Abstract
Hydroacoustic mapping has recently challenged the long-held view of a uniform abyssal seafloor by predicting substantial habitat heterogeneity in this environment. The RUBBLE expedition M205 validated these predictions in the Vema Fracture Zone (VFZ) with visual surveys at six locations using a towed deep-sea camera system. Exposed rock outcrops and varied hard substrates were consistently confirmed in areas of high hard-rock exposure, while moderately predicted areas contained a mix of sediments, cobbles, and pebbles; low-potential sites were almost entirely covered in sediment. Although a detailed quantification is beyond the scope of this report, visual correspondence supports the reliability of hydroacoustic predictions for abyssal habitats. Notably, this study pioneers the application of hydroacoustic-based seafloor characterization at abyssal depths—extending methods formerly focused on bathyal zones to the planet’s largest benthic environment and enabling direct ground-truthing of habitat models below 5,000 m. These findings highlight abyssal habitat heterogeneity, confirm the utility of hydroacoustic tools for broad-scale benthic mapping, and establish a baseline for future research on deep-sea biodiversity and ecological dynamics.
Introduction
Contrary to popular belief, the abyssal deep ocean floor, at depths of 3,000–6,000 m, is not just a uniform sediment expanse. The formation of abyssal plains through sediment accumulation from the ocean surface or continental margins via turbidity currents has led to the belief that these plains are featureless (Straume et al., 2019). This is particularly true for seafloors older than 10 million years, where thick sediment layers may hide irregularities in oceanic crust (Ramirez-Llodra et al., 2010; Straume et al., 2019).
However, multiple recent studies have contributed to dispelling the notion of a monotonous abyss (see, e.g., Vanreusel et al., 2010; Durden et al., 2015; Purser et al., 2016; Smith, 2020). Riehl et al. (2020) predicted extensive rocky habitats along fracture zones in the abyssal regions of the Atlantic Ocean. Deep-sea researchers have long recognized hard substrates on the abyssal plains. Polymetallic nodules, discovered during the Challenger expedition (1872–1876) (Murray and Renard, 1891), are common in the Pacific abyss, but less so in the Indian and Atlantic Oceans (Dutkiewicz et al., 2015). In some Pacific areas, nodules may cover from 10% to over 50% of the seafloor (Wedding et al., 2013; Parnum et al., 2023), providing significant rocky habitats (Clark et al., 2010; Amon et al., 2016; Gooday et al., 2024). Seamounts also add to the rocky substrates at abyssal depths, with approximately 200,000 seamounts, mostly in the Pacific (Pitcher et al., 2007). Surveys using camera systems, manned submersibles, remotely operated vehicles (ROVs), and autonomous underwater vehicles have only covered a small proportion of the abyssal seafloor so far (Bell et al., 2025). Nevertheless, hard substrates have been found at abyssal depths in several ocean basins far from seamounts and mid-ocean ridges (Tilot et al., 2018). In addition to nodules, manganese (Mn) crusts have also been reported to provide important habitats for megafauna organisms (Thiel, 1992; Bitner et al., 2013; Purser et al., 2016; Brandt et al., 2018).
However, quantifying these features is challenging due to the limited survey coverage and the scarce high-resolution multibeam sonar mapping of the abyssal floor, which is essential for the identification of scarps and abrupt topographical features that may reveal rocky outcrops (Smith, 2020).
To address the lack of data on abyssal seafloor habitats, Riehl et al. (2020) used high-resolution multibeam sonar to survey a 2,700-km stretch of seafloor along the Vema Fracture Zone (VFZ), a transform fault crossing the Mid-Atlantic Ridge near 11° N. The ∼94,000-km2 survey area spans crustal ages from 0 to 100 Ma. By analyzing seafloor ruggedness and acoustic backscatter, the areas were classified into three habitat types: “sediment” (low hard-rock exposure potential), “transitional” (moderate hard-rock exposure potential), and “hard rock” (high hard-rock exposure potential). The young crust near the Mid-Atlantic Ridge (0–5 Ma) has a high hard-rock potential, with rocky habitats covering 30% of the area. Even the older crust (50–100 Ma) showed 5% rocky habitat. Transitional habitats, featuring hard surfaces, such as rock outcrops and polymetallic nodules, accounted for 33% of the area. This indicates that rocky habitats persist even on the ancient seafloor far from the ridge. Extrapolating to all Atlantic fracture zones, ∼264,000 km2 (9%) may have a high hard-rock exposure. Similar patterns are likely to extend across other ocean basins with mid-ocean ridges and fracture zones.
Why should we consider rocky habitats and habitat heterogeneity in the abyssal Atlantic Ocean and other oceans? Abyssal ecosystems are the largest benthic ecosystems on Earth, hosting immense biodiversity and genetic novelty (Shi et al., 2020; Lejzerowicz et al., 2021), which makes habitat mapping essential for understanding the distribution of life. Rocky substrates support distinct sessile communities, with diverse representations of sponges, corals, and anemones, many of which are long-lived and ecologically important (Purser et al., 2016; Mejía-Saenz et al., 2023). Mobile species such as crustaceans, polychaetes, sea cucumbers, and brittle stars are more common (Young, 2009; Durden et al., 2015; Simon-Lledó et al., 2019; Uhlenkott et al., 2023b). Habitat heterogeneity, particularly certain levels of rocky substrates in soft-sediment environments, enhances biodiversity (Bell et al., 2016) by providing niches, increasing food flux, offering prey or predator refuge, and influencing gene flow and speciation (Young, 2009; Durden et al., 2015; Vanreusel et al., 2016; Simon-Lledó et al., 2019). For example, Amon et al. (2016) found approximately 170 megafauna species in a 30-km × 30-km abyssal Pacific area, half reliant on Mn nodules, highlighting the importance of rocky habitats. If rocky habitats are more widespread than previously thought, then abyssal biodiversity is even more undersampled, necessitating targeted surveys of these poorly studied environments.
Finally, abyssal habitats face growing threats from climate change, mining, and pollution (Glover and Smith, 2003; Ramirez-Llodra, 2020), emphasizing the need for baseline data on habitat distribution and biodiversity patterns (Kuhn et al., 2020; Uhlenkott et al., 2020). The abyssal seafloor is not a homogeneous ecosystem, and damage to one area cannot be assumed to leave similar communities undisturbed elsewhere (Ramirez-Llodra et al., 2011; Sweetman et al., 2017; Stratmann et al., 2018; Levin et al., 2020; Riehl and De Smet, 2020). In this paper, we present new preliminary results from a recent expedition to the VFZ, validating the previously made predictions of the existence of exposed hard substrates and supporting hydroacoustic-based differentiation between areas of low, moderate, and high potential for rock exposure.
