 Veronica L. M. Coppolaro
Veronica L. M. Coppolaro Emma L. Ausen
Emma L. Ausen Lisa L. Loseto
Lisa L. Loseto Marianne Marcoux
Marianne Marcoux- 1Centre for Earth Observation Science, University of Manitoba, Winnipeg, MB, Canada
- 2Institute for the Study of Anthropic Impacts and Sustainability in the Marine Environment, National Research Council of Italy, Genova, Italy
- 3Fisheries and Oceans Canada, Freshwater Institute, Winnipeg, MB, Canada
Vessel traffic and underwater noise pollution are increasing in the Arctic. Marine mammals are sensitive to underwater noise from vessels which can negatively impact them at the individual and population levels. The marine region of Southampton Island, Nunavut, Canada, is a recognized key area for many marine mammal species and is under consideration to become a marine protected area. Given the increase in vessel traffic in the region, this study explores the potential impact of vessel traffic noise on the vocal behavior of walruses and belugas. This represents the first study to investigate walrus vocal behavior during exposure to vessels. Underwater acoustic data were collected near Southampton Island from June to November 2018. Vessel movements were tracked using the Automatic Identification System (AIS) data and compared with underwater recordings to identify noise sources by vessel type (ship or motorboat). Generalized linear mixed models were used to assess changes in walrus vocalization rates before, during, and after vessel encounters across vessel type. The results showed that walrus vocalization rates decreased during and after vessel encounters and were significantly lower in the presence of ships than motorboats. Belugas were never recorded during motorboat transits, which may indicate avoidance behavior. However, there was not enough data to investigate this hypothesis further. Our findings demonstrate that vessel traffic influences walrus vocal behavior and highlight the need for updated maritime navigation mitigation measures in the study area.
Introduction
Global vessel traffic has increased rapidly in recent decades, driven by a growing merchant fleet and an increase in international maritime transportation, which now accounts for 80% of the total volume of trade goods (UNCTAD, 2022). As a result, underwater noise from vessel traffic has also intensified and is now nearly ubiquitous, reaching even marine areas that are far from major shipping lanes (Clark et al., 2009; Duarte et al., 2021; Thomsen and Popper, 2024). Once largely inaccessible, the Arctic is losing sea ice at unprecedented rates, facilitating a fast increase in vessel traffic in the region (Pizzolato et al., 2016; Andrews et al., 2018; Dawson et al., 2018; Stevenson et al., 2019). Maritime traffic in the Arctic has never been this high, nor its underwater soundscape so altered by human activities (PAME, 2024).
The main bandwidth of vessel underwater noise ranges from 10 Hz to 1 kHz (Richardson et al., 1995; Malakoff, 2010), overlapping with the frequency range used by many marine mammal species, which is typically between 10 Hz and 20 kHz (Erbe et al., 2018; Duarte et al., 2021). Recognized as a pervasive pollutant, underwater noise can negatively affect marine ecosystems and has been shown to cause a multitude of impacts on marine mammals, including communication masking, temporary behavioral changes, hearing loss, stranding, stress-induced health problems, permanent abandonment of biologically important areas, and, over time, potential consequences at the population level (Erbe et al., 2018; Southall, 2021; Tervo et al., 2021; Pirotta et al., 2022; Sweeney et al., 2022). Arctic marine mammals can be especially vulnerable to vessel noise (Moore et al., 2012; Hauser et al., 2018), making it critical to understand their behavioral responses to such disturbances.
The Southampton Island Area of Interest (AOI) in Hudson Bay, Nunavut, is under consideration for designation as a marine protected area (MPA) (Loewen et al., 2020). Vessel traffic in this region is increasing rapidly and is expected to grow further (Andrews et al., 2018; Dawson et al., 2018). The local community of Salliq (ᓴᓪᓕᖅ)/Salliit (ᓴᓪᓖᑦ) – Coral Harbour has expressed concern over the impact of vessels on marine mammals, particularly Atlantic walrus (Odobenus rosmarus (Linnaeus, 1758)) and beluga (Delphinapterus leucas (Pallas, 1776)) (COSEWIC, 2017; Carter et al., 2019; Loewen et al., 2020). The community has identified vessel activity as a major driver of changes in species distribution and abundance and called for changes in local maritime operations (Carter et al., 2019; Dawson et al., 2020). Gaining a better knowledge of vessel traffic impacts on marine mammals in this proposed MPA is essential for informed conservation efforts. However, current data on marine mammal habitat use in the AOI are limited (Loewen et al., 2020; Coppolaro et al., 2024), and no studies have yet investigated the acoustic responses of marine mammal species to vessels in the area.
Underwater passive acoustic monitoring (PAM) systems enable autonomous and continuous monitoring of marine mammals (Sousa-Lima et al., 2013; Heenehan et al., 2019; Halliday et al., 2020; Kline et al., 2020; Castellote et al., 2021) as well as the natural and anthropogenic sounds in their environment. PAM data can greatly contribute to assessing how marine species respond to vessel traffic. In Hudson Bay, studies examining walrus and beluga responses to vessels are limited and primarily based on visual observations (Caron and Smith, 1990; Mansfield and St. Aubin, 1991; Born et al., 1995; Malcolm and Penner, 2011; Ausen et al., 2022; Higdon et al., 2022). To date, no acoustic studies have examined walrus responses to vessel noise, and little is known about beluga vocal reactions to motorboats (Lesage et al., 1999; Karlsen et al., 2002).
This study aims to (1) integrate Automatic Identification System (AIS) data with PAM recordings to document vessel traffic in the AOI, distinguishing between ships and motorboats, and (2) investigate the underwater vocal responses of walruses and belugas to vessel transits in the Southampton Island AOI.
Materials and methods
Acoustic data collection
In June 2018, a TR-ORCA hydrophone (Turbulent Research)1 was deployed in Evans Strait, in the southern part of the Southampton Island AOI, Nunavut, Canada, approximately 120 km from the community of Salliq (Figure 1). The hydrophone was deployed at a depth of 142 m as part of an oceanographic mooring anchored to the sea floor. The deployment was conducted under the University of Manitoba’s Southampton Island Marine Ecosystem Project (Mundy, 2022). Acoustic data were recorded from June 5 to November 30, 2018, using a duty cycle of 5 minutes per hour. However, many files from October and November were corrupted due to equipment malfunction. The hydrophone was programmed with a sampling rate of 192 kHz with no set gain.
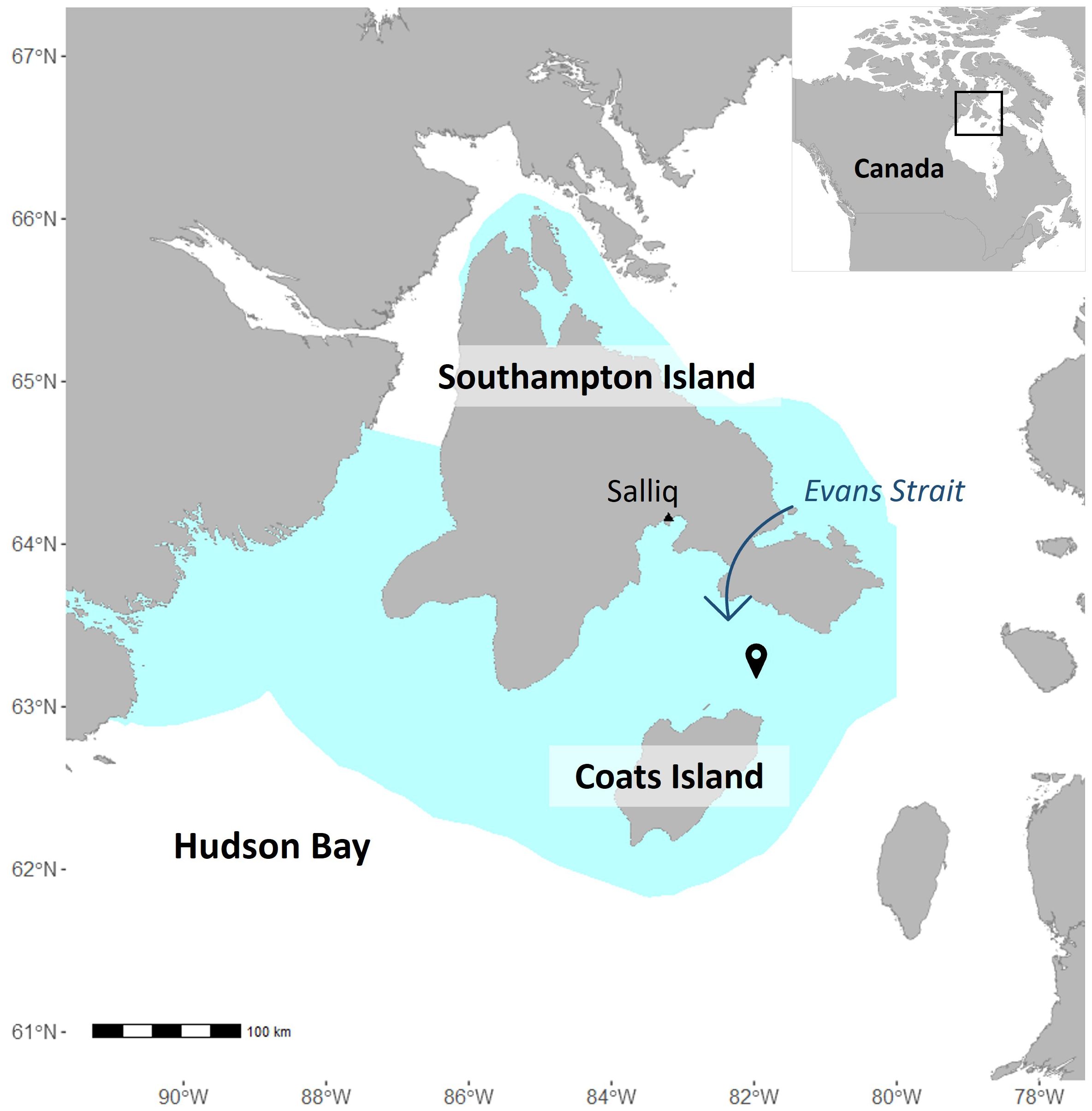
Figure 1. Map of the study area. The hydrophone (pin) was deployed in Evans Strait, Nunavut, Canada. The extent of the Southampton Island Area of Interest is represented in turquoise.
