 Natalia Servetto1,2*
Natalia Servetto1,2* Marleen De Troch3
Marleen De Troch3 Gastón Alurralde4,5
Gastón Alurralde4,5 Luciana Ferrero1,2
Luciana Ferrero1,2 M. Carla de Aranzamendi1,2Ricardo Sahade1,2*
M. Carla de Aranzamendi1,2Ricardo Sahade1,2*- 1Facultad de Ciencias Exactas Físicas y Naturales, Universidad Nacional de Córdoba, Córdoba, Argentina
- 2Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Instituto de Diversidad y Ecología Animal (IDEA), Ecosistemas Marinos Polares, Córdoba, Argentina
- 3Marine Biology, Ghent University, Gent, Belgium
- 4Department of Environmental Science, Stockholm University, Stockholm, Sweden
- 5Baltic Marine Environment Protection Commission Helsinki Commission (HELCOM) Secretariat, Helsinki, Finland
Introduction: Ocean acidification (OA), resulting from the absorption of increasing atmospheric CO2 by the oceans, represents a major threat to marine organisms. Despite growing concern, the biochemical responses of Antarctic species to OA remain poorly understood.
Methods: This study investigated the impact of OA (pH 7.70 ± 0.09) on the fatty acid (FA) composition of the Antarctic snail Neobuccinum eatoni over a two-month experimental period (December 2015–March 2016). Fatty acid profiles were analyzed in multiple tissues to assess potential alterations induced by low-pH (LpH) conditions.
Results: Significant tissue-specific changes in FA composition were detected, particularly in the mantle and gill. Under LpH exposure, notable modifications occurred in long-chain polyunsaturated fatty acids (LC-PUFAs) such as 22:5n-3, 22:6n-3, and 24:5n-6. Elevated LC-PUFA levels in the mantle suggested a compensatory response to oxidative stress, while shifts in the n-3/n-6 ratios in the gill pointed to potential alterations in immune and anti-inflammatory functions.
Discussion: Indicators of homeoviscous adaptation (HVA), including PUFA/SFA ratios and mean chain length (MCL), revealed biochemical strategies used by N. eatoni to maintain membrane fluidity under acidified conditions. This study provides the first evidence of FA-based responses to elevated pCO₂ in an Antarctic gastropod, highlighting the potential of fatty acids as sensitive biomarkers of physiological adaptation to environmental stressors.
1 Introduction
The Anthropocene, which began with the Industrial Revolution in the 18th century, is characterized by significant human impact on a global scale, marked by an unprecedented and rapid increase in atmospheric CO2 levels (Gingerich, 2019). Oceanic uptake of excess CO2 helps mitigate anthropogenic emissions at the expense of inducing ocean acidification (OA). This process alters the physicochemical properties of seawater, profoundly affecting marine organisms and ecosystem functions (Findlay and Turley, 2021). Key marine species are already experiencing significant impacts, threatening biodiversity and essential ecosystem services (Gattuso et al., 2015; Shi and Li, 2024; Teixidó et al., 2024).
The severity of OA impacts is expected to vary across regions, with high-latitude areas experiencing more intense effects at finer spatial scales. The Southern Ocean (SO) plays a disproportionately large role in global carbon uptake, accounting for 30–40% of anthropogenic CO2 absorption (Fisher et al., 2025). This, combined with naturally low calcium carbonate (CaCO3) levels and the increased solubility of CO2 in cold waters (Hancock et al., 2020), contributes to its already low buffering capacity, making the SO ecosystem particularly vulnerable to OA. Aragonite saturation is biologically important because it determines the availability of carbonate ions necessary for calcifying organisms to form and maintain their shells and skeletons. For example, pteropods and corals rely on supersaturated aragonite conditions to precipitate calcium carbonate structures, and declines in aragonite saturation due to ocean acidification can lead to shell dissolution and impaired growth (Fabry et al., 2008; Orr et al., 2005). These physiological effects on key species can cascade through the web, ultimately impacting ecosystem structure and function. For instance, projections based on an ensemble of ten Earth system models indicate that aragonite undersaturation events will begin to spread rapidly around 2030, affecting approximately 30% of the SO surface waters by 2060, and more than 70% by 2100 (Hauri et al., 2015). Moreover, from 2003 to 2022, CO2 absorption increased by 0.076 gC m−2 per month in the Atlantic region of the SO, largely due to enhanced westerly winds linked to the Antarctic Oscillation (AO) and events related to the El Niño Southern Oscillation (ENSO) (de Carvalho et al., 2025). This means that Antarctic biota are exposed to potentially accelerated and more severe OA conditions than elsewhere.
OA not only impedes biomineralization and leads to shell dissolution in calcifiers, but it also exerts broader impacts on marine biota through cascades of physiological and biochemical mechanisms, often involving energetic trade-offs and complex cellular adjustments (Johnson and Hofmann, 2017; Servetto et al., 2023, 2025). Under elevated partial pressure of carbon dioxide (pCO2), meeting these physiological and metabolic demands requires significant energy, which forces a reallocation of limited energy budgets away from other vital functions. Thus, beyond direct physiological impacts, reproduction, growth, and development can also be adversely affected (Kroeker et al., 2013; Turner et al., 2016). To meet this energetic demand, organisms generally rely on lipid reserves and adjust fatty acid (FA) composition (Gibbs et al., 2021), diverting resources toward processes such as enhanced glycolipid metabolism as observed in Crassostrea gigas under acid stress (Wang et al., 2025). These energetic trade-offs generally manifest as reductions in total lipid content and a shift in key fatty acid ratios (e.g., decreased polyunsaturated fatty acids (PUFAs)/saturated fatty acids (SFA)), providing a sensitive proxy for potential hidden costs of OA (Valles-Regino et al., 2015). The close link between FAs and fundamental physiological processes makes them powerful biomarkers for assessing health and stress responses in marine organisms (Capitão et al., 2017; Ericson et al., 2019). FAs are essential components of cellular membranes and key energy sources for metabolism (Arts and Kohler, 2009; Hedberg et al., 2023). Within immune and physiological functions, polyunsaturated fatty acids (PUFAs) such as docosahexaenoic acid (DHA, 22:6n-3) and eicosapentaenoic acid (EPA, 20:5n-3) play critical roles in lipid metabolism, reproduction, and anti-inflammatory processes (Corsolini and Borghesi, 2017; Schmitz and Ecker, 2008). Although less abundant, arachidonic acid (ARA, 20:4n-6) acts as a precursor of eicosanoids that regulate immune responses and other physiological pathways (Stanley-Samuelson et al., 1988). Ratios such as n-6/n-3 PUFAs provide further insight into stress and inflammation (Ericson et al., 2019; Van Anholt et al., 2004).
In addition, homeoviscous adaptation—where organisms adjust membrane fluidity in response to environmental change—is commonly evaluated through PUFA/SFA ratios and mean carbon chain length (MCL) (Bennett et al., 2018; Ericson et al., 2019). For instance, temperature-driven shifts in MCL have been documented in the sponge Rhopaloeides odorabile (Bennett et al., 2018), while reductions in saturated and monounsaturated fatty acids were observed in Artemia sinica and the gastropod Dicathais orbita under short-term acidifying CO2 conditions (Valles-Regino et al., 2015; Gao et al., 2018). These findings underscore the sensitivity of FA composition to environmental stress in aquatic organisms and ecosystems (Fadhlaoui and Lavoie, 2021). Gastropods are key components of benthic ecosystems, driving nutrient cycling, grazing, and serving as prey for higher trophic levels (Dennis et al., 2021). Their heavily calcified shells make them especially vulnerable to OA, which not only impairs calcification but also inhibits growth and development (Kroeker et al., 2013). Additionally, if the cost of coping with acidic conditions compromises lipid reserves (particularly essential FAs like EPA and DHA), it may reduce their nutritional value and jeopardize predator health and survival throughout the food web (Gladyshev et al., 2013; Zhukova, 2019). While research on Antarctic gastropods is still limited, studies on other Antarctic mollusks have shown that elevated CO2 levels can impair physiological performance (Cummings et al., 2011; Johnson and Hofmann, 2017; de Aranzamendi et al., 2021). For example, de Aranzamendi et al. (2021) examined the impact of OA on the Antarctic limpet Nacella concinna. During a 15-day controlled laboratory exposure to low pH, sublittoral individuals displayed downregulation of heat-shock protein genes (HSP70A and HSP70B), indicating a stress response to acidified conditions. These results suggest that OA alone can substantially disrupt the physiological functioning of N. concinna, potentially reducing their resilience under future OA scenarios.
This study focuses on Neobuccinum eatoni, an Antarctic gastropod found in shallow coastal areas to over 2000 meters deep. As an endemic species dominating Antarctic benthic ecosystems (Schiaparelli et al., 2006; Norkko et al., 2007), N. eatoni represents a relevant model species to assess the impacts of OA. Model projections under elevated CO2 emissions suggest a substantial decline in suitable habitat for N. eatoni, underscoring both its susceptibility to ocean acidification and the need to prioritize research on this species (González et al., 2024). This study examines the specific effects of OA on the FA composition of N. eatoni, aiming to determine the effect of OA on the FA composition of the Antarctic snail N. eatoni, by comparing individuals exposed to acidified conditions with those maintained under control conditions. Recognizing the established link between FA profiles and environmental stressors (including temperature, pH, and nutrient availability), the research focuses on key FA indicators associated with immune functions (such as the 22:6n-3/20:4n-6 and n-3/n-6 ratios), and HVA (including MCL and PUFA/SFA ratios). These metrics were analyzed across tissues (mantle, gill, gonads, and foot) with distinct metabolic roles and lipid requirements. Experimental exposures were conducted under current ambient CO2 levels and elevated concentrations projected for 2100 under the high-emission RCP8.5 scenarios (IPCC, 2019), simulating future ocean conditions.