Methods
The inactive remnant of the Vema transform fault, known as the Vema Fracture Zone (VFZ), is situated in the Mid-Atlantic Ridge at approximately 11° N and extends eastward for a considerable distance, reaching deep into the Gambia Basin. Although fracture zones are typically characterized as simple, deep valleys, the bathymetric charts of a significant portion of the VFZ created during the Vema-TRANSIT project using multibeam bathymetry data (Devey et al., 2018) revealed substantial variations in the seafloor topography throughout most of its extent. The unexpected seafloor heterogeneity observed in these data, which include diverse slope angles and spatial distributions, implies the presence of exposed rocky outcrops (Riehl et al., 2020)—an unanticipated habitat type within the broader abyssal landscape.
The echosounder-based seafloor categorization by Riehl et al. (2020) used a combination of backscatter intensity (BS) and the ruggedness index (RI), illustrated as bivariate choropleth maps, to predict the distributions of three principal habitat types: “sediment” was defined as low hard-rock exposure potential, “transitional” as medium hard-rock exposure potential, and “hard rock” as high hard-rock exposure potential. Their sediment habitat type (category A1) (see legend in Figure 1) encompassed flat regions that exhibited low values for both RI and BS signals. The intermediate or the transitional habitat type (categories A2, B1, and B2) (see legend in Figure 1) included areas characterized either by low BS with intermediate RI values or by low-to-moderate RI combined with medium BS—typically indicative of scattered hard-rock fragments or Mn nodules. Areas classified as having high hard-seafloor potential (categories A3, B3, and C1–C3) (see legend in Figure 1) were defined primarily by the presence of high BS values, regardless of terrain ruggedness. However, regions with low-to-moderate BS were also included in this category if they exhibited high RI (Riehl et al., 2020). In the present study, we followed the same habitat delineation method.
Figure 1
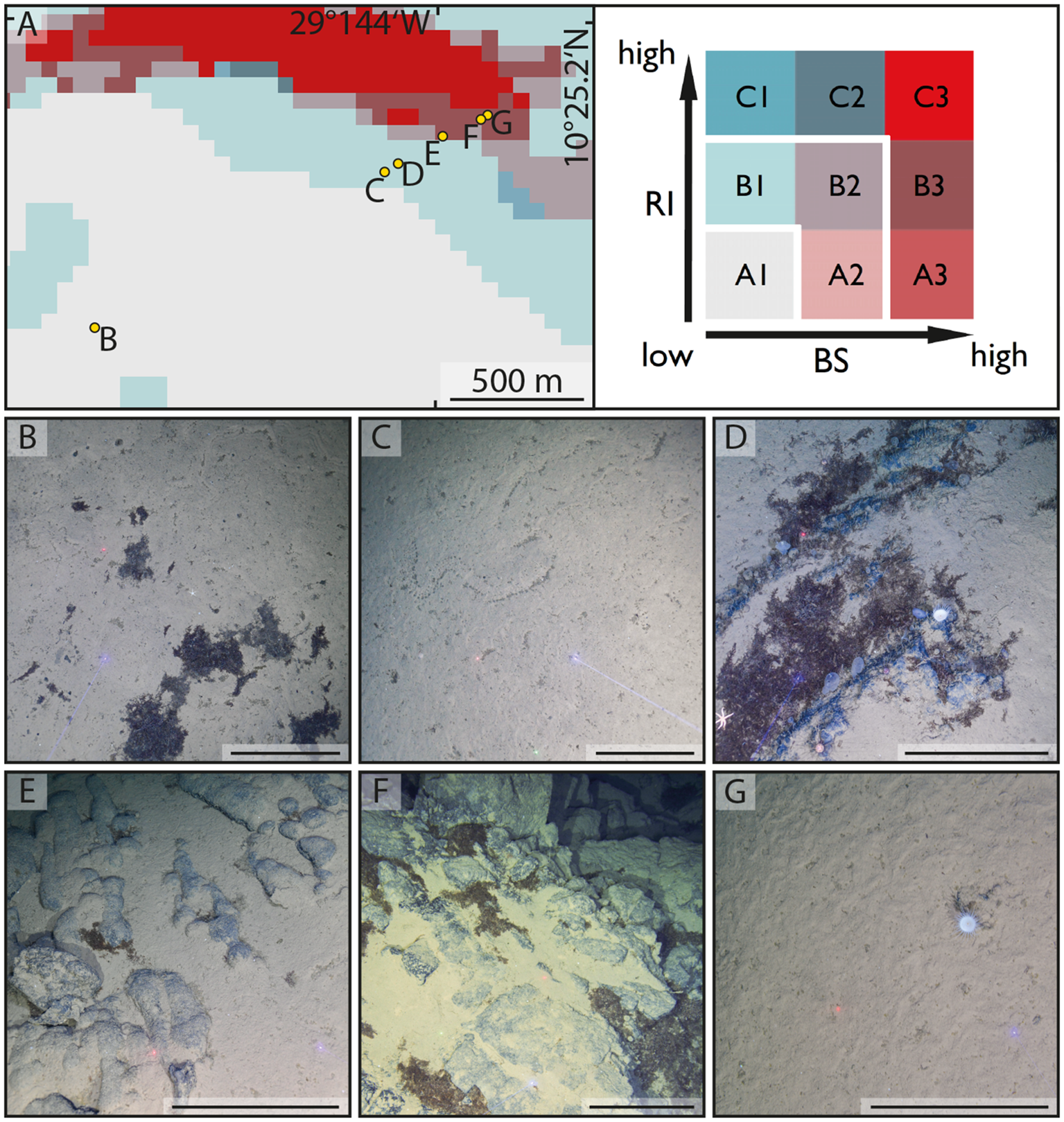
Seafloor habitat types encountered during the 205th voyage of R/V METEOR at the deep-sea mountain Mons Wehrmanni (Vema Fracture Zone) at sampling area SA3, station M205_41 (SA3-1). (A) Echosounder-based seafloor-type prediction using the combination of backscatter intensity (BS) and ruggedness index (RI), illustrated as a bivariate choropleth map following Riehl et al. (2020). (B) Deep-sea soft sediment with Lebensspuren and patches of sunken Sargassum debris typical for seafloor category A1 (low RI and low BS) and the principal habitat type “soft substrate.” (C) Sediment with Lebensspuren. (D) Stepped rocky outcrop with Sargassum debris and megafauna (anemone, sea urchins, sea star, sea squirts, and carnivorous sponges), both typical for category B1 (intermediate RI and low BS) and the “transitional” habitat type. (E) Pillow lava with partial sediment cover on a relatively gentle slope. (F) Rocky outcrop on a steeper slope, with patches of Sargassum debris and a relatively small area coverage of sediment, as frequently occurring on seafloor category B3 (intermediate RI and high BS) and the principal habitat type “high hard-rock exposure potential.” (G) Sediment-covered plateau (with solitary anemone), regularly encountered among the principal habitat type “high hard-rock exposure potential,” such as the seafloor category B3 (intermediate RI and high BS). Scales(B–G) = 50 cm.
