 Özay Köse
Özay Köse- Faculty of Fisheries and Aquaculture, Recep Tayyip Erdogan University, Rize, Türkiye
Introduction: This study investigated the effects of a high-fat diet (HFD) on hepatic steatosis and the protective role of quercetin (3,3′,4′,5,7-pentahydroxyflavon) in juvenile female rainbow trout (Oncorhynchus mykiss).
Methods: A total of 270 fish were randomly divided into three dietary groups and fed for 8 weeks: low-fat diet (LFD; 11.38% crude fat, 0 g/kg quercetin), high-fat diet (HFD; 22.53% crude fat, 0 g/kg quercetin), and high-fat diet supplemented with quercetin (HFD+Q; 22.33% crude fat, 0.20 g/kg quercetin). Growth performance, serum biochemical parameters, hepatic histomorphology, antioxidant enzyme gene expressions, and lipid metabolism-related gene expressions were evaluated.
Results: HFD and HFD+Q diets positively supported fish growth with a protein-sparing effect. However, it increased organosomatic indices, but this increase was limited to HFD+Q supplemented with quercetin. In this study, a high-fat diet (HFD) induced hepatic steatosis characterized by significant lipid accumulation, elevated non-esterified free fatty acid (NEFA) levels in liver tissue (p<0.05), increased serum levels of low-density lipoprotein (LDL), cholesterol, and triglycerides (TG), structural alterations in liver histomorphology such as hepatocyte vacuolization, nuclear degeneration, and sinusoidal dilation, enhanced activities of hepatic enzymes including alanine aminotransferase (ALT), aspartate aminotransferase (AST), alkaline phosphatase (ALP), and lactate dehydrogenase (LDH), as well as upregulation of cysteine-aspartate-specific protease-3 (caspase-3) mRNA expression (p<0.05). Quercetin prevented and alleviated the main metabolic and pathological changes induced by the HFD. Quercetin supplementation significantly reduced serum lipid profiles, hepatic lipid accumulation, NEFA levels, ALT, AST, ALP, and LDH enzyme activities, and downregulated caspase-3 gene expression compared to the HFD group (p<0.05). Gene expression analysis revealed that quercetin upregulated lipolysis and β-oxidation-related genes including peroxisome proliferator-activated receptor alpha (ppar-α), carnitine palmitoyltransferase-1a (cpt-1a), and hormone-sensitive lipase (hsl), while downregulating lipogenesis-associated genes fatty acid synthase (fas) and lipoprotein lipase (lpl) (p<0.05). Regarding antioxidant defense, quercetin supplementation decreased superoxide dismutase (sod) and glutathione S-transferase (gst) mRNA levels, increased catalase (cat) expression, but the suppression of glutathione peroxidase (gpx) expression persisted (p<0.05).
Discussion: These findings suggest that the HFD diet disrupts oxidative balance by increasing oxidative stress and impairing antioxidant systems, while quercetin ameliorates oxidative imbalance and mitigates hepatic damage. In conclusion, quercetin exerts hepatoprotective effects against HFD-induced hepatic steatosis by targeting lipid accumulation, oxidative stress, and apoptosis pathways, supporting its potential as a dietary supplement to prevent hepatic steatosis in aquaculture.
1 Introduction
Türkiye is the largest producer of Oncorhynchus mykiss in Europe and the second largest globally (Yıldırım and Çantaş, 2022). Oncorhynchus mykiss exceeding 3000 g, cultivated in Türkiye for fillet production, is marketed internationally under the brand name “Turkish Salmon” (Çakmak et al., 2024). In aquaculture practices in Türkiye, an all-female population is preferred to optimize growth and feed costs, with fish being reared in sea or freshwater cages for 12–16 months and fed high-fat diets (minimum 20–22%). A dietary lipid level of at least 22.8% is necessary to maintain better organoleptic properties and nutritional quality in triploid rainbow trout (Meng et al., 2023). Since protein supplementation in fish feeds is an expensive resource, it is not cost-effective to use protein as an energy source. Instead, high-fat diets are commonly utilized in aquaculture due to their cost efficiency and energy benefits (Zou et al., 2022). Replacing fish oil with specific levels of dietary and vegetable oils has been shown to promote rapid fish growth while reducing feed costs. In the diets of many carnivorous fish species, including Oncorhynchus mykiss, Salmo salar, Salvelinus fontinalis, Sparus aurata, and Dicentrarchus labrax, vegetable oils can be incorporated in specific proportions as fish oil substitute (Shahrooz et al., 2018; Xu et al., 2022) However, long-term use of high-fat diets can lead to excessive fat accumulation in the liver, adipose tissue, and other organs of cultured fish, resulting in impaired lipid metabolism and the development of fatty liver syndrome (steatosis) (Jin et al., 2019; Liu et al., 2022; Tao et al., 2018).
Steatosis, provokes pathological changes in the liver, being a significant threat to aquaculture (Zou et al., 2021). Fatty liver disease encompasses a spectrum of hepatic histological changes, ranging from simple hepatic steatosis to steatohepatitis, which is characterized by cell death and fibrosis. This condition results from the excessive accumulation of triglycerides (TG) in the liver (Ahmed et al., 2015; Pagliassotti, 2012). Hepatic steatosis arises due to an imbalance between lipogenesis and lipolysis, leading to increased production of reactive oxygen species (ROS), oxidative stress, inflammation, and apoptosis (Jia et al., 2020; Li et al., 2020). Lipolysis involves the hydrolysis of TG, releasing fatty acids and glycerol into circulation, while lipogenesis consists of a series of processes that facilitate TG biosynthesis and lipid droplet expansion within adipose tissue (Saponaro et al., 2015). In fish, hepatic lipid accumulation and metabolism are significantly influenced by dietary protein, lipid, and carbohydrate levels (N. Xie et al., 2021). High-fat diets (HFDs) adversely affect fish health by directly impacting TG and cholesterol levels in the liver (Xu et al., 2022). Recent studies have demonstrated that HFDs negatively affect cultured fish species, such as Oreochromis niloticus and Ctenopharyngodon idella, decreasing growth performance, affecting hematological parameters, impairing lipid metabolism and suppressing immune responses (Ma et al., 2018; Tang et al., 2019).
In Oncorhynchus mykiss growth, various plants such as Taraxacum officinale (Köse et al., 2024) and Camellia sinensis (Boran et al., 2015), as well as plant-derived bioactive compounds silymarin (Abdel-Latif et al., 2023) and quercetin (Sukhovskaya et al., 2023) have been successfully used as alternatives to chemical drugs. Quercetin exhibits antioxidant (Rajesh et al., 2015), anti-inflammatory, antiapoptotic, hepatoprotective, renoprotective, neuroprotective (Maalik et al., 2014), antibacterial, antiproliferative (Ruiz et al., 2015), anti-allergic (Chirumbolo, 2010), antiviral and immunomodulatory (Park et al., 2003) properties. Additionally, it modulates the expression of catalase (CAT) and superoxide dismutase (SOD), increases intracellular glutathione (GSH) levels (Ruiz et al., 2015), inhibits lipoprotein oxidation and cytotoxicity (Miean and Mohamed, 2001), and regulates the cell cycle (Rajesh et al., 2015). In cases of liver damage, quercetin effectively inhibits lipid peroxidation and mitigates oxidative stress-related consequences. It stabilizes malondialdehyde (MDA) levels, which contribute to toxic stress in cells, and reduces lipid peroxidation, thereby protecting hepatic tissue from further damage (Farmer and Davoine, 2007).
In this study, the effects of dietary quercetin supplementation (3,3′,4′,5,7-pentahydroxyflavone) on hepatic steatosis, growth performance, and antioxidant defense in high-fat diet-fed rainbow trout (Oncorhynchus mykiss) were investigated. The study assessed physiological and biochemical alterations induced by a high-fat diet and examined histological changes in the liver. Furthermore, gene expressions related to lipid metabolism, lipogenesis, and lipolysis was analyzed, including fatty acid synthase (fas), sterol regulatory element-binding protein-1 (serbp-1), peroxisome proliferator-activated receptor alpha (ppar-α), carnitine palmitoiltransferase-1 (cpt-1), hormone-sensitive lipase (hsl), and lipoprotein lipase (lpl). Antioxidant-related genes including catalase (cat), superoxide dismutase (sod), glutathione peroxidase (gpx), and glutathione-S-transferase (gst), as well as the apoptosis-related gene caspase-3, were also investigated. To date, no studies have specifically explored the effects of quercetin on high-fat diet-induced hepatic steatosis in juvenile female Oncorhynchus mykiss. While the antioxidant and hepatoprotective roles of quercetin are well-documented in other animal models, its effects on diet-induced lipid metabolic disorders and associated gene expression in Oncorhynchus mykiss remain unexamined. Therefore, this study aims to fill a critical gap in the aquaculture and fish physiology literature by evaluating the potential of this natural plant-derived compound as a preventive strategy against diet-induced hepatic lipid accumulation a growing concern in intensive aquaculture systems.
2 Materials and methods
This study was approved by the Recep Tayyip Erdoğan University Experimental Animals Ethics Committee (Decision No: 2023/30, Date: 13.06.2023) and conducted in accordance with its guidelines.
2.1 Experimental fish, acclimation, and rearing conditions
A total of 540 female rainbow trout (Oncorhynchus mykiss) were obtained from a local fish farm (Biberoglu Kardesler Trout Production Farm, Rize/Pazar, Türkiye) and transported to the Recep Tayyip Erdoğan University Iyidere R&D Unit. Upon arrival at the facility, the fish (mean: 37.12 ± 3.50 g) were fasted for 24 hours to acclimate to the experimental conditions. Following this acclimation period, they were fed once daily to apparent satiation for 14 consecutive days using a commercial diet containing 45% crude protein and 20% crude lipid.
At the end of the acclimation period, all fish were fasted for 48 hours and weighed. From the total of 540 fish, 270 individuals were randomly selected and distributed equally into three experimental groups: Low-Fat Diet (LFD), High-Fat Diet (HFD), and High-Fat Diet supplemented with Quercetin (HFD+Q). Each group consisted of three replicate tanks (n = 30 fish per tank), and the initial body weights were 38.40 ± 0.15 g, 38.36 ± 0.11 g, and 38.36 ± 0.17 g for the LFD, HFD, and HFD+Q groups, respectively.
The feeding trial began on March 1, 2024, and lasted for 8 weeks. Fish were maintained in 100 L fiberglass tanks supplied with aeration and a water flow rate of 0.2 L/sec. Natural daylight was used. Fish were fed daily at 2% of their body weight, divided into three meals (9:00 AM, 1:00 PM, and 5:00 PM). Environmental parameters were recorded daily as follows: temperature 14.12 ± 1.78°C, pH 7.74 ± 0.18, conductivity 140.34 ± 9.35 µS/cm, total dissolved solids (TDS) 70.2 ± 4.69 mg/L, and dissolved oxygen 7.72 ± 0.62 mg/L. Biometric measurements (total length and body weight) were taken every 15 days and expressed as mean ± standard deviation (± SD) using the International System of Units (SI).
2.2 Diet formulation and preparation
The formulation and proximate composition of the trial diets are shown in Table 1. The three experimental diets were formulated based on their lipid content: Low-Fat Diet (LFD) containing 11.38% lipid, High-Fat Diet (HFD) with 22.53% lipid, and High-Fat Diet supplemented with Quercetin (>95% purity, Sigma Chemical Co., USA) (HFD+Q) containing 22.33% lipid. The protein levels of these diets were 45.91%, 45.96%, and 45.94%, respectively. Additionally, the HFD+Q diet was supplemented with 0.20 g kg1 quercetin (3,3′,4′,5,7-pentahydroxyflavone). Since no previous study has investigated the effects of quercetin-supplemented diets on steatosis in Oncorhynchus mykiss, the quercetin level used in this study was selected based on the effective dose range (150–600 mg/kg) reported in Channa argus (Kong et al., 2022) and Cyprinus carpio (Armobin et al., 2023), as well as studies in mice indicating that dietary supplementation with 250 mg/kg quercetin posed no toxicity risk (Cunningham et al., 2022).
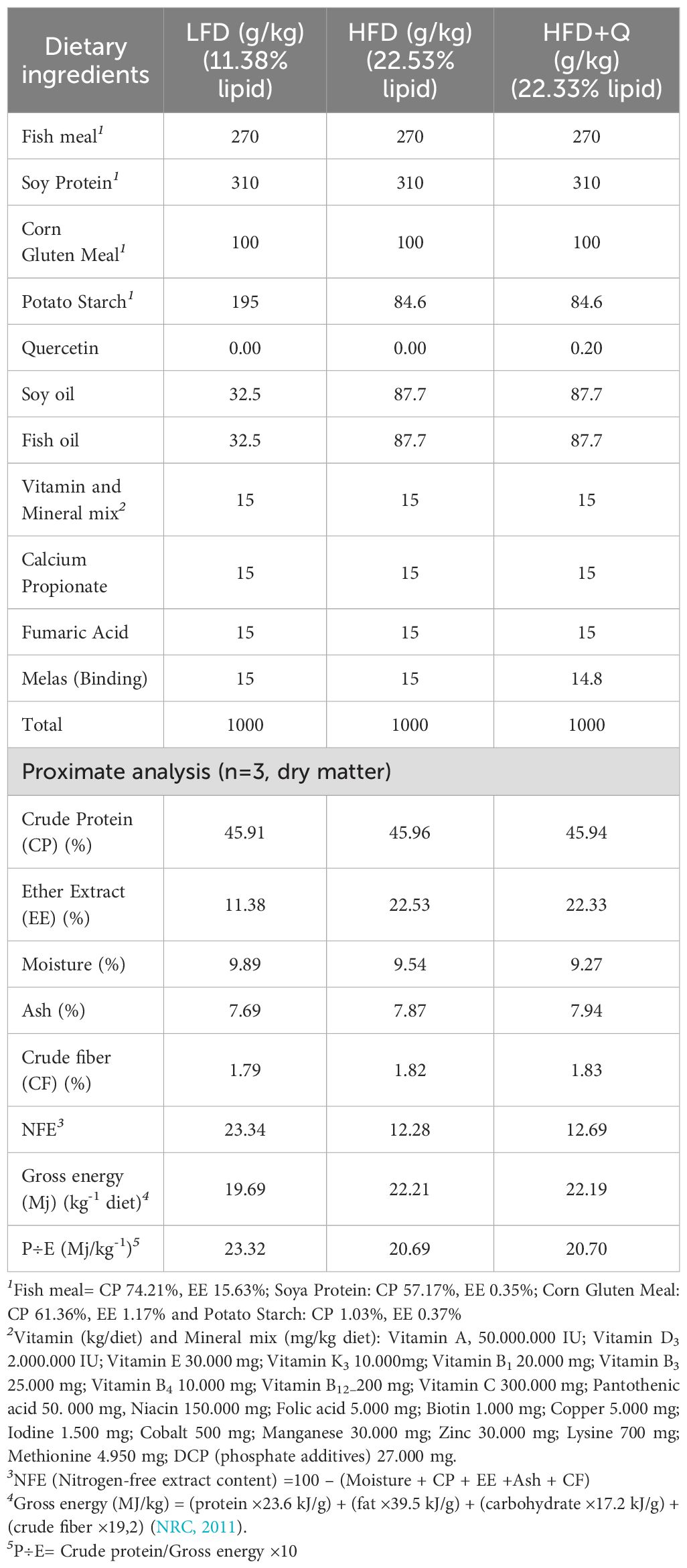
Table 1. Formulation and proximate compositions of experimental diets (g kg-1).