The central hypothesis posits that N. eatoni exposed to elevated CO2 levels will exhibit significant alterations in FA composition, reflecting changes in immune function and membrane fluidity compared to controls (individuals under current ambient conditions). Using FA profiles, the study addresses this question: could FA composition serve as a potential biomarker of stress in snails exposed to OA? By exploring these questions, the research aims to elucidate the mechanisms by which OA affects marine organisms at the biochemical level, enhancing our understanding of the potential impacts on Antarctic marine life.
2 Materials and methods
2.1 Study area
The experiment was carried out for 66 days in Potter Cove (PC) (62°14′S., 58°40′W; King George/25 de Mayo Island - South Shetlands - Antarctica) during the summer campaign of 2015-2016 (from December to March) (Figure 1). N. eatoni was collected by scuba diving at approximately 15 m depth, and they were immediately placed in seawater containers and transported to the experimental aquarium at the Argentinian research station Carlini. Prior to altering the pCO2, the snails were placed for seven days in an individual acclimation tank [with a continuous flow of seawater, maintained at in situ natural conditions (approximately 8.03)].
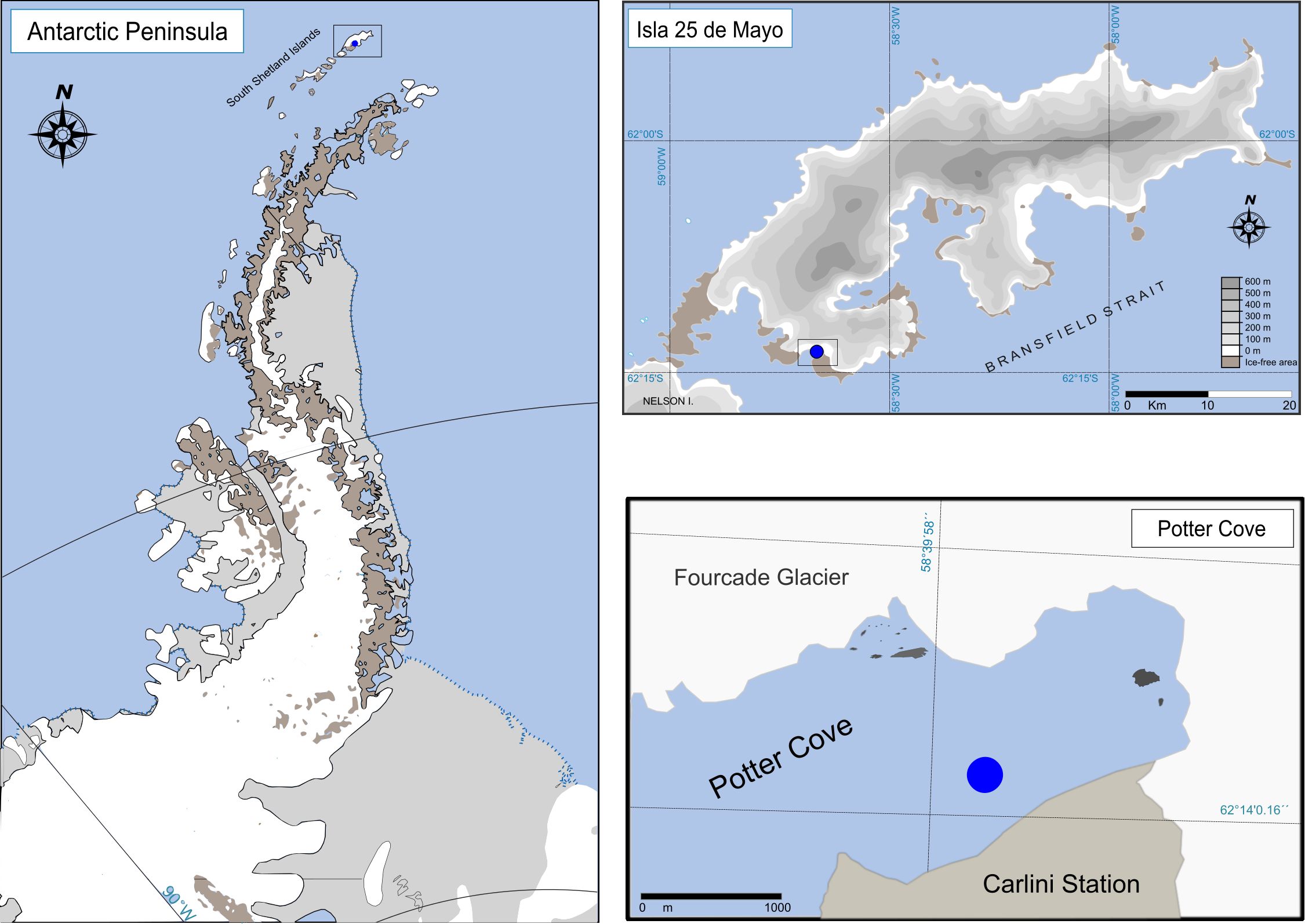
Figure 1. Study area. Antarctic Peninsula showing the location of 25 de Mayo Island. 25 de Mayo Island, with the location of Potter Cove indicated, and Potter Cove detailing the sampling area.
2.2 Experimental design
We implemented an experimental CO2-manipulation system following the same experimental design utilized in previous studies on zoo-benthic Antarctic species exposed to OA (de Aranzamendi et al., 2021; Servetto et al., 2021, 2023, 2025) (Figure 2). Seawater was continuously supplied from the field to two main header tanks (150 L each), designated as the acidified treatment (hereafter referred to as low pH, LpH) and the control. pH levels were continuously monitored using glass electrodes (LL Ecotrodeplus, Metrohm) connected to a pH controller (Consort R3610, Turnhout, Belgium). The LpH tank was gradually acidified by bubbling CO2 gas (99.9% purity) until reaching a target pH of 7.68 ± 0.17, representing ~1000 µatm pCO2, a reduction of approximately 0.3–0.4 pH units relative to the control. The control tank was maintained to reflect the natural pH variability recorded at the snails’ collection depth (15 m). It continuously received unaltered seawater directly supplied from the PC. Individual snails (N = 6; two individuals per tank) were placed in separate subsidiary smaller aquaria (~6 L), each connected to a continuous flow of seawater supplied from either the LpH or control head tank (Figure 2). The experimental sample size (5–6 cm) was carefully selected to minimize potential impacts on the individuals. As these snails are Antarctic organisms and little is known about their population in PC (Sahade et al., 2015; personal observation), a precautionary approach was taken by limiting the number of specimens collected. To minimize evaporation and reduce gas exchange with the air, each tank was covered with a methacrylate lid. Electrodes were calibrated daily using Tris buffers of known pH values, following standard procedures (SOP6a of Dickson et al., 2007). Weekly, 50 mL water samples were collected from the head tanks and fixed with HgCl2 for subsequent total alkalinity (TA) analysis. TA was quantified via Gran titration (Smith and Kinsey, 1978) employing a sample exchanger coupled to a TitroLine alpha plus titration system (SI Analytics, Mainz, Germany) equipped with an A157–1 M-DIN-ID pH electrode, and conducted in accordance with standard operating procedure SOP 3a (Dickson et al., 2007). Carbonate chemistry parameters were calculated with the CO2SYS spreadsheet (Pierrot et al., 2006), using dissociation constants for carbonate determined by Mehrbach et al. (1973) and refitted by Dickson and Millero (1987). A summary of the physicochemical parameters of seawater is provided in Table 1. Temperature and salinity were not experimentally manipulated; instead, natural conditions from the cove were maintained throughout the study. Nevertheless, both parameters were continuously monitored during the experimental period to account for natural oscillation and support data interpretation (Table 1).
