 Diego Cardeñosa
Diego Cardeñosa- Department of Biological Sciences, Florida International University, North Miami, FL, United States
Small-bodied hammerhead sharks (Sphyrna corona, S. media, and S. vespertina) are among the most threatened and understudied elasmobranchs, yet little is known about their distribution or current status. Here, I developed and validated species-specific environmental DNA (eDNA) assays to enable rapid, non-invasive detection of these species in remote and turbid coastal environments. Assays targeting mitochondrial NADH2 and Control Region sequences were tested in vitro for specificity, sensitivity, and quantitative performance. All assays showed high specificity with no cross-amplification of non-target species, and strong linearity (R² > 0.99), with efficiencies between 0.848 and 0.908. In situ validation within Uramba/Bahía Málaga National Natural Park (Colombia) confirmed the presence of all three species. The scalloped bonnethead was the most frequently detected species, aligning with acoustic telemetry, fishery-independent surveys, and local ecological knowledge. Detections of S. media and S. vespertina were less frequent but consistent with known species ranges and habitat use. These findings underscore the park’s significance as a conservation hotspot for small hammerheads and demonstrate the utility of eDNA for monitoring rare sharks. This study provides a scalable molecular toolkit for detecting data-deficient elasmobranchs and supports evidence-based conservation planning in coastal regions threatened by overfishing and habitat degradation.
1 Introduction
More than three-quarters of tropical and subtropical elasmobranchs are threatened with extinction mainly due to overfishing and habitat loss (Dulvy et al., 2021). Many of these species inhabit remote regions with limited scientific monitoring and weak fisheries management, meaning that conservation actions often arrive too late to prevent extinctions (White et al., 2019; Dulvy et al., 2021). Coastal environments have been identified as current shark mortality hotspots, where unselective fishing gear such as gillnets and longlines are commonly used (Worm et al., 2024). The widespread use of gillnets, trawl nets, and longlines by small-scale fisheries in these areas, while lower in volume compared to industrial operations (Carvalho et al., 2011), frequently involves some of the world’s most threatened elasmobranchs, particularly small coastal shark species, and thus represents a significant conservation concern (Roff et al., 2018; MacNeil et al., 2020; Simpfendorfer et al., 2023).
While large hammerhead sharks, such as the scalloped hammerhead (Sphyrna lewini), smooth hammerhead (S. zygaena), and great hammerhead (S. mokarran), have received increasing research attention and protection (Gallagher and Klimley, 2018), their smaller congeners remain largely understudied. These include the scalloped bonnethead (S. corona), scoophead shark (S. media), bonnethead shark (S. tiburo), Pacific bonnethead shark (S. vespertina), shovelhead shark (S. alleni), and golden hammerhead (S. tudes). These smaller Sphyrna species are endemic to the Atlantic and Eastern Pacific coasts of North, Central, and South America yet little is known about their ecology, distribution, or current conservation status. Small body size is itself ecologically relevant: these species tend to be restricted to shallow coastal habitats, where exposure to fisheries is high, and their limited reproductive output makes them especially vulnerable. Such patterns have been documented in analyses of life history and habitat among sharks, rays, and chimaeras, especially for species in shallow, coastal environments (Garcia et al., 2008; Dulvy et al., 2014). The International Union for the Conservation of Nature (IUCN) Red List of Threatened Species has assessed, based on the available data, S. corona and S. media as Critically Endangered and S. tiburo as Endangered (Pollom et al., 2020b; 2020a; 2021). Alarmingly, S. corona and S. media have already experienced local extinctions in Mexico, where they were last recorded in 1994 and 2007, respectively (Pérez-Jiménez, 2014).
However, recent findings by Herrera et al. (2024) provide new insights of the movement ecology of the scalloped bonnethead shark. Their study identified an important conservation site for this species along the Colombian Pacific coast using acoustic telemetry. The scalloped bonnethead appears to exhibit high site fidelity to small coastal areas, a trait that presents both an opportunity and a challenge for conservation: such residency enables localized protection but also makes populations extremely vulnerable to sustained fishing pressure (Herrera et al., 2024). To prevent further local extinctions, effective efforts are needed to identify and protect areas like those described by Herrera et al. (2024), where viable populations persist and targeted conservation actions can have the greatest impact.
Environmental DNA (eDNA) has emerged as a powerful conservation tool for detecting rare and threatened species (Budd et al., 2021; Ruiz‐Ramos et al., 2022; Faure et al., 2023), especially in environments where traditional non-invasive survey methods are limited or ineffective such as Baited Remove Underwater Video Stations (BRUVS) or visual surveys. This molecular approach is particularly valuable in remote or turbid habitats, where visual or fishing surveys in large geographical scales are impractical or too invasive. Environmental DNA allows for the non-invasive detection of species presence through genetic traces left in the environment, making it especially suitable for cryptic or low-abundance species (Jerde et al., 2011; Xia et al., 2021) like these small hammerhead sharks. When paired with traditional ecological knowledge and spatially explicit management strategies, eDNA can support the rapid identification of new conservation hotspots and provide early warning signals of population declines, enhancing our ability to act before species are lost (Spear et al., 2021; Hata et al., 2022).
Therefore, the goal of this study was to develop and apply species-specific eDNA assays to detect some of the most threatened coastal elasmobranchs. By focusing on species with limited distributional data and high extinction risk, this study aimed to provide a non-invasive, scalable method to assess their presence in poorly studied regions. Specifically, I designed and validated eDNA assays targeting highly threatened small-bodied hammerhead sharks (S. corona, S. media, S. vespertina) to evaluate their occurrence in coastal habitats of the Eastern Pacific. These tools are intended to support ongoing conservation efforts by enabling rapid biodiversity assessments, identifying critical habitats, and informing spatial management strategies.