The dietary protein sources included fish meal, soy protein, and corn gluten meal, while lipids were provided by fish oil and soybean oil, and carbohydrates by potato starch. Diets were prepared at the Recep Tayyip Erdoğan University Iyidere R&D Unit Feed Laboratory following previously reported protocols (Köse et al., 2024; Köse et al., 2021; Kose and Karabulut, 2022), with minor modifications to ensure quercetin stability and uniformity. These modifications involved first preparing quercetin as a premix using an oil-based carrier to achieve stable and uniform incorporation within the diet. Specifically, an equal mixture (1:1) of fish oil and soybean oil, as utilized in the feed formulation, was employed as the carrier oil, into which 0.20 g/kg quercetin powder was gradually dispersed to form a homogeneous liquid premix. During feed production, this premix was gradually and evenly incorporated into the main feed mixture and mixed at high speed to guarantee uniform distribution of quercetin throughout the diet.
2.3 Verification of quercetin incorporation via HPLC
To confirm the actual concentration and assess the homogeneity of quercetin distribution in the HFD+Q diet, a high-performance liquid chromatography (HPLC-UV) method was applied. The analysis was carried out using a reverse-phase HPLC system (Agilent 1260 Infinity, Agilent Technologies, USA) equipped with a C18 column (250 mm × 4.6 mm, 5 μm). The column temperature was maintained at 30 °C, and the flow rate was set to 1.0 mL/min. The mobile phase consisted of solvent A (0.1% formic acid in water) and solvent B (acetonitrile), and was applied with the following gradient program: 25% B (0–2 min), linear increase to 60% B (2–10 min), held at 60% B (10–15 min), return to 25% B (15–17 min), and re-equilibration at 25% B (17–20 min). The injection volume was 20 µL, and detection was performed at 370 nm. Considering the target dietary quercetin concentration of approximately 200 mg/kg, the calibration curve was extended to a range of 0.1–5.0 µg/mL. Standard solutions were prepared from a 1 mg/mL quercetin stock solution in methanol, and the linearity of the curve was confirmed (R² > 0.989). Feed samples (n = 5) were collected from different locations of the HFD+Q batch. Each sample (1.0 g) was finely ground and extracted with 25 mL of 80% methanol (acidified with 0.1% formic acid) using ultrasonic-assisted extraction for 30 minutes at room temperature. The extracts were centrifuged at 3075 × g for 10 minutes, and the supernatants were filtered through a 0.45 µm PTFE syringe filter. To ensure accurate quantification and prevent detector saturation, the final extracts were diluted 1:10 with mobile phase before injection. The results indicated that quercetin concentrations ranged from 198.24 mg/kg to 200.42 mg/kg among the samples, with a mean value of 199.36 ± 0.86 mg/kg and a coefficient of variation (CV) of 0.46%. These findings confirm the successful and homogeneous incorporation of quercetin into the feed.
2.4 Sampling procedures
When the feeding trial was completed, the fish were fasted for 24 hours. Then, 10 fish were randomly selected from each tank and anesthetized with clove oil at a dose of 2–5 mg/L. All analyses conducted in the present study were performed using samples obtained exclusively from these individuals.
2.5 Serum biochemistry analysis
Blood samples were taken from the caudal vein of the fish and collected in gel tubes with Klot Activator and coagulant, kept for 1 hour and centrifuged (1180 × g, 10 min) to separate serum. Serum was collected into 2-mL cryotubes and stored at -18°C for later analysis. Serum parameters alanine aminotransferase (ALT), aspartate aminotransferase (AST), alkaline phosphatase (ALP), gamma-glutamyl transferase (GGT), lactate dehydrogenase (LDH), low density lipoprotein (LDL), high density lipoprotein (HDL), cholesterol, and triglyceride (TG)) were determined in the Biochemistry Laboratory of Recep Tayyip Erdogan University Training and Research Hospital using the same method and device (Beckman Coulter AU5800, Beckman Coulter Inc. Brea, California, USA) as described in our previous study (Köse et al., 2024).
2.6 Non-esterified free fatty acid and liver histology
Non-esterified free fatty acid (NEFA) of liver was determined according to commercial kit instructions (Non-esterified Free Fatty Acids (NEFA/FFA) Colorimetric Assay Kit, Cat no: EEA017, İnvitrogen Thermo Fisher Scientific, USA). Briefly, 1 g of liver tissue was washed with homogenization medium at +4°C, blood cells were removed, dried with filter paper and homogenized with extraction solution at a 12:1 ratio. Then oscillate at +4°C for 2 hours to extract the NEFA, centrifuged (10000×g 10 min at +4°C) and the supernatant was removed. The assay procedure was performed, and the samples were read with a microtiter plate reader (Varioskan LUX, Thermo Fisher Scientific, USA) at 715nm wavelength.
For histological investigations, liver tissue was fixed with 10% Neutral Buffered Formalin (NBF). Tissue tracking was performed with LEICA-TP 1020, tissue blocking with LEICA HistoCore Arcadia H, tissue cutting with LEICA RM2125RT (Leica Microsystems Limited, Switzerlan), and Hematoxylin and Eosin staining with MLT FS-16 Combo Automated Slide Stainer (MLT Limited Liability Company, Moscow, Dubna, Russia). Briefly, the tissues were treated with alcohol and xylene, paraffinized at 65°C, embedded in blocks, sliced with a 5 μm, stained with hematoxylin & Eosin, and coated with entellene. Electronic imaging was obtained with a high-resolution video camera (TaupCam YW3609EH, CMOS 4k UHD) integrated into a light microscope (Leica DM 500) and TaupTek ToupView software version 4.11 (ToupTek, Hangzhou, Zhejiang, P.R Cihna). The liver was morphologically evaluated for hepatocyte status, vacuolization, hepatocyte nucleus diameter, hepatocyte cell diameter, necrotic areas, hepatocellular degeneration and apoptotic hepatocyte nuclei (Aydın and Gümüş, 2020; Caballero et al., 2004; Köse et al., 2024; Martínez-Llorens et al., 2012). Histopathological evaluation of liver tissues was performed using a semi-quantitative relative scoring system, conceptually like the Non-Alcoholic Fatty Liver Disease Activity Score (NAS) commonly applied in mammalian models. Although fish liver histology differs in some aspects from mammals, steatosis-related pathological changes such as vacuolization and hepatocyte degeneration are shared features, allowing conceptual adaptation of the NAS system for fish models. The assessment was carried out by two independent observers who were blinded to the group assignments and not directly involved in the study. Each liver sample was evaluated for four key histological parameters: glycogen-type hepatocellular vacuolization, lipid-type vacuolization, hepatocellular degeneration, and alterations in sinusoidal structures. These features were scored using the following scale: (−) none, (+) very little, (++) little, (+++) moderate, (++++) much, and (+++++) very much. This scoring method has been previously applied in Oncorhynchus mykiss by Boran et al. (2010); Köse et al. (2024), and in chronic viral hepatitis by (Tuncel and Alpaslan Duman, 2021), supporting its validity for structured and comparative liver histopathology analysis in fish.
2.7 Proximate composition analysis
Proximate analysis of each feed ingredient, experimental feeds, and fish liver were performed in triplicate according to the procedures of the Association of Official Analytical Chemists (AOAC, 2005). Crude Protein was determined by the Kjeldahl method (method 978.04). N×5.58, N×5.50, N×5.62, and N×5.60 were applied as conversion factors for fish and fish derivatives, soybean and soybean derivatives, corn and corn derivatives, and potato starch, respectively (Mariotti et al., 2008). Ether extract was determined by Soxhlet method (method 930.09), and Crude fiber was determined by Weende method (method 978.10). Dry matter was determined by drying the samples at 105°C to constant weight. Ash content was determined by burning the samples in porcelain crucibles at 550°C for 5 hours (method 930.05).
2.8 Gene expression analysis by qRT-PCR
For gene expression tests, fish liver samples were placed in cryotubes containing Hibrigen RNA Stabilization Solution (Cat. No: MG-STBL-01, Hibrigen Biyoteknoloji R&D San. and Tic. A.Ş. Gebze, Kocaeli, Türkiye) and stored in a refrigerator at +4°C. Cryotubes were kept in the refrigerator for 8 hours by turning upside and down every hour and then frozen at -20°C. Samples were analyzed in the laboratory of Sugenomics Biotechnology (OSTIM Technical University, Ankara, Türkiye).
Total RNA was extracted from the liver tissue using a commercial kit (Genomics RNA Isolation Reagent, Cat: NA08A100, Su Genomics Biotechnology OSTIM Technical University, Ankara, Türkiye). Isolated RNAs were precipitated with lithium chloride (LiCl). Vazyme DNAse I RNAse-free (Cat: EN401-01, Red Maple Hi-tech Industry Park, Nanjing, PRC) was used to remove DNA from LiCl precipitated RNA samples. RNA quality and quantity were assessed using agarose gel electrophoresis (2%, M:100bp). RNA was reverse transcribed into cDNA using the SuScript cDNA Synthesis Kit (Cat: RT01A025, Sugenomics Biotechnology OSTIM Technical University, Ankara, Türkiye).
The used primer sequences of the gene region to determine gene expression levels are presented in Table 2. The relative abundance of mRNA was normalized by β-actin expression. qRT-PCR was performed using the CFX96 Real-Time PCR Detection System (Bio-Rad Laboratories, Inc, Hercules CA, USA). The reaction mix contained 10 µl 2×SuYBRGreen qPCR Mastermix, 10 µM forward and reverse primer, 2 µl cDNA, 6 µl nuclease-free water. Thermal cycling was initiated with a 5-minute incubation at 95°C using hot-start activation. 45 cycles were performed, each consisting of 15 s heating at 95°C and 30 s at each specific annealing temperature (Table 2). Ct values were obtained using Bio-Rad CFX Maestro software (Bio-Rad Laboratories, Inc, CA, USA). The standard 2-ΔΔCt method was used to measure gene expression levels (Schmittgen and Livak, 2008).
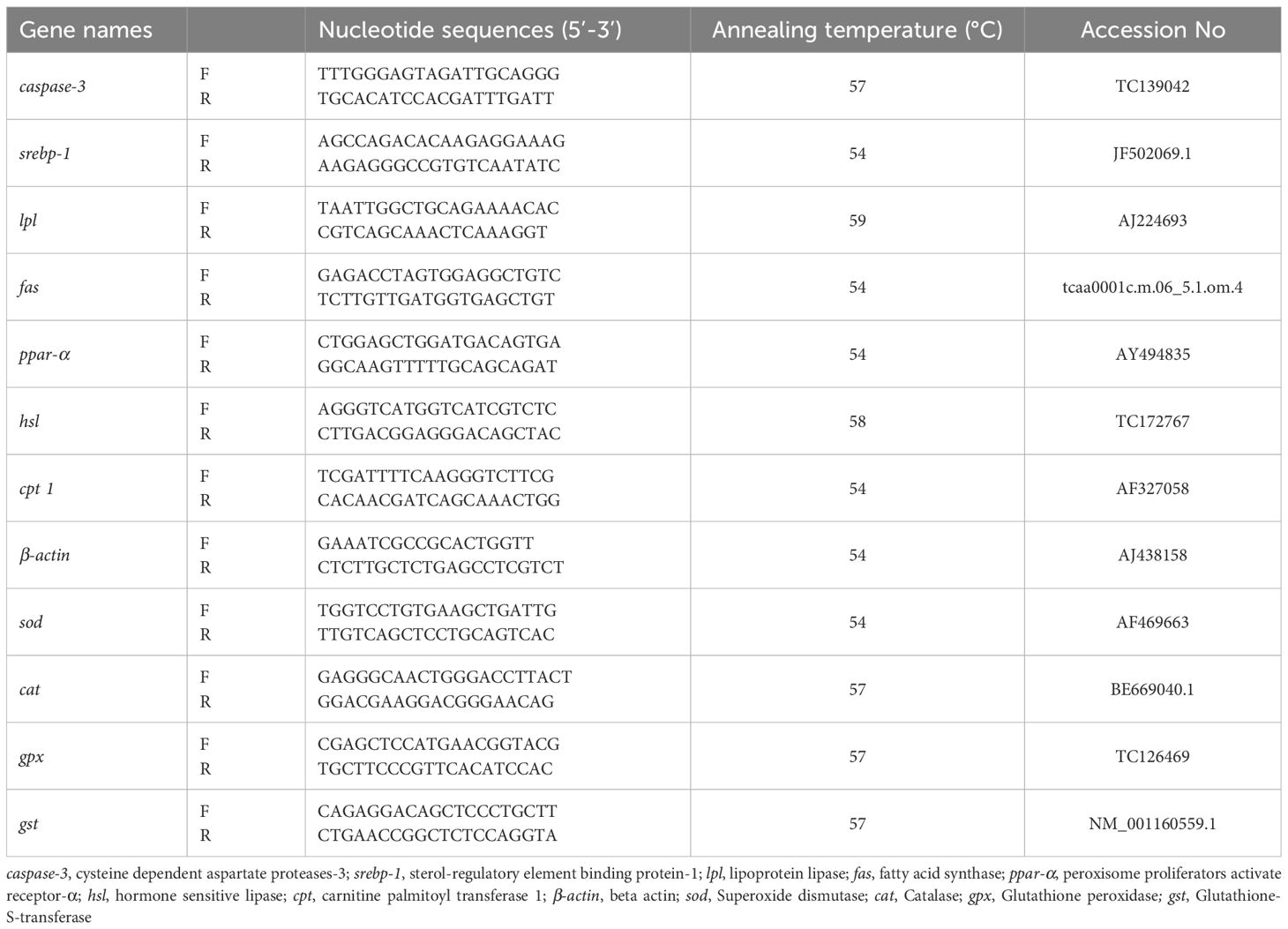
Table 2. Nucleotide sequences of the PCR primers used to evaluate mRNA abundance by qRT-PCR.
2.9 Growth performance and somatic indices calculations
Feed Intake was calculated per unit of metabolic weight of the fish (). Where () is the average daily feed intake per fish, while () is the geometric mean of the fish body weight and was calculated as follows. Where is İnitial body weigh is final body weight. İt was represented Feed Conversion Ratio with FCR, weight gain rate with WGR, condition factor with CF, viscerosomatic index with VSI, hepatosomatic index with HSI, intraperitoneal fat ratio with IPF, body weight with , body length with , viscera weight with , whole body weight with , liver weight with and intraperitoneal fat weight with .
2.10 Statistical analysis
Data are expressed as mean ± SD. Normality of the data was assessed using the Shapiro–Wilk test, and homogeneity of variances was tested using the Brown–Forsythe test. One-way analysis of variance (ANOVA) followed by the Holm–Sidak post hoc test was performed to identify significant differences among groups. Statistical significance was set at p< 0.05. All statistical analyses were conducted using SigmaPlot 14.0 (Systat Software Inc., USA).
3 Results
3.1 Growth, feed utilization and organosomatic indices
The mean values of feed utilization and organosomatic index data calculated after the 8-week feeding trial are presented in Table 3. Results showed that the high fat diet groups (HFD and HFD+Q) had significantly higher body weight (FBW), weight gain ratio (WGR), and feed intake (FI) compared to the low-fat diet (LFD) group (p<0.05). For the same parameters, the HFD+Q values were higher than those of the HFD group, but not statistically significant. FCR value was significantly higher in the LFD group compared to HFD and HFD+Q groups (p<0.05). Quercetin supplementation in the HFD+Q group significantly prevented the increase in HSI and IPF values observed in the HFD group (p < 0.05). There was no significant difference between the LFD and HFD+Q groups, but the LFD group was significantly different from the HFD group (p<0.05).