AIS data collection
To assess vessel traffic in the study area, AIS data and ship information were downloaded from the Arctic Ship Traffic Data (ASTD) database of the Protection of the Arctic Marine Environment (PAME)2. Data covering the Canadian Exclusive Economic Zone were downloaded for the period that goes from June to November 2018. The Level 2 dataset included vessel location and time, identification number, and type, classified using the Statcode 5 ship type coding system (PAME, 2024). Because pleasure craft and motorboats are not required to carry AIS transceivers, motorboat traffic could not be captured through this dataset.
Data analyses
Acoustic analysis
The hydrophone recorded 4,287 5-minute audio files, of which 28.8% were corrupted. A total of 3054 audio files were analyzed using the sound analysis software Raven Pro, version 1.6.5 (Raven Pro, 2024). Spectrograms were created for each file using a Hann window of 20,000 samples with 50% overlap, resulting in a frequency resolution of 9.6 Hz and a time resolution of 0.05 s. Visual analysis was conducted by scrolling through the files using a 30-second time window and a frequency range up to 1.4 kHz. Contrast and brightness were adjusted as needed. Species identification was based on comparisons with published information on walrus (Stirling et al., 1983; Sjare et al., 2003; Mouy et al., 2012) and beluga vocalizations (Sjare and Smith, 1986; Chmelnitsky and Ferguson, 2012; Garland et al., 2015; Booy et al., 2023). Although beluga clicks were excluded from the main acoustic analysis, their presence was investigated in each recording and contributed to validating the presence of belugas in combination with their whistles and pulsed calls. Additional reference sounds were sourced from online libraries such as the Discovery of Sound in the Sea (DOSITS)3 audio gallery and the Macaulay library of the Cornell Lab of Ornithology4. Expert consultation with specialists in acoustics of Arctic marine mammals further supported species identification and contributed to control for observer bias.
Vessel underwater noise was detected and analyzed using the same software and methodology applied to marine mammal sounds. Each vessel noise event was divided into three stages of noise exposure: before, during and after. The file immediately preceding the detection of vessel noise was labelled before; all the consecutive files containing vessel noise were labelled during; and the first file after the noise ceased, which did not include vessel noise, was labelled after. If marine mammal vocalizations were present in any of the three stages, the event was classified as vessel encounter. Species calls were counted for each vessel encounter and noise exposure stage. A single call was defined as one distinct vocalization, except in the case of walrus knocks, which typically occur in trains - each train was counted as one call, regardless of its duration or number of consecutive knocks.
Vessel noise was categorized as either ship or motorboat based on acoustic characteristics, such as main frequency, bandwidth, and tonal components (Richardson et al., 1995; Sorensen et al., 2010; Simard et al., 2016; Kuzin et al., 2022). Vessel type categorization was supported by a comparison of our recordings to online audio libraries dedicated to vessel underwater noise, such as Hear my ship5. Vessel noise classified as ship typically exhibited broadband frequencies ranging from around 50 Hz to 160 Hz. In contrast, motorboat noise was characterized by a narrow-band signal centered around 200 Hz (occasionally up to 400 Hz) with tonal harmonics at higher frequencies. Moreover, motorboat spectrograms often displayed rapid variations in frequency, indicative of changes in vessel speed or course direction (see, e.g., Reis et al., 2019).
AIS data analysis
AIS data were uploaded into ArcMap (Version 10.8.2) and cleaned to remove anomalies. AIS data points from June to November 2018 were overlaid with a polygon encompassing Southampton Island and nearby communities in Hudson Bay, referred to as the Vessel traffic Study Area (VSA). All points were then merged into a single shapefile. To map vessel movements, points were converted into tracks using the tool Points to Line and vessel identification numbers were used to generate separate tracks for each vessel. For vessels making multiple trips through the Southampton Island AOI, track lines were segmented into distinct trips based on the vessel’s entry and exit across the VSA boundaries. Statistical analyses of AIS data were conducted in R (R Core Team, 2022).
A comparison between vessel AIS tracks and underwater noise recordings was performed to validate vessel type during the acoustic analysis. A 10-km radius polygon was drawn around the hydrophone mooring location in ArcMap. Since the goal of the study was to detect walrus and beluga reactions to vessel noise, this radius was chosen based on the average underwater propagation range of walrus vocalizations (Sjare and Stirling, 1996; Sjare et al., 2003). This range represents an upper limit for most beluga vocalization (Simard et al., 2010; Vergara et al., 2021) and vessel underwater noise propagation (McKenna et al., 2012; Hermannsen et al., 2014; Jansen and De Jong, 2017). Each time a vessel crossed this 10-km area, the event was classified as a transit. For each vessel trip, the entry and exit times into the AOI were recorded, along with notes on whether the vessel passed through Evans Strait or made a stop in Salliq. The timing of vessel transits was then compared to the date and time of the acoustic detections of vessel underwater noise for validation.
Statistical analysis
Generalized linear mixed models (GLMMs) were run in RStudio (RStudio Team, 2024) employing the glmer.nb function from the lme4 package (Bates et al., 2015) to assess the effect of vessel encounters on call detection rates. Separate models were run for walrus and beluga encounters. Based on preliminary analyses, a negative binomial distribution was selected. The number of call detections was set as the response variable and modelled as a function of two predictor variables considered as fixed effects: vessel type (ship or motorboat) and noise exposure stage (before, during, or after). Vessel encounter events were included as random effects. To account for differences in encounter duration, the logarithm of the number of consecutive files containing vessel noise was included in the function as an offset. For each species, model optimization was conducted for both fixed and random structures. Model diagnostics were performed using the DHARMA package (Hartig, 2016), and model selection was based on the Akaike’s information criterion corrected for small sample size (AICc) (Burnham and Anderson, 2002).
Results
Walrus grunts and knocks, as well as beluga pulsed calls and whistles were detected during the study. Beluga were recorded in 8.1% and walrus in 5.9% of the total files. Vessel underwater noise was detected in 5.6% of the files. Beluga vocalizations were primarily recorded in June and November (Figure 2). Walrus vocal activity was low in June and increased in July, remaining relatively constant during the remainder of the monitoring period (Figure 2). Vessel noise was present throughout the entire study period, increasing throughout the summer and decreasing after October (Figure 2).
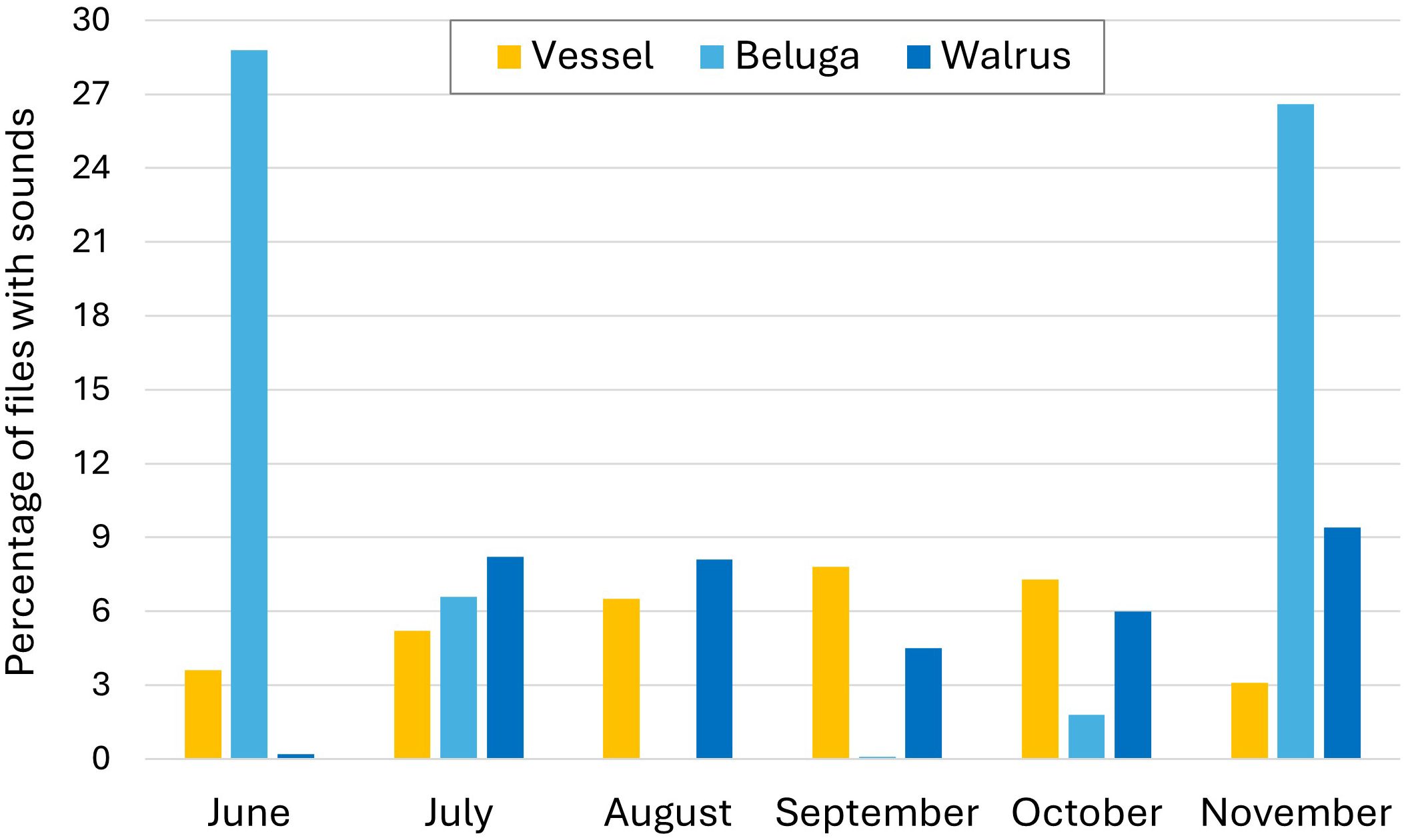
Figure 2. Monthly occurrence of vessel noise, and beluga and walrus sounds as recorded in Evans Strait, Nunavut, Canada, in 2018. The Y axis displays the percentage of recordings containing sounds over the total recorded files for each category.