2 Methodology
2.1 Assay design
Mitochondrial NADH dehydrogenase subunit 2 (ND2) and Control Region (CR) sequences from all hammerhead sharks (Family Sphyrnidae), other elasmobranch species common to the Eastern Pacific, and several distantly related co-occurring species (Supplementary Material S1, S2) were downloaded from GenBank and aligned (i.e., MUSCLE alignment) in Geneious Prime v.2025.1.2. Short regions (100–200 bp) containing at least three nucleotide mismatches at the 3′ end of the forward or primer were identified by eye to distinguish Sphyrna corona (ND2), S. media (ND2), and S. vespertina (CR) from all other species. Given the lack of CR sequences for S. corona, and S. media in GenBank, DNA from tissue samples previously collected in the field were extracted using the Qiagen DNeasy Blood & Tissue kit following the manufacturer’s instructions. CR amplification was conducted using the primers Pro-L 5′-AGGGRAAGGAGGGTCAAACT-3′) and 12S rRNA (heavy strand primer: 282 H 5′-AAGGCTAGGACCAAACCT-3′, Keeney et al., 2003). Each 25 µL qPCR reaction contained 12.5 µL of GoTaq Hotstart Green Master Mix (Promega), 1.5 µL of each primer (10 µM), 7.5 µL of molecular-grade water, and 2.0 µL of genomic DNA, following the thermal conditions described by Duncan et al. (2006). All PCR products were cleaned using ExoSAP-IT (Affymetrix, Inc., Santa Clara, CA, USA) and sequenced twice using the Big Dye Terminator v3.1 cycle sequencing kit (Applied Biosystems, Foster City, CA, USA). Sequencing was performed on an ABI 3730 DNA Analyzer (Applied Biosystems). All forward and reverse sequences were checked by eye and priming sites were trimmed using Geneious Pro v. 3.6.1 (http://www.geneious.com). Trimmed sequences were uploaded to GenBank with Accession Numbers: XXXXXXX).
Primers were designed manually following standard criteria: length 18–25 bp, GC content ~40–60%, avoidance of long homopolymers (>4 identical bases), minimal 3′ complementarity, and amplicon length restricted to 90–180 bp to suit degraded eDNA templates (Kutyavin et al., 2000; Bustin et al., 2009; Wilcox et al., 2013). Melting temperatures (Tm) were checked with IDT OligoAnalyzer (https://www.idtdna.com/calc/analyzer) to ensure ≤2°C difference between primer pairs, and to confirm the absence of strong hairpins (i.e., hairpin Tm equal or above annealing temperature), self-dimers, or heterodimers (i.e., ΔG< -9 kcal/mole). Corresponding hydrolysis probes were designed with Tm values ≥6 °C higher than their primer pairs (Table 1).

Table 1. Primer and probe sequences for each species-specific assay, including melting temperatures (Tm), amplicon size, and the mitochondrial locus targeted by each assay.
2.2 In vitro validation
Specificity was assessed using synthetic gBlocks containing the target amplicon sequences of each target species (Supplementary Material S3) each at 1 × 105 copies/µL, and genomic DNA extractions of co-occurring elasmobranchs (i.e., scalloped hammerhead S. lewini, great hammerhead S. mokarran, smooth hammerhead S. zygaena, bull shark Carcharhinus leucas, Pacific smalltail shark C. cerdale, and longtail stingray Hypanus longus). Each primer–probe assay was tested across a gradient of annealing temperatures (58–64 °C) on a MIC qPCR thermocycler (Biomolecular Systems) to determine the highest temperature yielding specific amplification of the target species (Table 1). Each 25 µL qPCR reaction contained 12.5 µL of TaqMan Environmental Master Mix 2.0 (Applied Biosystems), 1.0 µL of each primer (10 µM), 1.0 µL of hydrolysis probe (5 µM), 8.5 µL of molecular-grade water, and 1.0 µL of gBlock DNA (1 × 105 copies/µL). Thermal cycling consisted of an initial denaturation at 95 °C for 10 min followed by 50 cycles of 95 °C for 15 s, assay-specific annealing temperature for 30 s (Table 1), and 72 °C for 30 s. Fluorescence was measured during the extension phase of each cycle. A no-template control (NTC) was included in every run to monitor contamination.
Once specificity was confirmed, assay efficiency, sensitivity (Limit of Detection, LOD), and quantitative performance (Limit of Quantification, LOQ) were evaluated following Klymus et al. (2020). Ten-fold serial dilutions of each target gBlock were prepared from 1 × 107 to 10 copies/µL. Efficiency was assessed in triplicate for each dilution under the optimized qPCR conditions. Standard curves were generated by plotting quantification cycle (Cq) values against log-transformed DNA concentrations (Figure 1). The MIC software was used to calculate efficiency, slope, intercept, and R².