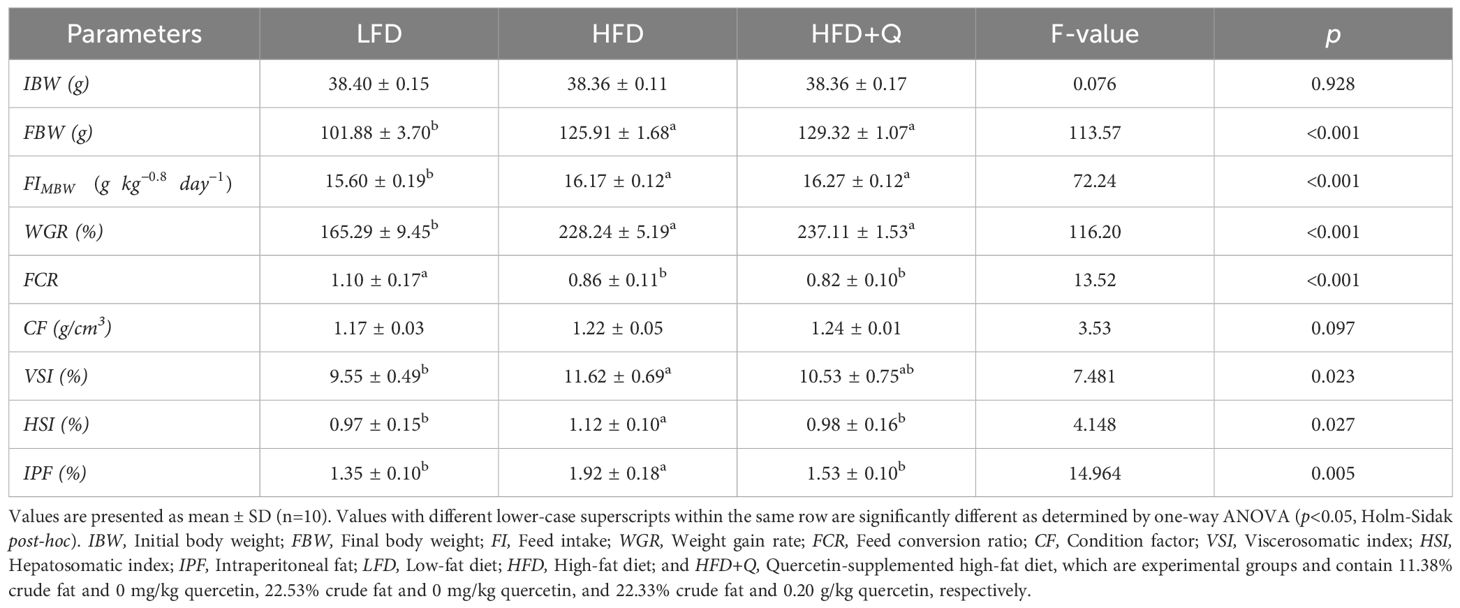
Table 3. Feed intake, growth, feed conversion ratio, and organosomatic indices of Oncorhynchus mykiss fed low fat, high fat and quercetin supplemented high fat diets for eight weeks.
3.2 Serum lipid profile, proximate composition of fish muscle and liver, and non-esterified fatty acids content
To investigate the effects of the LFD, HFD, and HFD+Q diets on the lipid profile of Oncorhynchus mykiss, LDL, HDL, cholesterol, and TG levels in serum were measured (Table 4). High dietary fat in the HFD and HFD+Q groups resulted in significantly higher serum lipid profile values compared to the LFD group (p<0.05). In the HFD+Q group, LDL, cholesterol, and TG levels were significantly lower than those in the HFD group due to the protective effect of quercetin (p < 0.05). Additionally, HDL levels were significantly higher in the HFD+Q group compared to the HFD group (p < 0.05), indicating that quercetin exerted a beneficial regulatory effect on HDL metabolism.
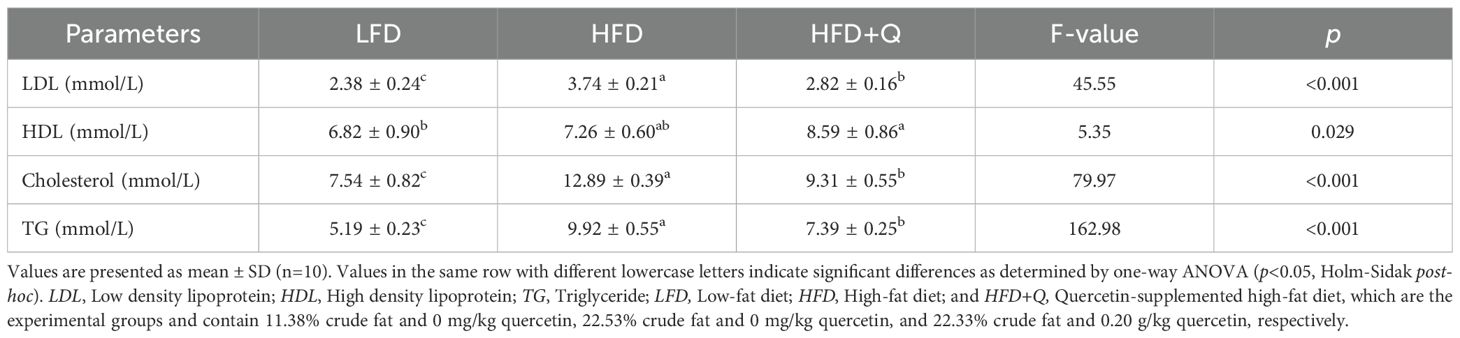
Table 4. Serum lipid profile of Oncorhynchus mykiss fed low-fat, high-fat, and quercetin-supplemented high-fat diets for eight weeks.
Non-esterified free fatty acids (NEFA) in liver tissue and the proximate levels of crude fat (ether extract) in fish muscle and liver tissues were also determined (Figure 1A–C). High dietary fat was manifested by increased levels of NEFA in the liver and an upward trend in crude fat levels of both liver and fish muscle. The HFD group exhibited significantly higher NEFA, and liver crude fat ratios compared to the HFD+Q and LFD groups, while the higher muscle crude fat ratio was significant only when compared to LFD (p<0.05). Quercetin supplementation in the HFD+Q group prevented excessive accumulation, resulting in lower levels of NEFA, liver crude fat, and muscle crude fat compared to the HFD group.
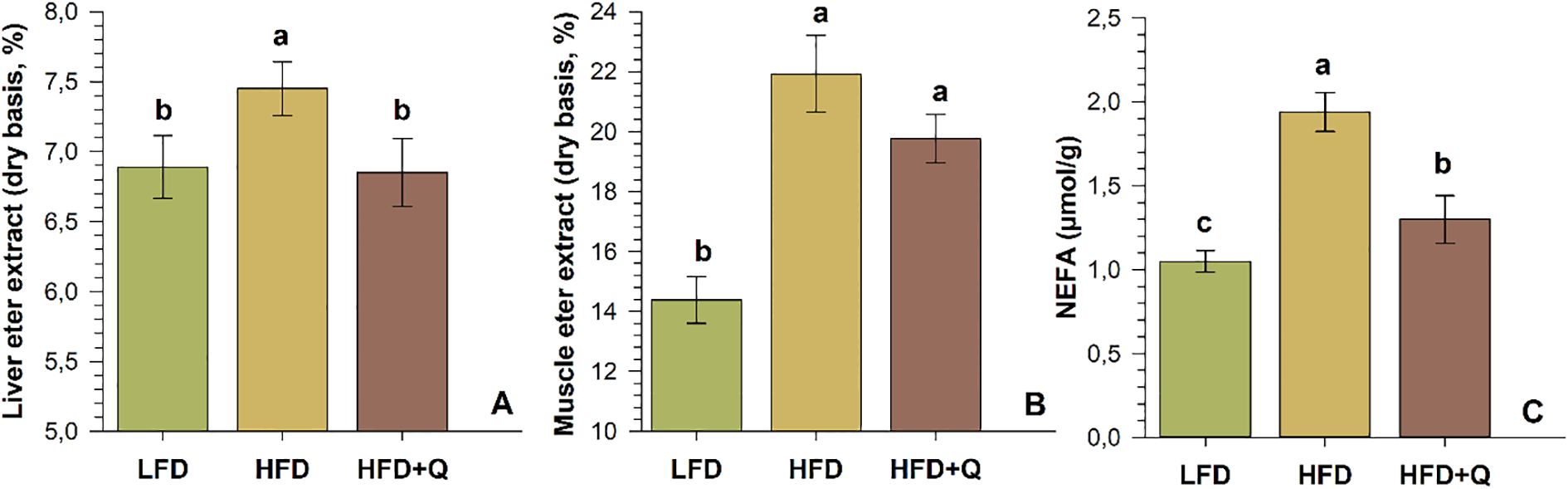
Figure 1. Crude fat and NEFA levels in Oncorhynchus mykiss fed low-fat (LFD), high-fat (HFD) and quercetin-supplemented high-fat (HFD+Q) diets for eight weeks. (A) Crude liver fat (ether extract, dry basis), (B) Fish-muscle crude fat (ether extract, dry basis), (C) Non-esterified free fatty acids (NEFA, µmol/g) in liver tissue. Values are presented as mean ± SD (n=10). Different letters above the columns indicate significant differences as determined by one-way ANOVA (p<0.05, Holm-Sidak post-hoc).
3.3 Liver histology, serum liver enzymes, and caspase-3 gene expression level
To investigate the effects of low-fat (LFD), high-fat (HFD), and quercetin-supplemented high-fat diets (HFD+Q), liver sections of Oncorhynchus mykiss were stained with H&E (Figure 2). The liver was scored for hepatocellular vacuolization (glycogen type and lipid type), hepatocytes and hepatocyte nuclei, hepatocellular degeneration zone, and sinusoidal spaces (Table 5). Levels of ALT, AST, ALP, GGT, and LDH were analyzed in serum; the apoptosis-related caspase-3 gene and apoptotic expression were determined in liver tissue (Figure 3).
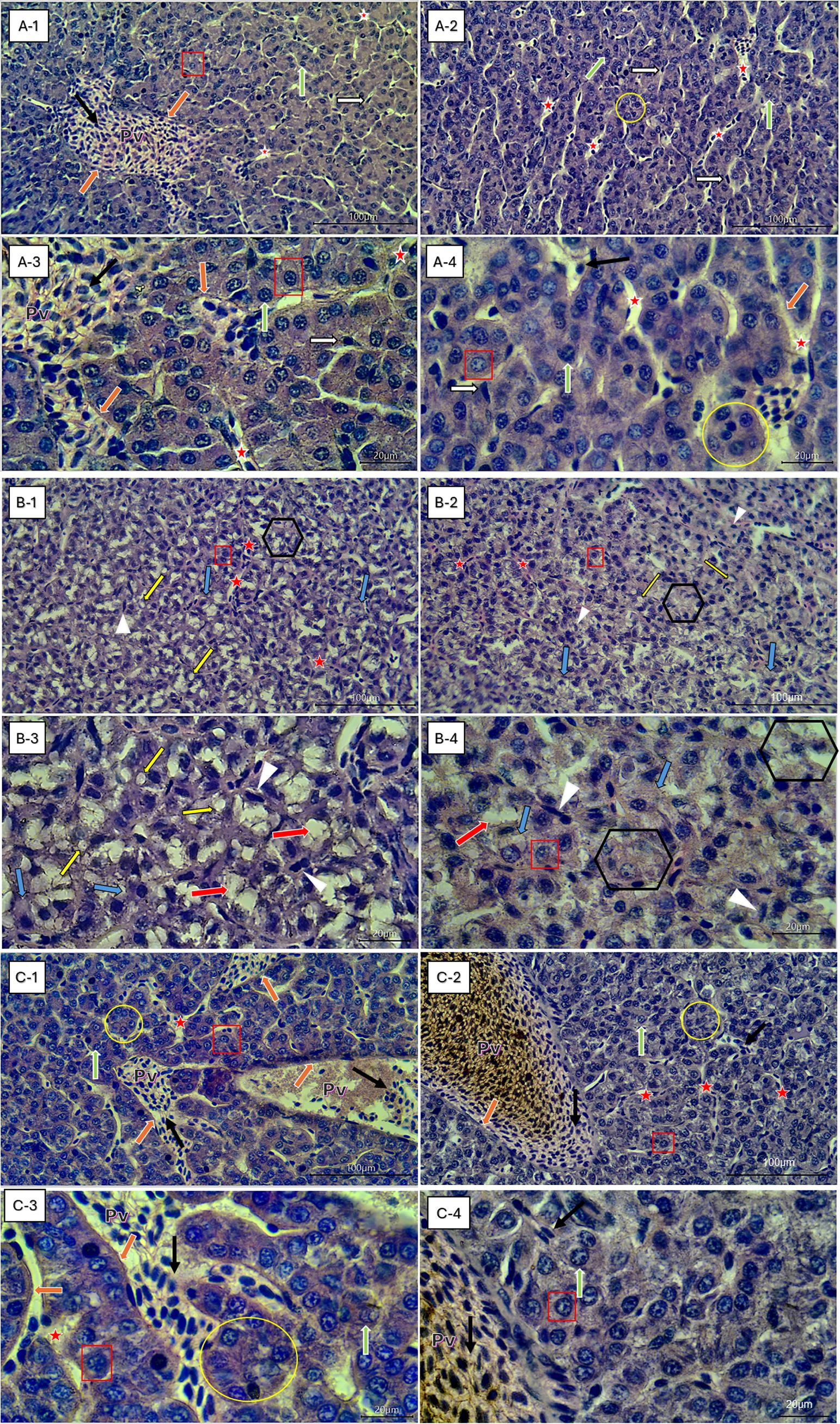
Figure 2. Representative liver histological sections (H&E staining) of Oncorhynchus mykiss fed different diets: low-fat diet (LFD), high-fat diet (HFD), and quercetin-supplemented high-fat diet (HFD+Q). (A1–A4) LFD (11.38% fat); (B1–B4) HFD (22.53% fat); (C1–C4) HFD+Q (22.33% fat + 0.20 g/kg quercetin). Images at 40× (A1, A2, B1, B2, C1, C2) and 100× (A3, A4, B3, B4, C3, C4) magnification. Pv – Portal vein; Red Asterisk – Blood vessel sinusoids; Black-Framed White Arrow – Kupffer cells; Red Square – Healthy hepatocytes with visible nuclei; White-Framed Green Arrow – Healthy hepatocyte nuclei; White-Framed Orange Arrow – Epithelial cells surrounding the portal vein; Black Arrow – Erythrocytes; Yellow Circle – Regular polygonal hepatocyte arrangements, Black-Framed Yellow Arrows – Lipid droplets; Black-Framed Blue Arrows – Swollen hepatocytes; White Arrowheads – Degenerative hepatocyte nuclei; White-Framed Red Arrows – Glycogen-type hepatocellular vacuolization; Black Hexagons – Degenerative hepatocyte zones.
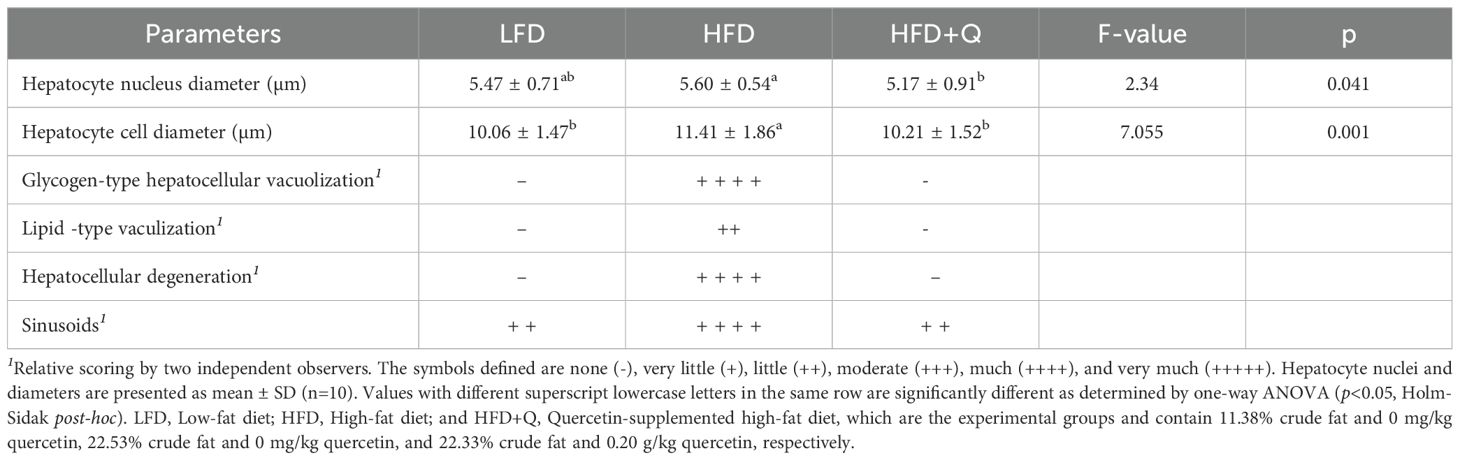
Table 5. Post-trial liver histological changes in the liver of Oncorhynchus mykiss fed low fat, high fat, and quercetin supplemented high fat diets for eight weeks.