AIS data analysis revealed that 23 vessels entered the Southampton Island AOI during the monitoring period (Table 1, Figure 3). Most were general cargo ships and chemical tankers; no bulk carriers were detected (Table 1). Seven vessels, primarily general cargo ships, stopped in Salliq. Of the vessels transiting Evans Strait, 70% did not stop in Salliq but continued to other Hudson Bay communities, including Qamani’tuaq (Baker Lake), Igluligaarjuk (Chesterfield Inlet), Kangiqtiniq (Rankin Inlet), Tikirarjuaq (Whale cove), and Naujaat (Repulse Bay). A total of 50 vessel transits were detected in Evans Strait during the study period, 34 of which passed within the 10-km area around the hydrophone (Table 1). Ship traffic peaked between July and October, with only one or two trips detected in June and November (Figure 4). General cargo ships transited Evans Strait throughout the monitoring period, while chemical tankers were detected only between July and October (Figure 4).
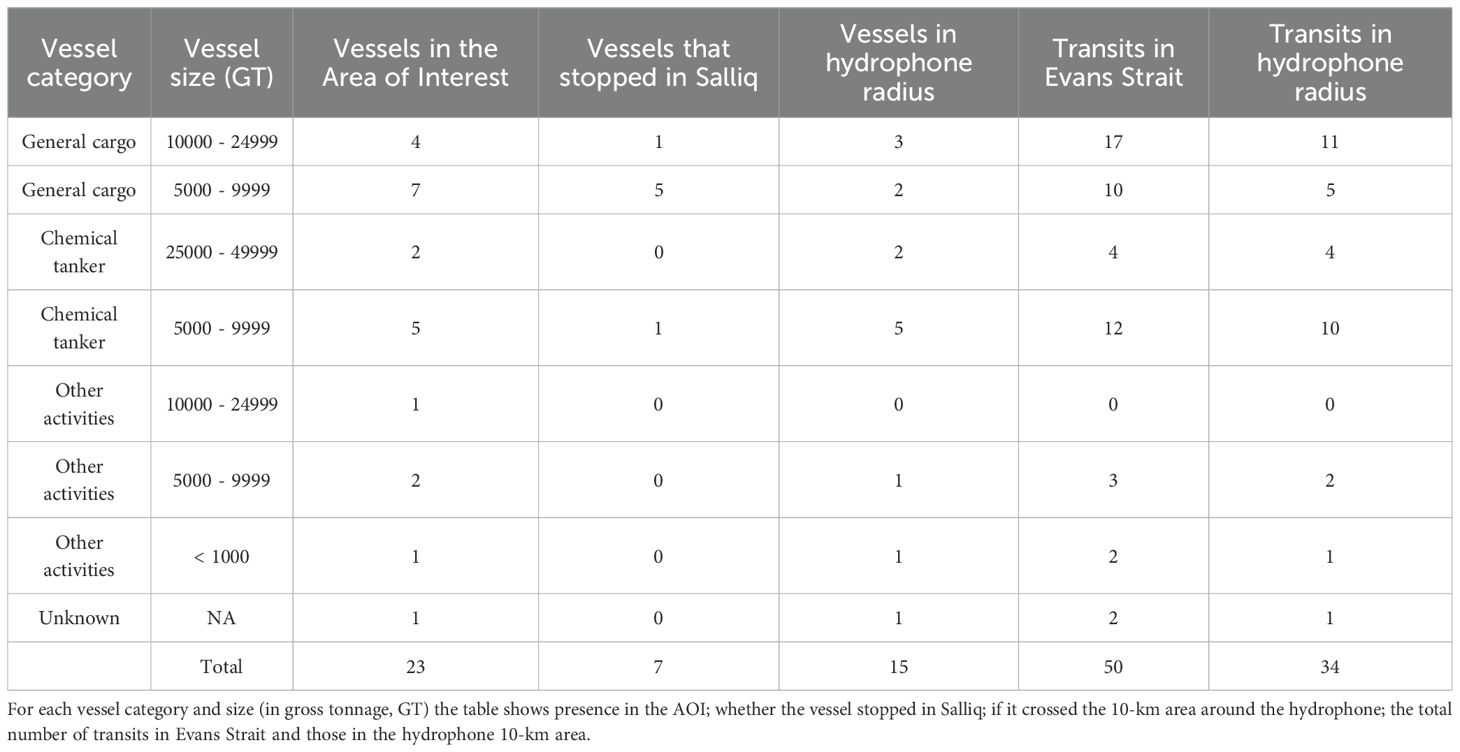
Table 1. Vessel traffic in the Southampton Island Area Of Interest (AOI) as derived from the AIS dataset from June to November 2018.
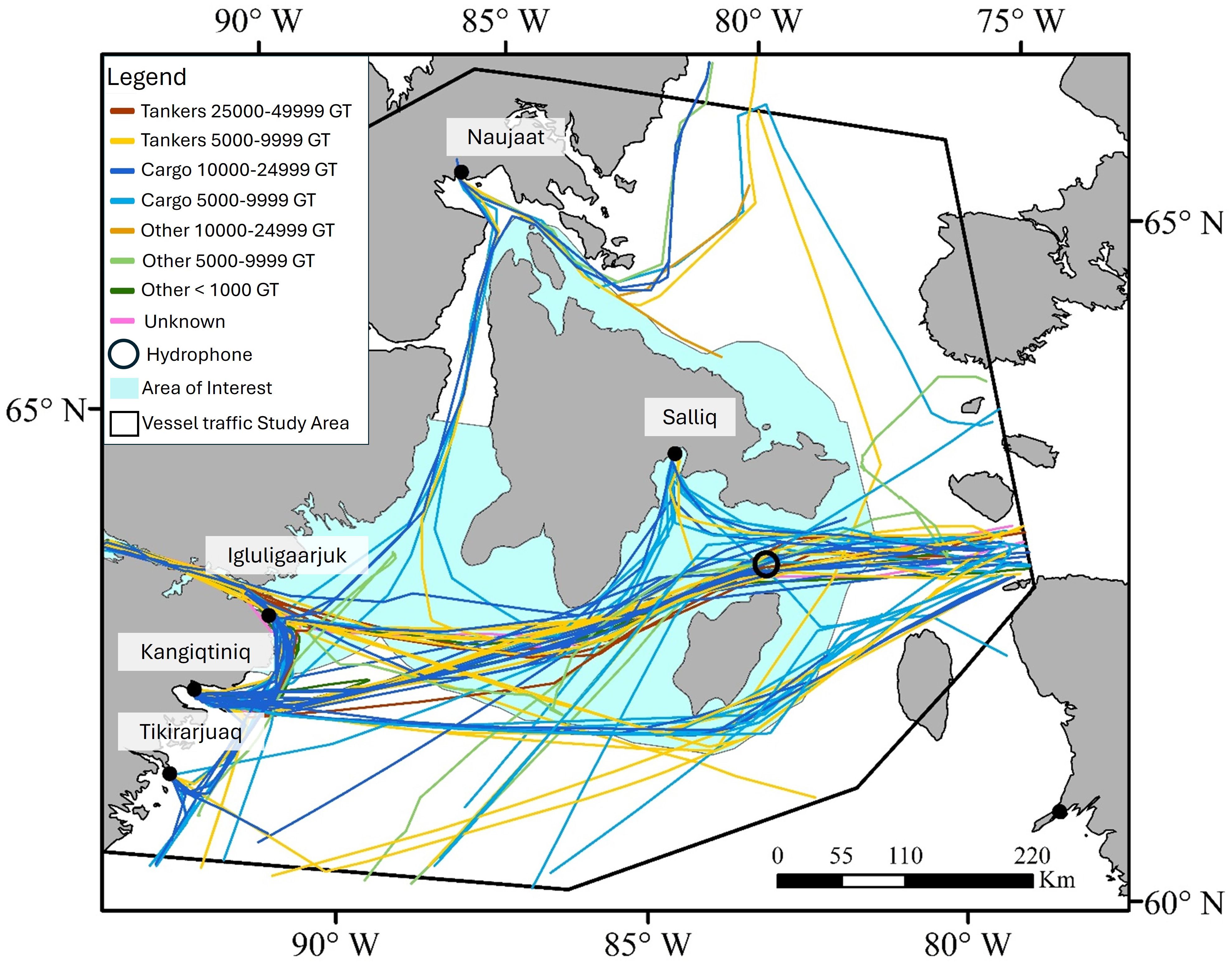
Figure 3. Vessel traffic map in the Southampton Island Area of Interest (AOI) from June to November 2018. Vessel category and size (in gross tonnage, GT) are indicated by different colors. Vessel type names have been shortened for convenience. The circle shows the 10-km radius around the hydrophone; the turquoise area represents the AOI extent; and the black line indicates the boundaries of the Vessel traffic Study Area.