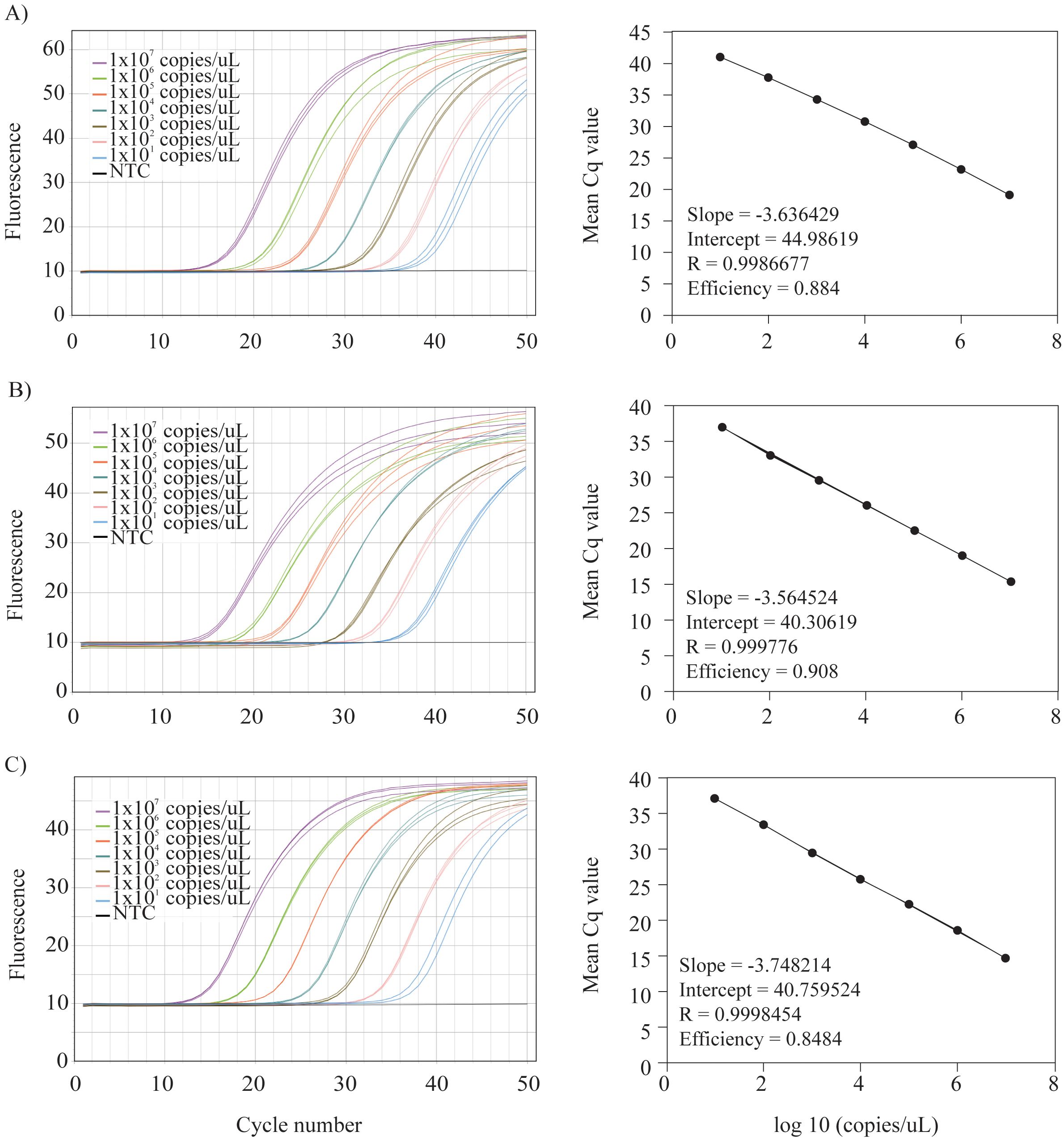
Figure 1. In vitro validation of eDNA assays. (A) Ten-fold dilution series for the S. corona assay with the standard curve depicting slope, intercept, R2, and efficiency of the assay. (B) Ten-fold dilution series for the S. media assay with the standard curve depicting slope, intercept, R2, and efficiency of the assay. (C) Ten-fold dilution series for the S. tiburo assay with the standard curve depicting slope, intercept, R2, and efficiency of the assay.
LOD and LOQ were determined by running 20 replicates each at 10, 5, and 1 copy/µL. The LOD was defined as the lowest concentration at which ≥ 95% of replicates amplified. The LOQ was defined as the lowest concentration at which ≥ 95% of replicates amplified and the coefficient of variation (CV) of Cq values was ≤ 35%, indicating acceptable quantitative precision. CV was calculated as:
2.3 In situ validation
To validate assays with field samples, 10 L of seawater were collected at 2 m depth from five sites within Uramba Bahía Málaga National Natural Park (Valle del Cauca, Colombia; Figure 2) using a novel eDNA sampling methodology to minimize field contamination. Automated pumps (Aquatic Labs) and factory-sterile 0.45 µm polyethersulfone cartridge filters (Waterra; 600 cm²) housed in sealed casings were used for the field sampling. The use of sealed 0.45 µm PES cartridge filters (Waterra) followed established eDNA practices (Spens et al., 2017; Hunter et al., 2019; Govindarajan et al., 2022) and was adopted here to improve filtration efficiency in turbid tropical waters. While controlled comparisons of filter types were beyond the scope of this study, the consistency of detection across species demonstrates the utility of this approach for species monitoring.
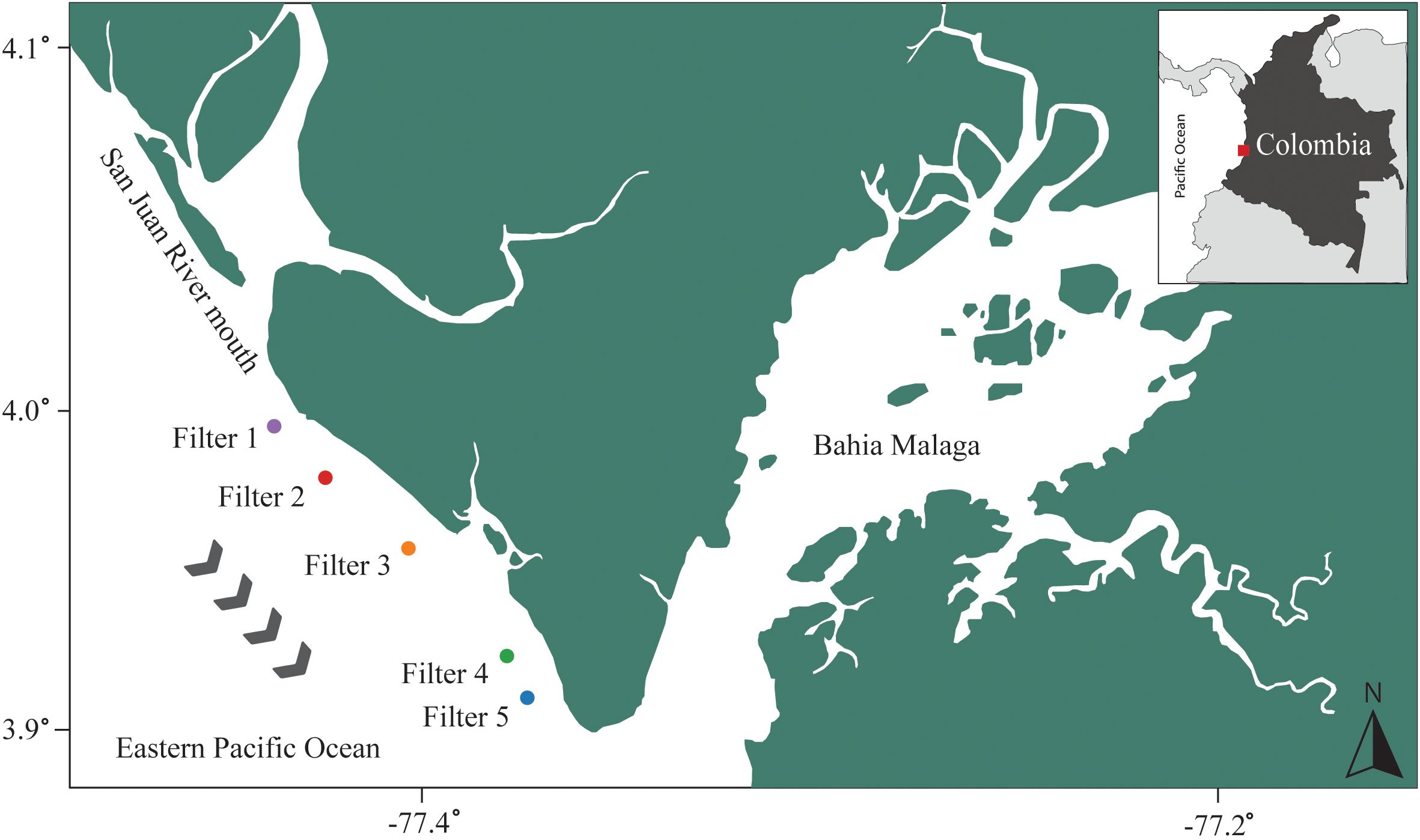
Figure 2. Map of field sampling locations within the Uramba/Bahia Malaga National Natural Park, Colombia. Gray arrows depict the direction of the current during sampling.