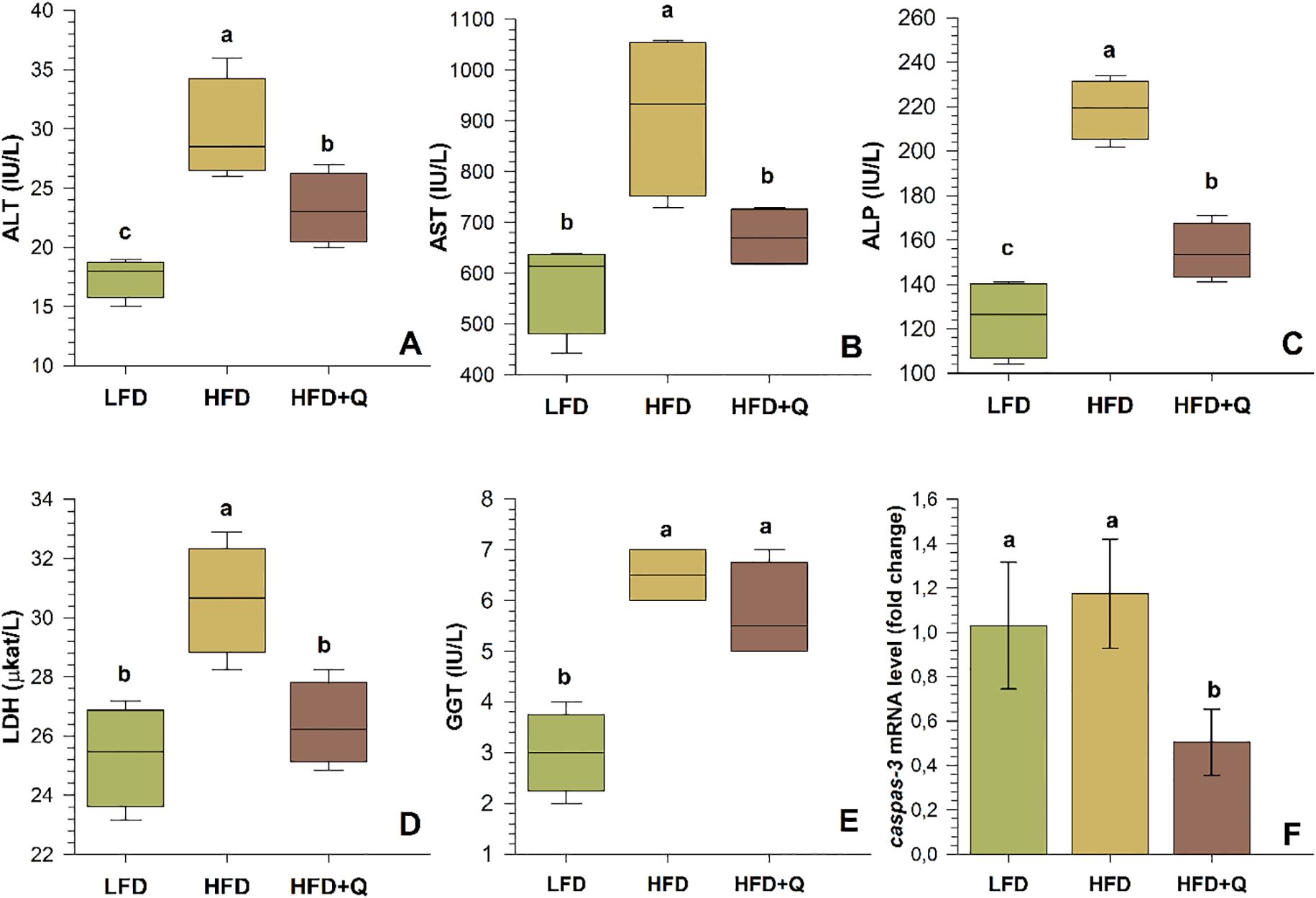
Figure 3. Serum liver enzyme levels and mRNA levels of the apoptosis-related caspase-3 gene in liver tissue of Oncorhynchus mykiss fed a low-fat diet (LFD), high-fat diet (HFD) and quercetin-supplemented high-fat diet (HFD+Q) for eight weeks. (A) Alanine aminotransferase (ALT), (B) Aspartate aminotransferase (AST), (C) Alkaline phosphatase (ALP), (D) Lactate dehydrogenase (LDH), (E) Gamma glutamyl transferase (GGT), (F) Cysteine-aspartate-specific protease-3 (caspase-3). Values are expressed as mean ± SD (n=10). Different letters above the columns indicate significant differences as determined by one-way ANOVA (p<0.05, Holm-Sidak post-hoc).
Histological examination revealed that the LFD group exhibited a normal liver architecture. Hepatocyte cords were well-organized, nuclear integrity was preserved, and no signs of steatosis or degeneration were observed (Figures 2A1-A4). In contrast, the HFD group displayed marked histopathological alterations in liver tissue. These included both glycogen-type and lipid-type vacuolization, hepatocyte swelling, nuclear degeneration, displaced nuclei, expanded sinusoidal spaces, and the presence of apoptotic cells. These structural abnormalities were accompanied by a pronounced disruption of the polygonal hepatocellular organization, indicating severe hepatic damage (Figures 2B1-B4). In the HFD+Q group, the histological structure appeared preserved. No vacuolization was observed, the sinusoidal architecture remained well-organized, and hepatocyte morphology and nuclear integrity were clearly maintained (Figures 2C1-C4). Furthermore, while significant increases in hepatocyte and nuclear diameters were observed in the HFD group, quercetin supplementation in the HFD+Q group prevented these increases, resulting in significantly lower values for both parameters compared to the HFD group, suggesting a protective effect against cellular hypertrophy (Table 5). At the molecular level, caspase-3 gene expression in liver tissue was significantly upregulated in the HFD group. However, this upregulation was markedly suppressed in the HFD+Q group, which showed significantly lower expression levels of the apoptosis-related caspase-3 gene compared to both HFD and LFD groups (p<0.05) (Figure 3F).
Serum liver enzymes were found to be higher in HFD and HFD+Q diet groups compared to LFD diet groups. The histological changes observed in the HFD group were consistent with the significant increase in serum ALT, AST, ALP, LDH, and GGT values (p<0.05). Quercetin supplementation in the HFD+Q group prevented the elevation of these enzymes, resulting in lower ALT, AST, ALP, and LDH levels compared to the HFD group (p<0.05).
3.4 Expressions of genes related to antioxidant function in the liver
The mRNA levels of antioxidant genes in the liver are shown in Figure 4. In the HFD group, sod mRNA and gst mRNA levels were high, and the sod mRNA level was significantly different from the LFD and HFD+Q groups (p<0.05). On the contrary, cat mRNA and gpx mRNA levels showed a decreasing trend in the HFD group, and this trend was significantly different from the LFD group (p<0.05). HFD+Q group was lower than LFD for sod mRNA and cat mRNA. There was no significant difference, but gpx mRNA and gst mRNA levels were significantly lower than LFD (p>0.05).
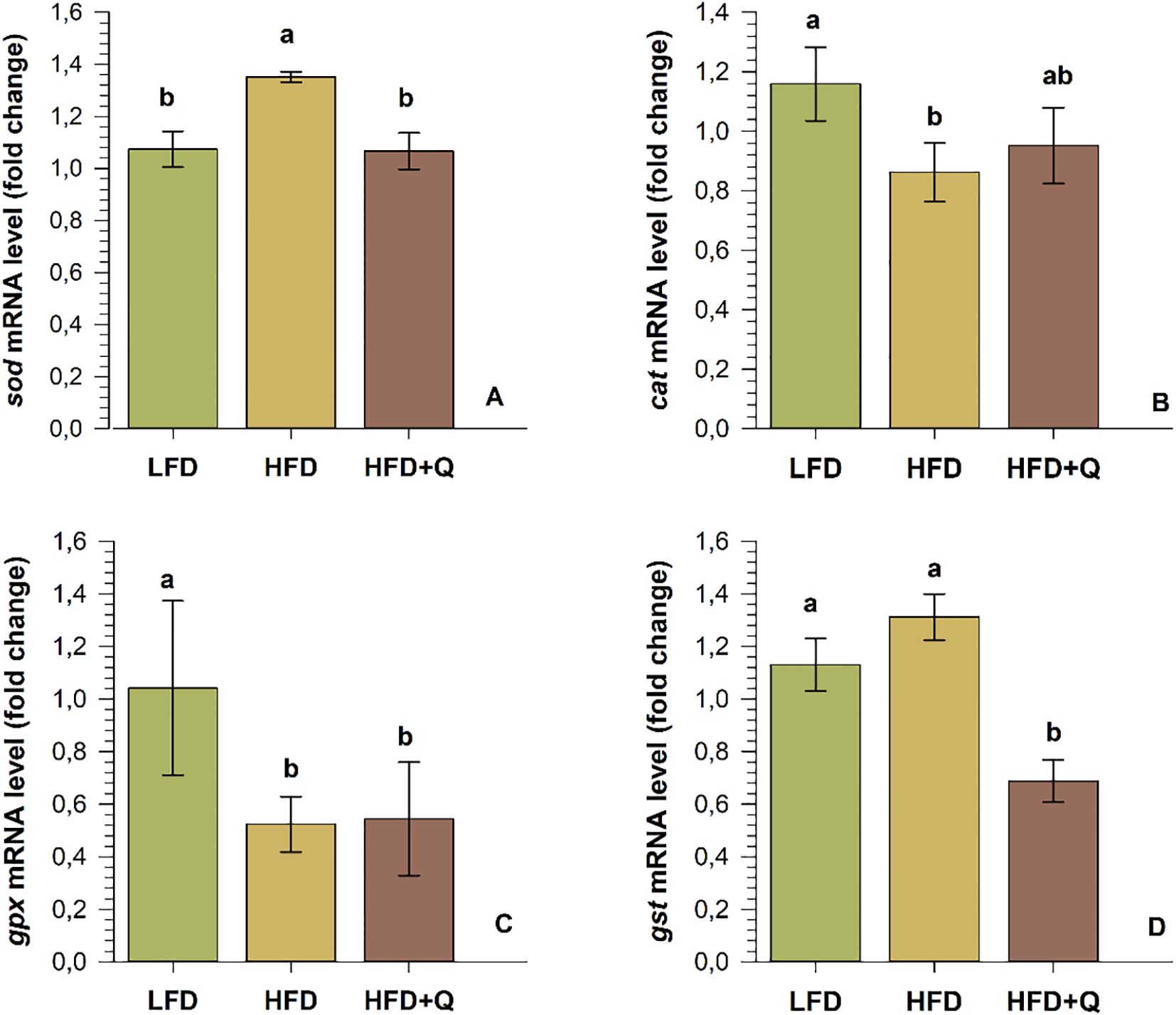
Figure 4. The mRNA levels of antioxidant function-related genes in the liver of Oncorhynchus mykiss fed low-fat (LFD), high-fat (HFD) and quercetin-supplemented high-fat diets (HFD+Q) for eight weeks. (A) superoxide dismutase (sod), (B) catalase (cat), (C) glutathione peroxidase (gpx) and (D) Glutathione-S-transferase (gst). Values are expressed as mean ± SD (n=10). Different letters above the columns indicate significant differences as determined by one-way ANOVA (p<0.05, Holm-Sidak post-hoc).
3.5 Expression of liver genes linked to lipid metabolism and regulatory factors
The relative mRNA levels of selected genes related to lipid metabolism in the liver are presented in Figure 5. The ppar-α mRNA level did not show a significant change in the LFD and HFD+Q diets, while the HFD diet group was significantly down-regulated (p<0.05). The srebp-1 mRNA levels showed opposite changes in the HFD and HFD+Q diet groups compared to the LFD group. While it was down-regulated in the HFD diet group, it was up-regulated in the HFD+Q diet group, and all groups were significantly different from each other (p<0.05). The lpl mRNA and fas mRNA levels were significantly up-regulated in the HFD diet group (p<0.05), while there was no significant change in the HFD+Q and LFD diet groups (p>0.05). The mRNA levels of hsl and cpt-1a were significantly down-regulated in the HFD diet group (p<0.05), while there was no significant change between HFD+Q and LFD (p>0.05).
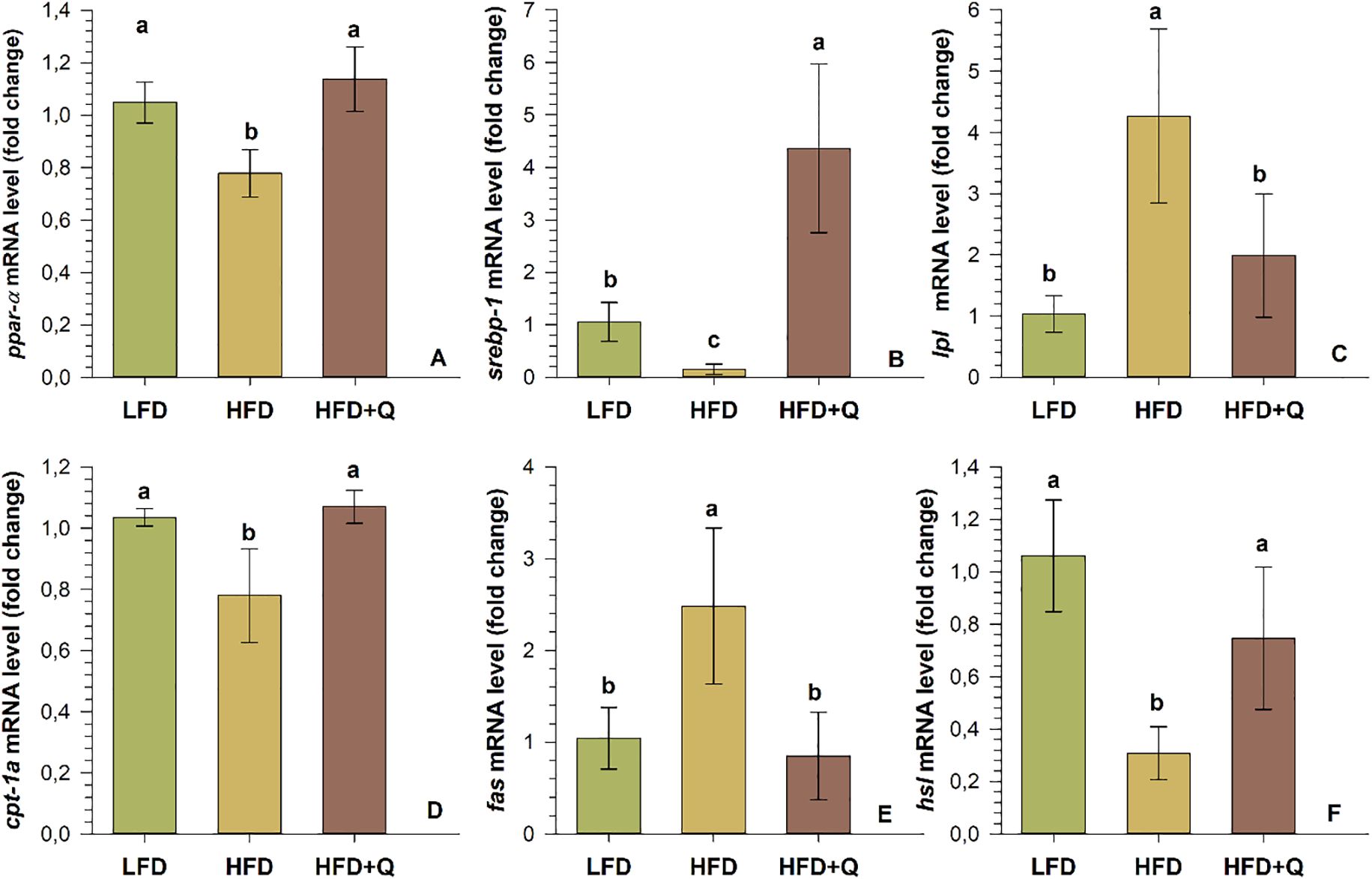
Figure 5. Expression of lipogenesis (srebp-1, lpl and fas) and lipolysis (ppar-α, cpt-1a and hsl) pathway genes in liver tissue of Oncorhynchus mykiss fed low-fat (LFD), high-fat (HFD) and quercetin-supplemented high-fat diets for 8 weeks. (A) peroxisome proliferator activated receptors-α (ppaR- α), (B) sterol regulatory element binding protein (srebp-1), (C) lipoprotein lipase (lpl), (D) carnitine palmitoyltransferase-1a (cpt-1a), (E) fatty acid synthase (fas), (F) Hormone sensitive lipase (hsl). Values are expressed as mean ± SD (n=10). Different letters in the columns indicate significant differences as determined by one-way ANOVA (p<0.05, Holm-Sidak post-hoc).