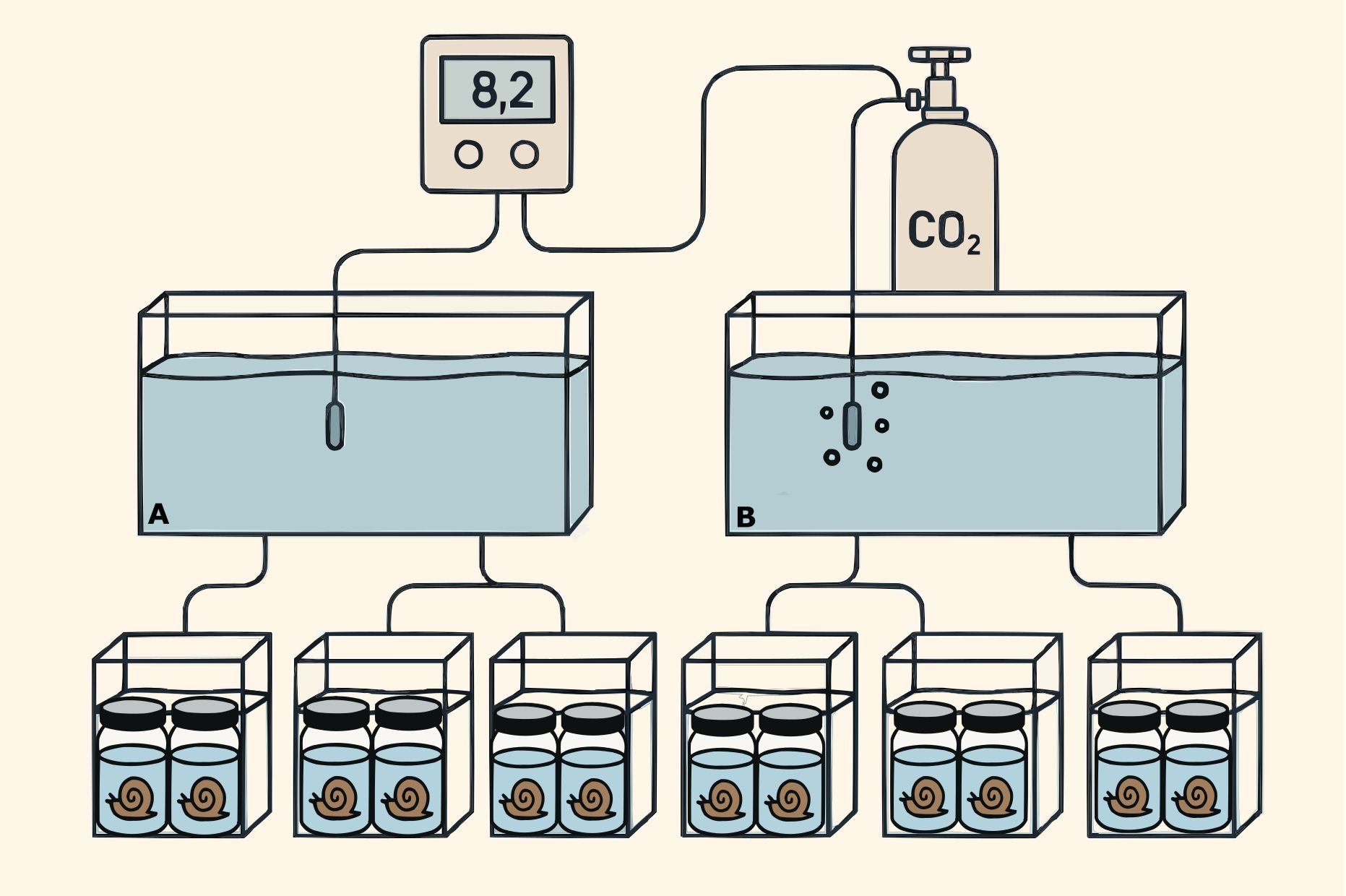
Figure 2. Experimental setup (following Servetto et al., 2021). Neobuccinum eatoni were collected by scuba divers in the Antarctic summer campaign 2015–2016 and acclimated until the start of experimentation. Seawater was continuously supplied to two head tanks: (A) control and (B) low pH head tank (7.70 ± 0.09). From each head tank, seawater was delivered to three smaller aquaria where the animals were placed separately. After exposure, tissue samples from the snail (mantle, gill, gonads, and foot) were taken for fatty acid analyses.
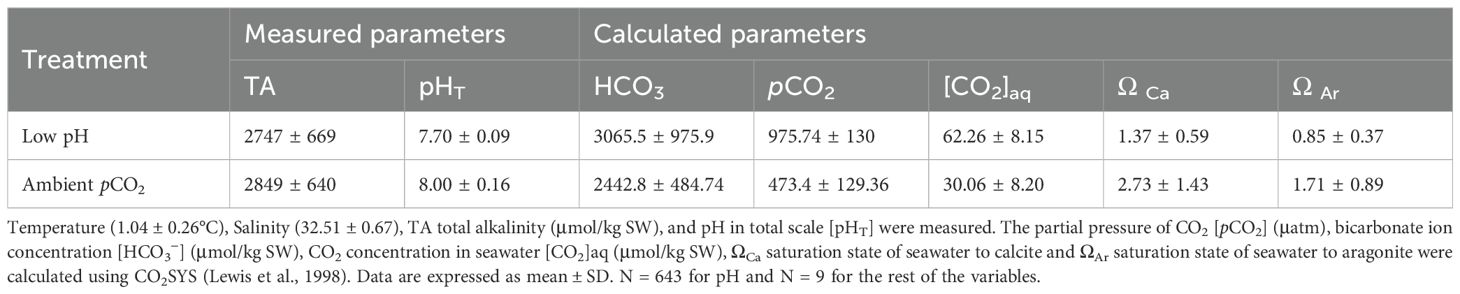
Table 1. Summary of seawater physicochemical conditions during the experiment with the Antarctic snail Neobuccinum eatoni (Servetto et al., 2023).
After two months of incubation under experimental conditions, the animals were dissected. Different tissues were selected for the FA analysis based on their functional and metabolic roles. Samples of mantle, gill, gonads, and foot were separated, stored at –80°C, and subsequently transported to Ghent University (Ghent, Belgium) for FA analysis. Results are expressed as both percentages and in μg·mg−¹ dry weight (DW).
2.3 Fatty acid profiling
FA methyl esters (FAME) were prepared via a direct transesterification procedure with 2.5% (v:v) sulfuric acid in methanol as described by De Troch et al. (2012) to achieve total FA analysis. An internal standard (FA 19:0 5 µg) was added to the freeze-dried tissue samples (~ 10 mg). FAME was extracted twice with hexane. FA composition was carried out using a gas chromatograph (GC) (HP 7890B. Agilent Technologies, Diegem. Belgium) equipped with a flame ionization detector (FID) and connected to an Agilent 5977A Mass Selective (MS) Detector (Agilent Technologies). The GC was equipped with a PTV injector (CIS-4. Gerstel. Mülheim an der Ruhr. Germany). A 60 m×0.25 mm×0.20 μm film thickness HP88 fused silica capillary column (Agilent Technologies) was used for the GC analysis at a constant Helium flow rate (2 mL min-1). The injection sample volume was 2 μL, and the oven temperature program was set as Boyen et al. (2020) described. The signal obtained with the FID detector was used to generate quantitative data on all compounds (MassHunter Quantitative Analysis Software, Agilent Technologies). Chromatogram peaks were identified based on their retention times. Quantification was done through the external standards (Supelco 37 Component FAME Mix, Sigma-Aldrich). Mean FA chain length (MCL) was calculated using the equation from Guerzoni et al. (2001):
where FAP is the percentage of fatty acid; C number of carbon atoms.
QC/QA procedures included repeated measurements of water chemistry parameters using calibrated instruments, verification of FA identification against reference standards, and cross-validation of statistical outputs. Residuals and diagnostic plots were systematically examined to ensure model assumptions were met, and all analyses were independently reproduced to confirm consistency.
2.4 Data analysis
To verify the consistency of experimental conditions, a t-test was performed on the measured water parameters (pH, CaCO3, total alkalinity (TA), and temperature) to detect any significant differences between control and LpH treatments. FA profiles of different tissues were analyzed using Principal Component Analysis (PCA), while Non-Metric Multidimensional Scaling (nMDS) was conducted separately for each tissue type. The homogeneity of variances was assessed using Bartlett’s test, and homoscedasticity and normality were further evaluated through residual analysis. Differences in FA composition among treatments were tested using an ANOVA (with tanks nested within treatments to account for potential tank effects). Additionally, a t-test was performed to compare the LpH treatment and control groups within each tissue after 66 days of the experimental condition. All statistical analyses were conducted using R version 3.6 for Microsoft Windows (R Core Team, 2020), with significance set at p < 0.05.
3 Results
3.1 Experimental conditions
The experimental setup successfully verified the targeted pH conditions (7.70 ± 0.09) despite variability in the incoming natural seawater throughout the experiment. The treatment was consistently maintained at a lower pH (7.70) compared to the control (8.00) (t-test, T = 4.94, p < 0.01) (Table 1). Additionally, the average bicarbonate ion concentration (HCO3−) and aragonite saturation state (ΩAr) differed significantly between the two conditions over the 66-day exposure period (T = -7.95, p < 0.01 and T = 2.60, p = 0.0266, respectively). In contrast (Table 1). Similarly, pCO2 (µatm) was significantly higher in the LpH treatment compared to ambient conditions (mean ± SD: 975.74 ± 61.25 vs. 473.43 ± 40.24; T = -9.45, p < 0.0001), and aqueous CO2 concentration ([CO2]aq, µmol kg−¹) also increased significantly under elevated pCO2 (62.26 ± 7.49 vs. 30.06 ± 3.99; T = -9.65, p < 0.0001). TA did not vary significantly between treatments for either variable (T = -1.53, p > 0.1407).
3.2 Fatty acid composition
A total of 20 FAs were identified in N. eatoni, grouped as saturated (SFA), monounsaturated (MUFA), and polyunsaturated (PUFA) fatty acids (Table 2). PUFA was the predominant group across all tissues (Supplementary Figure S1). FA composition varied among tissues and treatments, with the foot and mantle showing similar ranges, while the gonads exhibited the highest MUFA content (19.43–40.40%).