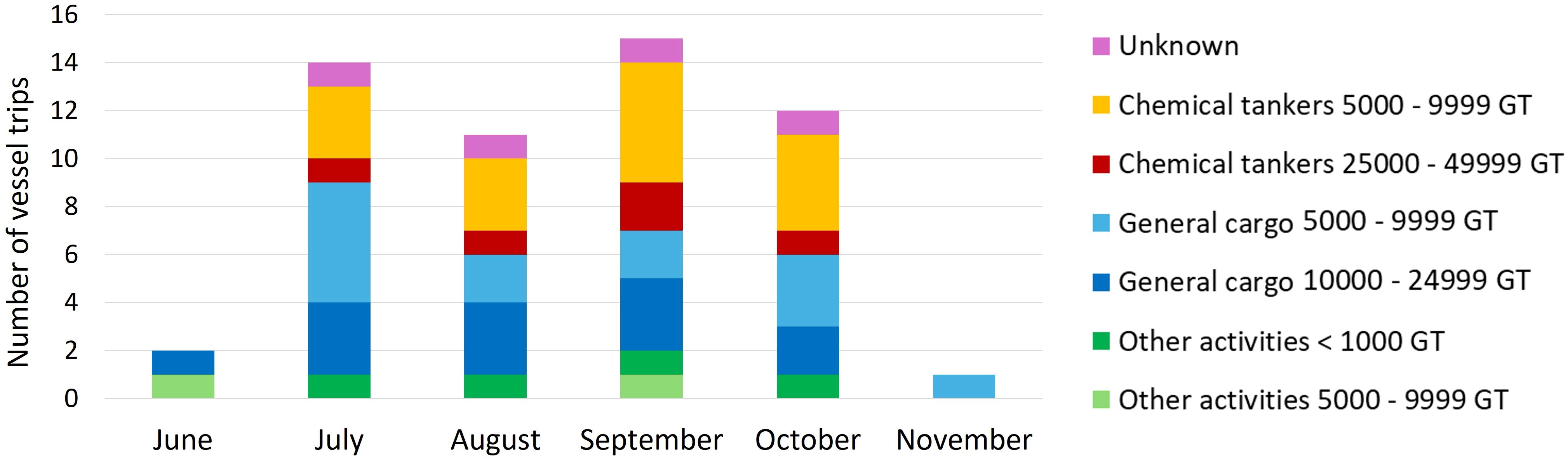
Figure 4. Monthly vessel trips in Evans Strait, Nunavut, Canada, as derived from AIS data for the period between June and November 2018.
Acoustic analysis detected 42 vessel transits during the entire monitoring period, of which 15 were classified as motorboats and 27 as ships based on their noise signature. When comparing AIS and PAM data, 15 of the 27 acoustically detected vessels classified as ships matched AIS vessel tracks that passed within the 10-km radius around the hydrophone. None of the vessels acoustically classified as motorboats corresponded with any AIS tracks. A total of 31 vessel encounters with walruses and 5 encounters with belugas were recorded (Table 2). No vessel encounters for either species were recorded in November and no beluga encounter with motorboats was detected for the entire monitoring period (Table 2).
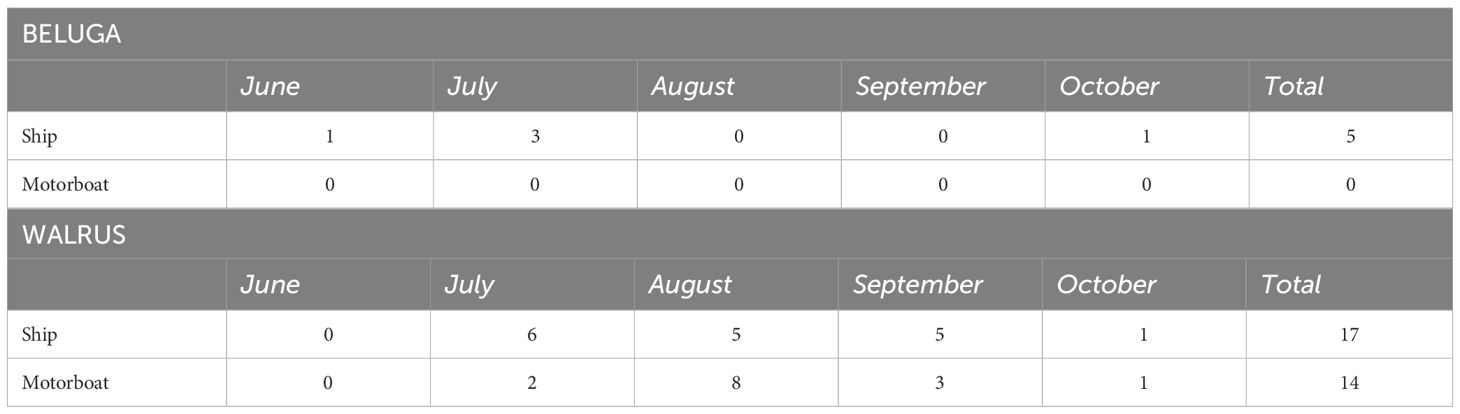
Table 2. Species encounters with vessels as derived from the acoustic recordings in Evans Strait, Nunavut, Canada, between June and November 2018.
Due to insufficient data, GLMMs could not be performed to assess the effect of vessel encounters on beluga call detection rates. As a result, modeling was limited to the walrus acoustic dataset. Among the models tested with different combinations of predictor variables, the model that included both noise exposure stage and vessel type, and the model with vessel type alone were equally supported as the best models based on AICc values (Table 3).
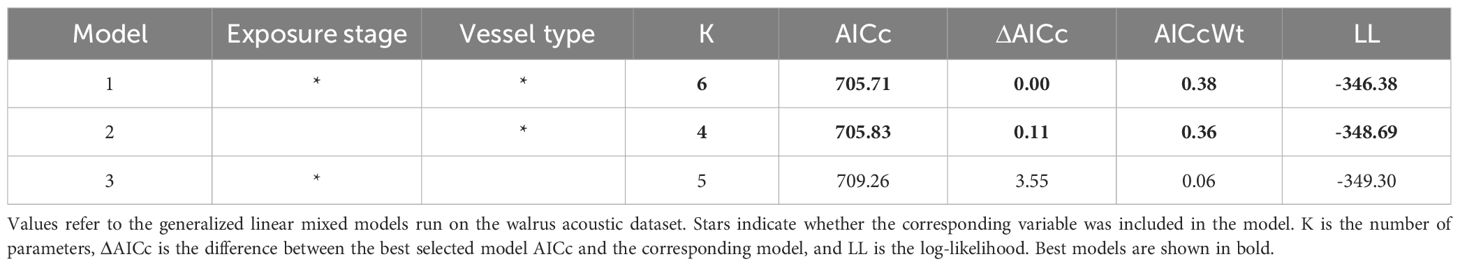
Table 3. Model comparison based on the Akaike’s information criterion corrected for small sample size (AICc) and corresponding weights (AICcWt).
Walrus vocalization rates decreased during the transit of both vessel types, with significantly lower estimates during ship encounters compared to motorboat encounters (p<0.05) (Table 4). Walrus vocalization rates were highest before vessel transits and decreased during and after the encounters (p<0.1) (Figure 5, Table 4). No statistically significant difference was found between vocalization rates recorded during and after vessel encounters (Figure 5, Table 4).
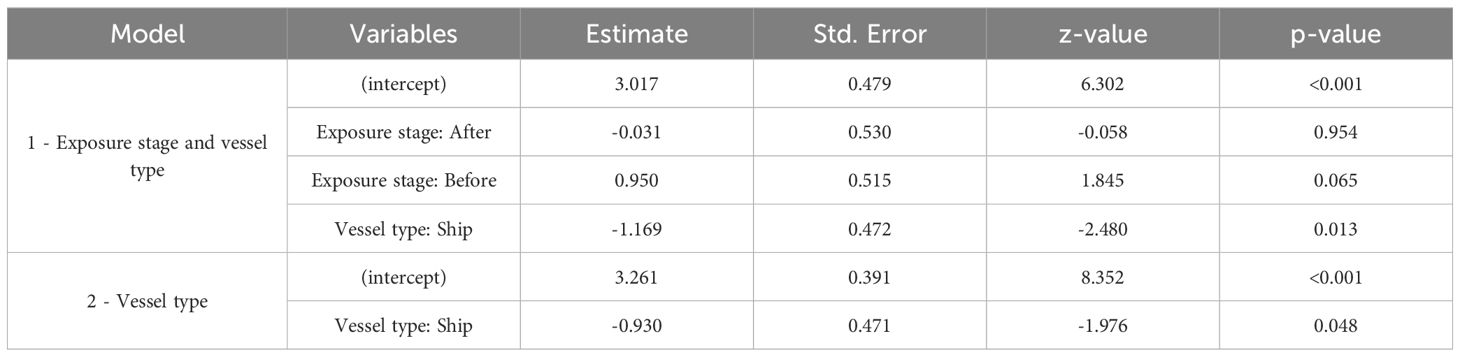
Table 4. Resulting parameters of the generalized linear mixed models of the effect of noise exposure stage and vessel type on walrus vocalization rates.
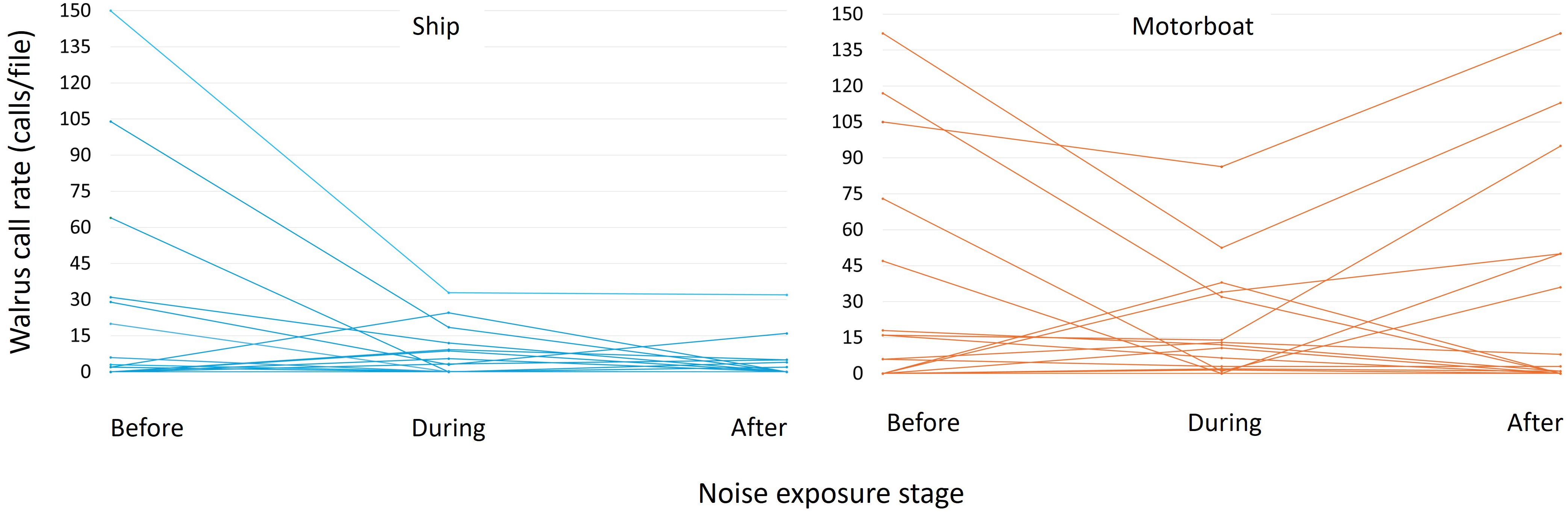
Figure 5. Walrus vocalization rates (calls per file) during ship (left) and motorboat (right) encounters in Evans Strait, Nunavut, Canada, from June to November 2018. The x-axis shows noise exposure stage.
