99)00141-1
Lehman P. W. (1981). Comparison of chlorophyll a and carotenoid pigments as predictors of phytoplankton biomass. Mar. Biol. 65, 237–244. doi: 10.1007/BF00397117
Lévy M., Ferrari R., Franks P. J. S., Martin A. P., and Rivière P. (2012). Bringing physics to life at the submesoscale. Geophysical Res. Lett. 39, L14602. doi: 10.1029/2012GL052756
Ma L., Xiao W., Laws E. A., Bai X., Chiang K. P., Liu X., et al. (2021). Responses of phytoplankton communities to the effect of internal wave-powered upwelling. Limnology Oceanography 66, 1083–1098. doi: 10.1002/lno.11666
McGillicuddy D. J., Anderson L. A., Bates N. R., Bibby T., Buesseler K. O., Carlson C. A., et al. (2007). Eddy/Wind interactions stimulate extraordinary mid-ocean plankton blooms. Science 316, 1021–1026. doi: 10.1126/science.1136256
Melin F. (2013). GMIS-MERIS Monthly climatology sea surface Chlorophyll-a concentration (9km) in mg m^-3. Eur. Commission Joint Res. Centre (JRC). Available online at: http://data.europa.eu/89h/a59f5362-a8af-4e85-b2e7-7052d244045e (Accessed July 05, 2025).
Messié M. and Chavez F. P. (2015). Seasonal regulation of primary production in eastern boundary upwelling systems. Prog. Oceanography 134, 1–18. doi: 10.1016/j.pocean.2014.10.011
Messié M., Ledesma J., Kolber D. D., Michisaki R. P., Foley D. G., and Chavez F. P. (2009). Potential new production estimates in four eastern boundary upwelling ecosystems. Prog. Oceanography 83, 151–158. doi: 10.1016/j.pocean.2009.07.018
Monteiro P. M. S., Dewitte B., Scranton M. I., Paulmier A., and Van Der Plas A. K. (2011). The role of open ocean boundary forcing on seasonal to decadal-scale variability and long-term change of natural shelf hypoxia. Environ. Res. Lett. 6, 25002. doi: 10.1088/1748-9326/6/2/025002
Pareja-Roman L. F., Miles T., and Glenn S. (2024). Coastal upwelling modulates winds and air-sea fluxes, impacting offshore wind energy. Front. Energy Res. 12. doi: 10.3389/fenrg.2024.1470712
Piontkovski S., Al-Azri A., and Al-Hashmi K. (2011). Seasonal and interannual variability of chlorophyll-a in the Gulf of Oman compared to the open Arabian sea regions. Int. J. Remote Sens. 32, 7703–7715. doi: 10.1080/01431161.2010.527393
Roman M., Smith S., Wishner K., Zhang X., and Gowing M. (2000). Mesozooplankton production and grazing in the Arabian Sea. Deep Sea Res. Part II: Topical Stud. Oceanography 47, 1423–1450.