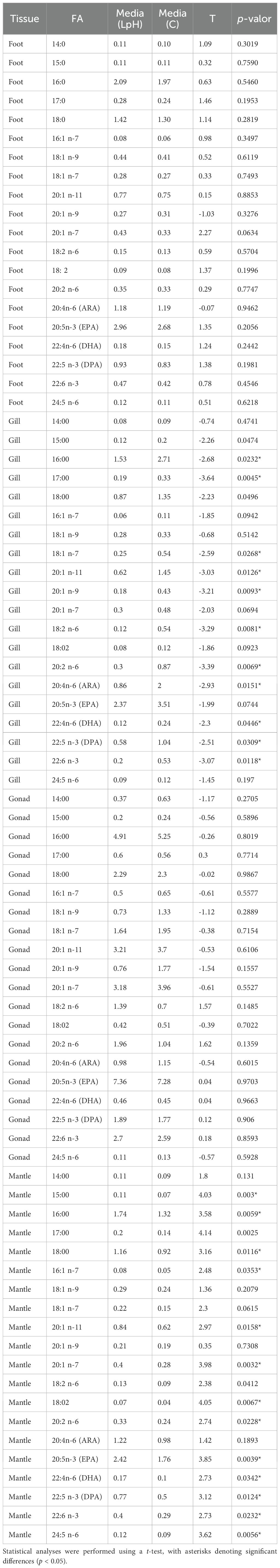
Table 2. Total concentrations (mg/µg DW) in various tissues of the Antarctic snail Neobuccinum eatoni under control conditions and low pH exposure (LpH).
Multivariate analyses supported these tissue-specific patterns. Principal Component Analysis (PCA) revealed a clear separation along PC1, differentiating the gonad’s FA composition from that of gill, foot, and mantle tissues (Figure 3). Within this framework, differences between control and LpH conditions were most evident in the gills. Similarly, non-metric Multidimensional Scaling (nMDS) indicated a marked distinction between control and LpH treatments in gill and mantle tissues, while this separation was less pronounced in the other tissues (Figure 4).
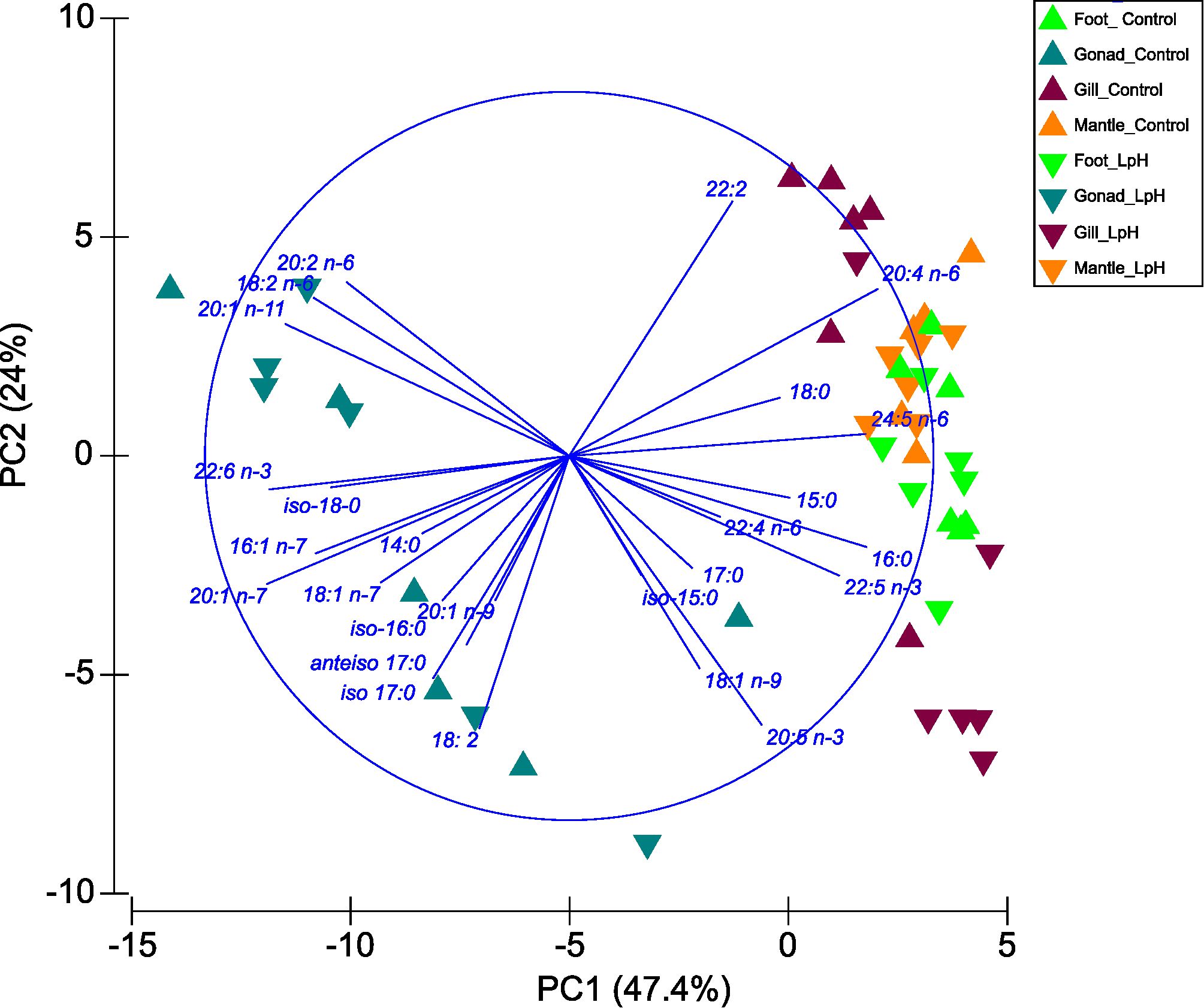
Figure 3. Principal Component Analysis (PCA) illustrating the relative (%) composition of fatty acids in different tissues (gonad, gill, mantle, and foot) of the Antarctic snail Neobuccinum eatoni under acidified conditions (LpH = low pH; (7.70 ± 0.09)) compared to control conditions.
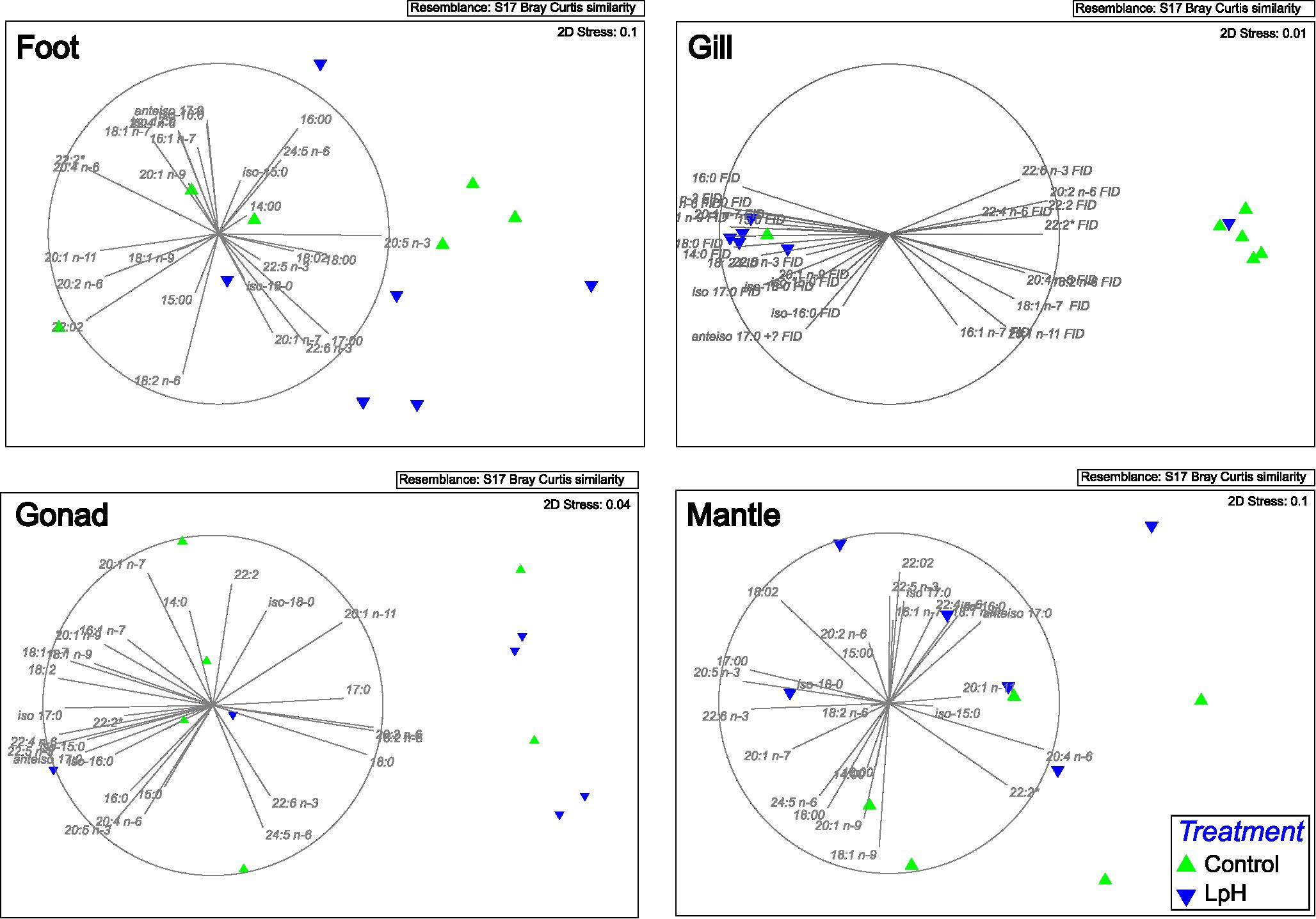
Figure 4. Non-metric Multidimensional Scaling (MDS) analysis depicting tissue fatty acid composition variability. LpH = low pH (7.70 ± 0.09).