Because all water was filtered through factory-sterile, single-use 0.45-µm PES cartridge filters (Waterra; 600 cm²) housed inside sealed casings, the filter membrane was never exposed to ambient air or deck splashes prior to pumping. A custom-made adapter mating the pump to the cartridge housing prevents water or air from contacting the membrane except under active suction (Supplementary Material S4). The automated pumps drew water directly from the water column into the sealed housing; housings were not opened on deck. After filtration, housings were sealed with Parafilm and frozen for transport. Under this closed, in-line configuration, a conventional “field blank” (pouring molecular-grade water through an open apparatus in the field) would not meaningfully interrogate any exposure pathway experienced by the filters, because the membrane is not accessible to aerosols or incidental splashes prior to initiating flow. Moreover, manipulating bottled water on deck can itself introduce exogenous DNA. Therefore, contamination control relied on (i) pre-field sterilization of adapters and pumps with 10% bleach (≥ 10 min contact) followed by multiple rinses with ultrapure water, (ii) physical separation of pre- and post-PCR spaces with UV/bleach decontamination, (iii) no-template controls (NTCs) on every run, and (iv) Sanger sequence confirmation of all putative positives to verify taxonomic identity. Filters remained sealed until processing in a clean laboratory.
In the laboratory, filters were aseptically opened and cut into six sections and extracted separately to maximize DNA recovery while maintaining high DNA concentration (Hinlo et al., 2017; Spens et al., 2017; Govindarajan et al., 2022). Each section was placed in a separate sterile 5 mL tube for full-filter DNA extraction using the Qiagen DNeasy Blood & Tissue Kit with minor modifications (Govindarajan et al., 2022). Briefly, 900 µL Buffer ATL and 100 µL proteinase K were added to each tube, followed by incubation at 56 °C for 3 h with periodic vortexing. After incubation, 1,000 µL Buffer AL and ethanol were added, and lysates were sequentially loaded onto a single spin column in six steps. Washes followed the manufacturer’s protocol, and DNA was eluted in 80 µL AE buffer per tube. DNA extracts from the six sections were pooled and purified using the Zymo OneStep PCR Inhibitor Removal Kit prior to qPCR. Although only modest elution volumes were used, pooling across all sections ensures that the full surface area of the filter was processed. Each pooled extract was run in eight technical replicates using the optimized assay conditions.
All pre-PCR procedures (filter handling, DNA extraction) were performed in a designated clean laboratory physically separated from the PCR workspace. PCRs were prepared in a UV-sterilized AirClean 600 PCR Workstation (AirClean Systems). Pipettes and tips were UV-sterilized prior to use, and all surfaces were cleaned with 10% bleach followed by Milli-Q water before each run to minimize contamination.
For a detection to be classified as positive, the following criteria from Budd et al. (2021) were applied: (1) at least one qPCR technical replicate must yield amplification from a single field sample; (2) no amplification may occur in the NTC; and (3) the amplified fragment must show 100% sequence identity to the target species as confirmed by Sanger sequencing. All qPCR products were cleaned using ExoSAP-IT (Affymetrix, Inc., Santa Clara, CA, USA) and sequenced twice using the Big Dye Terminator v3.1 cycle sequencing kit (Applied Biosystems, Foster City, CA, USA). Sequencing was performed on an ABI 3730 DNA Analyzer (Applied Biosystems). All forward and reverse sequences were checked by eye and priming sites were trimmed using Geneious Pro v. 3.6.1 (http://www.geneious.com). Trimmed sequences were compared to BLAST (GenBank) database to check for 100% matches with each of the target species.
3 Results
In vitro validation demonstrated that all assays were highly specific to their target species, with no amplification observed in non-target species after 50 cycles at the most stringent annealing temperatures (Table 1). All no-template controls confirmed the absence of reagent contamination. Standard curves estimated efficiencies between 0.848 and 0.908 with R² values > 0.99 for all assays (Figure 1). Sensitivity (LOD) and quantitative performance (LOQ) were estimated at 10 copies/µL for the S. corona and S. media assays, and 5 copies/µL for the S. vespertina assay.
In situ validation confirmed the presence of all three species within the Uramba/Bahía Málaga National Natural Park, Colombia (Figure 3). Sphyrna corona was the most frequently detected, with positive amplification in four of the five filters and 0–7 positive technical replicates per filter (out of eight; Figure 3). Sphyrna vespertina was detected in three filters with 0–3 positive replicates per filter, while S. media was detected in three filters with one positive replicate per filter (Figure 3). No species were detected in any replicates of filter 1, the northernmost sampling site (Figure 2). All sequences of positive field detections returned 100% identity to S. corona, S. media, and S. vespertina.