From October 23 to November 28, 2024, the deep-sea exploration mission “RUBBLE” (Rocky seafloor—Underestimated Driver for Benthic Biodiversity Living in the Abyss and its Evolution) was conducted on the research vessel METEOR. The expedition utilized bathymetric maps and backscatter data from a previous voyage to the VFZ (Devey et al., 2018), along with habitat predictions derived from this information (Riehl et al., 2020). One of RUBBLE’s primary objectives was to verify seafloor-type predictions using deep-sea camera surveys. The full cruise track and overview station maps have been published in the cruise report (Riehl and Shipboard scientific crew, 2025) and are freely available.
Seafloor imaging was conducted during expedition M205 aboard R/V METEOR using the Ocean Floor Observation System (OFOS), a passively towed sled equipped with cameras and light. The system was deployed to map benthic habitats, document faunal communities and traces of bioturbation (Lebensspuren), and provide georeferenced data for selecting sampling sites and ground-truthing hydroacoustic observations. The survey areas were chosen to capture a representative range of seafloor habitats, based on available multibeam echosounder maps and predictions of habitat distribution, and were further adjusted prior to each deployment according to the prevailing weather, sea state, and current conditions to minimize ship movement and maintain stable towing. Track lines were generally planned downslope to reduce the risk of seafloor contact and to ensure that the system maintained a constant altitude above the seabed.
The OFOS was equipped with multiple imaging systems, including an Ocean Imaging System (Nikon D7100 DSLR, 24.1 MP), a high-definition video camera (Canon HF-G10, 1080p at 25 fps, 10× optical zoom), a Nikon D850 DSLR (46 MP) with a 35-mm objective mounted in a deep-sea housing, and a GoPro HERO13 Black (5.3-K resolution, 27.6 MP) mounted in a titanium housing with a glass dome. Still and video imaging was supported by four dimmable LED lights (two Sealite Sphere 5150 and two Teledyne Bowtech) together with a GEOMAR-developed pressure-neutral LED array. To provide spatial scale, two to three parallel laser beams were mounted around the video camera at equal distances of 50 cm. An Applied Microsystems SV Plus V2 probe was installed within the sled frame to record the conductivity, sound velocity, temperature, and pressure at 3-s intervals, with the sensor mounted approximately 0.5 m above the base of the frame. The frame was outfitted with additional weights and two hydrodynamic fins to increase stability during towing.
Deployment was conducted from the vessel’s starboard-side crane using an 18-mm coaxial cable that enabled simultaneous real-time data transmission and power supply. During descent, the first 100 m of cable was payed out at 0.5 m/s, with the remaining descent generally at 1 m/s until the system was positioned several tens of meters above the seafloor. Telemetry, cameras, and lights were activated during the descent, and once near-bottom, the control of winch operations was transferred from the deck to the OFOS laboratory. R/V METEOR does not have automatic pitch compensation; therefore, ship motion was corrected manually through continuous communication between the winch operator, the bridge, and the scientific team in the OFOS control room. During surveys, the OFOS was towed 2 m above the bottom at speeds of 0.4–0.6 kn. Still images were acquired at 2-s intervals to ensure a high overlap suitable for creating photo mosaics and 3D reconstructions of the seabed.
The position of the OFOS was continuously monitored using the Ultra-Short Baseline acoustic positioning system Posidonia6000, which required the attachment of a pinger to the frame prior to each dive. The USBL provided point-cloud positioning data that were averaged during post-processing, enabling georeferencing of imagery and revisiting the survey tracks. Live video from the Canon HF-G10 was transmitted to the ship’s control room, bridge, meeting room, and intranet to allow real-time monitoring, annotation of habitat variation and faunal occurrences, and navigation support for maintaining altitude above bottom. A Blackmagic Design ATEM Mini Extreme ISO video mixer was used to record the live feed together with the ship and gear metadata, logbook entries, and geospatial information on both the vessel and OFOS position (processed in QGIS and Delph RoadMap). The OIS, Nikon D850, and GoPro cameras operated autonomously and stored imagery internally. After each deployment, the data were offloaded immediately, and all cameras were synchronized to Coordinated Universal Time (UTC) prior to surveys to ensure temporal consistency across datasets.
During expedition M205, a total of 177,038 seabed photographs were acquired across 14 survey stations (Table 1). The functionality of the individual imaging components varied with deployment, but image acquisition was consistent and provided a high-resolution dataset suitable for quantitative analysis of seafloor habitats, associated megabenthic communities, and seabed structures. A more detailed description of the OFOS is provided in Chapter 5.3 of the cruise report (Riehl and Shipboard scientific crew, 2025).