4 Discussion
The HFD induced fatty liver is common worldwide, and the primary pathological symptoms are hepatic lipid accumulation and steatosis. Nowadays, excessive addition of fat to fish feed causes frequently fatty liver in fish, which has a negative impact on their growth and development (S. Xie et al., 2021). Therefore, alternative medications or bioactive chemicals that can improve fish livers are extremely important. This study determined that HFD induced degenerative consequences in Oncorhynchus mykiss liver, and quercetin administration prevented the development of these pathological changes. The effect of quercetin on genes associated with lipid metabolism and liver antioxidant status in liver cells was investigated in response to a high fat diet. The effects of a quercetin-supplemented and a low-fat diet on fish development were compared with the high fat diet.
4.1 Growth, feed utilization, and organosomatic indices
Dietary lipid levels have sensitive effects on protein conservation and growth. It was reported that the high dietary lipids reduce growth in some fish without a protein conservation effect (Akpınar et al., 2012; Du et al., 2005; Sevgili et al., 2014) or increasing dietary lipid levels may not have a significant effect on growth (McGoogan and Gatlin, 1999; Wang et al., 2015). On the other hand, the dietary lipid content has improved the growth performance of semi-adult triploid rainbow trout (Meng et al., 2019). Similarly, there were positive effects on Salmo gairdneri (Beamish and Medland, 1986), Salmo salar (Hillestad and Johnsen, 1994), and Oncorhynchus mykiss (Meng et al., 2019). Quercetin-supplemented feeds promoted the growth of Paralichthys olivaceus and Channa argus and reduce the FCR (Kong et al., 2022; Shin et al., 2010a). Fish avoid low-energy or unbalanced diets, and their feed intake decreases (Meng et al., 2019). Carnivorous fish convert protein to energy more efficiently than lipids and carbohydrates, and can adjust their feed intake to meet their digestible energy requirements (Du et al., 2005). Even though there are variation of dietary lipid levels’ effect on growth, the current study has shown that HFD and HFD+Q diets support growth effect that provides protein savings with better FBW, WGR, FCR, FI, and CF values compared to the LFD diet. Moreover, the protein levels of all diets were equal, the growth of fish in the HFD and HFD+Q diets was better than the LFD diet. The low FBW, WGR, CF, and high FCR values in the LFD diet group can be explained by the fish’s low FI due to not preferring the feed or by using protein as an energy source for growth because of insufficient dietary fat.
It was reported that VSI, HSI and IPF levels increased on the HFD diet, but this increase was limited with quercetin treatment on Megalobrama amblycephala (Abasubong et al., 2023), Ictalurus punctatus (Desouky et al., 2020), and Oxyeleotris marmorata (Yong et al., 2015) fish. The present results support the previous studies. High lipids in the diet reduce growth performance due to lipid accumulation in internal organs (Desouky et al., 2020; Han et al., 2014); however, in this study, the growth did not negatively affect although HFD and HFD+Q diets increased the organosomatic indices. Studies on Oncorhynchus mykiss with varying dietary lipid ratios (Liu et al., 2021; Meng et al., 2019) are also consistent with the current findings. The effects of dietary lipid levels on fish growth are complex, and the findings showed variation; thus, it may be related to fish species, dietary components, dietary protein and lipid levels, feeding regimes, and environmental conditions.
4.2 Serum lipid profile, proximate composition of fish muscle and liver, and non-esterified fatty acids content
Endogenous and exogenous variables change the proximate composition of farmed fish. Protein and ash content are mostly determined by the size of the fish (endogenous factors), while fat content is influenced by exogenous factors such as diet (Shearer, 1994). High dietary lipid can cause excessive fat accumulation in the visceral cavity and tissues (López et al., 2006). NEFA is the primary reason of liver lipotoxicity (Brent, 2010), and quercetin may weaken its effects by reducing the level of lipotoxicity Lateolabrax maculatus (Xia et al., 2024), Rachycentron canadum (Wang et al., 2005), Epinephelus coioides (Luo et al., 2005), and Oncorhynchus mykiss (Jobling et al., 1998). In this study, lipid accumulation in fish muscle and liver was increased in the HFD and HFD+Q diets compared to the LFD diet. It also increased in liver NEFA. These increases were more limited in the HFD+Q supplemented with quercetin, and they are consistent with the previous studies.
The liver synthesizes cholesterol, which is transferred to other tissues via LDL. The HDL transports cholesterol from peripheral tissues to the liver, where it is processed and excreted with the bile regulating blood cholesterol levels (Asgary et al., 2000). Cholesterol levels can indicate disorders of lipid and lipoprotein metabolism, and liver diseases (Mensinger et al., 2005). In mammals, the increase in plasma/liver LDL, TG, cholesterol levels, and the decrease in HDL levels are significant risk factors for steatosis (Desouky et al., 2020). Quercetin inhibits cholesterol biosynthesis by blocking fatty acid synthesis activity (Yamamoto and Oue, 2006). Quercetin lower TG levels by inhibiting the enzymes involved in TG biosynthesis and fat adipogenesis (Jia et al., 2019; Kuipers et al., 2018). Moreover, it can also lower cholesterol levels by reducing cholesterol absorption in the intestines and affecting lipoprotein biosynthesis (Jia et al., 2019). In this study, LDL, HDL, TG, and cholesterol levels increased with the HFD diet. Quercetin prevented the elevation of LDL, TG, and cholesterol levels, while positively regulating HDL levels. In a study on Pseudosciaena crocea with high dietary lipids (Wang et al., 2015), the reported TG and cholesterol levels were similar, but the changes in LDL and HDL were different. Another study in Oreochromis niloticus found that the addition of quercetin reduced serum LDL and TG levels (Abdo et al., 2024). In contrast, another study in Oreochromis niloticus found that HDL and cholesterol levels increased while LDL and TG levels decreased (Zhai and Liu, 2013). Total cholesterol levels of Paralichthys olivaceus fed supplemental diets containing 0.25% or 0.50% quercetin were reported to be significantly lower than those fed diets without quercetin (Shin et al., 2010b). Studies conducted on mice (Nakamura et al., 2000; Yugarani et al., 1992) have reported different results from the present study. Differences in animal species, dietary quercetin levels, dietary contents and study duration may explain the differences between studies. As a result, quercetin may reduce TG and LDL levels, increase HDL levels, control liver NEFA content, cholesterol levels and liver crude fat content. Quercetin may also be beneficial in avoiding pathological changes in fatty liver (steatosis).
4.3 Liver histology, serum liver enzymes, and caspase-3 gene expression
Long-term consumption of a high-fat diet (HFD) disrupts the balance between catabolism and anabolism, leading to the accumulation of lipids and fatty acids. This imbalance can result in impaired lipid homeostasis and contribute to the development of pathological conditions characterized by excessive lipid accumulation in organs such as the liver (Du et al., 2013; Li et al., 2014; Yan et al., 2015; Zhang et al., 2021; Zhou et al., 2022). Hepatocyte surface area, hepatocyte nucleus diameter, hepatocyte count, cytoplasmic lipid, and glycogen content are commonly used parameters to assess metabolic activity in the liver (Strüssmann and Takashima, 1990). Previous studies have shown that HFD increases liver volume in Oreochromis niloticus and Lateolabrax maculatus, leading to structural disruptions in liver tissue, the formation of prominent vacuoles between lipid droplets and cells, and these adverse effects were mitigated by quercetin treatment (Xia et al., 2024). In this study, fish fed with an HFD exhibited hepatocellular glycogen and lipid-type vacuolization, hepatocyte nucleus degeneration, swollen hepatocytes, nuclear displacement, increased nucleus diameter, and expanded sinusoidal spaces. However, quercetin prevented these pathological changes, supporting findings from previous studies.
An enzymatic approach to liver disease aids in diagnosis, monitoring disease progression, and identifying subclinical manifestations. Five key serum enzymes-ALT, AST, ALP, LDH, and GGT-are commonly utilized in the assessment of liver function (Choi, 2003). However, rather than serving as direct indicators of liver function, serum levels of these enzymes primarily reflect hepatobiliary damage, ranging from inflammation to necrosis (Bayraktar and Coşar, 2019). AST and ALT are intracellular enzymes released into the bloodstream due to hepatocyte injury, whereas ALP plays a role in fundamental physiological processes and is considered an indicator of fish health status (Zhou et al., 2015). To differentiate liver-derived ALP elevation, it is more informative to evaluate it in conjunction with GGT levels (Ersoy, 2012). LDH, a cytoplasmic enzyme, serves as a marker of cellular damage, while GGT functions as an indicator of cellular antioxidant deficiency and liver disease (Morovvati and Armand, 2019). The elevation of GGT alongside other liver enzymes strongly suggests hepatic origin (Ersoy, 2012). In the present study, high-fat diet (HFD) consumption led to increased serum levels of ALT, AST, ALP, LDH, and GGT, indicating hepatocellular damage. Quercetin supplementation prevented the elevation of ALT, AST, ALP, and LDH levels in the HFD+Q group compared to the HFD group; however, it had limited impact on GGT levels. This finding may reflect GGT’s more complex regulation or delayed responsiveness to intervention. The elevation of liver enzymes indicates hepatocellular damage and impaired liver function at the cellular level and can be interpreted as a biochemical manifestation of steatosis (fatty liver) in fish. In this study, the histopathological changes observed in the liver of the HFD group—including hepatocellular glycogen and lipid-type vacuolization, hepatocyte nuclear degeneration, swollen hepatocytes, nuclear displacement, increased nuclear diameter, and expanded sinusoidal spaces—support this interpretation. These structural alterations suggest that a high-fat diet may induce metabolic stress and morphological disruption in the liver, thereby promoting the development of steatosis. A clear correlation can therefore be established between the elevated serum liver enzyme levels and the histological findings. Previous studies have reported that quercetin administration following acute cyclophosphamide exposure in rats significantly reduced ALT, AST, and ALP levels (Khalaf and Salih, 2023). Similarly, quercetin treatment has been shown to lower ALT and AST levels in Cyprinus carpio (Ghafarifarsani et al., 2022a, 2022b). In Oreochromis niloticus exposed to Abamectin (ABM), quercetin supplementation mitigated the elevation of ALT and AST levels, demonstrating a strong hepatoprotective effect (Mansour et al., 2022). Furthermore, in patients with non-alcoholic fatty liver disease (NAFLD), quercetin treatment for two weeks significantly decreased elevated ALT, AST, and GGT levels (Prysyazhnyuk and Voloshyn, 2017). Collectively, these findings align with the present study, suggesting that quercetin may exert a potential therapeutic effect on liver health.
Caspase-3 is recognized as a critical regulator of apoptosis. It is frequently activated through proteolytic cleavage within apoptotic cells, triggering apoptosis via both extrinsic and intrinsic pathways (Earnshaw et al., 1999; McArthur and Kile, 2018). The inactivation of caspase-3 has been reported to provide protection against hepatic cell death and fibrogenesis (Thapaliya et al., 2014). Studies have demonstrated a significant upregulation of caspase-3 in the liver tissue of mice fed with an HFD (Gadallah and Almasari, 2023). Conversely, quercetin has been reported to induce apoptosis due to its toxic effects and upregulate caspase-3 expression3 (Lee et al., 2015). In addition to its role in apoptosis, caspase-3 also participates in non-apoptotic cellular processes, including cell differentiation, cell cycle regulation, and cell migration (Schwerk and Schulze-Osthoff, 2003; Zhao et al., 2006). The involvement of caspase-3 in these processes is attributed to its proteolytic activity, which facilitates the controlled degradation of specific intracellular proteins, thereby regulating various cellular functions (Du et al., 2004). In this study, HFD consumption led to a significant upregulation of caspase-3 gene expression, while quercetin supplementation prevented this upregulation. Normal hepatic histoarchitecture was preserved in the LFD and HFD+Q groups, whereas the HFD group exhibited marked histological abnormalities. The unchanged caspase-3 levels and preserved histological structure in the LFD group, the significant upregulation of caspase-3 expression accompanied by disrupted histoarchitecture in the HFD group, and the significant downregulation of caspase-3 along with the preservation of histoarchitecture in the HFD+Q group collectively indicate that caspase-3 exerts apoptotic activity in the liver and that quercetin exhibits a hepatoprotective effect against this process.