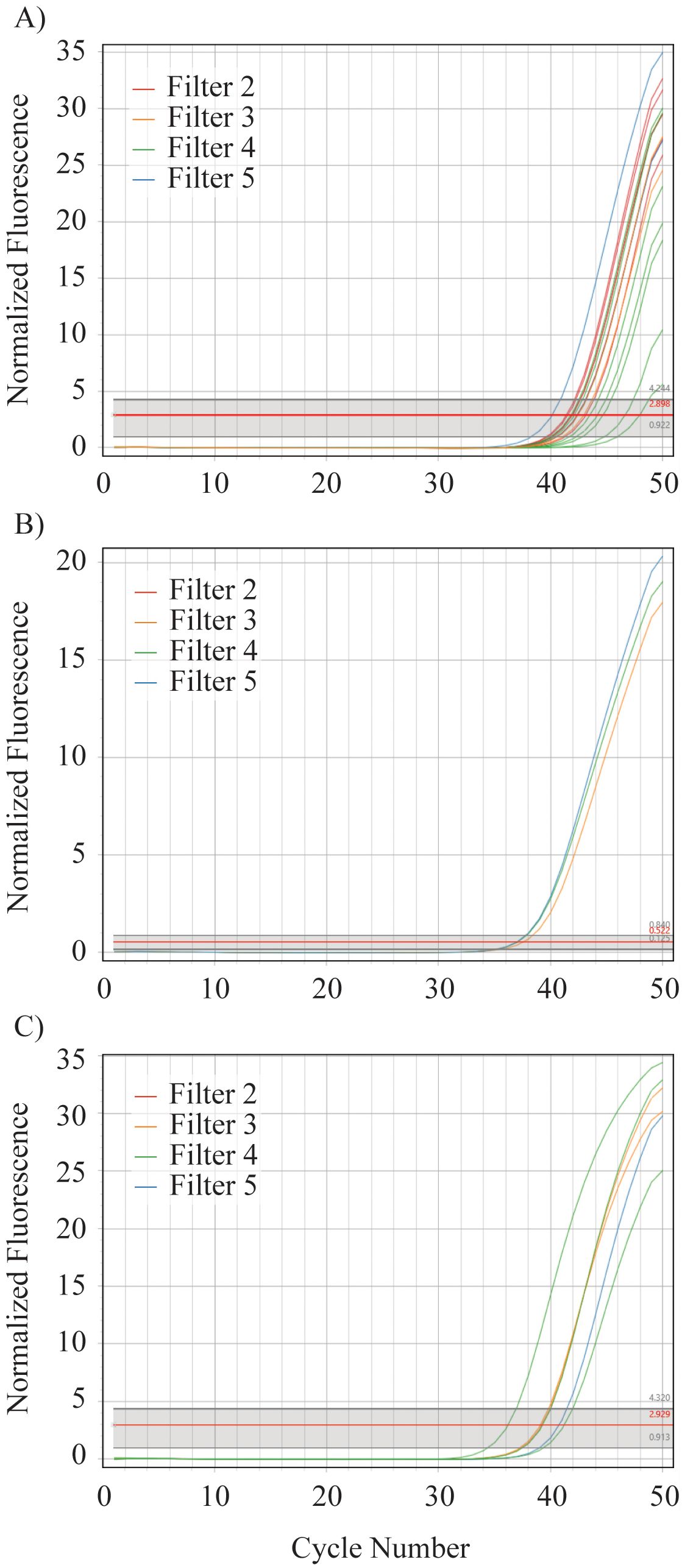
Figure 3. Positive field amplifications for (A) S. corona, (B) S. media, and (C) S. tiburo. Filter 1 is not included as it yielded no positive amplifications.
Consistent with the low contamination risk of the closed filtration system, no NTCs amplified, and all positives were sequence-confirmed; additionally, the absence of detections at the northernmost site (filter 1) provided an internal negative control supporting the lack of field contamination.
4 Discussion
This study presents the first successful development and application of species-specific eDNA assays for three small-bodied, highly threatened hammerhead sharks, Sphyrna corona, S. media, and S. vespertina. In vitro validation confirmed high specificity and sensitivity of all assays, with no cross-amplification of non-target species and no contamination in negative controls. All positive field detections returned 100% sequence identity to the target species.
Because eDNA persistence in seawater is typically limited to hours to days (Collins et al., 2018; Allan et al., 2021) detections reported here are most consistent with recent local presence. Nonetheless, water movement may transport DNA away from the precise location of individuals, so detections should be interpreted as evidence of local occurrence within the broader sampling area rather than pinpointing exact animal locations.
Beyond field applications, the assays were designed with mismatches at the 3′ end of the forward primer, a feature that enhances specificity and makes them adaptable to other contexts. For example, they could be applied to species identification in fisheries and trade monitoring (Cardeñosa et al., 2018), or used with alternative chemistries (e.g., SYBR Green) when hydrolysis probes are impractical.
Standard curves showed high linearity (R² > 0.99; Figure 1), indicating consistent quantification across a wide range of DNA concentrations and affirming the assays’ reliability for quantifying DNA in the sample. However, amplification efficiencies ranged from 0.848 to 0.908, slightly below the optimal range of 0.9–1.1 typically recommended for qPCR-based diagnostics (Goldberg et al., 2016; Klymus et al., 2020). While these values remain acceptable for ecological applications, they may reflect minor inefficiencies potentially caused by potential secondary structure formation in some primer–probe sets, which could affect the accuracy of absolute quantification. Although assay efficiency might improve at lower annealing temperatures, this would come at the cost of reduced specificity and an increased risk of non-target amplification.