Table 1
| Station (ship) | Station | Event time (UTC) | Action | Lat (dec) [ship] | Long (dec) [ship] | Lat (dec) [OFOS] | Long (dec) [OFOS] | Depth (m) |
|---|---|---|---|---|---|---|---|---|
| M205_1 | SA1-1 | 29.10.2024 09:50 | Start | 10.5295667 | −26.7481 | 10.5289788 | −26.7491945 | 5,098 |
| M205_1 | SA1-1 | 29.10.2024 14:31 | End | 10.5403167 | −26.7213167 | 10.5373957 | −26.7245087 | 5,647 |
| M205_4 | SA1-4 | 29.10.2024 20:26 | Start | 10.5380667 | −26.7536 | 10.5377683 | −26.7547942 | 5,206 |
| M205_4 | SA1-4 | 30.10.2024 00:45 | End | 10.5478333 | −26.7315667 | 10.545994 | −26.7329237 | 5,681 |
| M205_21 | SA2-1 | 01.11.2024 23:10 | Start | 10.3460333 | −27.2146833 | 10.3455527 | −27.2156822 | 4,865 |
| M205_21 | SA2-1 | 02.11.2024 05:10 | End | 10.3533167 | −27.1771667 | 10.3518 | −27.1796 | 5,654 |
| M205_22 | SA2-4 | 02.11.2024 11:40 | Start | 10.4389 | −27.2333333 | 10.4385037 | −27.233627 | 5,220 |
| M205_22 | SA2-4 | 02.11.2024 14:53 | End | 10.4503 | −27.2206667 | 10.4475742 | −27.2226813 | 5,606 |
| M205_41 | SA3-1 | 06.11.2024 10:06 | Start | 10.4093667 | −29.2529 | 10.4082725 | −29.2537135 | 5,462 |
| M205_41 | SA3-1 | 06.11.2024 15:22 | End | 10.4251333 | −29.22545 | 10.4227457 | −29.227686 | 5,835 |
| M205_42 | SA3-2 | 06.11.2024 21:43 | Start | 10.3603167 | −29.22665 | 10.3590595 | −29.2273402 | 5,100 |
| M205_42 | SA3-2 | 07.11.2024 01:07 | End | 10.36465 | −29.2042 | 10.3635 | −29.2053 | 5,560 |
| M205_62 | SA4-1 | 10.11.2024 17:48 | Start | 10.4714667 | −30.9837667 | 10.4707235 | −30.9847887 | 5,057 |
| M205_62 | SA4-1 | 10.11.2024 23:49 | End | 10.4903167 | −30.94855 | 10.4875148 | −30.9527645 | 5,511 |
| M205_64 | SA4-3 | 11.11.2024 07:32 | Start | 10.4558667 | −30.9678833 | 10.4550512 | −30.9690338 | 5,207 |
| M205_64 | SA4-3 | 11.11.2024 11:20 | End | 10.4715 | −30.9492333 | 10.4686207 | −30.9514342 | 5,572 |
| M205_80 | SA5-1 | 14.11.2024 23:24 | Start | 10.2238167 | −34.4850667 | 10.2225552 | −34.4850197 | 4,849 |
| M205_80 | SA5-1 | 15.11.2024 05:13 | End | 10.2539333 | −34.4645 | 10.2504008 | −34.4662342 | 5,105 |
| M205_82 | SA5-3 | 15.11.2024 13:11 | Start | 10.2651167 | −34.4749333 | 10.2636508 | −34.47521 | 5,112 |
| M205_82 | SA5-3 | 15.11.2024 19:06 | End | 10.297 | −34.4460667 | 10.2890315 | −34.4504515 | 5,606 |
| M205_98 | SA5-6 | 18.11.2024 16:24 | Start | 10.2008667 | −34.50265 | 10.1995205 | −34.5033 | 4,292 |
| M205_98 | SA5-6 | 18.11.2024 22:18 | End | 10.2269667 | −34.4743167 | 10.2236165 | −34.4772222 | 4,884 |
| M205_102 | SAX-1 | 19.11.2024 15:59 | Start | 10.0637167 | −34.6706167 | 10.0627262 | −34.6718223 | 5,278 |
| M205_102 | SAX-1 | 19.11.2024 22:39 | End | 10.1027167 | −34.6514167 | 10.0981853 | −34.6541502 | 5,366 |
| M205_108 | SAX-5 | 20.11.2024 13:57 | Start | 10.04465 | −34.6678833 | 10.0440268 | −34.6690983 | 5,338 |
| M205_108 | SAX-5 | 20.11.2024 17:45 | End | 10.0628333 | −34.6530167 | 10.0598068 | −34.6557713 | 5,406 |
| M205_115 | SAX-12 | 22.11.2024 05:05 | Start | 10.0308 | −34.3488833 | 10.0295708 | −34.3504687 | 5,429 |
| M205_115 | SAX-12 | 22.11.2024 12:07 | End | 10.0708833 | −34.3148667 | 10.0672737 | −34.3169832 | 5,408 |
Station data of the Ocean Floor Observation System (OFOS) deployed during the 205th voyage of R/V METEOR.
The raw and processed image data are available on the Zenodo repository (European Organization For Nuclear Research and OpenAIRE, 2013): https://doi.org/10.5281/zenodo.16993854.
Results
For each of the OFOS dives, hard substrates were recorded in areas where a high hard-rock exposure potential was predicted (Figures 1–3). In these areas, a variety of different types of hard substrates were observed, ranging from outcropping, often apparently Mn-encrusted, through rounded pillow lavas of varying sizes (Figures 1E, 2D, 3E, F), to debris deposits probably of basaltic nature (Figure 1F). The hard substrates that we observed were frequently partially covered by sediments in less exposed areas (Figures 1E, F). Areas of high hard-rock exposure potential also included largely sedimented areas where rocky substrates were visible only in relatively small patches (Figure 1G). The frequent occurrence of hard-substrate-dependent megafauna organisms, such as certain sponges (Figure 2C) and anemones (Figure 1G), in sedimented areas indicates that only a relatively thin layer of sediment blankets an underlying rocky substrate.
Figure 2

Seafloor habitat types encountered during the 205th voyage of R/V METEOR at the deep-sea mountain Mons RUBBLE (Vema Fracture Zone) at sampling area SA4, station M205_64 (SA4-3). (A) Echosounder-based seafloor-type prediction using the combination of backscatter intensity (BS) and ruggedness index (RI), illustrated as a bivariate choropleth map following Riehl et al. (2020) (see Figure 1 for legend). (B) Deep-sea sediment with patches of sunken Sargassum debris typical for seafloor category A1 (low RI and low BS) and the principal habitat type “soft substrate.” (C) Sediment with Sargassum debris and a glass sponge (Hexactinellida) commonly occurring in areas of transitional habitats and intermediate exposure potential for hard substrates (e.g., category B1—intermediate RI and low BS). (D) Rocky outcrop (partly represented by pillow lava) with sediment plateaus on top and below, as well as sessile megafauna (anemones and sea squirt), as typical for category C1 (high RI and low BS) and the “high hard-rock exposure potential” habitat type. Scales(B–D) = 50 cm.
Figure 3

Seafloor habitat types encountered during the 205th voyage of R/V METEOR at the deep-sea mountain Mons Wehrmanni (Vema Fracture Zone) at sampling area SA3, station M205_42 (SA3-2). Examples of the “transitional” habitat type, represented by category B1 [intermediate ruggedness index (RI) and low backscatter intensity (BS)] in (B–D) and of the principal habitat type “high hard-rock exposure potential,” represented by seafloor category C1 (high RI and low BS) in (E, F). (A) Echosounder-based seafloor-type prediction using the combination of BS and RI, illustrated as a bivariate choropleth map following Riehl et al. (2020) (see Figure 1 for legend). (B) Manganese nodules embedded in soft sediment. (C) Sediment with locomotion Lebensspuren and a sea cucumber (Holothuroidea). (D) Stepped rocky outcrop with Sargassum debris and sessile megafauna (anemones). (E) Outcropping pillow lava with partial sediment cover on a relatively gentle slope. (F) Rocky outcrop containing pillow lava (inhabited by motile and sessile megafauna: anemones) and sedimented areas on a steeper slope with patches of Sargassum debris. Scales(B–F) = 50 cm.