4.4 Expressions of genes related to antioxidant function in the liver
Under conditions of physiological homeostasis, animal cells produce reactive oxygen species (ROS) and maintain a balance of various antioxidant defenses through several opposing mechanisms. The disruption of the balance between these two processes can lead to oxidative stress. Although it is clear that antioxidant defense mechanisms are directly related to the dietary base, there is very little information on how the level and type of dietary nutrients affect oxidative status in fish (Azaza et al., 2020). Superoxide dismutases (SODs) are enzymes primarily responsible for maintaining the oxidation-reduction balance. The SODs use the superoxide anion in a dismutation reaction to produce hydrogen peroxide (H2O2), which is converted to oxygen and water by catalase (CAT) and glutathione peroxidase (GPx) (Gottfredsen et al., 2014). A study in Danio rerio found that quercetin supplementation dramatically increased liver SOD levels (Wang et al., 2020). Similarly, higher SOD activity and upregulation of sod mRNA expression were reported in the liver of Oncorhynchus mykiss fed a high-fat diet (HFD) (26.06%) (Liu et al., 2021). The CAT is mainly found in peroxisomes and is responsible for reducing H2O2 produced by peroxisomal fatty acid metabolism, GPx reduces both hydrogen and lipid peroxides (Li et al., 2010). Low levels of cat mRNA and gpx mRNA despite high levels of sod mRNA may indicate dysregulation H2O2 detoxification in the SOD dismutation reaction. The decrease in CAT and GPx activity may increase H2O2 levels in fish liver hepatocytes, thereby increasing the peroxidation of biomacromolecules and oxidative damage (Shadegan and Banaee, 2018). Glutathione S-transferase (GST) is thought to form glutathione conjugates with peroxyradicals, although this function has not been confirmed in fish (Tocher et al., 2002). The 5.3 mg kg-1 dose of mycotoxin was reported to increase the GST value in carp liver (Matejova et al., 2017). Increased levels of SOD, CAT, GPx and GST have been reported in the literature with quercetin treatment (Ghafarifarsani et al., 2022b; Jia et al., 2019; Kim et al., 2012; Nasirin et al., 2023; Wang et al., 2020; Xu et al., 2019). In this study, enzymatic antioxidant activities (SOD, CAT, GPx, GST) were not directly assessed; however, the gene expression levels of antioxidant-related genes were determined in the liver in response to HFD and quercetin-supplemented HFD diets. The results revealed that a high-fat diet significantly upregulated sod mRNA expression in liver tissue while markedly downregulating cat mRNA and gpx mRNA levels. The upregulation of sod mRNA in response to oxidative stress induced by HFD may be associated with an increased need for the liver to detoxify superoxide radicals. However, despite this upregulation, the downregulation of cat mRNA and gpx mRNA suggests an insufficient detoxification of H2O2, potentially exacerbating oxidative stress. The suppression of cat mRNA and gpx mRNA in the HFD group indicates that a high-fat diet may exert an inhibitory effect on the antioxidant system, possibly depleting the glutathione system. The significant downregulation of gpx mRNA in the HFD+Q group suggests that the effect of quercetin supplementation on this pathway warrants further investigation. Although the HFD group exhibited an upward trend in gst mRNA levels compared to the LFD group, this increase was not statistically significant, potentially representing an adaptive response aimed at enhancing the detoxification of lipid peroxidation products due to elevated H2O2 levels. Conversely, the reduction in gst mRNA levels observed in the HFD+Q group suggests that quercetin may mitigate oxidative stress without exerting a pro-oxidant effect. In conclusion, a high-fat diet may disrupt oxidative balance by increasing oxidative stress burden and suppressing cat mRNA and gpx mRNA expression, thereby impairing the maintenance of redox homeostasis. Quercetin supplementation may exert a hepatoprotective effect either by directly reducing oxidative stress or by modulating antioxidant defense mechanisms through alternative pathways. Future studies integrating enzymatic activity assessments with gene expression analyses are essential to achieve a more comprehensive understanding of oxidative stress and antioxidant defense mechanisms.
4.5 Expression of liver genes linked to lipid metabolism and regulatory factors
Quercetin has a regulatory effect on the expression of lipid metabolism-related genes in hepatic mRNA in obese individuals (Jung et al., 2013; Wang et al., 2008). Quercetin treatment has been shown to promote fat hydrolysis by stimulating lipolysis through increased expression of hsl mRNA (Seo et al., 2015). The expression of lpl varies in a tissue-specific manner (Li et al., 2013). The expression level of lpl mRNA is influenced by dietary lipid levels (Liu et al., 2021), and while hepatic lpl has been reported to prevent diet-induced obesity and hepatic steatosis, liver-specific overexpression of lpl mRNA paradoxically leads to hepatic steatosis (Chiu et al., 2010). Furthermore, increased dietary intake and fatty acid levels have been associated with elevated lpl mRNA expression (Kleveland et al., 2006; Liang et al., 2002; Zheng et al., 2010). One of the causes of steatosis is reduced free fatty acid β-oxidation. The expression of cpt-1a and ppar-α mRNA, which play a rate-limiting role in β-oxidation, has been reported to be downregulated in Oryzias latipes and Megalobrama amblycephala (Lu et al., 2017, 2014; Matsumoto et al., 2010), whereas contrasting findings have been reported in zebrafish (Dai et al., 2015) and Ctenopharyngodon idellus (Li et al., 2016). The srebp-1 is a transcription factor that regulates the expression of genes involved in lipogenesis. Several genes responsible for fatty acid desaturation are regulated by this transcription factor (Kawakami et al., 2005; Shimano, 2001). The fas serves as a key marker of fat formation in the lipogenesis process, whereas hsl is another important marker involved in fat lipolysis (Moseti et al., 2016; Sekar and Thirumurugan, 2022). In this study, to investigate the mechanism behind the lipid-lowering effect of quercetin and its hepatoprotective properties, the relative expression of some selected genes involved in the lipogenesis (srepb-1, fas, lpl) and lipolysis (ppar-α, cpt-1a and hsl) pathways were analyzed. Interestingly, contrary to expectations, hepatic srebp-1 mRNA expression was downregulated in the HFD group. Although srebp-1 is an important transcription factor that promotes lipogenesis, this downregulation may be related to feedback inhibition mechanisms triggered by excessive hepatic lipid accumulation. In steatotic hepatocytes, the cellular overload of triglycerides and free fatty acids may have suppressed srebp-1 transcription to limit further fat synthesis. In the HFD+Q group, srebp-1 mRNA expression increased to levels comparable to or slightly higher than the LFD group. However, the expression of downstream target genes such as fas remained low, suggesting that the increase in srebp-1 did not translate into enhanced lipogenesis. This indicates that quercetin may exert a regulatory rebalancing effect on metabolic signaling pathways without promoting lipid accumulation. These findings emphasize that the effect of quercetin is not merely a simple suppression of lipid synthesis but rather a normalization of disrupted transcriptional networks under metabolic stress. The HFD diet was found to increase lpl mRNA expression. This may contribute to steatosis by promoting the uptake of free fatty acids generated through lpl-mediated hydrolysis of triacylglycerol in the liver by extrahepatic tissues. The decreased expression of cpt-1α and ppar-α mRNA suggests a reduction in β-oxidation, which may accelerate lipid accumulation and explain the abnormal histoarchitecture observed in the liver of Oncorhynchus mykiss fed an HFD diet. On the other hand, the upregulation of cpt-1α and ppar-α mRNA in the HFD+Q group suggests an adaptive response against hepatotoxicity by enhancing β-oxidation. HFD treatment induced fat formation by upregulating fas mRNA, while the downregulation of hsl mRNA prevented the breakdown of dietary fatty acids, leading to abnormal liver architecture. In conclusion, quercetin treatment may exert its hepatoprotective effect by modulating lipid metabolism through the regulation of lipogenesis and lipolysis pathways.
5 Conclusion
his study demonstrated that feeding female Oncorhynchus mykiss a high-fat diet (HFD) induced significant metabolic and pathological alterations, including hepatic steatosis, dysregulated lipid metabolism, oxidative stress, hepatocellular damage, and impaired serum biochemical parameters such as elevated ALT, AST, and TG levels. Quercetin supplementation prevented fat accumulation in the liver, preserved its normal microscopic structure, limited cell death (apoptosis) by downregulating casp3 gene expression, modulated antioxidant gene expression to improve oxidative balance, and maintained serum biochemical parameters at normal levels. Additionally, quercetin suppressed the expression of lipogenesis-related genes (fas and lpl), while upregulating genes involved in fat breakdown and energy production (ppar-α, cpt-1α, and hsl). These findings suggest that quercetin may effectively prevent liver damage caused by a high-fat diet through modulation of key molecular mechanisms. In conclusion, quercetin alleviated metabolic and pathological disturbances induced by high-fat feeding and demonstrated potential as a natural feed additive to support liver health in aquaculture. Future studies should evaluate dose–response relationships, long-term safety, and potential synergistic effects with other plant-derived bioactive compounds. Furthermore, to gain a more comprehensive and mechanistic understanding of these biological processes, advanced studies incorporating additional molecular markers and protein-level analyses related to oxidative stress signaling pathways, cellular redox regulation, and apoptotic mechanisms are needed.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author/s.
Ethics statement
The animal study was approved by Recep Tayyip Erdoğan University Experimental Animals Ethics Committee (decision number: 2023/30, date: June 13, 2023). The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
ÖK: Validation, Methodology, Writing – original draft, Conceptualization, Investigation, Data curation, Resources, Visualization, Formal Analysis, Writing – review & editing, Project administration.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This study has been supported by the Recep Tayyip Erdoğan University Development Foundation (Grant number: 02025007021617). This study was supported by Recep Tayyip Erdogan University Scientific Research Projects Coordination Unit with the grant ID: FBA-2023-15.
Acknowledgments
The author, Dr. OK, would like to thank Dr. Salih KUMRU for proofreading the article’s native language, and would like to thank Yusuf DEMİR for his help with record keeping during data collection.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abasubong K. P., Jiang G.-Z., Guo H., Wang X., Li X.-F., Yan-zou D., et al. (2023). High-fat diet alters intestinal microbiota and induces endoplasmic reticulum stress via the activation of apoptosis and inflammation in blunt snout bream. Fish Physiol. Biochem. 49, 1079–1095. doi: 10.1007/s10695-023-01240-2
Abdel-Latif H. M. R., Shukry M., Noreldin A. E., Ahmed H. A., El-Bahrawy A., Ghetas H. A., et al. (2023). Milk thistle (Silybum marianum) extract improves growth, immunity, serum biochemical indices, antioxidant state, hepatic histoarchitecture, and intestinal histomorphometry of striped catfish, Pangasianodon hypophthalmus. Aquaculture 562, 738761. doi: 10.1016/j.aquaculture.2022.738761
Abdo H. S., Mohammady E. Y., Tonsy H. D., Ibrahim A., and Hassaan M. S. (2024). The potential synergistic action of quercetin and/or Pediococcus acidilactici on Nile tilapia, Oreochromis niloticus performance. Aquaculture 581, 740353. doi: 10.1016/j.aquaculture.2023.740353
Ahmed A., Wong R. J., and Harrison S. A. (2015). Nonalcoholic fatty liver disease review: diagnosis, treatment, and outcomes. Clin. Gastroenterol. Hepatol. 13, 2062–2070. doi: 10.1016/j.cgh.2015.07.029
Akpınar Z., Sevgili H., Demir A., Özgen T., Emre Y., and Tufan Eroldoğan O. (2012). Effects of dietary lipid levels on growth, nutrient utilization, and nitrogen and carbon balances in shi drum (Umbrina cirrosa L.). Aquac. Int. 20, 131–143. doi: 10.1007/s10499-011-9447-7
AOAC (2005). “Official methods of analysis of AOAC INTERNATIONAL,”. Eds. Hortwitz W. and Latimer G. W. (AOAC. AOAC IN TER NA TIONAL, Maryland, USA).
Armobin K., Ahmadifar E., Adineh H., Samani M. N., Kalhor N., Yilmaz S., et al (2021). Quercetin application for common carp (Cyprinus carpio): I. Effects on growth performance, humoral immunity, antioxidant status, immune-related genes, and resistance against heat stress. Aquac. Nutr. 2023, 1–10. doi: 10.1155/2023/1168262
Asgary S., Naderi G. H., Sarrafzadegan N., Mohammadifard N., Mostafavi S., and Vakili R. (2000). Antihypertensive and antihyperlipidemic effects of Achillea wilhelmsii. Drugs Exp. Clin. Res. 26, 89–93.
Aydın B. and Gümüş E. (2020). Use of corn distiller’s dried grains with solubles as a feed ingredient for rainbow trout (Oncorhynchus mykiss) feeds: Growth, digestibility, liver and intestine histology. Iran. J. Fish. Sci. 19, 2573–2592. doi: 10.22092/ijfs.2020.122503
Azaza M. S., Saidi S. A., Dhraief M. N., and EL-feki A. (2020). Growth performance, nutrient digestibility, hematological parameters, and hepatic oxidative stress response in juvenile nile tilapia, oreochromis niloticus, fed carbohydrates of different complexities. Animals 10, 1913. doi: 10.3390/ani10101913
Bayraktar V. and Coşar A. M. (2019). Karaciğer enzim yüksekliklerine yaklaşım. Güncel Gastroenteroloji 23, 41–47.
Beamish F. W. H. and Medland T. E. (1986). Protein sparing effects in large rainbow trout, Salmo gairdneri. Aquaculture 55, 35–42. doi: 10.1016/0044-8486(86)90053-0
Boran H., Altinok I., and Capkin E. (2010). Histopathological changes induced by maneb and carbaryl on some tissues of rainbow trout, Oncorhynchus mykiss. Tissue Cell 42, 158–164. doi: 10.1016/j.tice.2010.03.004
Boran H., Ciftci C., Er A., Kose O., Kurtoglu I. Z., and Kayis S. (2015). Evaluation of Antibacterial Activity of Green Tea (Camellia sinensis L.) Seeds Against Some Fish Pathogens in Rainbow Trout (Oncorhynchus mykiss, Walbaum). Turkish J. Fish. Aquat. Sci. 15, 49–57. doi: 10.4194/1303-2712-v15_1_06
Brent A. N. (2010). Hepatic lipotoxicity and the pathogenesis of nonalcoholic steatohepatitis. Hepatology 52, 774–788. doi: 10.1002/hep.23719
Caballero M. J., Izquierdo M. S., Kjørsvik E., Fernández A. J., and Rosenlund G. (2004). Histological alterations in the liver of sea bream, Sparus aurata L., caused by short- or long-term feeding with vegetable oils. Recovery of normal morphology after feeding fish oil as the sole lipid source. J. Fish Dis. 27, 531–541. doi: 10.1111/j.1365-2761.2004.00572.x
Çakmak E., Özel O. T., Batır E., and Evin D. (2024). A new approach in rainbow trout (Oncorhynchus mykiss Walbaum 1792) industry: “Turkish salmon” production and marketing trends. Ege J. Fish. Aquat. Sci. 41, 69–81. doi: 10.12714/egejfas.41.1.10
Chirumbolo S. (2010). The role of quercetin, flavonols and flavones in modulating inflammatory cell function. Inflamm. Allergy - Drug Targets 9, 263–285. doi: 10.2174/187152810793358741
Chiu H. K., Qian K., Ogimoto K., Morton G. J., Wisse B. E., Agrawal N., et al. (2010). Mice lacking hepatic lipase are lean and protected against diet-induced obesity and hepatic steatosis. Endocrinology 151, 993–1001. doi: 10.1210/en.2009-1100
Choi J. W. (2003). Association between elevated serum hepatic enzyme activity and total body fat in obese humans. Ann. Clin. Lab. Sci. 33, 257–264.