Under LpH conditions, opposite patterns were observed in gill and mantle tissues. In the gills, total SFA, MUFA, PUFA, and LC-PUFA contents were significantly higher in the control group than in LpH-exposed snails (Figure 5; Supplementary Table S1). The PUFA/SFA and MCL ratios followed the same trend, whereas the 20:5n-3/20:4n-6 and n-3/n-6 ratios were elevated under LpH. EPA was the most abundant FA in the gills, although no significant difference was detected between treatments. Several other FAs, including 20:2n-6, ARA, DHA, 22:5n-3, and MUFAs such as 18:1n-7, 20:1n-11, and 20:1n-9, were significantly higher in control samples (Table 2).
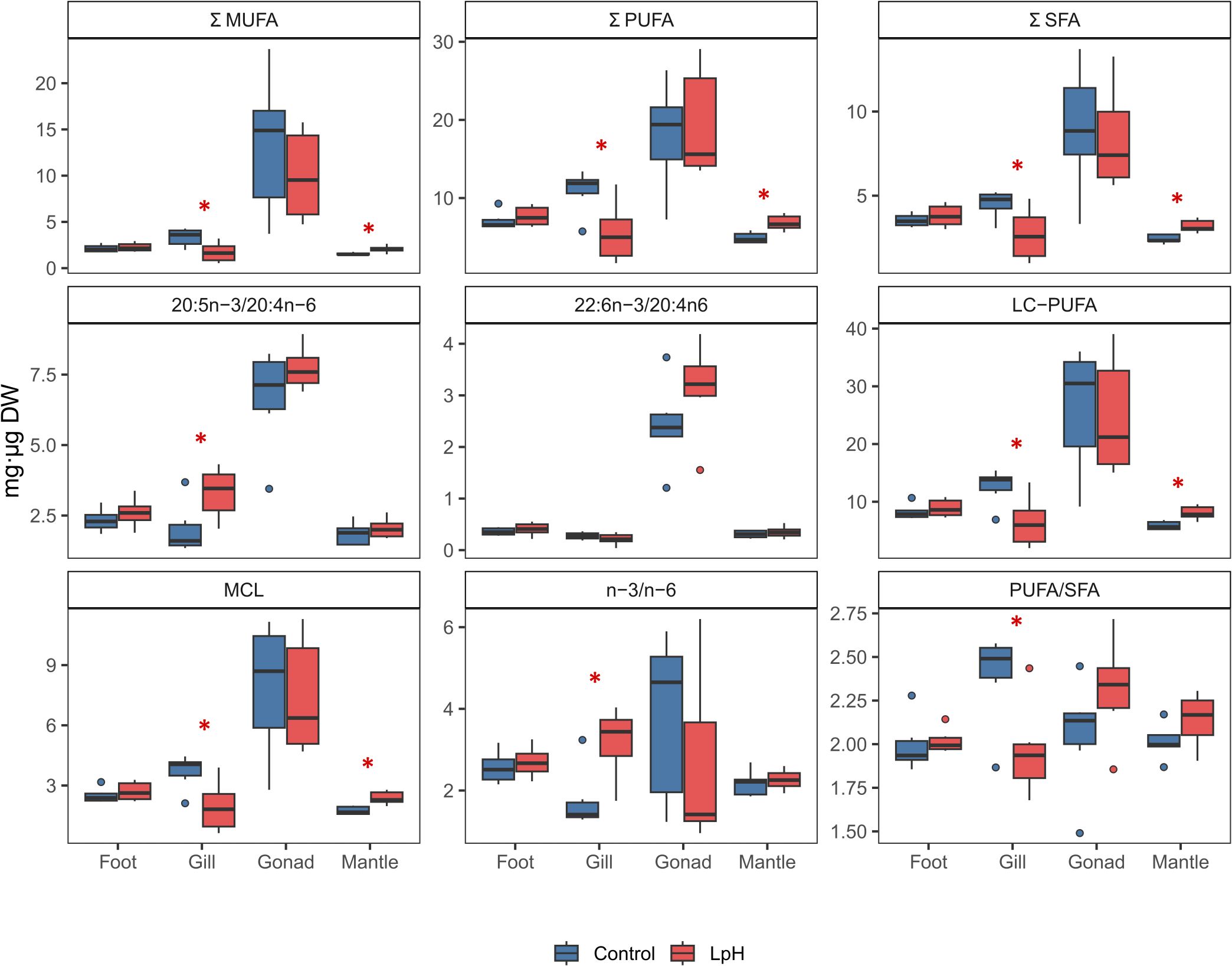
Figure 5. Fatty acid composition (mg/µg DW) across various Antarctic snail Neobuccinum eatoni tissues under low pH (LpH) (7.70 ± 0.09) and control conditions, with asterisks denoting significant differences (p < 0.05).
Conversely, in the mantle, SFA, MUFA, PUFA, and LC-PUFA levels were significantly higher under LpH exposure. The PUFA/SFA ratio and MCL indicator also increased in LpH samples compared to controls (Figure 5; Supplementary Table S1). Most mantle FAs were significantly altered by acidification, with EPA, DHA, 22:5n-3, 22:6n-3, and 24:5n-6 showing particularly elevated levels under LpH conditions (Table 2; Supplementary Table S2).
4 Discussion
This study represents the first investigation into the response of N. eatoni to projected OA scenarios, providing novel evidence of tissue-specific sensitivity through FA composition analyses. Our findings demonstrate that exposure to LpH conditions induces significant alterations in FA profiles, with distinct patterns observed between gill and mantle tissues.
4.1 Tissue-specific fatty acid profiles
FA composition varied considerably among tissues, with PUFAs predominating across all samples. The gonads displayed the highest MUFA content, likely reflecting their reproductive function and energy storage role. In contrast, the gills and mantle were more responsive to OA exposure. In gills, total SFA, MUFA, PUFA, and LC-PUFA levels were significantly reduced under LpH conditions, indicating a reorganization of FA composition in response to environmental stress. Notably, ARA (20:4 n-6) levels also decreased, which could reduce the production of eicosanoids, potentially compromising inflammatory signaling and immune plasticity (Supplementary Table S3). Conversely, in the mantle, these same FA classes were elevated under LpH. The increased LC-PUFA content in the mantles may represent an adaptive response to mitigate oxidative damage and maintain membrane integrity under environmental stress. LC-PUFAs, particularly DHA and EPA, play a crucial role in modulating oxidative stress, as they are involved in the regulation of antioxidant defense mechanisms and inflammatory responses (Monroig et al., 2013). While this mechanism has been previously documented in marine microorganisms (Okuyama et al., 2008), studies have also shown the involvement of LC-PUFAs in oxidative stress responses in marine invertebrates, such as mollusks and crustaceans (Monroig et al., 2013; Lv et al., 2021).
The observed tissue-specific variations in DHA levels between the mantle and gill tissues of N. eatoni under LpH conditions may be attributed to the distinct physiological roles and environmental exposures of these tissues. The mantle, primarily responsible for shell formation and protection, exhibited elevated DHA levels under LpH conditions, suggesting an adaptive mechanism to maintain membrane fluidity and integrity in response to environmental stressors. This lipid remodeling could enhance the mantle’s resilience to oxidative stress induced by ocean acidification. However, this strategy may involve a metabolic cost, diverting resources from other physiological processes such as growth or reproduction.
In contrast, the gills, which are directly exposed to the external environment and are the main site for respiration and ion regulation, contained higher DHA concentrations in the control samples. Under LpH conditions, the reduced DHA levels in the gills likely reflect increased lipid peroxidation caused by elevated reactive oxygen species (ROS) (Fadhlaoui and Lavoie, 2021; Lushchak, 2021). Gills are known to be particularly susceptible to environmental stressors, including changes in pH, which can lead to oxidative damage and lipid degradation (Harms et al., 2014; Zutshi et al., 2019). This susceptibility necessitates efficient antioxidant defenses to mitigate oxidative damage and maintain cellular function (Mathieu-Resuge et al., 2020). Moreover, the loss of PUFA and increased SFA content in gills could reduce membrane fluidity, potentially impairing critical physiological functions such as ion transport, respiration efficiency, and filtration. These findings underscore the importance of DHA in modulating tissue-specific responses to environmental stress, reflecting the organism’s strategic allocation of lipid resources to maintain functionality under varying conditions.
In mantle tissue, EPA levels were elevated in individuals exposed to LpH conditions. These FAs are well known for their anti-inflammatory properties and their ability to modulate membrane-associated proteins, such as ion channels and transporters (Banaszak et al., 2024). Additionally, they play a crucial role in defining key biophysical properties of biological membranes, including organization, ion permeability, elasticity, and the formation of microdomains (Bruno et al., 2007). A similar pattern was observed by Silva et al. (2017) in Gibbula umbilicalis exposed to metal contamination, further supporting the idea that DHA and EPA serve as reliable biomarkers of environmental stressors, in that particular case, metal pollutants.