In areas of moderate hard-rock potential, we observed entirely sediment-covered surfaces (Figures 1C, 2C) alongside areas with cobbles and pebbles of varying densities on the seafloor (Figure 3B). In addition, boulders and outcropping rock were also observed in isolated instances, embedded in unconsolidated sediments (Figures 1D, 3D). Areas with moderate hard-rock potential often represent transitional zones between regions with low and high hard-substrate potential (see maps in Figures 1-3). Accordingly, the proportion of rocky seafloor was observed to increase toward the transition from moderate to high hard-rock potential (e.g., Figures 1C, D). As for areas of high hard-rock exposure potential, also here in several instances, hard-substrate-dependent megafauna organisms point at a relatively thin sediment blanket on top of a rocky substrate (Figure 1G).
Areas for which mostly sediments and low hard-substrate potential were predicted were mostly confirmed as free of hard substrates (Figures 1B, 2B) or were only locally covered with single rocks or sparsely covered with stones. The thickness of the sediment overlying the crust could not be studied using the methods applied in this study; however, in a few instances, the occasional occurrence of hard-substrate inhabitants indicates the presence of rocks or nodules just below the sediment surface (Figure 2B).
Discussion
The assumption of a homogeneous abyssal seafloor has long been challenged by reports showing that some abyssal regions exhibit higher faunal diversity than shallow-water environments (Ramirez-Llodra et al., 2010; McClain and Schlacher, 2015). The discovery of hard substrates in the abyss has significantly affected deep-sea ecology and biodiversity studies, providing new insights into the complexity and diversity of deep-sea ecosystems. Hard substrates such as seamounts, nodule fields, and abyssal hills have been found to host diverse and abundant communities of deep-sea organisms (Pitcher et al., 2007; Durden et al., 2015; Amon et al., 2016; Purser et al., 2016; Simon-Lledó et al., 2019). The presence of hard substrates introduced habitat heterogeneity into the abyssal environment, which was previously thought to be relatively uniform. This heterogeneity has been shown to have a significant ecological impact (Uhlenkott et al., 2023a). For example, a study on abyssal hills found that megafaunal biomass was significantly greater on hills than on adjacent plains, with the differences in assemblage and trophic compositions correlating with the sediment particle size distributions (Durden et al., 2015). This suggests that the local topography and hydrodynamics play a crucial role in shaping deep-sea communities. The discovery of hard substrates in the abyss has challenged some long-held paradigms in deep-sea ecology. Although the deep sea has long been recognized as biologically highly diverse at the local scale, the presence of hard substrates has revealed that the species turnover at regional and larger geographical scales may be more significant than previously thought (Lins et al., 2017). This finding emphasizes the need for a more nuanced understanding of the deep-sea biodiversity patterns and their influencing factors.
Understanding the deep-sea habitat heterogeneity and substrate composition across large deep-sea ecosystems, particularly in the abyss, is crucial for several reasons. Deep-sea habitat heterogeneity significantly influences the species diversity and distribution patterns (Vanreusel et al., 2010, 2016; Durden et al., 2015; Amon et al., 2016; Uhlenkott et al., 2022). Increased habitat heterogeneity has important implications for biodiversity and ecosystem functioning (Staudacher et al., 2018). Seafloor heterogeneity allows for diversified assemblages across different habitats, increasing the diversity and influencing the ecosystem processes at the regional scale (Zeppilli et al., 2016). Therefore, understanding deep-sea habitat heterogeneity is essential to accurately assess the biodiversity, the ecosystem functioning, and the potential impacts of human activities. This highlights the need for more sophisticated sampling campaigns to capture the full range of deep-sea habitat heterogeneity and biodiversity (Riehl et al., 2020; Smith, 2020). In addition, high-resolution seabed mapping and detailed analyses of the species distributions at the habitat scale are crucial for improving the management of goods and services delivered by deep-sea ecosystems (Zeppilli et al., 2016).
Herein, we report a repeated and unambiguous confirmation of exposed rock outcrops across a wide range of crustal ages along the eastern VFZ. Our camera surveys recorded habitat distributions that matched areas with high hard-rock potential, as predicted using hydroacoustic seafloor mapping (Riehl et al., 2020). These findings provide strong evidence of the accuracy of the geological predictions and mapping techniques in the context of biological habitat surveys. The frequent occurrence of hard substrates in areas with moderate potential is valuable for refining predictive models and for improving the accuracy of echosounder-based biodiversity surveys. Despite predictions of their potential presence, the absence of hard substrates in certain areas highlights the complexity of geological formations and the need for continuous refinement of the assessment techniques. This discrepancy can be explained by the fact that the sediment cover is too thin to make a difference in the multibeam and backscatter signals, which can occur several meters thick, or by the low resolution of the multibeam seafloor maps at water depths of 4,500–5,500 m, which makes it impossible to resolve smaller or patchily occurring seafloor structures, emphasizing the importance of ground-truthing and multifaceted approaches in such investigations.
This study represents a first step toward establishing the effectiveness of hydroacoustic-based seafloor characterization for the mapping and interpretation of abyssal landscapes as introduced by Riehl et al. (2020), advancing its use from bathyal and shallower to much deeper environments by successfully ground-truthing predictions at depths exceeding 5,000 m and expanding the usability of this method, as presented in an increasing number of studies (see, e.g., Uhlenkott et al., 2020; Fejer et al., 2021). Acoustic mapping techniques revealed pronounced habitat heterogeneity, and the visual confirmation of hard substrates robustly validates the hydroacoustic method within the VFZ. These results demonstrate that this approach can be reliably applied across other deep-sea regions, potentially revolutionizing our understanding of benthic ecosystem distribution and diversity. Accurate identification of hard-substrate areas has far-reaching implications for the planning of deep-sea research, conservation strategies, and the management of future human activities such as deep-sea mining, supporting effective stewardship of the largest and least explored marine habitats on Earth.
Conclusion
Utilizing a towed deep-sea camera system, hydroacoustics-based habitat predictions of the abyssal seafloor were corroborated, enhancing our understanding of deep-sea ecology and biodiversity. The findings of the RUBBLE campaign validated the hydroacoustic prediction method for habitat types on the abyssal seafloor proposed by Riehl et al. (2020). The acoustic predictions accurately identified the areas of hard substrate and soft sediment across the study region. Consequently, this mapping approach is a valuable tool for elucidating deep-sea benthic habitats on a broad spatial scale. Further ground-truthing with visual surveys and ROV-based sediment sampling would contribute to refining the acoustic classification and validating its applicability in other abyssal environments. ROV sampling is the preferred method for the investigation of faunal communities inhabiting hard substrates. These findings offer new perspectives on the unexplained high deep-sea diversity and emphasize the need for continued exploration and research in deep-sea environments to fully comprehend the complexity and diversity of these ecosystems. Such confirmations are crucial for baseline biodiversity research and environmental impact assessments as they provide empirical evidence of seafloor surface conditions. These exposed rock formations may serve as visible indicators of megabenthic epifauna, enabling biotope and biodiversity assessments using remote sensing methods. The degree to which these formations represent habitats and biodiversity remains to be determined.