Cunningham P., Patton E., VanderVeen B. N., Unger C., Aladhami A., and Enos R. T. (2021). Sub-chronic oral toxicity screening of quercetin in mice. BMC Complement. Med. Ther 22, 279. doi: 10.1186/s12906-022-03758-z
Dai W., Wang K., Zheng X., Chen X., Zhang W., Zhang Y., et al. (2015). High fat plus high cholesterol diet lead to hepatic steatosis in zebrafish larvae: a novel model for screening anti-hepatic steatosis drugs. Nutr. Metab. (Lond). 12, 42. doi: 10.1186/s12986-015-0036-z
Desouky H. E., Jiang G., Zhang D., Abasubong K. P., Yuan X., Li X., et al. (2020). Influences of glycyrrhetinic acid (GA) dietary supplementation on growth, feed utilization, and expression of lipid metabolism genes in channel catfish (Ictalurus punctatus) fed a high-fat diet. Fish Physiol. Biochem. 46, 653–663. doi: 10.1007/s10695-019-00740-4
Du Z.-Y., Liu Y.-J., Tian L.-X., Wang J.-T., Wang Y., and Liang G.-Y. (2005). Effect of dietary lipid level on growth, feed utilization and body composition by juvenile grass carp (Ctenopharyngodon idella). Aquac. Nutr. 11, 139–146. doi: 10.1111/j.1365-2095.2004.00333.x
Du Z.-Y., Ma T., Liaset B., Keenan A. H., Araujo P., Lock E.-J., et al. (2013). Dietary eicosapentaenoic acid supplementation accentuates hepatic triglyceride accumulation in mice with impaired fatty acid oxidation capacity. Biochim. Biophys. Acta - Mol. Cell Biol. Lipids 1831, 291–299. doi: 10.1016/j.bbalip.2012.10.002
Du J., Wang X., Miereles C., Bailey J. L., Debigare R., Zheng B., et al. (2004). Activation of caspase-3 is an initial step triggering accelerated muscle proteolysis in catabolic conditions. J. Clin. Invest. 113, 115–123. doi: 10.1172/JCI18330
Earnshaw W. C., Martins L. M., and Kaufmann S. H. (1999). Mammalian caspases: structure, activation, substrates, and functions during apoptosis. Annu. Rev. Biochem. 68, 383–424. doi: 10.1146/annurev.biochem.68.1.383
Farmer E. E. and Davoine C. (2007). Reactive electrophile species. Curr. Opin. Plant Biol. 10, 380–386. doi: 10.1016/j.pbi.2007.04.019
Gadallah A. A. and Almasari A. R. (2023). The effects of a high-fat diet on the liver of pregnant albino rats and their developing offspring. Egypt. J. Basic Appl. Sci. 10, 567–579. doi: 10.1080/2314808X.2023.2245618
Ghafarifarsani H., Hoseinifar S. H., Javahery S., and Van Doan H. (2022a). Effects of dietary vitamin C, thyme essential oil, and quercetin on the immunological and antioxidant status of common carp (Cyprinus carpio). Aquaculture 553, 738053. doi: 10.1016/j.aquaculture.2022.738053
Ghafarifarsani H., Hoseinifar S. H., Javahery S., Yazici M., and Van Doan H. (2022b). Growth performance, biochemical parameters, and digestive enzymes in common carp (Cyprinus carpio) fed experimental diets supplemented with vitamin C, thyme essential oil, and quercetin. Ital. J. Anim. Sci. 21, 291–302. doi: 10.1080/1828051X.2021.1965923
Gottfredsen R. H., Goldstrohm D. A., Hartney J. M., Larsen U. G., Bowler R. P., and Petersen S. V. (2014). The cellular distribution of extracellular superoxide dismutase in macrophages is altered by cellular activation but unaffected by the naturally occurring R213G substitution. Free Radic. Biol. Med. 69, 348–356. doi: 10.1016/j.freeradbiomed.2014.01.038
Han T., Li X., Wang J., Hu S., Jiang Y., and Zhong X. (2014). Effect of dietary lipid level on growth, feed utilization and body composition of juvenile giant croaker Nibea japonica. Aquaculture 434, 145–150. doi: 10.1016/j.aquaculture.2014.08.012
Hillestad M. and Johnsen F. (1994). High-energy/low-protein diets for Atlantic salmon: effects on growth, nutrient retention and slaughter quality. Aquaculture 124, 109–116. doi: 10.1016/0044-8486(94)90366-2
Jia R., Cao L.-P., Du J.-L., He Q., Gu Z.-Y., Jeney G., et al. (2020). Effects of high-fat diet on antioxidative status, apoptosis and inflammation in liver of tilapia (Oreochromis niloticus) via Nrf2, TLRs and JNK pathways. Fish Shellfish Immunol. 104, 391–401. doi: 10.1016/j.fsi.2020.06.025
Jia E., Yan Y., Zhou M., Li X., Jiang G., Liu W., et al. (2019). Combined effects of dietary quercetin and resveratrol on growth performance, antioxidant capability and innate immunity of blunt snout bream (Megalobrama amblycephala). Anim. Feed Sci. Technol. 256, 114268. doi: 10.1016/j.anifeedsci.2019.114268
Jin M., Pan T., Tocher D. R., Betancor M. B., Monroig Ó., Shen Y., et al. (2019). Dietary choline supplementation attenuated high-fat diet-induced inflammation through regulation of lipid metabolism and suppression of NFκB activation in juvenile black seabream (Acanthopagrus schlegelii). J. Nutr. Sci. 8, e38. doi: 10.1017/jns.2019.34
Jobling M., Koskela J., and Savolainen R. (1998). Influence of dietary fat level and increased adiposity on growth and fat deposition in rainbow trout, Oncorhynchus mykiss (Walbaum). Aquac. Res. 29, 601–607. doi: 10.1046/j.1365-2109.1998.00251.x
Jung C. H., Cho I., Ahn J., Jeon T., and Ha T. (2013). Quercetin reduces high-fat diet-induced fat accumulation in the liver by regulating lipid metabolism genes. Phyther. Res. 27, 139–143. doi: 10.1002/ptr.4687
Kawakami Y., Tsurugasaki W., Nakamura S., and Osada K. (2005). Comparison of regulative functions between dietary soy isoflavones aglycone and glucoside on lipid metabolism in rats fed cholesterol. J. Nutr. Biochem. 16, 205–212. doi: 10.1016/j.jnutbio.2004.11.005
Khalaf M. M. and Salih R. A. (2023). Investigating the potential hepatoprotective effect of quercetin in male rats Following acute exposure to cyclophosphamide. Iraqi J. Vet. Med. 47, 23–30. doi: 10.30539/ijvm.v47i2.1555
Kim S.-S., Shin S.-J., Han H.-S., Kim J.-D., and Lee K.-J. (2012). Effects of Dietary Spirulina pacifica on Innate Immunity and Disease Resistance against Edwardsiella tarda in Olive Flounder Paralichthys olivaceus. Isr. J. Aquac. - Bamidgeh 16, 7–14. doi: 10.46989/001c.20716
Kleveland E. J., Ruyter B., Vegusdal A., Sundvold H., Berge R. K., and Gjøen T. (2006). Effects of 3-thia fatty acids on expression of some lipid related genes in Atlantic salmon (Salmo salar L.). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 145, 239–248. doi: 10.1016/j.cbpb.2006.07.012
Kong Y., Tian J., Niu X., Li M., Li R., Chen X., et al. (2022). Effects of dietary quercetin on growth, antioxidant capacity, immune response and immune-related gene expression in snakehead fish, Channa argus. Aquac. Rep. 26, 101314. doi: 10.1016/j.aqrep.2022.101314
Köse Ö., Arıman Karabulut H., Kurtoğlu İ. Z., and Er A. (2021). Effects of feed rations containing egg powder in different proportions on growth performance, feed utilization, body composition and survival rate of rainbow trout (Oncorhynchus mykiss Walbaum, 1792). Ege J. Fish. Aquat. Sci. 38, 69–78. doi: 10.12714/egejfas.38.1.08
Kose O. and Karabulut H. A. (2022). Effects of dandelion (Taraxacum officinale) methanolic root leaf and flower extracts on growth performance, body composition and hematologic blood parameters of fingerling rainbow trout (Oncorhynchus mykiss Walbaum 1792). Fresenius Environ. Bull. 31, 4459–71.
Köse Ö., Karabulut H. A., and Er A. (2024). Dandelion Root Extract in Trout Feed and its Effects on the Physiological Performance of Oncorhynchus mykiss and Resistance to Lactococcus garvieae Infection. Ann. Anim. Sci. 24, 161–177. doi: 10.2478/aoas-2023-0072
Kuipers E. N., Dam A. D., Held N. M., Mol I. M., Houtkooper R. H., Rensen P. C. N., et al. (2018). Quercetin lowers plasma triglycerides accompanied by white adipose tissue browning in diet-induced obese mice. Int. J. Mol. Sci. 19, 1786. doi: 10.3390/ijms19061786
Lee Y. J., Park I. S., Song J. H., Oh M. H., Nam H. S., Cho M. K., et al. (2015). Quercetin exerts preferential cytotoxic effects on Malignant mesothelioma cells by inducing p53 expression, caspase-3 activation, and apoptosis. Mol. Cell. Toxicol. 11, 295–305. doi: 10.1007/s13273-015-0029-z
Li L., Chu X., Yao Y., Cao J., Li Q., and Ma H. (2020). (–)-hydroxycitric acid alleviates oleic acid-induced steatosis, oxidative stress, and inflammation in primary chicken hepatocytes by regulating AMP-activated protein kinase-mediated reactive oxygen species levels. J. Agric. Food Chem. 68, 11229–11241. doi: 10.1021/acs.jafc.0c04648
Li X.-F., Jiang G.-Z., Qian Y., Xu W.-N., and Liu W.-B. (2013). Molecular characterization of lipoprotein lipase from blunt snout bream Megalobrama amblycephala and the regulation of its activity and expression by dietary lipid levels. Aquaculture 416–417, 23–32. doi: 10.1016/j.aquaculture.2013.08.020
Li C. P., Li J. H., He S. Y., Li P., and Zhong X. L. (2014). Roles of Fas/Fasl, Bcl-2/Bax, and Caspase-8 in rat nonalcoholic fatty liver disease pathogenesis. Genet. Mol. Res. 13, 3991–3999. doi: 10.4238/2014.May.23.10
Li A., Yuan X., Liang X.-F., Liu L., Li B., Fang J., et al. (2016). Adaptations of lipid metabolism and food intake in response to low and high fat diets in juvenile grass carp (Ctenopharyngodon idellus). Aquaculture 457, 43–49. doi: 10.1016/j.aquaculture.2016.01.014
Li Z.-H., Zlabek V., Velisek J., Grabic R., Machova J., and Randak T. (2010). Modulation of antioxidant defence system in brain of rainbow trout (Oncorhynchus mykiss) after chronic carbamazepine treatment. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 151, 137–141. doi: 10.1016/j.cbpc.2009.09.006
Liang X.-F., Ogata H. Y., and Oku H. (2002). Effect of dietary fatty acids on lipoprotein lipase gene expression in the liver and visceral adipose tissue of fed and starved red sea bream Pagrus major. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 132, 913–919. doi: 10.1016/S1095-6433(02)00118-6
Liu Y.-C., Limbu S. M., Wang J.-G., Ren J., Qiao F., Zhang M.-L., et al. (2022). Dietary L-carnitine alleviates the adverse effects caused by reducing protein and increasing fat contents in diet juvenile largemouth bass (Micropterus salmoides). Aquac. Nutr. 2022, 1–14. doi: 10.1155/2022/6288972
Liu Z.-L., Zhao W., Hu W.-S., Zhu B., Xie J.-J., Liu Y.-J., et al. (2021). Lipid metabolism, growth performance, antioxidant ability and intestinal morphology of rainbow trout (Oncorhynchus mykiss) under cage culture with flowing water were affected by dietary lipid levels. Aquac. Rep. 19, 100593. doi: 10.1016/j.aqrep.2021.100593
López L. M., Torres A. L., Durazo E., Drawbridge M., and Bureau D. P. (2006). Effects of lipid on growth and feed utilization of white seabass (Atractoscion nobilis) fingerlings. Aquaculture 253, 557–563. doi: 10.1016/j.aquaculture.2005.08.007
Lu K.-L., Wang L.-N., Zhang D.-D., Liu W.-B., and Xu W.-N. (2017). Berberine attenuates oxidative stress and hepatocytes apoptosis via protecting mitochondria in blunt snout bream Megalobrama amblycephala fed high-fat diets. Fish Physiol. Biochem. 43, 65–76. doi: 10.1007/s10695-016-0268-5
Lu K.-L., Xu W.-N., Wang L.-N., Zhang D.-D., Zhang C.-N., and Liu W.-B. (2014). Hepatic β-oxidation and regulation of carnitine palmitoyltransferase (CPT) I in blunt snout bream megalobrama amblycephala fed a high fat diet. PloS One 9, e93135. doi: 10.1371/journal.pone.0093135
Luo Z., Liu Y.-J., Mai K.-S., Tian L.-X., Liu D.-H., Tan X.-Y., et al. (2005). Effect of dietary lipid level on growth performance, feed utilization and body composition of grouper Epinephelus coioides juveniles fed isonitrogenous diets in floating netcages. Aquac. Int. 13, 257–269. doi: 10.1007/s10499-004-2478-6
Ma Q., Li L.-Y., Le J.-Y., Lu D.-L., Qiao F., Zhang M.-L., et al. (2018). Dietary microencapsulated oil improves immune function and intestinal health in Nile tilapia fed with high-fat diet. Aquaculture 496, 19–29. doi: 10.1016/j.aquaculture.2018.06.080
Maalik A., Khan F., Mumtaz A., Mehmood A., Azhar S., Atif M., et al. (2014). Pharmacological applications of quercetin and its derivatives: A short review. Trop. J. Pharm. Res. 13, 1561–1566. doi: 10.4314/tjpr.v13i9.26
Mansour A. T., Mahboub H. H., Amen R. M., El-Beltagy M. A., Ramah A., Abdelfattah A. M., et al. (2022). Ameliorative Effect of Quercetin against Abamectin-Induced Hemato-Biochemical Alterations and Hepatorenal Oxidative Damage in Nile Tilapia, Oreochromis niloticus. Animals 12, 3429. doi: 10.3390/ani12233429
Mariotti F., Tomé D., and Mirand P. P. (2008). Converting nitrogen into protein—Beyond 6.25 and jones’ Factors. Crit. Rev. Food Sci. Nutr. 48, 177–184. doi: 10.1080/10408390701279749
Martínez-Llorens S., Baeza-Ariño R., Nogales-Mérida S., Jover-Cerdá M., and Tomás-Vidal A. (2012). Carob seed germ meal as a partial substitute in gilthead sea bream (Sparus aurata) diets: Amino acid retention, digestibility, gut and liver histology. Aquaculture 338–341, 124–133. doi: 10.1016/j.aquaculture.2012.01.029
Matejova I., Faldyna M., Modra H., Blahova J., Palikova M., Markova Z., et al. (2017). Effect of T-2 toxin-contaminated diet on common carp (Cyprinus carpio L.). Fish Shellfish Immunol. 60, 458–465. doi: 10.1016/j.fsi.2016.11.032
Matsumoto T., Terai S., Oishi T., Kuwashiro S., Fujisawa K., Yamamoto N., et al. (2010). Medaka as a model for human nonalcoholic steatohepatitis. Dis. Model. Mech. 3, 431–440. doi: 10.1242/dmm.002311
McArthur K. and Kile B. T. (2018). Apoptotic caspases: multiple or mistaken identities? Trends Cell Biol. 28, 475–493. doi: 10.1016/j.tcb.2018.02.003
McGoogan B. B. and Gatlin D. M. (1999). Dietary manipulations affecting growth and nitrogenous waste production of red drum, Sciaenops ocellatus I. Effects of dietary protein and energy levels. Aquaculture 178, 333–348. doi: 10.1016/S0044-8486(99)00137-4
Meng Y., Liu X., Guan L., Bao S., Zhuo L., Tian H., et al. (2023). Does dietary lipid level affect the quality of triploid rainbow trout and how should it be assessed? Foods 12, 15. doi: 10.3390/foods12010015
Meng Y., Qian K., Ma R., Liu X., Han B., Wu J., et al. (2019). Effects of dietary lipid levels on sub-adult triploid rainbow trout (Oncorhynchus mykiss): 1. Growth performance, digestive ability, health status and expression of growth-related genes. Aquaculture 513, 734394. doi: 10.1016/j.aquaculture.2019.734394
Mensinger A. F., Walsh P. J., and Hanlon R. T. (2005). Blood biochemistry of the oyster toadfish. J. Aquat. Anim. Health 17, 170–176. doi: 10.1577/H04-021.1
Miean K. H. and Mohamed S. (2001). Flavonoid (Myricetin, quercetin, kaempferol, luteolin, and apigenin) content of edible tropical plants. J. Agric. Food Chem. 49, 3106–3112. doi: 10.1021/jf000892m
Morovvati H. and Armand N. (2019). Assessment of changes in serum concentrations of liver function (ALT, ALP, AST, LDH, GGT) after the intake of narcissus bulbs. J. Fasa Univ. Med. Sci. 9, 1736–1742.