The assays also demonstrated strong sensitivity, with LODs estimated at 10 copies/µL for S. corona and S. media, and 5 copies/µL for S. vespertina, meaning that trace levels of DNA from these species can be consistently detected. The LOQs met the criteria of ≥95% amplification success and ≤35% Cq variation across replicates, supporting the use of these assays for semi-quantitative comparisons. However, caution is warranted when interpreting very low detection signals near the LOD, especially under field conditions where DNA degradation, inhibition, or patchy distribution could influence results (Goldberg et al., 2016; Spear et al., 2021). For future applications, these performance metrics support the use of the assays for presence/absence detection and site-level comparisons, but not yet for estimating absolute abundance or biomass without additional calibration against known density data (Cardás et al., 2020; Spear et al., 2021). Still, the robust specificity, high sensitivity, and reproducibility of these assays provide a strong foundation for broader monitoring programs and conservation assessments of these rare species.
The taxonomy of the Sphyrna tiburo species complex has been recently revised with the formal description of S. alleni (Gonzalez et al., 2024) and the taxonomic reevaluation of S. vespertina (Aroca et al., 2022). As currently understood, S. tiburo is restricted to the coastlines of the United States and Mexico, while S. alleni occurs from Belize to southern Brazil in the western Atlantic (Gonzalez et al., 2024), and S. vespertina ranges from Mexico to northern Peru along the eastern Pacific (Aroca et al., 2022). Although the assays presented in this study were designed to target small-bodied hammerheads from the eastern Pacific, the assay used for S. vespertina was originally developed using S. tiburo reference sequences due to the absence of voucher-confirmed S. vespertina sequences in GenBank at the time of design. Nonetheless, comparative alignment of available sequences revealed no mismatches at the primer or probe annealing sites among S. tiburo, S. alleni, and S. vespertina. This indicates that the assay is likely to detect any of these three taxa across their respective ranges. Therefore, while the current application was focused on the eastern Pacific coast of Colombia, these assays can also be used to detect S. tiburo, S. alleni, and S. media along the Atlantic and Pacific coastlines of the Americas where these species occur.
The detection of all three target species within Uramba/Bahía Málaga National Natural Park underscores the area’s importance as a conservation hotspot for small-bodied hammerhead sharks. Sphyrna corona was the most frequently detected species, found in four of five sampling sites and in seven of eight technical replicates at filter 4 (Figures 2, 3). This high detection rate supports recent telemetry data indicating strong site fidelity by S. corona to this particular location within the park (Herrera et al., 2024). While such residency presents an opportunity for localized protection, it also increases vulnerability to sustained fishing pressure. The presence of S. corona within the boundaries of the National Park, despite its apparent extirpation from parts of its former range (e.g., Mexico; Pérez-Jiménez, 2014), further highlights the urgency of prioritizing this area for targeted conservation efforts.
In contrast, S. vespertina and S. media were each detected at only three sites, and typically in low replicate numbers (Figure 3). These patterns may reflect lower local abundances, more transient habitat use, or species-specific ecological preferences. Notably, all three species were absent from filter 1, the northernmost sampling site, potentially due to environmental factors such as reduced salinity from freshwater input by the nearby San Juan River (Figure 2), or anthropogenic disturbances (e.g., higher fishing pressure; Herrera et al., 2024). The detection patterns observed here were consistent with fishery-independent surveys (authors, unpublished data), and mirrored local ecological knowledge from fishing communities, who perceive S. corona as more common than S. media or S. vespertina. Similarly, the number of positive qPCR replicates per filter aligned with preliminary catch data and acoustic detections, suggesting potential for semi-quantitative inference. Previous studies have used the proportion of positive replicates per filter as a proxy for relative abundance in rare or low-density species (Wilcox et al., 2013; Mauvisseau et al., 2019; Shelton et al., 2022). Although further research is needed, these results suggest the assays presented here may hold promise for future abundance monitoring and trend detection in these data-deficient elasmobranchs.
Although these results highlight the conservation importance of the Uramba/Bahía Málaga National Park, the field sampling design had two key limitations. First, all sites were located within the National Park, which precludes direct comparisons with adjacent areas such as the nearby San Juan River estuary. Second, sampling was conducted at a single time point, limiting my ability to capture potential seasonal or tidal variation in species occurrence. Future studies incorporating multi-season and multi-region surveys will be essential to assess the temporal stability of detections and to evaluate broader distributional patterns of these species.
Despite the limitations of the field sampling design, this study highlights the power of eDNA to detect low-abundance elasmobranchs in remote and turbid environments where traditional methods such as BRUVS or net surveys are logistically difficult or ecologically disruptive. The combination of species-specific assays and rigorous sequence verification ensures both high sensitivity and taxonomic accuracy, minimizing false positives and enabling reliable detection of cryptic species. Given the limited resources available for monitoring most threatened coastal sharks, these tools can provide an accessible and scalable approach to support rapid biodiversity assessments and guide spatial protections. Overall, my findings emphasize the urgent need for proactive conservation interventions focused on lesser-known shark species. While large-bodied hammerheads have attracted increasing attention from researchers and policymakers, their smaller congeners remain overlooked despite their comparable extinction risk. By providing a field-validated method to track their presence, and potentially relative abundance, this study contributes a critical toolset for reversing data deficiencies and facilitating informed management of some of the world’s most threatened elasmobranchs.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Ethics statement
The animal study was approved by Angel Rayo Florida International University. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
DC: Writing – original draft, Investigation, Funding acquisition, Resources, Formal Analysis, Writing – review & editing, Visualization, Methodology, Data curation, Project administration, Conceptualization, Validation.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. Primary funding came from The Explorers Club, Save Our Seas Foundation, the National Philanthropic Trust, and National Geographic Society.
Acknowledgments
I want to thank Maria Alejandra Herrera for her co-PI role in the Cachuda Project and for assisting with the water sample collection in the field. Thanks to the communities of Puerto España, La Barra, Ladrilleros, Juan Chaco and La Plata for their help in the field and for their commitment with the Cachuda Project and its conservation objectives. I want to thank National Natural Parks of Colombia for their support. Special thanks to The Explorers Club, the National Philanthropic Trust, and Save our Seas Foundation, for their funding support and believing in the conservation outcomes of the project.