Discussion
Our results indicate that vessel traffic disrupted walrus vocal behavior. A reduction in walrus vocalization rates was found during encounters with both types of vessels, with ships causing more changes than motorboats. Previous studies indicate that walruses are vulnerable to vessel traffic (Moore et al., 2012; Erbe et al., 2018; Hauser et al., 2018), and that vessel noise overlaps with their hearing range in both air and water (Kastelein et al., 2002; Reichmuth et al., 2020; Duarte et al., 2021). Walrus hearing sensitivity is centered at frequencies between 1–12 kHz (Kastelein et al., 2002), which allows them to hear the main frequencies of both ships and motorboats in this study.
Most research on walrus vocal behavior has focused on the Pacific walrus (Odobenus rosmarus divergens), with documentation of their reactions to vessels being limited to visual observations of behavioral changes. The few studies on Atlantic walrus disturbance from vessels consist mainly of visual monitoring at haul-out sites (Born et al., 1995; Øren et al., 2018; DFO, 2019; Higdon et al., 2022). Reported reactions range from signs of short-term disturbance, such as head-raising and diving, to stampedes, shifts in feeding areas, and long-term abandonment of haul-out sites (Salter, 1979; Fay et al., 1984; Mansfield and St. Aubin, 1991; Born et al., 1995; COSEWIC, 2017; Higdon et al., 2022). To our knowledge, this is the first study investigating walrus acoustic responses to vessel transits.
Walrus populations subject to hunting are particularly susceptible to motorboat approaches (Malme et al., 1989; Higdon and Stewart, 2018; Øren et al., 2018). The community of Salliq relies on walrus for both subsistence and income (COSEWIC, 2017; Carter et al., 2019; Loewen et al., 2020), with motorboats commonly used for subsistence harvesting and sport hunting (COSEWIC, 2017; Minister of Justice, 2018). In this study, however, walrus vocalization rates were found to change significantly more during ship transits compared to motorboats, suggesting that motorboat traffic may have a lesser impact on walrus vocal behavior compared to ships. This difference may depend on the recurrence of exposure to the two vessel types, with motorboats being a long-established means of transportation in the study area, while consistent ship traffic is a more recent phenomenon (Dawson et al., 2018; Carter et al., 2019; Dawson et al., 2021). Repeated exposure to a stimulus can cause a decrease in the amplitude of marine mammal responses, a process known as habituation or acclimation (Groves and Thompson, 1970; Romero, 2004; Wright et al., 2007; Götz and Janik, 2011). The weaker response recorded for walruses during motorboat transits compared to ships may therefore reflect a process of associative learning from repeated events, as reported in other marine mammals (Born et al., 1995; Erbe et al., 2018; Harris et al., 2018). However, reduced behavioral reactions does not necessarily imply reduced stress and physiological responses (Wright et al., 2007), hence weaker responses should be carefully interpreted when developing disturbance mitigation measures.
This study could not determine whether the decrease in walrus vocalization rates during vessel encounters resulted from abandonment of the area or from a tendency to remain silent in those circumstances. Evans Strait is used by walruses for mating and foraging, while the southern coast of Southampton Island and the northern shore of Coats Island are important walrus haul-out sites (Carter et al., 2019; Loewen et al., 2020; Higdon et al., 2022; Coppolaro et al., 2024). Observed deviations from walrus undisturbed vocal behavior may be indicative of disturbance already occurring and should therefore be further investigated, especially in trafficked walrus hotspots such as the study area.
Future acoustic monitoring efforts could benefit from optimizing the recording duty cycle to align with the objectives of the survey and the acoustic behavior of the focal species. Specifically, longer listening durations or duty cycles with shorter intervals may enhance the detection of walrus and beluga vocalizations, improving assessments of acoustic presence, diel acoustic patterns, and call rate estimates (Thomisch et al., 2015). Such adjustments may also facilitate more accurate evaluations of marine mammal responses to anthropogenic underwater noise by better capturing potential changes in their vocal behavior. This would be particularly valuable in combination with effective assessments of noise exposure through measurements such as sound pressure levels. Furthermore, integrating PAM with AIS data and satellite telemetry from tagged animals would provide a more comprehensive understanding of vessel traffic dynamics and their impacts on marine mammals within the AOI (see, for example, Martin et al., 2024).
Belugas are notoriously a vocal species (Au et al., 1985; Sjare and Smith, 1986; Chmelnitsky and Ferguson, 2012; Panova et al., 2019). In this study, beluga vocalizations were detected more frequently than walrus sounds overall; however, detections were primarily concentrated during the months corresponding to beluga migration through the area (Carter et al., 2019; Loewen et al., 2020). In contrast, walrus vocalizations were spread across the entire monitoring period, reflecting the different habitat use of the two species. While the walrus is a resident species of the Southampton Island AOI, belugas mainly transit the area in early summer and autumn (Carter et al., 2019; Loewen et al., 2020; Coppolaro et al., 2024).
During this study, beluga vocalizations were never recorded during motorboat encounters and rarely during ship transits. Although most beluga vocalizations were recorded outside the main shipping season, AIS data and recordings of ship and motorboat noise confirmed temporal overlap between vessel transits and overall beluga acoustic activity in the study area. We hypothesize that the complete absence of beluga vocalizations during motorboat encounters may indicate an avoidance response, either physical or vocal. Previous studies have documented a decrease in beluga vocalizations in response to motorboat traffic (Lesage et al., 1999; Karlsen et al., 2002), as well as vocal interruptions in areas frequented by orcas (Orcinus orca) (Castellote et al., 2022). During motorboat transits, belugas may have reduced vocalizations to avoid detection by what could be perceived as a potential predator. Belugas were also known to leave an area when hunted from motorboats (Caron and Smith, 1990; Mymrin et al., 1999; Malcolm and Penner, 2011). Since belugas are subject to hunting in the AOI (Hoover et al., 2013; Carter et al., 2019; Loewen et al., 2020), the absence of beluga vocalizations during motorboat encounters in this study may also reflect their physical displacement. However, the data collected in this study were insufficient to assess beluga responses to vessel encounters. To investigate the aforementioned hypotheses, future research should include high-frequency analyses of beluga vocalizations—particularly ultrasonic burst pulses (Vergara et al., 2025)—which were beyond the scope of this work.
In this study, the combined use of PAM and AIS data allowed for the inclusion of both ship and motorboat information in the vessel traffic analysis. Including motorboats was deemed important for the purpose of this study due to both their common use and the growing presence of pleasure craft in the area (Dawson et al., 2018; Carter et al., 2019). Each monitoring methodology has its advantages and limitations. PAM allows for the detection of vessels without AIS instrumentation on board, which generally consists of motorboats, and of ships with AIS systems turned off (Kline et al., 2020). The acoustic analysis identified more vessel transits than those extracted from the AIS dataset, primarily due to the inclusion of motorboats exclusively in the PAM dataset. For ships, the detection discrepancy between the AIS and PAM datasets may be caused by the use of the exploratory 10-km hydrophone range. The selected range may have resulted in an underestimation of the number of AIS-tracked vessels that passed sufficiently close to the hydrophone to be acoustically detected. Also, AIS signal irregularities, equipment turned off, and the use of Class B units on some of the vessels may have contributed to inaccurate ship positions relative to the hydrophone (Corsi et al., 2023). Future studies would benefit from in situ measurements of the hydrophone sensitivity, local underwater sound propagation models, and from comparison tests between AIS and acoustic datasets using different distances for the hydrophone range (Aulanier et al., 2016). These would enable estimates of vessel and marine mammal distances, as well as a more accurate determination of the hydrophone detection radius to use for comparison with AIS data. Moreover, the use of multi-channel hydrophones would increase the number of synchronized listening points and enable localization of vocalizing individual positions in relation to vessels.
When applied to vessel traffic studies and compared to the use of AIS data, PAM limitations include a higher effort in extracting vessel position and speed, which often requires arrays or multichannel hydrophones, and the lack of certain navigation information such as vessel type (Zhu et al., 2018; Tesei et al., 2020). Manual analysis of recordings is time consuming, hence automated techniques are critical to significantly accelerate acoustic data analyses for vessel traffic studies (Reis et al., 2019; Vieira et al., 2020). The comparison between PAM and AIS data in this study also served to validate the classification method used to distinguish ships from motorboats based on their underwater acoustic signature. Overall, combining these two techniques demonstrated how passive acoustics can integrate standard marine traffic monitoring.
Vessel underwater noise was recorded throughout the entire monitoring period. Ship presence in early June and November indicates that local ice conditions allowed for marine traffic in the area, reflecting the prolonged ice-free season in the Arctic (Parkinson, 2014; Crawford et al., 2021). Interestingly, bulk carriers were not detected in the AIS data, despite the rapid growth of bulk carrier traffic in the Arctic (PAME, 2024). Most ships crossing Evans Strait in 2018 did not service the community of Salliq but transited the area to reach other Hudson Bay communities and the Baker Lake and Rankin Inlet mines. To reduce the impact of vessel traffic on marine mammals, vessels not servicing Salliq could be re-routed south of Coats Island, hence avoiding crossing biologically important areas. Agnico Eagle Mines Ltd. (2020) adopted such a rerouting plan for their sealift operations to the aforementioned mines, following the low-impact shipping corridors plan presented for the area by Dawson et al. (2018; 2020).