Additionally, independent of the OA experiment, we observed differences in FA concentrations between the gonads and the other three analyzed tissues. These differences are primarily driven by the higher abundance of MUFAs in the gonads, where MUFAs represent the predominant FA class. The elevated MUFA suggests a critical role in reproductive processes, likely associated with energy storage and the maintenance of membrane fluidity during gametogenesis. In N. eatoni, this pattern may indicate a physiological adaptation to meet the energetic and structural requirements of gamete development. For example, Kapranova et al. (2019) reported that in the mussel Mytilus galloprovincialis, MUFA concentrations peak in male gonads at the onset of stage 2, whereas SFAs predominate in female gonads throughout stages 1 to 4. This pattern is consistent with our findings in N. eatoni, where the predominance of MUFAs in gonads suggests a comparable role of these fatty acids in supporting reproductive processes. These findings highlight the necessity for further research to elucidate the specific FA composition patterns associated with the reproductive cycle of N. eatoni and their potential ecological implications. Overall, the comparison underscores that differences in gonadal FA profiles across species can inform interpretations of reproductive status and resilience in N. eatoni under changing environmental conditions.
4.2 Immune modulation and n-3/n-6 balance
In addition to their structural and energetic roles, fatty acids contribute to immune regulation. In particular, the balance between n-3 and n-6 PUFAs influences the production of pro- and anti-inflammatory eicosanoids, as both groups compete for the same desaturation and elongation pathways (Monroig and Kabeya, 2018). In our study, the n-3/n-6 ratio remained largely unchanged in mantle, foot, and gonadal tissues, suggesting a stable immune–lipid balance under OA conditions. However, in gill tissues, this ratio was significantly elevated in the LpH group, indicative of a potential shift toward an anti-inflammatory lipid profile under acidified conditions.
Similar findings have been reported in Crassostrea gigas, where OA exposure increased n-3 PUFA content in gills, influencing immune performance and disease susceptibility (Wang et al., 2016). In N. eatoni, this tissue-specific shift may serve as a compensatory immune response to the heightened oxidative and ionic challenges posed by OA. However, whether this adjustment enhances or compromises immune competency remains to be further investigated.
4.3 Homeoviscous adaptation
Homeoviscous adaptation (HVA) refers to modifications in the chemical and mechanical properties of the lipid bilayer that help preserve membrane fluidity under changing stress conditions (Parrish, 2013). In this study, HVA biomarkers—specifically PUFA/SFA ratios and mean chain length (MCL)—revealed effects of elevated CO2 exposure in both mantle and gill tissues. In the gills, a higher PUFA/SFA index under control conditions suggests a decrease in membrane fluidity in response to acidification, potentially indicating an adaptive mechanism for maintaining membrane function. This aligns with known functions of PUFAs enhancing membrane fluidity, and SFAs contributing to membrane stability (Hąc-Wydro and Wydro, 2007). Elevated seawater pCO2 in treatment also influenced MCL in both tissues. The mantle exhibited higher MCL under LpH conditions, suggesting a reduced membrane fluidity as a compensatory response to acidification (Guerzoni et al., 2001). These findings are evidence that OA can modulate HVA pathways in polar invertebrates. In addition, under stress, HVA, through the regulation of lipid class ratios (e.g., ST/PL ratio), may be more energy-efficient than modifications in PUFA/SFA ratios and MCL. HVA pathways have been previously reported as adaptive responses to abiotic stressors in other marine organisms (Somero, 2022). For example, the impact of increasing seawater CO2 and temperature on HVA has been studied in marine sponges (Bennett et al., 2018). They found that certain species used the additional carbon to enhance sterol biosynthesis, which likely reflects an HVA mechanism, providing a potential pathway where elevated CO2 helps to mitigate thermal stress (Bennett et al., 2018).
4.4 Biochemical drivers of FA shifts
While our results support the use of FA profiles as tissue-specific biomarkers of physiological stress in N. eatoni, a more mechanistic understanding of the biochemical underpinnings driving these shifts under pCO2/pH variability is essential. FA composition in marine invertebrates is not static, but rather dynamically regulated through enzymatic pathways such as desaturation and elongation, mediated by desaturases and elongases whose activities are often sensitive to environmental stressors (Monroig et al., 2013; Bell et al., 2016). Changes in pH and CO2 availability can alter cellular acid-base balance, indirectly affecting enzymatic efficiency and gene expression linked to lipid metabolism (Tocher, 2010). Moreover, the remodeling of membrane lipid composition, particularly increases in LC-PUFAs like EPA and DHA, may reflect adaptive strategies to maintain membrane fluidity and functionality under acidified conditions, as observed in both invertebrate and vertebrate systems (Mourente et al., 2022). In gill tissues, reductions in PUFAs under LpH (7.70 ± 0.09) may be associated with impaired membrane-bound processes such as ion transport or respiration, whereas the mantle’s elevated LC-PUFA content could suggest compensatory regulation aimed at preserving cellular performance. Collectively, these tissue-specific FA shifts likely represent a balance between maintaining membrane integrity, oxidative defense, and energetic allocation, highlighting the complex trade-offs faced by N. eatoni under OA stress. Such tissue-specific responses align with broader findings across phyla, including mollusks and crustaceans, where FA remodeling serves as a plastic response to environmental change (Arts and Kohler, 2009; Soudant et al., 2020). Thus, interpreting FA shifts not solely as stress indicators, but as reflections of underlying biochemical and physiological strategies, provides a richer framework for understanding organismal resilience to ocean acidification.
5 Conclusion
This study provides new evidence of tissue-specific alterations in the FA composition of N. eatoni under LpH exposure. Significant changes were observed, particularly in mantle and gill tissues, with shifts in n-3/n-6 ratios and lipid-related indicators (e.g., PUFA/SFA ratio, MCL), suggesting possible effects on membrane properties and immune-related pathways. The depletion of ARA in gills and elevation of LC-PUFAs in the mantle highlight tissue-specific trade-offs between immune regulation and membrane resilience. While our study did not directly measure physiological processes, these FA patterns may serve as biochemical signals of tissue remodeling under OA conditions. Overall, these findings highlight that different tissues of N. eatoni respond distinctly to acidified seawater, offering valuable baseline information on potential sensitivity to OA. Given the scarcity of data on Antarctic gastropods, this work represents a step toward understanding their responses, but longer-term and integrative studies are needed to clarify the physiological and ecological implications, particularly regarding organismal performance and population resilience in polar environments. Considering the ecological and reproductive implications, longer-term and integrative studies are needed to clarify the consequences for organismal performance, survival, and population resilience in polar environments.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
NS: Formal analysis, Investigation, Methodology, Writing – original draft, Writing – review & editing, Funding acquisition, Project administration. MD: Conceptualization, Supervision, Writing – review & editing, Methodology. GA: Formal analysis, Methodology, Writing – review & editing, Conceptualization, Investigation. LF: Formal analysis, Methodology, Writing – review & editing. MD: Methodology, Writing – review & editing. RS: Conceptualization, Formal analysis, Funding acquisition, Investigation, Supervision, Writing – review & editing, Project administration.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This study was supported by PADI Foundation (#81356), Dirección Nacional del Antártico (DNA)/Instituto Antártico Argentino (IAA), Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Alfred Wegener Institute (AWI, Germany), and Universidad Nacional de Córdoba. Partial funding was provided by PICT-2018-02125, PICT-2021-I-GRF1, PICT-2020-SERIEA-02956, and the EU project CoastCarb, Marie Curie Action RISE (H2020-MCSA-RISE 872690). The research presented here was conducted with infrastructure funded by EMBRC Belgium-FWO International Research Infrastructure I001621N.
Acknowledgments
The authors extend their sincere thanks to the staff of the Carlini Station. Special thanks to Dr. Bruno Vlaeminck (UGent, Marine Biology) for his assistance with lab work.