Conflict of interest
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2025.1688088/full#supplementary-material
References
Allan E. A., Zhang W. G., Lavery A. C., and Govindarajan A. F. (2021). Environmental DNA shedding and decay rates from diverse animal forms and thermal regimes. Environ. DNA 3, 492–514. doi: 10.1002/edn3.141
Aroca A. K., Tavera J., and Torres Y. (2022). Molecular and morphological evaluation of the bonnethead shark complex Sphyrna tiburo (Carcharhiniformes: Sphyrnidae). Environ. Biol. Fish. 105, 1643–1658. doi: 10.1007/s10641-022-01358-x
Budd A. M., Cooper M. K., Port A. L., Schils T., Mills M. S., Deinhart M. E., et al. (2021). First detection of critically endangered scalloped hammerhead sharks (Sphyrna lewini) in Guam, Micronesia, in five decades using environmental DNA. Ecol. Indic. 127, 107649. doi: 10.1016/j.ecolind.2021.107649
Bustin S. A., Benes V., Garson J. A., Hellemans J., Huggett J., Kubista M., et al. (2009). The MIQE guidelines: minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 55, 611–622. doi: 10.1373/clinchem.2008.112797
Cardás J. B., Deconinck D., Márquez I., Torre P. P., Garcia-Vazquez E., and MaChado-Schiaffino G. (2020). New eDNA based tool applied to the specific detection and monitoring of the endangered European eel. Biol. Conserv. 250, 108750. doi: 10.1016/j.biocon.2020.108750
Cardeñosa D., Quinlan J., Shea K. H., and Chapman D. D. (2018). Multiplex real-time PCR assay to detect illegal trade of CITES-listed shark species. Sci. Rep. 8, 1–10.
Carvalho N., Edwards-Jones G., and Isidro E. (2011). Defining scale in fisheries: Small versus large-scale fishing operations in the Azores. Fish. Res. 109, 360–369. doi: 10.1016/j.fishres.2011.03.006
Collins R. A., Wangensteen O. S., O’Gorman E. J., Mariani S., Sims D. W., and Genner M. J. (2018). Persistence of environmental DNA in marine systems. Commun. Biol. 1, 185. doi: 10.1038/s42003-018-0192-6
Dulvy N. K., Fowler S. L., Musick J. A., Cavanagh R. D., Kyne P. M., Harrison L. R., et al. (2014). Extinction risk and conservation of the world’s sharks and rays. eLife 3, e00590–e00590. doi: 10.7554/eLife.00590
Dulvy N. K., Pacoureau N., Rigby C. L., Pollom R. A., Jabado R. W., Ebert D. A., et al. (2021). Overfishing drives over one-third of all sharks and rays toward a global extinction crisis. Curr. Biol. 31, 4773–4787. doi: 10.1016/j.cub.2021.08.062
Duncan K. M., Martin A. P., Bowen B. W., and Couet H. D. G. (2006). Global phylogeography of the scalloped hammerhead shark (Sphyrna lewini).. Mol. Ecol. 15, 2239–2251. doi: 10.1111/j.1365-294x.2006.02933.x
Faure N., Manel S., Macé B., Arnal V., Guellati N., Holon F., et al. (2023). An environmental DNA assay for the detection of Critically Endangered angel sharks (Squatina spp.). Aquat. Conserv.: Mar. Freshw. Ecosyst. 1–10. doi: 10.1002/aqc.3954
Gallagher A. J. and Klimley A. P. (2018). The biology and conservation status of the large hammerhead shark complex: the great, scalloped, and smooth hammerheads. Rev. Fish. Biol. Fish. 28, 777–794. doi: 10.1007/s11160-018-9530-5
Garcia V. B., Lucifora L. O., and Myers R. A. (2008). The importance of habitat and life history to extinction risk in sharks, skates, rays and chimaeras. Proc. R. Soc B.: Biol. Sci. 275, 83–89. doi: 10.1098/rspb.2007.1295
Goldberg C. S., Turner C. R., Deiner K., Klymus K. E., Thomsen P. F., Murphy M. A., et al. (2016). Critical considerations for the application of environmental DNA methods to detect aquatic species. Methods Ecol. Evol. 7, 1299–1307. doi: 10.1111/2041-210x.12595
Gonzalez C., Postaire B., Driggers W., Caballero S., and Chapman D. (2024). Sphyrna alleni sp. nov., a new hammerhead shark (Carcharhiniformes, Sphyrnidae) from the Caribbean and the Southwest Atlantic. Zootaxa 5512, 491–511. doi: 10.11646/zootaxa.5512.4.2
Govindarajan A. F., McCartin L., Adams A., Allan E., Belani A., Francolini R., et al. (2022). Improved biodiversity detection using a large-volume environmental DNA sampler with in situ filtration and implications for marine eDNA sampling strategies. Deep Sea Res. Part Oceanogr. Res. Pap. 189, 103871. doi: 10.1016/j.dsr.2022.103871
Hata H., Ogasawara K., and Yamashita N. (2022). Population decline of an endangered unionid, Pronodularia Japanensis, in streams is revealed by eDNA and conventional monitoring approaches. Hydrobiologia 849, 2635–2646. doi: 10.1007/s10750-022-04852-6
Herrera M., Cardeñosa D., Papastamatiou Y., Vaudo J., Bermúdez-Rivas C., and Shivji M. (2024). High residency of a Critically Endangered hammerhead shark to a small area: implications for marine protected area management and design. Mar. Ecol. Prog. Ser. 743, 47–63. doi: 10.3354/meps14658
Hinlo R., Gleeson D., Lintermans M., and Furlan E. (2017). Methods to maximise recovery of environmental DNA from water samples. PloS One 12, e0179251. doi: 10.1371/journal.pone.0179251
Hunter M. E., Ferrante J. A., Meigs-Friend G., and Ulmer A. (2019). Improving eDNA yield and inhibitor reduction through increased water volumes and multi-filter isolation techniques. Sci. Rep. 9, 5259. doi: 10.1038/s41598-019-40977-w