Disruption of individual activities can lead to long-term impacts on fitness and populations (Pirotta et al., 2018; 2022). As the ongoing reduction in sea ice is predicted to further boost Arctic marine traffic (Dawson et al., 2018; Stevenson et al., 2019; Rodríguez et al., 2024), additional studies are needed to better understand marine mammal reactions to vessel traffic in the Southampton Island AOI. For a marine protected area to be effective in supporting species health and conservation, measures must be implemented to minimize species disturbance (Williams et al., 2015). Vessel slow-downs and rerouting to avoid spatial and temporal overlap with key areas, especially during ecologically important periods, have been shown to reduce the impact of underwater noise pollution on several species (Pine et al., 2018; Williams et al., 2019; Findlay et al., 2023). Therefore, vessel traffic regulations and underwater noise mitigation strategies are deemed essential to support the establishment of a MPA around Southampton Island.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
Ethical approval was not required for the study involving animals in accordance with the local legislation and institutional requirements because of the use of non-invasive passive acoustic monitoring techniques.
Author contributions
VC: Data curation, Software, Investigation, Project administration, Conceptualization, Resources, Visualization, Writing – original draft, Validation, Writing – review & editing, Methodology, Formal analysis. EA: Data curation, Investigation, Methodology, Validation, Writing – review & editing, Formal analysis, Software, Visualization. LL: Writing – review & editing. MM: Supervision, Investigation, Conceptualization, Resources, Methodology, Writing – review & editing, Project administration.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This research was partially funded by the Marine Environmental Observation, Prediction and Response Network of Centres of Excellence (MEOPAR-NCE), Polar Knowledge Canada, and a National Science and Engineering Research Council of Canada (NSERC) ship time grant for the Southampton Island Marine Ecosystem Project (SIMEP), a NSERC-matched grant for the 2017-2018 Belmont Forum and BiodivERsA joint call for research proposals under the BiodivScen ERA-Net CO-FUND programme to the project ACCES: De-icing of Arctic Coasts: Critical or new opportunities for marine biodiversity and Ecosystem Services. Furthermore, this study was supported in part by the Marine Environmental Quality (MEQ) program of Fisheries and Oceans Canada (DFO) Winnipeg, and by the Churchill Marine Observatory (CMO), which was funded by the Canada Foundation for Innovation and other partners, including the Arctic Research Foundation (ARF). This work represents a contribution to the Canada Excellence Research Chair (CERC) unit at the University of Manitoba.
Acknowledgments
The authors would like to thank the Aiviit Hunters and Trappers Organization and the Hamlet of Salliq for the support received for this project (Nunavut Research License #0301518N-M). They thank CJ Mundy (University of Manitoba) for his leadership in the Southampton Island Marine Ecosystem Project. The authors are grateful for the help received with the moorings and instrumentation by Sergei Kirillov and Keesha Petersen (University of Manitoba) and the crew and scientists of the RV William Kennedy. Thanks to Jody Willis (Fisheries and Oceans Canada) for her contribution to this project and to CJ Mundy (University of Manitoba) for reviewing a draft of this work.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Footnotes
- ^ https://turbulentresearch.com/tr-orca
- ^ https://pame.is/ourwork/arctic-shipping/current-shipping-projects/astd/
- ^ https://dosits.org/galleries/audio-gallery/
- ^ https://www.macaulaylibrary.org/
- ^ https://hearmyship.fer.hr/
References
Agnico Eagle Mines Ltd. (2020). Sealift Season 2020 – Agnico Eagle Nunavut Operations. Meadowbank / Whale Tail, Nunavut: Agnico Eagle Mines Ltd.
Andrews J., Babb D., and Barber D. G. (2018). Climate change and sea ice: Shipping in Hudson Bay, Hudson Strait, and Foxe Basin, (1980–2016). Elem. Sci. Anth. 6, 13–26. doi: 10.1525/elementa.281
Au W. W., Penner R. H., Carder D. A., and Scronce B. (1985). Demonstration of adaptation in beluga whale echolocation signals. J. Acoust. Soc Am. 77, 726–730. doi: 10.1121/1.392341
Aulanier F., Simard Y., Roy N., Bandet M., and Gervaise C. (2016). Groundtruthed probabilistic shipping noise modeling and mapping: Application to blue whale habitat in the Gulf of St. Lawrence. Proc. Mtgs. Acoust. 27, 070006. doi: 10.1121/2.0000258
Ausen E. L., Marcoux M., Chan W. S., and Barber D. G. (2022). Beluga (Delphinapterus leucas) response to personal watercraft and motorized whale watching vessels in the Churchill River estuary. Front. Mar. Sci. 9. doi: 10.3389/fmars.2022.837425
Bates D., Mächler M., Bolker B., and Walker S. (2015). Fitting linear mixed-effects models using lme4. Journal of Statistical Software, 67(1), 1–48. doi: 10.18637/jss.v067.i01
Booy K. V., Vergara V., Loseto L., Mikus M. A., and Marcoux M. (2023). Geographic variation in simple contact calls of Canadian beluga whales (Delphinapterus leucas). Marine Mammal Science, 1–15. doi: 10.1111/mms.13095
Born E. W., Gjertz I., and Reeves R. R. (1995). Population assessment of Atlantic walrus (Odobenus rosmarus rosmarus L.). Norsk Polarinstitutt.
Burnham K. P. and Anderson D. R. (2002). Model selection and multimodel inference: A practical information-theoretic approach (2nd ed.) (New York, NY: Springer).
Caron L. M. J. and Smith T. G. (1990). Philopatry and site tenacity of belugas, Delphinapterus leucas, hunted by the Inuit at the Nastapoka estuary, eastern Hudson Bay. Can. Bull. Aquat. Sci. 69), 69–79.
Carter N. A., Dawson J., and Weber M. (2019). Arctic corridors and northern voices: Governing marine transportation in the Canadian Arctic (Coral Harbour, Nunavut community report). Available online at: http://hdl.handle.net/10393/38505 (Accessed May 14, 2020).
Castellote M., Mooney A., Andrews R., Deruiter S., Lee W., Ferguson M., et al. (2021). Beluga whale (Delphinapterus leucas) acoustic foraging behavior and applications for long term monitoring. PloS One 16, e0260485. doi: 10.1371/journal.pone.0260485
Castellote M., Stafford K., Small R., Whiting A., and Frost K. J. (2022). Beluga (D. leucas), harbor porpoise (P. phocoena), and killer whale (O. orca) acoustic presence in Kotzebue Sound, Alaska: Silence speaks volumes. Front. Mar. Sci. 3. doi: 10.3389/frsen.2022.940247
Chmelnitsky E. G. and Ferguson S. H. (2012). Beluga whale, Delphinapterus leucas, vocalizations from the Churchill River, Manitoba, Canada. J. Acoust. Soc Am. 131, 4821–4835. doi: 10.1121/1.4707501
Clark C. W., Ellison W. T., Southall B. L., Hatch L., van Parijs S. M., Frankel A., et al. (2009). Acoustic masking in marine ecosystems: Intuitions, analysis, and implication. Mar. Ecol. Prog. Ser. 395, 201–222. doi: 10.3354/meps08402
Coppolaro V. L. M., Gupta K., Loseto L., Mundy C. J., and Marcoux M. (2024). Seasonal and diurnal marine mammal presence in the proposed marine protected area of Southampton Island, Hudson Bay, Nunavut as revealed by passive acoustic monitoring. Arctic Sci. 10, 520–544. doi: 10.1139/as-2023-0076
Corsi M., Francalanci C., Giacomazzi P., Ravanelli P., and Tilia S. (2023). Extracting standard naval routes from AIS data: Insights from an in-depth analysis of the Arctic region. ITAIS 2023 Proc. 19.
COSEWIC (2017). COSEWIC assessment and status report on the Atlantic walrus Odobenus rosmarus rosmarus, High Arctic population, Central-Low Arctic population and Nova Scotia–Newfoundland–Gulf of St. Lawrence population in Canada (Committee on the Status of Endangered Wildlife in Canada). Available online at: https://www.Canada.ca/en/environment-climate-change/services/species-risk-public-registry/cosewic-assessments-status-reports/atlantic-walrus-2017.html (Accessed February 12, 2020).
Crawford A., Stroeve J., Smith A., and Jahn A. (2021). Arctic open-water periods are projected to lengthen dramatically by 2100. Commun. Earth Environ. 2, 109. doi: 10.1038/s43247-021-00183-x
Dawson J., Carter N., van Luijk ,. N., Parker C., Weber M., Cook A., et al. (2020). Infusing Inuit and local knowledge into the low impact shipping corridors: an adaptation to increased shipping activity and climate change in Arctic Canada. Environ. Sci. Policy 105, 19–36. doi: 10.1016/j.envsci.2019.11.013
Dawson J., Carter N., Van Luijk ,. N., Cook A., Weber M., Orawiec A., et al. (2021). Tourism vessels and low impact shipping corridors in Arctic Canada: trends, risks, community perspectives and management strategies (Ottawa, ON: Universitéd’Ottawa/University of Ottawa). doi: 10.20381/D3DD-YK49
Dawson J., Pizzolato L., Howell S. E. L., Copland L., and Johnston M. (2018). Temporal and spatial patterns of ship traffic in the Canadian Arctic from 1990 to 2015. Arctic 71, 15–26. doi: 10.14430/arctic4698
DFO (2019). Mitigation buffer zones for Atlantic walrus (Odobenus rosmarus rosmarus) in the Nunavut Settlement Area, (2018/055; DFO Can. Sci. Advis. Sec. Sci. Resp.