The development of science-based societal solutions to protect ecosystems and predict how the effects of point-source human disturbance (e.g., mining and pollution) propagate regionally (Mitchell et al., 2020) in deep-sea ecosystems has been largely restricted to seamounts and hydrothermal vents (Chiu et al., 2020; Fleming et al., 2022). Uncovering habitat heterogeneity on the abyssal seafloor on the large scale via remote sensing provides an opportunity to employ metacommunity theory in this largely overlooked but spatially dominant seascape and may eventually allow testing classical metacommunity assumptions, such as limited dispersal and species turnover dominated by environmental filtering (Mullineaux et al., 2018).
Statements
Data availability statement
The original contributions presented in the study are publicly available. This data can be found here: https://zenodo.org/records/16993854.
Author contributions
TR: Conceptualization, Funding acquisition, Investigation, Project administration, Resources, Supervision, Writing – original draft, Writing – review & editing. LB: Data curation, Formal Analysis, Investigation, Methodology, Visualization, Writing – review & editing. AB: Resources, Supervision, Validation, Writing – review & editing. MH: Data curation, Methodology, Writing – review & editing. PA: Data curation, Investigation, Visualization, Writing – review & editing. FM: Data curation, Writing – review & editing. NA: Supervision, Visualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. The financing of OFOS missions was made possible through generous private philanthropic donations. We thank the German Science Foundation (DFG) and the Federal Ministry for Education and Research (BMBF) for funding the expedition M205 (GPF 21-1_060). Cruise report: https://www.pangaea.de/expeditions/bybasis/Meteor%20%281986%29. The donors were not involved in the study design, collection, analysis, interpretation of data, the writing of this article, or the decision to submit it for publication.
Acknowledgments
We extend our gratitude to Captain Rainer Hammacher and his crew for their outstanding support, teamwork, and fostering a positive atmosphere throughout cruise M205. The exceptional efforts of the R/V METEOR crew were essential to the success of the expedition. This work was made possible by funding for the “RUBBLE” project from the German Research Foundation (DFG) and the Federal Ministry of Education and Research (BMBF) through the Expert Panel Research Vessels (“Gutachterpanel Forschungsschiffe” (GPF)) under project ID GPF 21-1_060. In addition, financing OFOS missions was made possible by generous private philanthropic donations. OFOS was provided by the GEOMAR Helmholtz Centre for Ocean Research Kiel. During the OFOS deployments, Anchita Casaubon, Angelina Eichsteller, Julia Jacoby, and Henry Knauber helped obtain an initial impression of the seafloor by providing live annotations. We are greatly thankful to Mike Wilsenack for providing us with a camera system on short notice. The RUBBLE campaign was logistically and financially supported by the Senckenberg Ocean Species Alliance (SOSA), particularly Ruth Wasmund, Alica Torkov, and Carly Rospert. This is SOSA contribution #89 and RUBBLE contribution #2. The invaluable support by the logistics team of the German Centre for Marine Biodiversity Research (DZMB) is greatly appreciated. Karmen Yoshida provided administrative assistance, without which the campaign would not have been possible. Finally, the first author would like to express his deep appreciation and gratitude to his family, whose selfless support made the expedition possible. The authors used written content produced by a generative AI technology; specifically, the AI-powered writing assistant Paperpal was used. The application of AI was limited to text polishing (Paperpal’s grammar checker, paraphraser, plagiarism checker and manuscript checker). The authors verified the factual accuracy of any content created using the generative AI technologies.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that Generative AI was used in the creation of this manuscript. The authors used written content produced by a generative AI technology; specifically, the AI-powered writing assistant Paperpal was used. The application of AI was limited to text polishing (Paperpal’s grammar checker, paraphraser, plagiarism checker and manuscript checker). Additionally, the abstract and summary in common language were written with the help of AI. The authors verified the factual accuracy of any content created using the generative AI technologies.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1
Amon D. J. Ziegler A. F. Dahlgren T. G. Glover A. G. Goineau A. Gooday A. J. et al . (2016). Insights into the abundance and diversity of abyssal megafauna in a polymetallic-nodule region in the eastern Clarion-Clipperton Zone. Sci. Rep.6, 30492. doi: 10.1038/srep30492
2
Bell J. B. Alt C. H. S. Jones D. O. B. (2016). Benthic megafauna on steep slopes at the Northern Mid-Atlantic Ridge. Mar. Ecol.37, 1290–1302. doi: 10.1111/maec.12319
3
Bell K. L. C. Johannes K. N. Kennedy B. R. C. Poulton S. E. (2025). How little we’ve seen: A visual coverage estimate of the deep seafloor. Sci Adv.11, eadp8602. doi: 10.1126/sciadv.adp8602
4
Bitner M. A. Melnik V. P. Zezina O. N. (2013). New paedomorphic brachiopods from the abyssal zone of the north-eastern Pacific Ocean. Zootaxa3613, 281–288. doi: 10.11646/zootaxa.3613.3.6
5
Brandt A. Frutos I. Bober S. Brix S. Brenke N. Guggolz T. et al . (2018). Composition of abyssal macrofauna along the Vema Fracture Zone and the hadal Puerto Rico Trench, northern tropical Atlantic. Deep-Sea Res. Pt. II148, 35–44. doi: 10.1016/j.dsr2.2017.07.014
6