Jerde C. L., Mahon A. R., Chadderton W. L., and Lodge D. M. (2011). Sight-unseen” detection of rare aquatic species using environmental DNA. Conserv. Lett. 4, 150–157. doi: 10.1111/j.1755-263x.2010.00158.x
Keeney D. B., Heupel M., Hueter R. E., and Heist E. J. (2003). Genetic heterogeneity among blacktip shark, Carcharhinus limbatus, continental nurseries along the U.S. Atlantic and Gulf of Mexico. Mar. Biol. 143, 1039–1046. doi: 10.1007/s00227-003-1166-9
Klymus K. E., Ramos D. V. R., Thompson N. L., and Richter C. A. (2020). Development and testing of species-specific quantitative PCR assays for environmental DNA applications. J. Vis. Exp. 1–25. doi: 10.3791/61825
Kutyavin I. V., Afonina I. A., Mills A., Gorn V. V., Lukhtanov E. A., Belousov E. S., et al. (2000). 3′-Minor groove binder-DNA probes increase sequence specificity at PCR extension temperatures. Nucleic Acids Res. 28, 655–661. doi: 10.1093/nar/28.2.655
MacNeil M. A., Chapman D. D., Heupel M., Simpfendorfer C. A., Heithaus M., Meekan M., et al. (2020). Global status and conservation potential of reef sharks. Nature 583, 801–806. doi: 10.1038/s41586-020-2519-y
Mauvisseau Q., Burian A., Gibson C., Brys R., Ramsey A., and Sweet M. (2019). Influence of accuracy, repeatability and detection probability in the reliability of species-specific eDNA based approaches. Sci. Rep. 9, 580. doi: 10.1038/s41598-018-37001-y
Pérez-Jiménez J. C. (2014). Historical records reveal potential extirpation of four hammerhead sharks (Sphyrna spp.) in Mexican Pacific waters. Rev. Fish. Biol. Fish. 24, 671–683. doi: 10.1007/s11160-014-9353-y
Pollom R., Avalos C., Bizzarro J., Burgos-Vázquez M., Cevallos A., Charvet P., et al. (2020a). “The IUCN Red List of Threatened Species 2020: e.T60201A3091753,” in Sphyrna media. Available online at: https://dx.doi.org/10.2305/IUCN.UK.2020-3.RLTS.T60201A3091753.en.
Pollom R., Carlson J., Charvet P., Avalos C., Bizarro J., Blanco-Parra M., et al. (2021). “The IUCN Red List of Threatened Species 2021: e.T39387A205765567,” in Sphyrna tiburo. Available online at: https://dx.doi.org/10.2305/IUCN.UK.2021-3.RLTS.T39387A205765567.en.
Pollom R., Jiménez J. P., Bizzarro J., Burgos-Vázquez M., Cevallos A., Espinoza M., et al. (2020b). “The IUCN Red List of Threatened Species 2020: e.T44591A124434064,” in Sphyrna corona. Available online at: https://dx.doi.org/10.2305/IUCN.UK.2020-3.RLTS.T44591A124434064.en.
Roff G., Brown C. J., Priest M. A., and Mumby P. J. (2018). Decline of coastal apex shark populations over the past half century. Commun. Biol. 223, 1–11. doi: 10.1038/s42003-018-0233-1
Ruiz-Ramos D. V., Meyer R. S., Toews D., Stephens M., Kolster M. K., and Sexton J. P. (2022). Environmental DNA (eDNA) detects temporal and habitat effects on community composition and endangered species in ephemeral ecosystems: A case study in vernal pools. Environ. Dna 1–17. doi: 10.1002/edn3.360
Shelton A. O., Ramón-Laca A., Wells A., Clemons J., Chu D., Feist B. E., et al. (2022). Environmental DNA provides quantitative estimates of Pacific hake abundance and distribution in the open ocean. Proc. R. Soc B. 289, 20212613. doi: 10.1098/rspb.2021.2613
Simpfendorfer C. A., Heithaus M. R., Heupel M. R., MacNeil M. A., Meekan M., Harvey E., et al. (2023). Widespread diversity deficits of coral reef sharks and rays. Science 380, 1155–1160. doi: 10.1126/science.ade4884
Spear M. J., Embke H. S., Krysan P. J., and Zanden M. J. V. (2021). Application of eDNA as a tool for assessing fish population abundance. Environ. DNA 3, 83–91. doi: 10.1002/edn3.94
Spens J., Evans A. R., Halfmaerten D., Knudsen S. W., Sengupta M. E., Mak S. S. T., et al. (2017). Comparison of capture and storage methods for aqueous macrobial eDNA using an optimized extraction protocol: advantage of enclosed filter. Methods Ecol. Evol. 8, 635–645. doi: 10.1111/2041-210x.12683
White W. T., Charles C., Kyne P. M., and Harris M. (2019). Lost before found: A new species of whaler shark Carcharhinus obsolerus from the Western Central Pacific known only from historic records. PloS One 14, e0209387–24. doi: 10.1371/journal.pone.0209387
Wilcox T. M., McKelvey K. S., Young M. K., Jane S. F., Lowe W. H., Whiteley A. R., et al. (2013). Robust detection of rare species using environmental DNA: the importance of primer specificity. PloS One 8, e59520. doi: 10.1371/journal.pone.0059520
Worm B., Orofino S., Burns E. S., D’Costa N. G., Feitosa L. M., Palomares M. L. D., et al. (2024). Global shark fishing mortality still rising despite widespread regulatory change. Science 383, 225–230. doi: 10.1126/science.adf8984
Keywords: Sphyrna corona, Sphyrna media, Sphyrna vespertina, Sphyrna tiburo, Sphyrna alleni, Eastern Pacific
Citation: Cardeñosa D (2025) Ghosts of the current: environmental DNA assays to detect conservation priority areas for three critically endangered hammerhead sharks. Front. Mar. Sci. 12:1688088. doi: 10.3389/fmars.2025.1688088
Received: 18 August 2025; Accepted: 30 September 2025;
Published: 15 October 2025.
Edited by:
Patti Virtue, University of Tasmania, AustraliaReviewed by:
Wang Minxiao, Chinese Academy of Sciences (CAS), ChinaMarcelo Merten Cruz, Federal University of Pará, Brazil
Copyright © 2025 Cardeñosa. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Diego Cardeñosa, ZGNhcmRlbm9AZml1LmVkdQ==