Conflict of interest
The authors declare that no commercial or financial relationships were present that could be perceived as a potential conflict of interest in the conduct of this research.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2025.1645755/full#supplementary-material
References
Banaszak M., Dobrzyńska M., Kawka A., Górna I., Woźniak D., Przysławski J., et al. (2024). Implementation of and systems-level barriers to cancer clinical trials: a qualitative study. JCO Precis Oncol. (2024) 8:1–8. doi: 10.1200/PO.23.00518
Bell M. V., Tocher D. R., and Sargent J. R. (2016). Biosynthesis of polyunsaturated fatty acids in aquatic ecosystems: general pathways and species differences. Prog. Lipid Res. 62, 1–17. doi: 10.1016/j.plipres.2016.01.001
Bennett H., Bell J. J., Davy S. K., Webster N. S., and Francis D. S. (2018). Elucidating the sponge stress response; lipids and fatty acids can facilitate survival under future climate scenarios. Glob. Chang. Biol. 24, 3130–3144. doi: 10.1111/gcb.14116
Boyen J., Fink P., Mensens C., Hablützel P. I., and De Troch M. (2020). Fatty acid bioconversion in harpacticoid copepods in a changing environment: a transcriptomic approach. Philos. Trans. R. Soc Lond. B Biol. Sci. 375, 20190645. doi: 10.1098/rstb.2019.0645
Bruno M. J., Koeppe R. E., and Andersen O. S. (2007). Docosahexaenoic acid alters bilayer elastic properties. PNAS 104, 9638–9643. doi: 10.1073/pnas.0701015104
Capitão A., Lyssimachou A., Castro L. F. C., and Santos M. M. (2017). Obesogens in the aquatic environment: an evolutionary and toxicological perspective. Environ. Int. 106, 153–169. doi: 10.1016/j.envint.2017.06.003
Corsolini S. and Borghesi N. (2017). A comparative assessment of fatty acids in Antarctic organisms from the Ross Sea: Occurrence and distribution. Chemosphere 174, 747–753. doi: 10.1016/j.chemosphere.2017.02.031
Cummings V., Hewitt J., Van Rooyen A., Currie K., Beard S., Thrush S., et al. (2011). Ocean acidification at high latitudes: potential effects on functioning of the Antarctic bivalve Laternula elliptica. PloS One 6, e16069. doi: 10.1371/journal.pone.0016069
de Aranzamendi M. C., Servetto N., Movilla J., Bettencourt R., and Sahade R. (2021). Ocean acidification effects on the stress response in a calcifying Antarctic coastal organism: The case of Nacella concinna ecotypes. Mar. pollut. Bull. 166, 112218. doi: 10.1016/j.marpolbul.2021.112218
de Carvalho G. T., Pezzi L. P., Lefèvre N., Rodrigues C. C. F., Santini M. F., and Mejia C. (2025). Spatio-temporal variability in CO2 fluxes in the Atlantic sector of the southern ocean. Atmosphere 16, 319. doi: 10.3390/atmos16030319
Dennis M. M., Molnár K., Kriska G., and Lőw P. (2021). Mollusca: gastropoda. Histol. Invertebr., 87–132. doi: 10.1002/9781119507697.ch4
De Troch M., Boeckx P., Cnudde C., Van Gansbeke D., Vanreusel A., Vincx M., et al. (2012). Bioconversion of fatty acids at the basis of marine food webs: insights from a compound-specific stable isotope analysis. Mar. Ecol. Prog. Ser. 465, 53–67. doi: 10.3354/meps09920
Dickson A. G., Sabine C. L., and Christian J. R. (2007). Guide to best practices for ocean CO2 measurements (North Pacific Marine Science Organization, Canada: PICES Special Publication), 191.
Dickson A. G. and Millero F. J. (1987). A comparison of the equilibrium constants for the dissociation of carbonic acid in seawater media. Deep-Sea Res. I: Oceanogr. Res. Pap. 34, 1733–1743. doi: 10.1016/0198-0149(87)90021-5
Ericson J. A., Hellessey N., Kawaguchi S., Nichols P. D., Nicol S., Hoem N., et al. (2019). Near-future ocean acidification does not alter the lipid content and fatty acid composition of adult Antarctic krill. Sci. Rep. 9 (1), 12375. doi: 10.1038/s41598-019-48665-5
Fabry V. J., Seibel B. A., Feely R. A., and Orr J. C. (2008). Impacts of ocean acidification on marine fauna and ecosystem processes. ICES J. Mar. Sci. 65, 414–432. doi: 10.1093/icesjms/fsn048
Fadhlaoui M. and Lavoie I. (2021). Effects of temperature and glyphosate on fatty acid composition, antioxidant capacity, and lipid peroxidation in the gastropod lymneae sp. Water 13 (8), 1039. doi: 10.3390/w13081039
Findlay H. S. and Turley C. (2021). Ocean acidification and climate change, in: Climate Change: Observed Impacts on Planet Earth. 3rd ed. (Elsevier), 251–279. doi: 10.1016/B978-0-12-821575-3.00013-X
Fisher B. J., Poulton A. J., Meredith M. P., Baldry K., Schofield O., and Henley S. F. (2025). Climate-driven shifts in Southern Ocean primary producers and biogeochemistry in CMIP6 models. Biogeosciences 22, 975–994. doi: 10.5194/bg-22-975-2025
Gao Y., Zheng S. C., Zheng C. Q., Shi Y. C., Xie X. L., Wang K. J., et al. (2018). The immune-related fatty acids are responsive to CO2-driven seawater acidification in a crustacean brine shrimp Artemia sinica. Dev. Comp. Immunol. 81, 342–347. doi: 10.1016/j.dci.2017.12.022
Gattuso J. P., Magnan A., Billé R., Cheung W. W., Howes E. L., Joos F., et al. (2015). Contrasting futures for ocean and society from different anthropogenic CO2 emissions scenarios. Science 349, aac4722. doi: 10.1126/science.aac4722
Gibbs M. C., Parker L. M., Scanes E., Byrne M., O’Connor W. A., and Ross P. M. (2021). Energetic lipid responses of larval oysters to ocean acidification. Mar. pollut. Bull. 168, 112441. doi: 10.1016/j.marpolbul.2021.112441
Gingerich P. D. (2019). Temporal scaling of carbon emission and accumulation rates: modern anthropogenic emissions compared to estimates of PETM onset accumulation. Paleoceanogr. Paleoclimatol. 34, 329–335. doi: 10.1029/2018PA003379
Gladyshev M. I., Sushchik N. N., and Makhutova O. N. (2013). Production of EPA and DHA in aquatic ecosystems and their transfer to the land. PGs 107, 117–126. doi: 10.1016/j.prostaglandins.2013.03.002
González R., Pertierra L. R., Guerrero P. C., and Díaz A. (2024). High vulnerability of the endemic Southern Ocean snail Neobuccinum eatoni (Buccinidae) to critical projected oceanographic changes. Sci. Rep. 15, 3799. doi: 10.1038/s41598-024-80353-x
Guerzoni M. E., Lanciotti R., and Cocconcelli P. S. (2001). Alteration in cellular fatty acid composition as a response to salt, acid, oxidative and thermal stresses in Lactobacillus helveticus. Microbiology 147, 2255–2264. doi: 10.1099/00221287-147-8-2255
Hąc-Wydro K. and Wydro P. (2007). The influence of fatty acids on model cholesterol/phospholipid membranes. Chem. Phys. Lipids. 150, 66–81. doi: 10.1016/j.chemphyslip.2007.06.213
Hancock A. M., King C. K., Stark J. S., McMinn A., and Davidson A. T. (2020). Effects of ocean acidification on Antarctic marine organisms: A meta-analysis. Ecol. Evol. 10, 4495–4514. doi: 10.1002/ece3.6205
Harms L., Frickenhaus S., Schiffer M., Mark F. C., Storch D., Held C., et al. (2014). Gene expression profiling in gills of the great spider crab Hyas araneus in response to ocean acidification and warming. BMC Genomics 15, 1–17. doi: 10.1186/1471-2164-15-789
Hauri C., Doney S. C., Takahashi T., Erickson M., Jiang G., and Ducklow H. W. (2015). Two decades of inorganic carbon dynamics along the West Antarctic Peninsula. BG 12, 6761–6779. doi: 10.5194/bg-12-6761-2015
Hedberg P., Lau D. C. P., Albert S., and Winder M. (2023). Variation in fatty acid content among benthic invertebrates in a seasonally driven system. Limnol. Oceanogr. Lett. 8, 751–759. doi: 10.1002/lol2.10333
IPCC (2019). IPCC Special Report on the Ocean and Cryosphere in a Changing Climate Pö̈rtner H.-O., Roberts D.C., Masson-Delmotte V., Zhai P., Tignor M., Poloczanska E., Mintenbeck K., Alegría A., Nicolai M., et al (eds.) Cambridge University Press, Cambridge, UK and New York, NY, USA, 755 pp. doi: 10.1017/9781009157964
Johnson K. M. and Hofmann G. E. (2017). Transcriptomic response of the Antarctic pteropod Limacina helicina Antarctica to ocean acidification. BMC Genomics 18, 1–16. doi: 10.1186/s12864-017-4161-0
Kapranova L. L., Nekhoroshev M. V., Malakhova L. V., Ryabushko V. I., Kapranov S. V., and Kuznetsova T. V. (2019). Fatty acid composition of gonads and gametes in the Black Sea bivalve mollusk Mytilus galloprovincialis Lam. at different stages of sexual maturation. J. Evol. Biochem. Phys. 55, 448–455. doi: 10.1134/S0022093019060024
Kroeker K. J., Kordas R. L., Crim R., Hendriks I. E., Ramajo L., Singh G. S., et al. (2013). Impacts of ocean acidification on marine organisms: quantifying sensitivities and interaction with warming. Glob. Change Biol. 19, 1884–1896. doi: 10.1111/gcb.12179
Lushchak V. I. (2021). Interplay between bioenergetics and oxidative stress at normal brain aging. Aging as a result of increasing disbalance in the system oxidative stress–energy provision. Pflugers Arch. 473, 713–722. doi: 10.1007/s00424-021-02531-4
Lv D., Zhang F., Ding J., Chang Y., and Zuo R. (2021). Effects of dietary n-3 LC-PUFA on the growth performance, gonad development, fatty acid profile, transcription of related genes and intestinal microflora in adult sea urchin (Strongylocentrotus intermedius). Aquac. Res. 52, 1431–1441. doi: 10.1111/are.14997
Mathieu-Resuge M., Le Grand F., Schaal G., Lluch-Cota S. E., Racotta I. S., and Kraffe E. (2020). Specific regulations of gill membrane fatty acids in response to environmental variability reveal fitness differences between two suspension-feeding bivalves (Nodipecten subnodosus and Spondylus crassisquama). Conserv. Physiol. 8. doi: 10.1093/conphys/coaa079
Mehrbach C., Culberson C. H., Hawley J. E., and Pytkowicx R. M. (1973). Measurement of the apparent dissociation constants of carbonic acid in seawater at atmospheric pressure. Limnol. Oceanogr. 18, 897–907. doi: 10.4319/lo.1973.18.6.0897
Monroig Ó. and Kabeya N. (2018). Desaturases and elongases involved in polyunsaturated fatty acid biosynthesis in aquatic invertebrates: a comprehensive review. Fish. Sci. 84, 911–928. doi: 10.1007/s12562-018-1254-x
Monroig Ó., Tocher D. R., and Navarro J. C. (2013). Biosynthesis of polyunsaturated fatty acids in marine invertebrates: recent advances in molecular mechanisms. Mar. Drugs 11, 3998–4018. doi: 10.3390/md11103998
Mourente G., Bell J. G., and Tocher D. R. (2022). Polyunsaturated fatty acids and eicosanoids in marine invertebrates: recent advances and perspectives. Prog. Lipid Res. 86, 101166. doi: 10.1016/j.plipres.2022.101166
Norkko A., Thrush S. F., Cummings V. J., Gibbs M. M., Andrew N. L., Norkko J., et al. (2007). Trophic structure of coastal Antarctic food webs associated with changes in sea ice and food supply. Ecology 88, 2810–2820. doi: 10.1890/06-1396.1
Okuyama H., Orikasa Y., and Nishida T. (2008). Significance of antioxidative functions of eicosapentaenoic and docosahexaenoic acids in marine microorganisms. Appl. Environ. Microbiol. 74, 570–574. doi: 10.1128/AEM.02256-07
Orr J. C., Fabry V. J., Aumont O., Bopp L., Doney S. C., Feely R. A., et al. (2005). Anthropogenic ocean acidification over the twenty-first century and its impact on calcifying organisms. Nature 437, 681–686. doi: 10.1038/nature04095
Parrish C. C. (2013). Lipids in marine ecosystems. Int. Sch. Res. Notices., 2356–7872. doi: 10.5402/2013/604045
Pierrot D., Lewis E., and Wallace D. W. R. (2006). CO2SYS DOS Program Developed for CO2 System Calculations (Oak Ridge, TN: ORNL/CDIAC-105. Carbon Dioxide Information Analysis Center, Oak Ridge National Laboratory, US Department of Energy).