Chiu M.-C. Ao S. He F. Resh V. H. Cai Q. (2020). Elevation shapes biodiversity patterns through metacommunity-structuring processes. Sci Total Environ.743, 140548. doi: 10.1016/j.scitotenv.2020.140548
7
Clark M. R. Rowden A. A. Schlacher T. Williams A. Consalvey M. Stocks K. I. et al . (2010). The ecology of seamounts: structure, function, and human impacts. Annu. Rev. Mar. Sci2, 253–278. doi: 10.1146/annurev-marine-120308-081109
8
Devey C. W. Augustin N. Yeo I. A. (2018). Raw multibeam EM122 data and data products: SONNE cruise SO237 (North Atlantic). doi: 10.1594/PANGAEA.893352
9
Durden J. M. Bett B. J. Jones D. O. B. Huvenne V. A. I. Ruhl H. A. (2015). Abyssal hills – hidden source of increased habitat heterogeneity, benthic megafaunal biomass and diversity in the deep sea. Prog. Oceanography137, 209–218. doi: 10.1016/j.pocean.2015.06.006
10
Dutkiewicz A. Müller R. D. O’Callaghan S. Jónasson H. (2015). Census of seafloor sediments in the world’s ocean. Geology. 43, 795–798. doi: 10.1130/G36883.1
11
European Organization For Nuclear Research and OpenAIRE (2013). Zenodo: research. doi: 10.25495/7GXK-RD71
12
Fejer A. Cecino G. Flynn A. (2021). Habitat mapping for ecosystem-based management of deep-sea mining. Mar. Technol. Soc. J.55, 53–64. doi: 10.4031/MTSJ.55.6.4
13
Fleming B. F. Beaulieu S. E. Mills S. W. Gaggiotti O. E. Mullineaux L. S. (2022). Ecological connectivity in Pacific deep-sea hydrothermal vent metacommunities. Mar. Ecol. Prog. Series. 731, 267-278. doi: 10.3354/meps14182
14
Glover A. G. Smith C. R. (2003). The deep-sea floor ecosystem: current status and prospects of anthropogenic change by the year 2025. Environ. Conserv.30, 219–241. doi: 10.1017/S0376892903000225
15
Gooday A. J. Holzmann M. Barrenechea-Angeles I. Lim S.-C. Pawlowski J. (2024). New xenophyophores (Foraminifera, Monothalamea) from the eastern Clarion-Clipperton Zone (equatorial Pacific). Zootaxa5419, 151–188. doi: 10.11646/zootaxa.5419.2.1
16
Kuhn T. Uhlenkott K. Vink A. Rühlemann C. Martínez Arbizu P. (2020). “ Manganese nodule fields from the Northeast Pacific as benthic habitats,” in Seafloor Geomorphology as Benthic Habitat. Eds. HarrisP. T.BakerE. ( Amsterdam, Oxford, Cambridge: Elsevier Science Ltd), 933–947. doi: 10.1016/B978-0-12-814960-7.00058-0
17
Lejzerowicz F. Gooday A. J. Barrenechea Angeles I. Cordier T. Morard R. Apothéloz-Perret-Gentil L. et al . (2021). Eukaryotic biodiversity and spatial patterns in the clarion-clipperton zone and other abyssal regions: insights from sediment DNA and RNA metabarcoding. Front. Mar. Sci8. doi: 10.3389/fmars.2021.671033
18
Levin L. A. Amon D. J. Lily H. (2020). Challenges to the sustainability of deep-seabed mining. Nat. Sustainability. 3, 1–11. doi: 10.1038/s41893-020-0558-x
19
Lins L. Leliaert F. Riehl T. Pinto Ramalho S. Alfaro Cordova E. Morgado Esteves A. et al . (2017). Evaluating environmental drivers of spatial variability in free-living nematode assemblages along the Portuguese margin. Biogeosciences14, 651–669. doi: 10.5194/bg-14-651-2017
20
McClain C. R. Schlacher T. A. (2015). On some hypotheses of diversity of animal life at great depths on the sea floor. Mar. Ecol.26, 849–872. doi: 10.1111/maec.12288
21
Mejía-Saenz A. Simon-Lledó E. Partridge L. S. Xavier J. R. Jones D. O. B. (2023). Rock outcrops enhance abyssal benthic biodiversity. Deep Sea Res. Part I: Oceanographic Res. Papers195, 103999. doi: 10.1016/j.dsr.2023.103999
22
Mitchell E. G. Durden J. M. Ruhl H. A. (2020). First network analysis of interspecific associations of abyssal benthic megafauna reveals potential vulnerability of abyssal hill community. Prog. Oceanography187, 102401. doi: 10.1016/j.pocean.2020.102401
23
Mullineaux L. S. Metaxas A. Beaulieu S. E. Bright M. Gollner S. Grupe B. M. et al . (2018). Exploring the ecology of deep-sea hydrothermal vents in a metacommunity framework. Front. Mar. Sci5, 49. doi: 10.3389/fmars.2018.00049
24
Murray J. Renard A. F. (1891). Report on deep-sea deposits based on the specimens collected during the voyage of HMS Challenger in the years 1872 to 1876 ( Neill and Company for her Majesty's Stationery Office, Edinburgh). Available online at: https://books.google.de/books?hl=de&lr=&id=rPlCAQAAIAAJ&oi=fnd&pg=PR9&dq=Report+on+Deep-Sea+Deposits+Based+on+the+Specimens+Collected+During+the+Voyage+of+H.M.S.+Challenger&ots=wtS6wR3CPR&sig=WfON91FTV49LcOcjHr-IfnRhQGM.
25
Parnum I. M. Flynn A. Donnelly D. M. Clarke M. (2023). Underwater acoustic surveys in the eastern Clarion Clipperton Zone. J. Acoustical Soc. America154, A275–A275. doi: 10.1121/10.0023511
26
Pitcher T. J. Morato T. Hart P. J. B. Clark M. R. Haggan N. Santos R. S. (Eds.) (2007). Seamounts: Ecology, Fisheries & Conservation (Oxford, UK: Blackwell Publishing Ltd). doi: 10.1002/9780470691953
27
Purser A. Marcon Y. Hoving H.-J. T. Vecchione M. Piatkowski U. Eason D. et al . (2016). Association of deep-sea incirrate octopods with manganese crusts and nodule fields in the Pacific Ocean. Curr. Biol.26, R1268–R1269. doi: 10.1016/j.cub.2016.10.052
28
Ramirez-Llodra E. (2020). “Deep-sea ecosystems: biodiversity and anthropogenic impacts,” in The Law of the Seabed, ed. C. Banet (Leiden; Boston: Brill Nijhoff), 36–60. Available at: https://brill.com/view/book/edcoll/9789004391567/BP000013.xml (Accessed August 26, 2020)
29
Ramirez-Llodra E. Brandt A. Danovaro R. De Mol B. Escobar E. German C. R. et al . (2010). Deep, diverse and definitely different: unique attributes of the world’s largest ecosystem. Biogeosciences7, 2851–2899. doi: 10.5194/bg-7-2851-2010
30
Ramirez-Llodra E. Tyler P. A. Baker M. C. Bergstad O. A. Clark M. R. Escobar E. et al . (2011). Man and the last great wilderness: human impact on the deep sea. PloS One6, e22588. doi: 10.1371/journal.pone.0022588
31