Duarte C. M., Chapuis L., Collin S. P., Costa D.P., Devassy R.P., Eguiluz V.M., et al. (2021). The soundscape of the Anthropocene ocean. Science 371, eaba4658. doi: 10.1126/science.aba4658
Erbe C., Dunlop R., and Dolman S. (2018). “Effects of noise on marine mammals,” in Effects of Anthropogenic Noise on Animals, vol. 66 . Eds. Slabbekoorn H., Dooling R. J., Popper A. N., and Fay R. R. (New York, NY: Springer New York). doi: 10.1007/978-1-4939-8574-6
Fay F. H., Kelly B. P., Gehnrich P. H., et al. (1984). Modern populations, migrations, demography, trophics, and historical status of the Pacific walrus. Available online at: https://www.govinfo.gov/app/details/GOVPUB-I-e4afaac7a5139b5907a945b920ebc663 (Accessed June 23, 2023).
Findlay C. R., Rojano-Doñate L., Tougaard J., Johnson M.P., and Madsen P.T. (2023). Small reductions in cargo vessel speed substantially reduce noise impacts to marine mammals. Sci. Adv. 9. doi: 10.1126/sciadv.adf2987
Garland E. C., Berchok C. L., and Castellote M. (2015). Beluga whale (Delphinapterus leucas) vocalizations and call classification from the eastern Beaufort Sea population. Journal of the Acoustical Society of America, 137, 3054. doi: 10.1121/1.4919338
Götz T. and Janik V. M. (2011). Repeated elicitation of the acoustic startle reflex leads to sensitisation in subsequent avoidance behaviour and induces fear conditioning. BMC Neurosci. 12, 30. doi: 10.1186/1471-2202-12-30
Groves P. M. and Thompson R. F. (1970). Habituation: a dual-process theory. Psychol. Rev. 77, 419–450. doi: 10.1037/h0029810
Halliday W. D., Pine M. K., and Insley S. J. (2020). Underwater noise and Arctic marine mammals: review and policy recommendations. Environ. Rev. 28, 438–448. doi: 10.1139/er-2019-0033
Harris C. M., Thomas L., Falcone E. A., Hildebrand J., Houser D., Kvadsheim P.H., et al. (2018). Marine mammals and sonar: dose-response studies, the risk-disturbance hypothesis and the role of exposure context. J. Appl. Ecol. 55, 396–404. doi: 10.1111/1365-2664.12955
Hartig F. (2016). DHARMa: residual diagnostics for hierarchical (multilevel/mixed) regression models. Available online at: https://cran.r-project.org/web/packages/DHARMa/vignettes/DHARMa.html (Accessed June 14, 2023).
Hauser D. D. W., Laidre K. L., and Stern H. L. (2018). Vulnerability of Arctic marine mammals to vessel traffic in the increasingly ice-free Northwest Passage and Northern Sea Route. Proc. Natl. Acad. Sci. U.S.A. 115, 7617–7622. doi: 10.1073/pnas.1803543115
Heenehan H., Stanistreet J. E., Corkeron P. J., Bouveret L., and Chalifour J. (2019). Caribbean Sea soundscapes: monitoring humpback whales, biological sounds, geological events, and anthropogenic impacts of vessel noise. Front. Mar. Sci. 6. doi: 10.3389/fmars.2019.00347
Hermannsen L., Beedholm K., Tougaard J., and Madsen P. T. (2014). High frequency components of ship noise in shallow water with a discussion of implications for harbor porpoises (Phocoena phocoena). J. Acoust. Soc Am. 136, 1640–1653. doi: 10.1121/1.4893908
Higdon J. W. and Stewart D. B. (2018). State of circumpolar walrus (Odobenus rosmarus) populations. Prepared by Higdon Wildlife Consulting and Arctic Biological Consultants for the WWF Arctic Programme.
Higdon J. W., Stewart D. B., and Stewart R. E. A. (2022). Atlantic walrus (Odobenus rosmarus rosmarus) in northern Hudson Bay—Status, research needs, and monitoring opportunities, prepared for Oceans North under contract to Higdon Wildlife Consulting (Jeff W. Higdon) Winnipeg, Manitoba, Canada.
Hoover C., Bailey M., Higdon J., Ferguson S. H., and Sumaila R. (2013). Estimating the economic value of narwhal and beluga hunts in Hudson Bay, Nunavut. Arctic, 66(1), 1–16. doi: 10.14430/arctic4261
Jansen E. and De Jong C. (2017). Experimental assessment of underwater acoustic source levels of different ship types. IEEE J. Oceanic Eng. 42, 439–448. doi: 10.1109/JOE.2016.2644123
Karlsen J. D., Bisther A., Lydersen C., Haug T., and Kovacs K. M. (2002). Summer vocalisations of adult male white whales (Delphinapterus leucas) in Svalbard, Norway. Polar Biol. 25, 808–817. doi: 10.1007/s00300-002-0415-6
Kastelein R. A., Mosterd P., Van Santen ,. B., Hagedoorn M., and de Haan D. (2002). Underwater audiogram of a Pacific walrus (Odobenus rosmarus divergens) measured with narrow-band frequency-modulated signals. J. Acoust. Soc Am. 112, 2173–2182. doi: 10.1121/1.1508783
Kline L. R., DeAngelis A. I., McBride C., Rodgers G.G., Rowell T.J., Smith J., et al. (2020). Sleuthing with sound: understanding vessel activity in marine protected areas using passive acoustic monitoring. Mar. Policy 120, 104138. doi: 10.1016/j.marpol.2020.104138
Kuzin D., Statsenko L., and Smirnova M. (2022). Automated sea vehicle classification system based on neural network. 2022 Int. Conf. Ocean Stud. (ICOS), 87–90. doi: 10.1109/ICOS55803.2022.10033384
Lesage V., Barrette C., Kingsley M. C. S., and Sjare B. (1999). The effect of vessel noise on the vocal behavior of belugas in the St. Lawrence River estuary, Canada. Mar. Mamm. Sci. 15, 65–84. doi: 10.1111/j.1748-7692.1999.tb00782.x
Loewen T., Hornby C., Johnson M., Chambers C., Dawson K., MacDonell D., et al. (2020). Ecological and biophysical overview of the Southampton Island ecologically and biologically significant area in support of the identification of an Area of Interest (DFO Can. Sci. Advis. Sec. Res. Doc.
Malakoff D. (2010). A push for quieter ships. Science 328, 1502–1503. doi: 10.1126/science.328.5985.1502
Malcolm C. D. and Penner H. C. (2011). “Behavior of belugas in the presence of whale-watching vessels in Churchill, Manitoba and recommendations for local beluga-watching activities,” in Polar Tourism: Human, Environmental and Governance Dimensions Putnam Valley, NY: Cognizant Communication Corporation, 54–79.
Malme C. I., Miles P. R., Miller G. W., Richardson W. J., Roseneau D. G., Thomson D. H., et al. (1989). Analysis and ranking of the acoustic disturbance potential of petroleum industry activities and other sources of noise in the environment of marine mammals in Alaska (BBN Report No. 6945; OCS Study MMS 89 0006). BBN Systems & Technologies Corporation for the U.S. Minerals Management Service.
Mansfield A. W. and St. Aubin D. J. (1991). Distribution and abundance of Arctic walrus in the Southampton Island region. Can. Field Nat. 105, 95–100. doi: 10.5962/p.357954
Martin M. J., Halliday W. D., Ferguson S. H., Young B. G., Charish R., Dawson J., et al. (2024). Exposure of satellite tagged bowhead whales (Balaena mysticetus) to transiting vessels in the Eastern Canadian Arctic. Mar. Mamm. Sci. 40, e13125. doi: 10.1111/mms.13125
McKenna M. F., Ross D., Wiggins S. M., and Hildebrand J. A. (2012). Underwater radiated noise from modern commercial ships. J. Acoust. Soc Am. 131, 92–103. doi: 10.1121/1.3664100
Minister of Justice (2018). Consolidated federal laws of Canada, Marine Mammal Regulations SOR/93-56 (Justice Laws Website). Available online at: https://laws-lois.justice.gc.ca/eng/regulations/sor-93-56/page-2.html (Accessed May 20, 2023).
Moore S. E., Reeves R. R., Southall B. L., Ragen T. J., Suydam R. S., and Clark C. W. (2012). A new framework for assessing the effects of anthropogenic sound on marine mammals in a rapidly changing Arctic. BioScience 62, 289–295. doi: 10.1525/bio.2012.62.3.10
Mouy X., Hannay D., Zykov M., and Martin B. (2012). Tracking of Pacific walruses in the Chukchi Sea using a single hydrophone. Journal of the Acoustical Society of America, 131, 1349–1358. doi: 10.1121/1.3675008
Mymrin N. I, The Communities of Novoe Chaplino, Sireniki, Uelen, and Yanrakinnot, and Huntington H. P. (1999). Traditional knowledge of the ecology of beluga whales (Delphinapterus leucas) in the Northern Bering Sea, Chukotka, Russia. Arctic, 52(1), 62–70. doi: 10.14430/arctic910
Øren K., Kovacs K. M., Yoccoz N. G., and Lydersen C. (2018). Assessing site-use and sources of disturbance at walrus haul-outs using monitoring cameras. Polar Biol. 41, 1737–1750. doi: 10.1007/s00300-018-2313-6
PAME (2024). The increase in Arctic shipping: 2013–2023 (Arctic Shipping Status Report (ASSR) 1) (Protection of the Arctic Marine Environment (PAME). Available online at: https://oaarchive.arctic-council.org/bitstreams/b045fa91-01ab-425b-a30f-ba41f047b092/download (Accessed February 20, 2025).