R Core Team. (2020). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Available at: https://www.R-project.org.
Schiaparelli S., Lörz A. N., and Cattaneo-Vietti R. (2006). Diversity and distribution of mollusc assemblages on the Victoria Land coast and the Balleny Islands, Ross Sea, Antarctica. Antarct. Sci. 18, 615–631. doi: 10.1017/S0954102006000654
Schmitz G. and Ecker J. (2008). The opposing effects of n-3 and n-6 fatty acids. Prog. Lipid Res. 47, 147–155. doi: 10.1016/j.plipres.2007.12.004
Sahade R., Lagger C., Torre L., Momo F., Monien P., Schloss I., et al. (2015). Climate change and glacier retreat drive shifts in an Antarctic benthic ecosystem. Sci. Adv. 1, e1500050. doi: 10.1126/sciadv.1500050
Servetto N., de Aranzamendi M. C., Bettencourt R., Held C., Abele D., Movilla J., et al. (2021). Molecular mechanisms underlying responses of the Antarctic coral Malacobelemnon daytoni to ocean acidification. Mar. Environ. Res. 170, 105430. doi: 10.1016/j.marenvres.2021.105430
Servetto N., De Troch M., Gazeau F., de Aranzamendi C., Alurralde G., González G., et al. (2025). Fatty acid response of calcifying benthic Antarctic species to ocean acidification and warming. Mar. pollut. Bull. 217, 118111. doi: 10.1016/j.marpolbul.2025.118111
Servetto N., Ruiz M. B., Martínez M., Harms L., de Aranzamendi M. C., Alurralde G., et al. (2023). Molecular responses to ocean acidification in an Antarctic bivalve and an ascidian. Sci. Total Environ. 903, 166577. doi: 10.1016/j.scitotenv.2023.166577
Shi Y. and Li Y. (2024). Impacts of ocean acidification on physiology and ecology of marine invertebrates: a comprehensive review. Aquat. Ecol. 58, 207–226. doi: 10.1007/s10452-023-10058-2
Silva C. O., Simões T., Novais S. C., Pimparel I., Granada L., Soares A. M., et al. (2017). Fatty acid profile of the sea snail Gibbula umbilicalis as a biomarker for coastal metal pollution. Sci. Total Environ. 586, 542–550. doi: 10.1016/j.scitotenv.2017.02.015
Smith S. V. and Kinsey D. (1978). “Calcification and organic carbon metabolism as indicated by carbon dioxide,” in Coral Reef: Research Methods, UNESCO Monographs on Oceanographic Methology, vol. 5 . Eds. Stoddart D.-R. and Johannes R. E., 462–484.
Somero G. N. (2022). Solutions: how adaptive changes in cellular fluids enable marine life to cope with abiotic stressors. Mar. Life Sci. Technol. 4, 389–413. 3. doi: 10.1007/s42995-022-00140-3
Soudant P., Gonçalves J. G., Quere C., and Rittschof D. (2020). Environmental effects on fatty acid composition and functional implications in marine bivalves. Front. Physiol. 11. doi: 10.3389/fphys.2020.576237
Stanley-Samuelson D. W., Jurenka R. A., Cripps C., Blomquist G. J., and de Renobales M. (1988). Fatty acids in insects: composition, metabolism, and biological significance. Arch. Insect Biochem. Physiol. 9, 1–33. doi: 10.1002/arch.940090102
Teixidó N., Carlot J., Alliouane S., Ballesteros E., De Vittor C., Gambi M. C., et al. (2024). Functional changes across marine habitats due to ocean acidification. Glob. Change Biol. 30, e17105. doi: 10.1111/gcb.17105
Tocher D. R. (2010). Fatty acid requirements in ontogeny of marine and freshwater fish. Aquac. Res. 41, 717–732. doi: 10.1111/j.1365-2109.2008.02150.x
Turner L. M., Ricevuto E., Massa Gallucci A., Lorenti M., Gambi M. C., and Calosi P. (2016). Metabolic responses to high pCO2 conditions at a CO2 vent site in juveniles of a marine isopod species assemblage. Mar. Biol. 163, 1–11. doi: 10.1007/s00227-016-2984-x
Valles-Regino R., Tate R., Kelaher B., Savins D., Dowell A., and Benkendorff K. (2015). Ocean warming and CO2-induced acidification impact the lipid content of a marine predatory gastropod. Mar. Drugs 13, 6019–6037. doi: 10.3390/md13106019
Van Anholt R. D., Spanings F. A. T., Koven W. M., Nixon O., and Wendelaar Bonga S. E. (2004). Arachidonic acid reduces the stress response of gilthead seabream Sparus aurata L. J. Exp. Biol. 207, 3419–3430. doi: 10.1242/jeb.01166
Wang Q., Cao R., Ning X., You L., Mu C., Wang C., et al. (2016). Effects of ocean acidification on immune responses of the Pacific oyster Crassostrea gigas. Fish Shellfish Immunol. 49, 24–33. doi: 10.1016/j.fsi.2015.12.025
Wang S. T., Yang Q., Liu M. K., Li L., Wang W., Zhang S. D., et al. (2025). Comparative transcriptomic analysis reveals a differential acid response mechanism between estuarine oyster (Crassostrea ariakensis) and Pacific oyster (Crassostrea gigas). Ecotoxicol. Environ. Saf. 297, 118210. doi: 10.1016/j.ecoenv.2025.118210
Zhukova N. V. (2019). Fatty acids of marine mollusks: Impact of diet, bacterial symbiosis and biosynthetic potential. Biomolecules 9, 857. doi: 10.3390/biom9120857
Keywords: Southern Ocean, gastropod, CO2 anthropogenic emissions, lipid biochemistry, benthos
Citation: Servetto N, De Troch M, Alurralde G, Ferrero L, de Aranzamendi MC and Sahade R (2025) Effects of ocean acidification on fatty acid composition in the Antarctic snail Neobuccinum eatoni. Front. Mar. Sci. 12:1645755. doi: 10.3389/fmars.2025.1645755
Received: 12 June 2025; Accepted: 13 October 2025;
Published: 29 October 2025.
Edited by:
Eva Chatzinikolaou, Hellenic Centre for Marine Research (HCMR), GreeceReviewed by:
M. Roberto García-Huidobro, Universidad Santo Tomás, ChileGurucharan Sudarshan, Ben-Gurion University of the Negev, Israel
Copyright © 2025 Servetto, De Troch, Alurralde, Ferrero, de Aranzamendi and Sahade. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ricardo Sahade, cnNhaGFkZUB1bmMuZWR1LmFy; Natalia Servetto, bnNlcnZldHRvQG1pLnVuYy5lZHUuYXI=