Riehl T. De Smet B. (2020). Macrostylis metallicola spec. nov. — An isopod with geographically clustered genetic variability from a polymetallic-nodule area in the Clarion-Clipperton Fracture Zone. PeerJ8, 1–44. doi: 10.7717/peerj.8621
32
Riehl T. Shipboard scientific crew (2025). Rocky seafloor — Underestimated Driver for Benthic Biodiversity Living in the Abyss and its Evolution. Bonn: Begutachtungspanel Forschungsschiffe. doi: 0.48433/cr_m205
33
Riehl T. Wölfl A.-C. Augustin N. Devey C. W. Brandt A. (2020). Discovery of widely available abyssal rock patches prompts rethinking origins of deep-sea biodiversity. Proc. Natl. Acad. Sci. U.S.A117, 15450–15459. doi: 10.1073/pnas.1920706117
34
Shi J. Lei Y. Li Q. Lyu M. Li T. (2020). Molecular diversity and spatial distribution of benthic foraminifera of the seamounts and adjacent abyssal plains in the tropical Western Pacific Ocean. Mar. Micropaleontology156, 101850. doi: 10.1016/j.marmicro.2020.101850
35
Simon-Lledó E. Bett B. J. Huvenne V. A. I. Schoening T. Benoist N. M. A. Jones D. O. B. (2019). Ecology of a polymetallic nodule occurrence gradient: Implications for deep-sea mining. Limnology Oceanography64, 1883–1894. doi: 10.1002/lno.11157
36
Smith C. R. (2020). The heterogeneous abyss. Proc. Natl. Acad. Sci. U.S.A. 177, 16729-16731. doi: 10.1073/pnas.2010215117
37
Staudacher K. Rennstam Rubbmark O. Birkhofer K. Malsher G. Sint D. Jonsson M. et al . (2018). Habitat heterogeneity induces rapid changes in the feeding behaviour of generalist arthropod predators. Funct. Ecol.32, 809–819. doi: 10.1111/1365-2435.13028
38
Stratmann T. Lins L. Purser A. Marcon Y. Rodrigues C. F. Ravara A. et al . (2018). Abyssal plain faunal carbon flows remain depressed 26 years after a simulated deep-sea mining disturbance. Biogeosciences15, 4131–4145. doi: 10.5194/bg-15-4131-2018
39
Straume E. O. Gaina C. Medvedev S. Hochmuth K. Gohl K. Whittaker J. M. et al . (2019). GlobSed: updated total sediment thickness in the world’s oceans. Geochemistry Geophysics Geosystems20, 1756–1772. doi: 10.1029/2018GC008115
40
Sweetman A. K. Thurber A. R. Smith C. R. Levin L. A. Mora C. Wei C.-L. et al . (2017). Major impacts of climate change on deep-sea benthic ecosystems. Elementa: Sci Anthropocene5, 1–23. doi: 10.1525/elementa.203
41
Thiel H. (1992). Deep-sea environmental disturbance and recovery potential. Internationale Rev. der gesamten Hydrobiologie und Hydrographie77, 331–339. doi: 10.1002/iroh.19920770213
42
Tilot V. Ormond R. Moreno Navas J. Catalá T. S. (2018). The benthic megafaunal assemblages of the CCZ (Eastern Pacific) and an approach to their management in the face of threatened anthropogenic impacts. Front. Mar. Sci.5. doi: 10.3389/fmars.2018.00007
43
Uhlenkott K. Meyn K. Vink A. Martínez Arbizu P. (2023a). A review of megafauna diversity and abundance in an exploration area for polymetallic nodules in the eastern part of the Clarion Clipperton Fracture Zone (North East Pacific), and implications for potential future deep-sea mining in this area. Mar. Biodivers.53, 1–19. doi: 10.1007/s12526-022-01326-9
44
Uhlenkott K. Simon-Lledó E. Vink A. Martínez Arbizu P. (2022). Investigating the benthic megafauna in the eastern Clarion Clipperton Fracture Zone (north-east Pacific) based on distribution models predicted with random forest. Sci. Rep.12, 8229. doi: 10.1038/s41598-022-12323-0
45
Uhlenkott K. Simon-Lledó E. Vink A. Martínez Arbizu P. (2023b). Habitat heterogeneity enhances megafaunal biodiversity at bathymetric elevations in the Clarion Clipperton Fracture Zone. Mar. Biodivers.53, 1–13. doi: 10.1007/s12526-023-01346-z
46
Uhlenkott K. Vink A. Kuhn T. Martínez Arbizu P. (2020). Predicting meiofauna abundance to define preservation and impact zones in a deep-sea mining context using random forest modelling. J. Appl. Ecol. 57, 1210–1221. doi: 10.1111/1365-2664.13621
47
Vanreusel A. Fonseca G. Danovaro R. Da Silva M. C. Esteves A. M. Ferrero T. et al . (2010). The contribution of deep-sea macrohabitat heterogeneity to global nematode diversity. Mar. Ecol.31, 6–20. doi: 10.1111/j.1439-0485.2009.00352.x
48
Vanreusel A. Hilario A. Ribeiro P. A. Menot L. Martínez Arbizu P. (2016). Threatened by mining, polymetallic nodules are required to preserve abyssal epifauna. Sci. Rep.6, 26808. doi: 10.1038/srep26808
49
Wedding L. M. Friedlander A. M. Kittinger J. N. Watling L. Gaines S. D. Bennett M. et al . (2013). From principles to practice: a spatial approach to systematic conservation planning in the deep sea. Proc. R. Soc B.280, 20131684. doi: 10.1098/rspb.2013.1684
50
Young C. M. (2009). “ Communities on Deep-Sea Hard Bottoms,” in Marine Hard Bottom Communities: Patterns, Dynamics, Diversity, and Change. Ed. WahlM. ( Springer, Berlin, Heidelberg), 39–60. doi: 10.1007/b76710_3
51
Zeppilli D. Pusceddu A. Trincardi F. Danovaro R. (2016). Seafloor heterogeneity influences the biodiversity–ecosystem functioning relationships in the deep sea. Sci. Rep.6, 26352. doi: 10.1038/srep26352
Summary
Keywords
abyss, deep sea, habitat heterogeneity, hard substrates, Atlantic Ocean
Citation
Riehl T, Budke L, Brandt A, Henseler M, Martínez Arbizu P, Momtazi F and Augustin N (2025) Confirmation of hard-substrate predictions in the abyssal Vema Fracture Zone. Front. Mar. Sci. 12:1674707. doi: 10.3389/fmars.2025.1674707
Received
28 July 2025
Accepted
08 September 2025
Published
26 September 2025
Volume
12 - 2025
Edited by
Ana Colaço, University of the Azores, Portugal
Reviewed by
Angelo Fraga Bernardino, Federal University of Espirito Santo, Brazil; Michael Vecchione, National Oceanic and Atmospheric Administration (NOAA), United States
Updates
Copyright
© 2025 Riehl, Budke, Brandt, Henseler, Martínez Arbizu, Momtazi and Augustin.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Torben Riehl, torben.riehl@senckenberg.de
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.