Moseti D., Regassa A., and Kim W.-K. (2016). Molecular regulation of adipogenesis and potential anti-adipogenic bioactive molecules. Int. J. Mol. Sci. 17, 124. doi: 10.3390/ijms17010124
Nakamura Y., Ishimitsu S., and Tonogai Y. (2000). Effects of quercetin and rutin on serum and hepatic lipid concentrations, fecal steroid excretion and serum antioxidant properties. J. Heal. Sci. 46, 229–240. doi: 10.1248/jhs.46.229
Nasirin C., Najm M. A. A., Chen T.-C., Dhamija A., Lionardo A., Bokov D. O., et al. (2023). The protective effects of quercetin on the physiological responses in malathion-exposed common carp, Cyprinus carpio. Trop. Anim. Health Prod. 55, 22. doi: 10.1007/s11250-022-03429-8
NRC (2011). Nutrient requirements of fish and shrimp (Washington, D.C: National Academies Press). doi: 10.17226/13039
Pagliassotti M. J. (2012). Endoplasmic reticulum stress in nonalcoholic fatty liver disease. Annu. Rev. Nutr. 32, 17–33. doi: 10.1146/annurev-nutr-071811-150644
Park C., So H.-S., Shin C.-H., Baek S.-H., Moon B.-S., Shin S.-H., et al. (2003). Quercetin protects the hydrogen peroxide-induced apoptosis via inhibition of mitochondrial dysfuntion in H9c2 cardiomyoblast cells. Biochem. Pharmacol. 66, 1287–1295. doi: 10.1016/S0006-2952(03)00478-7
Prysyazhnyuk V. P. and Voloshyn O. I. (2017). Effects of comprehensive treatment with quercetin administration on biochemical blood parameters and pro-and anti-inflammatory cytokines in nonalcoholic fatty liver disease patients. Pharma Innov. J. 6, 386–389.
Rajesh E., Sankari L., Malathi L., and Krupaa J. (2015). Naturally occurring products in cancer therapy. J. Pharm. Bioallied Sci. 7, 181. doi: 10.4103/0975-7406.155895
Ruiz L. M., Salazar C., Jensen E., Ruiz P. A., Tiznado W., Quintanilla R. A., et al. (2015). Quercetin affects erythropoiesis and heart mitochondrial function in mice. Oxid. Med. Cell. Longev. 2015, 1–12. doi: 10.1155/2015/836301
Saponaro C., Gaggini M., Carli F., and Gastaldelli A. (2015). The subtle balance between lipolysis and lipogenesis: A critical point in metabolic homeostasis. Nutrients 7, 9453–9474. doi: 10.3390/nu7115475
Schmittgen T. D. and Livak K. J. (2008). Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 3, 1101–1108. doi: 10.1038/nprot.2008.73
Schwerk C. and Schulze-Osthoff K. (2003). Non-apoptotic functions of caspases in cellular proliferation and differentiation. Biochem. Pharmacol. 66, 1453–1458. doi: 10.1016/S0006-2952(03)00497-0
Sekar M. and Thirumurugan K. (2022). Autophagy: a molecular switch to regulate adipogenesis and lipolysis. Mol. Cell. Biochem. 477, 727–742. doi: 10.1007/s11010-021-04324-w
Seo Y.-S., Kang O.-H., Kım S.-B., Mun S.-H., Kang D.-H., Yang D.-W., et al. (2015). Quercetin prevents adipogenesis by regulation of transcriptional factors and lipases in OP9 cells. Int. J. Mol. Med. 35, 1779–1785. doi: 10.3892/ijmm.2015.2185
Sevgili H., Kurtoğlu A., Oikawa M., Öztürk E., Dedebali N., Emre N., et al. (2014). High dietary lipids elevate carbon loss without sparing protein in adequate protein-fed juvenile turbot (Psetta maxima). Aquac. Int. 22, 797–810. doi: 10.1007/s10499-013-9708-8
Shadegan M. R. and Banaee M. (2018). Effects of dimethoate alone and in combination with Bacilar fertilizer on oxidative stress in common carp, Cyprinus carpio. Chemosphere 208, 101–107. doi: 10.1016/j.chemosphere.2018.05.177
Shahrooz R. O., Agh N., Jafari N., Kalantari A., Jalili R., and Karimi A. (2018). Effects of fish oil replacement with vegetable oils in rainbow trout (Oncorhynchus mykiss) fingerlings diet on growth performance and foregut histology. Turkish J. Fish. Aquat. Sci. 18, 825–832. doi: 10.4194/1303-2712-V18_6_09
Shearer K. D. (1994). Factors affecting the proximate composition of cultured fishes with emphasis on salmonids. Aquaculture 119, 63–88. doi: 10.1016/0044-8486(94)90444-8
Shimano H. (2001). Sterol regulatory element-binding proteins (SREBPs): transcriptional regulators of lipid synthetic genes. Prog. Lipid Res. 40, 439–452. doi: 10.1016/S0163-7827(01)00010-8
Shin H. S., Yoo J. H., Min T. S., Lee J., and Choi C. Y. (2010a). Effect of Quercetin on the Activity and mRNA Expression of Antioxidant Enzymes and Physiological Responses in Olive Flounder (Paralichthys olivaceus) Exposed to Cadmium. Asian-Australasian J. Anim. Sci. 23, 742–749. doi: 10.5713/ajas.2010.10006
Shin H. S., Yoo J. H., Min T. S., Lee K.-Y., and Choi C. Y. (2010b). The effects of quercetin on physiological characteristics and oxidative stress resistance in olive flounder, paralichthys olivaceus. Asian-Australasian J. Anim. Sci. 23, 588–597. doi: 10.5713/ajas.2010.90624
Strüssmann C. A. and Takashima F. (1990). Hepatocyte nuclear size and nutritional condition of larval pejerrey, Odontesthes bonariensis (Cuvier et Valenciennes). J. Fish Biol. 36, 59–65. doi: 10.1111/j.1095-8649.1990.tb03519.x
Sukhovskaya I. V., Lysenko L. A., Fokina N. N., Kantserova N. P., and Borvinskaya E. V. (2023). Survival, growth performance, and hepatic antioxidant and lipid profiles in infected rainbow trout (Oncorhynchus mykiss) fed a diet supplemented with dihydroquercetin and arabinogalactan. Animals 13. doi: 10.3390/ani13081345
Tang T., Yi Hu, Peng M., Chu W., Yajun Hu, and Zhong L. (2019). Effects of high-fat diet on growth performance, lipid accumulation and lipid metabolism-related MicroRNA/gene expression in the liver of grass carp (Ctenopharyngodon idella). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 234, 34–40. doi: 10.1016/j.cbpb.2019.04.006
Tao Y.-F., Qiang J., Bao J.-W., Chen D.-J., Yin G.-J., Xu P., et al. (2018). Changes in physiological parameters, lipid metabolism, and expression of microRNAs in genetically improved farmed tilapia (Oreochromis niloticus) with fatty liver induced by a high-fat diet. Front. Physiol. 9. doi: 10.3389/fphys.2018.01521
Thapaliya S., Wree A., Povero D., Inzaugarat M. E., Berk M., Dixon L., et al. (2014). Caspase 3 inactivation protects against hepatic cell death and ameliorates fibrogenesis in a diet-induced NASH model. Dig. Dis. Sci. 59, 1197–1206. doi: 10.1007/s10620-014-3167-6
Tocher D. R., Mourente G., van der Eecken A., Evjemo J. O., Diaz E., Bell J. G., et al. (2002). Effects of dietary vitamin E on antioxidant defence mechanisms of juvenile turbot (Scophthalmus maximus L.), halibut (Hippoglossus hippoglossus L.) and sea bream (Sparus aurata L.). Aquac. Nutr. 8, 195–207. doi: 10.1046/j.1365-2095.2002.00205.x
Tran-Duy A., van Dam A. A., and Schrama J. W. (2012). Feed intake, growth and metabolism of Nile tilapia (Oreochromis niloticus) in relation to dissolved oxygen concentration. Aquac. Res. 43, 730–744. doi: 10.1111/j.1365-2109.2011.02882.x
Tuncel F. and Alpaslan Duman A. (2021). Kronik viral hepatitli olgularda kombine non-alkolik yağlı karaciğer hastalığının sıklığı ve önemi. Mersin Üniversitesi Sağlık Bilim. Derg. 14, 77–86. doi: 10.26559/mersinsbd.883097
Wang Y., Ausman L. M., Russell R. M., Greenberg A. S., and Wang X.-D. (2008). Increased apoptosis in high-fat diet–induced nonalcoholic steatohepatitis in rats is associated with c-jun NH2-terminal kinase activation and elevated proapoptotic bax. J. Nutr. 138, 1866–1871. doi: 10.1093/jn/138.10.1866
Wang X., Li Y., Hou C., Gao Y., and Wang Y. (2015). Physiological and molecular changes in large yellow croaker (Pseudosciaena crocea R.) with high-fat diet-induced fatty liver disease. Aquac. Res. 46, 272–282. doi: 10.1111/are.12176
Wang J.-T., Liu Y.-J., Tian L.-X., Mai K.-S., Du Z.-Y., Wang Y., et al. (2005). Effect of dietary lipid level on growth performance, lipid deposition, hepatic lipogenesis in juvenile cobia (Rachycentron canadum). Aquaculture 249, 439–447. doi: 10.1016/j.aquaculture.2005.04.038
Wang J., Zhang C., Zhang J., Xie J., Yang L., Xing Y., et al. (2020). The effects of quercetin on immunity, antioxidant indices, and disease resistance in zebrafish (Danio rerio). Fish Physiol. Biochem. 46, 759–770. doi: 10.1007/s10695-019-00750-2
Xia T., Mao X., Zhang J., Rahimnejad S., and Lu K. (2024). Effects of quercetin and hydroxytyrosol supplementation in a high-fat diet on growth, lipid metabolism, and mitochondrial function in spotted seabass (Lateolabrax maculatus). Aquaculture 582, 740538. doi: 10.1016/j.aquaculture.2023.740538
Xie S., Lin Y. Y., Wu T., Tian L., Liang J., and Tan B. (2021). Dietary lipid levels affected growth performance, lipid accumulation, inflammatory response and apoptosis of Japanese seabass (lateolabrax japonicus). Aquac. Nutr. 27, 807–816. doi: 10.1111/anu.13225
Xie N., Wen H., Xie S., Jiang M., Yu L., Wu F., et al. (2021). Adaptations of hepatic lipid and glucose metabolism in response to high-macronutrient diets in juvenile grass carp. Aquac. Nutr. 27, 1738–1749. doi: 10.1111/anu.13311
Xu Z., Li X., Yang H., Liang G., Gao B., and Leng X. (2019). Dietary quercetin improved the growth, antioxidation, and flesh quality of grass carp (Ctenopharyngodon idella). J. World Aquac. Soc 50, 1182–1195. doi: 10.1111/jwas.12663
Xu D., Xiang X., Li X., Xu N., Zhang W., Mai K., et al. (2022). Effects of dietary vegetable oils replacing fish oil on fatty acid composition, lipid metabolism and inflammatory response in adipose tissue of large yellow croaker (Larimichthys crocea). J. Mar. Sci. Eng. 10, 1760. doi: 10.3390/jmse10111760
Yamamoto Y. and Oue E. (2006). Antihypertensive effect of quercetin in rats fed with a high-fat high-sucrose diet. Biosci. Biotechnol. Biochem. 70, 933–939. doi: 10.1271/bbb.70.933
Yan J., Liao K., Wang T., Mai K., Xu W., and Ai Q. (2015). Dietary Lipid Levels Influence Lipid Deposition in the Liver of Large Yellow Croaker (Larimichthys crocea) by Regulating Lipoprotein Receptors, Fatty Acid Uptake and Triacylglycerol Synthesis and Catabolism at the Transcriptional Level. PloS One 10, e0129937. doi: 10.1371/journal.pone.0129937
Yıldırım Ö. and Çantaş İ.B. (2022). Investigation of production and economic indicators of rainbow trout farming in Türkiye. Acta Aquat. Turc. 18, 461–474. doi: 10.22392/actaquatr.1101098
Yong A. S. K., Ooi S., Shapawi R., Biswas A. K., and Kenji T. (2015). Effects of dietary lipid increments on growth performance, feed utilization, carcass composition and intraperitoneal fat of marble goby, oxyeleotris marmorata, juveniles. Turkish J. Fish. Aquat. Sci. 15, 653–660. doi: 10.4194/1303-2712-v15_3_10
Yugarani T., Tan B. K. H., Teh M., and Das N. P. (1992). Effects of polyphenolic natural products on the lipid profiles of rats fed high fat diets. Lipids 27, 181–186. doi: 10.1007/BF02536175
Zhai S.-W. and Liu S.-L. (2013). Effects of dietary quercetin on growth performance, serum lipids level and body composition of tilapia (Oreochromis niloticus). Ital. J. Anim. Sci. 12, e85. doi: 10.4081/ijas.2013.e85
Zhang Y., Jun-Xian W., Ren J., Jie W., Qiao F., Zhang M., et al. (2021). Activation of peroxisome proliferator-activated receptor-α stimulated lipid catabolism in liver of largemouth bass, Micropterus salmoides. Aquac. Nutr. 27, 2749–2758. doi: 10.1111/anu.13400
Zhao X., Wang D., Zhao Z., Yingyi X., Sengupta S., Yijin X., et al. (2006). Caspase-3-dependent activation of calcium-independent phospholipase A2 enhances cell migration in non-apoptotic ovarian cancer cells. J. Biol. Chem. 281, 29357–29368. doi: 10.1074/jbc.M513105200
Zheng K., Zhu X., Han D., Yang Y., Lei W., and Xie S. (2010). Effects of dietary lipid levels on growth, survival and lipid metabolism during early ontogeny of Pelteobagrus vachelli larvae. Aquaculture 299, 121–127. doi: 10.1016/j.aquaculture.2009.11.028
Zhou C., Li P., Han M., and Gao X. (2022). Daidzein stimulates fatty acid-induced fat deposition in C2C12 myoblast cells via the G protein-coupled receptor 30 pathway. Anim. Biotechnol. 33, 851–863. doi: 10.1080/10495398.2020.1842749
Zhou C., Lin H., Ge X., Niu J., Wang J., Wang Y., et al. (2015). The Effects of dietary soybean isoflavones on growth, innate immune responses, hepatic antioxidant abilities and disease resistance of juvenile golden pompano Trachinotus ovatus. Fish Shellfish Immunol. 43, 158–166. doi: 10.1016/j.fsi.2014.12.014
Zou C., Du L., Wu J., Gan S., Li Q., Sarath Babu V., et al. (2022). Saikosaponin d alleviates high-fat-diet induced hepatic steatosis in hybrid grouper (Epinephelus lanceolatus♂ × Epinephelus fuscoguttatus♀) by targeting AMPK/PPARα pathway. Aquaculture 553, 738088. doi: 10.1016/j.aquaculture.2022.738088
Keywords: antioxidant status, gene expression, lipolysis, lipogenesis, liver histology
Citation: Köse Ö (2025) Quercetin: a natural remedy against high-fat diet-induced liver steatosis in Oncorhynchus mykiss. Front. Mar. Sci. 12:1656703. doi: 10.3389/fmars.2025.1656703
Received: 30 June 2025; Accepted: 28 July 2025;
Published: 15 August 2025.
Edited by:
Gladstone Sagada, Victory Farms, KenyaReviewed by:
Yuwen Dong, University of Pennsylvania Division of Gastroenterology and Hepatology, United StatesAfef Amri, Institut National des Sciences et Technologies de la Mer, Tunisia
Copyright © 2025 Köse. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Özay Köse, b3pheS5rb3NlQGVyZG9nYW4uZWR1LnRy
†ORCID: Özay Köse, orcid.org/0000-0002-3565-160X