Saad M. A. H. (1978). Seasonal Variations of some Physico-Chemical Conditions of Shatt al-Arab Estuary, Iraq. Estuar. Coast. Mar. Sci. 6, 503–513. doi: 10.1016/0302-3524(78)90027-0
Salim M., Subeesh M. P., Scott J., Song H., Marshall J., and Al Shehhi M. R. (2024). Role of tidal mixing on ocean exchange through the Strait of Hormuz. Environ. Res. Commun. 6, 071006. doi: 10.1088/2515-7620/ad578c
Sheppard C., Al-husiani M., Al-jamali F., Al-yamani F., Baldwin R., Bishop J., et al. (2010). The Gulf : A young sea in decline. Mar. Pollut. Bull. 60 (1), 13–38. doi: 10.1016/j.marpolbul.2009.10.017
Sommer U., Adrian R., De Senerpont Domis L., Elser J. J., Gaedke U., Ibelings B., et al. (2012). Beyond the plankton ecology group (PEG) model: Mechanisms driving plankton succession. Annu. Rev. ecology evolution systematics 43, 429–448. doi: 10.1146/annurev-ecolsys-110411-160251
Subeesh M. P., Song H., Addad Y., Scott J. R., Marshall J., and Al Shehhi M. R. (2025). Seasonality of internal tides in the Strait of Hormuz: observations and modeling. J. Geophysical Research: Oceans 130, e2024JC021007. doi: 10.1029/2024JC021007
Swift S. A. and Bower A. S. (2003). Formation and circulation of dense water in the Persian/Arabian Gulf. J. Geophys. Res: Oceans, 108 (C1), 4–1. doi: 10.1029/2002JC001360
Teixeira I. G., Crespo B., Nielsen T. G., and Figueiras F. G. (2014). Stratification-mixing cycles and plankton dynamics in a shallow estuary (Limfjord, Denmark). J. Plankton Res. 36, 475–489. doi: 10.1093/plankt/fbt107
Vasou P., Vervatis V., Krokos G., Hoteit I., and Sofianos S. (2020). Variability of water exchanges through the Strait of Hormuz. Ocean Dynamics 70, 1053–1065. doi: 10.1007/s10236-020-01384-2
Vörös L. and Padisák J. (1991). Phytoplankton biomass and chlorophyll-a in some shallow lakes in Central Europe. Hydrobiologia 215, 111–119. doi: 10.1007/BF00014715
Watanabe T. K., Watanabe T., Yamazaki A., Pfeiffer M., Garbe-Schönberg D., and Claereboudt M. R. (2017). Past summer upwelling events in the Gulf of Oman derived from a coral geochemical record. Sci. Rep. 7, 4568. doi: 10.1038/s41598-017-04865-5
Wiggert J. D., Hood R. R., Banse K., and Kindle J. C. (2005). Monsoon-driven biogeochemical processes in the Arabian Sea. Prog. Oceanography 65, 176–213. doi: 10.1016/j.pocean.2005.03.008
Wiggert J. D. and Murtugudde R. G. (2007). The sensitivity of the southwest monsoon phytoplankton bloom to variations in aeolian iron deposition over the Arabian Sea. J. Geophysical Research: Oceans 112 (C5), C05005. doi: 10.1029/2006JC003514
Wilkerson F. P., Lassiter A. M., Dugdale R. C., Marchi A., and Hogue V. E. (2006). The phytoplankton bloom response to wind events and upwelled nutrients during the CoOP WEST study. Deep-Sea Res. Part II: Topical Stud. Oceanography 53, 3023–3048. doi: 10.1016/j.dsr2.2006.07.007
Yi X., Hünicke B., and Zorita E. (2021). Evolution of the Arabian sea upwelling from the last millennium to the future as simulated by Earth system models. Climate 9, 72. doi: 10.3390/cli9050072
Zar J. H. (2014). Biostatistical analysis. 5th ed (London, United Kingdom: Pearson Education Limited).
Keywords: chlorophyll-a, upwelling, lagged correlation, nutrient, remineralization, Arabian Gulf, northwestern Arabian Sea
Citation: Ismail KA and Salim M (2025) Regional phytoplankton responses to upwelling in the Arabian Gulf and northwestern Arabian Sea adjacent to the Omani coast. Front. Mar. Sci. 12:1641995. doi: 10.3389/fmars.2025.1641995
Received: 05 June 2025; Accepted: 28 July 2025;
Published: 13 August 2025.
Edited by:
Fahad Al Senafi, Kuwait University, KuwaitReviewed by:
Ali H. Abdulhussain, Kuwait University, KuwaitMaria Saburova, Kuwait Institute for Scientific Research, Kuwait
Copyright © 2025 Ismail and Salim. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Kaltham A. Ismail, a2FsdGhhbS5pc21haWxAdWtmLmFjLmFl