Panova E., Agafonov A., Belikov R., and Melnikova F. (2019). Characteristics and microgeographic variation of whistles from the vocal repertoire of beluga whales (Delphinapterus leucas) from the White Sea. J. Acoustical Soc. America 146, 681–692. doi: 10.1121/1.5119249
Parkinson C. L. (2014). Spatially mapped reductions in the length of the Arctic sea ice season. Geophysical Res. Lett. 41, 4316–4322. doi: 10.1002/2014GL060434
Pine M. K., Hannay D. E., Insley S. J., Halliday W.D., and Juanes F. (2018). Assessing vessel slowdown for reducing auditory masking for marine mammals and fish of the western Canadian Arctic. Mar. pollut. Bull. 135, 290–302. doi: 10.1016/j.marpolbul.2018.07.031
Pirotta E., Booth C. G., Calambokidis J., Costa D.P., Fahlbusch J.A., Friedlaender A.S., et al. (2022). From individual responses to population effects: Integrating a decade of multidisciplinary research on blue whales and sonar. Anim. Conserv. 25, 796–810. doi: 10.1111/acv.12785
Pirotta E., Booth C. G., Costa D. P., Fleishman E., Kraus S.D., Lusseau D., et al. (2018). Understanding the population consequences of disturbance. Ecol. Evol. 8, 9934–9946. doi: 10.1002/ece3.4458
Pizzolato L., Howell S. E. L., Dawson J., Laliberté F., and Copland L. (2016). The influence of declining sea ice on shipping activity in the Canadian Arctic. Geophysical Res. Lett. 43, 12146–12154. doi: 10.1002/2016GL071489
Raven Pro Interactive sound analysis software (Version 1.6.5) (2024). K Lisa Yang Center for Conservation Bioacoustics at the Cornell Lab of Ornithology. Available online at: https://ravensoundsoftware.com/ (Accessed February 20, 2025).
R Core Team (2022). R: A language and environment for statistical computing (Version 4.2.2) (R Foundation for Statistical Computing). Available online at: https://www.R-project.org/ (Accessed February 20, 2025).
Reichmuth C., Sills J. M., Brewer A., Triggs L., Ferguson R., Ashe E., et al. (2020). Behavioral assessment of in-air hearing range for the Pacific walrus (Odobenus rosmarus divergens). Polar Biol. 43, 767–772. doi: 10.1007/s00300-020-02667-6
Reis C. D. G., Padovese L. R., and De Oliveira M. C. F. (2019). Automatic detection of vessel signatures in audio recordings with spectral amplitude variation signature. Methods Ecol. Evol. 10, 1501–1516. doi: 10.1111/2041-210X.13245
Richardson W. J., Greene C. R. Jr., Malme C. I., Malme C.I., and Thomson D.H. (1995). Marine Mammals and Noise. 1st ed (San Diego, CA: Elsevier Science and Technology).
Rodríguez J. P., Klemm K., Duarte C. M., and Eguíluz V. M. (2024). Shipping traffic through the Arctic Ocean: Spatial distribution, seasonal variation, and its dependence on the sea ice extent. iScience 27, 110236. doi: 10.1016/j.isci.2024.110236
Romero L. M. (2004). Physiological stress in ecology: Lessons from biomedical research. Trends Ecol. Evol. 19, 249–255. doi: 10.1016/j.tree.2004.03.008
RStudio Team (2024). RStudio: Integrated Development for R (Version 2024.09.0.375) (RStudio PBC). Available online at: http://www.rstudio.com/ (Accessed February 20, 2025).
Salter R. E. (1979). Site utilization, activity budgets, and disturbance responses of Atlantic walruses during terrestrial haul-out. Can. J. Zoology 57, 1169–1180. doi: 10.1139/z79-149
Simard Y., Roy N., Giard S., Gervaise C., Conversano M., and Menard N. (2010). Estimating whale density from their whistling activity: Example with St. Lawrence beluga. Appl. Acoustics 71, 1081–1086. doi: 10.1016/j.apacoust.2010.05.013
Simard P., Wall K. R., Mann D. A., Wall C.C., and Stallings C.D. (2016). Quantification of boat visitation rates at artificial and natural reefs in the eastern Gulf of Mexico using acoustic recorders. PloS One 11, e0160695. doi: 10.1371/journal.pone.0160695
Sjare B. L. and Smith T. G. (1986). The vocal repertoire of white whales, Delphinapterus leucas, summering in Cunningham Inlet, Northwest Territories. Can. J. Zoology 64, 407–415. doi: 10.1139/z86-063
Sjare B. and Stirling I. (1996). The breeding behavior of Atlantic walruses, Odobenus rosmarus rosmarus, in the Canadian High Arctic. Can. J. Zool. 74, 897–911. doi: 10.1139/z96-103
Sjare B., Stirling I., and Spencer C. (2003). Structural variation in the songs of Atlantic walruses breeding in the Canadian High Arctic. Aquat. Mamm. 29, 297–318. doi: 10.1578/016754203101024121
Sorensen E., Ou H. H., and Zurk L. M. (2010). Passive acoustic sensing for detection of small vessels. MTS/IEEE Seattle OCEANS 2010, 1–6. doi: 10.1109/OCEANS.2010.5664542
Sousa-Lima R. S., Norris T. F., Oswald J. N., and Fernandes D. P. (2013). A review and inventory of fixed autonomous recorders for passive acoustic monitoring of marine mammals. Aquat. Mamm. 39, 23–53. doi: 10.1578/AM.39.1.2013.23
Southall B. L. (2021). Evolutions in marine mammal noise exposure criteria. Acoust. Today 17, 52. doi: 10.1121/AT.2021.17.2.52
Stevenson T. C., Davies J., Huntington H. P., and Sheard W. (2019). An examination of trans-Arctic vessel routing in the Central Arctic Ocean. Mar. Policy 100, 83–89. doi: 10.1016/j.marpol.2018.11.031
Stirling I., Calvert W., and Cleator H. (1983). Underwater vocalizations as a tool for studying the distribution and relative abundance of wintering pinnipeds in the High Arctic. Arctic, 36(3), 262–274. doi: 10.14430/arctic2267
Sweeney S. O., Terhune J. M., Frouin-Mouy H., and Rouget P. A. (2022). Assessing potential perception of shipping noise by marine mammals in an arctic inlet. J. Acoust. Soc Am. 151, 2310–2325. doi: 10.1121/10.0009956
Tervo O. M., Blackwell S. B., Ditlevsen S., Conrad A.S., Samson A.L., Garde E., et al. (2021). Narwhals react to ship noise and airgun pulses embedded in background noise. Biol. Lett. 17, 20210220. doi: 10.1098/rsbl.2021.0220
Tesei A., Meyer F., and Been R. (2020). Tracking of multiple surface vessels based on passive acoustic underwater arrays. J. Acoust. Soc Am. 147, EL87–EL92. doi: 10.1121/10.0000598
Thomisch K., Boebel O., Zitterbart D., Samaran F., Van Parijs S., van Opzeeland I., et al. (2015). Effects of subsampling of passive acoustic recordings on acoustic metrics. Journal of the Acoustical Society of America 138 (1), 267–278. doi: 10.1121/1.4922703
Thomsen F. and Popper A. N. (2024). Refocusing aquatic noise: Shifting from single to combined anthropogenic pressures. J. Acoust. Soc Am. 155, 3568–3572. doi: 10.1121/10.0026203
UNCTAD (2022). Review of Maritime Transport 2022-Navigating stormy waters (United Nations Conference on Trade and Development). Available online at: https://unctad.org/rmt2022 (Accessed April 17, 2023).
Vergara V., Mikus M., Chion C., Lagrois D., Marcoux M., and Michaud R. (2025). Effects of vessel noise on beluga (Delphinapterus leucas) call type use: ultrasonic communication as an adaptation to noisy environments? Biol. Open 14 , bio061783. doi: 10.1242/bio.061783
Vergara V., Wood J., Lesage V., Ames A., Mikus M., and Michaud R. (2021). Can you hear me? Impacts of underwater noise on communication space of adult, sub-adult and calf contact calls of endangered St. Lawrence belugas (Delphinapterus leucas). Polar Res. 40, 5521. doi: 10.33265/polar.v40.5521
Vieira M., Amorim M. C. P., Sundelöf A., Prista N., and Fonseca P. J. (2020). Underwater noise recognition of marine vessels passages: two case studies using hidden Markov models. ICES J. Mar. Sci. 77, 2157–2170. doi: 10.1093/icesjms/fsz194
Williams R., Erbe C., Ashe E., and Clark C. W. (2015). Quiet(er) marine protected areas. Mar. pollut. Bull. 100, 154–161. doi: 10.1016/j.marpolbul.2015.09.012
Williams R., Veirs S., Veirs V., Ashe E., and Mastick N. (2019). Approaches to reduce noise from ships operating in important killer whale habitats. Mar. pollut. Bull. 139, 459–469. doi: 10.1016/j.marpolbul.2018.05.015
Wright A. J., Soto N. A., Baldwin A. L., Bateson M., Beale C.M., Clark C., et al. (2007). Do marine mammals experience stress related to anthropogenic noise? Int. J. Comp. Psychol. 20, 274–316. doi: 10.46867/IJCP.2007.20.02.01
Zhu C., Garcia H., Kaplan A., SChinault M., Handegard N. O., Godø O. R., et al. (2018). Detection, localization and classification of multiple mechanized ocean vessels over continental-shelf scale regions with passive ocean acoustic waveguide remote sensing. Remote Sens. 10, 1699. doi: 10.3390/rs10111699
Keywords: passive acoustic monitoring (PAM), walrus, beluga, underwater noise, Arctic, vessel traffic, automatic identification system (AIS), marine protected area (MPA)
Citation: Coppolaro VLM, Ausen EL, Loseto LL and Marcoux M (2025) Vessel traffic disrupts walrus vocal behavior in a proposed marine protected area. Front. Mar. Sci. 12:1658328. doi: 10.3389/fmars.2025.1658328
Received: 02 July 2025; Accepted: 13 October 2025;
Published: 31 October 2025.
Edited by:
John Man Kon Wong, Ministry of Environment and Climate Change, QatarReviewed by:
Joseph G. Schnitzler, University of Veterinary Medicine Hannover, GermanyPhilippe Blondel, University of Bath, United Kingdom
Copyright © 2025 Coppolaro, Ausen, Loseto and Marcoux. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Veronica L. M. Coppolaro, Y29wcG9sYXZAbXl1bWFuaXRvYmEuY2E=